 María B. Sánchez
María B. Sánchez
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Microbiol., 30 June 2015
Sec. Infectious Agents and Disease
Volume 6 - 2015 | https://doi.org/10.3389/fmicb.2015.00658
This article is part of the Research TopicA Multidisciplinary Look at Stenotrophomonas maltophilia: An Emerging Multi-Drug-Resistant Global Opportunistic PathogenView all 12 articles
Stenotrophomonas maltophilia is an environmental bacterium found in the soil, associated with plants and animals, and in aquatic environments. It is also an opportunistic pathogen now causing an increasing number of nosocomial infections. The treatment of S. maltophilia is quite difficult given its intrinsic resistance to a number of antibiotics, and because it is able to acquire new resistances via horizontal gene transfer and mutations. Certainly, strains resistant to quinolones, cotrimoxale and/or cephalosporins—antibiotics commonly used to treat S. maltophilia infections—have emerged. The increasing number of available S. maltophilia genomes has allowed the identification and annotation of a large number of antimicrobial resistance genes. Most encode inactivating enzymes and efflux pumps, but information on their role in intrinsic and acquired resistance is limited. Non-typical antibiotic resistance mechanisms that also form part of the intrinsic resistome have been identified via mutant library screening. These include non-typical antibiotic resistance genes, such as bacterial metabolism genes, and non-inheritable resistant phenotypes, such as biofilm formation and persistence. Their relationships with resistance are complex and require further study.
Stenotrophomonas maltophilia is an opportunistic nosocomial pathogen that has caused an increasing number of infections in recent years (Brooke, 2012). It is associated with a number of clinical syndromes, such as endocarditis, urinary infections, and respiratory infections, including pneumonia in patients with cystic fibrosis and the immunocompromised (Falagas et al., 2009; Looney et al., 2009).
S. maltophilia shows low susceptibility to many antibiotics, including those commonly used to treat the infections it causes. It is therefore crucial that new antibiotic targets be found, and the appearance of resistance during treatment be predicted. The analysis of resistance mechanisms and the identification of antibiotic resistance genes can help in this. Bioinformatic studies have identified genes showing homology to known antibiotic resistance genes, although their exact functions remain to be confirmed. In recent years, deep sequencing technologies have allowed the complete sequencing of two clinical S. maltophilia strains, K279a and D457 (Accession numbers: NC_010943.1 and NC_017671.1) (Crossman et al., 2008; Lira et al., 2012), and two environmental strains, R551-3 and JV3 (Accession numbers: NC_011071.1 and CP002986.1). The assembly of the sequences of several other strains is currently underway, and results should be available in the near future. Genome sequencing has revealed much of the genome to be conserved across different S. maltophilia strains (Rocco et al., 2009; Alavi et al., 2014). In addition, most of the genes associated with resistance in S. maltophilia have been found present in all strains examined. However, the genomes show also sequence variability, associated to normal evolution (mutation frequency) or induced in some situations. Antibiotic pressure increases the sequence variability in resistance or related genes, as regulators. The use of quinolones in S. maltophilia allows the appearance of mutants overexpressing efflux pumps, first SmeDEF, and when this is not present, SmeVWX. In both cases the overexpression is mainly associated to mutations in their regulators, SmeT and SmeRv, respectively (Garcia-Leon et al., 2014b).
The analysis of transposon mutant libraries allows the identification of genes which, if deleted, alter susceptibility to antibiotics, including genes that might appear to have no clear relationship with antibiotic resistance. The drawback of this type of analysis is that, since essential genes cannot be deleted, their putative involvement in antibiotic susceptibility cannot be confirmed. This methodology has, however, been successfully used with Pseudomonas aeruginosa and Escherichia coli (Girgis et al., 2009; Alvarez-Ortega et al., 2010). When S. maltophilia was thus examined, genes involved in biofilm formation appeared as being related to antibiotic resistance (Huang et al., 2006; Kang et al., 2015).
Transformation with genomic libraries allows the effect of gene overexpression to be investigated; genes whose overexpression (usually in heterologous microorganisms) exerts a direct effect on antibiotic susceptibility can therefore be identified. Only those genes which exert a direct effect on antibiotic susceptibility identified in this way. The use of a S. maltophilia D457R chromosomal DNA library allowed the identification of genes related to antibiotic resistance, and the subsequent cloning and characterization of the SmeDEF efflux pump (Alonso and Martinez, 2000; Alonso et al., 2000).
Evolution assays can provide information on the genes involved in acquired resistance. Such information could be useful for the rational design of better treatments since it might help predict the appearance of resistance.
The reduced susceptibility of S. maltophilia to antibiotics has been associated with intrinsic resistance factors common to all S. maltophilia strains, such as low membrane permeability, the presence of multidrug resistance (MDR) efflux pumps, antibiotic-modifying enzymes, and the quinolone resistance gene Smqnr (Crossman et al., 2008; Sanchez et al., 2009) (Table 1). Resistance can also be acquired via the acquisition of mutations or resistance genes through horizontal gene transfer (HGT). Microorganisms sharing the same environment can provide these exogenous genes. It has been postulated that other unknown mechanisms may also help account for the S. maltophilia antibiotic resistance phenotype. The intrinsic resistome has been defined as the group of chromosomal genes involved in the intrinsic resistance present in the strains of a bacterial species prior to exposure to an antibiotic and which is not due to HGT (Fajardo et al., 2008). The intrinsic resistome involves known and unknown genes related to antibiotic resistance, which might include genes involved in cell metabolism (Olivares et al., 2013). To date, most studies have focused on classical antibiotic resistance genes, such as those coding for efflux pumps or modifying enzymes (intrinsic resistance), and on the appearance during clinical treatment of mutants showing low susceptibility to antibiotics (acquired resistance). Our knowledge of the S. maltophilia intrinsic resistome is, however, limited.
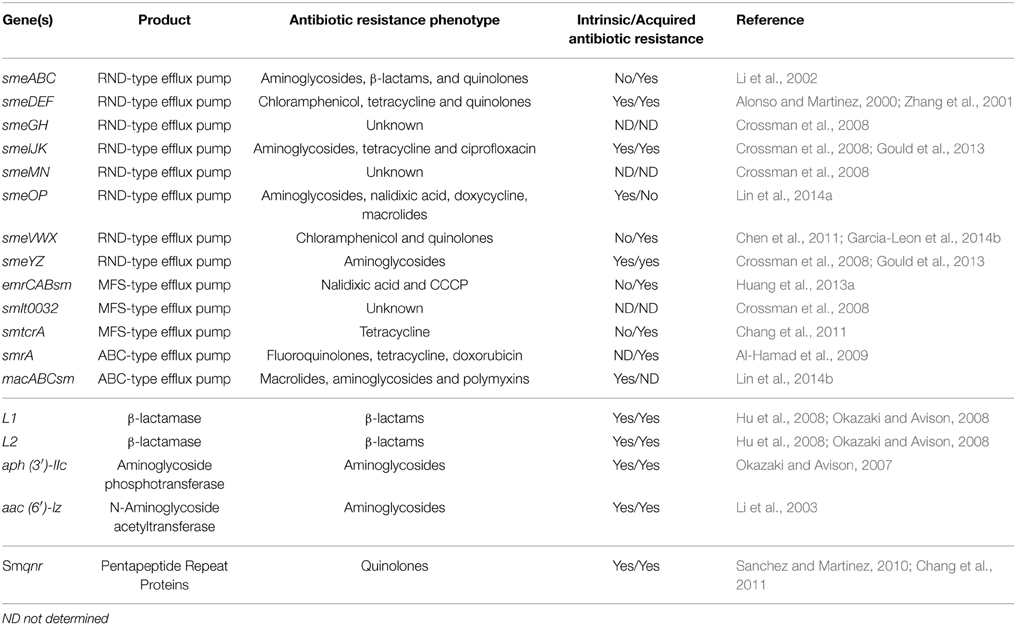
Table 1. Summary of known antibiotic resistances genes in S. maltophilia, antibiotic resistance profile and their role in intrinsic and acquired resistance.
Like all Gram negative bacteria, S. maltophilia shows low membrane permeability—the consequence of it having two cell membranes and a peptidoglycan wall. The outer membrane is an efficient barrier. Mutants showing altered outer membrane permeability or which have a different lipopolysaccharide structure show modified susceptibility to antibiotics (Vaara, 1993; Rahmati-Bahram et al., 1996).
Low susceptibility to antibiotics is often related to the presence of active efflux pumps. Such pumps have been identified in S. maltophilia K279a, including eight MDR efflux pumps belonging to the putative resistance nodulation cell division (RND) family, two belonging to the major facilitator superfamily (MFS), and two ATP-binding cassette (ABC) pumps (Crossman et al., 2008). In Gram negative bacteria, RND efflux pumps are composed of three proteins: an inner membrane protein, which binds the substrate, an outer membrane protein (porin), and a membrane fusion protein (MFP), which binds the outer and inner proteins in the periplasmic space. In general, the genes coding for the porin, MFP and inner protein are located in the same operon. Some exceptions in which there is no porin-coding gene have been identified. In addition, MDR efflux pumps are modulated by a regulator protein encoded by a gene located upstream and divergently transcribed from the efflux pump operon. In general, most efflux pump machinery is expressed at low levels (Li et al., 2002; Lin et al., 2014a). Overexpression is associated with low antibiotic susceptibility, and is sometimes related to mutations in regulator genes. Such mutations have been identified both in vitro and in vivo (Alonso and Martinez, 2001; Cho et al., 2012; Gould et al., 2013; García-León et al., 2015), supporting the idea that in vitro evolution studies may be able predict mutations appearing in vivo during the treatment of patients.
All the proteins of the efflux pumps SmeABC, SmeDEF and SmeVWX, which belong to the RND family, are encoded in the same operon following the typical genomic arrangement. The roles of these efflux pumps in intrinsic and acquired resistance have been extensively characterized (Alonso and Martinez, 2000; Li et al., 2002; Chen et al., 2011). SmeABC is involved in acquired resistance to β-lactams, aminoglycosides and quinolones, but has no influence on intrinsic resistance. The deletion of the smeC gene (porin) affects susceptibility to several antibiotics (Li et al., 2002), suggesting its possible relationship with other efflux pumps. SmeDEF is involved in both intrinsic and acquired resistance to chloramphenicol, tetracycline and quinolones, as well as acquired resistance to non-antibiotic compounds such as triclosan (Sanchez et al., 2005; Hernandez et al., 2011). SmeVWX has a role in acquired resistance to the same antibiotics (Alonso and Martinez, 2001; Zhang et al., 2001; Chen et al., 2011; Garcia-Leon et al., 2014b). In acquired resistance, the overexpression of the SmeDEF and SmeVWX efflux pumps is related to mutations in the regulators SmeT and SmeRv, respectively (Sanchez et al., 2002; Garcia-Leon et al., 2014b).
Other S. maltophilia efflux pumps have recently been studied, including SmeIJK and SmeYZ, which also belong to the RND family. Both have a role in intrinsic and acquired resistance, SmeJK to aminoglycosides, tetracycline and ciprofloxacin, and SmeZ to aminoglycosides (Crossman et al., 2008). In addition, their overexpression provides resistance to levofloxacin (Gould et al., 2013). Neither of these efflux pumps has a known associated porin. The efflux pump SmeOP, another RND family member, confers low susceptibility to aminoglycosides, nalidixic acid, doxycycline, macrolides and certain not antibiotic compounds, such as carbonyl cyanide 3-chlorophenylhydrazone (CCCP), crystal violet, sodium dodecyl sulfate (SDS), and tetrachlorosalicylanilide (TCS). In the acquired resistance setting, however, it provides protection only against CCCP and TCS (Lin et al., 2014a). The TolCsm porin has been associated with the SmeOP efflux pump. The tolCsm gene is located upstream of the smeOP operon, in another operon known as smeRo-pcm-tolC. The ΔtolCsm phenotype increases susceptibility to several compounds (Huang et al., 2013b), although no correlation is seen with the ΔsmeOP phenotype. This suggests that the TolCsm porin is not exclusive to the SmeOP efflux pump (Huang et al., 2013b; Lin et al., 2014a).
Bioinformatic analyses have also identified two putative MFS-type tripartite efflux transporters (Crossman et al., 2008). One of these, emrCABsm, shows high homology with emrAB of E. coli (Lomovskaya and Lewis, 1992). In S. maltophilia, this pump is encoded by an operon of four genes that cover the three efflux pump components and a MarR-type regulator, emrRsm, which is expressed in the same direction. ΔemrRsm mutants show low susceptibility to nalidixic acid and CCCP due to the overexpression of the efflux pump, indicating emrRsm to act as a repressor (Huang et al., 2013a).
Although ABC-type transporters play a major role in Gram positive bacteria, they have also been found in Gram negative organisms, e.g., MsbA and MacAB in E. coli, VcaM in Vibrio cholerae, MacAB in Neisseria gonorrhoeae, and SmdAB in Serratia marcescens (Lin et al., 2014b). Two ABC efflux pumps, SmrA and MacABCsm, have also been described in S. maltophilia (Al-Hamad et al., 2009; Lin et al., 2014b). The SmrA pump has only been studied in the heterologous microorganism E. coli, in which it provides resistance to fluoroquinolones, tetracycline, doxorubicin and multiple dyes; its role in S. maltophilia remains unknown (Al-Hamad et al., 2009). The other ABC-type efflux pump, MacABCsm, is associated with intrinsic resistance to macrolides, aminoglycosides and polymyxins. Interestingly, ΔmacCsm mutant bacteria show lower susceptibility to polymyxins, aminoglycosides and macrolides than ΔmacAB mutants, suggesting that the MacABCsm efflux pump uses an alternative, still-unidentified porin (Lin et al., 2014b). This efflux pump is constitutively expressed, contributing toward oxidative and envelope stress tolerances and biofilm formation. The original function of the MacABCsm efflux pump may therefore have seen it involved in metabolism or adaptation to environmental changes (Lin et al., 2014b).
Bioinformatic analyses have predicted the existence of additional pumps. For example, the SmtcrA gene, which codes for a putative MFS pump, has been associated with tetracycline resistance. Whether any other components are required to make this pump work remains unknown (Chang et al., 2011).
The presence of active extrusion mechanisms cannot, however, explain the low susceptibility of S. maltophilia to all antibiotics, and indeed the S. maltophilia genome codes for several modifying enzymes responsible for ß-lactam and aminoglycoside resistance phenotypes. S. maltophilia possesses two inducible ß-lactamases: L1, a Zn2+-dependent metalloenzyme which can hydrolyze nearly all classes of β-lactams (though not monobactams), and L2, a serine active-site cephalosporinase (Avison et al., 2001). The expression of both enzymes is regulated by ampR (a LysR type regulator located upstream of the L2 gene), and induced by the presence of ß-lactam antibiotics. AmpR acts as a weak repressor of the L2 gene in the absence of the inducer, and as an activator in its presence. With respect to L1, AmpR is required both for basal and induced expression (Lin et al., 2009). Other expression-regulating mechanisms also influence one or the other of the two enzyme genes, without affecting the regulated expression of the other. However, the exact mechanism of this additional regulation system remains to be elucidated (Avison et al., 2002; Okazaki and Avison, 2008). Finally, the expression of these enzymes in S. maltophilia is also subject to a complex regulation network. The deletion of the ampN-ampG operon, which encodes a permease transporter, prevents the induction of β-lactamases (Huang et al., 2010), while the inactivation of mrcA, which is predicted to encode penicillin-binding protein 1 (PBP1a), or of ampD1, which encodes a cytoplasmic N-acetyl-muramyl-L-alanine amidase, causes the hyperproduction of L1/L2 β-lactamase (Yang et al., 2009; Lin et al., 2011).
S. maltophilia also encodes two aminoglycoside modifying enzymes, conferring low susceptibility to aminoglycoside antibiotics (with the exception of gentamicin). Gene aph (3′)-IIc encodes an aminoglycoside phosphotransferase (Okazaki and Avison, 2007), while aac (6′)-Iz codes for an N-aminoglycoside acetyltransferase. The latter has three alleles—aac(6′)-Iz, aac(6′)-Iaz and aac(6′)-Iam—that show more than 80% similarity (Li et al., 2003; Tada et al., 2014). The presence of other inactivating enzymes (2′N-acetyltransferase, streptomycin 3″phosphotransferase/kinase, spectinomycin phosphotransferase, and chloramphenicol acetyltransferase) might be responsible for the susceptibility phenotype of S. maltophilia (Crossman et al., 2008). More studies are needed, however, to confirm their function.
S. maltophilia shows low susceptibility to synthetic antibiotics such as quinolones. Mutations in topoisomerases, the quinolone target, have been related to the main quinolone resistant mechanism in all bacteria. However, no topoisomerase mutations have ever been identified in S. maltophilia (Ribera et al., 2002; Valdezate et al., 2002; Garcia-Leon et al., 2014b). In contrast, S. maltophilia quinolone resistance is owed to efflux pumps (Alonso and Martinez, 2000; Li et al., 2002; Chen et al., 2011; Garcia-Leon et al., 2014b) and to the quinolone resistance protein SmQnr. This protein has been associated with both intrinsic and acquired resistance in S. maltophilia by a still unknown mechanism (Sanchez and Martinez, 2010; Chang et al., 2011). Qnr forms a dimer with a structure similar to double stranded DNA (Vetting et al., 2006; Xiong et al., 2011). Then, it has been proposed that SmQnr would bind topoisomerases protecting them, similarly to what was described for Qnr encoded in plasmid (Tran and Jacoby, 2002; Tran et al., 2005).
There is limited information on the antibiotic resistance mechanisms operating in clinical S. maltophilia strains. To date, overexpression of efflux pumps SmeABC, SmeDEF and SmeVWX, and the presence of class 1 integrons with antibiotic resistance genes have been associated with low susceptibility in clinical strains (Alonso and Martinez, 2001; Liaw et al., 2010; Cho et al., 2012).
In addition to the antibiotic resistance genes described, microorganisms may possess a non-inheritable resistance mechanism known as phenotypic resistance. Some (or indeed all) of a bacterial population, may temporarily appear less susceptible to an antibiotic, without the appearance of any genomic differences. The factors responsible for phenotypic resistance might be good targets for novel treatments. However, our knowledge of the genes responsible for phenotypic resistance remains limited.
Biofilms are complex structures composed of an exopolysaccharide matrix, DNA and proteins, in which bacteria lie. They often affect clinical equipment such as catheters and other devices, from which they can be difficult to remove. The reduction in susceptibility to antibiotics afforded by biofilms is due to the difficulty of making the antibiotic come into contact with the bacteria, and these bacteria having a metabolic status different to that of their non-biofilm counterparts (a consequence of differences in the availability of nutrients and oxygen, etc.).
Different factors involved in biofilm formation have been studied in S. maltophilia. The deletion of different genes related to the regulation and structure of flagella, and to exopolysaccharide synthesis, affects biofilm formation (Huang et al., 2006; Kang et al., 2015). Other biofilm components, such as extracellular DNA, have been analyzed in microorganisms such as P. aeruginosa and Salmonella enterica (Mulcahy et al., 2008; Johnson et al., 2013), but the literature contains no information for S. maltophilia.
Bacteria showing low susceptibility to antibiotics but which are genetically identical to the susceptible strain can become persistent. Different genes involved in E. coli persistence include those coding for toxin/antitoxin systems and the PhoU regulator (Olivares et al., 2013). Although S. maltophilia persistence increases in chronic infections (Brooke, 2012), the mechanisms responsible for this phenotype remain unknown.
Post-transcriptional and post-translational regulation or modification can also alter antibiotic resistance. In S. maltophilia, however, little is known about this kind of regulation. Non-coding small RNAs (sRNA) and the RNA-binding Hfq protein have been related to post-transcriptional gene expression, and in E. coli an sRNA and Hfq have been associated with antibiotic susceptibility (Moon and Gottesman, 2009). In S. maltophilia, 60 sRNA candidates and a hfq gene have been identified, and a Δhfq mutant has been associated with changes in antibiotic susceptibility, biofilm production, motility and the expression of several sRNAs (Roscetto et al., 2012). However, further studies are needed to determine the role of sRNAs in antibiotic susceptibility.
The study of the intrinsic resistome could provide novel antibiotic targets and help predict events during treatment. The intrinsic resistome has been studied in P. aeruginosa (Fajardo et al., 2008) and E. coli (Tamae et al., 2008; Girgis et al., 2009; Liu et al., 2010), and many genes whose deletion affects the antibiotic susceptibility phenotype have been identified. Recently, the screening of an S. maltophilia insertion mutant library identified smeT, which codes for a well-known regulator of the SmeDEF efflux pump, and mutS, which has a role in the DNA mismatch repair system. Other genes, e.g., the 23S gene, with no obvious role in antibiotic susceptibility may however influence it (Bernardini, 2014). Further analyses are required to precisely determine the role and mechanism of action of these genes in S. maltophilia antibiotic resistance.
The presence of this great number of influencing genes and antibiotic resistance mechanisms in S. maltophilia renders the treatment of its infections complicated. Several antibiotics are currently in use, including synthetic antibiotics and antibiotic combinations, that help prevent the appearance of resistant mutants. A trimethoprim/sulfamethoxazole (cotrimoxazole) combination is used as a last treatment option. Cotrimoxazole resistance in S. maltophilia has been associated with the genes sul1 and sul2. These have been linked to the presence of class 1 integrons in plasmids in the main, but also in the chromosomal genome (Barbolla et al., 2004; Toleman et al., 2007). The presence of these genes, however, cannot explain all the cases of cotrimoxazole resistance recorded. Porin TolCsm deletion also increases cotrimoxazole susceptibility (Huang et al., 2013b), but further studies are required to determine whether other porins or efflux pumps are also involved.
How to avoid the antibiotic resistance? This has been a problem since the beginning of antibiotics use. The search of new antibiotics, inhibitors of efflux pumps (Leitner et al., 2011), new targets among genes form intrinsic resistome or use of combination of known antibiotics, as trimethoprim/sulfamethoxazole (described above), are some of the new strategies to avoid not only resistant strains but also the appearance of resistant mutants.
In summary, S. maltophilia possesses a great many antibiotic resistance mechanisms. Most of the genes involved were present in S. maltophilia before any use of antibiotics. For example, the efflux pump SmeDEF is associated with the ability of S. maltophilia to colonize plants, and its regulator SmeT is induced by plant-produced flavonoids (Garcia-Leon et al., 2014a); thus, the main function of the genes encoding them is unlikely to be the provision of antibiotic resistance. Other mechanisms might appear in the future, depending on antibiotic pressure, the emergence of mutations, and gene acquisition events. While resistance may benefit bacteria in the presence of antibiotics, in other situations it could impair growth, as has been described for the overexpression of the efflux pump SmeDEF (Alonso et al., 2004). Further, the fitness cost of acquired resistance in S. maltophilia determines whether new mechanisms are kept.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Adrian Burton is thanked for English editing assistance with the manuscript.
Alavi, P., Starcher, M. R., Thallinger, G. G., Zachow, C., Muller, H., and Berg, G. (2014). Stenotrophomonas comparative genomics reveals genes and functions that differentiate beneficial and pathogenic bacteria. BMC Genomics 15:482. doi: 10.1186/1471-2164-15-482
Al-Hamad, A., Upton, M., and Burnie, J. (2009). Molecular cloning and characterization of SmrA, a novel ABC multidrug efflux pump from Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 64, 731–734. doi: 10.1093/jac/dkp271
Alonso, A., and Martinez, J. L. (2000). Cloning and characterization of SmeDEF, a novel multidrug efflux pump from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 44, 3079–3086. doi: 10.1128/AAC.44.11.3079-3086.2000
Alonso, A., and Martinez, J. L. (2001). Expression of multidrug efflux pump SmeDEF by clinical isolates of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 45, 1879–1881. doi: 10.1128/AAC.45.6.1879-1881.2001
Alonso, A., Morales, G., Escalante, R., Campanario, E., Sastre, L., and Martinez, J. L. (2004). Overexpression of the multidrug efflux pump SmeDEF impairs Stenotrophomonas maltophilia physiology. J. Antimicrob. Chemother. 53, 432–434. doi: 10.1093/jac/dkh074
Alonso, A., Sanchez, P., and Martinez, J. L. (2000). Stenotrophomonas maltophilia D457R contains a cluster of genes from gram-positive bacteria involved in antibiotic and heavy metal resistance. Antimicrob. Agents Chemother. 44, 1778–1782. doi: 10.1128/AAC.44.7.1778-1782.2000
Alvarez-Ortega, C., Wiegand, I., Olivares, J., Hancock, R. E., and Martinez, J. L. (2010). Genetic determinants involved in the susceptibility of Pseudomonas aeruginosa to beta-lactam antibiotics. Antimicrob. Agents Chemother. 54, 4159–4167. doi: 10.1128/AAC.00257-10
Avison, M. B., Higgins, C. S., Ford, P. J., Von Heldreich, C. J., Walsh, T. R., and Bennett, P. M. (2002). Differential regulation of L1 and L2 beta-lactamase expression in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 49, 387–389. doi: 10.1093/jac/49.2.387
Avison, M. B., Higgins, C. S., Von Heldreich, C. J., Bennett, P. M., and Walsh, T. R. (2001). Plasmid location and molecular heterogeneity of the L1 and L2 beta-lactamase genes of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 45, 413–419. doi: 10.1128/AAC.45.2.413-419.2001
Barbolla, R., Catalano, M., Orman, B. E., Famiglietti, A., Vay, C., Smayevsky, J., et al. (2004). Class 1 integrons increase trimethoprim-sulfamethoxazole MICs against epidemiologically unrelated Stenotrophomonas maltophilia isolates. Antimicrob. Agents Chemother. 48, 666–669. doi: 10.1128/AAC.48.2.666-669.2004
Bernardini, A. (2014). Papel de la Respuesta a Choque Térmico en la Resistencia a Quinolonas de Stenotrophomonas maltophilia. Ph.D. thesis, Universidad Autónoma de Madrid.
Brooke, J. S. (2012). Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin. Microbiol. Rev. 25, 2–41. doi: 10.1128/CMR.00019-11
Chang, Y. C., Tsai, M. J., Huang, Y. W., Chung, T. C., and Yang, T. C. (2011). SmQnrR, a DeoR-type transcriptional regulator, negatively regulates the expression of Smqnr and SmtcrA in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 66, 1024–1028. doi: 10.1093/jac/dkr049
Chen, C. H., Huang, C. C., Chung, T. C., Hu, R. M., Huang, Y. W., and Yang, T. C. (2011). Contribution of resistance-nodulation-division efflux pump operon smeU1-V-W-U2-X to multidrug resistance of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 55, 5826–5833. doi: 10.1128/AAC.00317-11
Cho, H. H., Sung, J. Y., Kwon, K. C., and Koo, S. H. (2012). Expression of Sme efflux pumps and multilocus sequence typing in clinical isolates of Stenotrophomonas maltophilia. Ann. Lab. Med. 32, 38–43. doi: 10.3343/alm.2012.32.1.38
Crossman, L. C., Gould, V. C., Dow, J. M., Vernikos, G. S., Okazaki, A., Sebaihia, M., et al. (2008). The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol. 9, R74. doi: 10.1186/gb-2008-9-4-r74
Fajardo, A., Martinez-Martin, N., Mercadillo, M., Galan, J. C., Ghysels, B., Matthijs, S., et al. (2008). The neglected intrinsic resistome of bacterial pathogens. PLoS ONE 3:e1619. doi: 10.1371/journal.pone.0001619
Falagas, M. E., Kastoris, A. C., Vouloumanou, E. K., and Dimopoulos, G. (2009). Community-acquired Stenotrophomonas maltophilia infections: a systematic review. Eur. J. Clin. Microbiol. Infect. Dis. 28, 719–730. doi: 10.1007/s10096-009-0709-5
Garcia-Leon, G., Hernandez, A., Hernando-Amado, S., Alavi, P., Berg, G., and Martinez, J. L. (2014a). A function of the major quinolone resistance determinant of Stenotrophomonas maltophilia SmeDEF is the colonization of the roots of the plants. Appl. Environ. Microbiol. 80, 4559–4565. doi: 10.1128/AEM.01058-14
García-León, G., Ruiz De La Alegría Puig, C., García De La Fuente, C., Martínez-Martínez, L., Martínez, J. L., and Sánchez, M. B. (2015). High-level quinolone resistance is associated to the overexpression of SmeVWX in Stenotrophomonas maltophilia clinical isolates. Clin. Microbiol. Infect. 21, 464–467. doi: 10.1016/j.cmi.2015.01.007
Garcia-Leon, G., Salgado, F., Oliveros, J. C., Sanchez, M. B., and Martinez, J. L. (2014b). Interplay between intrinsic and acquired resistance to quinolones in Stenotrophomonas maltophilia. Environ. Microbiol. 16, 1282–1296. doi: 10.1111/1462-2920.12408
Girgis, H. S., Hottes, A. K., and Tavazoie, S. (2009). Genetic architecture of intrinsic antibiotic susceptibility. PLoS ONE 4:e5629. doi: 10.1371/journal.pone.0005629
Gould, V. C., Okazaki, A., and Avison, M. B. (2013). Coordinate hyperproduction of SmeZ and SmeJK efflux pumps extends drug resistance in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 57, 655–657. doi: 10.1128/AAC.01020-12
Hernandez, A., Ruiz, F. M., Romero, A., and Martinez, J. L. (2011). The binding of triclosan to SmeT, the repressor of the multidrug efflux pump SmeDEF, induces antibiotic resistance in Stenotrophomonas maltophilia. PLoS Pathog. 7:e1002103. doi: 10.1371/journal.ppat.1002103
Hu, R. M., Huang, K. J., Wu, L. T., Hsiao, Y. J., and Yang, T. C. (2008). Induction of L1 and L2 beta-lactamases of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 52, 1198–1200. doi: 10.1128/AAC.00682-07
Huang, T. P., Somers, E. B., and Wong, A. C. (2006). Differential biofilm formation and motility associated with lipopolysaccharide/exopolysaccharide-coupled biosynthetic genes in Stenotrophomonas maltophilia. J. Bacteriol. 188, 3116–3120. doi: 10.1128/JB.188.8.3116-3120.2006
Huang, Y. W., Hu, R. M., Chu, F. Y., Lin, H. R., and Yang, T. C. (2013a). Characterization of a major facilitator superfamily (MFS) tripartite efflux pump EmrCABsm from Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 68, 2498–2505. doi: 10.1093/jac/dkt250
Huang, Y. W., Hu, R. M., and Yang, T. C. (2013b). Role of the pcm-tolCsm operon in the multidrug resistance of Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 68, 1987–1993. doi: 10.1093/jac/dkt148
Huang, Y. W., Lin, C. W., Hu, R. M., Lin, Y. T., Chung, T. C., and Yang, T. C. (2010). AmpN-AmpG operon is essential for expression of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 54, 2583–2589. doi: 10.1128/AAC.01283-09
Johnson, L., Horsman, S. R., Charron-Mazenod, L., Turnbull, A. L., Mulcahy, H., Surette, M. G., et al. (2013). Extracellular DNA-induced antimicrobial peptide resistance in Salmonella enterica serovar Typhimurium. BMC Microbiol. 13:115. doi: 10.1186/1471-2180-13-115
Kang, X. M., Wang, F. F., Zhang, H., Zhang, Q., and Qian, W. (2015). Genome-wide identification of genes necessary for biofilm formation of nosocomial pathogen stenotrophomonas maltophilia reveals orphan response regulator FsnR is a critical modulator. Appl. Environ. Microbiol. 81, 1200–1209. doi: 10.1128/AEM.03408-14
Leitner, I., Nemeth, J., Feurstein, T., Abrahim, A., Matzneller, P., Lagler, H., et al. (2011). The third-generation P-glycoprotein inhibitor tariquidar may overcome bacterial multidrug resistance by increasing intracellular drug concentration. J. Antimicrob. Chemother. 66, 834–839. doi: 10.1093/jac/dkq526
Li, X. Z., Zhang, L., Mckay, G. A., and Poole, K. (2003). Role of the acetyltransferase AAC(6′)-Iz modifying enzyme in aminoglycoside resistance in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 51, 803–811. doi: 10.1093/jac/dkg148
Li, X. Z., Zhang, L., and Poole, K. (2002). SmeC, an outer membrane multidrug efflux protein of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 46, 333–343. doi: 10.1128/AAC.46.2.333-343.2002
Liaw, S. J., Lee, Y. L., and Hsueh, P. R. (2010). Multidrug resistance in clinical isolates of Stenotrophomonas maltophilia: roles of integrons, efflux pumps, phosphoglucomutase (SpgM), and melanin and biofilm formation. Int. J. Antimicrob. Agents 35, 126–130. doi: 10.1016/j.ijantimicag.2009.09.015
Lin, C. W., Huang, Y. W., Hu, R. M., Chiang, K. H., and Yang, T. C. (2009). The role of AmpR in regulation of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. Res. Microbiol. 160, 152–158. doi: 10.1016/j.resmic.2008.11.001
Lin, C. W., Huang, Y. W., Hu, R. M., and Yang, T. C. (2014a). SmeOP-TolCSm efflux pump contributes to the multidrug resistance of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 58, 2405–2408. doi: 10.1128/AAC.01974-13
Lin, C. W., Lin, H. C., Huang, Y. W., Chung, T. C., and Yang, T. C. (2011). Inactivation of mrcA gene derepresses the basal-level expression of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 66, 2033–2037. doi: 10.1093/jac/dkr276
Lin, Y. T., Huang, Y. W., Liou, R. S., Chang, Y. C., and Yang, T. C. (2014b). MacABCsm, an ABC-type tripartite efflux pump of Stenotrophomonas maltophilia involved in drug resistance, oxidative and envelope stress tolerances and biofilm formation. J. Antimicrob. Chemother. 69, 3221–3226. doi: 10.1093/jac/dku317
Lira, F., Hernandez, A., Belda, E., Sanchez, M. B., Moya, A., Silva, F. J., et al. (2012). Whole-genome sequence of Stenotrophomonas maltophilia D457, a clinical isolate and a model strain. J. Bacteriol. 194, 3563–3564. doi: 10.1128/JB.00602-12
Liu, A., Tran, L., Becket, E., Lee, K., Chinn, L., Park, E., et al. (2010). Antibiotic sensitivity profiles determined with an Escherichia coli gene knockout collection: generating an antibiotic bar code. Antimicrob. Agents Chemother. 54, 1393–1403. doi: 10.1128/AAC.00906-09
Lomovskaya, O., and Lewis, K. (1992). Emr, an Escherichia coli locus for multidrug resistance. Proc. Natl. Acad. Sci. U.S.A. 89, 8938–8942. doi: 10.1073/pnas.89.19.8938
Looney, W. J., Narita, M., and Muhlemann, K. (2009). Stenotrophomonas maltophilia: an emerging opportunist human pathogen. Lancet Infect. Dis. 9, 312–323. doi: 10.1016/S1473-3099(09)70083-0
Moon, K., and Gottesman, S. (2009). A PhoQ/P-regulated small RNA regulates sensitivity of Escherichia coli to antimicrobial peptides. Mol. Microbiol. 74, 1314–1330. doi: 10.1111/j.1365-2958.2009.06944.x
Mulcahy, H., Charron-Mazenod, L., and Lewenza, S. (2008). Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathog. 4:e1000213. doi: 10.1371/journal.ppat.1000213
Okazaki, A., and Avison, M. B. (2007). Aph(3′)-IIc, an aminoglycoside resistance determinant from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 51, 359–360. doi: 10.1128/AAC.00795-06
Okazaki, A., and Avison, M. B. (2008). Induction of L1 and L2 beta-lactamase production in Stenotrophomonas maltophilia is dependent on an AmpR-type regulator. Antimicrob. Agents Chemother. 52, 1525–1528. doi: 10.1128/AAC.01485-07
Olivares, J., Bernardini, A., Garcia-Leon, G., Corona, F., B Sanchez, M., and Martinez, J. L. (2013). The intrinsic resistome of bacterial pathogens. Front. Microbiol. 4:103. doi: 10.3389/fmicb.2013.00103
Rahmati-Bahram, A., Magee, J. T., and Jackson, S. K. (1996). Temperature-dependent aminoglycoside resistance in Stenotrophomonas (Xanthomonas) maltophilia; alterations in protein and lipopolysaccharide with growth temperature. J. Antimicrob. Chemother. 37, 665–676. doi: 10.1093/jac/37.4.665
Ribera, A., Domenech-Sanchez, A., Ruiz, J., Benedi, V. J., Jimenez De Anta, M. T., and Vila, J. (2002). Mutations in gyrA and parC QRDRs are not relevant for quinolone resistance in epidemiological unrelated Stenotrophomonas maltophilia clinical isolates. Microb. Drug Resist. 8, 245–251. doi: 10.1089/10766290260469499
Rocco, F., De Gregorio, E., Colonna, B., and Di Nocera, P. P. (2009). Stenotrophomonas maltophilia genomes: a start-up comparison. Int. J. Med. Microbiol. 299, 535–546. doi: 10.1016/j.ijmm.2009.05.004
Roscetto, E., Angrisano, T., Costa, V., Casalino, M., Forstner, K. U., Sharma, C. M., et al. (2012). Functional characterization of the RNA chaperone Hfq in the opportunistic human pathogen Stenotrophomonas maltophilia. J. Bacteriol. 194, 5864–5874. doi: 10.1128/JB.00746-12
Sanchez, M. B., Hernandez, A., and Martinez, J. L. (2009). Stenotrophomonas maltophilia drug resistance. Future Microbiol. 4, 655–660. doi: 10.2217/fmb.09.45
Sanchez, M. B., and Martinez, J. L. (2010). SmQnr contributes to intrinsic resistance to quinolones in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 54, 580–581. doi: 10.1128/AAC.00496-09
Sanchez, P., Alonso, A., and Martinez, J. L. (2002). Cloning and characterization of SmeT, a repressor of the Stenotrophomonas maltophilia multidrug efflux pump SmeDEF. Antimicrob. Agents Chemother. 46, 3386–3393. doi: 10.1128/AAC.46.11.3386-3393.2002
Sanchez, P., Moreno, E., and Martinez, J. L. (2005). The biocide triclosan selects Stenotrophomonas maltophilia mutants that overproduce the SmeDEF multidrug efflux pump. Antimicrob. Agents Chemother. 49, 781–782. doi: 10.1128/AAC.49.2.781-782.2005
Tada, T., Miyoshi-Akiyama, T., Dahal, R. K., Mishra, S. K., Shimada, K., Ohara, H., et al. (2014). Identification of a novel 6′-N-aminoglycoside acetyltransferase, AAC(6′)-Iak, from a multidrug-resistant clinical isolate of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 58, 6324–6327. doi: 10.1128/AAC.03354-14
Tamae, C., Liu, A., Kim, K., Sitz, D., Hong, J., Becket, E., et al. (2008). Determination of antibiotic hypersensitivity among 4,000 single-gene-knockout mutants of Escherichia coli. J. Bacteriol. 190, 5981–5988. doi: 10.1128/JB.01982-07
Toleman, M. A., Bennett, P. M., Bennett, D. M., Jones, R. N., and Walsh, T. R. (2007). Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerging Infect. Dis. 13, 559–565. doi: 10.3201/eid1304.061378
Tran, J. H., and Jacoby, G. A. (2002). Mechanism of plasmid-mediated quinolone resistance. Proc. Natl. Acad. Sci. U.S.A. 99, 5638–5642. doi: 10.1073/pnas.082092899
Tran, J. H., Jacoby, G. A., and Hooper, D. C. (2005). Interaction of the plasmid-encoded quinolone resistance protein Qnr with Escherichia coli DNA gyrase. Antimicrob. Agents Chemother. 49, 118–125. doi: 10.1128/AAC.49.1.118-125.2005
Vaara, M. (1993). Outer membrane permeability barrier to azithromycin, clarithromycin, and roxithromycin in gram-negative enteric bacteria. Antimicrob. Agents Chemother. 37, 354–356. doi: 10.1128/AAC.37.2.354
Valdezate, S., Vindel, A., Echeita, A., Baquero, F., and Canto, R. (2002). Topoisomerase II and IV quinolone resistance-determining regions in Stenotrophomonas maltophilia clinical isolates with different levels of quinolone susceptibility. Antimicrob. Agents Chemother. 46, 665–671. doi: 10.1128/AAC.46.3.665-671.2002
Vetting, M. W., Hegde, S. S., Fajardo, J. E., Fiser, A., Roderick, S. L., Takiff, H. E., et al. (2006). Pentapeptide repeat proteins. Biochemistry 45, 1–10. doi: 10.1021/bi052130w
Xiong, X., Bromley, E. H., Oelschlaeger, P., Woolfson, D. N., and Spencer, J. (2011). Structural insights into quinolone antibiotic resistance mediated by pentapeptide repeat proteins: conserved surface loops direct the activity of a Qnr protein from a Gram-negative bacterium. Nucleic Acids Res. 39, 3917–3927. doi: 10.1093/nar/gkq1296
Yang, T. C., Huang, Y. W., Hu, R. M., Huang, S. C., and Lin, Y. T. (2009). AmpDI is involved in expression of the chromosomal L1 and L2 beta-lactamases of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 53, 2902–2907. doi: 10.1128/AAC.01513-08
Keywords: Stenotrophomonas maltophilia, antibiotic resistance, multidrug resistance, intrinsic resistome, phenotypic resistance
Citation: Sánchez MB (2015) Antibiotic resistance in the opportunistic pathogen Stenotrophomonas maltophilia. Front. Microbiol. 6:658. doi: 10.3389/fmicb.2015.00658
Received: 04 March 2015; Accepted: 16 June 2015;
Published: 30 June 2015.
Edited by:
Joanna S. Brooke, DePaul University, USAReviewed by:
Paras Jain, Albert Einstein College of Medicine, New York, USACopyright © 2015 Sánchez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María B. Sánchez, Departamento de Biotecnología Microbiana, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas, Darwin 3, Cantoblanco, 28049 Madrid, Spain,YnNhbmNoZXpAY25iLmNzaWMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.