


95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 04 June 2013
Sec. Microbiological Chemistry and Geomicrobiology
Volume 4 - 2013 | https://doi.org/10.3389/fmicb.2013.00140
This article is part of the Research Topic Microbial enzymes that oxidize hydrocarbons View all 11 articles
Alkanes comprise a substantial fraction of crude oil and refined fuels. As such, they are prevalent within deep subsurface fossil fuel deposits and in shallow subsurface environments such as aquifers that are contaminated with hydrocarbons. These environments are typically anaerobic, and host diverse microbial communities that can potentially use alkanes as substrates. Anaerobic alkane biodegradation has been reported to occur under nitrate-reducing, sulfate-reducing, and methanogenic conditions. Elucidating the pathways of anaerobic alkane metabolism has been of interest in order to understand how microbes can be used to remediate contaminated sites. Alkane activation primarily occurs by addition to fumarate, yielding alkylsuccinates, unique anaerobic metabolites that can be used to indicate in situ anaerobic alkane metabolism. These metabolites have been detected in hydrocarbon-contaminated shallow aquifers, offering strong evidence for intrinsic anaerobic bioremediation. Recently, studies have also revealed that alkylsuccinates are present in oil and coal seam production waters, indicating that anaerobic microbial communities can utilize alkanes in these deeper subsurface environments. In many crude oil reservoirs, the in situ anaerobic metabolism of hydrocarbons such as alkanes may be contributing to modern-day detrimental effects such as oilfield souring, or may lead to more beneficial technologies such as enhanced energy recovery from mature oilfields. In this review, we briefly describe the key metabolic pathways for anaerobic alkane (including n-alkanes, isoalkanes, and cyclic alkanes) metabolism and highlight several field reports wherein alkylsuccinates have provided evidence for anaerobic in situ alkane metabolism in shallow and deep subsurface environments.
Alkanes are carbon and hydrogen containing molecules that are abundant across the globe, found primarily in fossil energy deposits. They are most abundant in crude oil reservoirs, but can also be found associated with shale and coal seams (Formolo et al., 2008; Strąpoć et al., 2011). Straight chain alkanes (n-alkanes), branched alkanes (also known as isoalkanes), and cyclic alkanes form the saturate fraction of crude oils, which can comprise ~20% (by wt.) in a heavy oil and up to ~50% (by wt.) in a typical light oil (Hunt, 1996). The smallest alkane is methane, a C1 compound, while the largest alkane extends beyond C100 in paraffinic (waxy) oils (Hsieh et al., 2000). Under ambient conditions, alkanes ranging from C1–C4 are gases, those ranging from C5–C16 are liquids, and alkanes >C17 are solids (Hunt, 1996). In addition to their prevalence in petroleum mixtures, alkanes can also be found in some plants and animals, where they play a protective role or function as signaling molecules among species (Thom et al., 2007; Kunst and Samuels, 2009).
Alkanes are characterized by non-polar sigma bonds, a property that renders them relatively unreactive to most chemical transformations unless high temperatures, UV light, or specialized catalysts are applied (Carey, 2007). In contrast, these compounds are readily susceptible to microbiological transformation under ambient conditions. Understanding the biodegradation of all classes of hydrocarbons including alkanes is especially important, as the transport, storage, and usage of fossil fuel as our primary global energy source has resulted in widespread contamination of hydrocarbons into natural surface or near surface ecosystems such as groundwater aquifers. Thus, the microbial degradation of hydrocarbons represents an important fate process to mitigate such contamination. In deeper subsurface environments such as oil reservoirs, it has also been suggested that anaerobic microbial activity contributed to oil biodegradation over geological time, giving rise to heavy oil reservoirs (Huang and Larter, 2005; Head et al., 2010). Thus, understanding the processes underlying hydrocarbon biodegradation in reservoirs is important because these processes can potentially lead to detrimental effects such as heavy oil generation or reservoir souring (Voordouw et al., 2009). However, such activity can also lead to the development of beneficial biotechnologies such as microbially-enhanced energy recovery by purposefully stimulating hydrocarbon biodegradation to yield methane as an alternate energy source in mature oil reservoirs or other fossil energy environments like coal or shale deposits (Formolo et al., 2008; Gieg et al., 2008; Gray et al., 2010; Strąpoć et al., 2011). Therefore, understanding the biodegradation of hydrocarbons such as alkanes under a variety of electron-accepting conditions is important for both bioremediation and microbial-based energy recovery applications.
Alkanes have long been known to be biodegradable under aerobic conditions by microorganisms. In addition to utilizing oxygen as an electron acceptor, aerobes use oxygen as a co-reactant to activate the stable carbon-carbon bonds of alkanes using monooxygenases (Rojo, 2009). The monooxygenases incorporate oxygen into the alkane molecule typically at the terminal carbon, forming an alcohol. This metabolite is then transformed to an aldehyde, which is subsequently converted to a carboxylic acid. The latter product is then readily metabolized via β-oxidation reactions to products that can enter central metabolic pathways (Rojo, 2009). The reader is referred to other papers in this volume for more details on the aerobic microbial oxidation of alkanes.
The anaerobic oxidation of alkanes has also been documented in a few early reports (ZoBell, 1946; Muller, 1957; Jack et al., 1985) but was largely discounted due to the overwhelming evidence for and widespread prevalence of aerobic alkane biodegradation processes that can readily occur in surface soils or in aerated environments. However, mounting evidence over the last two decades has shown that petroleum mixtures spilled into natural environments such as groundwater aquifers results in a rapid depletion of oxygen, creating anaerobic conditions. Thus, much attention has now also been focused on examining the potential for anaerobic hydrocarbon metabolism primarily in the interest of applying bioremediation for site cleanup.
Research conducted in the 1990's examining the anaerobic biodegradation of alkylbenzenes such as toluene and xylene using samples collected from hydrocarbon-contaminated sites revealed that anaerobes activate hydrocarbons by a novel carbon-carbon addition mechanism known as “fumarate addition.” In this mechanism, the methyl group of an alkylbenzene such as toluene is added to fumarate (a C4-dicarboxylic acid present in central microbial metabolic pathways) forming benzylsuccinic acid (Biegert et al., 1996; Beller and Spormann, 1997). The enzyme carrying out this reaction, benzylsuccinate synthase, is known to be part of the radical SAM superfamily of enzymes (Frey, 2001; Widdel and Grundmann, 2010). Following this discovery, many more details regarding the mechanism, enzymes, and genes involved in this activation step (and further downstream metabolic reactions) have been elucidated for many aromatic hydrocarbons, including methylated mono- and polycyclic aromatic hydrocarbons [e.g., reviewed by Foght (2008)]. Subsequently, investigators examining anaerobic n-alkane metabolism found that this class of hydrocarbons could also be activated by carbon-carbon bond addition to fumarate, forming alkylsuccinates (Kropp et al., 2000; Rabus et al., 2001). Fumarate addition occurs primarily at the C2 subterminal carbon position of a given n-alkane (Figure 1A). Alkylsuccinates are then further transformed by a postulated carbon skeleton rearrangement followed by decarboxylation yielding branched fatty acids that can readily enter β-oxidation and other central metabolic pathways (Wilkes et al., 2002; Cravo-Laureau et al., 2005; Davidova et al., 2005; Callaghan et al., 2006; Widdel and Grundmann, 2010) (Figure 1A). Recently, the genes responsible for the initial “alkylsuccinate synthase” reaction were identified, known as the masD or assA genes (Callaghan et al., 2008; Grundmann et al., 2008). Laboratory-based studies examining the anaerobic degradation of pure n-alkanes have demonstrated that alkanes ranging from C3 to C16 are susceptible to fumarate addition, as the corresponding alkylsuccinates have been identified in culture fluids (Table 1). A recent laboratory study has also shown that n-alkanes up to C26 in whole crude oil can be degraded via fumarate addition under sulfate-reducing conditions (Aitken et al., 2013). Further, the assA gene has been detected in anaerobic cultures incubated with pure n-alkanes as short as C3 and as long as C28 (Callaghan et al., 2010). It has been suggested that fumarate addition may even be possible for anaerobic methane oxidation (Thauer and Shima, 2008; Beasley and Nanny, 2012). An alternate mechanism to fumarate addition has also been suggested involving carboxylation at the C3 position of an n-alkane, followed by ethyl group removal by an unknown mechanism (So et al., 2003). Studies with a nitrate-reducing enrichment culture degrading n-hexadecane suggested a similar mechanism (Callaghan et al., 2009). Even though recent research has shown that alkanes are biodegraded under methanogenic conditions in laboratory incubations (e.g., Zengler et al., 1999; Siddique et al., 2006; Gieg et al., 2008; Jones et al., 2008; Siddique et al., 2011; Zhou et al., 2012; Aitken et al., 2013; Cheng et al., 2013), alkylsuccinates have not been detected as metabolites during the metabolism of n-alkanes, prompting the hypothesis that a different pathway may be occurring under these highly reduced conditions (Mbadinga et al., 2011; Aitken et al., 2013).
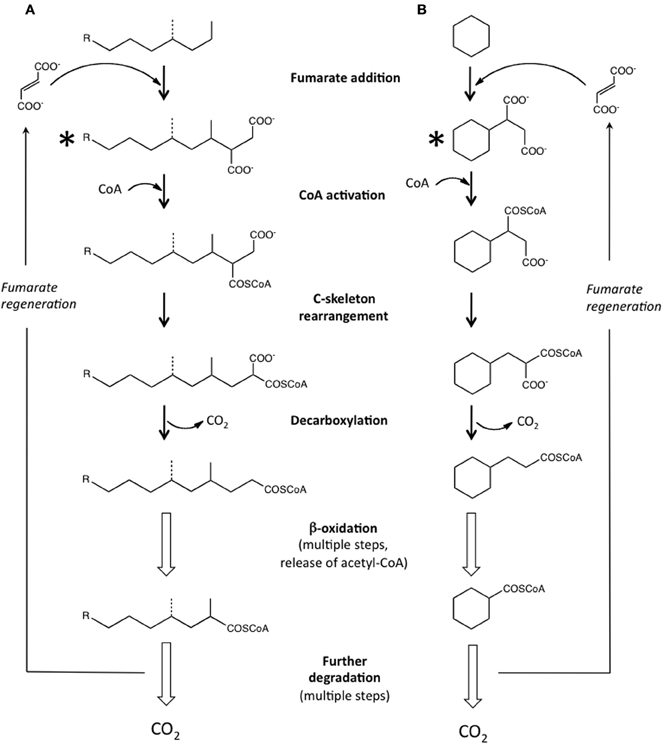
Figure 1. The anaerobic biodegradation of alkanes via fumarate addition and subsequent reactions. (A) Pathway for the biodegradation of n-alkanes and potentially for isoalkanes (dotted lines) based on literature reports [e.g., as overviewed by Widdel and Grundmann (2010)]. (B) Proposed pathway for the biodegradation of cyclic alkanes using (after Rios-Hernandez et al., 2003; Musat et al., 2010). Compounds marked with asterisks indicate fumarate addition metabolites that are most diagnostic of in situ anaerobic biodegradation of alkanes.
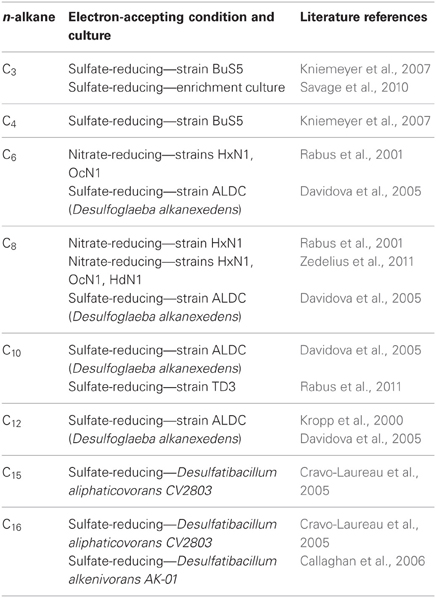
Table 1. Overview of literature studies wherein fumarate addition metabolites (alkylsuccinates) were detected in anaerobic laboratory incubations with n-alkanes added as pure substrates.
A few studies have now shown that cyclic alkanes may be also be activated by fumarate addition (Figure 1B). In studies examining the biotransformation of hydrocarbons in whole crude oil by a nitrate-reducer (strain HxN1) capable of utilizing C6–C8 n-alkanes, Wilkes et al. (2003) detected metabolites showing that cyclopentane and methylcyclopentane were biotransformed to the corresponding fumarate addition products in co-metabolic reactions with hexane as the primary carbon source. Using sediments from a gas condensate-contaminated site, Rios-Hernandez et al. (2003) demonstrated sulfate-dependent ethylcyclopentane metabolism, and identified a putative fumarate addition metabolite from this cyclic alkane. In anoxic laboratory incubations prepared from the same contaminated sediments amended with sulfate and methylcyclohexane-d14, the corresponding d-labeled fumarate addition metabolite was also tentatively identified from this cyclic alkane (Gieg et al., 2009). Unfortunately, the exact location on the alkylated cyclic alkanes to which fumarate was added (e.g., alkyl group or ring carbon) could not be determined in the studies. Musat et al. (2010) showed that the model unsubstituted cyclic alkane cyclohexane was also activated by a carbon-carbon addition to fumarate under nitrate-reducing conditions, a process that was also coupled to anaerobic ammonium oxidation. A proposed pathway for the anaerobic biodegradation of a model cyclic alkane (cyclohexane) via fumarate addition and subsequent reactions (Rios-Hernandez et al., 2003; Musat et al., 2010) is shown in Figure 1B.
Comparatively little has been reported on the anaerobic biotransformation of isoalkanes. Branched alkanes such as pristane and phytane have frequently been used as biomarkers with which to compare the extent of biodegradation of alkanes in crude oils, as these kinds of compounds are typically more recalcitrant than n-alkanes (Huang and Larter, 2005). However, there is some evidence that branched alkanes can be utilized under anaerobic conditions. Two studies carried out under anaerobic conditions showed that the branched alkane pristane was susceptible to biodegradation under nitrate-reducing or methanogenic conditions but no metabolites were reported (Bregnard et al., 1997; Grossi et al., 2000). Using oil sands tailings ponds samples as an inoculum, Abu Laban et al. (2012) recently showed that C7 and C8 isoalkanes could also be metabolized under sulfate-reducing or methanogenic conditions. In the study, metabolites consistent with the mass spectral (MS) profiles of the corresponding alkylsuccinates were detected, showing for the first time that isoalkanes are also susceptible to fumarate addition (Figure 1A).
Given the overwhelming evidence that microbes are able to biodegrade hydrocarbons, relying on such activity for the remediation of contaminated sites is an attractive and relatively inexpensive clean-up option. However, documenting that such anaerobic hydrocarbon biodegradation is occurring in fuel-contaminated sites or in fossil energy reservoirs remains a challenge. For example, multiple lines of evidence, both chemical and microbiological, are required to offer support that in situ hydrocarbon bioremediation is occurring in contaminated aquifers (NRC, 1993; Gieg and Suflita, 2005; Beller et al., 2008; Weiss and Cozzarelli, 2008; Bombach et al., 2010; Morasch et al., 2011; Jeon and Madsen, 2012). In conjunction with the elucidation of anaerobic hydrocarbon metabolic pathways, the idea of using anaerobic metabolites to indicate that in situ anaerobic hydrocarbon biodegradation is occurring emerged as a powerful tool for proving in situ bioremediation. Beller et al. (1995) first proposed that these metabolites can be effective indicators of in situ anaerobic hydrocarbon biodegradation since they are uniquely anaerobic and are specific to their hydrocarbon substrate; indeed they detected these metabolites in anoxic hydrocarbon-impacted groundwater but not in uncontaminated areas. Beller (2000) proposed a list of criteria that define a signature metabolite for use as an in situ bioremediation indicator including that the metabolite should be (1) actively produced during biodegradation, (2) relatively chemically and biologically stable so that it can be detected, (3) a true intermediate rather than a cometabolic by-product, (4) absent in the contaminant mixture, (5) absent in uncontaminated environments, and (6) be of no commercial use.
In addition to the initial reports by Beller and colleagues, many other studies have now shown that fumarate addition metabolites of alkylbenzenes (e.g., toluene, ethylbenzene, xylenes, and trimethylbenzenes) can be detected in hydrocarbon-contaminated groundwater [see reviews by Beller (2000); Gieg and Suflita (2005), and Callaghan (2012)]. With the discovery that methylnaphthalenes and other multi-ringed aromatics (including heterocycles) can also be metabolized by fumarate addition, the corresponding succinates have also been detected in field studies (Griebler et al., 2004; Bombach et al., 2010; Jobelius et al., 2011; Morasch et al., 2011), attesting to the usefulness of this method for demonstrating the in situ anaerobic biodegradation of many hydrocarbon classes. Furthermore, primer sets based on the catabolic gene(s) encoding benzylsuccinate synthase have been developed by several investigators and have been used successfully to indicate the metabolic potential for microbial communities in contaminated field samples to anaerobically biodegrade alkylbenzenes (e.g., Beller et al., 2002, 2008; Winderl et al., 2007; Callaghan et al., 2010; Oka et al., 2011; von Netzer et al., 2013). It should be noted that downstream metabolites of anaerobic hydrocarbon biodegradation can also be indicators of in situ biodegradation, especially if they are detected in contaminated but not in unrelated, uncontaminated groundwater samples (Cozzarelli et al., 1995; Gieg and Suflita, 2002). However, these compounds are typically not specific to a parent substrate (for example, benzoate, often detected in contaminated groundwaters, is a central intermediate in the anaerobic metabolism of many aromatic compounds) or may be produced via aerobic or anaerobic processes (such as toluates from xylenes) (Beller, 2000; Gieg and Suflita, 2005). In anaerobic alkane metabolism, downstream metabolites include fatty acids, which may be produced via a variety of other cellular pathways. Thus, the fumarate addition metabolites are most diagnostic of the in situ anaerobic metabolism of specific hydrocarbons including alkanes. It should be noted, however, that there are some disadvantages to using signature metabolites as sole indicators of in situ bioremediation (Morasch et al., 2011). For example, these products may be below analytical detection limits, knowledge of biodegradation pathways for various hydrocarbons is required (e.g., in order to determine which metabolites to look for), and commercially-available authentic standards for many of the fumarate addition products are not available for analytical comparisons with metabolites found in field samples. Thus, using signature fumarate addition metabolites to determine whether in situ hydrocarbon metabolism is occurring at a given site should ideally be done in conjunction with other methods of site assessment (Bombach et al., 2010; Morasch et al., 2011).
In field bioremediation studies, the biodegradation of BTEX compounds has been a focus because these compounds are highly water-soluble (150–1780 mg/L; Heath et al., 1993) thus can rapidly migrate away from the spill source. Furthermore, benzene is among the most highly regulated hydrocarbons due to its carcinogenicity (Maltoni et al., 1989). However, in many fuel mixtures that spill into aquifers (such as gasoline, diesel, gas condensate, jet fuel), the saturate fraction (alkanes, isoalkanes, and cyclic alkanes) is the most abundant (e.g., up to 80% by wt., Heath et al., 1993). Since many alkanes have appreciable water solubilites (in the tens of mg/L), some alkanes can also migrate in groundwater away from the spill source. Many alkanes have known toxicity (Ritchie et al., 2001), thus their presence in contaminated aquifers is also of concern and their levels are often regulated (Nascarella et al., 2002).
To date, though, relatively few studies have reported on the detection of signature anaerobic alkane metabolites indicative of in situ anaerobic alkane in hydrocarbon-contaminated near surface environments. Gieg and Suflita (2002) documented that alkylsuccinates can be detected in hydrocarbon contaminated aquifers alongside benzylsuccinates, providing evidence that alkanes are susceptible to in situ anaerobic biodegradation and that alkylsuccinates can also be used as indicator metabolites. In 5 of the 6 sites examined for these signature compounds, alkylsuccinates were detected in contaminated groundwater samples, ranging from C3 to C11 alkylsuccinates depending on the site interrogated (Gieg and Suflita, 2002; Table 2). Several of the detected metabolites had MS fragments with two mass units less than those predicted to arise from straight chain (or branched) alkane metabolism, suggesting the formation of fumarate addition products from cyclic alkanes. Using sediments from a gas condensate-contaminated site, Rios-Hernandez et al. (2003) demonstrated that ethylcyclopentane could be biodegraded under sulfate-reducing conditions in laboratory incubations, and identified a putative fumarate addition metabolite that formed during the biodegradation of this compound. The gas chromatographic (GC) retention time and MS profile of this incubation-derived metabolite matched with that of a GC peak detected in several of the samples from the contaminated site, supporting the notion that cyclic alkanes can be anaerobically biotransformed in situ. Parisi et al. (2009) also detected alkylsuccinates from C5 to C9 n- or cyclic alkanes in groundwater samples collected from an aquifer contaminated with a variety of fuel mixtures while Gieg et al. (2009) detected C5–C9 alkylsuccinates in several anoxic groundwater samples prior to conducting push-pull tests in a jet fuel contaminated aquifer, further attesting to the in situ anaerobic biodegradation of alkanes (Table 2). In a gas condensate-contaminated site wherein alkylsuccinates were detected in groundwater samples (Gieg and Suflita, 2002; Rios-Hernandez et al., 2003), Callaghan et al. (2010) detected the presence of the alkylsuccinate synthase gene (assA gene), augmenting the metabolite findings. Primer sets for this gene were also used to probe samples collected from a handful of other hydrocarbon-contaminated aquifers in the US and in Germany, and indeed this gene was detected (Table 2; Callaghan et al., 2010; von Netzer et al., 2013). Recently, assA gene sequences were also detected in deep sea core samples collected from areas near the Deepwater Horizon oil spill, indicating that the extant organisms in deep-sea sediments have the potential to biodegrade alkanes and can potential contribute to bioremediation of such oil spills (Kimes et al., 2013). Although the alkylsuccinates themselves were not detected in these latter field sites, the detection of the gene catalyzing their formation in contaminated sites reveals a widespread potential for in situ anaerobic alkane biodegradation and usefulness of probing for the requisite metabolic genes. However, there are currently some limitations in the use of the assA gene. For example, this gene appears to be highly diverse, as a single primer set has not yet been designed that can be used to detect/amplify this gene (Callaghan et al., 2010; Aitken et al., 2013; von Netzer et al., 2013). Further, the assA genes show high similarity with the bssA genes, thus primer pairs designed to detect assA may also amplify the bssA genotypes (Callaghan et al., 2010). However, as stated above, such limitations can be overcome by coupling gene analysis with metabolite analysis and other indicators of biodegradation at field sites to provide strong evidence for intrinsic bioremediation of hydrocarbons like alkanes at contaminated sites. Table 2 summarizes the reports to date wherein alkylsuccinates (and/or alkylsuccinate synthase genes) have been detected in hydrocarbon-contaminated sites, providing evidence in support of in situ anaerobic alkane biodegradation.
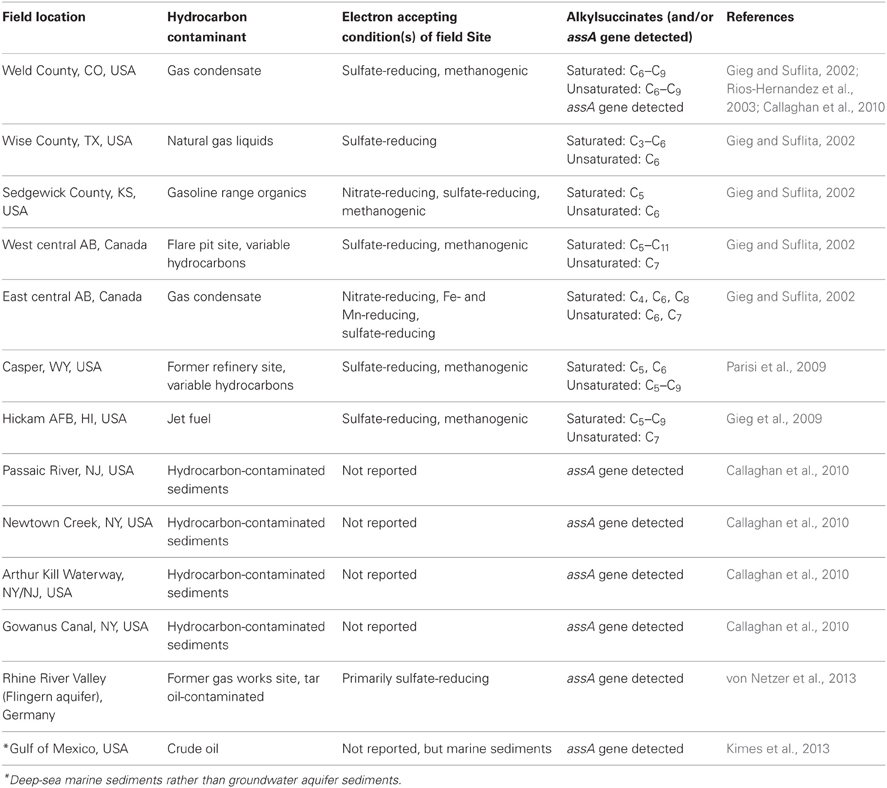
Table 2. Summary of studies reporting the detection of alkylsuccinates [and/or alkylsuccinate synthase (assA) gene] in hydrocarbon- contaminated aquifer samples*.
Geochemical data suggesting the biodegradation of crude oil in reservoirs over geological time generating heavy oil has prompted an examination of these oils and fluids of oil reservoirs in general for evidence of anaerobic hydrocarbon metabolites. Upon examining 77 crude oils from across the globe, Aitken et al. (2004) detected known anaerobic metabolites of naphthalenes in 52 samples (including 2-naphthoic acid; 5,6,7,8-tetrahydro-2-naphthoic acid; decahydro-2-naphthoic acid), offering the first metabolic evidence that anaerobic in situ biodegradation of crude oil can occur in reservoirs. Although alkylsuccinates were also sought, none were detected in the assays. In a later study, Duncan et al. (2009) examined the metabolite profiles of several produced water samples (e.g., water removed along with oil from production wells during oil recovery) collected from high temperature Alaska North Slope (ANS) oilfields. The authors were able to detect some low molecular weight alkylsuccinates derived from C1–C4 alkanes in the produced water samples (Table 3). Downstream, branched alkanoic acids related to these alkylsuccinates were also detected (Duncan et al., 2009). Operationally, these oilfields were pressurized with natural gas to aid in oil recovery, providing the oilfield microbial community with a continuous source of low molecular weight alkanes that were likely ultimately biotransformed to the corresponding alkylsuccinates (Duncan et al., 2009). In a subsequent study, Gieg et al. (2010) examined samples collected from two different high temperature ANS oilfields, and also detected several fumarate addition metabolites including C1–C3 alkylsuccinates (Table 3). This latter study further showed that the methanogenic microbial communities present in crude oil reservoirs are able to subsist on hydrocarbons supporting the notion that oil biodegradation within fossil energy reservoirs can occur under anaerobic conditions.
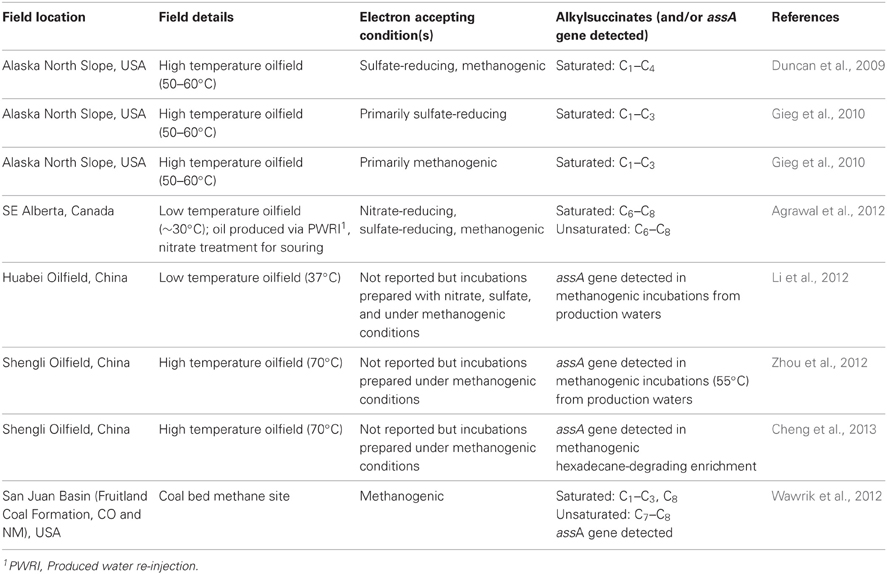
Table 3. Summary of studies reporting the detection of alkylsuccinates [and/or alkylsuccinate synthase (assA) gene] in fossil energy reservoir samples.
A low temperature (~30°C) heavy oil field in Alberta, Canada (Medicine Hat Glauconitic C field) has been under study for over 5 years to test the effects of nitrate treatment on souring caused by sulfate-reducing bacteria (Lambo et al., 2008; Voordouw et al., 2009; Callbeck et al., 2011; Agrawal et al., 2012). Oil analysis conducted on various samples from this field showed that toluene served as a major electron donor driving the microbial activities within the subsurface reservoir environment (Lambo et al., 2008). This result was subsequently confirmed in laboratory studies conducted under nitrate-reducing and sulfate-reducing conditions using produced water and heavy crude oil from the field. However, the laboratory incubation tests also showed that other alkylbenzenes and alkanes present in the oil served as substrates for the oilfield microbial community (Agrawal et al., 2012). Notably, several alkanes were depleted in sulfate-reducing enrichments, and numerous metabolites indicative of fumarate addition to both aromatics and alkanes were detected, including methyl- and dimethylbenzylsuccinates and alkylsuccinates derived from C6–C9 alkanes, confirming the anaerobic biodegradation of hydrocarbons in the oil. These laboratory-based studies prompted an investigation of produced water samples from the heavy oil field for the presence of alkylsuccinates, and indeed several of these signature anaerobic alkane metabolites were detected, ranging from C5 to C8 alkylsuccinates (Table 3). These data suggested that the anaerobic microbial communities present within this low temperature reservoir can utilize alkanes as a carbon and energy source, potentially leading to souring when sulfate is present. Although alkylsuccinates were not specifically identified in other oilfields (Li et al., 2012; Zhou et al., 2012; Cheng et al., 2013), recently identified alkylsuccinate synthase genes in incubations prepared from oilfield fluids, showing that microbes residing in the sampled oilfield harbor the potential for in situ anaerobic alkane metabolism, again attesting to the usefulness of catabolic gene probes for determining in situ alkane biodegradation potential (Table 2).
Wawrik et al. (2012) recently conducted a study examining the biological conversion of coal to methane in a major coal-bed methane producing region in the USA (San Juan Basin) using a series of enrichments, functional gene assays, hydrocarbon metabolite analyses, and microbial community profiling of produced water samples. Coal is a complex, organic carbon-rich mixture derived from ancient wetland plants that were buried and transformed by heat and pressure over geological time (Strąpoć et al., 2011). While methane produced from coal beds is typically thermogenic, increasing evidence (such as the isotopic signature of methane) is showing that biologically produced methane is also occurring in many coal-bed regions (Strąpoć et al., 2011). Although not as clearly defined as crude oil, coal can contain many substrates such as alkanes (Orem et al., 2007; Strąpoć et al., 2011) that may be used by microbial communities living in coal beds to produce methane in real time, presumably via pathways similar to those shown in Figure 1. Using a functional gene array (GeoChip 3.1), Wawrik et al. (2012) detected the assA gene in several produced water samples collected from the San Juan Basin, showing that the indigenous microbial community has the metabolic potential for in situ anaerobic alkane biodegradation. In addition, they also detected C1–C3, C7, and C8 alkylsuccinates in several of the produced waters (Table 3). These data collectively showed for the first time that alkanes (and other substrates) associated with the coal can be metabolized by the associated microbial community, presumably leading to biological methane production in such fossil energy resources (Wawrik et al., 2012).
Tools of analytical chemistry including gas chromatography-mass spectrometry (GC-MS) and high performance liquid chromatography coupled with tandem MS (LC-MS-MS) have generally been used to detect hydrocarbon metabolites in samples collected from contaminated groundwater wells or produced water wellheads of oil or coal reservoirs. Detailed descriptions of these approaches have been described in several previous publications (e.g., Beller, 2002; Gieg and Suflita, 2002, 2005; Alumbaugh et al., 2004; Jobelius et al., 2011). Briefly, for metabolite analysis by GC-MS, at least a liter of water should be collected into clean vessels (preferably glass) and acidified immediately to pH ~2 to preserve samples and protonate metabolites. Samples can then be processed in the laboratory using liquid-liquid extraction with an organic solvent such as dichloromethane, diethyl ether, or ethylacetate. Concentrated organic solvent extracts are concentrated, then derivatized by methylation or silylation prior to analysis by GC-MS (Gieg and Suflita, 2002). Depending on the method of derivatization, alkylsuccinates have diagnostic MS fragment ions that can be used to readily determine their presence in a given water sample. For example, trimethylsilyl esters of alkylsuccinates have diagnostic fragment ions of m/z 262, 217, 172, 147, and 73 (Gieg and Suflita, 2002, 2005). Metabolite concentrations can be estimated by comparison to calibration curves prepared from commercially available or readily synthesized alkylsuccinates (e.g., methylsuccinate or n-octylsuccinate). Detection limits for alkylsuccinates by GC-MS analysis are in the nanomolar range (Gieg and Suflita, 2002). Hydrocarbon metabolite analysis by LC-MS-MS analysis has the advantage of requiring little to no sample preparation prior to analysis thus is far less labor-intensive (Beller, 2002). Further, the technique requires only small amounts of sample (~1 mL). Parent and/or daughter ions of various metabolites are typically monitored (Beller, 2002; Alumbaugh et al., 2004). Beller (2002) found the detection limits for benzylsuccinates to be ~0.3 μg/L using LC tandem MS, while Alumbaugh et al. (2004) found that sample pre-treatment using solid-phase extraction lowered the LC-MS-MS detection level of benzylsuccinates by at least an order of magnitude (0.006–0.029 μg/L). Although high sensitivity LC-MS techniques (e.g., LC/MS-ESI-Q-TOF) have been used to successfully detect benzylsuccinates in environmental samples (Jobelius et al., 2011; Kimes et al., 2013), the detection of alkylsuccinates remains elusive, possibly because these metabolites are present at levels below detection limits (Kimes et al., 2013). Other analytical techniques such as 2-dimensional GC analysis (GC × GC) coupled with MS, currently used for separating and quantifying complex mixtures of crude oil components and related compounds (such as naphthenic acids) (e.g., Rowland et al., 2011; Reddy et al., 2012), may find future application for hydrocarbon metabolite analysis.
Research on the topic of anaerobic alkane metabolism has increased in the last dozen years, revealing that alkanes can be activated by fumarate addition reactions as well as by other proposed mechanisms (yet to be understood). Of the saturate fraction of fuels, the anaerobic metabolism of n-alkanes has been the most frequently studied while less is known about how isoalkanes and cyclic alkanes are metabolized anaerobically, representing a knowledge gap that can be addressed by future research. As knowledge is gained regarding the key metabolic pathways for anaerobic alkane biodegradation, the identified metabolites can readily be sought in the natural environment to indicate biodegradation. Further, as newer fuel mixtures such as biofuels become more commonly used, metabolic insights gained from the study of alkanes and other hydrocarbons may help predict the kinds of metabolites that can serve as diagnostic indicators for the biotreatment of such alternative fuels. To date, a metabolite profiling approach using robust tools of analytical chemistry (GC-MS, LC-MS-MS) has been widely been used to garner evidence for the in situ anaerobic metabolism of aromatic hydrocarbons in contaminated aquifers, but has been used to a lesser extent for determining for in situ alkane biodegradation. Since alkanes (including n-alkanes, isoalkanes, and cyclic alkanes) comprise substantial components of many fuel mixtures that are spilled into aquifers, looking for their metabolic by-products should be included in more future site assessments to determine the prospects for in situ alkane bioremediation; details on how to look for such metabolites in environmental samples have been described (e.g., Gieg and Suflita, 2002; Duncan et al., 2009). Parallel assessments of the catabolic gene (e.g., assA gene) in subsurface microbial populations along with metabolite profiling offers very strong evidence for the in situ anaerobic biodegradation of alkanes (Callaghan et al., 2010; Wawrik et al., 2012), an approach that should also be part of future site assessments. Although many hydrocarbon biodegradation studies have been carried out in the context of petroleum contaminated site bioremediation, understanding anaerobic hydrocarbon biodegradation in fossil energy reservoirs has enormous economic implications related to detrimental effects (e.g., heavy oil generation and souring) and beneficial technologies (e.g., microbially enhanced energy recovery). Thus, more extensive research into hydrocarbon-related microbial metabolic processes including anaerobic alkane biodegradation in fossil energy resources such as crude oil reservoirs, and coal and shale deposits is warranted.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The preparation of this review was supported by funding from the Natural Sciences and Engineering Research Council of Canada (NSERC) via a Discovery Grant awarded to Lisa M. Gieg. Akhil Agrawal was supported through NSERC Industrial Research Chair funding awarded to Dr. Gerrit Voordouw (University of Calgary) that was in turn supported by the partner companies Aramco Services, Baker Hughes Incorporated, British Petroleum, Intertek/CML, the Computer Modelling Group Limited, ConocoPhillips Company, YPF SA, Shell Canada Limited, Suncor Energy Developments Inc., and Yara International ASA, as well as by Alberta Innovates-Energy and Environment Solutions.
Abu Laban, N., Semple, K., Young, R., Dao, A., and Foght, J. M. (2012). “Biodegradation of C7 and C8 iso-alkanes under methanogenic and sulfate-reducing conditions,” in Abstracts of the 14th International Symposium on Microbial Ecology (Copenhagen, DK).
Agrawal, A., Park, H. S., Nathoo, S., Gieg, L. M., Jack, T. R., Miner, K., et al. (2012). Toluene depletion in produced oil contributes to souring control in a fied subjected to nitrate injection. Environ. Sci. Technol. 46, 1285–1292. doi: 10.1021/es203748b
Aitken, C. M., Jones, D. M., and Larter, S. R. (2004). Anaerobic hydrocarbon biodegradation in deep subsurface oil reservoirs. Nature 341, 291–294. doi: 10.1038/nature02922
Aitken, C. M., Jones, D. M., Maguire, M. J., Gray, N. D., Sherry, A., Bowler, B. F. J., et al. (2013). Evidence that crude oil alkane activation proceeds by different mechanisms under sulfate-reducing and methanogenic conditions. Geochim. Cosmochim. Acta 109, 162–174. doi: 10.1016/j.gca.2013.01.031
Alumbaugh, R. E., Gieg, L. M., and Field, J. A. (2004). Determination of alkylbenzene metabolites in groundwater by solid-phase extraction and liquid chromatography-tandem mass spectrometry. J. Chromatog. A 1042, 89–97. doi: 10.1016/j.chroma.2004.05.031
Beasley, K. K., and Nanny, M. A. (2012). Potential energy surface for anaerobic oxidation of methane via fumarate addition. Environ. Sci. Technol. 46, 8244–8252. doi: 10.1021/es3009503
Beller, H. R. (2000). Metabolic indicators for detecting in situ anaerobic alkylbenzene degradation. Biodegradation 11, 125–139. doi: 10.1023/A:1011109800916
Beller, H. R. (2002). Analysis of benzylsuccinates in groundwater by liquid chromatography/ tandem mass spectrometry and its use for monitoring in situ BTEX biodegradation. Environ. Sci. Technol. 36, 2724–2728. doi: 10.1021/es025527l
Beller, H. R., Ding, W.-H., and Reinhard, M. (1995). Byproducts of anaerobic alkylbenzene metabolism useful as indicators of in situ bioremediation. Environ. Sci. Technol. 29, 2864–2870. doi: 10.1021/es00011a024
Beller, H. R., and Spormann, A. (1997). Anaerobic activation of toluene and o-xylene by addition to fumarate in denitrifying strain T. J. Bacteriol. 179, 670–676.
Beller, H. R., Kane, S. R., Legler, T. C., and Alvarez, P. J. J. (2002). A real-time polymerase chain reaction method for monitoring anaerobic, hydrocarbon-degrading bacteria based on a catabolic gene. Environ. Sci. Technol. 36, 3977–3984. doi: 10.1021/es025556w
Beller, H. R., Kane, S. R., Legler, T. C., McKelvie, J. R., Sherwood-Lollar, B., Pearson, F., et al. (2008). Comparative assessments of benzene, toluene, and xylene natural attenuation by quantitative polymerase chain reaction analysis of a catabolic gene, signature metabolites, and compound-specific isotope analysis. Environ. Sci. Technol. 42, 6065–6072. doi: 10.1021/es8009666
Biegert, T., Fuchs, G., and Heider, J. (1996). Evidence that anaerobic oxidation of toluene in the denitrifying bacterium Thauera aromatica is initiated by formation of benzylsuccinate from toluene and fumarate. Eur. J. Biochem. 238, 661–668. doi: 10.1111/j.1432-1033.1996.0661w.x
Bombach, P., Richnow, H. H., Kastner, M., and Rischer, A. (2010). Current approaches for the assessment of in situ biodegradation. Appl. Environ. Microbiol. 86, 839–852. doi: 10.1007/s00253-010-2461-2
Bregnard, T. P., Haner, A., Hohener, P., and Zeyer, J. (1997). Anaerobic degradation of pristane in nitrate-reducing microcosms and enrichment cultures. Appl. Environ. Microbiol. 63, 2077–2081.
Callaghan, A. V. (2012). Metabolomic investigations of anaerobic hydrocarbon-impacted environments. Curr. Opin. Biotechnol. doi: 10.1016/j.copbio.2012.08.012. [Epub ahead of print].
Callaghan, A. V., Davidova, I. A., Savage-Ashlock, K., Parisi, V. A., Gieg, L. M., Suflita, J. M., et al. (2010). Diversity of benzyl- and alkylsuccinate synthase genes in hydrocarbon-impacted environments and enrichment cultures. Environ. Sci. Technol. 44, 7287–7294. doi: 10.1021/es1002023
Callaghan, A. V., Gieg, L. M., Kropp, K. G., Suflita, J. M., and Young, L. Y. (2006). Comparison of mechanisms of alkane metabolism under sulfate-reducing conditions among two bacterial isolates and a bacterial consortium. Appl. Environ. Microbiol. 72, 4274–4282. doi: 10.1128/AEM.02896-05
Callaghan, A. V., Tierney, M., Phelps, C. D., and Young, L. Y. (2009). Anaerobic biodegradation of n-hexadecane by a nitrate-reducing consortium. Appl. Environ. Microbiol. 75, 1339–1344. doi: 10.1128/AEM.02491-08
Callaghan, A. V., Wawrik, B., Ní Chadhain, S. M., Young, L. Y., and Zylstra, G. J. (2008). Anaerobic alkane-degrading strain AK-01 contains two alkylsuccinate synthase genes. Biochem. Biophys. Res. Commun. 366, 142–148. doi: 10.1016/j.bbrc.2007.11.094
Callbeck, C. M., Dong, X., Chatterjee, I., Agrawal, A., Caffrey, S. M., Sensen, C., et al. (2011). Microbial community succession in a bioreactor modeling a souring low temperature oil reservoir subjected to nitrate injection. Appl. Microbiol. Biotech. 91, 799–810. doi: 10.1007/s00253-011-3287-2
Cheng, L., Rui, J., Li, Q., Zhang, H., and Lu, Y. (2013). Enrichment and dynamics of novel syntrophs in a methanogenic hexadecane-degrading culture from a Chinese oilfield. FEMS Microbiol. Ecol. 83, 757–766. doi: 10.1111/1574-6941.12031
Cozzarelli, I. M., Herman, J. S., and Baedecker, M. J. (1995). Fate of microbial metabolites of hydrocarbons in a coastal plain aquifer: the role of electron acceptors. Environ. Sci. Technol. 29, 458–469. doi: 10.1021/es00002a023
Cravo-Laureau, C., Grossi, V., Raphel, D., Matheron, R., and Hirschler-Rea, A. (2005). Anaerobic n-alkane metabolism by a sulfate-reducing bacterium, Desulfatibacillum aliphaticovorans strain CV2803T. Appl. Environ. Microbiol. 71, 3458–3467. doi: 10.1128/AEM.71.7.3458-3467.2005
Davidova, I. A., Gieg, L. M., Nanny, M., Kropp, K. G., and Suflita, J. M. (2005). Stable isotopic studies of n-alkane metabolism by a sulfate-reducing bacterial enrichment culture. Appl. Environ. Microbiol. 71, 8174–8182. doi: 10.1128/AEM.71.12.8174-8182.2005
Duncan, K. E., Gieg, L. M., Parisi, V. A., Tanner, R. S., Tringe, S. G., Bristow, J., et al. (2009). Biocorrosive thermophilic microbial communities in Alaskan North Slope oil facilities. Environ. Sci. Technol. 43, 7977–7984. doi: 10.1021/es9013932
Foght, J. (2008). Anaerobic biodegradation of aromatic hydrocarbons: pathways and prospects. J. Mol. Microbiol. Biotechnol. 15, 93–120. doi: 10.1159/000121324
Formolo, M. J., Salacup, J. M., Petsch, S. T., Martini, A. M., and Nusslein, K. (2008). A new model linking atmospheric methane sources to Pleistocene glaciation via methanogenesis in sedimentary basins. Geology 36, 139–142.
Frey, P. A. (2001). Radical mechanisms of enzymatic catalysis. Ann. Rev. Biochem. 70, 121–148. doi: 10.1146/annurev.biochem.70.1.121
Gieg, L. M., Alumbaugh, R. E., Field, J. A., Jones, J., Istok, J. D., and Suflita, J. M. (2009). Assessing in situ rates of anaerobic hydrocarbon bioremediation. Microb. Biotechnol. 2, 222–233. doi: 10.1111/j.1751-7915.2008.00081.x
Gieg, L. M., Davidova, I. A., Duncan, K. E., and Suflita, J. M. (2010). Methanogenesis, sulfate reduction and crude oil biodegradation in hot Alaskan oilfields. Environ. Microbiol. 12, 3074–3086. doi: 10.1111/j.1462-2920.2010.02282.x
Gieg, L. M., Duncan, K. E., and Suflita, J. M. (2008). Bioenergy production via microbial conversion of residual oil to natural gas. Appl. Environ. Microbiol. 74, 3022–3029. doi: 10.1128/AEM.00119-08
Gieg, L. M., and Suflita, J. M. (2002). Detection of anaerobic metabolites of saturated and aromatic hydrocarbons in petroleum-contaminated aquifers. Environ. Sci. Technol. 36, 3755–3762.
Gieg, L. M., and Suflita, J. M. (2005). “Metabolic indicators of anaerobic hydrocarbon biodegradation in petroleum-laden environments,” in Petroleum Microbiology. eds B. Ollivier, and M. Magot (Washington DC: ASM Press), 337–356.
Gray, N. D., Sherry, A., Hubert, C., Dolfing, J., and Head, I. M. (2010). Methanogenic degradation of petroleum hydrocarbons in subsurface environments: remediation, heavy oil formation, and energy recovery. Adv. Appl. Microbiol. 72, 137–161. doi: 10.1016/S0065-2164(10)72005-0
Griebler, C., Safinowski, M., Vieth, A., Richnow, H. H., and Meckenstock, R. U. (2004). Combined stable carbon isotope analysis and specific metabolites determination for assessing is situ degradation of aromatic hydrocarbons in a tar oil-contaminated aquifer. Environ. Sci. Technol. 38, 617–631.
Grossi, V., Raphel, D., Hirschler-Rea, A., Gilewicz, M., Mouzdahir, A., Bertrand, J.-C., et al. (2000). Anaerobic biodegradation of pristane by a marine sedimentary bacterial and/or archaeal community. Org. Geochem. 31, 769–772. doi: 10.1016/S0146-6380(00)00060-7
Grundmann, O., Behrends, A., Rabus, R., Amann, J., Halder, T., Heider, J., et al. (2008). Genes encoding the candidate enzyme encoding for anaerobic activation of n-alkanes in the denitrifying bacterium, strain HxN1. Environ. Microbiol. 10, 376–385. doi: 10.1111/j.1462-2920.2007.01458.x
Head, I. M., Larter, S. R., Gray, N. D., Sherry, A., Adams, J. J., Aitken, C. M., et al. (2010). “Hydrocarbon biodegradation in petroleum reservoirs,” in Handbook of Hydrocarbon and Lipid Microbiology, ed T. M. Timmis (Berlin: Springer-Verlag), 3098–3109.
Heath, J. S., Koblis, K., and Sager, S. L. (1993). Review of chemical, physical, and toxicologic properties of components of total petroleum hydrocarbons. J. Soil Contam. 2, 1–25.
Hsieh, M., Philp, R. P., and del Rio, J. C. (2000). Characterization of high molecular weight biomarkers in crude oils. Org. Geochem. 31, 1581–1588.
Huang, H., and Larter, S. R. (2005). “Biodegradation of petroleum in subsurface geological reservoirs,” in Petroleum Microbiology, eds B. Ollivier and M. Magot (Washington, DC: ASM Press), 91–121.
Hunt, J. M. (1996). Petroleum Geochemistry and Geology, 2nd Edn. New York, NY: W. H. Freeman and Company.
Jack, T. R., Lee, E., and Mueller, J. (1985). “Anaerobic gas production from crude oil,” in Microbes and Oil Recovery: International Bioresources Journal, Vol. 1, eds J. E. Zajic and E. C. Donaldson (El Paso, TX: Bioresources Publications), 167–180.
Jeon, C. O., and Madsen, E. L. (2012). in situ microbial metabolism of aromatic-hydrocarbon environmental pollutants. Curr. Opin. Biotechnol. doi: 10.1016/j.copbio.2012.09.001. [Epub ahead of print].
Jobelius, C., Ruth, B., Griebler, C., Meckenstock, R. U., Hollender, J., Reineke, A., et al. (2011). Metabolites indicate hot spots of biodegradation and biogeochemical gradients in a high-resolution monitoring well. Environ. Sci. Technol. 45, 474–481. doi: 10.1021/es1030867
Jones, D. M., Head, I. A., Gray, N. D., Adams, J. J., Rowan, A. K., Aitken, C. M., et al. (2008). Crude oil biodegradation via methanogenesis in subsurface petroleum reservoirs. Nature 451, 176–180. doi: 10.1038/nature06484
Kimes, N. E., Callaghan, A. V., Aktas, D. F., Smith, W. L., Sunner, J., Golding, B. T., et al. (2013). Metagenomic analysis and metabolite profiling of deep-sea sediments from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 4:50. doi: 10.3389/fmicb.2013.00050
Kniemeyer, O., Musat, F., Sievert, S. M., Knittel, K., Wilkes, H., Blumenberg, M., et al. (2007). Anaerobic oxidation of short-chain hydrocarbons by marine sulphate-reducing bacteria. Nature 449, 898–902. doi: 10.1038/nature06200
Kropp, K. G., Davidova, I. A., and Suflita, J. M. (2000). Anaerobic oxidation of n-dodecane by an addition reaction in a sulfate-reducing bacterial enrichment culture. Appl. Environ. Microbiol. 66, 5393–5398. doi: 10.1128/AEM.66.12.5393-5398.2000
Kunst, L., and Samuels, L. (2009). Plant cuticles shine: advances in wax biosynthesis and export. Curr. Opin. Plant Biol. 12, 721–727. doi: 10.1016/j.pbi.2009.09.009
Lambo, A. J., Noke, K., Larter, S. R., and Voordouw, G. (2008). Competitive, microbially-mediated reduction of nitrate with sulfide and aromatic oil components in a low-temperature, western Canadian oil reservoir. Environ. Sci. Technol. 42, 8941–8946.
Li, W., Wang, L.-Y., Duan, R.-Y., Liu, J.-F., Gu, J.-D., and Mu, B.-Z. (2012). Microbial community characteristics of petroleum reservoir production water amended with n-alkanes and incubated under nitrate-, sulfate-reducing and methanogenic conditions. Int. Biodeter. Biodeg. 69, 87–96. doi: 10.1016/j.ibiod.2012.01.005
Maltoni, C., Ciliberti, A., Cotti, G., Conti, B., and Belpoggi, F. (1989). Benzene, an experimental multipotential carcinogen: results of the long-term bioassays performed at the Bologna Institute of Oncology. Environ. Health Perspect. 82, 109–124.
Mbadinga, S. M., Wang, L.-Y., Zhou, L., Liu, J.-F., Gu, J.-D., and Mu, B.-Z. (2011). Microbial communities involved in anaerobic degradation of alkanes. Int. Biodeter. Biodeg. 65, 1–13. doi: 10.1016/j.ibiod.2010.11.009
Morasch, B., Hunkeler, D., Zopfi, J., Temime, B., and Hohener, P. (2011). Intrinsic biodegradation potential of aromatic hydrocarbons in an alluvial aquifer—potentials and limits of signature metabolite analysis and two stable isotope-based techniques. Water Res. 45, 4459–4469. doi: 10.1016/j.watres.2011.05.040
Muller, F. M. (1957). On methane fermentation of higher alkanes. Antonie Van Leeuwenhoek 23, 369–384. doi: 10.1007/BF02545890
Musat, F., Wilkes, H., Behrends, A., Woebken, D., and Widdel, F. (2010). Microbial nitrate-dependent cyclohexane degradation coupled with anaerobic ammonium oxidation. ISME J. 4, 1290–1301. doi: 10.1038/ismej.2010.50
Nascarella, M. A., Kostecki, P. T., Calabrese, E. J., and Click, D. (2002). “AEHS's 2001 survey of states' soil and groundwater cleanup standards,” in Contaminated Soil, Sediment, and Water (January/February), 15–68.
National Research Council (NRC). (1993). In Situ Bioremediation: When Does it Work? Washington, DC: National Academy Press.
Oka, A. R., Phelps, C. D., Zhu, X., Saber, D. L., and Young, L. Y. (2011). Dual biomarkers of anaerobic hydrocarbon degradation in historically contaminated groundwater. Environ. Sci. Technol. 45, 3407–3414. doi: 10.1021/es103859t
Orem, W. H., Tatu, C. A., Lerch, H. E., Rice, C. A., Bartos, T. T., Bates, A. L., et al. (2007). Organic compounds in produced waters from coalbed natural gas wells in the Powder River Basin, Wyoming, USA. Appl. Geochem. 22, 2240–2256.
Parisi, V. A., Brubaker, G. R., Zenker, M. J., Prince, R. C., Gieg, L. M., da Silva, M. L. B., et al. (2009). Field metabolomics and laboratory assessments of anaerobic intrinsic bioremediation of hydrocarbons at a petroleum-contaminated site. Microb. Biotechnol. 2, 202–212. doi: 10.1111/j.1751-7915.2009.00077.x
Rabus, R., Jarling, R., Lahme, S., Kuhner, S., Heider, J., Widdel, F., et al. (2011). Co-metabolic conversion of toluene in anaerobic n-alkane-degrading bacteria. Environ. Microbiol. 13, 2576–2586. doi: 10.1111/j.1462-2920.2011.02529.x
Rabus, R., Wilkes, H., Behrends, A., Armstroff, A., Fischer, T., Pierik, A. J., et al. (2001). Anaerobic initial reaction of n-alkanes in a denitrifying bacterium: evidence for (1-methylpentyl)succinate as initial product and for involvement of an organic radical in n-hexane metabolism. J. Bacteriol. 183, 1707–1715. doi: 10.1128/JB.183.5.1707-1715.2001
Reddy, C. M., Arey, J. S., Seewald, J. S., Sylva, S., Lemkau, K. L., Nelson, R. K., et al. (2012). Composition and fate of gas and oil released to the water column during the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. U.S.A. 109, 20229–20234. doi: 10.1073/pnas.1101242108
Rios-Hernandez, L. A., Gieg, L. M., and Suflita, J. M. (2003). Biodegradation of an alicyclic hydrocarbon by a sulfate-reducing enrichment from a gas condensate-contaminated aquifer. Appl. Environ. Microbiol. 69, 434–443.
Ritchie, G. D., Still, K. R., Alexander, W. K., Nordholm, A. F., Wilson, C. L., Rossi, J. 3rd., et al. (2001). A review of the neurotoxicity risk of selected hydrocarbon fuels. J. Toxicol. Environ. Health 4(Pt. B), 223–312. doi: 10.1080/10937400118874
Rojo, F. (2009). Degradation of alkanes by bacteria. Environ. Microbiol. 11, 2477–2490. doi: 10.1111/j.1462-2920.2009.01948.x
Rowland, S. J., West, C. E., Scarlett, A. G., Jones, D., and Frank, R. A. (2011). Identification of individual tetra− and pentacyclic naphthenic acids in oil sands process water by comprehensive two−dimensional gas chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 25, 1198–1204. doi: 10.1002/rcm.4977
Savage, K. N., Krumholz, L. R., Gieg, L. M., Parisi, V. A., Suflita, J. M., Allen, J., et al. (2010). Biodegradation of low-molecular-weight alkanes under mesophilic sulfate-reducing conditions: metabolic intermediates and community patterns. FEMS Microbiol. Ecol. 72, 485–495. doi: 10.1111/j.1574-6941.2010.00866.x
Siddique, T., Fedorak, P. M., and Foght, J. M. (2006). Biodegradation of short-chain n-alkanes in oil sands tailings ponds under methanogenic conditions. Environ. Sci. Technol. 40, 5459–5464. doi: 10.1021/es200649t
Siddique, T., Penner, T., Semple, K., and Foght, J. M. (2011). Anaerobic biodegradation of longer chain n-alkanes coupled to methane production in oil sands tailings. Environ. Sci. Technol. 45, 5892–5899. doi: 10.1021/es200649t
So, C. M., Phelps, C. D., and Young, L. Y. (2003). Anaerobic transformation of alkanes to fatty acids by a sulfate-reducing bacterium, strain Hxd3. Appl. Environ. Microbiol. 69, 3892–3900. doi: 10.1128/AEM.69.7.3892-3900.2003
Strąpoć, D., Mastalerz, M., Dawson, K., Macaladay, J. L., Callaghan, A. V., Wawrik, B., et al. (2011). Biogeochemistry of microbial coal-bed methane. Annu. Rev. Earth Planet. Sci. 39, 617–656.
Thauer, R. K., and Shima, S. (2008). Methane as fuel for anaerobic microorganisms. Ann. N.Y. Acad. Sci. 1125, 158–170. doi: 10.1196/annals.1419.000
Thom, C., Gilley, D. C., Hooper, J., and Esch, H. E. (2007). The scent of the waggle dance. PLoS Biol. 5:e228. doi: 10.1371/journal.pbio.0050228
von Netzer, F., Pilloni, G., Kleindienst, S., Krueger, M., Knittel, K., Grundger, T., et al. (2013). Enhanced gene detection assays for fumarate-adding enzymes allow uncovering anaerobic hydrocarbon degraders in terrestrial and marine systems. Appl. Environ. Microbiol. 79, 543–552. doi: 10.1128/AEM.02362-12
Voordouw, G., Grigoryan, A. A., Lambo, A., Lin, S., Park, H. S., Jack, T. R., et al. (2009). Sulfide remediation by pulsed injection of nitrate into a low temperature Canadian heavy oil reservoir. Environ. Sci. Technol. 43, 9512–9518. doi: 10.1021/es902211j
Wawrik, B., Mendivelso, M., Parisi, V. A., Suflita, J. M., Davidova, I. A., Marks, C. R., et al. (2012). Field and laboratory studies on the bioconversion of coal to methane in the San Juan Basin. FEMS Microbiol. Ecol. 81, 26–42. doi: 10.1111/j.1574-6941.2011.01272.x
Weiss, J. V., and Cozzarelli, I. M. (2008). Biodegradation in contaminated aquifers: incorporating microbial/molecular methods. Ground Water 46, 305–322. doi: 10.1111/j.1745-6584.2007.00409.x
Widdel, F., and Grundmann, O. (2010). “Biochemistry of the anaerobic degradation of non-methane alkanes,” in Handbook of Hydrocarbon and Lipid Microbiology, ed K. N. Timmis (Berlin: Springer-Verlag), 909–924.
Wilkes, H., Kuhner, S., Bolm, C., Fischer, T., Classen, A., Widdel, F., et al. (2003). Formation of n-alkane- and cycloalkane-derived organic acids during anaerobic growth of a denitrifying bacterium with crude oil. Org. Geochem. 34, 1313–1323.
Wilkes, H., Rabus, R., Fischer, T., Armstroff, A., Behrends, A., and Widdel, F. (2002). Anaerobic degradation of n-hexane in a denitrifying bacterium: further degradation of the initial intermediate (1-methylpentyl)succinate via C-skeleton rearrangement. Arch. Microbiol. 177, 235–243. doi: 10.1007/s00203-001-0381-3
Winderl, C., Schaefer, S., and Lueders, T. (2007). Detection of anaerobic toluene and hydrocarbon degraders in contaminated aquifers using benzylsuccinate synthase (bssA) genes as a functional marker. Environ. Microbiol. 9, 1035–1046. doi: 10.1111/j.1462-2920.2006.01230.x
Zedelius, J., Rabus, R., Grundmann, O., Werner, I., Brodkorb, D., Schreiber, F., et al. (2011). Alkane degradation under anoxic conditions by a nitrate-reducing bacterium with possible involvement of the electron acceptor in substrate activation. Environ. Microbiol. Rep. 3, 125–135. doi: 10.1111/j.1758-2229.2010.00198.x
Zengler, K., Richnow, H. H., Rossello-Mora, R., Michaelis, W., and Widdel, F. (1999). Methane formation from long-chain alkanes by anaerobic microorganisms. Nature 401, 266–269. doi: 10.1038/45777
Zhou, L., Li, K.-P., Mbadinga, S. M., Yang, S.-Z., Gu, J.-D., and Mu, B.-Z. (2012). Analyses of n-alkanes degrading community dynamics of a high-temperature methanogenic consortium enriched from production water of a petroleum reservoir by a combination of molecular techniques. Exotoxicology 21, 1680–1691. doi: 10.1007/s10646-012-0949-5
Keywords: anaerobic, metabolites, alkylsuccinates, alkanes, subsurface
Citation: Agrawal A and Gieg LM (2013) In situ detection of anaerobic alkane metabolites in subsurface environments. Front. Microbiol. 4:140. doi: 10.3389/fmicb.2013.00140
Received: 02 January 2013; Accepted: 17 May 2013;
Published online: 04 June 2013.
Edited by:
Amy V. Callaghan, University of Oklahoma, USAReviewed by:
Rachel N. Austin, Bates College, USACopyright © 2013 Agrawal and Gieg. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Lisa M. Gieg, Petroleum Microbiology Research Group, Department of Biological Sciences, University of Calgary, 2500 University Drive NW, Calgary, AB T2N 1N4, Canada e-mail:bG1naWVnQHVjYWxnYXJ5LmNh
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.