
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 25 May 2016
Sec. Plant Pathogen Interactions
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.00720
This article is part of the Research Topic Mycorrhiza in tropical and neotropical ecosystems View all 12 articles
 Martha V. T. Cely1
Martha V. T. Cely1 Admilton G. de Oliveira2
Admilton G. de Oliveira2 Vanessa F. de Freitas2Marcelo B. de Luca2
Vanessa F. de Freitas2Marcelo B. de Luca2 André R. Barazetti2Igor M. O. dos Santos2Barbara Gionco2Guilherme V. Garcia3Cássio E. C. Prete4
André R. Barazetti2Igor M. O. dos Santos2Barbara Gionco2Guilherme V. Garcia3Cássio E. C. Prete4 Galdino Andrade2*
Galdino Andrade2*Nutrient availability is an important factor in crop production, and regular addition of chemical fertilizers is the most common practice to improve yield in agrosystems for intensive crop production. The use of some groups of microorganisms that have specific activity providing nutrients to plants is a good alternative, and arbuscular mycorrhizal fungi (AMF) enhance plant nutrition by providing especially phosphorus, improving plant growth and increasing crop production. Unfortunately, the use of AMF as an inoculant on a large scale is not yet widely used, because of several limitations in obtaining a large amount of inoculum due to several factors, such as low growth, the few species of AMF domesticated under in vitro conditions, and high competition with native AMF. The objective of this work was to test the infectivity of a Rhizophagus clarus inoculum and its effectiveness as an alternative for nutrient supply in soybean (Glycine max L.) and cotton (Gossypium hirsutum L.) when compared with conventional chemical fertilization under field conditions. The experiments were carried out in a completely randomized block design with five treatments: Fertilizer, AMF, AMF with Fertilizer, AMF with 1/2 Fertilizer, and the Control with non-inoculated and non-fertilized plants. The parameters evaluated were AMF root colonization and effect of inoculation on plant growth, nutrient absorption and yield. The results showed that AMF inoculation increased around 20 % of root colonization in both soybean and cotton; nutrients analyses in vegetal tissues showed increase of P and nitrogen content in inoculated plants, these results reflect in a higher yield. Our results showed that, AMF inoculation increase the effectiveness of fertilizer application in soybean and reduce the fertilizer dosage in cotton.
Nutrient availability is crucial to plant growth and crop production. This is influenced by several factors such as the chemical and physical properties of soil, climate and crop type. Crop production in tropical soils requires large amounts of chemical fertilizers, which enhance nutrient release and availability for plant nutrition (Miransari, 2011). Soybean (Glycine max L.) is a legume plant, of Fabaceae family. It is cultivated on large scale because has good adaptability to different soil and climatic conditions. Brazil is the second largest world producer of soybeans after the United States, and the total planted area reached 30,105 thousand hectares. Cotton (Gossypium hirsutum L.) is a dicotyledonous plant of Malvaceae family, also cultivated in large scale, its cultivation is an economically important activity for the country and reached around of 1,102.8 thousand hectares of planted area (IBGE, 2014). Therefore, cotton and soybean are two important crops in Brazil and, represent around 58% of the total cultivated area (CONAB, 2014) requiring a large amounts of chemicals fertilizers.
The large use of chemical fertilizers has a serious impact on the environment (Tilman et al., 2002) and the agricultural practices influence soil microorganisms greatly, decreasing soil fertility and organic matter turnover (Altieri, 1999). However, the more crucial issue for modern agriculture is that the natural reservoir of some nutrients as phosphorus (P) is decreasing in the world, leading to increase in fertilizer prices in the last decade (Cordell et al., 2009). The challenge for crop production is change to sustainable practices, by finding alternatives for increasing nutrient availability for plant nutrition as organic fertilization. Some these alternatives for organic fertilization include the use de soil microorganisms (Barrios, 2007; Miransari, 2011). Soil microorganisms play an important role by contributing significantly to nutrient availability through biochemical transformations. Some of these microorganisms act directly on plant nutrition by establishing symbiotic associations with plant roots (Bardgett, 2005). The symbiosis between arbuscular mycorrhizal fungi (AMF) and plant roots is one of the most known beneficial interactions occurring in soil (Smith and Smith, 2011), playing an important role in crop production and nutrient turnover (Andrade, 2004).
Arbuscular mycorrhizal fungi increase the uptake of soil inorganic nutrients, mainly P (Neumann and George, 2010). In addition, other benefits related to AMF are the stabilization of soil aggregates (Rillig, 2004), increased resistance to water stress (Garg and Chandel, 2010) and protection against pathogens (Jung et al., 2012). The use of biofertilizer is considered a good alternative to replace or reduce chemical fertilizer use. In example, other symbiotic microorganisms have been successfully used in soybean, and currently, Bradyrhizobium and other genera of symbiotic N-fixing bacteria are extensively used as biofertilizer in intensive soybean culture (Deaker et al., 2004) but not for AMF inocula.
In recent years, interest in AMF has focused on finding a viable method to optimize the production of AMF inoculum to use as inoculant in crop systems (Gianinazzi and Vosátka, 2004; Ijdo et al., 2011). The AMF inoculation in field conditions was been evaluated by some authors as Romero and Bago (2010), Pellegrino et al. (2011, 2012), and Ortas (2012) showing a high potential to increase crops yields. However, the success of AMF inoculation in agricultural soils can be determined by many factors such as species compatibility, habitat niche availability for AMF and competition with native fungi (Verbruggen et al., 2013), these aspects need to be evaluated under local conditions for a more appropriate assessment of the viability of AMF use as biofertilizer in crops.
The potential of colonization in soil of in vitro Rhizophagus clarus inoculum was first assessment in cotton and soybean in greenhouse conditions. No differences were found between R. clarus in vitro and pot culture inoculums for root colonization, plant biomass and P uptake. These results showed the successful of this AMF isolate in pure culture and the potential of this species for large-scale inoculum production (Cely, 2014).
The objective of this work was to determine the effectiveness of AMF (R. clarus) inoculation in two crops soybean (Glycine max L.) and cotton (Gossypium hirsutum L.), assessing its effect on plant growth, nutrient uptake and yield when compared with conventional chemical fertilization under field conditions. Our hypothesis is that AMF inoculation can be an alternative for total nutrient supply or more effective nutrient absorption, when combined with chemical fertilization.
The experiments were carried out in Londrina city – PR, Brazil (23°55′46″ S and 51°19′11″ W) during summer (November to June). The climate is humid subtropical, with rainfall during all seasons, relative humidity around 69% and about 2,000 mm of annual precipitation, and the average summer temperature is around 29.5°C.
Two experimental areas were used (A1 and A2) with a Rhodic Ferralsol soil type according FAO (1994). Soil chemical composition and the number of indigenous AMF were determined before sowing by wet sieving and decanting (Gerdemann and Nicolson, 1963) (Table 1).
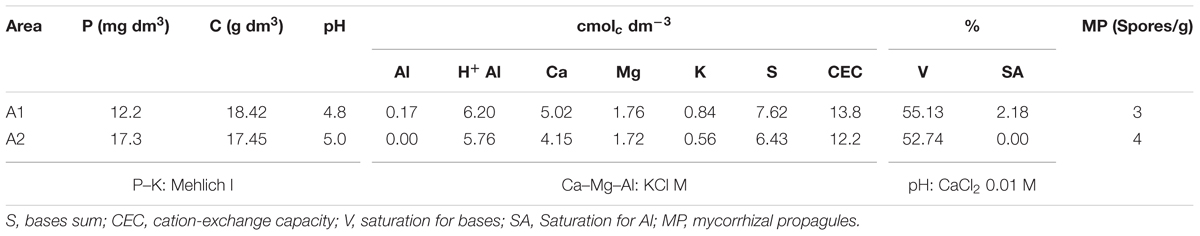
TABLE 1. Soil properties of the experimental areas.
The R. clarus inoculum was produced in vitro conditions. The monoxenic culture was obtained using carrot (Daucus carota L.) Ri T-DNA transformed roots as host organs (Supplementary Figure S1). The R. clarus cultures were maintained by continuous subculture of young colonized root fragments (every 4–5 weeks at 25°C, in the dark) in modified Strullu–Romand medium (MRS; Declerck et al., 1998). Petri dishes with massive growth (mycelia and spores) of R. clarus and colonized roots were used as crude inoculum. The inoculation methods consist in the seeds palletization with different propagules (colonized roots, hyphae fragments, and spores) from in vitro pure cultures of R. clarus helped by an organic matrix and turf. The procedure to obtain massive inoculum and seeds inoculation is described in the patent INPI BR 10 2014 017389 7 – July 15, 2014 (Andrade et al., 2014).
Two experiments were carried out with soybean, first in the harvest 2012/13 (E1), using a conventional soybean var. BRS 133 and the second in the harvest 2013/14 (E2) with a transgenic soybean var. BRS 359 RR. Both experiments were composed by the following treatments: Control (non-AMF inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 NPK 0:20:20); AMF (R. clarus inoculation plus 65 kg ha-1 KCl); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 NPK 0:20:20); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 NPK 0:20:20). The fertilizer dosage (200 kg ha-1 NPK 0:20:20) was according with agronomic recommendations and chemicals analyses of soil in experimental areas (Table 1). The nitrogen (N) supply in all treatments was a commercial inoculant (Rizo Plus® Rhizobacter) that contain two lines of Bradyrhizobium japonicum (SEMIA 5079 and SEMIA 5080) and its inoculation was according the manufacturer’s recommendation.
The treatments were arranged in a completely randomized block design with five replicates (Supplementary Figure S2). Each replicate consist in plots of 5 × 8 m (40 m2) with 10 rows with spacing 0.45 m and, a density of ten plants per linear meter (approx. 200,000 plants ha-1). The plots were separated by two lateral lines as edge.
Cotton experiment was carried out in the harvest 2013/14 (December–June) with the following treatments: Control (Non-AMF inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 PK 20:20 + 200 kg ha-1 urea); AMF (R. clarus inoculation plus 65 kg ha-1 KCl + 200 kg ha-1 urea); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 PK 20:20 + 200 kg ha-1 urea); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 PK 20:20 + 200 kg ha-1 urea). The cotton variety used was Bayer® FM 975WS and the fertilizer dosage (200 kg ha-1 PK 20:20 + 200 kg ha-1 urea) was according agronomic recommendation for experimental area based in chemical analyses of soil (Table 1). The treatments were arranged in a completely randomized block design with five replicates as described above for soybean experiments.
The effect of R. clarus inoculation in soybean and cotton experiments was assessed by the quantification of effective mycorrhizal colonization of roots and their effect in nutrient uptake (N and phosphorus), biomass production (shoot dry weight), and yield grain (soybean) and lint (cotton).
In soybean experiments, roots of 10 plants per plot were sampled randomly at 30 and 80 days after emergence (DAE) to evaluate the mycorrhizal colonization. In sampled plants at 80 DAE was made the evaluations of biomass and quantification of N and P in plant tissues for variety BRS 133. The percentage of mycorrhizal colonization was estimated by the grid-line method (Giovanetti and Mosse, 1980) after fresh roots were stained (Phillips and Hayman, 1970). N and P in shoot tissues were quantified according to Murphy and Riley (1962) and Sarruge and Haag (1974), respectively. For biomass quantification, plants were cut at the ground level; the total fresh shoot height was measured and shoot dry weight was determined after drying at 50°C for 72 h. For cotton, plants and roots were sampled at 120 DAE. Five plants per treatment of each plot were randomly collected, and evaluated for AMF colonization, fresh and dry shoot height, and N and P quantification.
Relative mycorrhizal dependency (RMD) was determined by the given below (Plenchette et al., 1983).
Soybean grains were harvested at 120 DAE. For yield estimation were sampled four linear meters (2 m2) in central area of each plot; after sampling the grains were cleaned, dried, and weighted. Cotton yield was estimated at 190 DAE by counting and collecting opened bolls in 20 plants in the central rows of each plot.
The statistical analyses of AMF root colonization were performed using the Friedman test at a significance level of p ≤ 0.05. Plant growth parameters, nutrient uptake and field were analyzed by analysis of variance (ANOVA) and the Tukey test (HSD) at a significance level of p ≤ 0.05. The analysis was carried with BioEstat 5.0 and STATISTICA 7.0 software.
The first evaluation (30 DAE) of AMF colonization for two soybean varieties (BRS 133 and BRS 359 RR) showed that R. clarus inoculation increased root colonization about 20% more than non-inoculated plants; although not statistically significant, this difference indicates that inoculation have a positive effect (Figures 1A and 2A). At 80 DAE the roots colonization showed higher values, around 70%, in inoculated plants with the addition of half dose of fertilizers (AMF + 1/2 Fertilizer), in this time these differences were statistically significant by Friedman test (p < 0,05) when compared with non-inoculated plants for two soybean varieties (Figures 1B and 2B). When analyze the AMF root colonization of two soybean varieties, is possible observing that the transgenic variety BRS 359 RR had a highest early colonization (around 50% at 30 DAE) that the conventional variety BRS 133 (around 30% at 30 DAE).
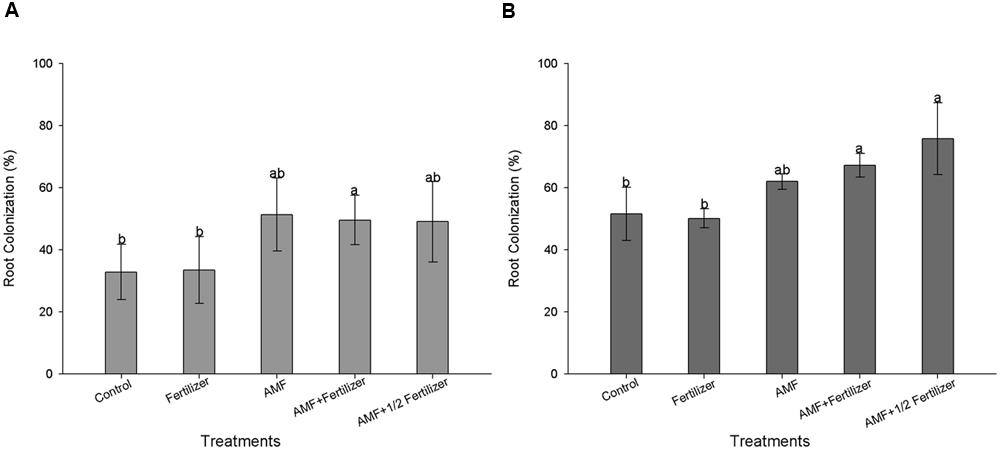
FIGURE 1. Root colonization of soybean variety BRS 133 at 30 days (A) and 80 days (B). Control (Non-arbuscular mycorrhizal fungi (AMF) inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 NPK 0:20:20); AMF (Rhizophagus clarus inoculation plus 65 kg ha-1 KCl); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 NPK 0:20:20); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 NPK 0:20:20). Columns followed by the same letter are not significantly different among treatments by Friedman test (n = 5) at p < 0.05. Bars represent standard error of means.
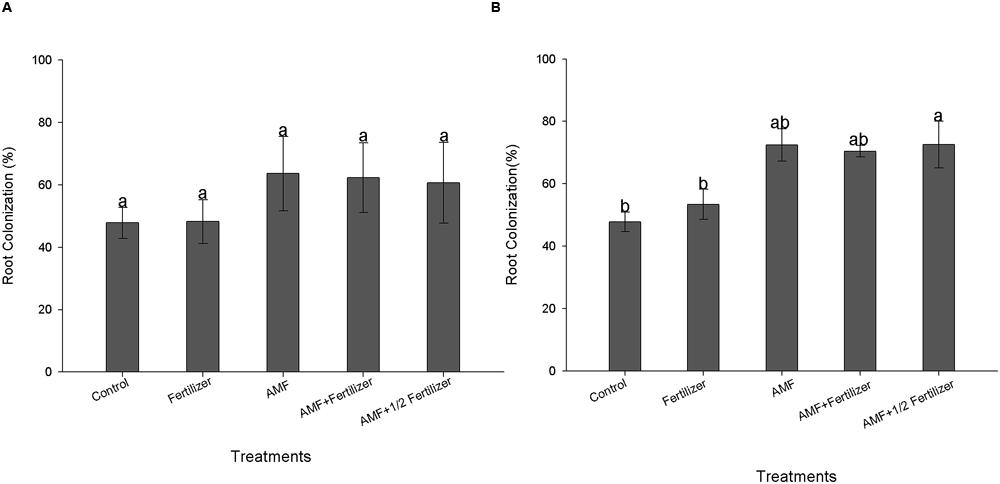
FIGURE 2. Root colonization of soybean variety BRS 359 RR at 30 days (A) and 80 days (B). Control (Non-AMF inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 NPK 0:20:20); AMF (R. clarus inoculation plus 65 kg ha-1 KCl); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 NPK 0:20:20); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 NPK 0:20:20). Columns followed by the same letter are not significantly different among treatments by Friedman test (n = 5) at p < 0.05. Bars represent standard error of means.
The response of soybean at R. clarus inoculation was assessment in variety BRS 133 at 80 DAE and are show in Table 2. No differences were observed in plant height between the control and fertilizer or inoculated treatments. Plant biomass and nutrients (N and P) uptake showed that R. clarus inoculation (AMF) had the same effect that the conventional fertilization (Fertilizer) and R. clarus inoculation with half dose of fertilizer (AMF + 1/2 Fertilizer). These treatments presented statistic differences regarding control (Control) according Tukey test (p < 0.05) but not among them. In the other hand, the highest values for these variables were observed in the treatment with R. clarus inoculation in combination with conventional fertilization (AMF + Fertilizer). This combination increased highly N and P uptake, around 24%, when compared with soybean only fertilized.
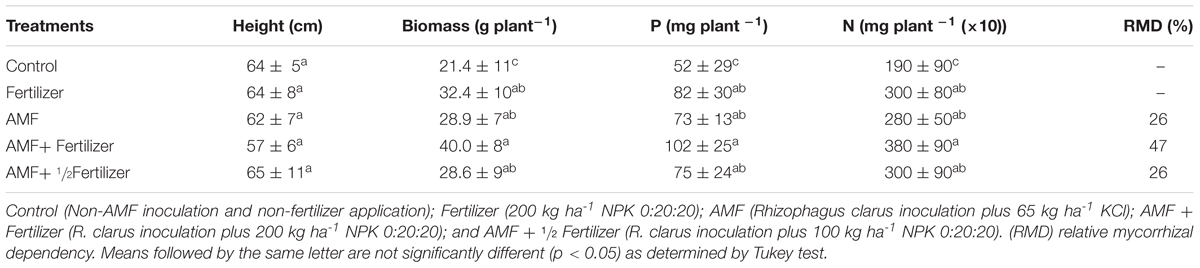
TABLE 2. Effect of AM inoculation on height, biomass and nutrients uptake in soybean plants at 80 DAE.
Reflecting the increase in nutrients uptake, R. clarus inoculum increased grain yield in cultivar BRS 133, the higher yield was observed in AMF+ Fertilizer treatment, the statistical analysis showed the yield could be equivalent between conventional fertilization, AMF and AMF + Fertilizer treatments (Figure 3A). Soybean BRS 359 showed the best grain yield in AMF + Fertilizer and AMF + 1/2Fertilizer treatments (Figure 3B). The effect of R. clarus inoculation showed high correlation between yield of soybean BRS 133 and P (r = 0.98; p = 0.01) and N (r = 0.96; p = 0.03) tissue contents (Figure 4).
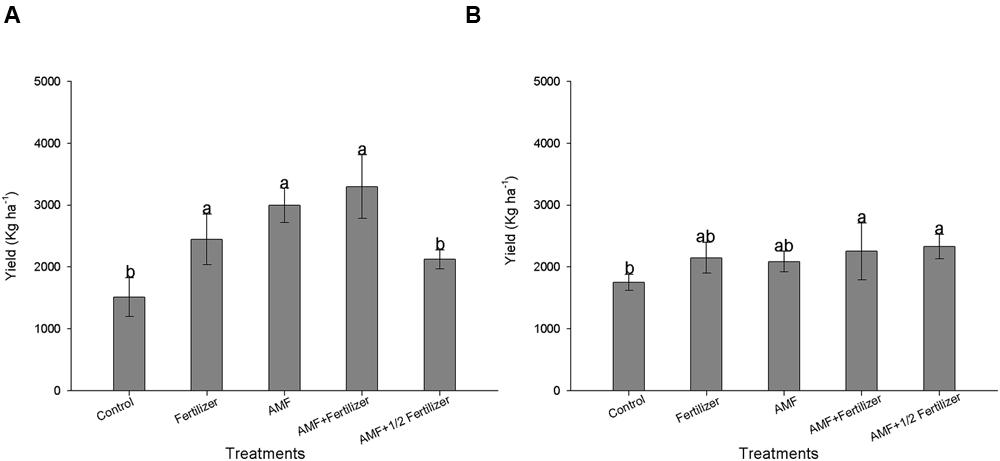
FIGURE 3. Effect of AMF inoculation on grain yield of soybean BRS 133 (A) and BRS 359 RR (B). Control (Non-AMF inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 NPK 0:20:20); AMF (R. clarus inoculation plus 65 kg ha-1 KCl); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 NPK 0:20:20); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 NPK 0:20:20). Columns followed by the same letter are not significantly different between treatments (p < 0.05) was determined by Tukey test. Bars represent standard error of means.
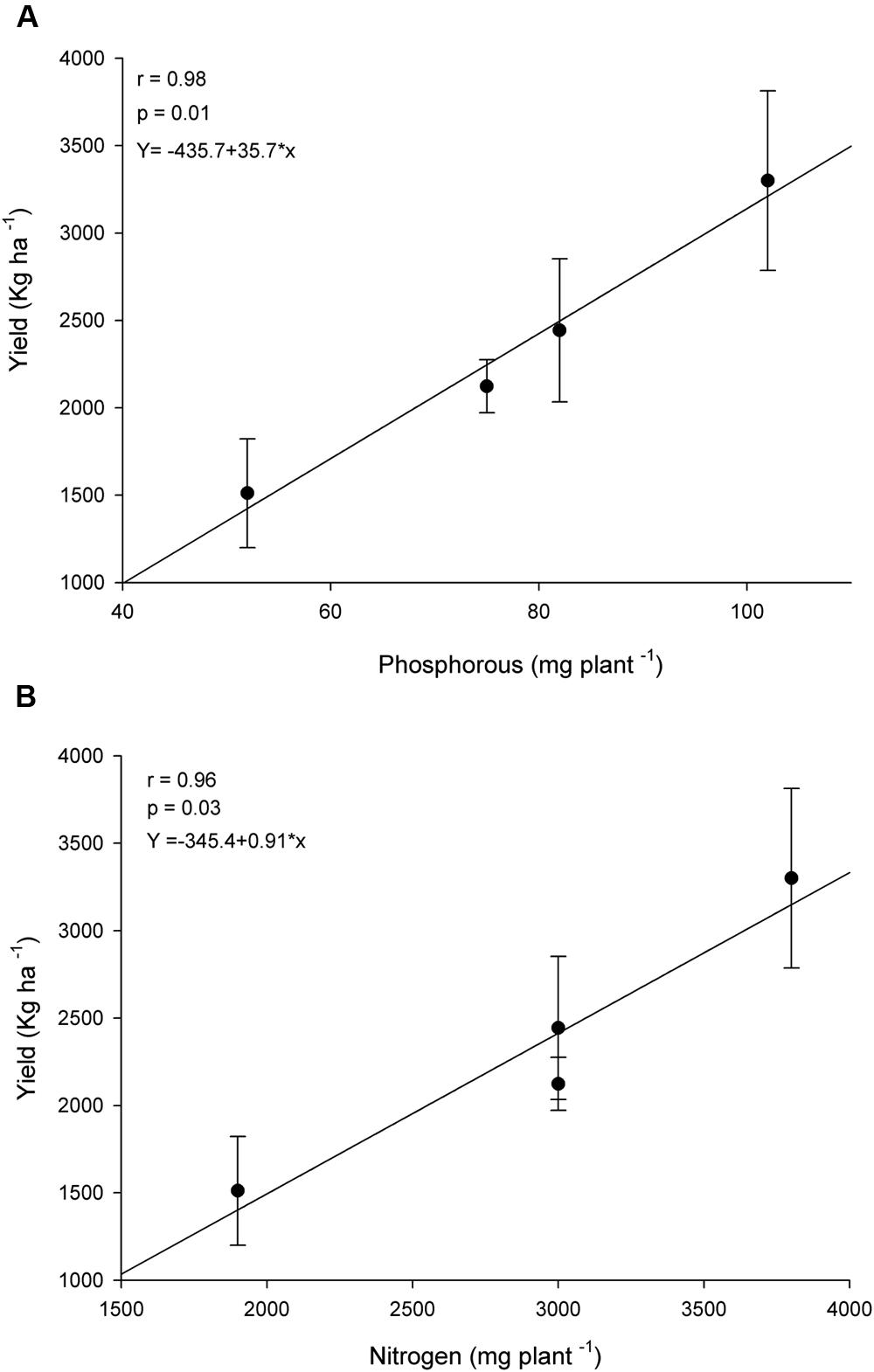
FIGURE 4. Correlation between shoot nutrients uptake and grain yield of soybean var. BRS133. (A) Phosphorous (P) uptake and grain yield. (B) Nitrogen (N) uptake and grain yield.
The R. clarus inoculation increased root colonization (80%) when compared with plants without inoculation (50%) at 120 DAE (Figure 5A) and, this difference in the colonization was statistically significant according Friedman test (p < 0.05), showing that just as soybean, the cotton inoculation with a R. clarus had a positive response.
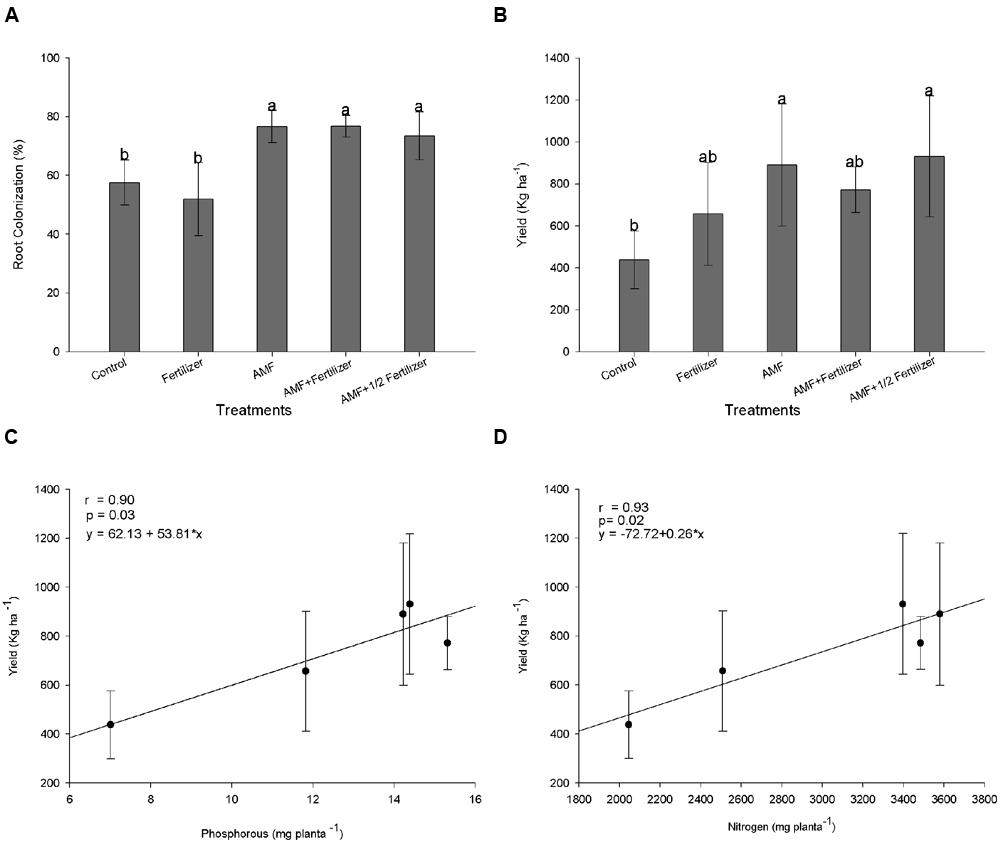
FIGURE 5. Effect of AMF inoculation in cotton. (A) Root colonization at 120 days after emergence (DAE); (B) lint cotton yield; (C) correlation between shoot P uptake and lint cotton yield; (D) correlation between shoot N uptake and lint cotton yield. Control (Non-AMF inoculation and non-fertilizer application); Fertilizer (200 kg ha-1 PK 20:20 + 200 kg ha-1 urea); AMF (R. clarus inoculation plus 65 kg ha-1 KCl + 200 kg ha-1 urea); AMF + Fertilizer (R. clarus inoculation plus 200 kg ha-1 PK 20:20 + 200 kg ha-1 urea); and AMF + 1/2 Fertilizer (R. clarus inoculation plus 100 kg ha-1 PK 20:20 + 200 kg ha-1 urea). Columns followed by the same letter are not significantly different (p < 0.05) between treatments by Friendman test (for root colonization) and Tukey test (for lint yield). Bars represent standard error of means for each treatment.
The AMF inoculation does not show a significant effect in plant height. Others parameters as plant biomass and nutrients uptake showed differences in control plants when compared with fertilizer and AMF + Fertilizer combinations. Statistical analysis of these parameters suggest that conventional fertilization in cotton have the same effect that only AMF inoculation, in other hand the fertilization in combination with AMF inoculation (AMF + Fertilizer and AMF + 1/2/Fertilizer) not differ among them (Table 3). Lint cotton yield was significantly higher in plans with AMF inoculation without fertilization and in treatment with AMF inoculation with half dose of fertilizer (Figure 5B). Nutrients uptake showing significantly high correlation with lint cotton yield, therefore for P uptake the correlation coefficient was r = 0.90 (Figure 5C), and for N uptake r = 0.96 (Figure 5D), both statistically significant (p < 0.05).

TABLE 3. Effect of R. clarus inoculation on total shoot height, biomass, P and N shoot uptake of cotton plants at 120 DAE.
The inoculation of R. clarus increased plant growth and yield of two varieties of soybean and cotton. Apparently, the inoculum produced in vitro was more competitive against native AMF, since inoculated plants showed increased AMF colonization and shoot uptake of P and N. Soybean and cotton showed different responses for R. clarus inoculum. First, in soybean, there was a triple interaction (Bradyrhizobium – R. clarus – plant root), and the inoculum tested was infective and effective, since symbiotic bacteria were already present.
The success of AMF inoculation in agricultural soils can be determined by many factors such as species compatibility, habitat niche availability for AMF and competition with native fungi (Verbruggen et al., 2013). Compatibility is an important point for AMF inoculation, where some isolates could be host “specialists,” while others “generalists” (Öpik and Moora, 2012). The inoculum of R. clarus tested showed a generalist nature, since it enhanced both plant growth and yield. Accordingly, AMF that are considered plant host generalists have a high establishment rate in several crops (Öpik and Moora, 2012); the results showed that soybean and cotton were effectively colonized, indicating a low specificity by the host plants for R. clarus.
In the experiments, mycorrhizal colonization in control plant was around 50% indicating that the agricultural soils support an active indigenous AMF community. The adaptation of R. clarus and its competition capacity against indigenous AMF were high. The problem in obtaining an effective AMF inoculum to use on large scale concerns these factors exactly; the inoculum showed good infectivity and high competition capacity under field conditions.
As well known, soil P availability is one the most important factors of AMF regulation, and this characteristic is directly related to the role of P uptake in the AMF symbiosis (Smith et al., 2003; Breuillin et al., 2010; Gutjahr and Parniske, 2013). Our results showed that in soybean and cotton, the moderate soil P availability in the experimental areas (12 and 17 mg dm-3) did not inhibit root colonization of the native AMF population and inoculum of R. clarus. The effectiveness of AMF inoculation in greenhouse experiments with phosphate fertilization showed that moderate phosphate availability can allow mycorrhizal colonization, promoting plant growth (Schroeder and Janos, 2005; Taffouo et al., 2013; Xie et al., 2014), and the same responses were found in a field conditions in soybean (Maddox and Soileau, 1991; Karaca et al., 2013).
On other hand, soil P availability can be determined by soil chemical characteristics that influence phosphate solubility. In acid soils, P is less available because of immobilization, even with fertilizer applications, making it unavailable to plants (Busman et al., 2002). Rhodic Ferralsol soils in the experimental area showed low pH, where they can adsorb phosphate, and AMF has an important role in enhancing P uptake and availability, including P from chemical fertilization.
Plants with high P requirements show a high RMD index (Plenchette et al., 1983). Cotton showed a higher RMD (45%) than did soybean (26%) when inoculated with R. clarus in the presence or absence of fertilizer. In contrast, when P was added at the recommended dose in combination with AMF inoculation, this index decreased to 41% in cotton and increased to 47% in soybean, suggesting that the gain in biomass was related to the availability of P from the fertilizer, which R. clarus provided for the plant roots. Thompson et al. (2012) obtained the same results.
The finding that P and N uptake increased in both crops may be related to R. clarus association as observed by other authors (Allen et al., 2003; Barea et al., 2005). AMF improved plant nutrition, leading to an increase in grain yield in soybean and cotton lint production, showing a positive correlation between plant nutrition and yield. Mahanta et al. (2014) also observed a positive linear relationship between P and yield in soybean when inoculated with AMF. The effect of R. clarus on cotton growth and yield found here agrees with Thompson et al. (2012) who found an increase in seed cotton yield with Glomus mosseae inoculation. The effect of AMF inoculation. However, this is the first time that R. clarus inoculum obtained under axenic conditions was tested under field conditions. Ceballos et al. (2013) showed that inoculation of Rhizophagus irregularis increased the cassava yield in field and suggest this practice as alternative for improve this crop in several countries.
The inoculum of R. clarus evaluated was very competitive against endogenous AMF and also increased plant growth and yield. R. clarus obtained in vitro and tested in the field was efficient in starting early AMF infection in seedlings, improving AM colonization in soybean and cotton. The inoculum of R. clarus helped plants to take up P from fertilizer and showed high potential for use in combination with conventional fertilization, for intensive agriculture system in large areas in tropical soils, increasing P absorption and more efficient fertilization use, this is fundamental for the actual challenge of crops production.
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The National Council of Scientific and Technological Development (CNPq) who made this study possible by awarding PIBIC, MSc., Ph.D. and Productivity in Research grants.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Dr. A. Leyva helped with English editing of the manuscript.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00720
Allen, M. F., Swenson, W., Querejeta, J. I., Egerton-Warburton, L. M., and Treseder, K. K. (2003). Ecology of mychorrhizae: a conceptual framework for complex interactions among plants and fungi. Annu. Rev. Phytopathol. 41, 271–303. doi: 10.1146/annurev.phyto.41.052002.095518
Altieri, M. A. (1999). The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 74, 19–31. doi: 10.1016/S0167-8809(99)00028-6
Andrade, G. (2004). “Role of functional groups of microorganisms on the rhizosphere microcosm dynamics,” in Plant Surface Microbiology, eds A. Varma, L. Abbott, D. Werne, and R. Hampp (New York, NY: Springer),k51–68.
Andrade, G., de Oliveira, A. G., and Cely, M. V. T. (2014). Processo de Produção e Inoculação de Fungos Micorrízicos Arbusculares. Brazil Patent No BR1020140173897. Londrina, PR: Instituto Nacional de Propriedade Intelectual.
Bardgett, R. D. (2005). “Organism interactions and soil processes,” in The Biology of Soil a Community and Ecosystem Approach, ed. R. D. Bardgett (New York: Oxford University Press), 57–85.
Barea, J. M., Azcón, R., and Aguilar, A. C. (2005). “Interactions between mycorrhizal fungi and bacteria to improve plant nutrient cycling and soil structure,” in Soil Biology, eds F. Buscot and A. Varma (New York, NY: Springer), 195–208.
Barrios, E. (2007). Soil biota, ecosystem services and land productivity. Ecol. Econom. 64, 269–285. doi: 10.1111/1462-2920.12559
Breuillin, F., Schramm, J., Hajirezaei, M., Ahkami, A., Favre, P., Druege, U., et al. (2010). Phosphate systemically inhibits development of arbuscular mycorrhiza in Petunia hybrid and represses genes involved in mycorrhizal functioning. Plant J. 64, 1002–1017. doi: 10.1111/j.1365-313X.2010.04385.x
Busman, L., Lamb, J., Randall, G., Rehm, G., Schmitt, M. (2002). Data From: The Nature of Phosphorus in Soils. Saint Paul, MN: Regents of the University of Minnesota.
Ceballos, I., Ruiz, M., Fernández, C., Peña, R., Rodríguez, A., and Sanders, I. R. (2013). The in vitro mass-produced model mycorrhizal fungus, Rhizophagus irregularis, significantly increases yields of the globally important food security crop cassava. PLoS ONE 8:e70633. doi: 10.1371/journal.pone.0070633
Cely, M. V. T. (2014). In vitro Rhizophagus Clarus Inoculum Production and Its Application in Agricultural Systems. Ph.D. thesis, Universidade Estadual de Londrina-Brazil, Londrina PR.
CONAB (2014). Data from: Companhia Nacional de Abastecimento, Acompanhamento da Safra Brasileira: Grãos, Safra 2013/14. Available at: http://www.conab.gov.br/OlalaCMS/uploads/arquivos/14_09_10_14_35_09_boletim_graos_setembro_2014.pdf
Cordell, D., Drangert, J., and White, S. (2009). The story of phosphorus: global food security and food for thought. Global Environ. Change 19, 292–305. doi: 10.1016/j.gloenvcha.2008.10.009
Deaker, R., Roughley, R. J., and Kennedy, I. R. (2004). Legume seed inoculation technology: a review. Soil Biol. Biochem. 36, 1275–1288. doi: 10.1016/j.ram.2015.06.006
Declerck, S., Strullu, D. G., and Plenchette, C. (1998). Monoxenic culture of the intraradical forms of Glomus sp. isolated from a tropical ecosystem: a proposed methodology for germplasm collection. Mycologia 90, 579–585. doi: 10.2307/3761216
Garg, N., and Chandel, S. (2010). Arbuscular mycorrhizal networks: process and functions, a review. Agron. Sustain. Dev. 30, 581–599. doi: 10.1051/agro/2009054
Gerdemann, J. W. T., and Nicolson, H. (1963). Spores of mycorrhizal endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 46, 235–244. doi: 10.1016/S0007-1536(63)80079-0
Gianinazzi, S., and Vosátka, M. (2004). Inoculum of arbuscular mycorrhizal fungi for production systems: science meets business. Can. J. Bot. 82, 1264–1271. doi: 10.1139/b04-072
Giovanetti, M., and Mosse, B. (1980). Evaluation of techniques for measuring vesicular-arbuscular micorrhizal infections in roots. New Phytol. 84, 489–500. doi: 10.1111/j.1469-8137.1980.tb04556.x
Gutjahr, C., and Parniske, M. (2013). Cell and developmental biology of arbuscular mycorrhiza symbiosis. Annu. Rev. Cell. Biol. 29, 593–617. doi: 10.1146/annurev-cellbio-101512-122413
IBGE (2014). Data from: Instituto Brasileiro de Geografia e Estatística, Levantamento Sistemático da Produção Agrícola. Available at: http://www.ibge.gov.br/home/estatistica/indicadores/agropecuaria/lspa/default.shtm
Ijdo, M., Cranenbrouck, S., and Declerck, S. (2011). Methods for large-scale production of AM fungi: past, present, and future. Mycorrhiza 21, 1–16. doi: 10.1007/s00572-010-0337-z
Jung, S. C., Martinez-Medina, A., Lopez-Raez, J. Á., and Pozo, M. J. (2012). Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 38, 651–664.
Karaca, H., Uygur, V., Özkan, A., and Kaya, Z. (2013). Effects of mycorrhizae and fertilization on soybean yield and nutrient uptake. Commun. Soil Sci. Plant 44, 2459–2471. doi: 10.1080/00103624.2013.809730
Maddox, J. J., and Soileau, J. M. (1991). Effects of phosphate fertilization, lime amendments and inoculation with VA-mycorrhizal fungi on soybeans in an acid soil. Plant. Soil 134, 83–93.
Mahanta, D., Rai, R. K., Mishra, S. D., Raja, A., Purakayastha, T. J., and Varghese, E. (2014). Influence of phosphorus and biofertilizers on soybean and wheat root growth and properties. Field Crops. Res. 166, 1–19. doi: 10.1016/j.fcr.2014.06.016
Miransari, M. (2011). Soil microbes and plant fertilization. Appl. Microbiol. Biotechnol. 92, 875–885. doi: 10.1007/s00253-011-3521-y
Murphy, J., and Riley, J. P. A. (1962). Modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 27, 31–36. doi: 10.1016/S0003-2670(00)88444-5
Neumann, E., and George, E. (2010). “Nutrient uptake: the arbuscular mycorrhiza fungal symbiosis as a plant nutrient acquisition strategy,” in Arbuscular Mycorrhizas: Physiology and Function, eds H. Koltai and Y. Kapulnik (New York, NY: Springer), 137–167.
Öpik, M., and Moora, M. (2012). Missing nodes and links in mycorrhizal networks. New Phytol. 194, 304–306. doi: 10.1111/j.1469-8137.2012.04121.x
Ortas, I. (2012). The effect of mycorrhizal fungal inoculation on plant yield, nutrient uptake and inoculation effectiveness under long-term field conditions. Field Crop. Res. 125, 35–48. doi: 10.1016/j.fcr.2011.08.005
Pellegrino, E., Bedini, S., Avio, L., Bonari, E., and Giovannetti, M. (2011). Field inoculation effectiveness of native and exotic arbuscular mycorrhizal fungi in a Mediterranean agricultural soil. Soil Biol. Biochem. 43, 367–376. doi: 10.1007/s00572-014-0600-9
Pellegrino, E., Turrini, A., Gamper, H. A., Cafa, G., Bonari, E., Young, J. P. W., et al. (2012). Establishment, persistence and effectiveness of arbuscular mycorrhizal fungal inoculants in the field revealed using molecular genetic tracing and measurement of yield components. New Phytol. 94, 810–822.
Phillips, J., and Hayman, D. (1970). Improved producers for clearing roots and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55, 158–160. doi: 10.1016/S0007-1536(70)80110-3
Plenchette, C., Fortin, J. A., and Furlan, V. (1983). Growth responses of several plant species to mycorrhizae in a soil of moderate P-fertility: mycorrhizal dependency under field conditions. Plant Soil 70, 199–209. doi: 10.1007/BF02374780
Rillig, M. C. (2004). Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil. Sci. 84, 355–363. doi: 10.4141/S04-003
Romero, C. C., and Bago, A. (2010). Inoculante Aséptico de Micorrización y Procedimientos de Aplicación en Condiciones In Vitro y Ex Vitro. Patent WO 2007/014974
Sarruge, J. R., and Haag, H. P. (1974). Analises Químicas em Plantas. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz.
Schroeder, M. S., and Janos, D. P. (2005). Plant growth, phosphorus nutrition, and root morphological responses to arbuscular mycorrhizas, phosphorus fertilization, and intraspecific density. Mycorrhiza 15, 203–221. doi: 10.1007/s00572-004-0324-3
Smith, S. E., and Smith, F. A. (2011). Mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales. Annu. Rev. Plant. Biol. 62, 227–250. doi: 10.1146/annurev-arplant-042110-103846
Smith, S. E., Smith, F. A., and Jakobsen, I. (2003). Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 133, 16–20. doi: 10.1104/pp.103.024380
Taffouo, V. D., Ngwene, B., Akoa, A., and Franken, P. (2013). Influence of phosphorus application and arbuscular mycorrhizal inoculation on growth, foliar nitrogen mobilization, and phosphorus partitioning in cowpea plants. Mycorrhiza 24, 361–368. doi: 10.1007/s00572-013-0544-5
Thompson, J. P., Seymour, N. P., and Clewett, T. G. (2012). Stunted cotton (Gossypium hirsutum L.) fully recovers biomass and yield of seed cotton after delayed root inoculation with spores of an arbuscular mycorrhizal fungus (Glomus mosseae). Australas. Plant Pathol. 41, 431–437. doi: 10.1007/s13313-012-0137-3
Tilman, D., Cassman, K. G., Matson, P. A., Naylor, R., and Polasky, S. (2002). Agricultural sustainability and intensive production practices. Nature 418, 671–677. doi: 10.1038/nature01014
Verbruggen, E., van der Heijden, M. G. A., Rillig, M. C., and Kiers, T. (2013). Mycorrhizal fungal establishment in agricultural soils: factors determining inoculation success. New Phytol. 197, 1104–1109. doi: 10.1111/j.1469-8137.2012.04348.x
Keywords: nutrient uptake, seed inoculation, rhizosphere, Glycine max L., Gossypium hirsutum L.
Citation: Cely MVT, de Oliveira AG, de Freitas VF, de Luca MB, Barazetti AR, dos Santos IMO, Gionco B, Garcia GV, Prete CEC and Andrade G (2016) Inoculant of Arbuscular Mycorrhizal Fungi (Rhizophagus clarus) Increase Yield of Soybean and Cotton under Field Conditions. Front. Microbiol. 7:720. doi: 10.3389/fmicb.2016.00720
Received: 13 March 2016; Accepted: 29 April 2016;
Published: 25 May 2016.
Edited by:
Mohamed Hijri, Université de Montréal, CanadaReviewed by:
Erik Limpens, Wageningen University, NetherlandsCopyright © 2016 Cely, de Oliveira, de Freitas, de Luca, Barazetti, dos Santos, Gionco, Garcia, Prete and Andrade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Galdino Andrade, YW5kcmFkZWdAdWVsLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.