 Kok-Gan Chan
Kok-Gan Chan Yi-Chia Liu
Yi-Chia Liu Chien-Yi Chang
Chien-Yi Chang- 1Division of Genetics and Molecular Biology, Institute of Biological Sciences, Faculty of Science, University of Malaya, Kuala Lumpur, Malaysia
- 2Division of Molecular Microbiology, School of Life Sciences, University of Dundee, Dundee, UK
- 3Centre for Bacterial Cell Biology, Medical School, Newcastle University, Newcastle upon Tyne, UK
- 4Interdisciplinary Computing and Complex BioSystems Research Group, School of Computing Science, Newcastle University, Newcastle upon Tyne, UK
Bacteria sense their own population size, tune the expression of responding genes, and behave accordingly to environmental stimuli by secreting signaling molecules. This phenomenon is termed as quorum sensing (QS). By exogenously manipulating the signal transduction bacterial population behaviors could be controlled, which may be done through quorum quenching (QQ). QS related regulatory networks have been proven their involvement in regulating many virulence determinants in pathogenic bacteria in the course of infections. Interfering with QS signaling system could be a novel strategy against bacterial infections and therefore requires more understanding of their fundamental mechanisms. Here we review the development of studies specifically on the inhibition of production of N-acyl-homoserine lactone (AHL), a common proteobacterial QS signal. The opportunistic pathogen, Pseudomonas aeruginosa, equips the alkylquinolone (AQ)-mediated QS which also plays crucial roles in its pathogenicity. The studies in QQ targeting on AQ are also discussed.
Introduction
Quorum sensing (QS) is an intercellular communication mechanism of bacteria used to coordinate the activities of individual cells in population level in response to surroundings through production and perception of diffusible signal molecules. The signal synthase, signal receptor, and signal molecules are three essential elements of the basic QS circuit machinery. Genes encoded signal generating proteins are also included among the QS target genes. This forms an autoinduction feedback loop to modulate generation of signal molecules (Fuqua et al., 1994; Ng and Bassler, 2009; Williams and Cámara, 2009). Several bacterial behaviors including virulence factors expression, secondary metabolites production, biofilm formation, motility, and luminescence are regulated by QS (Antunes et al., 2010; LaSarre and Federle, 2013). QS system is part of global regulation networks in bacterial cell related to cyclic-di-GMP second messenger signaling pathways (Srivastava and Waters, 2012), small RNA regulation (Vakulskas et al., 2015), two-components systems (Okkotsu et al., 2014), toxin–antitoxin (Kumar and Engelberg-Kulka, 2014), flagella regulation (Atkinson et al., 2008), and protein secretion systems (Zheng et al., 2010; Atkinson et al., 2011). Through complex regulatory networks bacteria are capable of expressing corresponding genes according to their own population size and of behaving in a coordinated manner.
Bacteria produce ranges of QS signal molecules. N-acyl-homoserine lactones (AHLs) are major signal molecules produced by Gram-negative bacteria (Ng and Bassler, 2009; LaSarre and Federle, 2013). Gram-positive bacteria use small linear (e.g., ComX in Bacillus subtilis) or circular peptides (e.g., autoinducing peptides AIP in Staphylococcus aureus) as signal molecules which are the ligands for the extracellular receptor of a two-component module (Ng and Bassler, 2009). There are other classes of QS molecules including diffusible signaling factor (DSF; cis-11-methyl-2-dodecenoic acid) in Xanthomonas, Xylella, Burkholderia, and Pseudomonas (Deng et al., 2011); α-hydroxyketones (AHKs) in Legionella pneumophila and Vibrio sp. (Tiaden et al., 2010); furanone molecules (autoinducer-2; AI-2) produced by Gram-negative and -positive bacteria as universal signals for inter- and intra-species communications (Kendall and Sperandio, 2007; LaSarre and Federle, 2013); aromatic molecules (autoinducer-3; AI-3) in enteric pathogens Salmonella and Escherichia coli O157:H7 served in inter-kingdom communications via the epinephrine signaling system in mammalian cells (Sperandio et al., 2003; Moreira et al., 2010) and alkylquinolone (AQ) in Pseudomonas sp. and Burkholderia sp (Heeb et al., 2011; Lee and Zhang, 2014). The fragments of peptidoglycan from bacterial cell wall have been suggested as novel QS signals for signaling the growth state of bacteria cell than population size (Dworkin, 2014).
Due to its key roles in bacterial population behaviors and pathogenicity, QS has been suggested to be the target for novel bacterial infection therapy (Zhang and Dong, 2004; Rasmussen, 2006; LaSarre and Federle, 2013). By reducing concentration of signals or interrupting the interactions of signal on receptor protein, the expression of QS-regulated genes can be disturbed and bacterial virulence can be subsequently attenuated (Dong et al., 2000, 2001). These approaches coined as quorum quenching (QQ) were considered as alternatives against bacterial infections (Cámara et al., 2002; Zhang and Dong, 2004; González and Keshavan, 2006). Several natural compounds and enzymes from prokaryotic (Dong et al., 2000; Wang, 2004; Chu et al., 2013) and eukaryotic organisms including plants (Vandeputte et al., 2011; Koh et al., 2013), murine (Yang et al., 2005), and human (Chun et al., 2004; Ozer et al., 2006) have been discovered and shown the capability of inhibiting bacterial QS systems. Broad advances on QQ studies and various types of QS inhibitors are also highlighted in several reviews (Chun et al., 2004; Yang et al., 2005; Ozer et al., 2006; Dong et al., 2007; Chu et al., 2013; Kalia, 2013; Koh et al., 2013; LaSarre and Federle, 2013; Nazzaro et al., 2013). Here we review the development of inhibiting of AHL synthesis. In the genome of Pseudomonas aeruginosa, an opportunistic human pathogen, QS is responsible for the regulation of around 10% of genes (Williams and Cámara, 2009). AQ–QS coupled with AHL-QS play crucial roles in virulence regulation. Thus the recent QQ studies on AQ–QS are also discussed.
AHL Synthesis and Inhibition
More than 30 different AHLs have been identified. AHLs generally consist of a homoserine lactone ring and of a fatty acyl side chain ranging from 4 to 18 carbons (Chhabra et al., 2005). The regulation of bioluminescence in Aliivibrio fischeri (former Vibrio fischeri) (Urbanczyk et al., 2007) is the archetypal example of AHL-QS. The genes for bioluminescence are encoded in an operon (luxICDABE) where luxI encodes the AHL synthase that generates N-(3-oxo-hexanoyl)-L-homoserine lactone (3-oxo-C6 HSL) and luxCDABE encode for proteins responsible for bioluminescence production. This operon is regulated by the LuxR protein, a transcriptional regulator. Binding of 3-oxo-C6-HSL by LuxR activates the expression of the luxICDABE operon which in turns results in the production of more of this signal molecule, through an autoinduction loop, and bioluminescence (Fuqua and Greenberg, 2002). S-adenosylmethionine (SAM) is the main amino acid substrate for the homoserine lactone ring, while the acyl chain of AHL is derived from a 6-carbon acyl-ACP (acyl carrier protein). The LuxI catalyzes the formation of an amide bond between the two substrates. SAM binds to the active site of LuxI, and then the acyl group from a specific acyl-ACP is transferred to the methionine on SAM and forms an amide bond. Lactonization of the intermediate results in the production of AHL and methylthioadenosine (MAT; Parsek et al., 1999). Two regions of LuxI are essential for the full function of the enzyme. Residues 25 to 104 are thought to be the region of the enzymatic catalysis, while residue 133 to 164 are involved in the selection of the appropriate acyl-ACP substrates (Hanzelka and Greenberg, 1996).
Homologs of luxI/luxR are involved in Gram-negative bacteria (Miller and Bassler, 2001; LaSarre and Federle, 2013). AHL synthases from more than 40 different bacteria share four conserved regions in their protein sequences and within them eight residues are completely conserved (Fuqua et al., 1994). The X-ray crystallography study of an AHL synthase EsaI, a LuxI-type protein producing 3-oxo-C6-HSL from Pantoea stewartii, has been revealed (Watson et al., 2002). A model for catalyzing the amide bond between the two substrates based on this structural study has been proposed. Many of the conserved residues across LuxI-type synthases lie on the same face of the EsaI and are localized in the active site in N-terminus. This supports a notion that SAM and acyl-ACP interact with this region. The presence or absence of a serine/threonine at residue 140 in AHL synthase may be crucial for selecting different acyl-ACP substrates. LasI, EsaI, and LuxI which mainly produce 3-oxo-AHLs have a conserved threonine at this position whilst the absence of threonine at position 140 in RhlI, CerI, SwrI, and AsaI seems to preferentially produce AHLs lacking 3-oxo or 3-hydroy moiety (Watson et al., 2002). The crystal structure of LasI, the synthase from P. aeruginosa mainly produces N-(3-oxododecanoyl)-homoserine lactone (3-oxo-C12-HSL), further reveals the mechanism of AHL synthesis. The conserved residues in LasI N-terminus form a binding packet for SAM and the structure of the acyl-ACP binding packet is formed as a tunnel which allow the binding with different acyl-ACPs with longer side chain in contrast to the restrictive hydrophobic packet in EsaI (Gould et al., 2004). The acceptance of various acyl-ACPs in LasI may explain the wild spectrum of AHL production profile in P. aeruginosa (Ortori et al., 2011).
N-acyl-homoserine lactone molecules act not only in bacterial communication but also in cross-kingdom conversation with eukaryotic cells (Joint et al., 2002; Twigg et al., 2013). AHLs modulate the behaviors of innate immune cells and interfere with the signaling pathways in epithelial cells (Holm and Vikström, 2014). Inhibition of AHLs production is the strategy to reduce the AHLs-mediated virulence factors and to prevent AHLs-promoted tissue damages and inflammation. To study the kinetic process of AHLs synthesis by RhlI from P. aeruginosa, the end products MTA and several substrate analogs including holo-ACP, sinefungin, D/L-S-adenosylhomocysteine, L-S-adenosylcysteine, and butyryl-SAM have been tested for their inhibition effects on AHLs production in vitro (Parsek et al., 1999). However their effects on QS and QS-related phenotypes in vivo are unclear (Rasmussen, 2006). More inhibitors targeting on the AHL synthesis precursors or their synthesis have been identified. A small molecule, triclosan, reduces AHL synthesis by inhibiting the precursor production from enoyl-ACP reductase (Hoang and Schweizer, 1999). The 5-MAT/S-adenosyl-homocysteine nucleosidase (MTAN), which plays crucial part in both AHL and AI-2 synthesis, was inhibited by immucillin A (ImmA) derivatives and DADMe-ImmA derivatives (Singh et al., 2005a,b, 2006). These inhibitors not only show the effective potency on AHL synthesis but also inhibit the central amino acid and fatty acid metabolisms by which may affect other cellular functions. This imposes selective pressure to bacterial cells and increases the risk of resistance. For example P. aeruginosa develops resistance to triclosan because of active efflux pumps (Schweizer, 2003). Thus, it is necessary to identify inhibitors specifically targeting on AHLs synthesis without interrupting metabolisms.
In Burkholderia glumae, the pathogen of rice grain rot, virulence factor biosynthesis and transportation, protein secretion and motility are controlled by N-octanoyl-HSL (C8-HSL) produced by TofI. Two acyl-HSL analogs have been identified to inhibit the C8-HSL mediated QS. One of the inhibitor, E9C-3oxoC6, competitively inhibits C8-HSL binding to its cognate regulator, TolR. The other inhibitor, J8-C8 (N-3-oxocyclohex-1-enyl octanamide), inhibits C8-HSL synthesis in a dose-dependent manner. Crystal structure analysis suggested that the TofI, which is similar to its homologues, LasI and EsaI, consists of two substrate binding sites (Watson et al., 2002; Gould et al., 2004; Chung et al., 2011). A putative pocket in apo-TofI structure has been identified which is bound by J8-C8. This study suggested J8-C8 occupies the acyl-ACP substrate-binding site of TofI and inhibits its function (Chung et al., 2011).
Besides rationally designed analogs, natural products isolated from various sources also exhibit QQ properties (Koh et al., 2013). Three pure botanic compounds, salicylic acid, tannic acid, and trans-cinnamaldehyde, showed the inhibition on P. aeruginosa AHL synthase. Further liquid chromatography–mass spectrometry (LC–MS) analysis suggested the trans-cinnamaldehyde specifically targets short-chain AHL synthase RhlI. The RhlI/R-QS in P. aeruginosa is involved in the production of pyocyanin, a green phenazine pigment functioning as a toxin and promoting biofilm formation. The trans-cinnamaldehyde inhibits pyocyanin production in a dose-dependent manner. Successful molecule docking suggested trans-cinnamaldehyde occupies the SAM substrate binding sites of RhlI counterpart, LasI in P. aeruginosa and the acyl-chain substrate binding site of EasI in Pa. stewartii (Chang et al., 2014). Furthermore cinnamaldehyde and its derivatives target on AI-2 based LuxR in Vibrio sp. and reduce the LuxR DNA binding ability which lead to the reduction of biofilm formation, sensitivity to starvation and antibiotics treatment, reduction of pigment and virulence factors production and attenuated pathogenicity (Brackman et al., 2008, 2011). These studies suggested the potentiality of cinnamaldehyde as a QQ compound against bacteria infections.
Quenching AQ-Mediated QS
A non-AHL signaling molecule produced by P. aeruginosa was described and termed as PQS (Pseudomonas quinolone signal) by Pesci et al. (1999). This molecule was chemically characterized as 2-heptyl-3-hydroxy-4-(1H)-quinolone, part of the 4-hydroxy-2-alkyl quinoline series (HAQ; Pesci et al., 1999). Other major molecules comprised the C7 and C9 long alkyl chain including 2-heptyl-4-quinolone (HHQ), 2-nonyl-4-quinolone (NHQ), 2-heptyl-4-quinolone N-oxide (HHQNO), 2-heptyl-4-hydroxyquinoline N-oxide (HQNO) and 2-nonyl-4-hydroxyquinoline N-oxide (NQNO) are also produced by P. aeruginosa. More than 50 different AQs have been found in P. aeruginosa. These molecules all belong to the family of 2-alkyl-4-quinolones (AQs) which have been previously studied for their antimicrobial properties (Heeb et al., 2011; Ortori et al., 2011).
2-heptyl-4-quinolone and Pseudomonas quinolone signal are the major AQ–QS signaling molecules in P. aeruginosa. The AQ–QS system consists of multiple genes. The pqsABCDE in an operon are essential for AQs synthesis. PqsA catalyzes the conversion from anthranilic acid produced by phnAB, which are adjacent to pqsABCDE operon, to anthraniloyl-coenzyme A (CoA). PqsD then condenses anthraniloyl moiety from anthraniloyl-CoA with malonyl-CoA to form intermediate 2-aminobenzoyl-acetyl-CoA (2-ABA-CoA) for subsequently synthesizing 2,4-dihydroxyquinolone (DHQ) with unknown function or 2-aminobenzoylacetate (2-ABA). The decarboxylating coupling of 2-ABA to an octanoate group linked to PqsBC produces HHQ (Bera et al., 2009; Dulcey et al., 2013). HHQ acts as a signaling molecule or can then be transformed to PQS by the mono-oxygenase, PqsH (Pesci et al., 1999; Diggle et al., 2006). PqsE is required for P. aeruginosa virulence in plant and animals infection models and biofilm formation (Rampioni et al., 2010). A recent study suggested PqsE is also involved in the HHQ synthesis as a thioesterase, hydrolyzing the 2-ABA-CoA to form 2-ABA. Although PqsE plays crucial role in P. aeruginosa pathogenicity, its catalytic role in AQ biosynthesis can be replaced by a broad specificity thioesterase, TesB (Drees and Fetzner, 2015). Two other genes, pqsH and pqsL, involved in AQ synthesis are located separately elsewhere on the chromosome in P. aeruginosa. PqsH, FAD-dependent monooxygenase, is required for the conversion of HHQ into PQS. PqsL is also a monooxygenase which is required for HQNO synthesis (Heeb et al., 2011). PQS or HHQ binds to and activates the LysR-type transcriptional regulator PqsR (also known as MvfR), which in turn induces the expression of the pqsABCDE operon and possibly the phnAB operon and triggers the typical QS autoinducing response enhancing AQ biosynthesis (Maddocks and Oyston, 2008; Heeb et al., 2011).
Alkylquinolone–quorum sensing cooperates with the AHL-QS systems, lasI/R and rhlI/R, in P. aeruginosa (Xiao et al., 2006; Heeb et al., 2011). PQS has been shown to regulate the expression of the AHL synthase gene rhlI (Pesci et al., 1999; McKnight et al., 2000). Furthermore, it has been demonstrated that PQS is essential for the activation of certain rhl-dependent genes (Diggle et al., 2003). The las QS system increases the expression of pqsR and pqsA as well as controlling the expression of pqsH indicating that las acts as a positive regulator of PQS (Déziel et al., 2004; Xiao et al., 2006). However, the biosynthesis of PQS only partially relies on the las system since PQS is still produced in the absence of lasR (Diggle et al., 2003). In contrast to the las QS system, the rhl system negatively regulates the PQS production (Xiao et al., 2006; Heeb et al., 2011).
Alkylquinolone–quorum sensing also plays important roles in pathogenicity. Mutants on AQ–QS showed reduced P. aeruginosa virulence in infection models (Cao et al., 2001; Diggle et al., 2003; Déziel et al., 2004). These results suggest that PQS or HHQ signaling pathway could be a novel target against P. aeruginosa infecyions. A sesquiterpene, farnesol produced by the fungus Candida albicans, decreases the level of pqsA expression by interfering with PqsR-mediated transcription activation and sequentially reduces the PQS and PQS-regulated pyocyanin production. However, in the PQS-defective lasR mutant farnesol restores PQS production by inducing PqsH via RhlI/C4-HSL activation (Cugini et al., 2007, 2010). Around 50% of strains isolated from lungs of late stage cystic fibrosis (CF) patients are deficient in lasR function (Winstanley and Fothergill, 2009). The fungal–bacterial communication via farnesol is still unclear and may provide a new target for mitigating bacterial chronic infection in CF lungs.
A monomeric enzyme, Hod (3-hydroxy-2-methyl-4(1H)-quinolone 2,4-dioxygenase) from Arthrobacter sp. strain Rue61a involved in the degradation of 2-methylquinoline (quinaldine), is able to cleave PQS to N-octanoylanthranilic acid and carbon monoxide but not HHQ. Exogenous Hod reduces the PQS production and PQS-regulated virulence factors, lectin A, pyrocyanin, and rhamnolipids in P. aeruginosa. Hod also attenuates the pathogenicity of P. aeruginosa in a plant leaf infection model (Pustelny et al., 2009). It has been reported that, a group of analogs of PQS precursor anthranilatic acid (AA) including methylanthranilate (MA), 2-amino-6-fluorobenzoic acid (6FABA), 2-amino-6-chlorobenzoic acid (6CABA), and 2-amino-4-chlor-obenzoic acid (4CABA), acts as competitors for PqsA active site and as inhibitors for HHQ and PQS production resulting in reduced pathogenicity of P. aeruginosa in mice infection models (Calfee et al., 2001; Lesic et al., 2007). Recently Soh et al. (2015) reports an environmental bacterium, Achromobacter xylosoxidans Q19 from rainforest soil, is capable of oxidizing PQS into 2-heptyl-2-hydroxy-1,2-dihydroquinoline-3,4-dione (HHQD), which was elucidated by mass spectrometry and nuclear magnetic resonance spectroscopy. The PQS oxidation to HHQD also occurs with less efficiency in the A. xylosoxidans and P. aeruginosa from CF lung which suggests that the hydroxylated PQS is a common molecule in soil and in CF lungs (Soh et al., 2015). However, the biological function of HHQD is still unknown.
Besides targeting on PQS molecules production or modification, a group of compounds with a benzamide-benzimidazole backbone targeting on MvfR (PqsR)-regulated pathways have been identified by using a whole-cell high-throughput screen (HTS) and structure–activity relationship (SAR) analysis. These compounds inhibit the production of pyocyanin in several clinically isolated P. aeruginosa strains; limit the formation of antibiotic-resistant cells; rescue mice macrophage from the bacterial cytotoxicity; attenuate the bacterial pathogenicity in acute thermal injury and lung infection murine models; reduce the accumulation of macrophage at the infection sites resulting in inhibition of inflammation evoked by bacteria and inhibit bacterial persistence for developing chronic infection in burned mice. One of the most effective compounds, M64 molecule, binds to MvfR and decreases the MvfR–DNA binding affinity by 10-folds which results in the reduction of MvfR-activated virulence factors. These compounds have been suggested as next generation therapeutic agents against bacterial infections (Starkey et al., 2014).
Conclusion
Here we summarize recent studies of AHL synthesis inhibition and QQ on P. aeruginosa AQ–QS (Table 1). Although QS is an ideal target to attenuate bacterial virulence and pathogenicity, several studies warn that bacteria rapidly evolve and spread the resistance against QS inhibitors like the fate of antibiotics (García-Contreras et al., 2013; Kalia et al., 2013). Furthermore Decho et al. (2010) suggested that the biologically or chemically modified signals might interact with unexpected signal receptors for unpredicted outcomes in the complex natural environments. Little is known about the effects of QS inhibitors or enzymes on the broader microorganisms’ communities or the fate of products from signal molecules degradation and modifications in the environments (Decho et al., 2010). Many questions remain for further investigation. Thus it is highly desired to further investigate the virulent roles of QS signals in infections and to explore the strategy to eliminate their production rather than to modify or to degrade them for diminishing unpredicted impacts to environments.
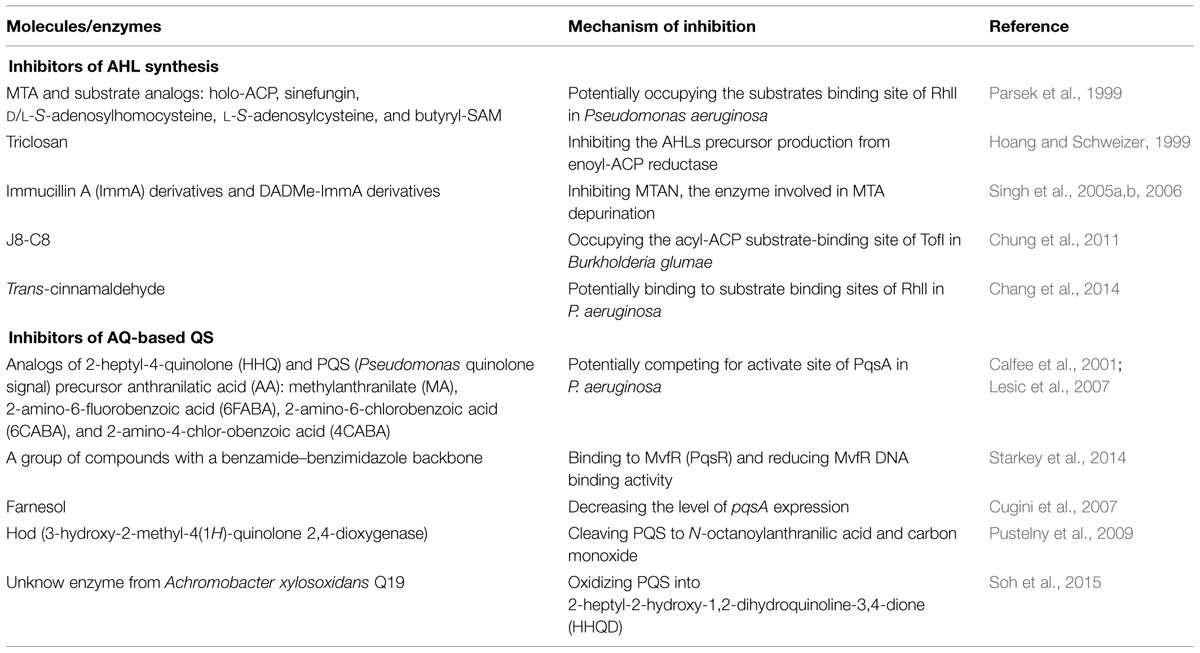
TABLE 1. Overview of inhibitors on N-acyl-homoserine lactone (AHL) synthesis and on alkylquinolone (AQ)-mediated quorum sensing (QS) system.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the University of Malaya High Impact Research Grants (UM C/625/1/HIR/MOHE/CHAN/01, A-000001-50001, and UM C/625/1/HIR/MOHE/CHAN/14/1, H-50001-A000027) awarded to K.-G.C.
References
Antunes, L. C. M., Ferreira, R. B. R., Buckner, M. M. C., and Finlay, B. B. (2010). Quorum sensing in bacterial virulence. Microbiology 156, 2271–2282. doi: 10.1099/mic.0.038794-0
Atkinson, S., Chang, C.-Y., Patrick, H. L., Buckley, C. M. F., Wang, Y., Sockett, R. E., et al. (2008). Functional interplay between the Yersinia pseudotuberculosis YpsRI and YtbRI quorum sensing systems modulates swimming motility by controlling expression of flhDC and fliA. Mol. Microbiol. 69, 137–151. doi: 10.1111/j.1365-2958.2008.06268.x
Atkinson, S., Goldstone, R. J., Joshua, G. W. P., Chang, C. Y., Patrick, H. L., Cámara, M., et al. (2011). Biofilm development on Caenorhabditis elegans by Yersinia is facilitated by quorum sensing-dependent repression of type III secretion. PLoS Pathog. 7:e1001250. doi: 10.1371/journal.ppat.1001250.g009
Bera, A. K., Atanasova, V., Robinson, H., Eisenstein, E., Coleman, J. P., Pesci, E. C., et al. (2009). Structure of PqsD, a Pseudomonas quinolone signal biosynthetic enzyme, in complex with anthranilate. Biochemistry 48, 8644–8655. doi: 10.1021/bi9009055
Brackman, G., Celen, S., Hillaert, U., Van Calenbergh, S., Cos, P., Maes, L., et al. (2011). Structure-activity relationship of cinnamaldehyde analogs as inhibitors of AI-2 Based quorum sensing and their effect on virulence of Vibrio spp. PLoS ONE 6:e16084. doi: 10.1371/journal.pone.0016084.s001
Brackman, G., Defoirdt, T., Miyamoto, C., Bossier, P., Van Calenbergh, S., Nelis, H., et al. (2008). Cinnamaldehyde and cinnamaldehyde derivatives reduce virulence in Vibrio spp. by decreasing the DNA-binding activity of the quorum sensing response regulator LuxR. BMC Microbiol. 8:149. doi: 10.1186/1471-2180-8-149
Calfee, M. W., Coleman, J. P., and Pesci, E. C. (2001). Interference with Pseudomonas quinolone signal synthesis inhibits virulence factor expression by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 98, 11633–11637. doi: 10.1073/pnas.201328498
Cámara, M., Williams, P., and Hardman, A. (2002). Controlling infection by tuning in and turning down the volume of bacterial small-talk. Lancet Infect. Dis. 2, 667–676. doi: 10.1016/S1473-3099(02)00447-4
Cao, H., Krishnan, G., Goumnerov, B., Tsongalis, J., Tompkins, R., and Rahme, L. G. (2001). A quorum sensing-associated virulence gene of Pseudomonas aeruginosa encodes a LysR-like transcription regulator with a unique self-regulatory mechanism. Proc. Natl. Acad. Sci. U.S.A. 98, 14613–14618. doi: 10.1073/pnas.251465298
Chang, C.-Y., Krishnan, T., Wang, H., Chen, Y., Yin, W. F., Chong, Y. M., et al. (2014). Non-antibiotic quorum sensing inhibitors acting against N-acyl homoserine lactone synthase as druggable target. Sci. Rep. 4, 7245. doi: 10.1038/srep07245
Chhabra, S. R., Philipp, B., Eberl, L., Givskov, M., Williams, P., and Cámara, M. (2005). Extracellular communication in bacteria. Topic Curr. Chem. 240, 279–315. doi: 10.1007/s10038-008-0301-3
Chu, Y.-Y., Nega, M., Wölfle, M., Plener, L., Grond, S., Jung, K., et al. (2013). A new class of quorum quenching molecules from Staphylococcus species affects communication and growth of gram-negative bacteria. PLoS Pathog. 9:e1003654. doi: 10.1371/journal.ppat.1003654.s003
Chun, C. K., Ozer, E. A., Welsh, M. J., Zabner, J., and Greenberg, E. P. (2004). Inactivation of a Pseudomonas aeruginosa quorum-sensing signal by human airway epithelia. Proc. Natl. Acad. Sci. U.S.A. 101, 3587–3590. doi: 10.1073/pnas.0308750101
Chung, J., Goo, E., Yu, S., Choi, O., Lee, J., Kim, J., et al. (2011). Small-molecule inhibitor binding to an N-acyl-homoserine lactone synthase. Proc. Natl. Acad. Sci. U.S.A. 108, 12089–12094. doi: 10.1073/pnas.1103165108
Cugini, C., Calfee, M. W., Farrow, J. M., Morales, D. K., Pesci, E. C., and Hogan, D. A. (2007). Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol. Microbiol. 65, 896–906. doi: 10.1111/j.1365-2958.2007.05840.x
Cugini, C., Morales, D. K., and Hogan, D. A. (2010). Candida albicans produced farnesol stimulates Pseudomonas quinolone signal production in LasR-defective Pseudomonas aeruginosa strains. Microbiology 156, 3096–3107. doi: 10.1099/mic.0.037911-0
Decho, A. W., Norman, R. S., and Visscher, P. T. (2010). Quorum sensing in natural environments: emerging views from microbial mats. Trends Microbiol. 18, 73–80. doi: 10.1016/j.tim.2009.12.008
Deng, Y., Wu, J., Tao, F., and Zhang, L.-H. (2011). Listening to a new language: DSF-based quorum sensing in Gram-negative bacteria. Chem. Rev. 111, 160–173. doi: 10.1021/cr100354f
Déziel, E., Gopalan, S., Tampakaki, A. P., Lépine, F., Padfield, K. E., Saucier, M., et al. (2004). The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl- l-homoserine lactones. Mol. Microbiol. 55, 998–1014. doi: 10.1111/j.1365-2958.2004.04448.x
Diggle, S. P., Cornelis, P., Williams, P., and Cámara, M. (2006). 4-Quinolone signalling in Pseudomonas aeruginosa: old molecules, new perspectives. Int. J. Med. Microbiol. 296, 83–91. doi: 10.1016/j.ijmm.2006.01.038
Diggle, S. P., Winzer, K., Chhabra, S. R., Worrall, K. E., Cámara, M., and Williams, P. (2003). The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol. 50, 29–43. doi: 10.1046/j.1365-2958.2003.03672.x
Dong, Y. H., Wang, L. H., Xu, J. L., Zhang, H. B., and Zhang, X. F. (2001). Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 411, 813–817. doi: 10.1038/35081101
Dong, Y. H., Wang, L. H., and Zhang, L. H. (2007). Quorum-quenching microbial infections: mechanisms and implications. Philos. Trans. Roy. Soc. B 362, 1201–1211. doi: 10.1073/pnas.96.9.4832
Dong, Y. H., Xu, J. L., and Li, X. Z. (2000). AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. U.S.A. 97, 3526–3531. doi: 10.1073/pnas.97.7.3526
Drees, S. L., and Fetzner, S. (2015). PqsE of Pseudomonas aeruginosa acts as pathway-specific thioesterase in the biosynthesis of alkylquinolone signaling molecules. Chem. Biol. 22, 611–618. doi: 10.1016/j.chembiol.2015.04.012
Dulcey, C. E., Dekimpe, V., Fauvelle, D.-A., Milot, S., Groleau, M.-C., Doucet, N., et al. (2013). The end of an old hypothesis: the Pseudomonas signaling molecules 4-hydroxy-2-alkylquinolines derive from fatty acids, not 3-ketofatty acids. Chem. Biol. 20, 1481–1491. doi: 10.1016/j.chembiol.2013.09.021
Dworkin, J. (2014). The medium is the message: interspecies and interkingdom signaling by peptidoglycan and related bacterial glycans. Annu. Rev. Microbiol. 68, 137–154. doi: 10.1146/annurev-micro-091213-112844
Fuqua, C., and Greenberg, E. P. (2002). Signalling: listening in on bacteria: acyl-homoserine lactone signalling. Nat. Rev. Mol. Cell Biol. 3, 685–695. doi: 10.1038/nrm907
Fuqua, W. C., Winans, S. C., and Greenberg, E. P. (1994). Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 176, 269–275.
García-Contreras, R., Maeda, T., and Wood, T. K. (2013). Resistance to quorum-quenching compounds. Appl. Environ. Microbiol. 79, 6840–6846. doi: 10.1128/AEM.02378-13
González, J. E., and Keshavan, N. D. (2006). Messing with bacterial quorum sensing. Microbiol. Mol. Biol. Rev. 70, 859–875. doi: 10.1128/MMBR.00002-06
Gould, T. A., Schweizer, H. P., and Churchill, M. E. A. (2004). Structure of the Pseudomonas aeruginosa acyl-homoserinelactone synthase LasI. Mol. Microbiol. 53, 1135–1146. doi: 10.1111/j.1365-2958.2004.04211.x
Hanzelka, B. L., and Greenberg, E. P. (1996). Quorum sensing in Vibrio fischeri: evidence that S-adenosylmethionine is the amino acid substrate for autoinducer synthesis. J. Bacteriol. 178, 5291–5294.
Heeb, S., Fletcher, M. P., Chhabra, S. R., Diggle, S. P., Williams, P., and Cámara, M. (2011). Quinolones: from antibiotics to autoinducers. FEMS Microbiol. Rev. 35, 247–274. doi: 10.1111/j.1574-6976.2010.00247.x
Hoang, T. T., and Schweizer, H. P. (1999). Characterization of Pseudomonas aeruginosa enoyl-acyl carrier protein reductase (FabI): a target for the antimicrobial triclosan and its role in acylated homoserine lactone synthesis. J. Bacteriol. 181, 5489–5497.
Holm, A., and Vikström, E. (2014). Quorum sensing communication between bacteria and human cells: signals, targets, and functions. Front. Plant Sci. 5:309–309. doi: 10.3389/fpls.2014.00309
Joint, I., Tait, K., Callow, M. E., Callow, J. A., Milton, D., Williams, P., et al. (2002). Cell-to-cell communication across the prokaryote-eukaryote boundary. Science 298, 1207. doi: 10.1126/science.1077075
Kalia, V. C. (2013). Quorum sensing inhibitors: an overview. Biotech. Adv. 31, 224–245. doi: 10.1016/j.biotechadv.2012.10.004
Kalia, V. C., Wood, T. K., and Kumar, P. (2013). Evolution of resistance to quorum-sensing inhibitors. Microb. Ecol. 68, 13–23. doi: 10.1007/s00248-013-0316-y
Kendall, M. M., and Sperandio, V. (2007). Quorum sensing by enteric pathogens. Curr. Opin. Gastroenterol. 23, 10–15. doi: 10.1097/MOG.0b013e3280118289
Koh, C.-L., Sam, C.-K., Yin, W. F., Tan, L. Y., Krishnan, T., Chong, Y. M., et al. (2013). Plant-derived natural products as sources of anti-quorum sensing compounds. Sensors 13, 6217–6228. doi: 10.3390/s130506217
Kumar, S., and Engelberg-Kulka, H. (2014). Quorum sensing peptides mediating interspecies bacterial cell death as a novel class of antimicrobial agents. Curr. Opin. Microbiol. 21, 22–27. doi: 10.1016/j.mib.2014.09.001
LaSarre, B., and Federle, M. J. (2013). Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 77, 73–111. doi: 10.1128/MMBR.00046-12
Lee, J., and Zhang, L. (2014). The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 6, 26–41. doi: 10.1007/s13238-014-0100-x
Lesic, B., Lépine, F., Déziel, E., Zhang, J., Zhang, Q., Padfield, K., et al. (2007). Inhibitors of pathogen intercellular signals as selective anti-infective compounds. PLoS Pathog. 3:e126. doi: 10.1371/journal.ppat.0030126.st003
Maddocks, S. E., and Oyston, P. C. F. (2008). Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 154, 3609–3623. doi: 10.1099/mic.0.2008/022772-0
McKnight, S. L., Iglewski, B. H., and Pesci, E. C. (2000). The Pseudomonas quinolone signal regulates rhl quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 182, 2702–2708. doi: 10.1128/JB.182.10.2702-2708.2000
Miller, M. B., and Bassler, B. L. (2001). Quorum sensing in bacteria. Ann. Rev. Microbiol. 55, 165–199. doi: 10.1146/annurev.micro.55.1.165
Moreira, C. G., Weinshenker, D., and Sperandio, V. (2010). QseC mediates Salmonella enterica serovar Typhimurium virulence in vitro and in vivo. Infect. Immun. 78, 914–926. doi: 10.1128/IAI.01038-09
Nazzaro, F., Fratianni, F., and Coppola, R. (2013). Quorum sensing and phytochemicals. Int. J. Mol. Sci. 14, 12607–12619. doi: 10.3390/ijms140612607
Ng, W.-L., and Bassler, B. L. (2009). Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 43, 197–222. doi: 10.1146/annurev-genet-102108-134304
Okkotsu, Y., Little, A. S., and Schurr, M. J. (2014). The Pseudomonas aeruginosa AlgZR two-component system coordinates multiple phenotypes. Front. Cell Infect. Microbiol. 4:82–82. doi: 10.3389/fcimb.2014.00082
Ortori, C. A., Dubern, J.-F., Chhabra, S. R., Cámara, M., Hardie, K., Williams, P., et al. (2011). Simultaneous quantitative profiling of N-acyl-L-homoserine lactone and 2-alkyl-4(1H)-quinolone families of quorum-sensing signaling molecules using LC-MS/MS. Anal. Bioanal. Chem. 399, 839–850. doi: 10.1007/s00216-010-4341-0
Ozer, E. A., Pezzulo, A., Shih, D. M., Chun, C., Furlong, C., Lusis, A. J., et al. (2006). Human and murine paraoxonase 1 are host modulators of Pseudomonas aeruginosa quorum-sensing. FEMS Microbiol. Lett. 253, 29–37. doi: 10.1016/j.femsle.2005.09.023
Parsek, M. R., Val, D. L., Hanzelka, B. L., Cronan, J. E., and Greenberg, E. P. (1999). Acyl homoserine-lactone quorum-sensing signal generation. Proc. Natl. Acad. Sci. U.S.A. 96, 4360–4365. doi: 10.1073/pnas.96.8.4360
Pesci, E. C., Milbank, J. B., Pearson, J. P., McKnight, S., Kende, A. S., Greenberg, E. P., et al. (1999). Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 96, 11229–11234. doi: 10.1073/pnas.96.20.11229
Pustelny, C., Albers, A., BUldt-Karentzopoulos, K., Parschat, K., Chhabra, S. R., Cámara, M., et al. (2009). Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem. Biol. 16, 1259–1267. doi: 10.1016/j.chembiol.2009.11.013
Rampioni, G., Pustelny, C., Fletcher, M. P., Wright, V. J., Bruce, M., Rumbaugh, K. P., et al. (2010). Transcriptomic analysis reveals a global alkyl-quinolone-independent regulatory role for PqsE in facilitating the environmental adaptation of Pseudomonas aeruginosa to plant and animal hosts. Environ. Microbiol. 12, 1659–1673. doi: 10.1111/j.1462-2920.2010.02214.x
Rasmussen, T. B. (2006). Quorum sensing inhibitors: a bargain of effects. Microbiology 152, 895–904. doi: 10.1099/mic.0.28601-0
Schweizer, H. P. (2003). Efflux as a mechanism of resistance to antimicrobials in Pseudomonas aeruginosa and related bacteria: unanswered questions. Genet. Mol. Res. 2, 48–62.
Singh, V., Evans, G. B., Lenz, D. H., Mason, J. M., Clinch, K., Mee, S., et al. (2005a). Femtomolar transition state analogue inhibitors of 5′-methylthioadenosine/S-adenosylhomocysteine nucleosidase from Escherichia coli. J. Biol. Chem. 280, 18265–18273. doi: 10.1074/jbc.M414472200
Singh, V., Lee, J. E., Núñez, S., Howell, P. L., and Schramm, V. L. (2005b). Transition state structure of 5′-methylthioadenosine/S-adenosylhomocysteine nucleosidase from Escherichia coli and its similarity to transition state analogues. Biochemistry 44, 11647–11659. doi: 10.1021/bi050863a
Singh, V., Shi, W., Almo, S. C., Evans, G. B., Furneaux, R. H., Tyler, P. C., et al. (2006). Structure and inhibition of a quorum sensing target from Streptococcus pneumoniae. Biochemistry 45, 12929–12941. doi: 10.1021/bi061184i
Soh, E. Y.-C., Chhabra, S. R., Halliday, N., Heeb, S., Müller, C., Birmes, F. S., et al. (2015). Biotic inactivation of the Pseudomonas aeruginosa quinolone signal molecule. Environ. Microbiol. doi: 10.1111/1462-2920.12857 [Epub ahead of print].
Sperandio, V., Torres, A. G., Jarvis, B., Nataro, J. P., and Kaper, J. B. (2003). Bacteria-host communication: the language of hormones. Proc. Natl. Acad. Sci. U.S.A. 100, 8951–8956. doi: 10.1073/pnas.1537100100
Srivastava, D., and Waters, C. M. (2012). A tangled web: regulatory connections between quorum sensing and cyclic di-GMP. J. Bacteriol. 194, 4485–4493. doi: 10.1128/JB.00379-12
Starkey, M., Lépine, F., Maura, D., Bandyopadhaya, A., Lesic, B., He, J., et al. (2014). Identification of anti-virulence compounds that disrupt quorum-sensing regulated acute and persistent pathogenicity. PLoS Pathog. 10:e1004321. doi: 10.1371/journal.ppat.1004321.s009
Tiaden, A., Spirig, T., and Hilbi, H. (2010). Bacterial gene regulation by alpha-hydroxyketone signaling. Trends Microbiol. 18, 288–297. doi: 10.1016/j.tim.2010.03.004
Twigg, M. S., Tait, K., Williams, P., Atkinson, S., and Cámara, M. (2013). Interference with the germination and growth of Ulva zoospores by quorum-sensing molecules from Ulva-associated epiphytic bacteria. Environ. Microbiol. 16, 445–453. doi: 10.1111/1462-2920.12203
Urbanczyk, H., Ast, J. C., Higgins, M. J., Carson, J., and Dunlap, P. V. (2007). Reclassification of Vibrio fischeri, Vibrio logei, Vibrio salmonicida and Vibrio wodanis as Aliivibrio fischeri gen. nov., comb. nov., Aliivibrio logei comb. nov., Aliivibrio salmonicida comb. nov. and Aliivibrio wodanis comb. nov. Int. J. Syst. Evol. Microbiol. 57, 2823–2829. doi: 10.1099/ijs.0.65081-0
Vakulskas, C. A., Potts, A. H., Babitzke, P., Ahmer, B. M. M., and Romeo, T. (2015). Regulation of bacterial virulence by Csr (Rsm) Systems. Microbiol. Mol. Biol. Rev. 79, 193–224. doi: 10.1128/MMBR.00052-14
Vandeputte, O. M., Kiendrebeogo, M., Rasamiravaka, T., Stevigny, C., Duez, P., Rajaonson, S., et al. (2011). The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology 157, 2120–2132. doi: 10.1099/mic.0.049338-0
Wang, L. H. (2004). Specificity and enzyme kinetics of the quorum-quenching N-acyl homoserine lactone lactonase (AHL-lactonase). J. Biol. Chem. 279, 13645–13651. doi: 10.1074/jbc.M311194200
Watson, W. T., Minogue, T. D., Val, D. L., Bodman, von, S. B., and Churchill, M. E. A. (2002). Structural basis and specificity of acyl-homoserine lactone signal production in bacterial quorum sensing. Mol. Cell 9, 685–694. doi: 10.1016/S1097-2765(02)00480-X
Williams, P., and Cámara, M. (2009). Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 12, 182–191. doi: 10.1016/j.mib.2009.01.005
Winstanley, C., and Fothergill, J. L. (2009). The role of quorum sensing in chronic cystic fibrosis Pseudomonas aeruginosa infections. FEMS Microbiol. Lett. 290, 1–9. doi: 10.1111/j.1574-6968.2008.01394.x
Xiao, G., He, J., and Rahme, L. G. (2006). Mutation analysis of the Pseudomonas aeruginosa mvfR and pqsABCDE gene promoters demonstrates complex quorum-sensing circuitry. Microbiology 152, 1679–1686. doi: 10.1099/mic.0.28605-0
Yang, F., Wang, L.-H., Wang, J., Dong, Y.-H., Hu, J. Y., and Zhang, L.-H. (2005). Quorum quenching enzyme activity is widely conserved in the sera of mammalian species. FEBS Lett. 579, 3713–3717. doi: 10.1016/j.febslet.2005.05.060
Zhang, L.-H., and Dong, Y.-H. (2004). Quorum sensing and signal interference: diverse implications. Mol. Microbiol. 53, 1563–1571. doi: 10.1111/j.1365-2958.2004.04234.x
Keywords: quorum sensing, quorum quenching, N-acyl-homoserine lactone, Pseudomonas quinolone signal, Pseudomonas aeruginosa, alkylquinolone
Citation: Chan K-G, Liu Y-C and Chang C-Y (2015) Inhibiting N-acyl-homoserine lactone synthesis and quenching Pseudomonas quinolone quorum sensing to attenuate virulence. Front. Microbiol. 6:1173. doi: 10.3389/fmicb.2015.01173
Received: 04 August 2015; Accepted: 09 October 2015;
Published: 19 October 2015.
Edited by:
Maria Tereza Dos Santos Correia, Universidade Federal de Pernambuco, BrazilReviewed by:
Efstathios D. Giaouris, University of the Aegean, GreeceShaohua Chen, Agency for Science, Technology and Research, Singapore
Copyright © 2015 Chan, Liu and Chang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chien-Yi Chang, Y2hpZW4teWkuY2hhbmdAbmNsLmFjLnVr