 Michelle Z. Tadra-Sfeir1
Michelle Z. Tadra-Sfeir1 Helisson Faoro1,2
Helisson Faoro1,2 Doumit Camilios-Neto3
Doumit Camilios-Neto3 Liziane Brusamarello-Santos1
Liziane Brusamarello-Santos1 Eduardo Balsanelli1
Eduardo Balsanelli1 Vinicius Weiss1
Vinicius Weiss1 Valter A. Baura1
Valter A. Baura1 Roseli Wassem4
Roseli Wassem4 Leonardo M. Cruz1
Leonardo M. Cruz1 Fábio De Oliveira Pedrosa1
Fábio De Oliveira Pedrosa1 Emanuel M. Souza1
Emanuel M. Souza1 Rose A. Monteiro1*
Rose A. Monteiro1*- 1Nitrogen Fixation group, Department of Biochemistry and Molecular Biology, Universidade Federal do Paraná, Curitiba, Brazil
- 2Instituto Carlos Chagas, Fundação Oswaldo Cruz, Fiocruz-PR, Curitiba, Brazil
- 3Department of Biochemistry and Biotechnology, Universidade Estadual de Londrina, Londrina, Brazil
- 4Department of Genetics, Universidade Federal do Paraná, Curitiba, Brazil
Herbaspirillum seropedicae is a diazotrophic bacterium which associates endophytically with economically important gramineae. Flavonoids such as naringenin have been shown to have an effect on the interaction between H. seropedicae and its host plants. We used a high-throughput sequencing based method (RNA-Seq) to access the influence of naringenin on the whole transcriptome profile of H. seropedicae. Three hundred and four genes were downregulated and seventy seven were upregulated by naringenin. Data analysis revealed that genes related to bacterial flagella biosynthesis, chemotaxis and biosynthesis of peptidoglycan were repressed by naringenin. Moreover, genes involved in aromatic metabolism and multidrug transport efllux were actived.
Introduction
Beneficial plant–bacterial interactions promote plant growth and development. During this process molecular changes occur in both partners, and signal molecules are involved in partner communication. In legume-Rhizobium interactions, flavonoids released by plant roots induce sets of genes involved in nodulation (Broughton et al., 2000). In addition, flavonoids seem to play a role in other plant bacterial associations. Naringenin stimulates lateral root crack (LRC) colonization of Arabidopsis thaliana by Azorhizobium caulinodans and H. seropedicae a process independent of the nod genes (Gough et al., 1997). This flavonoid, secreted by some plants, is a signal molecule that regulates gene expression in bacteria such as H. seropedicae (Tadra-Sfeir et al., 2011) and A. caulinodans (Webster et al., 1998).
The diazotroph H. seropedicae is frequently found in endophytic association with maize (Zea mays), rice (Oryza sativa), sorghum (Sorghum bicolor), sugar cane (Saccharum officinarum) and other plants. Inoculation of rice with H. seropedicae strains resulted in plant growth promotion and increase in productivity (Baldani et al., 2000; Gyaneshwar et al., 2002). This effect may be due in part to transfer of fixed nitrogen, since 15N dilution assays indicate significant N transfer to the host plant (Baldani et al., 2000), and production of phytohormones by the bacteria (Bastián et al., 1998). However, the bacterial genes necessary for the establishment of endophytic interaction and the molecular cues that direct their expression are largely unkown. Previously, we isolated 16 H. seropedicae mutant strains in genes regulated by the plant-derived flavonoid naringenin; 12 of these were downregulated and 4 upregulated. Four of these genes are involved in the synthesis of the outer membrane of the cell wall, suggesting that changes in the cell surface probably occur during the interaction between H. seropedicae and its host plants (Tadra-Sfeir et al., 2011).
To explore which other genes are regulated by naringenin, we determined the transcriptional profile of H. seropedicae grown in NFBHP malate medium in the presence or absence of naringenin using RNA-seq.
Materials and Methods
Bacterial Growth
H. seropedicae SmR1, a streptomycin resistant strain, was grown at 30°C and 120 rpm in NFbHPN medium (Klassen et al., 1997) in the presence (+Nar) or absence (–Nar) of 100 μM of naringenin containing streptomycin (80 μg.mL−1)for 6 h (optical density at 600 nm of 0.8).
Transcriptome Profiling Experiments Design and Analyses
The total RNA was isolated using RiboPure™-Bacteria Kit (Ambion) and treated with DNase I (Ambion) for removal of the remaining genomic DNA. Seven micrograms of total RNA were rRNA-depleted using two rounds of the MICROBExpress™ Bacterial mRNA Enrichment Kit (Ambion). The efficiency of the depletion was evaluated in agarose gel 1% and all RNA preparations were quantified with a Nanodrop 1000 spectrophotometer. After rRNA depletion, 500 ng of depleted rRNA was used to construct the sequencing libraries following standard protocols using the SOLiD Total RNA-Seq Kit (Life Technologies). The libraries were barcoded by using the SOLiD Transcriptome Multiplexing Kit (Life Technologies). The emulsion PCR and SOLiD sequencing were performed according to standard Life Technologies protocols. Two independent samples were used to prepare replicate libraries resulting in a total of 4 libraries. Mapping of the reads against the H. seropedicae genome sequence, data processing and statistical analysis were performed using the CLC Genomics Workbench 5.1 and the results were expressed in RPKM (Reads Per Kilobase of exon model per Million mapped reads) (Mortazavi et al., 2008). The sequence data are available in the ArrayExpress database (www.ebi.ac.uk/arrayexpress) under accession number E-MTAB-3435.
A gene was considered expressed when read coverage was equal to or higher than 3-fold, and differentially expressed when RPKM value was 2-fold higher/smaller in +Nar compared to -Nar and p-value higher than 0.05 by the Baggerley's test as implemented in CLC Workbench. The Baggerley's test (Baggerly et al., 2003) compares the proportion of counts in a group of replicates (+Nar) against those of another group of replicates (−Nar), comprising a weighted t-type test statistic. The samples are given different weights depending on their sizes (total counts). The weights are obtained by assuming a Beta distribution on the proportions in a group, and estimating these, along with the proportion of a binomial distribution, by the method of moments. RNAseq statistical analyses were also made using the R package DESeq, which performs a negative binomial distribution and a shrinkage estimator for the distribution's variance and size-factor normalization (Anders and Huber, 2010).
Motility Assay
The motility assay was performed on NFbHPN-malate semi-solid agar (0.25%) plates supplemented with 100 μM naringenin. Overnight culture of H. seropedicae was inoculated in the center of the plate and incubated at 30°C. The plates were photographed after 12 h and motility halos were measured by using ImageJ (Rasband, 1997). Ten replicates with 108 bacteria were inoculated in both plates.
RT-qPCR
For validation with RT-qPCR, total RNA was isolated from cultures grown in the presence and absence of naringenin (100 μM) using the Ribominus (Ambion), the cDNAs were synthesized using the High-capacity cDNA Reverse Transcription kit (Applied Biosystems), and quantified using the Power SYBR-Green PCR Master Mix on a Step One Plus Real Time-PCR System (Applied Biosystems). The Primer express 3.0 software was used to design the primers. The 16S rRNA gene was used as internal control, and the relative gene expression was determined using the 2−ΔΔCt method (Livak and Schmittgen, 2008).
Colonization Assay
H. seropedicae SmR1 cells were grown in NFbHPN medium at 30°C, 120 rpm, until OD600nm = 1. The culture was diluted to OD600nm = 0.2 in fresh medium, and grown in the same conditions in the presence or absence of 100 μM naringenin for 6 h. These cells were washed with saline buffer and 105 cells were inoculated per maize plantlet (samples in triplicate). Quantification of bacterial root endophytic populations was performed according to Balsanelli et al. (2013), every 12 h until 3 days after inoculation. Endophytic bacteria at time zero is too low to count (REF).
Results and Discussion
Early Herbaspirillum seropedicae Smr1 Maize Root Colonization Is Stimulated by Naringenin
Flavonoids constitute a large part of root exudates (Cesco et al., 2010), being involved in root colonization. Previously results showed that the plant-derived flavonoid naringenin regulated the expression of H. seropedicae genes. Maize roots were inoculated with H. seropedicae in the presence of naringenin (100 μM) to determine the effect of this flavonoid in colonization pattern. The results showed that the endophytic population in the first 36 h is higher in the presence of naringenin (Figure 1). After 36 h the endophytic population is the same in the absence and in the presence of naringenin. These results indicate that naringenin affects early endophytic colonization. Naringenin stimulation of root colonization was also observed during the colonization of Arabidopsis thaliana by H. seropedicae (Gough et al., 1997), and wheat by Azorhizobium caulinodans (Webster et al., 1998).
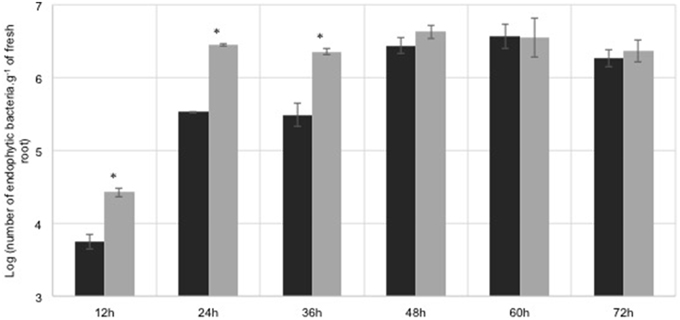
Figure 1. Maize root endophytic colonization by H. seropedicae wild-type. H. seropedicae SmR1 cells were grown in the presence (gray bars) or absence (black bars) of 100 μM naringenin for 6 h, and 105 cells were inoculated on maize plantlets. The number of root endophytic bacteria was determined after the periods indicated. Results are shown as means of Log10 (number of endophytic bacteria.g−1 of fresh root) ± standard deviation. Asterisk indicates significant differences at p < 0.0083 (Student t-test with Bonferroni correction) of endophytic colonization between naringenin treated and non-treated bacteria.
Changes in the Herbaspirillum seropedicae SmR1 Transcriptome in Response to Naringenin
RNA-seq profiling of H. seropedicae cells grown in the presence (+Nar) or absence (−Nar) of 100 μM of naringenin for 6 h was performed as described in Bacterial growth in Material and Methods. Sixty four million and fifty eight million reads were obtained for −Nar and +Nar conditions, respectively, and of those 2.7 million and 2.5 million were mapped uniquely to the H. seropedicae genome. As expected, biological replicates showed a very high level of correlation (r2 > 0.97) (Table 1), thus all the libraries of each condition were used for further analysis.

Table 1. Summary of RNA-seq data.
The genes that showed fold change greater than 2.0 (+Nar relative to −Nar) and a p = 0.05 were considered to be regulated by naringenin. Three hundred and four genes were downregulated and 77 were upregulated by naringenin by CLC Workbench. Fifty-three percent of these were also differentially regulated by the DESeq analysis (Table S1). The regulation of flhB, flgE, Hsero_2564 and murF genes are confirmed by qPCR (Table 2). Some genes with fold changes marginally lower than 2.0 fold or p higher than 0.05 were also considered regulated if neighborhood analysis suggested that they are part of an operon with genes regulated according to the previous criteria. The majority of the upregulated genes belong to the following functional gene categories: lipid transport and metabolism, energy production and conversion, inorganic ion transport and metabolism and unknown function (Figure 2A). The downregulated genes belong mainly to the categories aminoacid transport and metabolism, carbohydrate transport and metabolism, motility and unknown function (Figure 2B).
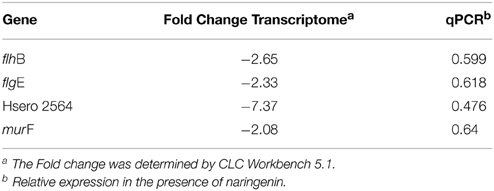
Table 2. Genes differentially expressed in the presence of naringenin.
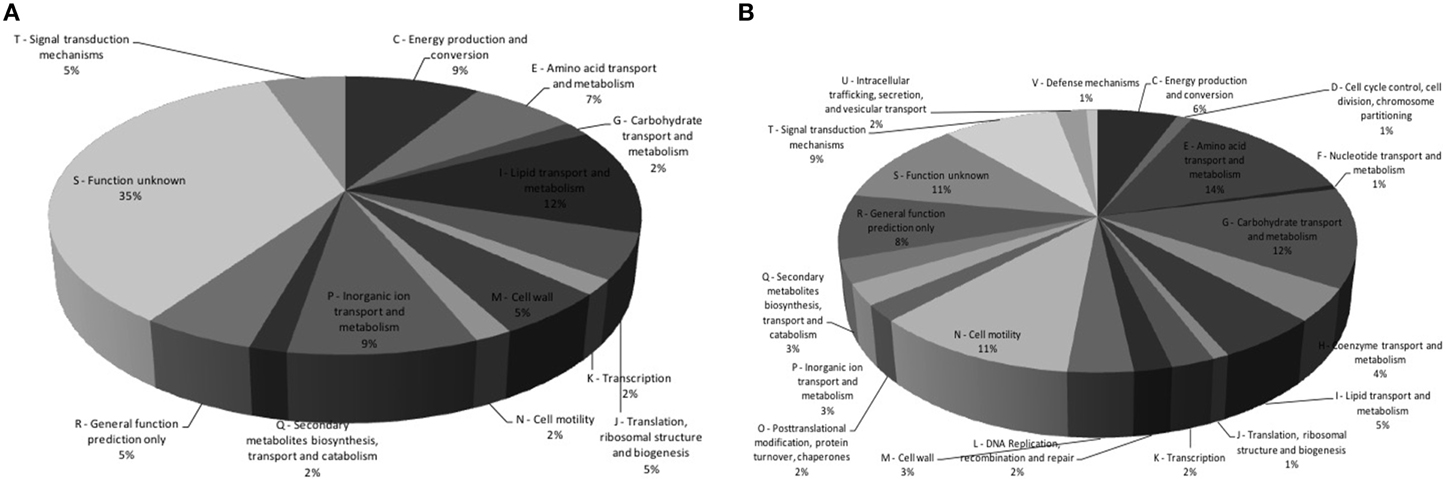
Figure 2. Functional classification of genes upregulated and downregulated by naringenin. Three hundred and four genes were upregulated (A) and 77 were downregulated (B) in the presence of naringenin. The genes were functional classificated by COG (Clusters if Orthologous Groups of proteins Tatusov et al., 1997) (http://www.ncbi.nlm.nih.gov/COG).
We found some groups of genes whose expression were regulated based on both CLC Workbench and DESeq analyses by naringenin that may be involved in plant-bacterial interaction. These genes are mainly involved with cell wall and motility. We also noticed an increase in the expression of genes that could be involved in naringenin degradation.
H. seropedicae Cell Wall Is Altered in the Presence of Naringenin
Peptidoglycan is an essential component for synthesis of the bacterial cell wall and the biosynthesis of this molecule is a complex process that proceeds in several stages (Ramos et al., 2004; Patin et al., 2010; Muchova et al., 2011). The assembly of the peptidoglycan involves, the Mur ligases (MurC, MurD, MurE, and MurF) that catalyze the first step of the synthesis the UDP-N-acetylmuramoyl-pentapeptide precursor (Patin et al., 2010). Three genes that encode the enzymes MurC, MurD, and MurF were downregulated in the presence of naringenin (Figure 3). Other mur genes were also inhibited by naringenin such as genes encoding the enzymes MraY and MurG (Figure 3). MraY catalyzes the transfer of the phospho-N-acetyl-muramoyl-pentapeptide from UDP-N-acetyl-muramoyl-pentapeptide to a membrane acceptor to form lipid I. The final step of the peptidoglycan subunit biosynthesis is the addition of N-acetylglucosamine (GlcNAc) to lipid I catalyzed by MurG producing lipid II (Muchova et al., 2011).
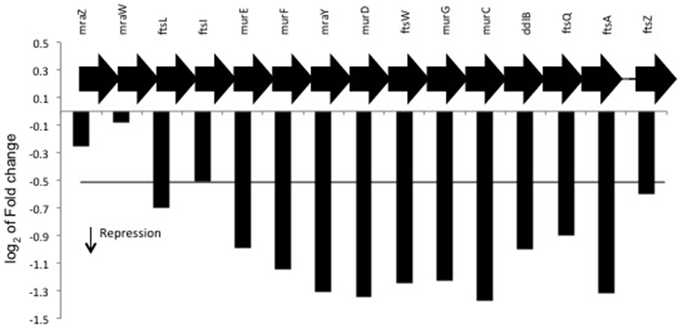
Figure 3. Differential expression of region of mur genes in the presence of naringenin. Region of mur genes in H. seropedicae genome. The value represents the log2 of fold change of genes.
Interestingly the genes ddlB and ftsQAZ were also downregulated by naringenin and found in the same operon as the mur genes (Figure 3, Figure S1A). D-Alanine–D-alanine ligase (coded by ddlB) is an enzyme involved in peptidoglycan biosynthesis and the proteins FtsQ, FtsA, and FtsZ are involved in septum formation in cell-division (Jofré et al., 2009). Mutations in ddlB and ftsQAZ genes in Azospirillum brasilense resulted in overproduction of exopolysaccharides, decreased bacterial tolerance to saline stress and alteration in cell morphology (Jofré et al., 2009).
A decrease in level of peptidoglycan synthesis enzyme GlmU was observed in the proteome of H. seropedicae grown in the presence of sugarcane extract (Cordeiro et al., 2013). In our study the expression of glmU was slightly decreased (−1.3-fold, p = 0) in the presence of naringenin, a decrease similar to that was observed by RT-PCR (-1.6-fold) in H. seropedicae grown in the presence of the sugarcane extract (Cordeiro et al., 2013). Peptidoglycan may act as elicitors of plant innate immunity, being recognized as a microbe-associated molecular pattern (MAMP) (Erbs and Newman, 2012). For example purified peptidoglycans from Xanthomonas campestris pv. Campestris and Agrobacterium tumefaciens act as MAMPs, inducing immune responses in Arabidopsis thaliana. The results suggest that the H. seropedicae is capable of controlling peptidoglycan synthesis in response to plant signals. A similar strategy has been described for Listeria monocytogenes that can N-deacetylate its peptidoglycan, avoiding the recognition and killing by host cells (Boneca et al., 2007). Agrobacterium tumefaciens also alters its peptidoglycan to reduce elicitation of plant defense (Erbs et al., 2008). Tadra-Sfeir et al. (2011) showed that the ampG gene of H. seropedicae is downregulated by naringenin), and the mutation in this gene alters the cell morphology. ampG codes for a muropeptide permease that is involved in the recycling of peptidoglycan. In the present transcriptome analysis the expression of this gene was slightly decreased (−1.2). It is possible that this difference is due to distinct growth condition used in the present work.
The genes rfbG galE rfbBC and wcaGA involved in lipopolyssacharide (LPS) biosynthesis were also downregulated by naringenin. Alterations in cell surface are common in other bacteria when they interact with plants or in the presence of plant compounds. Rhizobium sp. strain NGR234 synthesizes a new LPS in the presence of flavonoids and this LPS is important for the colonization of NGR234 in leguminous plants (Ardissone et al., 2011).
Chemotaxis and Flagella
Bacteria can sense the environment and rapidly respond to environmental changes through the action of specific signaling pathways. The chemotaxis signal begins with the binding of molecules on membrane receptors. Chemoreceptors are encoded by the tsr, tar, trg and tap genes, that code for methyl-accepting chemotaxis proteins (MCPs) (Pereira et al., 2004).
We identified forty-one genes involved in the chemotaxis transduction pathways in Herbaspirillum seropedicae genome. Twenty nine of these are found in five clusters and the other genes are monocistronics with 6 homologous to cheA, 5 to cheB, 10 to cheD, 5 to cheR, 1 to cheM, 9 to cheY, 1 to cheZ, and 5 to cheW (Pedrosa et al., 2011). Cluster I has five genes, Hsero_0623 (methyl-accepting chemotaxis transducer transmembrane protein), cheWRB and the Hsero_0627. Cluster III of H. seropedicae contains tar, cheRDBYZ and flhBA, three cheD-like and eight genes coding for methyl-accepting chemotaxis proteins (tsr, Hsero_0538, 1262, 1556, 3234, 4019, 4543, and 4615) all these were repressed in the presence of naringenin. In contrast the cheR and cheY genes of the cluster II cheWBRYA were activated in the presence of the flavonoid. Cluster IV has cheYAW genes and the cluster V contains eight genes, one encoding a methyl-accepting chemotaxis protein (Hsero_3022), one a chemotaxis signal transduction protein (Hsero_3021), one a response regulator protein (Hsero_3016), one an acyl dehydratase protein (Hsero_3015), and the genes cheRWAB. The expression of the gene clusters I, IV, and V did not change in the presence of naringenin. In Pseudomonas aeruginosa the che genes are also organized in five gene clusters, which have different responses depending on the stimulus (Ferrandez et al., 2002; Guvener et al., 2006).
In bacteria, flagella genes are regulated in response to environmental changes. These genes are found in operons that are divided in three temporally regulated transcriptional classes: early (class 1), middle (class 2), and late (class 3) (Komeda, 1986; Kutsukake et al., 1994). H. seropedicae has at least 46 genes involved in flagella biosynthesis, assembly and structure. H. seropedicae early genes homologous to the class 1 flhC and flhD, to middle genes class 2 flgA, flgBCDEFGHI, flhB, fliA, fliD, fliFGHIJK, fliOPQR, and late genes of class 3 tsr, cheA, cheRBYZ are all repressed in the presence of naringenin (Figure 4, Figure S1B). The FlhCD proteins are sigma 70 dependent transcriptional activators of class 2 promoters (Kutsukake et al., 1994; Liu and Matsumura, 1994) and FliA protein is a flagella alternative sigma factor – σ28 (Ohnishi et al., 1990). FliA has been shown to be involved in transcription of flagella, chemotaxis, and motility genes and the decrease in the expression of this protein could be responsible for the decrease in expression of middle and late flagella genes and chemotaxis genes (Iriarte et al., 1995).
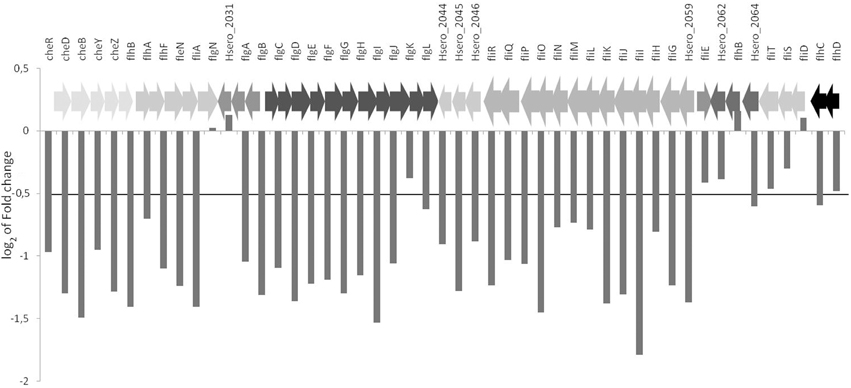
Figure 4. Differential expression of flagellar genes in the presence of naringenin. The value represents the log2 of fold change of genes. Genes of the same color are in the same operons.
Naringenin regulates flagellar genes expression in other bacteria. The transcriptome profile of Salmonella typhimurium LT2 revealed that 24 genes of pathogenicity island 1 and 17 genes involved in flagellar and motility were repressed in the presence of naringenin (Vikram et al., 2011). Flagellar genes were also downregulated in A. caulinodans by naringenin (Tsukada et al., 2009) and Pseudomonas syringae pv. tomato DC3000 by phloretin (Vargas et al., 2013). In Bacillus subtilis OKB105 chemotaxis and motility genes were downregulated in response to rice seedlings (Xie et al., 2015), suggesting that decrease of motility in the presence of root exudate maybe be involved in the establishment of interaction with the plant. Signal molecules released by plants direct the bacteria toward the root in a process dependent on chemotaxis and cell motility. At this initial stage the concentration of signal compounds are low and the bacteria follow a concentration gradient and bacteria motility depends on flagella-driven motility. Upon reaching root surface the bacteria would attach and reduce flagella gene expression and motility may depend on other means more appropriate for this new environment, such as type IV pilli-dependent swarming. Alternatively flavonoids such as naringenin acting as an anti-microbial agent targets the flagella, as suggested for P. syringae (Vargas et al., 2013).
Downregulation of flagella and chemotaxis genes indicated that naringenin might reduce H. seropedicae motility. This hypothesis was tested by measuring the motility of H. seropedicae in the presence of naringenin. The results demonstrated that naringenin impairs H. seropedicae motility (Figure 5B) without affecting the growth in liquid medium (Figure 5A).
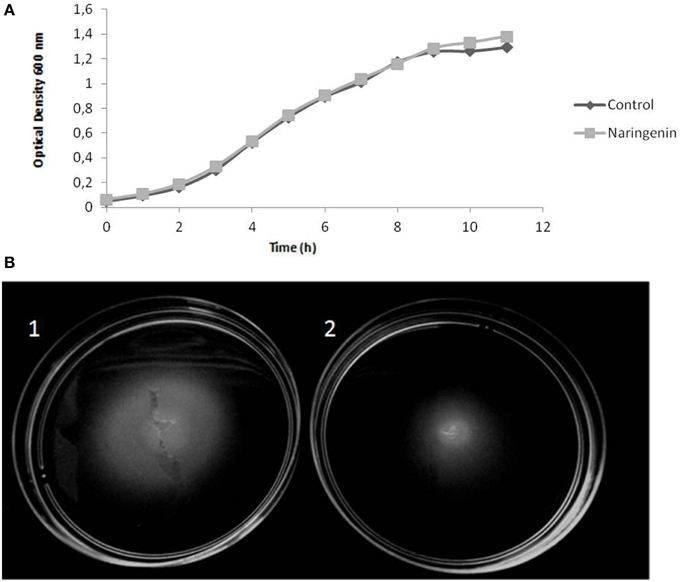
Figure 5. Motility of H. seropedicae SmR1 is reduced by the presence of naringenin. (A) Growth curve of H. seropedicae SmR1 in liquid NFbHPN-malate. (B) Motility of H. seropedicae SmR1 in semi-solid NFbHPN-malate in the absence (1) or presence (2) of 100 μM naringenin. Growth halos in semi-solid medium of ten replicates were analyzed by ImageJ software. Control = 1.00 ± 0.15; with naringenin = 0.70 ± 0.14, the values are arbitrary units in relation to the mean of the control condition.
Aromatic Compounds Metabolism is Induced by Naringenin
H. seropedicae genome sequence analysis showed the existence of genes involved in aromatic compounds metabolism such as catechol (cat), benzoate (ben), 4-hydroxybenzoate (pob), phenylacetate (paa), and protocatechuate (pca) metabolism (Pedrosa et al., 2011). These compounds can be metabolized to tricarboxylic acid intermediates.
In H. seropedicae the expression of pcaJIF and catCD genes increased by 3 -7 fold in the presence of naringenin, these genes are involved in the conversion of muconolactone to 3-oxoadipyl-Coa a pathway of the catechol metabolism. The expression of paaBC and Hsero_4130 also increased in the presence of naringenin. These three genes are probably organized in an operon with other paa genes. The paa, pca, and cat genes may be involved in naringenin intermediates degradation.
Another important system influenced by naringenin treatment was the multidrug efflux. H. seropedicae genome has five regions containing genes acrAB; these regions were named Cluster I, II, III, IV, and V. In this study, acrA and acrB of the cluster II were induced 7.5 and 3.2-fold, respectively. In S. typhimurium LT2 the genes acrAB were also induced 3-fold in the presence of naringenin (Vikram et al., 2010). Other genes involved in multidrug transport efflux were also induced by naringenin such as ompC (2.4-fold) and Hsero_1358 (3.15-fold). Multidrug transporter efflux pump provides low level of resistance to alkaline dyes, detergents and antibiotics. Induction of this efflux pump by naringenin suggests activation of the drug resistance system.
We found 35 ABC-transporter gene clusters differentially expressed in the presence of naringenin, being 29 downregulated and 6 upregulated (Table S1). The main group downregulated are amino acid and sugar transporter with a putative aromatic amino acid transporter system (~7-fold down-regulated) and a sugar transporter repressed 4-fold. Among the upregulated ABC transporter we found mainly anion transporters such as sulfate, phosphate and alkanesulfanates. A proteomics approach revealed that Bradyrhizobium japonicum strains differentially expressed three periplasmic amino acid binding proteins of ABC-transporter systems in the presence of genistein (Batista and Hungria, 2012). The reason for this effect is not known.
In this study we provided a comprehensive view of a H. seropedicae transcriptome in the presence of naringenin. We have defined in detail the RNA populations found in H. seropedicae in the presence and absence of the flavonoid. The data obtained from this study enabled us to infer some aspects of the metabolism of the bacteria in the presence of naringenin. Expression of genes related to bacterial flagella biosynthesis, flagella motor activity, and chemotaxis were repressed by naringenin, and this repression is predicted to have negative effects on flagella synthesis and bacterial motility. Biosynthesis of peptidoglycan is also inhibited by narigenin, whereas expression of a multidrug transport efflux pump is activated. The data suggest that in the presence of naringenin H. seropedicae triggers a concerted change in gene expression probably related to defense mechanisms.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the Brazilian Program of National Institutes of Science and Technology-INCT/Brazilian Research Council-CNPq/MCT, Fundação Araucaìria and CAPES. We would like to thank Dr. Marshall Geoffrey Yates for reading the manuscript. We are also thankful to Roseli A. Prado, Julieta Pie and Marilza D. Lamour for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.00491/abstract
References
Anders, S., and Huber, W. (2010). Differential expression analysis for sequence count data. Genome Biol. 11:R106. doi: 10.1186/gb-2010-11-10-r106
Ardissone, S., Kobayashi, H., Kambara, K., Rummel, C., Noel, K. D., Walker, G. C., et al. (2011). Role of BacA in lipopolysaccharide synthesis, peptide transport and nodulation by Rhizobium sp. NGR234. J. Bacteriol. 193, 2218–2228. doi: 10.1128/JB.01260-10
Baggerly, K., Deng, L., Morris, J., and Aldaz, C. (2003). Differential expression in SAGE: accounting for normal between-library variation. Bioinformatics 19, 1477–1483. doi: 10.1093/bioinformatics/btg173
Baldani, V. L. D., Baldani, J. I., and Dobereiner, J. (2000). Inoculation of rice plants with the endophytic diazotrophs Herbaspirillum seropedicae and Burkholderia spp. Biol. Fertil. Soils 30, 485–491. doi: 10.1007/s003740050027
Balsanelli, E., Tuleski, T. R., Baura, V. A., Yates, M. G., Chubatsu, L. S., Pedrosa, F. O., et al. (2013). Maize root lectins mediate the interaction with Herbaspirillum seropedicae via N-acetyl glucosamine residues of lipopolysaccharides. PLoS ONE 8:e77001. doi: 10.1371/journal.pone.0077001
Bastián, F., Cohen, A., Piccoli, P., Luna, V., Baraldi, R., and Bottini, R. (1998). Production of indole-3-acetic acid and gibberellins A1 and A3 by Acetobacter diazotrophicus and Herbaspirillum seropedicae in chemically-defined culture media. Plant Growth Reg. 24, 7–11.
Batista, J. S. S., and Hungria, M. (2012). Proteomics reveals differential expression of proteins related to a variety of metabolic pathways by genistein-induced Bradyrhizobium japonicum strains. J. Proteomics 75, 1211–1219. doi: 10.1016/j.jprot.2011.10.032
Boneca, I. G., Dussurget, O., Cabanes, D., Nahori, M. A., Sousa, S., Lecuit, M., et al. (2007). A critical role for peptidoglycan N-deacetylation in Listeria evasion from the host innate immune system. Proc. Natl. Acad. Sci. U.S.A. 104, 997–1002. doi: 10.1073/pnas.0609672104
Broughton, W. J., Jabbouri, S., and Perret, X. (2000). Keys to symbiotic 279 harmony. J. Bacteriol. 182, 5641–5652. doi: 10.1128/JB.182.20.5641-5652.2000
Cesco, S., Neumann, G., Tomasi, N., Pinton, R., and Weisskopf, L. (2010). Release of plant-borne flavonoids into the rhizosphere andtheir role in plant nutrition. Plant Soil 329, 1–25. doi: 10.1007/s11104-009-0266-9
Cordeiro, F. A., Tadra-Sfeir, M. Z., Huergo, L. F., Pedrosa, F. O., Monteiro, R. A., and Souza, E. M. (2013). Proteomic analysis of Herbaspirillum seropedicae cultivated in the presence of sugar cane extract. J. Proteome Res. 12, 1142–1150. doi: 10.1021/pr300746j
Erbs, G., and Newman, M. A. (2012). The role of lipopolysaccharide and peptidoglycan, two glycosylated bacterial microbe-associated molecular patterns (MAMPs), in plant innate immunity. Mol. Plant Pathol. 13, 95–104. doi: 10.1111/j.1364-3703.2011.00730.x
Erbs, G., Silipo, A., Aslam, S., De Castro, C., Liparoti, V., and Flagiello, A. (2008). Peptidoglycan and muropeptides from pathogens Agrobacterium and Xanthomonas elicit plant innate immunity: structure and activity. Chem. Biol. 15, 438–448. doi: 10.1016/j.chembiol.2008.03.017
Ferrandez, A., Hawkins, A. C., Summerfield, D. T., and Harwood, C. S. (2002). Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374–4383. doi: 10.1128/JB.184.16.4374-4383.2002
Gough, C., Galera, C., Vasse, J., Webster, G., Cocking, E. C., and Dénarié, J. (1997). Specific flavonoids promote intercellular root colonization of Arabidopsis thaliana by Azorhizobium caulinodans ORS571. Mol. Plant. Microbe Interact. 10, 560–570. doi: 10.1094/MPMI.1997.10.5.560
Guvener, Z. T., Tifrea, D. F., and Harwood, C. S. (2006). Two different Pseudomonas aeruginosa chemosensory signal transduction complexes localize to cell poles and form and remould in stationary phase. Mol. Microbiol. 61, 106–118. doi: 10.1111/j.1365-2958.2006.05218.x
Gyaneshwar, P., James, E. K., Reddy, P. M., and Ladha, J. K. (2002). Herbaspirillum colonization increases growth and nitrogen accumulation in aluminium-tolerant rice varieties. New Phytol. 154, 131–145. doi: 10.1046/j.1469-8137.2002.00371.x
Iriarte, M., Stainier, I., Mikulskis, A. V., and Cornelis, G. R. (1995). The fliA gene encoding sigma 28 in Yersinia enterocolitica. J. Bacteriol. 177, 2299–2304.
Jofré, E., Fischer, S., Príncipe, A., Castro, M., Ferrari, W., Lagares, A., et al. (2009). Mutation in a D-alanine D-alanine ligase of Azospirillum brasilense Cd results in an overproduction of exopolysaccharides and a decreased tolerance to saline stress. FEMS Microbiol. Lett. 290, 236–246. doi: 10.1111/j.1574-6968.2008.01421.x
Klassen, G., Pedrosa, F. O., Souza, E. M., Funayama, S., and Rigo, L. U. (1997). Effect of nitrogen compounds on nitrogenase activity in Herbaspirillum seropedicae strain SMR1. Can. J. Microbiol. 43, 887–891. doi: 10.1139/m97-129
Komeda, Y. (1986). Transcriptional control of flagellar genes in Escherichia coli K-12. J. Bacteriol. 168, 1315–1318.
Kutsukake, K., Minamino, T., and Yokoseki, T. (1994). Negative regulation by fliD, fliS, and fliT of the export of the flagellum-specific anti-sigma factor, FlgM, in Salmonella typhimurium. J. Bacteriol. 176, 7625–7629.
Liu, X., and Matsumura, P. (1994). The FlhD/FlhC complex, a transcriptional activator of the Escherichia coli flagellar class II operons. J. Bacteriol. 176, 7345–7351.
Livak, K. J., and Schmittgen, T. D. (2008). Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L., and Wold, B. (2008). Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 5, 621–628. doi: 10.1038/nmeth.1226
Muchova, K., Wilkinson, A. J., and Barak, I. (2011). Changes of lipid domains in Bacillus subtilis cells with disrupted cell wall peptidoglycan. FEMS Microbiol. Lett. 325, 92–98. doi: 10.1111/j.1574-6968.2011.02417.x
Ohnishi, K., Kutsukake, K., Suzuki, H., and Iino, T. (1990). Gene fliA encodes an alternative sigma factor specific for flagellar operons in Salmonella typhimurium. Mol. Gen. Genet. 221, 139–147. doi: 10.1007/BF00261713
Patin, D., Boniface, A., Kovac, A., Herve, M., and Dementin, S. (2010). Purification and biochemical characterization of Mur ligases from Staphylococcus aureus. Biochimie 92, 1793–1800. doi: 10.1016/j.biochi.2010.07.009
Pedrosa, F. O., Monteiro, R. A., Wassem, R., Cruz, L. M., Ayub, R. A., and Colauto, N. B. (2011). Genome of Herbaspirillum seropedicae strain SmR1, a specialized diazotrophic endophyte of tropical grasses. PLoS Genet. 7:e1002064. doi: 10.1371/journal.pgen.1002064
Pereira, M., Parente, J. A., Bataus, L. A., Cardos, D. D., Soares, R. B., and Soares, C. M. (2004). Chemotaxis and flagellar genes of Chromobacterium violaceum. Genet. Mol. Res. 3, 92–101.
Ramos, H. C., Rumbo, M., and Sirard, J. C. (2004). Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol. 12, 509–517. doi: 10.1016/j.tim.2004.09.002
Rasband, W. S. (1997). ImageJ. Bethesda, MD: U. S. National Institutes of Health. Available online at: http://imagej.nih.gov/ij/
Tadra-Sfeir, M. Z., Souza, E. M., Faoro, H., Müller-Santos, M., Baura, V. A., Tuleski, T. R., et al. (2011). Naringenin regulates expression of genes involved in cell wall synthesis in Herbaspirillum seropedicae. Appl. Environ. Microbiol. 77, 2180–2183. doi: 10.1128/AEM.02071-10
Tatusov, R. L., Koonin, E. V., and Lipman, D. J. (1997). A genomic perspective on protein families. Science 278, 631–637. doi: 10.1126/science.278.5338.631
Tsukada, S., Aono, T., Akiba, N., Lee, K. B., Liu, C. T., Toyazaki, H., et al. (2009). Comparative genome-wide transcriptional profiling of Azorhizobium caulinodans ORS571 grown under free-living and symbiotic conditions. Appl. Environ. Microbiol. 75, 5037–5046. doi: 10.1128/AEM.00398-09
Vargas, P., Farias, G. A., Nogales, J., Prada, H., Carvajal, V., Baron, M., et al. (2013). Plant flavonoids target Pseudomonas syringae pv. tomato DC3000 flagella and type III secretion system. Environ. Microbiol. Rep. 5, 841–850. doi: 10.1111/1758-2229.12086
Vikram, A., Jayaprakasha, G. K., Jesudhasan, P. R., Pillai, S. D., and Patil, B. S. (2010). Suppression of bacterial cell-cell signaling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 109, 515–527. doi: 10.1111/j.1365-2672.2010.04677.x
Vikram, A., Jesudhasan, P. R., Jayaprakasha, G. K., Pillai, S. D., Jayaraman, A., and Patil, B. S. (2011). Citrus flavonoid represses Salmonella pathogenicity island 1 and motility in S. Typhimurium LT2. Int. J. Food Microbiol. 145, 28–36. doi: 10.1016/j.ijfoodmicro.2010.11.013
Webster, G., Jain, V., Davey, M. R., Gough, C., Vasse, J., Dénarié, J., et al. (1998). The flavonoid naringenin stimulates the intercellular colonization of wheat roots by Azorhizobium caulinodans. Plant Cell Environ. 21, 373–383. doi: 10.1046/j.1365-3040.1998.00278.x
Keywords: H. seropedicae, naringenin, RNAseq, plant-bacteria interaction, transcription regulation
Citation: Tadra-Sfeir MZ, Faoro H, Camilios-Neto D, Brusamarello-Santos L, Balsanelli E, Weiss V, Baura VA, Wassem R, Cruz LM, De Oliveira Pedrosa F, Souza EM and Monteiro RA (2015) Genome wide transcriptional profiling of Herbaspirillum seropedicae SmR1 grown in the presence of naringenin. Front. Microbiol. 6:491. doi: 10.3389/fmicb.2015.00491
Received: 29 January 2015; Accepted: 04 May 2015;
Published: 21 May 2015.
Edited by:
Carolin Frank, University of California, Merced, USAReviewed by:
David Baltrus, University of Arizona, USAYumiko Sakuragi, University of Copenhagen, Denmark
Copyright © 2015 Tadra-Sfeir, Faoro, Camilios-Neto, Brusamarello-Santos, Balsanelli, Weiss, Baura, Wassem, Cruz, De Oliveira Pedrosa, Souza and Monteiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rose A. Monteiro, Department of Biochemistry and Molecular Biology, Centro Politecnico - Setor de Ciências Biológicas, Universidade Federal do Paraná, C. Postal 19046, CEP81531980, Curitiba, PR, Brazil,cm9zZWFkZWxlQGdtYWlsLmNvbQ==