
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 05 February 2016
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.00091
This article is part of the Research TopicNew frontiers in the search of antimicrobials agents from natural productsView all 54 articles
 Ariane F. Lacerda1,2
Ariane F. Lacerda1,2 Patrícia B. Pelegrini1*
Patrícia B. Pelegrini1* Daiane M. de Oliveira1,3
Daiane M. de Oliveira1,3 Érico A. R. Vasconcelos1,4
Érico A. R. Vasconcelos1,4 Maria F. Grossi-de-Sá1,2,3,5
Maria F. Grossi-de-Sá1,2,3,5Africa, Asia, and Latin America are regions highly affected by endemic diseases, such as Leishmaniasis, Malaria, and Chagas’ disease. They are responsible for the death of 1000s of patients every year, as there is not yet a cure for them and the drugs used are inefficient against the pathogenic parasites. During the life cycle of some parasitic protozoa, insects become the most important host and disseminator of the diseases triggered by these microorganisms. As infected insects do not develop nocive symptoms, they can carry the parasites for long time inside their body, enabling their multiplication and life cycle completion. Eventually, parasites infect human beings after insect’s transmission through their saliva and/or feces. Hence, host insects and general arthropods, which developed a way to coexist with such parasites, are a promising source for the prospection of anti-parasitic compounds, as alternative methods for the treatment of protozoa-related diseases. Among the molecules already isolated and investigated, there are proteins and peptides with high activity against parasites, able to inhibit parasite activity in different stages of development. Although, studies are still taking their first steps, initial results show new perspectives on the treatment of parasitic diseases. Therefore, in this report, we describe about peptides from host insect sources with activity against the three most endemic parasites: Leishmania sp., Plasmodium sp., and Trypanosomes. Moreover, we discuss the future application insect peptides as anti-parasitic drugs and the use of non-hosts insect transcriptomes on the prospection of novel molecules for the treatment of parasitic neglected diseases.
In 1970, the Rockefeller Foundation introduced the term “the Great Neglected Diseases,” corresponding to several illnesses caused by infectious microorganisms and parasites (viruses, bacteria, fungi, protozoa, and helminthes), endemic in poor populations of Asia, Africa, and America. Neglected diseases received this name due to the lack of interest by health science researchers and pharmaceutical companies on developing and commercializing products for their treatment or cure. Consequently, studies on this field are poorly invested (Souza, 2010).
However, during the last 20 years, the pattern of neglected diseases worldwide has changed, as mortality rates are decreasing, while morbidity still grows on the population. All neglected diseases are closely related to poverty, lack of hygiene, food and quality of life. Therefore, it is difficult to include long-time treatment programs, as most of the patients have no financial conditions to pay for the drugs (Paes and Silva, 1999; Brasil, 2010).
Chagas diseases, Leishmaniasis, and Malaria are some of these neglected diseases. Chagas disease, first reported in America during 1909, was classified as an enzootic illness affecting mostly the poor population of Latin America. Nevertheless, nowadays, it became a worldwide concern, as it spread through many countries in Africa, Europe, and Asia (Coura and Viñas, 2010; World Health Organization [WHO], 2014, 2015a). Migration processes of contaminated people to non-endemic regions and environmental changes in forestry areas, favoring trypanosome parasites to adapt into domestic environment, increased the identification of Chagas disease cases in North America, Western Pacific region and at least 16 countries of Europe. Therefore, new forms of parasite transmission were described, including blood transfusion, organ transplantation, and vertical transmission from mother to child (Coura and Viñas, 2010; World Health Organization [WHO], 2014). The constant migration of people from different countries during the last two decades enlarged epidemiological and social matters, leading to an evaluation of economic and political aspects for the prevention, treatment, and eradication of this disease (World Health Organization [WHO], 2015a).
Leishmaniasis is another neglected disease with high concern to many countries worldwide. In February, the World Health Organization [WHO] (2015b) estimated 1.3 million new cases of Leishmaniasis (visceral and cutaneous) and 20,000–30,000 deaths annually. The cutaneous form of Leishmaniasis occurs in different populations of the world, although higher rates are observed in South American and African countries. On the other hand, 90% of all cases of visceral Leishmaniasis is concentrated in Bangladesh, Brazil, Ethiopia, India, Sudan, and South Sudan (Alvar et al., 2012). In Brazil, Leishmaniasis is showing an expansion, generally associated with environmental modification caused by man, population displacements arising from endemic areas and insufficient infrastructure in water and sewer systems (Paes and Silva, 1999; Brasil, 2010).
Additionally, Malaria is one of the most important parasitic infectious disease of the world. It occurs in tropical and subtropical areas of 106 countries worldwide. Only in 2012, 627,000 1000 people in the Saharan Africa died of Malaria (Center for Disease Control and Prevention [CDC], 2014). Although an estimative from the World Malaria Report consider a significant decrease in the number of countries exposed to Malaria (from 140 to 106), every year, 100s of millions of new episodes are registered in different regions of the world (Public Health England, 2014; World Malaria Report, 2015).
Most of neglected diseases have low financial support, both from Private and Governmental funds. However, great amount of international funding is given to control new cases of Malaria in endemic countries, as well as applied on preventive Programs for poor populations. The amount invested on the control of Malaria worldwide raised from US$ 960 million in 2005 to US$ 2.5 billion in 2014 (World Malaria Report, 2015). The number increased 8% between 2013 and 2014, but the financial support invested on the prevention and treatment of Malaria are still not sufficient to contain the arrival of new cases.
Hence, part of the financial investments applied on Malaria, Leishmaniasis, and Chagas’ disease is spent on research for the discovery of novel molecules that can be used as new pharmaceutical products for the treatment of these illnesses. Recently, studies demonstrated that the use of peptides isolated from host mosquitoes and other invertebrates, called anti-parasitic peptides (APPs), are the key to inhibit or even kill protozoa cells infecting the human body (Li et al., 2006; Moreira et al., 2007; Tian et al., 2008; Fieck et al., 2010; Gao et al., 2010; Rangel et al., 2011; Marr et al., 2012).
Therefore, this report describes the first studies related to the activity of different peptides from insect hosts APPs against three parasite species: Trypanosoma sp., Leishmania sp., and Plasmodium sp. The results reported here show the great potential of using insect host proteins and peptides to treat Chagas disease, Leishmaniasis, and Malaria in human patients.
Chagas disease is a human trypanosomiasis endemic in Latin America countries, caused by the species Trypanosoma cruzi. The name was given by the scientist Carlos Chagas, in honor of the research developed by Oswaldo Cruz, during the beginning of the 20th century. The transmission occurs when a person is bitten by a triatomine insect containing T. cruzi parasites (Don and Chatelain, 2008). Common triatomines of Chagas disease vectors belong to Triatoma, Rhodnius, and Panstrongylus generi. In Central and South America, the most common triatomine is T. prolixus (Fieck et al., 2010).
Chagas’ disease can be classified into three different stages: acute, asymptomatic, and chronic. The acute stage consists of fever, facial edema, generalized lymphadenopathy, and hepatosplenomegaly. Usually, 5% of contaminated children die during this stage, but the illness can spontaneously undertake itself in 4–6 weeks, becoming asymptomatic. During this stage, also called indeterminate, the parasite passes to its second phase, which can last from 10 years to a lifetime. The last and most concerning stage is the chronic phase, once the disease can compromise the heart, leading to death by cardiac arrhythmia or congestive heart failure (Whitebread et al., 2005; Don and Chatelain, 2008). Therefore, Chagas disease has become one of the main causes of heart problems in Latin America countries, as well as gastrointestinal mega-syndromes in some patients (Chagas, 1981; Parada et al., 1997).
It is estimated that 11 million people in American countries are infected with T. cruzi, and 10–30% of them will develop the chronic stage of Chagas disease (Bern et al., 2007). There is no cure for Chagas disease and the available treatments correspond to the use of benznidazole or nifurtimox. Both are effective only in 50% of the cases, and for patients in the acute or early indeterminate stages. There is still no treatment, nor even a cure, for the late indeterminate and chronic stages of Chagas disease (Kennedy, 1997; Whitebread et al., 2005; Frearson et al., 2007).
Until now, there are also no vaccines developed to prevent infection by T. cruzi, neither effective vector eradication Programs to control parasite dissemination (Kevin, 2014). Thus, there is a strong need for the development of adequate therapeutic strategies to treat Chagas disease. Preliminary work described the use of peptides capable of lysing different cell types (Kehoe and Timmis, 1984). Therefore, the utilization of proteins and peptides from different sources became a strategic alternative for the control of parasites.
Another interesting feature is the inability of parasites to damage the cells of their insect hosts. Although, the infection with the parasite can cause several damages in human organs, it is harmless to triatomine insects (Rosendaal, 1997). In this way, investigating proteins and peptides from parasite’s insect hosts is a prospering alternative for the development of new tools against Chagas disease.
There are still, in literature, very few studies on the prospection and evaluation of the anti-trypanosomal effect of proteins and peptides from insect sources, especially for species causing Chagas disease. Among all classes of proteic molecules, only three have been studied for this purpose: apidaecins, cecropins, and melittins (Table 1).
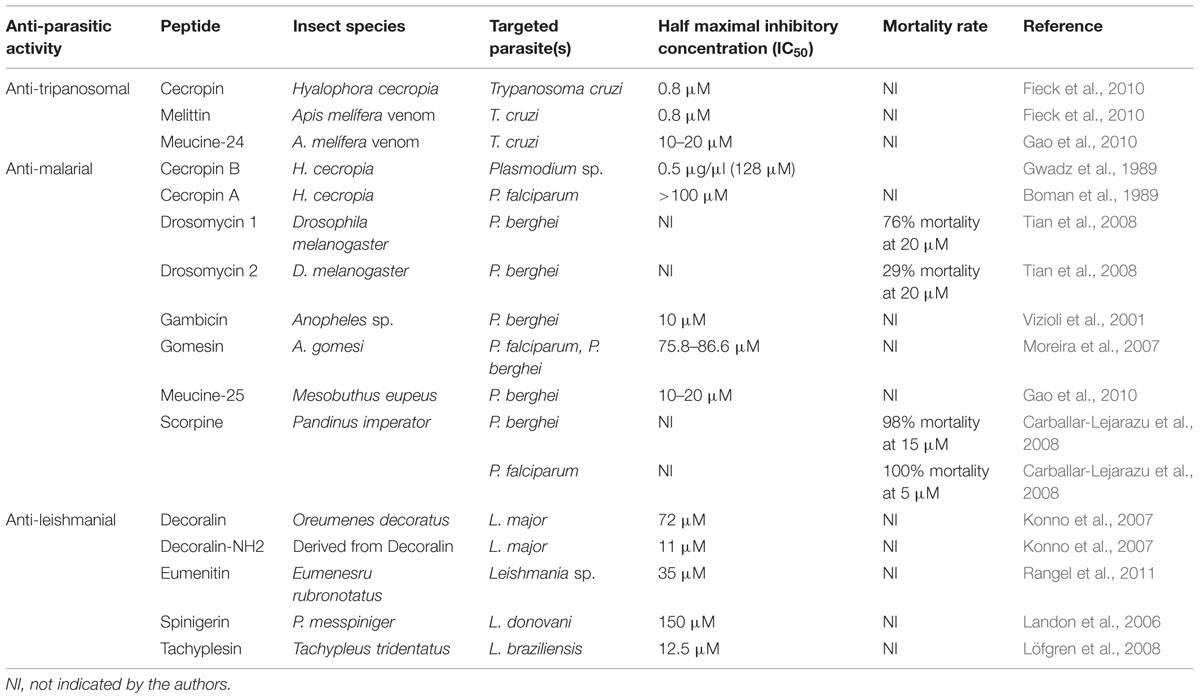
TABLE 1. Insect peptides with anti-parasitic activity.
Apidaecins correspond to a group of small prolin-rich peptides, ranging from 18 to 20 amino acid residues, previously isolated from the honeybees Apis melifera (Casteels et al., 1989; Li et al., 2006). They have been widely studied as potential molecules for therapeutic use due to their anti-bacterial activity and harmless impact against human and animal cells (Casteels and Tempst, 1994; Li et al., 2006). Recent studies showed that this class of peptides can also present anti-trypanocidal activity at an absolute lethal concentration (LC100) of 199 μM (Fieck et al., 2010). Moreover, when used in pairwise treatments, together with magainin II (from the Xenopus laevis), cecropin (from the silk worm) and melittin (also from honeybee), the anti-trypanocidal activity increased 10 folds, showing additive effects at a half maximal inhibitory concentration (IC50) of 0.70 μM (Fieck et al., 2010).
Cecropins include a class of small and basic peptides of about 31–37 amino acid residues. They were first isolated from the silk moth Hyalophora cecropia, and have been described as an important molecule in the cell-free immunity of insects, with activity against bacteria and fungi growth (Boman, 1991; Boman et al., 1991). Earlier studies demonstrated the performance of synthetic cecropins against T. cruzi. The synthetic peptides, called SB-37 and Shiva-1, with punctual mutations in their amino acid sequence, were able to kill trypomastigote forms of the parasite after 1-h exposure, in a dose-responsive manner (Jaynes et al., 1988). Interestingly, Shiva-1 peptide was 10 times more effective against Trypomastigotes than SB-37, in terms of parasite damage and lysis.
Another report showed that a Cecropin A displayed trypanostatic effects against Chagas diseases parasite, inhibiting its growth at a LC100 of 80 μM, although it was not able to kill T. Cruzi cells (Fieck et al., 2010). However, when coupled with other peptides, such as magainin, an antagonistic effect was observed, at a concentration of 0.1 and 1.0 μM for cecropin and magainin, respectively (Fieck et al., 2010).
Furthermore, melittin is a small 26-amino acid peptide isolated from honeybee venom with anti-microbial activity against yeasts and bacteria (Lubke and Garon, 1997; Klotz et al., 2004; Lazarev et al., 2005). It also has showed ability to inhibit protein kinase C, protein kinase II, and myosin kinase (Yang and Carrasquer, 1997). Previous reports described the potential use of melittin on the treatment of epilepsy, HIV infection, and cancer (Loftus, 2009; Hood et al., 2013; Verma et al., 2013). Further studies demonstrated that melittin present inhibitory activity against trypomastigotes (LD100 = 30 μM). Also, when coupled with apidaecin, the parasite inhibitory activity was increased. Nevertheless, when melittin was coupled with cecropin, it showed an antagonistic effect, contrary to the additive effect demonstrated with apidaecin (Fieck et al., 2010).
Leishmania is a genus of protozoa transmitted between mammals by blood-sucking sandflies. Mammal species, including dogs, mice and humans, are natural hosts for Leishmania and in some parts of the world, like India, humans are the main hosts (Kaye and Scott, 2011). The parasite’s live cycle encompasses a differentiation of procyclic promastigote form into infective metacyclic promastigote. This stage occurs inside sandflies, where the parasite will be ready for transmission at the stomodea valve’s insect. During blood-sucking, contaminated sandflies can inject metacyclic promastigotes into mammal tissues together with some parasite immunomodulatory elements. After phagocytized by host cells, parasites will accommodate inside human body. Then metacyclic promastigotes can become flagellate amastigote and replicates into host cells driving them to rupture, spreading amastigotes into host tissues and allowing infection of others phagocytes. The cycle is completed when infected phagocytes are sucked by another sand-fly and are converted in promastigotes into sand-fly’s midgut, re-starting a new cycle (Kaye and Scott, 2011).
There are more than 20 species of Leishmania related to several subtypes of chronic skin and viscera infections, classified as mucocutaneous disease, visceral disease, Leishmaniasis recidivism, and post-kala-azar dermal Leishmaniasis (McGwire and Satoskar, 2014). Around 1.3 million cases of Leishmaniasis are reported annually worldwide and there is an estimative that 350 million people in 88 countries are living at risk of developing Leishmaniasis (Kedzierski, 2010; World Health Organization [WHO], 2015b).
There is a huge number of drug treatment available to each form of Leishmaniasis. Pentavalent antimony is the agent of choice for the majority forms of such disease (Baiocco et al., 2009). Amphotericin B, Paromomycin, Pentamidine also have been used worldwide (Jaureguiberry et al., 2006; Chattopadhyay and Jafurulla, 2011; Sundar et al., 2011). However, these drugs are often harmful to the patient, the treatments need constant follow up and patients may not be 100% recovered (McGwire and Satoskar, 2014). Therefore, new strategies to treat Leishmaniasis is a clear need for millions of patients around the world.
Some APPs were reported as acting against Leishmania species and have been considered a low harmful alternative, despite of current drugs used to control Leishmania infections (Chadbourne et al., 2011). Marr et al. (2012) listed some leishmanicidal peptides, such as Temporins, Bombins, Magainins, and Cathelicidins (from amphibians and mammalians), and discussed their mechanism of action. Arthropods Leishmanicidal peptides are described in Table 1.
Hence, among the APPs from insect sources, there is Spinigerin. It is a cysteine-free peptide constitutively expressed by termite Pseudacanthotermes spiniger. Its minimal inhibitory concentration (MIC) in bioassays against filamentous fungi, yeast, gram-negative bacteria is <5 μM (Landon et al., 2006). When assayed against Leishmania donovani, spinigerin was able to cause apoptosis-like cell death (IC50, 150 μM). Landon et al. (2006) suggested that such peptide could cause such effects by inhibiting some Leshimanial trypanothione reductase, thus, arresting detoxification processes against reactive oxygen species (ROS) produced by the host cells.
Eumenitin is a class of small cationic peptides (15 amino acids) from the venom of the solitary eumenine Eumenes rubronotatus, which was isolated and characterized by Konno et al. (2006). Its activity against Gram-positive and Gram-negative bacteria, as well as its capacity to stimulate degranulation of mast cells are known since its discovery. However, eumenitin’s leishimanicidal activity was reported only in Rangel et al. (2011), when the peptide was assayed against Leishmania major, displaying anti-parasitic effects at IC50 of 35 μM.
Decoralin belongs to a class of small and linear cationic α-helical peptides not stabilized by disulfide bonds. It was isolated for the first time in Konno et al. (2007) from the venom of the solitary eumenine wasp Oreumenes decoratus. Such class of peptides is known to adopt an amphipathic α-helical conformation, which was reported as essential for its biological activity (Powers and Hancock, 2003). Hence, decoralin showed activity against L. major at micromolar concentrations (IC50 = 72 μM). But when the C-terminal of the peptide was amidated, the anti-parasitic activity demonstrated a IC50 sixfold lower, around 11 μM (Konno et al., 2007). Such enhance in its activity is speculated to be due a stabilization of α-helical conformation caused by C-terminal amidation, as well as an extra electrical charge provided by amidation (Sforça et al., 2004).
Tachyplesin is a class of anti-microbial and APPs from horseshoe-crab (Tachypleus tridentatus), described for the first time in Nakamura et al. (1988). This peptide is 17 amino acids long and have a beta conformation, been active against Gram-positive and negative bacteria, as well as against virus and cancer cells (Shen and Wu, 2014). Bioassays using Leishmania braziliensis demonstrated that tachyplesin could also inhibit this parasite’s growth at micromolar levels (12.5 μM; Löfgren et al., 2008).
Therefore, anti-leishmanicidal peptides from insect sources encompass a safe alternative to control Leishmania infection, once they are highly effective, but not as harmful as the current used drugs. The application of biotechnological tools for the large-scale production of such peptides are essential to aim this goal.
Malaria remains one of the most prevalent and debilitating parasitic infection across Africa, Asia, and America continents. According to the World Health Organization (WHO), approximately 207 million people were diagnosed with Malaria and 627,000 died worldwide from it during 2012 (Ramasamy, 2014). Four species are the main cause of Malaria: Plasmodium falciparum, P. vivax, P. malariae, and P. ovale, transmitted by over 70 species of Anopheles’ mosquitoes. Among the parasites, P. falciparum is the most severe and responsible for 90% of all malaria deaths (Grimberg and Mehlotra, 2011).
Plasmodium’s life cycle is complex, involving multiple developing stages and locations, both in mosquitos and humans. Although its complex infectious cycle offers multiple sites for the development of specific drugs, finding a universal drug against malaria has been a challenging task. The major problem with the currently available anti-malarial drugs is the consistent resistance developed by some parasites, well-reported in literature (Bloland, 2001; White, 2004; Carter and Hurd, 2010; Torrent et al., 2012). Recently, the emerging resistance toward the drug artemisinin decreased even more the chances for a cure of Malaria, once that it used as first-line treatment for uncomplicated cases caused by P. falciparum, in most endemic countries (Biamonte et al., 2013).
Therefore, the demand for novel therapeutic agents against Malaria is urgent, especially using new targets. Nowadays, the new generation of strategies involves the blockage of malaria transmission by employing genetically modified vectors, mosquito pathogens or symbionts that express anti-parasite molecules (Carter et al., 2013).
Additionally, APPs from the innate immune system of insect vectors have been used as sources to control sporogonic stages of the malaria parasites. There are three sources of APPs that are active against sporogonic forms of parasites: (1) Endogenous – peptides that form part of the natural mosquito immune repertoire; (2) Exogenous – peptides isolated from other species; and (3) Synthetic – rational design of novel compounds (Carter and Hurd, 2010).
As there is a bigger number of studies on Malaria, there are also more peptides presenting activity against Plasmodium species isolated from different arthropods (Table 1), including its insect host, Anopheles sp. (Gwadz et al., 1989; Vizioli et al., 2001; Kim et al., 2004; Bell, 2011).
Thus, during Anopheles’s life cycle, malaria mosquito vector (female anopheline) do not develop symptoms related to any infection. However, it was demonstrated that the insect produces three classes of anti-malarial peptides: cecropins, defensins, and gambicins. Recent reports showed that the three classes of peptides present activity against mosquito-stage and, in some cases, blood stage parasites (Bell, 2011). Thus, cecropins A and B demonstrated activity against ookinetes and oocysts of P. falciparum, P. berghei, P. knowlesia, and P. cymonolgi (Gwadz et al., 1989; Kim et al., 2004), while gambicin displayed 54–64% lethality against P. berghei ookinetes at 10–100 μM (Vizioli et al., 2001).
The first report of APPs was described in Boman et al. (1989). In this study, they analyzed the growth inhibition of P. falciparum cells using different concentrations of cecropin and melittin peptides, called CA (l–13) and H (l–13; Boman et al., 1989). During the same year, Gwadz et al. (1989) demonstrated the anti-malarial activity of Cecropin B (from giant silk moths) at a concentration of 0.5 μg/μl (128 μM). The peptide prevented normal development of oocysts from Plasmodium species in Anopheles gambiae insects previously infected with the parasite (Gwadz et al., 1989).
Other important source of anti-malarial peptides was isolated from scorpion and spiders’ venoms. Among these peptides, there is scorpine, isolated from the scorpion Pandinus imperator, whose structure resembles a hybrid between a defensin and a cecropin. This peptide was responsible for the mortality of 98% of P. berghei gametocytes at 15 μM and 100% reduction of P. falciparum parasitemia at 5 μM (Carballar-Lejarazu et al., 2008).
Meucine-24, a α-helical peptide with an N-terminal homologous to melittin and meucine-25 have been isolated from the venom of the scorpion Mesobuthus eupeus and showed ability to inhibit the development of P. berghei ookinetes at micromolar concentrations (IC50 10–20 μM; Gao et al., 2010).
Moreover, several insect defensins have displayed anti-malarial activity. Among them, there are two drosomycins isolated from the Drosophila melanogaster hemolymph. Drosomycin1 is able to inhibit the development of P. berghei ookinetes in 76% at 20 μM, while Drosomycin 2 is able to retard the development of the same parasite in 29% at 20 μM. (Tian et al., 2008). Similarly, gomesin, an insect defensin isolated from the hemocytes of the spider Acanthoscurria gomesiana, inhibited the in vitro growth of intra-erythrocytic forms of P. falciparum, causing a dramatic reduction of gametocyte exflagelation and oocyst population in both P. falciparum and P. berghei (Moreira et al., 2007).
As described here, multiple compounds can be pharmaceutically attractive to treat Malaria, once that current available anti-malarial drugs are inefficient against parasites. Therefore, novel peptides can open new perspectives for the development of new anti-parasitic drugs (Leavy, 2010; Vale et al., 2014).
The description of several anti-microbial peptides with activity against pathogenic bacteria, fungi, protozoa and other harmful organisms are well-described in literature. Those peptides are isolated from different sources, varying from human beings, to plants, insects, marine animals, and even microorganisms (Pelegrini et al., 2007; Lacerda et al., 2014; Vale et al., 2014; Kang et al., 2015). However, few of them show anti-parasitic activity, especially related to human neglected diseases (Li et al., 2006; Moreira et al., 2007; Tian et al., 2008; Fieck et al., 2010; Gao et al., 2010; Rangel et al., 2011; Marr et al., 2012). Among these diseases, Malaria has the most number of published articles on the discovery and characterization of anti-plasmodium proteins and peptides. Considering insect peptides with anti-parasitic activity, Malaria continues reaching the first position in tested molecules, although there are still very few studies demonstrating anti-malarial activities of insect peptides for future use as drugs (Boman et al., 1989; Gwadz et al., 1989; Vizioli et al., 2001; Kim et al., 2004; Moreira et al., 2007; Carballar-Lejarazu et al., 2008; Tian et al., 2008; Gao et al., 2010; Vale et al., 2014).
Recently, anti-malarial peptides from insect sources were classified into four groups: (i) α-helical peptides deprived of Cys residues; (ii) β-pleated peptides containing disulfide bridges; (iii) peptides rich in Pro, Gly, His, Arg, and Trp residues; and (iv) circular anti-microbial peptides (Vale et al., 2014). Out of these groups, only the first one contains peptides from insect sources with anti-parasitic activities against Plasmodium species.
Thus, α-helical peptides deprived of Cys residues correspond to linear cationic small peptides with α-helical structure, whose positive charges come from the presence of Lys and Arg residues. Another feature is the existence of at least 50% of their content composed by hydrophobic amino acid residues (Powers and Hancock, 2003). Cecropins (from H. cecropia) and Melittins (from A. mellifera venom) are some examples of α-helical peptides deprived of Cys residues, due to their α-helical domains and content of hydrophobic residues in the amino-terminal site, respectively (Raghuraman and Chattopadhyay, 2007; Tanaka et al., 2008).
For insect α-helical peptides deprived of Cys residues, the mode of action consists of poration of the lipid membrane through a disordered toroidally shape lining of peptides (Figure 1; Yang et al., 2001; Segunpta et al., 2008). The charged amino acid residues showed to be essential for electrostatic interactions with the lipid membrane, resulting in pore formation (Segunpta et al., 2008).
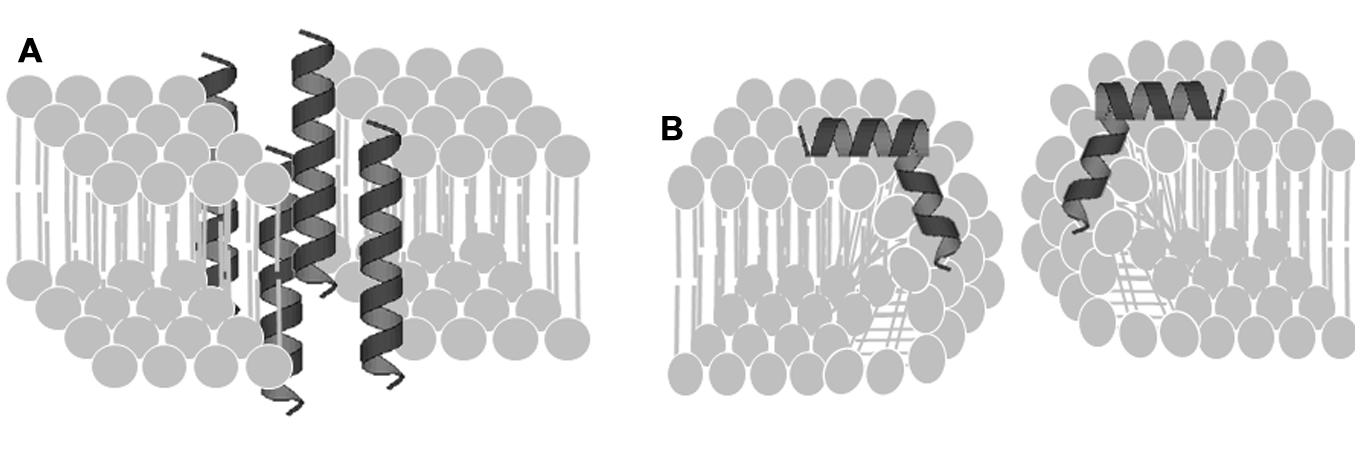
FIGURE 1. Mechanism of action of anti-malarial peptides. (A) Ordered toroidal pore formation. (B) Disoredered toroidal pore formation (Source: Segunpta et al., 2008 with modifications).
In general, the mechanism of action of anti-malarial peptides from insect sources are still poorly studied. However, the activities of distinct APPs obey many of the rules governing their ability to disrupt bacterial membranes. The interaction with APPs to the pathogen’s cell membrane appears to be acutely influenced by the respective charges and amphipathies of the reactants. In fact, a comparison between the peptides’ ability to inhibit the growth of malaria parasites and that of bacteria demonstrates a remarkable parallelism in the way that each modification affects both activities (Mor, 2009).
Transcriptome studies are important to understand the physiology and mechanism of action of parasitic molecules and have been one of the alternatives in seeking resolutions to certain diseases, such as Malaria (Tarun et al., 2008). In this report, transcriptome and proteome of malaria parasite liver stages displayed essential knowledge about how the parasite behaves inside host cells. Moreover, transcriptome studies are also being used to better understand parasite gene profile during infection, as demonstrated by Ngwa et al. (2013). The report showed changes in Plasmodium falciparum transcriptome pattern during its initial phase, describing that modifications were observed even during the first half hour after human transmission by the mosquito (Ngwa et al., 2013). Similarly, Hellgren et al. (2013) used transcriptome data from birds infected with P. relictum, which causes avian Malaria, to identify and characterize the MSP1 gene (SGS1 and GRW4). The analysis of those genes could led to the generation of a new vaccine for the prevention of avian Malaria, also becoming a relevant result for epidemiological studies (Hellgren et al., 2013).
The evaluation of insect transcriptomes, especially the ones that are hosts of human parasites, have been published for some species. Hence, analyses of Musca domestica transcriptome revealed new insights and genes involved in the prophenoloxidase system (proPO), a pathway related to melanization in arthropods. The melanin production occurs after infection, promoting protection against invasion of pathogens, such as bacteria (Li et al., 2015). Therefore, the novel information obtained from transcriptome studies allowed a better understanding on the mechanism of action and immune functions of the proPO system in M. domestica.
The inefficiency of therapeutic strategies to treat protozoa-related diseases and the frequent incidence of resistance developed by parasites led to the search for molecules that can be used as new pharmaceutical products for the treatment of parasitic diseases. In the last years, transcriptomes have been employed as promissory source for the prospection of active proteins and peptides. The availability of high-throughput transcriptomic technologies has contributed to considerably enhance the knowledge to understand host–parasite interactions and to identify new and effective treatments for parasitic diseases, especially neglected ones (Cantacessi et al., 2015; Davies et al., 2015). Furthermore, even transcriptome of phytopathogenic insects could be an interesting source for future prospections of proteins and peptides for therapeutic use on the treatment of Chagas disease, Leishmaniasis, and Malaria (Firmino et al., 2013; Fonseca et al., 2015).
Several researches are published every year on the prospection and discovery of new molecules from different sources with high potential use as drugs against bacterial, fungi, and parasitic infections (Lacerda et al., 2014; Breen et al., 2015; Kang et al., 2015; Tam et al., 2015). Insect peptides with anti-parasitic activity demonstrate to be an interesting tool on the treatment of neglected diseases, such as Malaria, Leishmaniasis, and Chagas disease.
Many reasons explain why neglected diseases are out of reach from eradication. Hygiene issues, social and economic status of the population and endemic areas full of viable parasite vectors are some of them (Alvar et al., 2006; European Alliance Against Malaria, 2007; Klein et al., 2012). Nevertheless, drug inefficiency and misuse, and serious side effects are the main causes of why these diseases are still increasing in number and area worldwide.
The mechanism of action of the drugs used nowadays for the treatment of some neglected diseases are yet not well-known, facilitating mutations triggering drug resistance by the parasites. Moreover, the poor efficient response of such drugs, combined with strong side effects, stimulates the development of alternatives for a better treatment of neglected diseases (Teixeira et al., 2014; Vale et al., 2014; Visser et al., 2014). As it is occurring with anti-bacterial drugs, the misuse of anti-parasitic drugs is also leading to resistance, increasing preoccupation about a growing arrival of new cases and inefficacy on the treatment of recurrent cases (White, 2004; Dondorp et al., 2009; Chakravarty and Sundar, 2010; Alsford et al., 2012; Whyllie et al., 2015).
Nowadays, anti-parasitic drug candidates containing peptides are still very few, with most of them at Clinical Trials stage (Fox, 2013). Nonetheless, the fact that pharmaceutical companies continue avoiding the application of peptides for the production of drugs goes beyond oral bioavailability and drug resistance issues. Peptide degradation by digestive enzymes and interactions with plasma proteins are some conditions that complicate the utilization of APPs for production of drugs. Furthermore, the cost of scaling-up the production of organic compounds is still high, contributing to expensive drugs into the market (Fox, 2013; Vale et al., 2014).
However, despite all disadvantages, since the 1990s, the scenario of drug production is changing. The advances on biotechnology applied for the development of new processes for the production of proteins and peptides for therapeutic use stimulated the formulation of different drugs with high specificity and activity toward their targets, at lower costs and using faster production methods (Leavy, 2010). Novel improvements on biocompatible carriers to extend peptide biocompatibility also contributed for enhancing target reach of peptide’s site of action (Costa et al., 2011; Maia et al., 2014). Similarly, use of nanotechnology and peptide encapsulation using biodegradable polymers allowed to increase the precision of drug target, as well as enabled long-acting release forms of peptides in the organism (Anthony and Freda, 2009; Stark, 2011). Therefore, by 2012, numerous peptide drugs were already approved in the US, reflecting in a market of 13 billion dollars, represented by 1.5% of total global drugs sales. Other countries also approved and released peptide drugs over the years, especially the Europeans Germany and UK, responsible for 63% of peptide therapeutics in the market, followed by France, Italy, Scandinavia, and Spain (Kaspar and Reichert, 2013; Vale et al., 2014).
Hence, although pharma companies’ main target were small molecules (below 500 Da), due to their ease of production and availability for oral application, small drug candidates showed low specificity for their targets, enhancing susceptibility to metabolic inactivation, which increase side effects, dose-response, and decrease their bioavailability (Craik et al., 2013). Peptides with average size of 5 kDa, however, displayed high specificity toward their targets, decreasing not only the dose of use, but also harmful side effects. For example, peptide drugs, such as NVB302, OP-145, Omiganan and Pexiganan, are now in Clinical Trials (Phases I, II, and III, respectively), as future effective alternatives for the treatment of Malaria (Fox, 2013). Therefore, peptide drugs for the treatment of neglected diseases is a reality for future years, with high chances of improvement and expanding in terms of variety and application.
As described before in this report, the activity of insect peptides against the parasites Trypanosoma sp., Leishmania sp., and Plasmodium sp. encourages advances in this research investigation, once there is still a great necessity on developing new effective drugs for the treatment of Chagas disease, Leishmaniasis, and Malaria, respectively. Although poorly explored, insect peptides demonstrate high potential for future application in therapeutics of infectious diseases, especially parasitic neglected ones.
Currently, Malaria, together with other infection disease, such as Tuberculosis and AIDS, receives annually significant amounts of international resources for research on the development of new strategies for their treatment and prevention. Therefore, it was recently removed from the “Neglected disease” group. Nevertheless, there is still no treatment 100% efficient for this disease, but only methods to prolong the life and decrease the pain of patients (Olliaro et al., 2012; Uniting to Combat NTDs, 2014).
Similarly, Chagas disease continues to represent the parasitic illness responsible for the highest number of deaths in Latin America, surpassing Malaria. Considering that 23% of all infected patients with T. cruzi are located in Brazil, this country stands out in the number of published articles about the disease, the parasite, its host vector and on the discovery of novel tools to treat Chagas’s disease (more than 1400 publications during the last 70 years). Out of 55 research institutes working with Chagas’s disease worldwide, six are locate in Brazil. However, yet, there are very few effective and low toxic drugs available for its treatment.
In the absence of appropriate drugs, Leishmaniasis has also represented a health concern, due to its geographic expansion and urbanization trend. Moreover, the control of human Leishmania cannot be done without understanding canine Leishmaniasis, as pet dogs hosting parasites can easily transmit the disease to humans (Papadopoulou et al., 2005).
Advances on molecular biotechnology allow the production of different proteins and peptides in their active form using several heterologous systems. Therefore, scaling up the production of insect peptides for the development of pharmaceutical drugs to be used on the treatment of parasitic diseases has become a routine technique. Similarly, transcriptome analyses can be a relevant tool on the discovery of novel peptides from insect species with activity against parasites. Therefore, it is clear that insect peptides present high potential to become the future drugs for decreasing the number of infectious diseases, such as Malaria, Leishmaniasis, and Chagas disease.
It is not expected that biotechnological peptides will take over chemical molecules on the development of new drugs. However, peptides are already becoming new options for the treatment of several diseases, from oncology to infectious ones, as they can cause synergic effects, enhancing the efficacy of drugs at lower concentration. Therefore, peptide drugs have reached a place of interest by pharmaceutical companies, where despite of their cost of production and biochemical challenges to overcome, peptides demonstrate to be unique in their mechanism of action and therapeutic use.
Peptide drugs for neglected diseases, such as Malaria, Leishmaniasis, and Chagas diseases, are still at early stages compared to other drugs developed for chronic and non-infectious diseases (Lax, 2010). But even at a slow pace, studies and researches on peptide drugs are demonstrating to be a reality for the treatment of neglected diseases. For the next 10–15 years, new peptide drugs are expected to be at the market for the treatment of Malaria and, hopefully, for other neglected diseases including Chagas and Leishmaniasis.
AL: contributed with informations about anti-malarial peptides, introduction of the text, revision, and edition of the entire manuscript. PP: contributed with informations about peptides against Chagas disease’s peptides, introduction of the text, revision, and edition of the entire manuscript. EV: contributed with informations about anti-Leishmanial peptides, conclusion, and English revision of the manuscript. DdO: contributed with informations about transcriptome studies in the manuscript. MG-d-S: contributed with English and content revision of the manuscript.
This article was finnanced by CAPES, EMBRAPA and CNPq.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Alsford, S., Eckert, S., Baker, N., Glover, L., Sanchez-Flores, A., Leung, K. F., et al. (2012). High-throughput decoding of antitrypanosomal drug efficacy and resistance. Nature 25, 232–236. doi: 10.1038/nature10771
Alvar, J., Vélez, I. D., Bern, C., Herrero, M., Desjeux, P., Cano, J., et al. (2012). Leishmaniasis worldwide and global estimates of tis incidence. PLoS ONE 7:e35671. doi: 10.1371/journal.pone.0035671
Alvar, J., Yactayo, S., and Bern, C. (2006). Leishmaniasis and poverty. Trends Parasitol. 22, 552–557. doi: 10.1016/j.pt.2006.09.004
Anthony, L., and Freda, P. U. (2009). From somatostatin to octreotide LAR: evolution of a somatostatin analogue. Curr. Med. Res. Opin. 25, 2989–2999. doi: 10.1185/03007990903328959
Baiocco, P., Colotti, G., Franceschini, S., and Ilari, A. (2009). Molecular basis of antimony treatment in leishmaniasis. J. Med. Chem. 52, 2603–2612. doi: 10.1021/jm900185q
Bell, A. (2011). Antimalarial peptides: the long and the short of it. Curr. Pharm. Des. 17, 2719–2731. doi: 10.2174/138161211797416057
Bern, C., Montgomery, S. P., Herwaldt, B. L., Rassi, A. Jr., Marin-Neto, J. A., Dantas, R. O., et al. (2007). Evaluation and treatment of Chagas disease in the United States: a systematic review. JAMA 298, 2171–2181. doi: 10.1001/jama.298.18.2171
Biamonte, M. A., Wanner, J., and Le Roch, K. G. (2013). Recent advances in malaria drug discovery. Bioorg. Med. Chem. Lett. 23, 2829–2843. doi: 10.1016/j.bmcl.2013.03.067
Boman, H. G. (1991). Antibacterial peptides: key components needed in immunity. Cell 65, 205–207. doi: 10.1016/0092-8674(91)90154-Q
Boman, H. G., Faye, I., Lee, J. Y., Gudmundsson, G. H., and Lidholm, D. A. (1991). Cell-free immunity in Cecropia. A model system for antibacterial proteins. Eur. J. Biochem. 201, 23–31. doi: 10.1111/j.1432-1033.1991.tb16252.x
Boman, H. G., Wadde, D., Boman, I. A., Wihlint, B., and Merrifield, R. B. (1989). Antibacterial and antimalarial properties of peptides that are cecropin-melittin hybrids. FEBS Lett. 259, 103–106. doi: 10.1016/0014-5793(89)81505-4
Brasil (2010). Doenças Infecciosas e Parasitárias: Guia de Bolso, 8 Rev Edn. Brasília: Ministério da Saúde, Secretaria de Vigilância em saúde, Departamento de Vigilância Epidemiológica, 444.
Breen, S., Solomon, P. S., Bedon, F., and Vincent, D. (2015). Surveying the potential of secreted antimicrobial peptides to enhance plant disease resistance. Front. Plant Sci. 6:900. doi: 10.3389/fpls.2015.00900
Cantacessi, C., Dantas-Torres, F., Nolan, M. J., and Otranto, D. (2015). The past, present, and future of Leishmania genomics and transcriptomics. Trends Parasitol. 31, 100–108. doi: 10.1016/j.pt.2014.12.012
Carballar-Lejarazu, R., Rodriguez, M. H., Hernnedez-Hernnedez, F. C., Ramos-Castañeda, J., Possanic, L. D., Zurita-Ortega, M., et al. (2008). Recombinant scorpine: multifunctional antimicrobial peptides with activity against different pathogens. Cell Mol. Life Sci. 65, 3081–3092. doi: 10.1007/s00018-008-8250-8
Carter, V., and Hurd, H. (2010). Choosing anti-Plasmodium molecules for genetically modifying mosquitoes: focus on peptides. Trends Parasitol. 26, 582–590. doi: 10.1016/j.pt.2010.07.005
Carter, V., Underhill, A., Baber, I., Sylla, L., Baby, M., Larget-Thiery, I., et al. (2013). Killer bee molecules: antimicrobial peptides as effector molecules to target sporonogic stages of Plasmodium. PLoS Pathog. 9:e1003790. doi: 10.1371/journal.ppat.1003790
Casteels, P., Ampe, C., Jacobs, F., Vaeck, M., and Tempst, P. (1989). Apidaecins: antibacterial peptides from honeybees. EMBO J. 8, 2387–2391.
Casteels, P., and Tempst, P. (1994). Apidaecin-type peptide antibiotics function through a non-poreforming mechanism involving stereospecificity. Biochem. Biophys. Res. Commun. 199, 339–345. doi: 10.1006/bbrc.1994.1234
Center for Disease Control and Prevention [CDC] (2014). Malaria Worldwide. Available at: http://www.cdc.gov/malaria/malaria_worldwide/ [accessed October 20, 2014].
Chadbourne, F. L., Raleigh, C., Ali, H. Z., Dennya, P. W., and Cobb, S. L. (2011). Studies on the antileishmanial properties of the antimicrobial peptides temporin A, B and 1Sa. J. Pept. Sci. 17, 751–755. doi: 10.1002/psc.1398
Chagas, C. (1981). Carlos Chagas: Coletanea de Trabalhos Científicos, Vol. 6. Brasilia: Editora Universidade de Brasília, 247–258.
Chakravarty, J., and Sundar, S. (2010). Drug resistance in Leishmaniasis. J. Glob. Infect Dis. 2, 167–176. doi: 10.4103/0974-777X.62887
Chattopadhyay, A., and Jafurulla, M. (2011). A novel mechanism for an old drug: amphotericin B in the treatment of visceral leishmaniasis. Biochem. Biophys. Res. Commun. 416, 7–12. doi: 10.1016/j.bbrc.2011.11.023
Costa, F., Carvalho, I. F., Montelaro, R. C., Gomes, P., and Martins, M. C. (2011). Covalent immobilization of antimicrobial peptides (AMPs) onto biomaterial surfaces. Acta Biomater. 7, 1431–1440. doi: 10.1016/j.actbio.2010.11.005
Coura, J. R., and Viñas, P. A. (2010). Chagas disease: a new worldwide challenge. Nature 56, S6–S7. doi: 10.1038/nature09221
Craik, D. J., Fairlie, D. P., Liras, S., and Price, D. (2013). The future of peptide-based drugs. Chem. Biol. Drug Des. 81, 136–147. doi: 10.1111/cbdd.12055
Davies, P. D., Bodmer, J. L., Felgner, P. L., and Doolan, D. L. (2015). Large screen approaches to identify novel malaria vaccine candidates. Vaccine 33, 7496–7505. doi: 10.1016/j.vaccine.2015.09.059
Don, R., and Chatelain, E. (2008). “Drug discovery for neglected diseases: view of a public–private partnership,” in Antiparasitc and Antibacterial Drug Discovery: From Molecules Targets to Drug Candidates, ed. P. M. Selzer (Blackwell: Wiley), 58–67.
Dondorp, A. M., Nosten, F., Yi, P., Das, D., Phyo, A. P., Tarning, J., et al. (2009). Artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 361, 455–467. doi: 10.1056/NEJMoa0808859
European Alliance Against Malaria (2007). Malaria & Poverty. London: European Alliance Against Malaria, 1–4.
Fieck, A., Hurwitz, I., Kang, A. S., and Durvasula, R. (2010). Trypanosoma cruzi: synergistic cytotoxicity of multiple amphipathic anti-microbial peptides to T. cruzi and potential bacterial hosts. Exp. Parasitol. 125, 342–347. doi: 10.1016/j.exppara.2010.02.016
Firmino, A. A., Fonseca, F. C., de Macedo, L. L., Coelho, R. R., Antonino de Souza, J. D. Jr., Togawa, R. C., et al. (2013). Transcriptome analysis in cotton boll weevil (Anthonomus grandis) and RNA interference in insect pests. PLoS ONE 8:e85079. doi: 10.1371/journal.pone.0085079
Fonseca, F. C., Firmino, A. A. P., Macedo, L. L., Coelho, R. R., de Souza Júnior, J. D., Silva-Junior, O, B., et al. (2015). Sugarcane giant borer transcriptome analysis and identification of genes related to digestion. PLoS ONE 10:e0118231. doi: 10.1371/journal.pone.0118231
Fox, J. L. (2013). Antimicrobial peptides stage a comeback. Nat. Biotechnol. 31, 379–382. doi: 10.1038/nbt.2572
Frearson, J. A., Wyatt, P. G., Gilbert, I. H., and Fairlamb, A. H. (2007). Target assessment for antiparasitic drug discovery. Trends Parasitol. 23, 589–595. doi: 10.1016/j.pt.2007.08.019
Gao, B., Xu, J., Rodriguez Mdel, C., Lanz-Mendonza, H., Hernández-Rivas, R., Du, W., et al. (2010). Characterization of two linear cationic antimalarial peptides in the scorpion Mesobuthus eupeus. Biochimie 92, 350–359. doi: 10.1016/j.biochi.2010.01.011
Grimberg, B. T., and Mehlotra, R. K. (2011). Expanding the antimalarial drug arsenal – now but how? Pharmaceuticals 4, 681–712. doi: 10.3390/ph4050681
Gwadz, R. W., Kaslow, D., Lee, J. Y., Maloy, W. L., Zasloff, M., and Miller, L. H. (1989). Effects of magainins and cecropins on the sporogonic development of malaria parasites in mosquitoes. Infect. Immun. 57, 2628–2633.
Hellgren, O., Kutzer, M., Bensch, S., Valkiunas, G., and Palinauskas, V. (2013). Identification and characterization of the merozoite surface protein 1 (msp1) gene in a host-generalist avian malaria parasite, Plasmodium relictum (lineages SGS1 and GRW4) with the use of blood transcriptome. Malar J. 12, 381. doi: 10.1186/1475-2875-12-381
Hood, J. L., Jallouk, A. P., Campbell, N., Ratner, L., and Wickline, S. A. (2013). Cytolytic nanoparticles attenuate HIV-1 infectivity. Antivir. Ther. 18, 95–103. doi: 10.3851/IMP2346
Jaureguiberry, P., Graby, G., and Caumes, E. (2006). Efficacy of short-course intramuscular pentamidine isethionate treatment on old world localized cutaneous leishmaniasis in 2 patients. Clin. Infect. Dis. 42, 1812–1813. doi: 10.1086/504430
Jaynes, J. M., Burton, C. A., Barr, S. B., Jeffers, G. W., Julian, G. R., and White, K. L. (1988). In vitro cytocidal effect of novel lytic peptides on Plasmodium falciparum and Trypanosoma cruzi. FASEB J. 2, 2878–2883.
Kang, H. K., Seo, C. H., and Park, Y. (2015). Marine peptides and their anti-infective activities. Mar. Drugs 13, 618–654. doi: 10.3390/md13010618
Kaspar, A. A., and Reichert, J. M. (2013). Future directions for peptide therapeutics developments. Drug Discov. Today 18, 807–817. doi: 10.1016/j.drudis.2013.05.011
Kaye, P., and Scott, P. (2011). Leishmaniasis: complexity at the host–pathogen interface. Nat. Rev. 9, 604–615. doi: 10.1038/nrmicro2608
Kedzierski, L. (2010). Leishmaniasis vaccine: where are we today? J. Glob. Infect. Dis. 2, 177–185. doi: 10.4103/0974-777X.62881
Kehoe, M., and Timmis, K. N. (1984). Cloning and expression in Escherichia coli of the streptolysin O determinant from Streptococcus pyogenes: characterization of the cloned streptolysin O determinant and demonstration of the absence of substantial homology with determinants of other thiol-activated toxins. Infect. Immun. 43, 804–810.
Kennedy, T. (1997). Managing the drug discovery/development interface. Drug Discov. Today 2, 436–444. doi: 10.1016/S1359-6446(97)01099-4
Kevin, M. B. (2014). Chagas disease in the 21th century: a public health success or an emerging threat? Parasite 21:11. doi: 10.1051/parasite/2014012
Kim, W., Koo, H., Richman, A. M., Seeley, D., Vizioli, J., Klocko, A. D., et al. (2004). Ectopic expression of a cecropin transgene in the human malaria vector mosquito Anopheles gambiae (Diptera: Culicidae): effects on susceptibility to Plasmodium. J. Med. Entomol. 41, 447–455. doi: 10.1603/0022-2585-41.3.447
Klein, N., Hurwitz, I., and Durvasula, R. (2012). Globalization of chagas disease: a growing concern in nonendemic countries. Epidemiol. Res. Int. 2012:13. doi: 10.1155/2012/136793
Klotz, S. A., Gaur, N. K., Rauceo, J., Lake, D. F., Park, Y., Hahm, K. S., et al. (2004). Inhibition of adherence and killing of Candida albicans with a 23-Mer peptide (Fn/23) with dual antifungal properties. Antimicrob. Agents Chemother. 48, 4337–4341. doi: 10.1128/AAC.48.11.4337-4341.2004
Konno, K., Hisada, M., Naoki, H., Itagaki, Y., Fontana, T., Rangel, M., et al. (2006). Eumenitin, a novel antimicrobial peptide from the venom of the solitary eumenine wasp Eumenesru bronotatus. Peptides 27, 2624–2631. doi: 10.1016/j.peptides.2006.04.013
Konno, K., Rangel, M., Oliveira, J. S., Cabrera, M. P. S., Fontana, R., Hirata, I. Y., et al. (2007). Decoralin, a novel linear cationic a-helical peptide from the venom of the solitary eumenine wasp Oreumenes decoratus. Peptides 28, 2320–2327. doi: 10.1016/j.peptides.2007.09.017
Lacerda, A. F., Vasconcelos, E. A. R., Pelegrini, P. B., and Grossi-de-Sa, M. F. (2014). Antifungal defensins and their role in plant defense. Front. Microbiol. 5:116. doi: 10.3389/fmicb.2014.00116
Landon, C., Meudal, H., Boulanger, N., Bulet, P., and Vovelle, F. (2006). Solution structures of stomoxyn and spinigerin, two insect antimicrobial peptides with anα-helical conformation. Biopolymers 81, 92–103. doi: 10.1002/bip.20370
Lax, R. (2010). The Future of Peptide Development in the Pharmaceutical Industry, Pharmanuacturing: the International Peptide Review. Available at: http://www.polypeptide.com/web/upload/medias/1401702726538c49464a6f5.pdf [accessed December 17, 2015].
Lazarev, V. N., Shkarupeta, M. M., Titova, G. A., Kostrjukova, E. S., Akopian, T. A., and Govorun, V. M. (2005). Effect of induced expression of an antimicrobial peptide melittinon Chlamydia trachomatis and Mycoplasma hominis infections in vivo. Biochem. Biophys. Res. Commun. 338, 946–950. doi: 10.1016/j.bbrc.2005.10.028
Leavy, O. (2010). Therapeutic antibodies: past, present and future. Nat. Rev. Immunol. 10:297. doi: 10.1038/nri2763
Li, D., Liang, Y., Wang, X., Wang, L., Qi, M., Yu, Y., et al. (2015). Transcriptomic analysis of musca domestica to reveal key genes of the prophenoloxidase-activating system. Gen. Immun. 5, 1827–1841. doi: 10.1534/g3.115.016899
Li, W. F., Ma, G. X., and Zhou, X. X. (2006). Apidaecin-type peptides: biodiversity, structure-function relationships and mode of action. Peptides 27, 2350–2359. doi: 10.1016/j.peptides.2006.03.016
Löfgren, S. E., Miletti, L. C., Steindel, M., Bachère, E., and Barracco, M. A. (2008). Trypanocidal and leishmanicidal activities of different antimicrobial peptides (AMPs) isolated from aquatic animals. Exp. Parasitol. 118, 197–202. doi: 10.1016/j.exppara.2007.07.011
Loftus, P. (2009). The buzz: targeting cancer with bee venom. Wall Street J. Available at: http://www.wsj.com/articles/SB10001424052970203803904574433382922095534 [accessed July 17, 2015].
Lubke, L. L., and Garon, C. F. (1997). The antimicrobial agent melittin exhibits powerful in vitro inhibitory effects on the Lyme disease spirochete. Clin. Infect. Dis. 25, S48–S51. doi: 10.1086/516165
Maia, F. R., Barbosa, M., Gomes, D. B., vale, N., Granja, P., Gomes, P., et al. (2014). Hydrogel depots for local co-delivery of osteoinductive peptides and mesenchymal stem cells. J. Control. Release 189, 158–168. doi: 10.1016/j.jconrel.2014.06.030
Marr, A. K., McGwire, B. S., and McMaster, W. R. (2012). Modes of action of Leishmanicidal antimicrobial peptides. Future Microbiol. 7, 1047–1059. doi: 10.2217/fmb.12.85
McGwire, B. S., and Satoskar, A. R. (2014). Leishmaniasis: clinical syndromes and treatment. Int. J. Med. 107, 7–14.
Mor, A. (2009). Multifunctional host defense peptides: antiparasitic activities. FEBS J. 276, 6474–6482. doi: 10.1111/j.1742-4658.2009.07358.x
Moreira, C. K., Rodrigues, F. G., Ghosh, A., Varotti, F. P., Miranda, A., Daffre, S., et al. (2007). Effect of the antimicrobial peptide gomesin against different life stages of Plasmodium spp. Exp. Parasitol. 116, 346–353. doi: 10.1016/j.exppara.2007.01.022
Nakamura, T., Furunaka, H., Miyata, T., Tokunaga, F., Muta, T., Iwanaga, S., et al. (1988). Tachyplesin, a class of antimicrobial peptide from the hemocytes of the horseshoe-crab (Tachypleus tridentatus) - isolation and chemical-structure. J. Biol. Chem. 263, 16709–16713.
Ngwa, C. J., Scheuermayer, M., Mair, G. R., Kern, S., Brügl, T., Wirth, C. C., et al. (2013). Changes in the transcriptome of the malaria parasite Plasmodium falciparum during the initial phase of transmission from the human to the mosquito. BMC Genomics 14:256. doi: 10.1186/1471-2164-14-256
Olliaro, P., Vaillant, M. T., Sundar, S., and Balaseagaram, M. (2012). More efficient ways of assessing treatments for neglected tropical diseases are required: innovative study designs, new endpoints, and markers of effects. PLoS Negl. Trop. Dis. 6:e1545. doi: 10.1371/journal.pntd.0001545
Paes, N. A., and Silva, L. A. (1999). Doenças infecciosas e parasitárias no Brasil: uma década de transição. Rev Panam Salud Publica/Pan. Am. J. Public Health 6, 99–109. doi: 10.1590/S1020-49891999000700004
Papadopoulou, C., Kostoula, A., Dimitriou, D., Panagiou, A., Bobojianni, C., and Antoniades, G. (2005). Human and canine leishmaniasis in asymptomatic and symptomatic population in Northwestern Greece. J. Infect. 50, 53–60. doi: 10.1016/j.jinf.2004.05.004
Parada, H., Carrasco, H. A., Anez, N., Fuenmayor, C., and Inglessis, I. (1997). Cardiac involvement is a constant finding in acute Chagas disease: a clinical, parasitological and histopathological study. Int. J. Cardiol. 60, 49–54. doi: 10.1016/S0167-5273(97)02952-5
Pelegrini, P. B., Quirino, B. F., and Franco, O. L. (2007). Plant cyclotides: na unusual class of defense compounds. Peptides 28, 1475–1481. doi: 10.1016/j.peptides.2007.04.025
Powers, J. P. S., and Hancock, R. E. W. (2003). The relationship between peptide structure and antibacterial activity. Peptides 24, 1681–1691. doi: 10.1016/j.peptides.2003.08.023
Public Health England (2014). Malaria: Guidance, Data and Analysis. Available at https://www.gov.uk/government/collections/malaria-guidance-data-and-analysis [accessed October 19, 2014].
Raghuraman, H., and Chattopadhyay, A. (2007). Melittin: a membrane-active peptide with diverse functions. Biosci. Rep. 27, 189–223. doi: 10.1007/s10540-006-9030-z
Ramasamy, R. (2014). Zoonotic malaria – global overview and research and policy needs. Front. Public Health 2:123. doi: 10.3389/fpubh.2014.00123
Rangel, M., Cabrera, M. P. S., Kazuma, K., Ando, K., Wang, X., Kato, M., et al. (2011). Chemical, and biological characterization of four new linear cationic α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 57, 1081–1092. doi: 10.1016/j.toxicon.2011.04.014
Rosendaal, J. A. (1997). Vector Control: Methods for Use by Individuals, and Communities, Chap. 3. Geneva: World Health Organization.
Segunpta, D., Leonatiadou, H., Mark, A. E., and Marrink, S.-J. (2008). Toroidal pores formed by antimicrobial peptides show significant disorder. Biochim. Biophys. Acta 1178, 2308–2317. doi: 10.1016/j.bbamem.2008.06.007
Sforça, M. L., Oyama, S. Jr., Canduri, F., Lorenzi, C. C. B., Pertinez, T. A., and Konno, K. (2004). How C-terminal carboxyamidation alters the mast cell degranulating activity of peptides from the venom of the eumenine solitary wasp. Biochemistry 43, 5608–5617. doi: 10.1021/bi0360915
Shen, D., and Wu, B. (2014). Structure, biological properties and utilities of marine-derived antimicrobial peptides. Curr. Organ. Chem. 18, 793–803.
Stark, W. J. (2011). Nanoparticles in biological systems. Angewandre Chem. Int. Edn. 50, 1242–1258. doi: 10.1002/anie.200906684
Sundar, S., Sinha, P. K., Rai, M., Verma, D. K., Nawin, K., and Alam, S. (2011). Comparison of short-course multidrug treatment with standard therapy for visceral leishmaniasis in India: an open label, non-inferiority, randomized controlled trial. Lancet 377, 477–486. doi: 10.1016/S0140-6736(10)62050-8
Tam, J. P., Wang, S., Wong, K. H., and Tan, W. L. (2015). Antimicrobial peptides from plants. Pharmaceuticals (Basel) 8, 711–757. doi: 10.3390/ph8040711
Tanaka, H., Ishibashi, J., Fujita, K., Nakajima, Y., Sagisaka, A., Tomimoto, K., et al. (2008). A genome-wide analysis of genes and gene families involved in innate immunity of Bombyx mori. Insect Biochem. Mol. Biol. 38, 1087–1110. doi: 10.1016/j.ibmb.2008.09.001
Tarun, A. S., Peng, X., Dumpit, R. F., Ogata, Y., Rivera-Silva, H., Camargo, N., et al. (2008). A combined transcriptome and proteome survey of malaria parasite liver stages. Proc. Natl. Acad. Sci. U.S.A. 105, 305–310. doi: 10.1073/pnas.0710780104
Teixeira, C., Vale, N., Pérez, B., Gomes, A., Gomes, J. R. B., and Gomes, P. (2014). “Recycling” classical drugs for malária. Chem. Rev. 114, 11164–11220. doi: 10.1021/cr500123g
Tian, C., Gao, B., Rodriguez Mdel, C., Lanz-Mendoza, H., Ma, B., and Zhu, S. (2008). Gene expression, antiparasitic activity, and functional evolution of the drosomycin family. Mol. Immunol. 45, 3909–3916. doi: 10.1016/j.molimm.2008.06.025
Torrent, M., Pulido, D., Rivas, L., and Andreu, D. (2012). Antimicrobial peptide action on parasites. Curr. Drug Targets 13, 1138–1147. doi: 10.2174/138945012802002393
Uniting to Combat NTDs (2014). NTD Report 2014: Delivering on Promises and Driving Progress. London: Uniting to Combat NTDs 46. Available at: http://unitingtocombatntds.org/sites/default/files/document/NTD%20Report%20Final%20%281%29.pdf
Vale, N., Aguiar, L., and Gomes, P. (2014). Antimicrobial peptides: a new class of antimalarial drugs. Front. Phamacol. 5:275. doi: 10.3389/fphar.2014.00275
Verma, N., Karmakar, M., Singh, K. P., and Smita, S. (2013). Structural and dynamic insights into S100B protein activity inhibition by melittin for the treatment of epilepsy. Int. J. Comp. App. NSAAILS 1, 55–60.
Visser, B. J., van Vugt, M., and Grobusch, M. P. (2014). Malaria: an update on current chemotherapy. Expert Opin. Pharmacother. 15, 2219–2254. doi: 10.1517/14656566.2014.944499
Vizioli, J., Bulet, P., Hoffmann, J. A., Kafatos, F. C., Muller, H. M., and Dimopoulos, G. (2001). Gambicin: a novel immune responsive antimicrobial peptide from the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. U.S.A. 98, 12630–12635. doi: 10.1073/pnas.221466798
White, N. J. (2004). Antimalarial drug resistance. J. Clin. Invest. 113, 1084–1092. doi: 10.1172/JCI21682
Whitebread, S., Hamon, J., Bojanic, D., and Urban, L. (2005). Keynote review: in vitro safety pharmacology profiling: an essential tool for successful drug development. Drug Discov. Today 10, 1421–1433.
Whyllie, S., Forth, B. J., Kelner, A., Sokolova, A. Y., Berriman, M., and Farlamb, A. H. (2015). Nitroheterocyclic drug resistance mechanisms in Ttrypanosoma brucei. J. Antimicrob. Chemother. [Epub ahead of print].
World Health Organization [WHO] (2014). Chagas Disease (American Trypanosomiasis): Fact Sheet n. 340. Available at: http://www.who.int/mediacentre/factsheets/fs340/en/ [accessed October 20, 2014].
World Health Organization [WHO] (2015a). Investing to Overcome the Global Impact of Neglected Tropical Diseases: The Who Report on Neglected Tropical Diseases. Geneva: World Health Organization, 211.
World Health Organization [WHO] (2015b). Leishmaniasis. Media Center. Fact Sheet n. 375. Available at: http://www.who.int/mediacentre/factsheets/fs375/en/ [accessed December 17th, 2015].
Yang, D., Chertov, O., and Oppenheim, J. J. (2001). Participation of mammalian defensins and cathelicidins in anti-microbial immunity: receptors and activities of human defensins and cathelicidin (LL-37). J. Leukoc. Biol. 69, 691–697.
Keywords: proteins, peptides, anti-parasitic, host insects, tropical diseases
Citation: Lacerda AF, Pelegrini PB, de Oliveira DM, Vasconcelos ÉAR and Grossi-de-Sá MF (2016) Anti-parasitic Peptides from Arthropods and their Application in Drug Therapy. Front. Microbiol. 7:91. doi: 10.3389/fmicb.2016.00091
Received: 22 November 2015; Accepted: 18 January 2016;
Published: 05 February 2016.
Edited by:
Luis Cláudio Nascimento Da Silva, University of Copenhagen, DenmarkReviewed by:
Robert Burman, Uppsala University, SwedenCopyright © 2016 Lacerda, Pelegrini, de Oliveira, Vasconcelos and Grossi-de-Sá. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patrícia B. Pelegrini, cGJwZWxlZ3JpbmlAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.