
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 27 October 2015
Sec. Food Microbiology
Volume 6 - 2015 | https://doi.org/10.3389/fmicb.2015.01197
This article is part of the Research Topic The controversial nature of the weissella genus: biotechnological, functional and safety aspects of its application in food and health View all 7 articles
 Hikmate Abriouel1*Leyre Lavilla Lerma1María del Carmen Casado Muñoz1Beatriz Pérez Montoro1Jan Kabisch2Rohtraud Pichner2Gyu-Sung Cho2Horst Neve2Vincenzina Fusco3Charles M. A. P. Franz2Antonio Gálvez1Nabil Benomar1
Hikmate Abriouel1*Leyre Lavilla Lerma1María del Carmen Casado Muñoz1Beatriz Pérez Montoro1Jan Kabisch2Rohtraud Pichner2Gyu-Sung Cho2Horst Neve2Vincenzina Fusco3Charles M. A. P. Franz2Antonio Gálvez1Nabil Benomar1Despite the use of several Weissella (W.) strains for biotechnological and probiotic purposes, certain species of this genus were found to act as opportunistic pathogens, while strains of W. ceti were recognized to be pathogenic for farmed rainbow trout. Herein, we investigated the pathogenic potential of weissellas based on in silico analyses of the 13 whole genome sequences available to date in the NCBI database. Our screening allowed us to find several virulence determinants such as collagen adhesins, aggregation substances, mucus-binding proteins, and hemolysins in some species. Moreover, we detected several antibiotic resistance-encoding genes, whose presence could increase the potential pathogenicity of some strains, but should not be regarded as an excluding trait for beneficial weissellas, as long as these genes are not present on mobile genetic elements. Thus, selection of weissellas intended to be used as starters or for biotechnological or probiotic purposes should be investigated regarding their safety aspects on a strain to strain basis, preferably also by genome sequencing, since nucleotide sequence heterogeneity in virulence and antibiotic resistance genes makes PCR-based screening unreliable for safety assessments. In this sense, the application of W. confusa and W. cibaria strains as starter cultures or as probiotics should be approached with caution, by carefully selecting strains that lack pathogenic potential.
Weissella species are non-spore forming, catalase-negative and Gram-positive bacteria that are non-motile, with the exception of Weissella (W.) beninensis. To date, the Weissella genus comprises 19 validly described species (February 20151). Most of these were isolated from and associated with fermented foods, e.g., W. confusa, W. cibaria (Björkroth et al., 2002), W. kimchii (Choi et al., 2002), W. koreensis (Lee et al., 2002), and W. viridescens (Comi and Iacumin, 2012). Weissella viridescens is considered the type species of the Weissella genus (Collins et al., 1994). As a member of the lactic acid bacteria (LAB), Weissella have complex nutritional requirements. Because of this, they inhabit nutrient-rich environments and can be isolated from a variety of such sources, including vegetables, meat, fish, raw milk, sewage, blood, soil, the gastrointestinal tracts of humans and animals, as well as the oral cavity and uro-genital tract of humans (Fusco et al., 2015).
From a technological point of view, Weissella plays an important role in fermentation processes such as the production of silage, as well as in food fermentations based on vegetables or meat as substrate (Björkroth et al., 2002; Santos et al., 2005).
Several weissellas, mainly belonging to the W. confusa and W. cibaria species, are being extensively studied for their ability to produce significant amounts of non-digestible oligosaccharides and extracellular polysaccharides, which can be used as prebiotics or for other applications in food, feed, clinical, and cosmetics industries.
Furthermore, several Weissella strains have been found to act as probiotics, mainly due to their antimicrobial activity, as is the case for certain bacteriocinogenic strains of W. paramesenteroides, W. hellenica, and W. cibaria (Fusco et al., 2015). For example, W. hellenica DS-12 isolated from flounder intestine has been used as probiotic in fish, due to its antimicrobial activity against fish pathogens, such as Edwardsiella, Pasteurella, Aeromonas, and Vibrio (Cai et al., 1998). Also, strains of W. cibaria were proposed as probiotics for oral health, inhibiting Streptococcus mutans glucan biofilm formation (Kang et al., 2006). Recently, weissellas were also shown to exhibit chemopreventive and anti-tumor effects (Kwak et al., 2014).
On the detrimental side, some weissellas were reported to be involved in disease outbreaks such as otitis, sepsis, endocarditis, and even fish mortality (Flaherty et al., 2003; Harlan et al., 2011; Lee et al., 2011; Welch and Good, 2013). Human infections caused by Weissella spp. are, however, rarely reported, and occur mostly in patients with impaired host defenses (Lee et al., 2011). Curiously, therefore, in the same species both beneficial and detrimental strains can be found. As an example, W. confusa causes sepsis and other serious infections in humans and animals, while it has a functional role in food fermentations and has also been suggested as a probiotic (Fusco et al., 2015).
The probiotic and pro-technological potential of weissellas therefore collides with the potential of these bacteria as human pathogens. As for enterococci, whose use for food and health application has been controversial (Franz et al., 2003; Ogier and Serror, 2008), a safety assessment of each strain that is intended to be used as starter culture or as probiotic, should thus be recommended.
Whole genome sequencing and sequence annotation is increasingly being used as valuable tool for assessing microbial food quality and safety aspects (Alkema et al., 2015), allowing the identification of new genes that may have an important impact on cell metabolism, fitness, and virulence.
Herein, we report the investigation of the pathogenic potential of weissellas based on in silico analyses of the 13 whole genome sequences (of 13 strains belonging to nine Weissella species) to date available.
Data sequences of genomes of the 13 strains belonging to nine Weissella species (Table 1) were retrieved from the National Centre for Biotechnology Information (NCBI2; accessed on February, 2015). All genome sequences available were analyzed for the presence of different virulence determinants (aggregation substances, adhesins, toxins, pili, hemolysins) and of antibiotic resistance genes, such as fosfomycin and methicillin resistance genes. The accession numbers of each target gene sequence are indicated in Tables 1–3 and Figures 1–5.
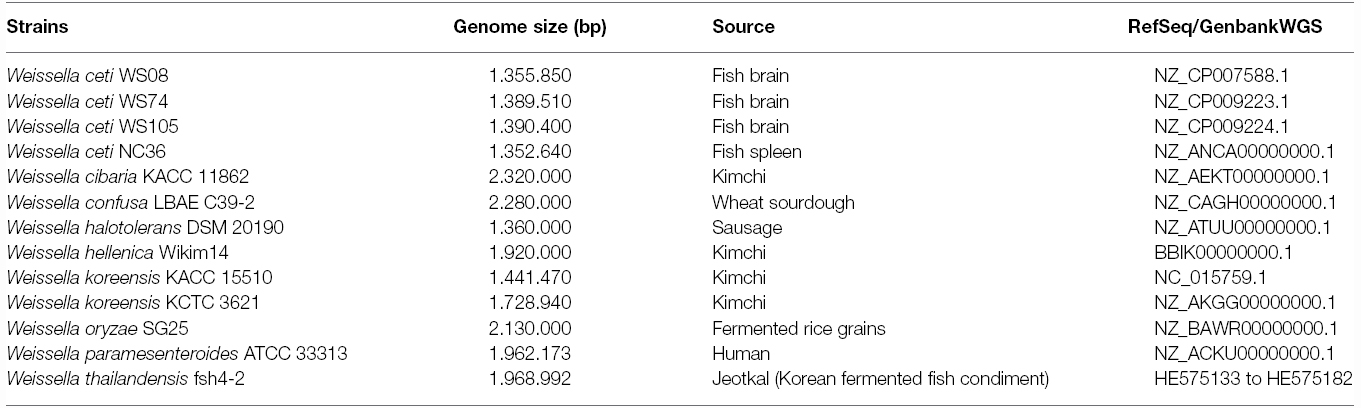
Table 1. Genome characteristics of Weissella species isolated from different sources.
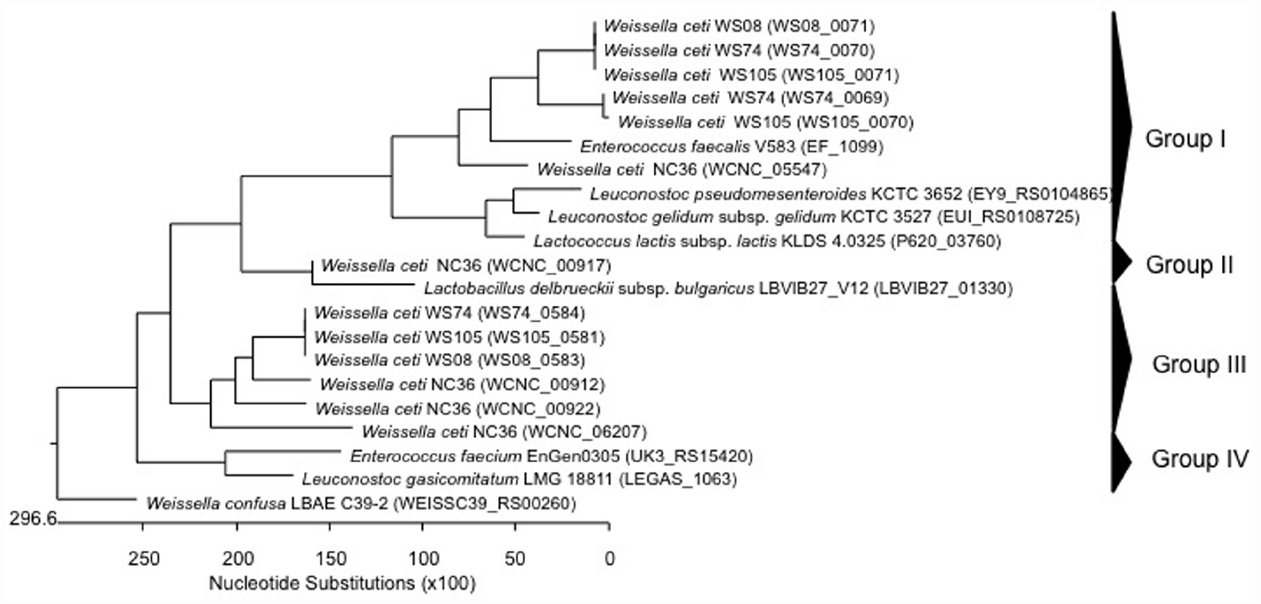
Figure 1. Phylogenetic relationships of Weissella species and Leuconostoc Lactobacillus Enterococcus inferred from the alignment of collagen adhesin encoding genes. The sequences were aligned and the most parsimonious phylogenetic trees were constructed using the CLUSTAL W of Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). The scale below indicates the number of nucleotide substitutions. Accession numbers are indicated in parentheses.
All selected target genes were subjected to phylogenetic analyses to determine phylogenetic relationships with those of closely related genera. Alignment of sequences was done using the CLUSTAL W module of the Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). Phylogenetic trees were reconstructed by the maximum parsimony method using MegAlign (Lasergene program, version 5.05).
The safety of many LAB species have been recognized as GRAS (Generally Regarded As Safe; for the USA; Food and Drug Administration, 1999) or have attained the QPS (Qualified Presumption of Safety; for the European Commission; European Food Safety Authority (EFSA), 2004) status. Indeed, many people refer to these bacteria as being innocuous, and even associated with health beneficial properties. However, it is well known that certain species within a genus, or even certain strains within a specific species, may have different health impacts, as has been pointed out for the enterococci (Franz et al., 2003; Ogier and Serror, 2008). So far, no Weissella species have QPS status (European Food Safety Authority (EFSA), 2015) Data on virulence factors present in Weissella species are quite scarce, and genomic analysis can therefore aid in detecting and describing the occurrence of virulence determinants that may be present at species or strain level within this genus. In this regard, Ladner et al. (2013) found in the genome sequence of W. ceti NC36, an emerging pathogen of farmed rainbow trout in the United States, the presence of several putative virulence factor genes, which did not have homologs encoded in any of the other sequenced Weissella genomes. In particular, these include five collagen adhesin genes (WCNC_00912, WCNC_00917, WCNC_00922, WCNC_05547, and WCNC_06207), a platelet-associated adhesin gene (WCNC_01820) and a gene for a mucus-binding protein (WCNC_01840; Ladner et al., 2013). The five collagen adhesin genes included those from W. ceti WS105 (three collagen adhesins), W. ceti WS74 (three collagen adhesins), W. ceti WS08 (two collagen adhesins), and W. confusa LBAE C39-2 (one collagen adhesin; Table 2). DNA sequences coding for collagen adhesin proteins in Weissella spp. and other LAB (Leuconostoc “Ln.”, Lactococcus “Lc.”, Enterococcus “E.”, and Lactobacillus “Lb.”) were aligned using CLUSTAL W of the Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA) and the most parsimonious phylogenetic tree was reconstructed. The dendrogram generated showed four groups (Figure 1). The first group contained two of the three collagen adhesin genes found in W. ceti strains, E. faecalis, Leuconostoc and Lactococcus, while genes for group II collagen adhesins were only present in W. ceti NC36 and Lb. delbrueckii. The third group presented the genes for the third collagen produced by W. ceti strains. In the last group (group IV) E. faecium and Leuconostoc collagen adhesin genes clustered together, however a W. confusa LBAE C39-2 collagen adhesin gene also formed a single lineage (Figure 1). In general, the close relatedness of Weissella, Enterococcus, and Leuconostoc collagen adhesion genes at the level of the primary protein sequence may suggest the same evolutionary origin of these proteins in these bacteria. Further genome sequencing of other Weissella species would be required to analyze this relatedness with enterococci and leuconostocs in more depth. The presence of virulence determinants commonly present in enterococci also in weissellas and leuconostocs may suggest a horizontal gene transfer event from the Enterococcus genus, the latter of which indeed includes strains that are commonly present in many different habitats (Franz et al., 2003).
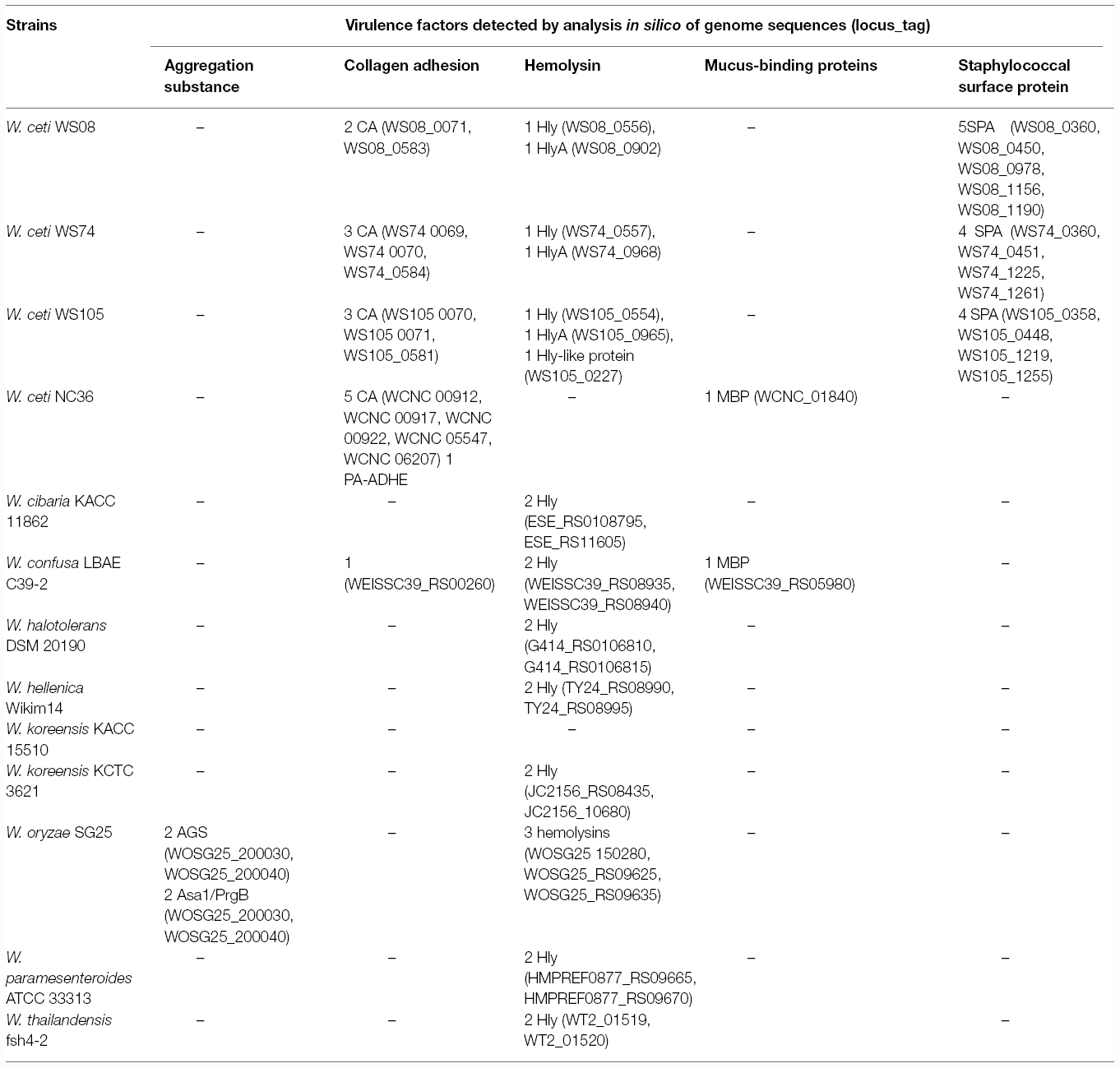
Table 2. Potential virulence genes of Weissella species as revealed by in silico screening of the annotated genome sequences.
The role of the genes above in the virulence of Weissella is still unknown. Further studies are needed to confirm whether these virulence genes are expressed or not, and if they are located on mobile genetic elements (e.g., transposons). However, the collagen adhesin protein (Ace) in Enterococcus was reported to be involved in adhesion to collagen and to contribute to infective endocarditis and urinary tract infection (Aymerich et al., 2006; Nallapareddy et al., 2011). In addition, the common localization of this gene on plasmids or transposon may pose a risk related with horizontal gene transfer to pathogenic bacteria. On the other hand, adherence is an important pre-requisite for the colonization of probiotics, providing a competitive advantage in different ecosystems. The presence of adhesin genes in weissellas that are not located on mobile genetic elements, and which are not transferable, should therefore not negatively reflect on the strains probiotic potential, and should not be an excluding factor for its use. It could be regarded as a key factor for the attachment of probiotic bacterial cell in the human host, and thus could be regarded as a colonization factor rather than a virulence factor (Dunne et al., 2004).
In silico analyses of Weissella genomes also showed the presence of genes coding for two unnamed aggregation substances (WOSG25_050600 and WOSG25_190240) as well as two aggregation substances named Asa1/PrgB (WOSG25_200030 and WOSG25_200040) in W. oryzae SG25 (Table 2). The sequences of the genes encoding the unnamed aggregation substances, the aggregation substances Asa1/PrgB from W. oryzae SG25 and those of other LAB (Leuconostoc and Enterococcus) were aligned using the CLUSTAL W of Lasergene program, and the most parsimonious phylogenetic tree was constructed. The results obtained showed that the aggregation substance genes found in W. oryzae SG25 clustered with those of Ln. pseudomesenteroides and E. faecalis (group I in dendrogram), the latter being the aggregation substance gene of E. faecalis pMG2200, which is located on a plasmid. This suggests that horizontal gene transfer from E. faecalis to other LAB such as Weissella and Leuconostoc (Figure 2A) probably occurred. Other aggregation substance genes of different strains of E. faecalis, Ln. mesenteroides, and Ln. pseudomesenteroides were very divergent, forming several clusters (Figure 2A). When aggregation substance gene sequences of Asa1/PrgB from W. oryzae SG25, E. faecalis, and Ln. pseudomesenteroides were clustered, the dendrogram showed aggregation substances Asa1/PrgB coding genes from W. oryzae SG25 and E. faecalis or Leuconostoc (Group I) to cluster very closely, also suggesting a horizontal gene transfer between these bacteria (Figure 2B). The presence of an asa1 gene was also reported in other potential probiotic LAB, such as Lb. casei SJRP35, Ln. citreum SJRP44, and Ln. mesenteroides subsp. mesenteroides SJRP58 isolated from Water-Buffalo Mozzarella Cheese (Jeronymo-Ceneviva et al., 2014), and also in Lc. lactis subsp. Lactis KT2W2L (Hwanhlem et al., 2013). As for adhesins, aggregation substances are considered virulence factors. However, the safety assessment of strains harboring adhesins or aggregation substances may highly depend on the localization of such genes on mobile genetic elements and on other safety aspects of the strain. However, the difference in infectivity between weissellas that are either probiotic or involved in infections such as endocarditis, cannot be explained by differences in adherence potential alone, but other factors involved in promoting disease must exist in pathogenic strains, as was similarly postulated by Vankerckhoven et al. (2007) for lactobacilli involved in human infections.
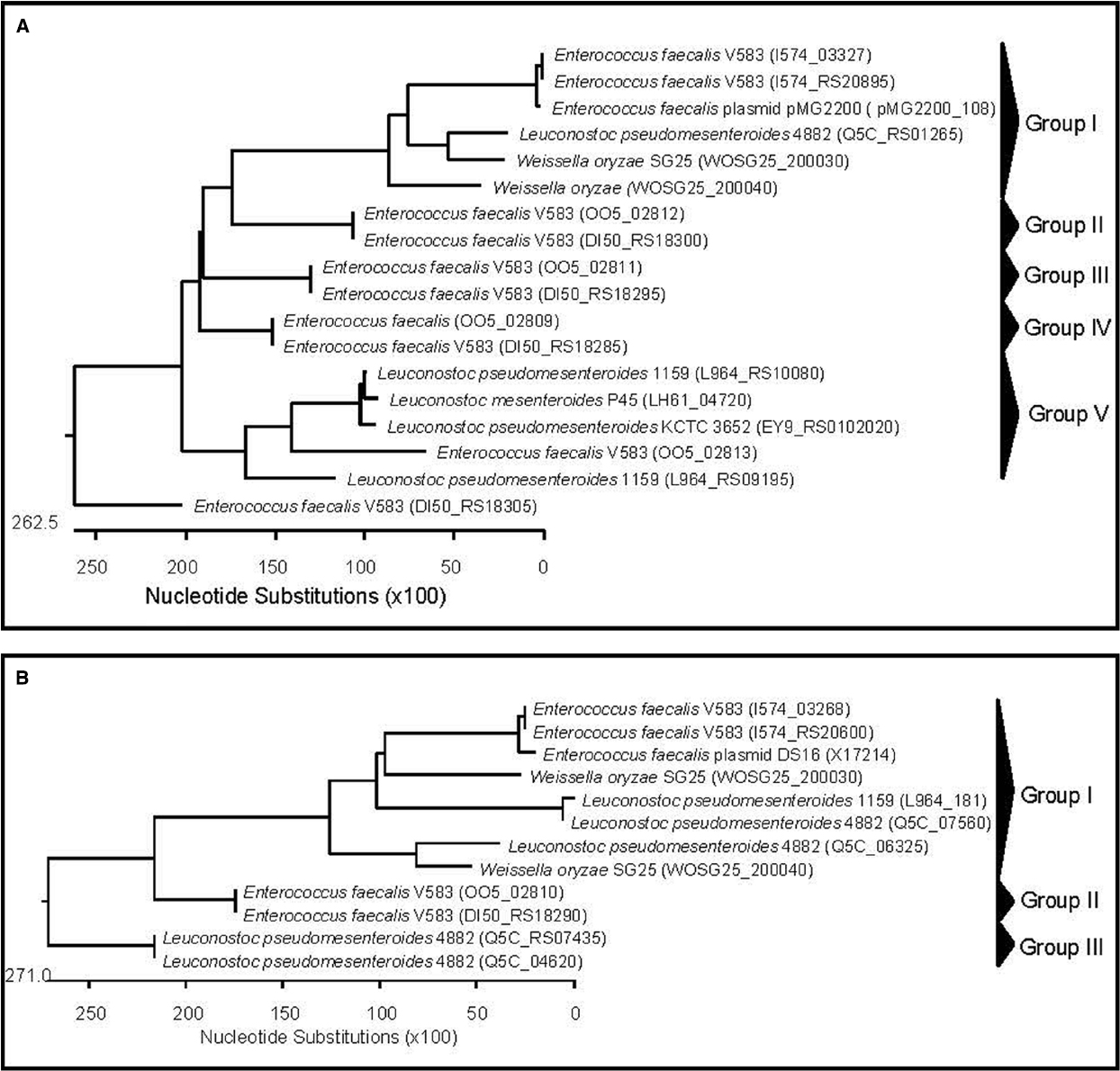
Figure 2. Phylogenetic relationships of Weissella species and Leuconostoc Enterococcus inferred from the alignment of aggregation substances (A) and aggregation substance Asa1/PrgB (B) encoding genes. The sequences were aligned and the most parsimonious phylogenetic trees were constructed using the CLUSTAL W of Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). The scale below indicates the number of nucleotide substitutions. Accession numbers are indicated in parentheses.
Genes for mucus-binding protein (Mub), which can serve as effector molecules involved in mechanisms of adherence of bacteria to the host, were detected in W. ceti NC36 and W. confusa LBAE C39-2 (Table 2) genome sequences. The phylogenetic tree of nucleotide sequences encoding mucus-binding proteins of weissellas, as well as of the closely related genera Leuconostoc, Lactobacillus, and Enterococcus, showed that the W. ceti NC36 mub gene clustered closely with Enterococcus sp. C1 mub in the same group together with sequences from Lb. delbrueckii and Leuconostoc. However, the mub of W. confusa LBAE C39-2 clustered with that of Lb. plantarum (Figure 3). The presence of mucus-binding protein may be a desirable feature in probiotic bacteria, as it may play an important role in the adhesion of the probiotic strain to host surfaces (Mack et al., 1999). However, this property is obviously problematic in potentially pathogenic strains.

Figure 3. Phylogenetic relationships of Weissella species and Leuconostoc Lactobacillus Enterococcus inferred from the alignment of mucus-binding protein encoding genes. The sequences were aligned and the most parsimonious phylogenetic trees were constructed using the CLUSTAL W of Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). The scale below indicates the number of nucleotide substitutions. Accession numbers are indicated in parentheses.
Regarding other virulence factors, such as the Serine-rich adhesin for platelets, which is also important in bacterial adhesion and is considered the central event in the pathogenesis of infective endocarditis (Sullam, 1994), this adhesin was only detected in the pathogen W. ceti NC36 (WCNC_RS02030; Table 2). On the other hand, the gene encoding the Serine-rich adhesin for platelets was detected also in probiotic lactobacilli such as Lb. johnsonii NCC533 and Lb. reuteri (Zhou and Wu, 2009), but not in leuconostocs. Concerning the staphylococcal surface protein A, which is involved in invasion and infectivity, several genes coding for a similar protein were detected in W. ceti WS08 (WS08_0360, WS08_0450, WS08_0978, WS08_1156, WS08_1190), W. ceti WS74 (WS74_0360, WS74_0451, WS74_1225, WS74_1261), and W. ceti WS105 (WS105_0358, WS105_0448, WS105_1219, WS105_1255). However, no homologous genes were detected in either leuconostocs or in lactobacilli (Table 2). Due to the low homology of virulence determinants in weissellas, it is difficult to detect such genes by PCR, thus genome sequencing would be the only way to detect and identify these determinants.
The presence of genes for hemolysin or hemolysin-like proteins in weissellas was revealed by in silico analysis of the annotated genomes sequences (Table 2). The ubiquitous occurrence of such genes in probiotic lactobacilli and bifidobacteria, and also in other LAB such as leuconostocs, may raise questions about their function and virulence potential. The role of these genes in LAB virulence is unknown, and their presence in the genome of a strain intended to be used as probiotic should not necessarily be an exclusion factor for targeting strains for probiotic use. As shown in Table 2, weissellas harbored in their genome 1–3 hemolysin encoding genes, i.e., genes for hemolysin A and also hemolysin-like proteins.
Studies on antibiotic resistance profiles in Weissella genus are very limited, and MIC breakpoints have not been defined by EFSA. Thus, to categorize weissellas as sensitive or resistant to different antibiotics of clinical relevance is a difficult task. These bacteria are known for their intrinsic resistance to antibiotics inhibiting cell wall biosynthesis such as vancomycin and fosfomycin, similar to other Gram-positive bacteria (Arca et al., 1997), and also to antibiotics that inhibit tetrahydrofolate biosynthesis such as sulfamethoxazol and trimethoprim (Liu et al., 2009). Furthermore, resistance to gentamicin, kanamycin, and norfloxacin was also reported in food-associated weissellas (Patel et al., 2012).
Reports about molecular detection of antibiotic resistance genes in weissellas are very scarce, possibly due to the high divergence of resistance genes. In this sense, only mef (A/E) drug efflux pump genes involved in the active efflux of macrolides (Clancy et al., 1996; Tait-Kamradt et al., 1997) were detected in W. cibaria of aquatic origin (Muñoz-Atienza et al., 2013). When we analyzed the in silico genome sequences of Weissella spp., different antibiotic resistance genes were detected, such as those coding for fosfomycin and methicillin resistance proteins in almost all the genomes of Weissella strains sequenced (Table 3) to date. For example, the fosB gene coding for fosfomycin resistance was detected in W. ceti WS08 (WS08_1256), W. ceti WS74 (WS74_1327) and W. ceti WS105 (WS105_1321), and nucleotide sequences of the genes were identical in all cases. In addition, other weissellas harbored multidrug transporters involved in fosfomycin and deoxycholate resistance, as was the case for W. ceti NC36 (WCNC_RS02205), W. cibaria KACC 11862 (ESE_RS0106205), W. confusa LBAE C39-2 (WEISSC39_RS10580), W. halotolerans DSM 20190 (G414_RS0105405), W. hellenica Wikim14 (TY24_RS09455), W. koreensis KCTC 3621 (JC2156_RS02465), W. oryzae SG25 (WOSG25_RS02065), and W. paramesenteroides ATCC 33313 (HMPREF0877_RS05895; Table 3). An alignment of nucleotide sequences encoding multidrug transporters involved in fosfomycin resistance of weissellas and other LAB by CLUSTAL W alignment showed sequences to cluster into two groups. High similarities in sequences again suggested an evolutionary relationship of weissellas, leuconostocs, enterococci, and lactobacilli (Figure 4) sequences. Fosfomycin is a broad antibacterial agent that targets several pathogens such as Haemophilus spp., Staphylococcus spp. and most of the enteric gram-negative bacteria. Although no cross resistances are known to occur for other antibiotics, there are no data on whether or not the fosB gene is transferable to other bacteria. Thus, weissellas harboring the fosB gene should be investigated in terms of the transferability of this gene, especially to pathogens. In the case of enterococci, these bacteria contain a conjugative plasmid that harbors the novel fosB transposon (ISL3-like transposon), as well as the Tn1546-like transposon (containing vanA and fosB, Qu et al. (2014)). Should weissellas harbor no transferable fosB gene, a potential probiotic use of these bacteria should be possible, especially when infection treatment with a combination of other antibiotics is possible.
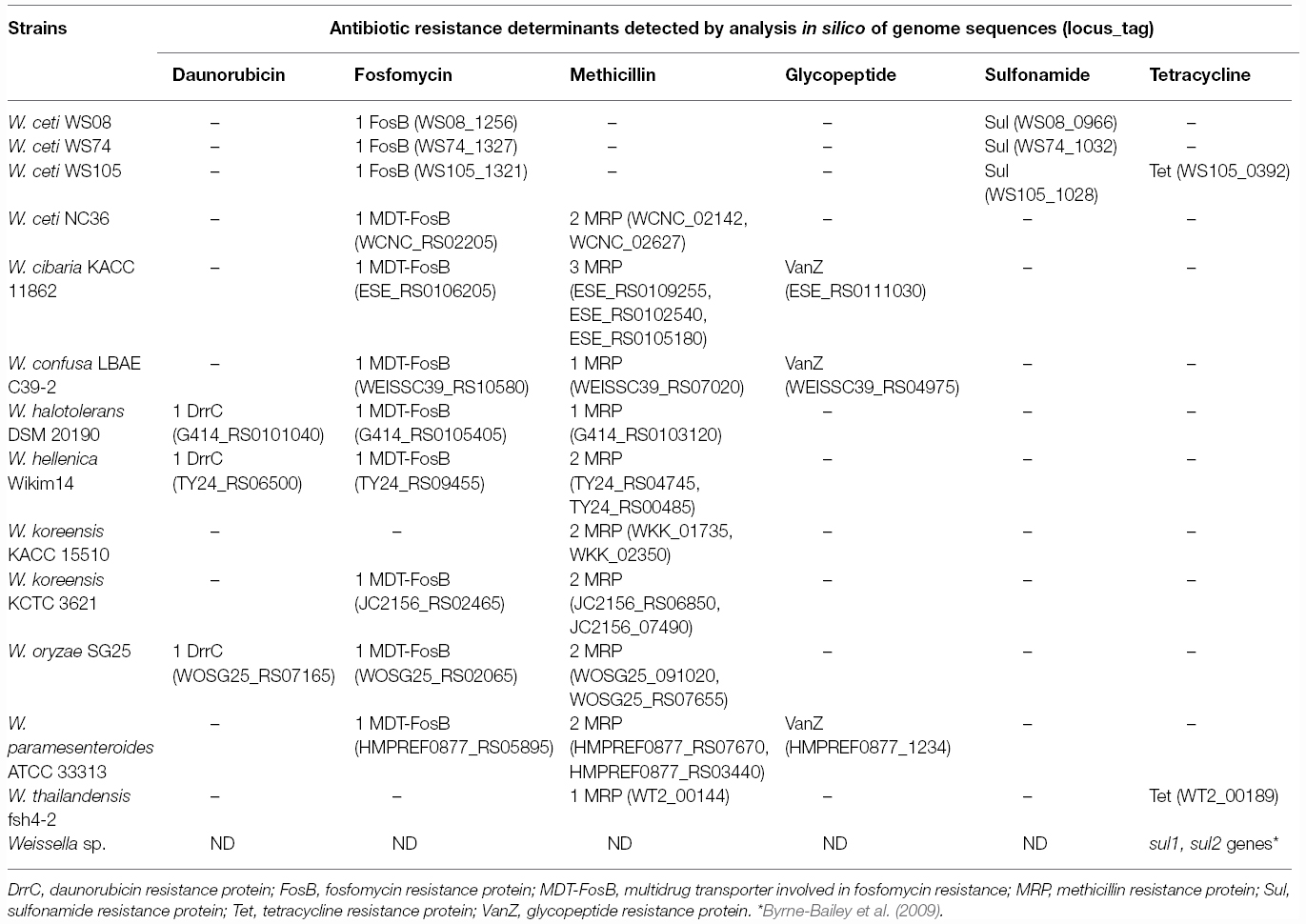
Table 3. Antibiotic resistance genes of Weissella species as revealed by in silico screening of the annotated genome sequences.

Figure 4. Phylogenetic relationships of Weissella species and Leuconostoc Lactobacillus Enterococcus inferred from the alignment of multidrug transporter involved in fosfomycin resistance encoding genes. The sequences were aligned and the most parsimonious phylogenetic trees were constructed using the CLUSTAL W of Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). The scale below indicates the number of nucleotide substitutions. Accession numbers are indicated in parentheses.
The intrinsic resistance of weissellas toward vancomycin may be attributed to the absence of D-Ala-D-lactate in their cell wall, which is the target of vancomycin. A similar situation exists in other LAB such as Lactobacillus, Pediococcus, and Leuconostoc species (Ammor et al., 2007). Thus, such resistance cannot be attributed to acquisition of resistance genes. Nevertheless, some weissellas harbored in their genomes the vanZ resistance gene, as is the case for W. cibaria KACC 11862 (ESE_RS0111030), W. confusa LBAE C39-2 (WEISSC39_RS04975), and W. paramesenteroides ATCC 33313 (HMPREF0877_1234). This gene confers resistance to teicoplanin and does not involve the incorporation of a substituent of D-alanine into the peptidoglycan precursors (Arthur et al., 1995; Table 3).
No data have been reported on methicillin resistance to date. However, analyzing the in silico genome sequences of weissellas available in NCBI database, methicillin resistance protein encoding genes were detected in all weissellas (sequenced genomes of nine species; Table 3). Methicillin resistance proteins are found in Staphylococcus spp. and especially in S. aureus, and the presence of genes encoding such proteins in Weissella species may suggest a horizontal gene transfer between the genera. The alignment of nucleotide sequences for methicillin resistance protein in weissellas and other Gram-positive bacteria (S. aureus, Leuconostoc, E. faecalis, and Lactococcus) clearly showed an evolutionary relationship between the methicillin-resistance sequences of Weissella and the other Gram-positive bacteria S. aureus, Leuconostoc, and E. faecalis (Figure 5). The dendrogram showed two main groups, in which the genes from Weissella spp. were distributed regardless of the species, strain or origin. A divergence of methicillin resistance genes could even be observed in a single strain of W. ceti NC36, which carried multiple methicillin resistance genes, varying considerably in nucleotide sequence (Figure 5). On the other hand, one of two genes encoding for methicillin resistance in W. cibaria KACC 11862 (ESE_RS0105180) formed a single lineage, as such being divergent from the other weissellas and other Gram-positive bacteria (Figure 5). Further studies should elucidate the functionality of the methicillin resistance genes of weissellas, and the transferability of these genes to other bacteria.
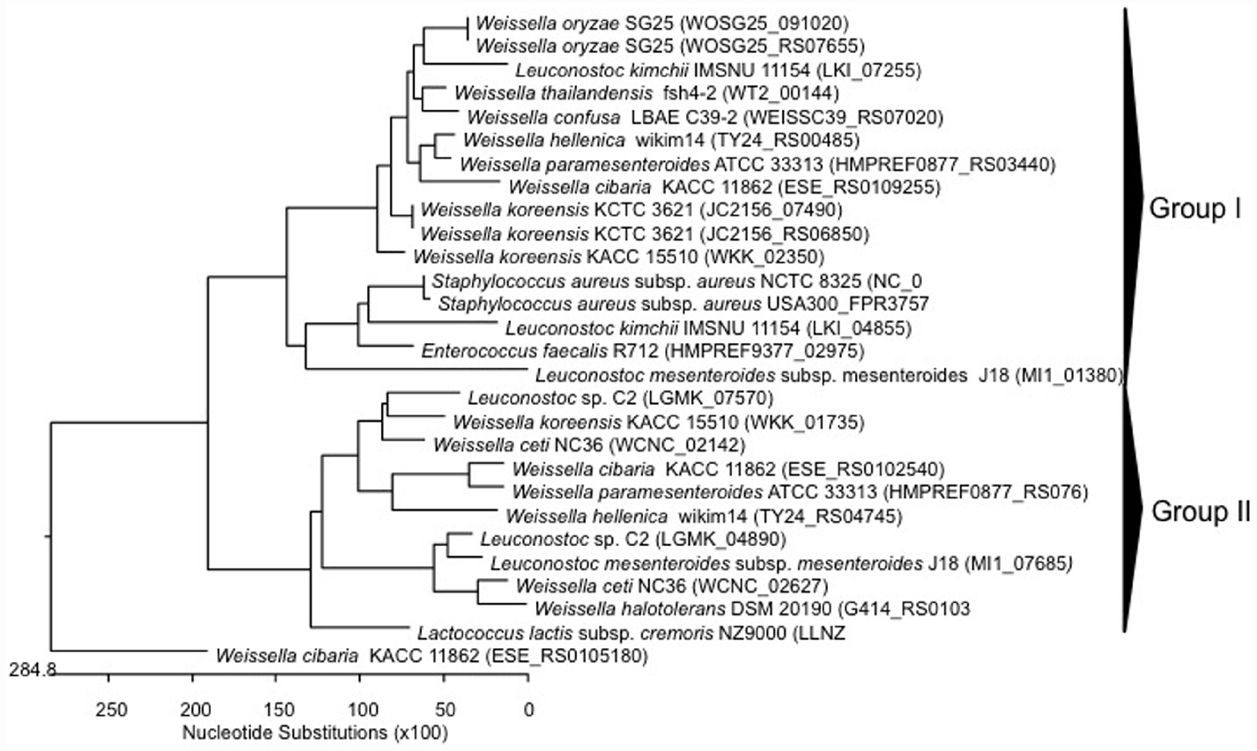
Figure 5. Phylogenetic relationships of Weissella species and Leuconostoc Lactobacillus Enterococcus inferred from the alignment of methicillin resistance protein encoding genes. The sequences were aligned and the most parsimonious phylogenetic trees were constructed using the CLUSTAL W of Lasergene program, version 5.05 (MegAlign, Inc., Madison, WI, USA). The scale below indicates the number of nucleotide substitutions. Accession numbers are indicated in parentheses.
Sulfonamide resistance could be due either to mutation in the chromosomal gene that mediates dihydropteroate synthesis, which is a folic acid precursor, or to the acquisition of resistance genes coding for resistant forms of the enzyme (Franklin and Snow, 2005). Resistance of weissellas to sulfonamides was reported by Byrne-Bailey et al. (2009) in Weissella spp. isolated from un-amended pig slurry and relied on the presence of sul1 and sul2 resistance genes. Figueiredo et al. (2012) isolated sulfonamide-resistant W. ceti that caused outbreaks characterized by acute haemorrhagic septicaemia and high mortality rates in rainbow trout. However, no genetic elements responsible for this resistance were described. Weissella genomes investigated in this study showed the presence of sulfonamide-resistance genes in W. ceti strains WS08 (WS08_0966), WS74 (WS74_1032), and WS105 (WS105_1028) isolated from rainbow trout (Figueiredo et al., 2012). Overall, resistance to vancomycin, fosfomycin, and sulfonamides appears to be a common trait in the genera Weissella and Leuconostoc. On the other hand, it is frequent that bacterial resistance to sulfonamides often co-exists with trimethoprim resistance, since this substance is used in combination with sulfonamides to minimize bacterial resistance. The known dfr trimethoprim resistance genes, which are usually associated with integrons (Skold, 2001) could, however, not be detected in any of the Weissella genome sequences analyzed here. Thus, the reported resistance of W. confusa (Fairfax et al., 2014; Medford et al., 2014) to trimethoprim may be caused by other modifications than those produced in the target enzyme dihydrofolate reductase (Dfr) that is encoded by the dfr-genes.
Weissellas are generally susceptible toward tetracycline. Screening of the published genomes revealed the presence of a gene encoding tetracycline (class C) resistance in W. ceti WS105 (WS105_0392) and in W. thailandensis fsh4-2 (WT2_00189; Table 3).
It is frequent to find resistance to the most widely used chemotherapeutic agent daunorubicin in probiotic bacteria such as lactobacilli (Lb. acidophilus NCFM, Lb. casei BD11) and bifidobacteria (Bifidobacterium animalis subsp. lactis Bb-12, Bifidobacterium longum subsp. infantis ATCC 15697). When the annotated genome sequences of weissellas were screened for daunorubicin resistance genes, ABC transporter-based resistance genes could be found in several Weissella spp. [W. halotolerans DSM 20190 (G414_RS0101040), W. hellenica Wikim14 (TY24_RS06500), and W. oryzae SG25 (WOSG25_RS07165)] (Table 3).
Antibiotic resistance is often based on unspecific mechanisms such as efflux pumps, which are widespread throughout evolution in different bacteria and can pump a variety of drugs out the cells (Piddock, 2006). When we screened the annotated genome sequences of Weissella spp., several efflux pumps were detected, such as multiple antibiotic resistance protein MarR, multidrug resistance SMR family protein, MFS multidrug transporter, ABC transporter, and a DedA family protein, which was recently shown to be associated with antibiotic resistance (Kumar and Doerrler, 2014).
Based on our genomic in silico investigation, the potential deleterious and beneficial effects for each of the Weissella species in view of their application as starter cultures or probiotics are discussed below, highlighting the controversial issue of strains of the same species being industrially important in some cases, while being problematic from a safety point of view in others.
Weissella ceti (Vela et al., 2011) was isolated from different organs of beaked whales (Mesoplodon bidens) such as the spleen, kidney, muscle, brain, and lymph. Several strains of W. ceti were reported as pathogens in fish, such as W. ceti WS08, W. ceti WS74, and W. ceti WS105 from farmed rainbow trout in Brazil (Figueiredo et al., 2014; Costa et al., 2015), and W. ceti NC36 from farmed rainbow trout in the United States (Ladner et al., 2013). This suggests that weissellosis is a rapidly emerging disease in farmed rainbow trout in different geographical locations. Recently, Welch and Good (2013) reported that Weissella spp. closely related with W. ceti strains on the basis of their 16S rRNA gene sequences (99% identity) from Chinese and Brazilian out breaks were involved in mortality of farmed rainbow trout in the USA. On the other hand, no reports were found on the application of W. ceti strains as starter cultures or as probiotics. In this study it was found that W. ceti genomes harbored several virulence factors and antibiotic resistance genes, which point toward a pathogenic potential. The industrial application of such strains thus seems problematic and should be considered carefully on a strain to strain basis in the background of a detailed safety evaluation for the presence of antibiotic resistances and virulence determinants.
Originally isolated from Thai fermented foods (Björkroth et al., 2002), W. cibaria was also isolated from other sources such as sourdough, fermented milk, cheese, fermented vegetables, fermented fish and meat, and also silage (Srionnual et al., 2007; reviewed by Fusco et al., 2015). However, clinical samples have also been a source of W. cibaria (Björkroth et al., 2002), with these bacteria being found in human saliva and the vagina, in human and animal feces and milk, and also in human blood and urine (reviewed by Fusco et al., 2015). Weissella cibaria has been targeted for use as starter culture in foods for different purposes such as for probiotic effects. In this sense, the dextran produced by dextransucrase from W. cibaria JAG8 had potential prebiotic effect for health benefits, stimulating the growth of probiotic bacteria (Tingirikari et al., 2014). Other beneficial effect of W. cibaria were suggested to be its capacity to produce higher levels of exopolysaccharides (EPS), which indicates more acid resistance, thus improving its probiotic capacity during passage throughout gastrointestinal tract (Park et al., 2013). Potentially probiotic W. cibaria strains were isolated from kimchi and also from goat milk (Elavarasi et al., 2014), and some strains were suggested to have anticancer activity, immune modulating activity, anti-inflammatory activity, antioxidant activity (patent related to the W. cibaria’s anticancer activity registered by Cha et al., 2008; Kang et al., 2011; Kwak et al., 2014), antiviral activity (the avian influenza virus; Rho et al., 2009) and anti-obesity effects being suggested to be more effective than the well-known probiotic bacterium Lb. rhamnosus GG (LGG; Ahn et al., 2013). It was also reported that the water-soluble polymers produced by W. cibaria inhibited biofilm formation by Streptococcus mutans, and thus production of such compounds could reduce oral plaque formation by approximately 20.7% in vivo and in vitro (Kang et al., 2006). Furthermore, the same authors reported that the hydrogen peroxide produced by W. cibaria inhibited the growth of the bacterial agent of periodontal disease, Fusobacterium nucleatum, and was effective in reducing the production of hydrogen sulfide and methanethiol responsible for the associated foul smell (Kang et al., 2006).
Several reports proposed different strains of W. cibaria as starter cultures in different fermentations, and these were based on its antimicrobial capacity. W. cibaria strains produce a variety of antagonistic substances, including organic acids and bacteriocins (i.e., weissellicin 110 produced by W. cibaria 110; Srionnual et al., 2007), as mentioned above. Wolter et al. (2014) reported that exopolysaccharide-producing W. cibaria MG1 might be a suitable starter culture for sourdough fermentation of buckwheat, quinoa and teff flour. W. cibaria was successfully tested in a defined semi-liquid sourdough starter (Gaggiano et al., 2007; Ricciardi et al., 2009), and due to its ability to grow at 45°C and produce EPS, which could be used as an alternative to additives for conditioning the textural properties of bread (Di Cagno et al., 2006), it was considered to be a suitable strain for this application. Weissella cibaria could also play an important role in meat fermentation. Thongsanit et al. (2009) proposed W. cibaria MSS2 as starter culture for the production of nham fermented sausage and for kimchi, due to the capacity of these bacteria to produce glutaminase, which is indispensable in food processing. Furthermore, the use of W. cibaria as starter for food fermentations promoted the formation of ornithine from arginine, which in turn may have beneficial health effects, such as anti-obesity effect due to high levels of ornithine in fermented foods (Yu et al., 2009).
On the other hand, W. cibaria was reported as an emerging pathogen being associated with bacteremia (Kulwichit et al., 2008) and dog ear otitis (Björkroth et al., 2002), and also as food spoilage organism in sliced vacuum-packed cooked ham (Han et al., 2010). In the present study, we showed that W. cibaria KACC 11862 harbored in its genome some virulence determinants (hemolysins) and antibiotic resistance genes (fosfomycin, methicillin, and glycopeptide; Tables 2–3). This argues for a detailed investigation on the virulence potential for this species on a strain basis.
Due to its previous confusion with viridans streptococci, most of infections caused by W. confusa were underestimated due to the misidentification of this bacterium by commercial identification systems. When the genus Weissella was described by Collins et al. (1993), it was shown that W. confusa played an important role in human and animal sepsis and also bacteremia (Green et al., 1990; Olano et al., 2001; Björkroth et al., 2002; Flaherty et al., 2007; Shin et al., 2007; Salimnia et al., 2010, 2011; Harlan et al., 2011; Kumar et al., 2011; Lee et al., 2011; Fairfax and Salimnia, 2012), thumb abscess (Bantar et al., 1991), endocarditis (Shin et al., 2007), osteomyelitis (Kulwichit et al., 2007), and recently it was also shown to be involved in infection of a prosthetic joint (Medford et al., 2014). Here, in silico analysis of the W. confusa LBAE C39-2 genome showed the presence of virulence determinant genes (encoding collagen adhesion, hemolysin, and mucus-binding proteins) and antibiotic resistance genes (fosfomycin, methicillin, and glycopeptide; Tables 2–3).
On the other hand, due to the widespread use of W. confusa in fermented foods (sourdough, cereals, vegetables, fermented milk, cheese; reviewed by Fusco et al., 2015), it was proposed as a starter culture, and also as probiotic which provides various beneficial health effects such as the inhibition of Helicobacter pylori, a bacterium that causes chronic inflammation and ulcers in the stomach (Nam et al., 2002). Furthermore, W. confusa strains isolated from human feces were proposed as potential probiotics (Lee et al., 2012). Other W. confusa strains (UI006 and UI007) isolated from traditional dairy foods from Nigeria were proposed as adjunct cultures for the dairy manufacture industry, because of their antagonistic activities against entero- and uro-pathogens (organic acids, ethanol, and hydrogen peroxide), and their lack of toxic compounds (Ayeni et al., 2009, 2011). In this regard Yang (2013) proposed W. confusa LK4 isolated from leek kimchi as a functional starter culture for fermentation of leeks. Due to the aforementioned role in human infections, the biotechnological use of W. confusa should also be carefully assessed on a strain to strain basis.
These bacteria isolated from a Korean fermented food “kimchi” (Lee et al., 2002) was used as starter culture in kimchi fermentations and production of functional foods, since W. koreensis OK1-6 has anti-obesity effects in high-fat diet (HF) induced obese mice (Park et al., 2012). Moon et al. (2012) reported the same anti-obesity effect for W. koreensis OK1-6, which produced ornithine from arginine, implying its functional role in reducing obesity. Also, Pi et al. (2014) showed that W. koreensis 521 was able to prevent and suppress obesity via the inhibition of pre-adipocyte mitogenesis and differentiation. On the other hand, the application of W. koreensis as starter culture together with Ln. citreum and baker’s yeast in sourdough to make whole wheat bread was also considered as successful, since a good texture and an extended shelf-life of bread could be obtained (Choi et al., 2012). The use of W. koreensis as starter culture in broken rice was also found to provide good organoleptic properties to jeungpyeon, which is a Korean fermented rice cake (Choi et al., 2013). Weissella koreensis was also proposed for use as probiotic in pigs, since dietary supplementation with W. koreensis WKG2 in growing pigs could improve the average daily gain (ADG) and could have a beneficial effect on the immune response during an inflammatory challenge (Wang et al., 2014). On the other hand, no negative effects on heath were yet attributed to this species. However, in our in silico genome investigation, we showed the presence of potential virulence determinants and antibiotic resistance genes in some strains of W. koreensis (Tables 2–3), again indicating that a safety assessment for strains targeted for biotechnological use should be done.
Regarding their health-promoting activities, several species of Weissella exhibited beneficial effects. W. kimchii is a hydrogen peroxide-producing species that has been proposed as probiotic to prevent vaginal infections against Candida albicans, Escherichia coli, S. aureus, and Streptococcus agalactiae (Lee, 2005; Lee et al., 2011). W. hellenica was isolated from different sources such as fermented vegetables, fermented sausage, fermented fish, cheeses, cow’s milk and flounder intestine (Leong et al., 2013; reviewed by Fusco et al., 2015), and is known for its potential probiotic activity due to its bacteriocin-producing capacity (weissellicins D, L, M, and Y). Strains from this species are active against several pathogens (Masuda et al., 2011; Leong et al., 2013; Chen et al., 2014) and have potential also as bio-protective culture to improve safety and shelf-life of foods like tofu (Chen et al., 2014). W. hellenica strains isolated from different sources (cheese, fermented sausage, fermented vegetables, cow’s milk, flounder intestine; reviewed by Fusco et al., 2015) were also reported as glucan (EPS) producers (Kim et al., 2008), which may have industrial applications. Finally, W. paramesenteroides isolated from soil, vegetables, cheese, fermented sausage, fermented sea food, fermented vegetables and also from feces, cow’s milk, and gut of rainbow (reviewed by Fusco et al., 2015) can have antibacterial activity by production of bacteriocins such as weissellin A from W. paramesenteroides DX (Papagianni and Papamichael, 2011), a bacteriocin from W. paramesenteroides DFR-8 (Pal and Ramana, 2010), and also non-proteinaceous antibacterial compounds (Pal and Ramana, 2009) with a broad inhibitory spectrum against Gram-positive and Gram-negative bacteria. These strains may therefore be interesting for their use as food biopreservatives. On the detrimental side, Han et al. (2010) showed that W. paramesenteroides was involved in sliced vacuum-packed cooked ham spoilage.
Due to their heterofermentative metabolism, some weissellas are also involved in food spoilage and lead to sensory defects of, e.g., meat products (Marsden et al., 2009). In this regards, W. viridescens plays an important role in meat spoilage, producing a greenish slime on meat surfaces as a result of its production of H2O2. In cooked hams, formation of cavities can result from the production of CO2 by these bacteria. Furthermore, taking into account data presented here from in silico analyses of Weissella spp. genomes, the presence of virulence determinants, especially hemolysins, were detected in all strains analyzed. Also antibiotic resistance genes, such as those coding for fosfomycin and methicillin resistances (Tables 2–3), occur in this species. Also in this case, therefore, the safety of strains intended for industrial use should be investigated for each strain in detail.
Considering the large number of health-promoting benefits which could arise from the use of strains of Weissella spp., such as antibacterial, anti-viral, anti-tumoral, anti-obesity, anti-inflammatory, and antioxidant activities, several weissellas could be targeted for use as starter cultures or probiotics. By contrast, specific strains of Weissella species have also been involved as pathogens in the etiology of different diseases such as bacteremia, endocarditis, sepsis, and may even cause mortality. In fact, the safety of this genus has not been deeply studied, as only some strains are considered as opportunistic pathogens. Thus, the application of strains in foods and feeds or for humans as probiotics should be done with caution. In this report we showed that screening of genome sequences revealed the presence of several virulence and antibiotic resistance genes, which could be the basis of the potential pathogenicity of some strains. However, the presence of single determinants should not be an exclusion criterium for weissellas that may have overall beneficial effects. Thus, selection of weissellas intended to be used as starters or as probiotics should be investigated carefully regarding their safety aspects, preferably by genome sequencing an annotation, since the heterogeneity in nucleotide sequences of well-known virulence and antibiotic resistance-genes make these undetectable by PCR methods. Generally, the application of W. confusa and W. cibaria strains as starter cultures or as probiotics should be approached with caution, carefully selecting strains which lack pathogenic potential and which do not possess transferable antibiotic resistance genes.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge research grants AGL2013-43571-P (Ministerio de Economía y Competitividad, MINECO, FEDER) and UJA2014/07/02 (Plan Propio UJA) and research group AGR230.
Ahn, S. B., Park, H. E., Lee, S. M., Kim, S. Y., Shon, M. Y., and Lee, W. K. (2013). Characteristics and immuno-modulatory effects of Weissella cibaria JW15 isolated from Kimchi, Korea traditional fermented food, for probiotic use. J. Biomed. Res. 14, 206–211. doi: 10.12729/jbr.2013.14.4.206
Alkema, W., Boekhorst, J., Wels, M., and van Hijum, S. A. F. T. (2015). Microbial bioinformatics for food safety and production. Brief. Bioinform. doi: 10.1093/bib/bbv034 [Epub ahead of print].
Ammor, M. S., Florez, A. B., and Mayo, B. (2007). Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 24, 559–570. doi: 10.1016/j.fm.2006.11.001
Arca, P., Reguera, G., and Hardisson, C. (1997). Plasmid-encoded fosfomycin resistance in bacteria isolated from the urinary tract in a multicentre Surrey. J. Antimicrob. Chemother. 40, 393–399. doi: 10.1093/jac/40.3.393
Arthur, M., Depardieu, F., Molinas, C., Reynolds, P., and Courvalin, P. (1995). The vanZ gene of Tn1546 from Enterococcus faecium BM4147 confers resistance to teicoplanin. Gene 154, 87–92. doi: 10.1016/0378-1119(94)00851-I
Ayeni, F. A., Adeniyi, B. A., Ogunbanwo, S. T., Tabasco, R., Paarup, T., Peláez, C., et al. (2009). Inhibition of uropathogens by lactic acid bacteria isolated from dairy foods and cow’s intestine in western Nigeria. Arch. Microbiol. 191, 639–648. doi: 10.1007/s00203-009-0492-9
Ayeni, F. A., Adeniyi, B. A., Ogunbanwo, S. T., Nader-Macias, M. E., and Ruas-Madiedo, P. (2011). Survival of Weissella confusa and Lactobacillus paracasei strains in fermented milks under cold storage and after freeze-drying. Milchwissenschaft 66, 61–64.
Aymerich, T., Martin, B., Garriga, M., Vidal-Carou, M. C., Bover-Cid, S., and Hugas, M. (2006). Safety properties and molecular strain typing of lactic acid bacteria from slightly fermented sausages. J. Appl. Microbiol. 100, 40–49. doi: 10.1111/j.1365-2672.2005.02772.x
Bantar, C. E., Relloso, S., Castell, F. R., Smayevsky, J., and Bianchini, H. M. (1991). Abscess caused by vancomycin-resistant Lactobacillus confusus. J. Clin. Microbiol. 29, 2063–2064.
Björkroth, K. J., Schillinger, U., Geisen, R., Weiss, N., Hoste, B., Holzapfel, W. H., et al. (2002). Taxonomic study of Weissella confusa and description of Weissella cibaria sp. nov., detected in food and clinical samples. Int. J. Syst. Evol. Microbiol. 52, 141–148. doi: 10.1099/00207713-52-1-141
Byrne-Bailey, K. G., Gaze, W. H., Kay, P., Boxall, A. B. A., Hawkey, P. M., and Wellington, E. M. H. (2009). Prevalence of sulfonamide resistance genes in bacterial isolates from manured agricultural soils and pig slurry in the United Kingdom. Antimicrob. Agents Chemother. 53, 696–702. doi: 10.1128/AAC.00652-07
Cai, Y., Benno, Y., Nakase, T., and Oh, T. K. (1998). Specific probiotic characterization of Weissella hellenica DS-12 isolated from flounder intestine. J. Gen. Appl. Microbiol. 44, 311–316. doi: 10.2323/jgam.44.311
Cha, S. K., Ahn, B. H., and Kim, J. R. (2008). Korea Food Research Institute, Assignee Weissella cibaria 148-2 Lactic Bacteria for Functional Healthy Effect and Makgeolli Containing the Same. Korea patent KR 1020080133488. Inventors Probiotic Properties of Lactic Acid Bacteria Isolated from Water-Buffalo Mozzarella Cheese.
Chen, C., Rui, X., Lu, Z., Li, W., and Dong, M. (2014). Enhanced shelf-life of tofu by using bacteriocinogenic Weissella hellenica D1501 as bioprotective cultures. Food Control 46, 203–209. doi: 10.1016/j.foodcont.2014.05.004
Choi, H., Kim, Y. W., Hwang, I., Kim, J., and Yoon, S. (2012). Evaluation of Leuconostoc citreum HO12 and Weissella koreensis HO20 isolated from kimchi as a starter culture for whole wheat sourdough. Food Chem. 134, 2208–2216. doi: 10.1016/j.foodchem.2012.04.047
Choi, H., Ok Park, J., and Yoon, S. (2013). Fermentation of broken rice using kimchi-derived Weissella koreensis HO20 and its use in Jeungpyeon. Food Sci. Biotechnol. 22, 1275–1283. doi: 10.1007/s10068-013-0213-7
Choi, H. J., Cheigh, C. I., Kim, S. B., Lee, J. C., Lee, D. W., Choi, S. W., et al. (2002). Weissella kimchii sp. nov., a novel lactic acid bacterium from kimchi. Int. J. Syst. Evol. Microbiol. 52, 507–511. doi: 10.1099/00207713-52-2-507
Clancy, J., Petitpas, J., Dib-Hajj, F., Yuan, W., Cronan, M., Kamath, A. V., et al. (1996). Molecular cloning and functional analysis of a novel macrolide-resistance determinant, mefA, from Streptococcus pyogenes. Mol. Microbiol. 22, 867–879. doi: 10.1046/j.1365-2958.1996.01521.x
Collins, M. D., Samelis, J., Metaxopoulos, J., and Wallbanks, S. (1993). Taxonomic studies on some Leuconostoc-like organisms from fermented sausages: description of a new genus Weissella for the Leuconostoc paramesenteroides group of species. J. Appl. Bacteriol. 75, 595–603. doi: 10.1111/j.1365-2672.1993.tb01600.x
Collins, M. D., Samelis, J., Metaxopoulos, J., and Wallbanks, S. (1994). Validation of the publication of new names and new combinations previously effectively published outside the IJSB List No. 49. Int. J. Syst. Bacteriol. 44, 370–371. doi: 10.1099/00207713-44-2-370
Comi, G., and Iacumin, L. (2012). Identification and process origin of bacteria responsible for cavities and volatile of flavour compounds in artisan cooked ham. Int. J. Food Sci. Tech. 47, 114–121. doi: 10.1111/j.1365-2621.2011.02816.x
Costa, F. A. A., Leal, C. A. G., Schuenker, N. D., Leite, R. C., and Figueiredo, H. C. P. (2015). Characterization of Weissella ceti infections in Brazilian rainbow trout, Oncorhynchus mykiss (Walbaum), farms and development of an oil-adjuvanted vaccine. J. Fish Dis. 38, 295–302. doi: 10.1111/jfd.12236
Di Cagno, R., De Angelis, M., Limitone, A., Minervini, F., Carnevali, P., Corsetti, A., et al. (2006). Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. J. Agric. Food Chem. 54, 9873–9881. doi: 10.1021/jf061393+
Dunne, C., Kelly, P., O’Halloran, S., Soden, D., Bennett, M., von Wright, A., et al. (2004). Mechanisms of adherence of a probiotic Lactobacillus strain during and after in vivo assessment in ulcerative colitis patients. Microb. Ecol. Health Dis. 16, 96–104. doi: 10.1080/08910600410032295
Elavarasi, V., Pugazhendhi, A., Poornima Priyadharsani, T. K., Valsala, H., and Thamaraiselvi, K. (2014). Screening and characterization of Weissella cibaria isolated from food source for probiotic properties. Int. J. Comput. Appl. 1, 29–32.
European Food Safety Authority (EFSA). (2004). EFSA Scientific Colloquium Summary Report. Brussels: QPS
European Food Safety Authority (EFSA). (2015). Statement on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA. 2: suitability of (taxonomic) units notified to EFSA until March 2015. EFSA J. 13, e4138. doi: 10.2903/j.efsa.2015.4138
Fairfax, M. R., Lephart, P. R., and Salimnia, H. (2014). Weissella confusa: problems with identification of an opportunistic pathogen that has been found in fermented foods and proposed as a probiotic. Front. Microbiol. 5:254. doi: 10.3389/fmicb.2014.00254
Fairfax, M. R., and Salimnia, H. (2012). “Beware of unusual organisms masquerading as skin contaminants,” in Sepsis–An Ongoing and Significant Challenge, ed. L. Azevedo (Rijeka: Intech Open), 275–286.
Figueiredo, H. C., Costa, F. A., Leal, C. A., Carvalho-Castro, G. A., and Leite, R. C. (2012). Weissella sp. outbreaks in commercial rainbow trout (Oncorhynchus mykiss) farms in Brazil. Vet. Microbiol. 156, 359–366. doi: 10.1016/j.vetmic.2011.11.008
Figueiredo, H. C., Leal, C. A., Dorella, F. A., Carvalho, A. F., Soares, S. C., Pereira, F. L., et al. (2014). Complete genome sequences of fish pathogenic Weissella ceti Strains WS74 and WS105. Genome Announc. 2, 1014–1014. doi: 10.1128/genomeA.01014-14
Flaherty, J. D., Levett, P. N., Dewhirst, F. E., Troe, T. E., Warren, J. R., and Johnson, S. (2003). Fatal case of endocarditis due to Weissella confusa. J. Clin. Microbiol. 41, 2237–2239. doi: 10.1128/JCM.41.5.2237-2239.2003
Flaherty, J. D., Levett, P. N., Dewhirst, F. E., Troe, T. E., Warren, J. R., and Johnson, S. (2007). Fatal case of endocarditis due to Weissella confusa. J. Clin. Microbiol. 41, 2237–2239. doi: 10.1128/JCM.41.5.2237-2239.2003
Food and Drug Administration. (1999). Federal Food, Drug, and Cosmetic Act. Washington, DC: US Food and Drug Administration.
Franklin, T. J., and Snow, G. A. (2005). Biochemistry and Molecular Biology of Antimicrobial Drug Action, 6th Edn. New York, NY: Springer Science And Business Media Incorporated.
Franz, C. M. A. P., Stiles, M. E., Schleifer, K.-H., and Holzapfel, W. H. (2003). Enterococci in foods—a conundrum for food safety. Int. J. Food Microbiol. 88, 105–122. doi: 10.1016/S0168-1605(03)00174-0
Fusco, V., Quero, G. M., Cho, G., Kabisch, J., Meske, D., Neve, H., et al. (2015). The genus Weissella: taxonomy, ecology and biotechnological potential. Front. Microbiol. 6:155. doi: 10.3389/fmicb.2015.00155
Gaggiano, M., Di Cagno, R., De Angelis, M., Arnault, P., Tossut, P., Fox, P. F., et al. (2007). Defined multi-species semi-liquid ready-to-use sourdough starter. Food Microbiol. 24, 15–24. doi: 10.1016/j.fm.2006.04.003
Green, M., Wadowsky, R. M., and Barbadora, K. (1990). Recovery of vancomycin-resistant gram-positive cocci from children. J. Clin. Microbiol. 28, 484–488.
Han, Y., Xu, X., Yun Jiang, Y., Zhou, G., Sun, X., and Xu, B. (2010). Inactivation of food spoilage bacteria by high pressure processing: evaluation with conventional media and PCR–DGGE analysis. Food Res. Int. 43, 1719–1724. doi: 10.1016/j.foodres.2010.05.012
Harlan, N. P., Kempker, R. R., Parekh, S. M., Burd, E. M., and Kuhar, D. T. (2011). Weissella confusa bacteremia in a liver transplant patient with hepatic artery thrombosis. Transpl. Infect. Dis. 13, 290–293. doi: 10.1111/j.1399-3062.2010.00579.x
Hwanhlem, N., Biscola, V., El-Ghaish, S., Jaffrès, E., Dousset, X., Haertlé, T., et al. (2013). Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in Southern Thailand as potential bio-control agents: purification and characterization of bacteriocin produced by Lactococcus lactis subsp. lactis KT2W2L. Probiotics Antimicrob. Proteins 5, 264–278. doi: 10.1007/s12602-013-9150-2
Jeronymo-Ceneviva, A. B., de Paula, A. T., Silva, L. F., Todorov, S. D., Franco, B. D., and Penna, A. L. (2014). Probiotic properties of lactic acid bacteria isolated from water-buffalo mozzarella cheese. Probiotics Antimicrob Proteins 6, 141–156. doi: 10.1007/s12602-014-9166-2
Kang, M. S., Chung, J., Kim, S. M., Yang, K. H., and Oh, J. S. (2006). Effect of Weissella cibaria isolates on the formation of Streptococcus mutans biofilm. Caries Res. 40, 418–425. doi: 10.1159/000094288
Kang, M.-S., Lim, H.-S., Kim, S.-M., Lee, H.-C., and Oh, J.-S. (2011). Effect of Weissella cibaria on Fusobacterium nucleatum-induced interleukin-6 and interleukin-8 production in KB cells. J. Bacteriol. Virol. 41, 9–18. doi: 10.4167/jbv.2011.41.1.9
Kim, M. J., Seo, H. N., Hwang, T. S., Lee, S. H., and Park, D. H. (2008). Characterization of exopolysaccharide (EPS) produced by Weissella hellenica SKkimchi3 isolated from kimchi. J. Microbiol. 46, 535–541. doi: 10.1007/s12275-008-0134-y
Kulwichit, W., Nilgate, S., Chatsuwan, T., Krajiw, S., Unhasuta, C., and Chongthaleong, A. (2007). Accuracies of Leuconostoc phenotypic identification: a comparison of API systems and conventional phenotypic assays. BMC Infect. Dis. 7:69. doi: 10.1186/1471-2334-7-69
Kulwichit, W., Nilgate, S., Krajiw, S., Unhasuta, C., Chatsuwan, T., and Chongthaleong, A. (2008). “Weissella spp.: lactic acid bacteria emerging as a human pathogen,” in Proceedings of the 18th European Congress of Clinical Microbiology and Infectious Diseases, Barcelona, 19–22.
Kumar, A., Augustine, D., Sudhindran, S., Kurian, A. M., Dinesh, K. R., Karim, S., et al. (2011). Weissella confusa: a rare cause of vancomycin-resistant Gram-positive bacteraemia. J. Med. Microbiol. 60, 1539–1541. doi: 10.1099/jmm.0.027169-0
Kumar, S., and Doerrler, W. T. (2014). Members of the conserved DedA family are likely membrane transporters and are required for drug resistance in Escherichia coli. Antimicrob. Agents Chemother. 58, 923–930. doi: 10.1128/AAC.02238-13
Kwak, S.-H., Cho, Y.-M., Noh, G.-M., and Om, A.-S. (2014). Cancer preventive potential of kimchi lactic acid bacteria (Weissella cibaria, Lactobacillus plantarum). J. Cancer Prev. 19, 253–258. doi: 10.15430/JCP.2014.19.4.253
Ladner, J. T., Welch, T. J., Whitehouse, C. A., and Palacios, G. F. (2013). Genome sequence of Weissella ceti NC36, an emerging pathogen of farmed rainbow trout in the United States. Genome Announc. 1, e00187-12. doi: 10.1128/genomeA.00187-12
Lee, J. S., Lee, K. C., Ahn, J. S., Mheen, T. I., Pyun, Y. R., and Park, Y. H. (2002). Weissella koreensis sp. nov., isolated from kimchi. Int. J. Syst. Evol. Microbiol. 52, 1257–1261. doi: 10.1099/00207713-52-4-1257
Lee, K. W., Park, J. Y., Jeong, H. R., Heo, H. J., Han, N. S., and Kim, J. H. (2012). Probiotic properties of Weissella strains isolated from human faeces. Anaerobe 18, 96–102. doi: 10.1016/j.anaerobe.2011.12.015
Lee, M. R., Huang, Y. T., Liao, C. H., Lai, C. C., Lee, P. I., and Hsueh, P. R. (2011). Bacteraemia caused by Weissella confusa at a university hospital in Taiwan 1997–2007. Clin. Microbiol. Infect. 17, 1226–1231. doi: 10.1111/j.1469-0691.2010.03388.x
Lee, Y. (2005). Characterization of Weissella kimchii PL9023 as a potential probiotic for women. FEMS Microbiol. Lett. 250, 157–162. doi: 10.1016/j.femsle.2005.07.009
Leong, K. H., Chen, Y. S., Lin, Y. H., Pan, S. F., Yu, B., Wu, H. C., et al. (2013). Weissellicin L, a novel bacteriocin from sian-sianzih-isolated Weissella hellenica 4–7. J. Appl. Microbiol. 115, 70–76. doi: 10.1111/jam.12218
Liu, J. Y., Li, A. H., Ji, C., and Yang, W. M. (2009). First description of a novel Weissella species as an opportunistic pathogen for rainbow trout Oncorhynchus mykiss (Walbaum) in China. Vet. Microbiol. 136, 314–320. doi: 10.1016/j.vetmic.2008.11.027
Mack, D. R., Michail, S., Wei, S., McDougall, L., and Hollingsworth, M. A. (1999). Probiotics inhibit enteropathogenic Escherichia coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. 276, 941–950.
Marsden, J., Ahmed-Kotrola, N., Saini, J., Fung, D. Y. C., and Phebus, R. (2009). “Cooked meats, poultry and their products,” in Meat Products, ed. R. Fernandes (Leatherhead: Leatherhead Food International Ltd.), 53–82.
Masuda, Y., Zendo, T., Sawa, N., Perez, R. H., Nakayama, J., and Sonomoto, K. (2011). Characterization and identification of weissellicin Y and weissellicin M, novel bacteriocins produced by Weissella hellenica QU 13. J. Appl. Microbiol. 112, 99–108. doi: 10.1111/j.1365-2672.2011.05180.x
Medford, R., Patel, S. N., and Evans, G. A. (2014). A confusing case—Weissella confusa prosthetic joint infection: a case report and review of the literature. Can. J. Infect. Dis. Med. Microbiol. 25, 173–175.
Moon, Y. J., Soh, J. R., Yu, J. J., Sohn, H. S., Cha, Y. S., and Oh, S. H. (2012). Intracellular lipid accumulation inhibitory effect of Weissella koreensis OK1-6 isolated from kimchi on differentiating adipocyte. J. Appl. Microbiol. 113, 652–658. doi: 10.1111/j.1365-2672.2012.05348.x
Muñoz-Atienza, E., Gómez-Sala, B., Araújo, C., Campanero, C., del Campo, R., Hernández, P. E., et al. (2013). Antimicrobial activity, antibiotic susceptibility and virulence factors of lactic acid bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiol. 13:15. doi: 10.1186/1471-2180-13-15
Nallapareddy, S. R., Singh, K. V., Sillanpaa, J., Zhao, M., and Murray, B. E. (2011). Relative contributions of Ebp Pili and the collagen adhesion ace to host extracellular matrix protein adherence and experimental urinary tract infection by Enterococcus faecalis OG1RF. Infect. Immun. 79, 2901–2910. doi: 10.1128/IAI.00038-11
Nam, H., Ha, M., Bae, O., and Lee, Y. (2002). Effect of Weissella confusa strain PL9001 on the adherence and growth of Helicobacter pylori. Appl. Environ. Microbiol. 68, 4642–4645. doi: 10.1128/AEM.68.9.4642-4645.2002
Ogier, J.-C., and Serror, P. (2008). Safety assessment of dairy microorganisms: the Enterococcus genus. Int. J. Food Microbiol. 126, 291–301. doi: 10.1016/j.ijfoodmicro.2007.08.017
Olano, A., Chua, J., Schroeder, S., Minari, A., Lasalvia, M., and Hall, G. (2001). Weissella confusa (Basonym: Lactobacillus confusus) bacteremia: a case report. J. Clin. Microbiol. 39, 1604–1607. doi: 10.1128/JCM.39.4.1604-1607.2001
Pal, A., and Ramana, K. V. (2009). Isolation and preliminary characterization of a non bacteriocin antimicrobial compound from Weissella paramesenteroides DFR-8 isolated from cucumber (Cucumis sativus). Process. Biochem. 44, 499–503. doi: 10.1016/j.procbio.2009.01.006
Pal, A., and Ramana, K. V. (2010). Purification and characterization of bacteriocin from Weissella paramesenteroides dfr-8, an isolate from cucumber (Cucumis sativus). J. Food Biochem. 34, 932–948. doi: 10.1111/j.1745-4514.2010.00340.x
Papagianni, M., and Papamichael, E. M. (2011). Purification, amino acid sequence and characterization of the class IIa bacteriocin weissellin A, produced by Weissella paramesenteroides DX. Bioresour. Technol. 102, 6730–6734. doi: 10.1016/j.biortech.2011.03.106
Park, J. A., Tirupathi Pichiah, P. B., Yu, J. J., Oh, S. H., Daily, J. W. III, and Cha, Y. S. (2012). Anti-obesity effect of kimchi fermented with Weissella koreensis OK1-6 as starter in high-fat diet-induced obese C57BL/6J mice. J. Appl. Microbiol. 113, 1507–1516. doi: 10.1111/jam.12017
Park, J. H., Ahn, H. J., Kim, S. G., and Chung, C. H. (2013). Dextran-like exopolysaccharide-producing Leuconostoc and Weissella from kimchi and its ingredients. Food Sci. Biotechnol. 22, 1047–1053. doi: 10.1007/s10068-013-0182-x
Patel, A. R., Lindström, C., Patel, A., Prajapati, J. B., and Holst, O. (2012). Probiotic properties of exopolysaccharide producing lactic acid bacteria isolated from vegetables and traditional Indian fermented foods. Int. J. Ferment. Foods 1, 87–101.
Pi, K., Lee, K., Kim, Y., and Lee, E.-J. (2014). The inhibitory effect of Weissella koreensis 521 isolated from kimchi on 3T3-L1 adipocyte differentiation. Int. J. Med. Health Biomed. Pharm. Eng. 8, 7–10.
Piddock, L. J. (2006). Multidrug-resistance efflux pumps—not just for resistance. Nat. Rev. Microbiol. 4, 629–636. doi: 10.1038/nrmicro1464
Qu, T.-T., Shi, K.-R., Ji, J.-S., Yang, Q., Du, X.-X., Wei, Z.-Q., et al. (2014). Fosfomycin resistance among vancomycin-resistant enterococci owing to transfer of a plasmid harbouring the fosB gene. Int. J. Antimicrob. Agents 43, 361–365. doi: 10.1016/j.ijantimicag.2013.11.003
Rho, J. B., Poo, H., Choi, Y. K., Kim, C. J., and Sung, M. H. (2009). New Lactic Acid Bacteria Having its Inhibitory Effect on Avian Influenza Virus Infection and Composition Containing the Same. United States Patent US 20120156172 A1. Inventors.
Ricciardi, A., Parente, E., and Zotta, T. (2009). Modelling the growth of Weissella cibaria as a function of fermentation conditions. J. Appl. Microbiol. 107, 1528–1535. doi: 10.1111/j.1365-2672.2009.04335.x
Salimnia, H., Alangaden, G. J., Bharadwaj, R., Painter, T. M., Chandrasekar, P. H., and Fairfax, M. R. (2010). Weissella confusa: an unexpected cause of vancomycin-resistant Gram-positive bacteremia in immune compromised hosts. Transpl. Infect. Dis. 12, 526–528. doi: 10.1111/j.1399-3062.2010.00539.x
Salimnia, H., Alangaden, G. J., Bharadwaj, R., Painter, T. M., Chandrasekar, P. H., and Fairfax, M. R. (2011). Weissella confusa: an unexpected cause of vancomycin-resistant Gram-positive bacteremia in immunocompromised hosts. Transpl. Infect. Dis. 13, 294–298. doi: 10.1111/j.1399-3062.2010.00586.x
Santos, E. M., Jaime, I., Rovira, J., Lyhs, U., Korkeala, H., and Björkroth, J. (2005). Characterization and identification of lactic acid bacteria in “morcilla de Burgos”. Int. J. Food Microbiol. 97, 285–296. doi: 10.1016/j.ijfoodmicro.2004.04.021
Shin, J. H., Kim, D. I., Kim, H. R., Kim, D. S., Kook, J. K., and Lee, J. N. (2007). Severe infective endocarditis of native valves caused by Weissella confusa detected incidentally on echocardiography. J. Infect. 54, 149–151. doi: 10.1016/j.jinf.2006.09.009
Skold, O. (2001). Resistance to trimethoprim and sulfonamides. Vet. Res. 32, 261–273. doi: 10.1051/vetres:2001123
Srionnual, S., Yanagida, F., Lin, L. H., Hsiao, K. N., and Chen, Y. S. (2007). Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from Plaa-Som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 73, 2247–2250. doi: 10.1128/AEM.02484-06
Sullam, P. M. (1994). Host-pathogen interactions in the development of bacterial endocarditis. Curr. Opin. Infect. Dis. 4, 304–309. doi: 10.1097/00001432-199406000-00004
Tait-Kamradt, A., Clancy, J., Cronan, M., Dib-Hajj, F., Wondrack, L., Yuan, W., et al. (1997). mefE is necessary for the erythromycin-resistant M phenotype in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 41, 2251–2255.
Thongsanit, J., Tanikawa, M., Yano, S., Tachiki, T., and Wakayama, M. (2009). Identification of glutaminase-producing lactic acid bacteria isolated from Nham, a traditional Thai fermented food and characterisation of glutaminase activity of isolated Weissella cibaria. Ann. Microbiol. 59, 715–720. doi: 10.1007/BF03179213
Tingirikari, J. M., Kothari, D., Shukla, R., and Goyal, A. (2014). Structural and biocompatibility properties of dextran from Weissella cibaria JAG8 as food additive. Int. J. Food Sci. Nutr. 65, 686–691. doi: 10.3109/09637486.2014.917147
Vankerckhoven, V., Moreillon, P., Piu, S., Giddey, M., Huys, G., Vancanneyt, M., et al. (2007). Infectivity of Lactobacillus rhamnosus and Lactobacillus paracasei isolates in a rat model of experimental endocarditis. J. Med. Microbiol. 56, 1017–1024. doi: 10.1099/jmm.0.46929-0
Vela, A. I., Fernández, A., de Quirós, Y. B., Herráez, P., Domínguez, L., and Fernández-Garayzábal, J. F. (2011). Weissella ceti sp. nov., isolated from beaked whales (Mesoplodon bidens). Int. J. Syst. Evol. Microbiol. 61, 2758–2762. doi: 10.1099/ijs.0.028522-0
Wang, J. P., Yoo, J. S., Jang, H. D., Lee, J. H., Cho, J. H., and Kim, I. H. (2014). Effect of dietary fermented garlic by Weissella koreensis powder on growth performance, blood characteristics, and immune response of growing pigs challenged with Escherichia coli lipopolysaccharide. J. Anim. Sci. 89, 2123–2131. doi: 10.2527/jas.2010-3186
Welch, T. J., and Good, C. M. (2013). Mortality associated with weissellosis (Weissella sp.) in USA farmed rainbow trout: potential for control by vaccination. Aquaculture 388–391, 122–127. doi: 10.1016/j.aquaculture.2013.01.021
Wolter, A., Hager, A. S., Zannini, E., Galle, S., Gänzle, M., Waters, D. M., et al. (2014). Evaluation of exopolysaccharide producing Weissella cibaria MG1 strain for the production of sourdough from various flours. Food Microbiol. 37, 44–50. doi: 10.1016/j.fm.2013.06.009
Yang, J. S. (2013). Selection of functional lactic acid bacteria as starter cultures for the fermentation of Korean leek (Allium tuberosum Rottler). J. Prob. Health 1, 4. doi: 10.4172/2329-8901.S1.012
Yu, J. J., Park, H. J., Kim, S. G., and Oh, S. H. (2009). Isolation, identification, and characterization of Weissella strains with high ornithine producing capacity from kimchi. Korean J. Microbiol. 45, 339–345.
Keywords: Weissella, in silico analysis, genome, virulence, antibiotic resistance
Citation: Abriouel H, Lavilla Lerma L, Casado Muñoz M.C, Pérez Montoro B, Kabisch J, Pichner R, Cho G-S, Neve H, Fusco V, Franz CMAP, Gálvez A and Benomar N (2015) The controversial nature of the Weissella genus: technological and functional aspects versus whole genome analysis-based pathogenic potential for their application in food and health. Front. Microbiol. 6:1197. doi: 10.3389/fmicb.2015.01197
Received: 17 September 2015; Accepted: 15 October 2015;
Published: 27 October 2015.
Edited by:
Andrea Gomez-Zavaglia, Centro de Investigación y Desarrollo en Criotecnología de Alimentos, ArgentinaReviewed by:
Zhao Chen, Clemson University, USACopyright © 2015 Abriouel, Lavilla Lerma, Casado Muñoz, Pérez Montoro, Kabisch, Pichner, Cho, Neve, Fusco, Franz, Gálvez and Benomar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hikmate Abriouel, aGlrbWF0ZUB1amFlbi5lcw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.