 Jeanette E. Bröms
Jeanette E. Bröms Anders Sjöstedt* and Moa Lavander
Anders Sjöstedt* and Moa Lavander
- Department of Clinical Microbiology, Clinical Bacteriology, and Laboratory for Molecular Infection Medicine Sweden, Umeå University, Umeå, Sweden
Francisella tularensis is a highly virulent gram-negative intracellular bacterium that causes the zoonotic disease tularemia. Essential for its virulence is the ability to multiply within host cells, in particular monocytic cells. The bacterium has developed intricate means to subvert host immune mechanisms and thereby facilitate its intracellular survival by preventing phagolysosomal fusion followed by escape into the cytosol, where it multiplies. Moreover, it targets and manipulates numerous host cell signaling pathways, thereby ameliorating the otherwise bactericidal capacity. Many of the underlying molecular mechanisms still remain unknown but key elements, directly or indirectly responsible for many of the aforementioned mechanisms, rely on the expression of proteins encoded by the Francisella pathogenicity island (FPI), suggested to constitute a type VI secretion system. We here describe the current knowledge regarding the components of the FPI and the roles that have been ascribed to them.
Introduction
Francisella tularensis is a highly infectious gram-negative intracellular bacterium that causes the disease tularemia (Oyston et al., 2004). Two main subspecies, F. tularensis subsp. tularensis and F. tularensis subsp. holarctica (also known as type A and type B, respectively), are responsible for the majority of infections and disease, which are particularly severe if caused by type A strains. Importantly, F. tularensis has the ability to infect a multitude of hosts, including mammals, fish, amphibians, protozoa and arthropods (reviewed in Mörner and Addison, 2001; Keim et al., 2007). Although primarily a vector-transmitted disease, F. tularensis infections can also occur through inhalation of contaminated materials, ingestion and skin abrasions (Dennis et al., 2001). From the site of infection, the bacteria will spread to regional lymph nodes, liver and spleen, where they rapidly grow and overwhelm the host’s immune defenses (Fortier et al., 1991).
The pathogenicity of F. tularensis is believed to depend on its ability to replicate within host cells, particularly macrophages (Tärnvik, 1989). F. tularensis enters host macrophages by triggering the formation of asymmetric, spacious pseudopod loops (Clemens et al., 2005). It then modifies the endocytic pathway of the macrophage, preventing phagolysosomal fusion, upon which it escapes into the cytosol and multiplies (Golovliov et al., 2003a; Clemens et al., 2004; Santic et al., 2005a). Several factors that regulate or facilitate phagosome escape and intracellular survival of F. tularensis have been identified (Dean et al., 2009; Meibom and Charbit, 2009; Qin et al., 2009; Mohapatra et al., 2010), however, the underlying molecular mechanisms still remain largely unknown. A gene cluster frequently encountered in screens aimed at identifying elements important for intracellular growth and/or virulence, is the Francisella pathogenicity island (FPI; reviewed in Nano et al., 2004; Nano and Schmerk, 2007; Meibom and Charbit, 2009), which has been suggested to encode a type VI secretion system (de Bruin et al., 2007; Nano and Schmerk, 2007; Bingle et al., 2008; Ludu et al., 2008; Barker et al., 2009; Bröms et al., 2009).
Note: F. tularensis subsp. novicida will be referred to as F. novicida and the FPI nomenclature proposed by Ludu et al. (2008) will be used throughout (Table 1; Figure 1).
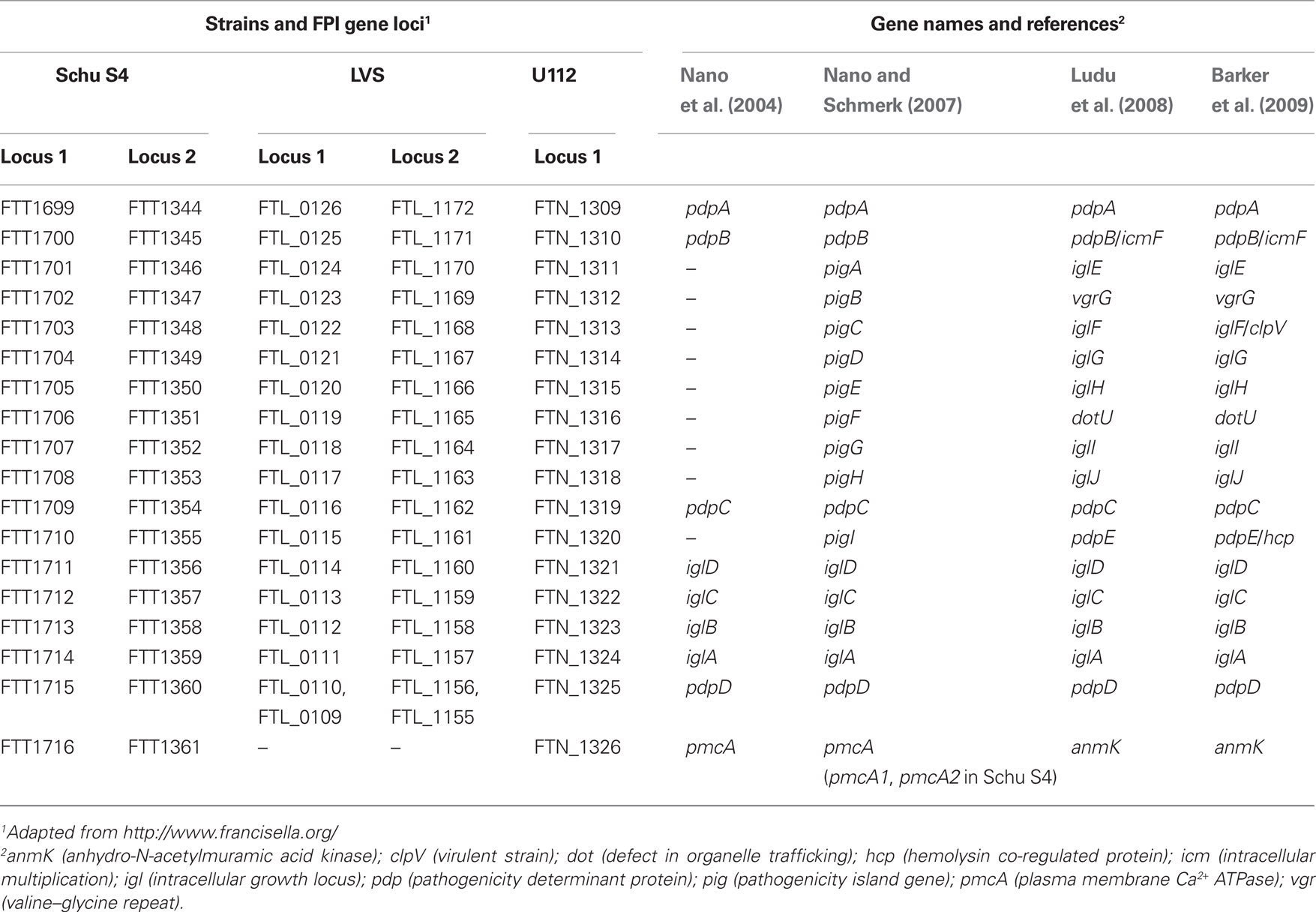
Table 1. FPI gene nomenclature.
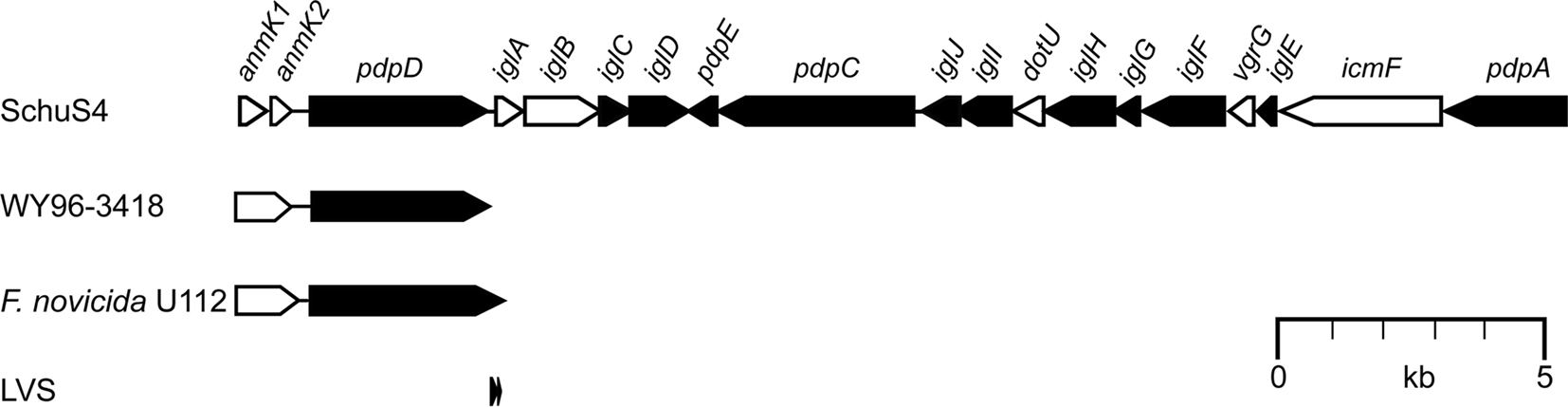
Figure 1. The Francisella pathogenicity island (FPI). A schematic representation of the Francisella tularensis Schu S4 variant of the FPI with a consensus nomenclature adapted from Ludu et al. (2008). The ORFs of the FPI are essentially the same in all subspecies, with the exception of the anmK–pdpD region. In type A strains like Schu S4 (clade A.I) or WY96-3418 (clade A.II), pdpD encodes a 1,195 aa protein, which is somewhat larger in F. novicida (1,245 aa). The anmK gene product (371 aa in F. novicida) is truncated in WY96-3418, and may be expressed as two separate ORFs in Schu S4 (anmK1 and anmK2). F. tularensis type B strains, e.g., LVS, lack anmK and most of the pdpD gene due to a large deletion within this region. The ORFs are drawn to scale, and their relative sizes indicated. Black arrows indicate gene products with no significant homology to other proteins; while white arrows represent products with homologs in other bacterial systems (see text for details).
The Francisella Pathogenicity Island
Duplication of the FPI in Strains of F. Tularensis
The 33-kb pathogenicity island, designated FPI (Figure 1), consists of 16–19 ORFs and its crucial role in the virulence of F. tularensis is well established (Golovliov et al., 2003b; Nano et al., 2004; Twine et al., 2005; Tempel et al., 2006; Santic et al., 2007, 2009; Weiss et al., 2007; Bönquist et al., 2008; Ludu et al., 2008; Vonkavaara et al., 2008; Bröms et al., 2009; Kraemer et al., 2009; Schmerk et al., 2009b; åhlund et al., 2010; Bröms et al., unpublished). It was first identified in 2004 (Nano et al., 2004) by bioinformatic analysis of the partly sequenced genomes of F. tularensis subsp. tularensis strain Schu S4 (Karlsson et al., 2000; Prior et al., 2001) and F. novicida strain U112 (Nano et al., 2004). All subsequently sequenced Francisella genomes have confirmed the presence of the FPI, suggesting that it is part of the core genome (Larsson et al., 2009). However, in contrast to the rest of the genome, the FPI has lower G + C content, indicating horizontal gene transfer (Nano et al., 2004).
Interestingly, the FPI is duplicated in all the subsp. of F. tularensis (F. tularensis subsp. holarctica, mediasiatica, and tularensis), but is present in a single copy in F. novicida and F. philomiragia (Nano et al., 2004; Larsson et al., 2009).
Larsson et al. (2009) recently suggested that this duplication event was facilitated by the insertion of ISFtu1 insertion sequence element adjacent to the FPI and an rRNA operon of an ancestral genome, which enabled duplication of the FPI by non-reciprocal recombination. Interestingly, as deduced from the genomic locations of the units, one of them was identified as ancestral whereas the second, more recently acquired, has integrated in various locations in the different genomes. As a consequence of the duplication, FPI null mutants are easier to generate in F. novicida, which is also the species most extensively used to analyze the role of individual FPI proteins. Noteworthy, there is a gene cluster in F. novicida, encompassing orfs FTN_0040–FTN_0052, which show homology to the FPI, albeit rudimentary, and with a different organization of the “true” FPI genes. The similarities are particularly pronounced for the core components IglA, IglB, PdpB/IcmF, and DotU where FTN_0042, FTN_0043, FTN_0040, and FTN_0051, respectively, show high (IglA and IglB) or intermediate (PdpB and DotU) protein sequence homology (Larsson et al., 2009). Further, protein-BLAST (http://blast.ncbi.nlm.nih.gov/) reveals that there are genes encoding products with weak homology to PdpA, PdpC, PdpD, IglD, IglH, and IglI, interspersed with orfs that reveal no FPI similarity. This gene cluster has not been widely discussed, nor its functionality investigated and its role in F. novicida virulence remains unknown.
Interestingly, combined work by several groups suggests that the second FPI locus may be functionally redundant in strains of F. tularensis, as transposon or deletion mutants for which only one of the two gene copies have been targeted are unaffected for virulence (Golovliov et al., 2003b; Kawula et al., 2004; Qin and Mann, 2006; Nano and Schmerk, 2007), in contrast to mutants for which both gene copies have been removed by targeted mutagenesis (Golovliov et al., 2003b; Bönquist et al., 2008; Bröms et al., 2009). In fact, Schu S4 can tolerate the loss of one of its two FPI loci (targeted deletions of regions encompassing FTT_1699–FTT_1715 or FTT_1344–FTT_1360), without substantial loss of virulence toward mice infected by the intradermal route (A. Sjöstedt and W. Conlan, unpublished). Contradicting these results is a study by Su et al. (2007), which suggests that both gene copies of iglA, iglB, or iglC are required for full virulence of LVS in a mouse model of respiratory tularemia and a study by Maier et al., demonstrating that HimarFT insertion mutants in one of the iglB, iglC, or iglD loci result in only partial loss of J774 monolayer integrity upon infection, in contrast to when parental LVS was used (Maier et al., 2007). Thus, it cannot be ruled out that the duplication event, and hence doubling of gene dosage, may provide some benefit to the pathogenic F. tularensis type A and B strains, and that this may have contributed to their differentiation from the human non-pathogenic, or rarely pathogenic, F. novicida and F. philomiragia strains, although there is little data to support this. It should be noted that FPI mutants even in highly virulent subspecies of F. tularensis, e.g., in the Schu S4 strain, are avirulent. Thus, despite that one CFU of the wild-type strain kills mice irrespective of route of inoculation, the lethal dose of the iglB, iglC, and iglD Schu S4 mutants are >>107 CFU. Presumably, the lack of phagosomal escape renders them avirulent (Twine et al., 2005; Kadzhaev et al., 2009). Moreover, they are non-immunogenic since mice infected with the iglC mutant of Schu S4 are not protected against a subsequent infection with a subspecies tularensis strain (Twine et al., 2005).
Genetic Differences within the FPI Cluster
The ORFs of the FPI are essentially conserved in all subspecies, with the exception of the anmK–pdpD region (Figure 1). In F. novicida, anmK (originally denoted pmcA) is suggested to encode a 371 aa gene product, AnmK, which shows high identity to members of COG2377 (predicted molecular chaperone distantly related to HSP70-fold metalloproteases) (Nano and Schmerk, 2007). However, in type A strains like Schu S4 and FSC033 (clade A.I), anmK may be expressed as two separate ORFs due to the presence of premature stop codons at positions 190 and 328, followed by an ATG start codon at position 194. In type A strains like WY96-3418 and B38 (clade A.II), AnmK has a premature stop codon at position 328, suggesting that this group encodes a truncated form of AnmK (Nano and Schmerk, 2007). How these genetic differences affect AnmK function has not been addressed so far, but AnmK of F. novicida has been shown to contribute to virulence, albeit to a small degree (Nano and Schmerk, 2007; Ludu et al., 2008). Supporting the notion that AnmK is not critical for Francisella pathogenicity is the fact that F. tularensis type B strains, virulent strains as well as LVS, lack the anmK gene due to a deletion that extends from 107 bp upstream of anmK and encompasses most of the adjacent pdpD gene (Nano et al., 2004; Champion et al., 2009). In F. novicida, pdpD encodes a 1,245 aa (1,195 aa in type A strains) outer membrane protein that has been shown to be important for virulence in mice (Ludu et al., 2008), while the deletion within type B strains is presumed to result in two minor ORFs (corresponding to residues 992–1124 and 1129–1195 of PdpD from type A strains) that are likely to be non-functional (Ludu et al., 2008; Champion et al., 2009). It has been speculated that the presence or absence of pdpD may account, in part, for the strain-to-strain specific difference in virulence between type A and type B strains (Nano et al., 2004).
FPI Nomenclature – Naming of Components
Over the years, the nomenclature of the FPI genes has been altered in attempts to adapt to the increasing knowledge of the function of individual proteins (Table 2). Already in 1997, the first FPI gene (later named iglC) was identified, and its expression shown to be prominently induced during intracellular growth (Golovliov et al., 1997). The gene was later recognized as a member of a four gene operon, which, due to its requirement for intramacrophage growth, was denoted iglABCD (intracellular growth locus) (Gray et al., 2002; Golovliov et al., 2003b, Figure 1; Tables 1 and 2). In 2004, at the time of the discovery of the FPI, a handful of the predicted ORFs were named, including pmcA (today anmK), which share homology to molecular chaperones, and the large ORFs pdpA, pdpB, pdpC, and pdpD (pathogenicity determinant protein) (Nano et al., 2004; Table 1). Later, pdpB was named IcmF, based on its homology to this crucial T6SS core component (Ludu et al., 2008). In 2007, the remaining ORFs of the island were recognized as most likely functional genes and thus denoted pigA to I (pathogenicity island gene) (Nano and Schmerk, 2007; Table 1). Subsequently, limited homologies have been identified between pigB, pigC, pigF, pigI, and the T6SS core components vgrG, clpV, dotU, and hcp, respectively, so these Francisella genes have been renamed accordingly (Ludu et al., 2008; Barker et al., 2009; Table 1). Also the remaining pig genes, i.e., pigA, pigD, pigE, pigG, and pigH have been renamed iglE, iglG, iglH, iglI, and iglJ respectively (Ludu et al., 2008; Barker et al., 2009; Table 1), suggesting that these genes, similar to iglABCD, are required for intracellular growth. However, with the exception of iglI of F. novicida (Barker et al., 2009), there is no published data to support that this is the case (Table 3). In fact, our data strongly suggest that iglG is dispensable for intracellular growth of F. tularensis strain LVS in both J774 macrophages and mouse peritoneal exudate (PEC) cells (Bröms et al., unpublished) (Table 3). Moreover, an iglI null mutant of LVS is able to efficiently grow within J774 cells, which is in contrast to the results observed by Barker et al. (2009) for F. novicida (above) (Bröms et al., unpublished; Table 3). This is an exciting finding, since it suggests that the phenotypes of FPI mutants may be species-specific, but it clearly complicates the use of a common FPI nomenclature; thus, the name IglG (Intracellular growth locus G) and IglI (Intracellular growth locus I) may perhaps be appropriate for F. novicida, but evidently not for LVS. Also, we do not see any obvious reason to distinguish pdpA from the iglABCD genes required for intracellular growth, since a pdpA mutant of F. novicida also clearly exhibits very limited replication in host cells (Schmerk et al., 2009a). Moreover, the homologies reported between some of the Francisella proteins and T6SS core components, e.g., PigC and ClpV (Barker et al., 2009), are weak and do not provide any convincing evidence for their relatedness, nor do they provide support when it comes to assigning a function to the specific FPI protein (Bröms et al., unpublished).
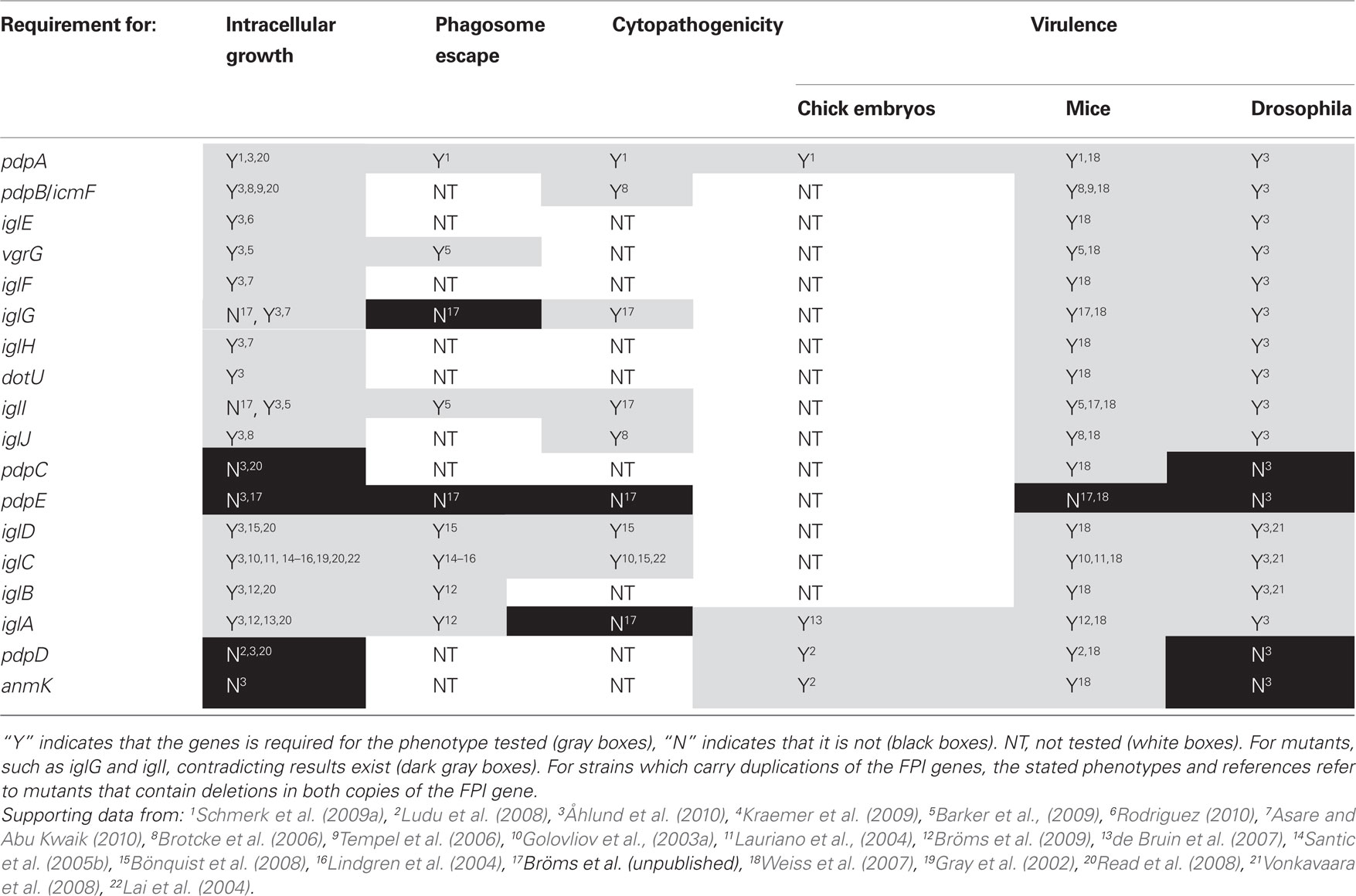
Table 2. Effects of FPI gene mutations on Francisella pathogenicity.
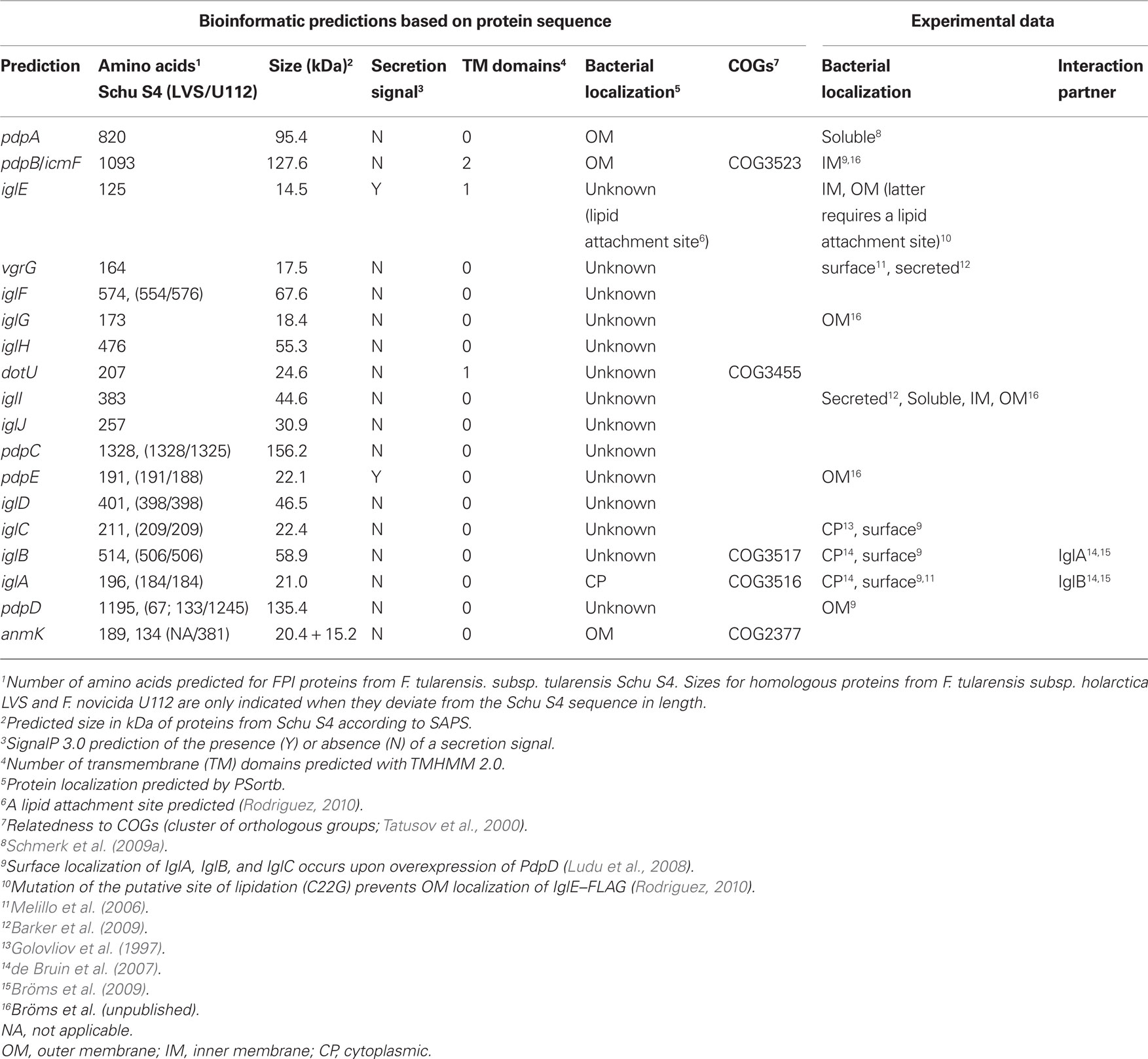
Table 3. In silico analysis of FPI proteins, demonstrated protein localization and protein–protein interactions.
For these reasons, we believe that the existing FPI nomenclature should be modified to better reflect the current knowledge of FPI protein function, but also to take into account that for many of these proteins we still have no assigned functions. In fact, since the Francisella T6SS is so different from all other T6SS described so far (Bingle et al., 2008) it is not unlikely that some of the FPI proteins will exhibit functions distinct from their homologous T6SS core components. For simplicity, we therefore propose that all FPI genes should be named igl, regardless of function, but this will be discussed more thoroughly elsewhere.
The FPI – Its Role in Phagosome Biogenesis, Intracellular Growth and Virulence
Similar to other intracellular pathogens like Listeria monocytogenes, Shigella flexneri, and Rickettsia spp. (Ray et al., 2009), Francisella rapidly escapes from the phagosome into the host cell cytosol where it replicates (Clemens et al., 2004; Santic et al., 2005a,b; Bönquist et al., 2008). The Francisella containing phagosome (FCP) transiently acquires early (EEA-1 and Rab5) and late (Rab7, CD63, LAMP-1, and LAMP-2) endosomal markers, but no/little cathepsin D, suggesting limited fusion to lysosomes (Clemens et al., 2004; Santic et al., 2005a, 2008; Bönquist et al., 2008). Inhibition of the proton ATPase pump within 15–60 min post infection has been shown to result in a significant delay in phagosomal escape (Chong et al., 2008; Santic et al., 2008), indicating the importance of acidification for FCP disruption and bacterial escape. Importantly, these results have recently been challenged by Clemens et al. (2009).
A vast number of studies have identified the FPI as a key player in the ability of F. tularensis to modulate phagosome maturation and escape into the host cell cytosol. This is likely to be the consequence of T6SS-mediated translocation of bacterial effector protein(s) directly involved in modulation of phagosome biogenesis and lysis of the phagosomal membrane. However, none of the FPI-encoded proteins required for escape so far hold any properties of cytolysins, pore-forming toxins, or hydrolytic enzymes commonly used by intracellular pathogens to disrupt the phagosomal membrane (Ray et al., 2009). This suggests that Francisella may employ novel mechanisms to promote bacterial escape into the cytosol, or merely that the effector proteins are encoded outside of the FPI cluster.
Since FPI mutants that fail to escape also fail to multiply, it has been difficult to determine whether the encoded gene products also contribute to multiplication, or to later stages of infection. Interestingly, FPI genes reach their maximal expression at the end of cytosolic replication, which may suggest a role for the FPI during late stages of infection, or perhaps, may provide an advantage (“ready and armed”) to bacteria that are about to infect new cells upon lysis of their current host cell (Wehrly et al., 2009).
IglC is the best characterized FPI protein so far, together with MglA, a positive regulator encoded outside of the FPI. Both have been shown to be essential for the ability of F. tularensis to modulate the biogenesis of the phagosome to avoid lysosomal fusion (Santic et al., 2005b; Bönquist et al., 2008), to escape into the host cytosol (Lindgren et al., 2004; Santic et al., 2005b), and to induce apoptosis of J774 cells (Lai et al., 2004; Table 3). Consequently, mglA and iglC mutations diminish intracellular survival and growth in many different macrophage-like cell lines (Gray et al., 2002; Golovliov et al., 2003a; Lauriano et al., 2003; Lindgren et al., 2004; Santic et al., 2005b), as well as ameba (Lauriano et al., 2004) and insect cells (Read et al., 2008; Vonkavaara et al., 2008; Santic et al., 2009; Table 3). To date, most of the genes located in the FPI, such as iglA, iglB, iglC, iglD, iglI, iglG, pdpA, pdpB, and pdpD, have been demonstrated to be required either for intramacrophage growth and/or virulence of at least one subspecies (Golovliov et al., 2003b; Lai et al., 2004; Nano et al., 2004; Tempel et al., 2006; de Bruin et al., 2007; Santic et al., 2007; Weiss et al., 2007; Bönquist et al., 2008; Chong et al., 2008; Ludu et al., 2008; Bröms et al., 2009; Schmerk et al., 2009b; Bröms et al., unpublished; Table 3). Interestingly, some gene products, such as IglG, have been shown to be required for virulence but dispensable for intracellular growth, while PdpE appears to be dispensable for both processes (Bröms et al., unpublished; Table 3). Noteworthy, many of the earlier FPI studies lacked the tools for precise genetic analysis and proper complementation controls, and this should be taken into account when discussing the phenotypes of Francisella strains with lesions in FPI genes.
FPI-Mediated Manipulation of the Host Inflammatory Response
The pathogenesis of tularemia appears to be critically dependent on the ability of F. tularensis to modulate the host immune response by a number of mechanisms, including degradation of serum antibodies (Crane et al., 2009), induction of immunosuppressive cytokines and prostaglandin E2 in the lungs of infected mice (Bosio et al., 2007; Woolard et al., 2008), suppression of responses to secondary stimuli in vivo (Bosio and Dow, 2005), inhibition of the respiratory burst in human neutrophils (McCaffrey and Allen, 2006), impairment of phosphatidylinositol 3-kinase-mediated (Parsa et al., 2006) and IFN-γ-mediated signaling (Parsa et al., 2008) and downregulation of several TLRs in monocytes (Telepnev et al., 2003), rapid induction of apoptosis in J774 cells and mouse tissue macrophages (Lai et al., 2004; Wickstrum et al., 2009), activation of the ASC inflammasome (Mariathasan et al., 2004; Fernandes-Alnemri et al., 2010; Jones et al., 2010; Rathinam et al., 2010), as well as rapid activation of Ras (Al-Khodor and Abu Kwaik, 2010). Several of these mechanisms are directly or indirectly linked to FPI proteins since specific FPI mutants or mutants within mglA show phenotypes deviant from the parent strain.
Already within 15 min after monocytic ingestion of wild-type F. novicida, recruitment of PKCα and PKCβI to the SOS2/GrB2 complex occurs, leading to activation of Ras (Figure 2). None of these events take place after infection with an iglC mutant of F. novicida, instead rapid activation of caspase-3 and cell death follows (Al-Khodor and Abu Kwaik, 2010; Figure 2). Thus, Ras activation is advantageous to the bacterium since it allows a prolonged replication in the cytosol. As it occurs so rapidly, the phagosomal localization of the iglC mutant may not necessarily be the reason for the aberrant Ras activation, instead, the effect may directly or indirectly relate to the function of IglC.
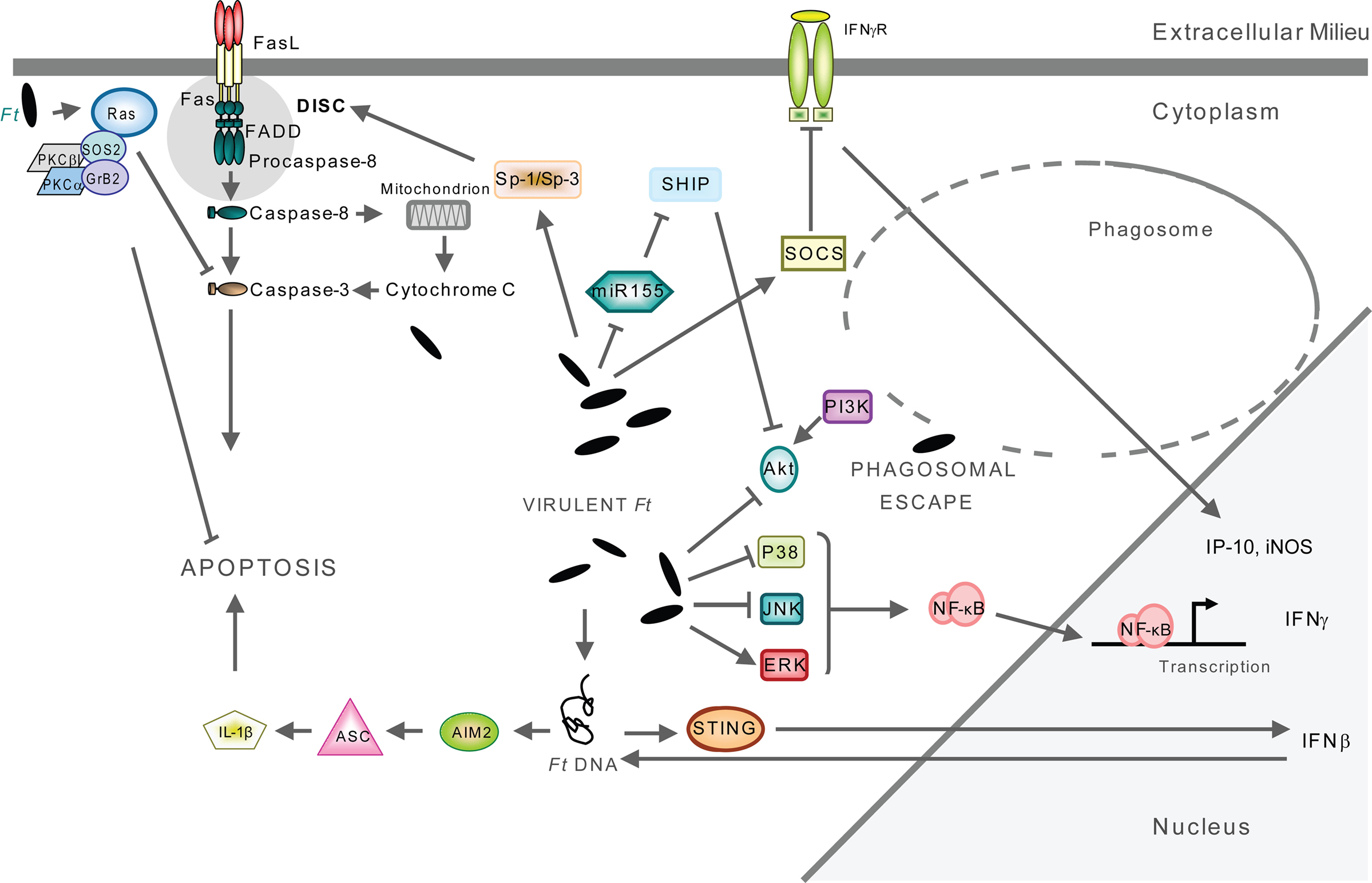
Figure 2. Signaling triggered by F. tularensis located in the cytosol. Host cell signaling occurs after F. tularensis escapes from the phagosome to reside and multiply within the cytoplasm of the monocytic cells. The infection eventually leads to apoptosis of the host cell. The engagement of a multitude of signaling pathways have been described, for details see references in the text. Only pathways that have been experimentally verified are included in the figure.
The proinflammatory response to F. tularensis in monocytic cells is TLR2-dependent (Cole et al., 2006, 2007; Katz et al., 2006; Li et al., 2006; Thakran et al., 2008; Abplanalp et al., 2009). Moreover, it has been demonstrated that Francisella colocalizes with TLR2 and MyD88 inside macrophages (Cole et al., 2007; Figure 3), signals through TLR2 from within the phagosome (Cole et al., 2008; Figure 3), and that phagosome-contained F. tularensis demonstrate a distinct signaling pattern (Cole et al., 2008). In the latter case, macrophages infected with an iglC mutant showed increased expression of a subset of TLR2-dependent, proinflammatory genes, but decreased expression of other genes encoding, e.g., IFN-γ, iNOS, and IP-10 (Cole et al., 2008). The latter subset was dependent on IFN-β, indicating that this cytokine is required for full induction of the macrophage proinflammatory response (Figure 2). While the iglC mutant greatly increased IL-1β mRNA expression in wild-type macrophages, IL-1β secretion was not observed (Cole et al., 2008). Thus, TLR2-dependent expression of IFN-β, IFN-β-stimulated genes, and IL-1β secretion requires bacterial escape from the phagosome. Noteworthy, in the absence of IFN-β, intracellular bacterial numbers were increased showing the central role of the cytokine in modulating the host response and controlling F. tularensis infection (Cole et al., 2008).
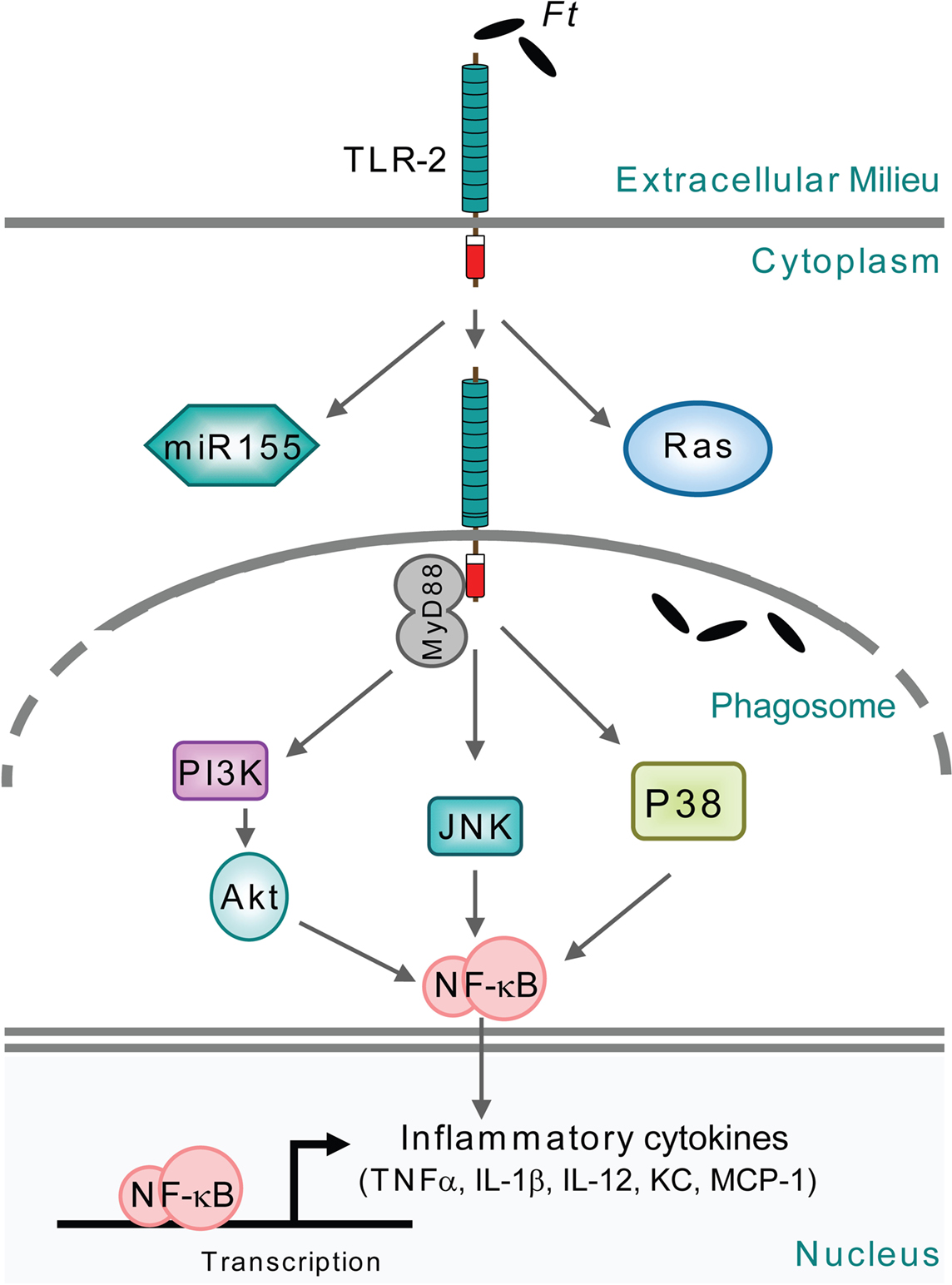
Figure 3. Signaling triggered by F. tularensis located in the phagosome. The signaling occurs during the brief period after uptake of the bacteria by monocytic cells and before they escape into the cytoplasm. Moreover, the signaling also occurs during infection with FPI mutants, e.g., within iglC, which are unable to escape from the phagosome.
A detailed characterization of the F. tularensis-induced IFN-β signaling has been revealed in a series of recent publications, resulting in the identification of F. novicida as a prototypic agent for AIM2-mediated inflammasome activation (Fernandes-Alnemri et al., 2010; Jones et al., 2010; Rathinam et al., 2010; Ulland et al., 2010). AIM-2 activation involves recognition of cytosolic DNA, which triggers formation of the AIM2 inflammasome by inducing AIM2 oligomerization (Fernandes-Alnemri et al., 2009; Figure 2). The AIM2-DNA complex then recruits and subsequently oligomerizes the caspase-1-activating adaptor protein ASC into the large ASC pyroptosome, which in turn activates caspase-1 leading to cleavage of IL-1β and subsequent apoptosis (Fernandes-Alnemri et al., 2007; Figure 2). By infecting AIM2-deficient mice with F. novicida, it was concluded that cytosolic invasion by F. tularensis leads to STING-mediated production of type I interferons, such as IFN-β, which is a prerequisite for the subsequent AIM2 activation, leading to caspase-1 activation, IL-1β and IL-18 secretion and inflammasome-mediated cell death (Gavrilin et al., 2006; Mariathasan et al., 2006; Henry et al., 2007; Cole et al., 2008; Figure 2). This chain of events critically depends on a cytosolic localization of bacteria as shown by infections with FPI mutants unable to escape the phagosome, such as iglC, or mutants within mglA, the positive regulator encoded outside of the FPI (Gavrilin et al., 2006; Cole et al., 2008). Similarly, F. novicida vgrG and iglI mutants, both unable to escape from the phagosome, were recently shown to be defective for IL-1β release (Barker et al., 2009). Thus, for F. novicida, there appears to be a strong correlation between the degree of phagosomal escape and inflammasome activation. In view of this, it was somewhat surprisingly that one of the studies found that the endosomal-acidification inhibitor bafilomycin completely prevented the production of type I interferons, implying the phagosome as the source of DNA for inflammasome activation (Fernandes-Alnemri et al., 2010). However, the divergent findings may be reconciled if it is assumed that the amounts of F. novicida DNA initially released into the cytosol upon escape per se are not sufficient to activate AIM2 but still can trigger production of type I interferons such as IFN-β. This will lead to monocyte activation and enhanced bacterial degradation in the phagosome, resulting in increased DNA release into the cytosol. Thereby, the accumulation of cytosolic DNA may reach a threshold required for AIM2 inflammasome activation. The role of bafilomycin in this context may be to prevent the type I interferon-enhanced killing of F. tularensis in the phagosome.
Francisella tularensis infection leads to interference of several signaling pathways crucial for the control of intracellular infection. One such example is the IFN-γ signaling pathway, which is essential although alone not sufficient to control F. tularensis infection of monocytic cells (Anthony et al., 1989; Lindgren et al., 2004; Edwards et al., 2009). The role of this pathway has been analyzed by studies of the IFN-γ-induced STAT1 activation in human and murine monocytic cells (Parsa et al., 2008; Roth et al., 2009). The interference with IFN-γ per se was dependent on upregulation of SOCS3, a well-known inhibitor of IFN-γ-mediated signaling, and accompanied by the suppression of IP-10 and iNOS production, resulting in increased intracellular bacterial survival (Figure 2). This signaling interference is independent of phagosomal escape and replication since mutants in mglA or iglD were still able to efficiently suppress IFNγ-induced STAT1 (Parsa et al., 2008). The important role of IFN-γ has been corroborated by determining its role when added to F. tularensis-infected cell cultures. Then, it was observed that treatment of mouse PEC cells conferred a static effect on wild-type LVS bacteria, whereas the treatment had a bactericidal effect on the iglC mutant demonstrating that the effect of IFN-γ is dependent on the subcellular localization of Francisella (Lindgren et al., 2004). There are conflicting data on the mechanisms effectuated by IFN-γ since one study reported that IFN-γ treatment did not affect the phagosomal escape and subcellular localization of Schu S4 in murine bone marrow-derived macrophages or human monocyte-derived macrophages (Edwards et al., 2009); two others that the subcellular localization of LVS was marginally affected in murine PEC (Lindgren et al., 2004), but not in murine J774A.1 cells (Bönquist et al., 2008); and a fourth that F. novicida was completely confined to the phagosome of human monocyte-derived macrophages (Santic et al., 2005a). Some of the discrepancies may relate to the use of different infection models and strains of F. tularensis, but additional studies are required to clarify this.
Mitogen-activated protein kinase (MAPK) pathways, such as p42/p44 (Erk1/2), c-Jun terminal kinase (JNK) and p38, of monocytic cells are also modulated by F. tularensis during infection. Hrstka et al. (2005) observed that apoptosis induced by LVS was associated with reduced p38 MAPK activity and activation of p42/p44 kinases (Figure 2). Moreover, Telepnev et al. (2005) showed that LVS is able to rapidly downregulate the initial activation of p38 and c-Jun, and hence cytokine secretion and that IglC is critical for this process. Thus, unlike the iglC mutant, LVS eventually downregulates the initial proinflammatory response in monocytic cells.
Moreover, phagosomal escape has been shown to be accompanied by marked downregulation of the phosphoinositol 3 kinase PI3K/Akt pathway and subsequent suppression of NF-kB-mediated production of inflammatory cytokines (Parsa et al., 2006; Rajaram et al., 2006; Figure 2). Suppression does not occur when an mglA mutant is used; instead it leads to increased proinflammatory production and lower IL-10 release (Rajaram et al., 2006; Figure 3). The pathway is under negative control of the inositol phosphatase SHIP, which becomes phosphorylated upon infection by F. tularensis (Parsa et al., 2006; Figure 2). Interestingly, the microRNA-155 (MiR-155) that targets and negatively regulates SHIP is induced by F. novicida but not Schu S4 (Figure 2), thereby leading to an enhanced proinflammatory response in the former case (Cremer et al., 2009). Furthermore, the expression of the PI3K/Akt pathway members is preferentially downregulated in macrophages infected with F. tularensis Schu S4 compared to F. novicida (Butchar et al., 2008).
The PI3K/Akt pathway is also related to host-cell death. Phagosomal escape of F. novicida leads to activation of Sp-1/Sp-3, upregulation of Fas and recruitment of the DISC complex (Rajaram et al., 2009), which in turn activates caspases-3 and -8 leading to apoptosis (Lai and Sjöstedt, 2003; Santic et al., 2009; Figure 2). Since activation of PI3K/Akt promotes phagosome–lysosome fusion (Rajaram et al., 2009), this should limit intracellular bacterial growth and host-cell death. In corroboration of this scenario, it was found that constitutive activation of Akt, or deletion of SHIP, promotes phagolysosomal fusion and limits bacterial replication in the host cytosol, as well as preventing subsequent induction of Fas expression and cell death (Rajaram et al., 2009).
Akt is also involved in phagosome biogenesis through activation of Rab5 (Barbieri et al., 1998) and the assembly of NADPH oxidase leading to triggering of the oxidative burst (Chen et al., 2003; Hoyal et al., 2003). Thereby, modulation of the PI3K/Akt pathway by F. tularensis may serve multiple purposes; downmodulation of the proinflammatory response and the oxidative burst, induction of host-cell death and limiting the biogenesis of the FCP.
The exact roles of the FPI proteins in modulating essentially all of the aforementioned pathways described is enigmatic since it is possible that the bacterium, once in the cytosol, mediates the effects via non-FPI-dependent mechanisms, e.g., via signaling through cytosolic receptors. Thus, a possible scenario is that the essential role of the FPI proteins simply is to enable the phagosomal escape.
Regulation of the FPI – a complex story
Studies in pathogens such as Burkholderia pseudomallei, Salmonella enterica, Vibrio cholerae, Pseudomonas aeruginosa, and enteroaggregative Escherichia coli have led to the identification of regulatory determinants and mechanisms underlying expression of T6SS gene clusters. These complex transcriptional regulatory networks involve two-component systems (Ishikawa et al., 2009; Syed et al., 2009), quorum sensing systems (Deziel et al., 2005; Ishikawa et al., 2009; Lesic et al., 2009), alternative sigma factor σ54 (Pukatzki et al., 2006; Ishikawa et al., 2009; Syed et al., 2009), transcriptional activators (Deziel et al., 2005; Dudley et al., 2006; Lesic et al., 2009; Sun et al., 2010), and histone-like proteins (Lucchini et al., 2006; Castang et al., 2008). In addition, upregulation of most T6S clusters has been shown to depend on contact with, or intracellular growth within, the host cell during infection (Das et al., 2000; Parsons and Heffron, 2005; Chugani and Greenberg, 2007; Shalom et al., 2007). In F. tularensis, this is consistent with the demonstration that FPI genes are induced during growth within macrophages (Golovliov et al., 1997; Baron and Nano, 1998; Twine et al., 2006; de Bruin et al., 2007; Chong et al., 2008; Wehrly et al., 2009). To date, the environmental stimuli and mechanisms that govern regulation of the FPI genes and thus virulence of Francisella spp. are as yet unknown, although iron depletion (Deng et al., 2006; Lenco et al., 2007), and oxidative stress (Lenco et al., 2005) have been shown to induce expression of FPI genes.
Combined data from several research groups suggest the involvement of at least six regulatory proteins in controlling FPI gene expression; MglA, SspA, FevR (PigR), MigR, Hfq, and PmrA (Figure 4; Baron and Nano, 1998; Lauriano et al., 2004; Brotcke et al., 2006; Charity et al., 2007; Mohapatra et al., 2007; Buchan et al., 2009; Meibom et al., 2009). Some of these also regulate expression of genes outside the FPI, however, whereas MglA, SspA, and FevR appear to regulate the same set of genes, PmrA and Hfq control separate groups of genes (Brotcke et al., 2006; Charity et al., 2007; Guina et al., 2007; Mohapatra et al., 2007; Brotcke and Monack, 2008; Meibom et al., 2009).
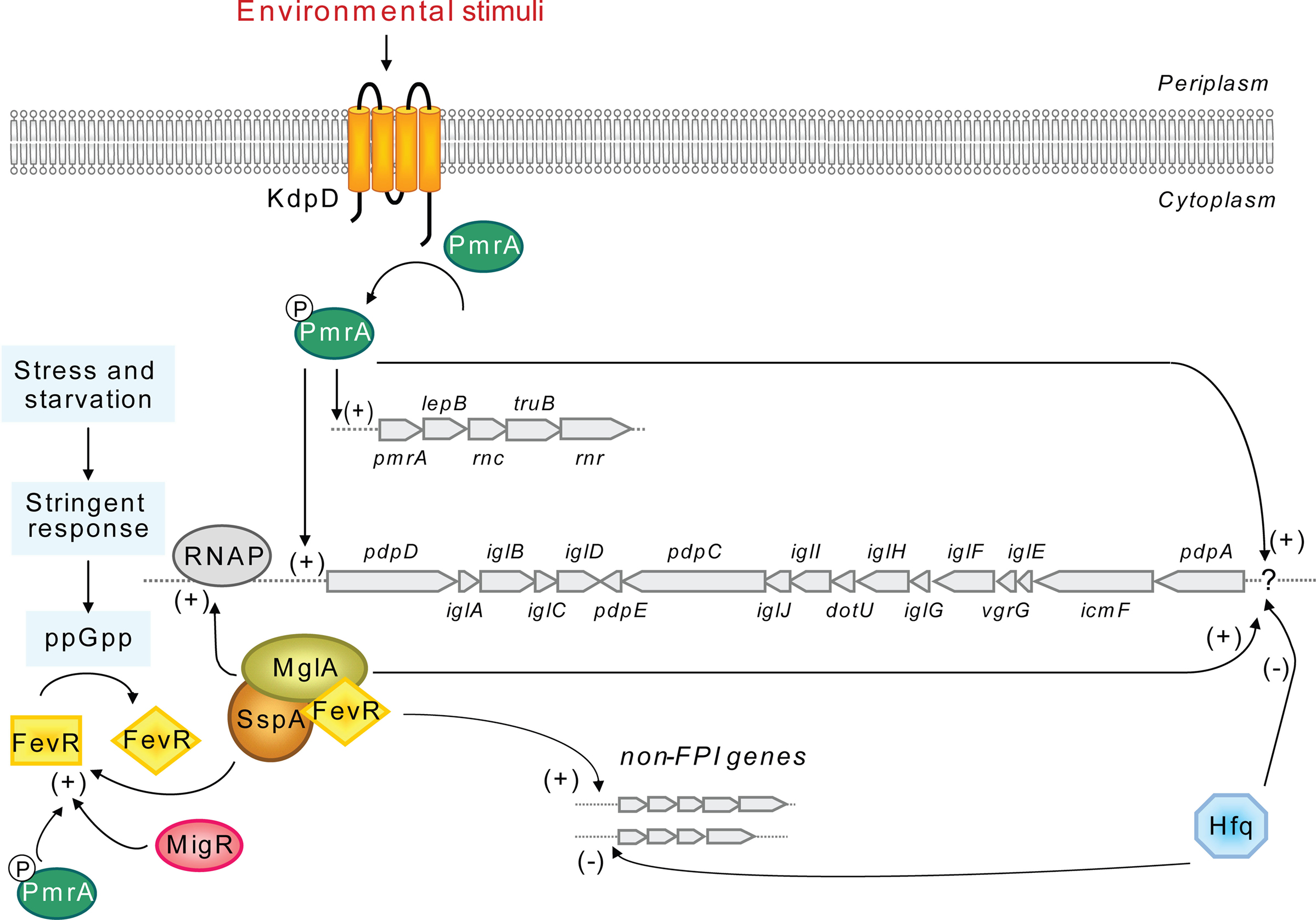
Figure 4. A model for the regulation of the FPI locus. Preceding expression of the FPI locus is an unknown environmental stimulus that leads to KdpD-mediated phosphorylation of PmrA, allowing it to bind to promoter regions. The promoter upstream of pdpD has been experimentally confirmed, while the promoter upstream of pdpA (?) is hypothetical. PmrA binding may recruit the MglA/SspA/FevR complex, which directly interacts with the RNA polymerase (RNAP) to initiate transcription. For FevR to efficiently bind to MglA/SspA, it first needs to be activated by the alarmone ppGpp of the stringent response, while fevR expression requires PmrA, MglA, SspA as well as MigR. PmrA and MglA/SspA/FevR also regulate many additional genes outside of the FPI cluster, mainly in a positive manner, however their target genes appear to be different. At an unknown signal, the Hfq repressor specifically inhibits the expression of the putative pdpA to pdpE operon, presumably by binding to the predicted promoter upstream of pdpA. Hfq also regulates expression of genes outside of the FPI, mainly by acting as a repressor.
Macrophage growth locus (MglA) and SspA are members of the stringent starvation protein A (SspA) family (Baron and Nano, 1998) and interact to form heterodimers that bind to the RNA polymerase, inducing the transcription of numerous genes, including fevR (Brotcke et al., 2006; Charity et al., 2007). FevR (Francisella effector of virulence regulation) has been shown to physically interact with MglA/SspA in a manner facilitated by the alarmone ppGpp, although there are conflicting data concerning this interaction (Brotcke and Monack, 2008; Charity et al., 2009). FevR positively autoregulates its own expression (Charity et al., 2009) and exhibits some homology to the helix-turn-helix region of the MerR family of transcriptional activators, suggesting that it may bind to target DNA (Brotcke and Monack, 2008). MigR (Macrophage intracellular growth regulator) may exert its positive effect on FPI gene expression indirectly through FevR, since it has been shown to be required for fevR expression, in contrast to that of mglA, sspA, or pmrA (Buchan et al., 2009). Moreover, fevR expression also requires the presence of MglA, SspA as well as PmrA (Brotcke et al., 2006; Charity et al., 2007; Mohapatra et al., 2007; Figure 4).
Importantly, current data suggest a connection between MglA/SspA/FevR and a two-component regulatory system consisting of PmrA/KdpD (Bell et al., 2010). PmrA, which positively regulates its own transcription and that of the FPI, was originally assumed to be an orphan response regulator (Mohapatra et al., 2007; Sammons-Jackson et al., 2008). However, Bell et al. (2010) recently demonstrated that PmrA, upon phosphorylation by the histidine kinase KdpD, efficiently binds to regulated promoters of the pmrA operon and FPI-encoded pdpD. Moreover, using coimmunoprecipitation experiments it was revealed that PmrA, MglA, and SspA are part of the same protein complex (Bell et al., 2010). Thus, although the sequence of protein binding to the regulatory complex is not clear, phosphorylated PmrA may bind to FPI gene promoters recruiting free or RNA polymerase-bound MglA, SspA and FevR, to initiate FPI gene transcription (Figure 4).
Finally, Hfq differs from all the other FPI regulators identified so far in that it seems to act primarily as a repressor of gene expression, both for genes located within the FPI as well as genes located outside (Meibom et al., 2009). Moreover, Meibom et al. (2009) were able to show that only a subset of FPI genes (pdpA to iglJ) are negatively regulated by Hfq (Figure 4), leading the authors to speculate that there is differential regulation of the two putative divergent operons in the FPI (pdpD to iglD and pdpA to pdpE). The G + C content of these two regions are significantly different from one another: 30.6% (pdpD to iglD) vs 26.6% (pdpA to pdpE), compared to an average of 33% for the Francisella chromosome (Nano and Schmerk, 2007), which supports the notion of two different origins for these regions. Nevertheless, this operon organization must still be shown experimentally and putative promoter(s), in addition to that located within the intergenic region of pdpD and anmK (Bell et al., 2010), identified. Importantly, the intergenic regions between pdpD and iglA (80 nt) and iglJ and pdpC (39 nt) are also, in theory, large enough to harbor promoters, suggesting the possibility of multiple overlapping promoters within the FPI (Nano and Schmerk, 2007).
Clearly, much more work is needed in order to understand the intricate regulatory network behind Francisella FPI gene expression, especially how the many regulators are able to coordinate this process. The fact that these also regulate genes outside of the FPI suggests there is cross-talk between T6S and other virulence determinants, which adds further complexity to the situation. Defining how these networks are coordinated and identifying the environmental signals that induce FPI expression and T6S may also help to identify the elusive effector proteins secreted by this system. All together, this will shed light on the critical role of the FPI in Francisella pathogenesis.
T6S – What can we Learn about the F. Tularensis System from other Pathogens?
To date, there is limited information about the functions of the putative T6SS encoded by the Francisella FPI gene cluster. However, as a phenomenon highly related to pathogenesis, bacterial secretion has received great interest from the scientific society. Today several comprehensive studies on T6S in various pathogens exist, which have contributed to the understanding of these complex secretion systems. An overview is presented below.
Type VI Secretion System Gene Clusters are Ubiquitous Among Gram-Negative Bacteria
When first discovered in V. cholerae, the T6SS gene clusters were denoted IAHP (IcmF associated homologous protein), since they include a gene with homology to icmF of the Legionella pneumophila Dot/Icm type IV secretion system (T4SS) (Das and Chaudhuri, 2003). Since then, similar clusters encompassing 15–25 ORFs have been identified by in silico methods in approximately 100 bacterial species (∼ ¼ of all bacterial genomes sequenced to date), some encoding more than one system (Bingle et al., 2008; Shrivastava and Mande, 2008; Boyer et al., 2009). Subsequently, the importance of T6SSs for virulence has been established for pathogens like V. cholerae, B. mallei, S. typhimurium, Edwardsiella tarda, Aeromonas hydrophila, enteroaggregative E. coli, and P. aeruginosa (Folkesson et al., 2002; Dudley et al., 2006; Mougous et al., 2006; Pukatzki et al., 2006; Schell et al., 2007; Zheng and Leung, 2007; Suarez et al., 2008). Importantly, T6SSs are not restricted to pathogens, since they are also found in symbiotic bacteria as well as bacteria with no known association with eukaryotes, the latter suggesting that they could be used by bacteria to adapt to environmental conditions and/or acquire food nutrients (Bingle et al., 2008; Shrivastava and Mande, 2008; Boyer et al., 2009).
There is large heterogeneity among T6SS gene clusters, which are suggested to form four to five major phylogenetic groups (Bingle et al., 2008; Boyer et al., 2009). However, the phylogenetic analysis by Bingle et al. (2008), based exclusively on IglA (DUF770) and IglB (DUF877) sequences, does not include the FPI in any of these groups, instead it is suggested to form an outgroup, indicating that it is unique from all other T6SSs. This is further accentuated by the fact that this is the only bioinformatic approach that actually identifies the FPI as a putative T6SS cluster. The current understanding of T6SSs reveals that they are related to both bacterial T4SSs and the cell-puncturing devices utilized by bacteriophages for DNA delivery (below).
Core Components and Substrates of T6SSs
T6SSs are, like T4SSs, complex multi-subunit machineries that can deliver macromolecules from the bacterial cytoplasm and into a eukaryotic host cell, traversing both bacterial and eukaryotic membranes. Such machineries include structural components of the secreton, secreted effector proteins, energizers of the secretion process as well as regulators that control the expression of these components. Most gene products of T6SS gene clusters are likely to be structural components, since when mutated, T6S is abolished. For example, in E. tarda 13 out of 16 genes are required for functional secretion, however their specific roles are largely unknown (Dudley et al., 2006; Schell et al., 2007; Zheng and Leung, 2007; Suarez et al., 2008; Wu et al., 2008; Pukatzki et al., 2009).
Due to the heterogeneity between the T6SS gene clusters, and the fact that empiric evidence exist from only a limited number of bacteria, it is not a trivial task to define the core T6SS components. However, most systems encode homologs to V. cholerae IcmF, DotU, ClpV, VipA, VipB, VgrG, and Hcp proteins (Boyer et al., 2009).
While contributing to stabilization of the L. pneumophila T4SS, but not being essential for its function (Sexton et al., 2004), IcmF–DotU homologs are absolutely required for functional T6S. These membrane proteins have been shown to interact and are found in all phylogenetic groups of T6SSs, suggesting that they are essential components of the secretion apparatus (Zheng and Leung, 2007; Boyer et al., 2009; Ma et al., 2009). In Francisella, the suggested IcmF homolog PdpB has been shown to be essential for functional T6S of IglI (Barker et al., 2009).
Another defining trait is a bicistronic locus encoding homologs of VipA and VipB (denoted IglA and IglB in Francisella). These are only found in T6SS clusters and are suggested to form a functional pair, since they have been shown to interact in a number of bacterial species including F. tularensis and to depend on each other for stability (de Bruin et al., 2007; Bröms et al., 2009; Aubert et al., 2010). Interestingly, VipA–VipB of V. cholerae and the corresponding proteins BcsLB–BcsKC from B. cenocepacia assemble into tubular structures suggested to span the bacterial membranes (Bönemann et al., 2009, 2010; Aubert et al., 2010). Whether this also occurs for Francisella IglA and IglB is not known, especially since it lacks a ClpV homolog, with which the V. cholerae and B. cenocepacia homologs have been shown to interact. ClpV is a ClpB-like AAA + family adenosine triphosphatase (ATPase) which is required for secretion of T6SS substrates. It forms a hexameric complex at the inner membrane, and requires an intact ATPase domain to support secretion suggesting that it is the main energizer of T6S (Schlieker et al., 2005; Mougous et al., 2006; Zheng and Leung, 2007; Bönemann et al., 2009). Intriguingly, ClpV is required for assembly of BcsLB–BcsKC tubules and for disassembly of VipA–VipB tubules. The latter is hypothesized to enable transport of VipA–VipB across the inner membrane before reassembling into the tubular structure. Further, the VipA–VipB tubule structurally resembles the T4 bacteriophage tail sheath. This sheath forms an outer layer to the rigid internal tail tube that, when the sheath is contracted, penetrates the outer bacterial membrane and intermembrane peptidoglycan (Kostyuchenko et al., 2005; Leiman et al., 2009). It is conceivable that the mechanism for T6S occurs in a similar manner, since also other T6SS components bear resemblance to phage proteins (below).
The two most well studied T6SS components are VgrG (valine–glycine repeat protein G) and Hcp (hemolysin co-regulated protein). Principally, a functional T6SS is defined by its ability to export these proteins into the extracellular milieu and they represent a class of secreted substrates that also are structural components of the secretion machinery. Hcp, which is the major secreted protein of T6SSs, forms hexameric donut-shaped rings that stack on top of each other to form tubules. These have large inner diameters, forming a conduit for delivery of macromolecules, and show great resemblance to phage tail tube structures (Mougous et al., 2006; Ballister et al., 2008; Leiman et al., 2009; Pell et al., 2009). VgrG contains two domains that structurally resemble the bacteriophage T4 gp27 and gp5 proteins (Figure 5). These assemble into (gp27)3–(gp5)3 needle complexes where the gp5 trimer forms the needle that pierces the bacterial membrane. Presumably, a VgrG trimer forms a similar structure with the ability to puncture both bacterial and eukaryotic membranes (Dudley et al., 2006; Mougous et al., 2006; Pukatzki et al., 2006, 2007, 2009; Schell et al., 2007; Zheng and Leung, 2007; Suarez et al., 2008; Wu et al., 2008; Chow and Mazmanian, 2010).
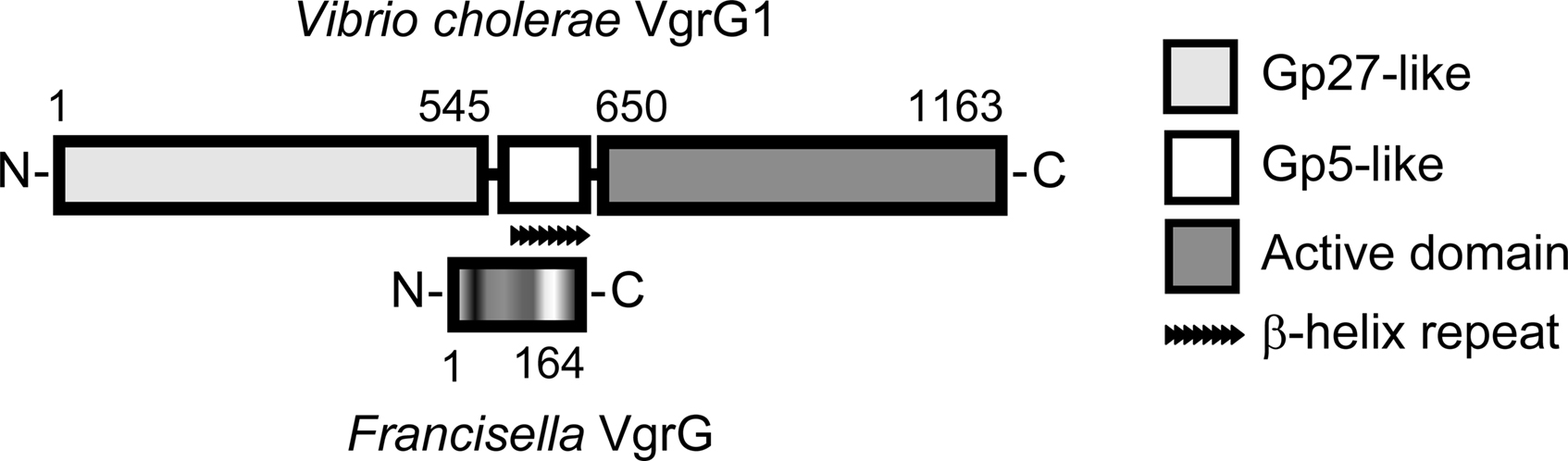
Figure 5. A comparison of V. cholerae VgrG1 and Francisella VgrG. VgrG1 schematics are as presented by Pukatzki et al. (2007). The gp27- and gp5-like domains are homologous to bacteriophage T4 tail-spike complexes utilized to pierce bacterial membranes. VgrG1 is an evolved VgrG, with a C-terminal extension encoding an actin cross-linking domain. The significantly smaller VgrG of Francisella lacks a C-terminal active domain and aligns to the central parts of VgrG1, showing homology to the predicted cell-puncturing parts of this protein.
Apart from the needle complex domains, some evolved VgrGs have C-terminal functional extensions that upon delivery into the host cell can interfere with cellular functions, e.g., the actin cross-linking domain of V. cholerae VgrG1 (Pukatzki et al., 2007). Besides Hcp and VgrG, there is a small number of T6SS exported substrates that are not themselves required for functional secretion and hence are more similar to the “classical” effectors of T3SSs and T4SSs, e.g., EvpP of E. tarda (Zheng and Leung, 2007; Pukatzki et al., 2009).
A Hypothetical Model for Type VI Secretion
Based on the current knowledge of T6S, and guided by the structural resemblance to phage tail-spike organelles, models of how the system functions have been proposed (Filloux, 2009; Pukatzki et al., 2009; Bönemann et al., 2010). A set of core components are suggested to form a pore complex in the bacterial plasma membrane, through which Hcp and VgrG are delivered to the periplasm. Here, Hcp assembles into a tubular structure, capped by the VgrG trimer and sheathed by the VipA–VipB tubule. Upon elongation of the Hcp tubule and contraction of the VipA–VipB sheath, VgrG pushes through the bacterial membrane. Depending on the localization of the bacterium, the VgrG tip may also puncture either the outer or the vesicular membrane of the host cell, allowing the delivery of the VgrG effector domain in the target cell cytosol. It is not known whether the active domain remains attached to VgrG or if it is cleaved off before it reaches its final destination. Further, the VgrG tip is too narrow to allow secretion and hence VgrG detaches from the Hcp tubule to allow Hcp and other components with effector functions to be delivered into the host cell cytosol. The hexameric ATPase ClpV has been suggested to localize to the membrane pore complex where it energizes the T6S process, controlling disassembly/assembly and export of T6S components.
Evidence of a Francisella Pathogenicity Island Encoded Secretion System
As discussed above, many of the components of the FPI are required for pathogenicity-related phenotypes such as phagosome escape, intracellular replication, cytopathogenicity and virulence in animal models (summarized in Table 2). Although information concerning the specific roles these proteins play in Francisella pathogenicity is scarce, they are likely to form part of a T6SS or a T6SS-like system (de Bruin et al., 2007; Bingle et al., 2008; Ludu et al., 2008; Barker et al., 2009).
In Silico Analysis of the Pathogenicity Island Gene Products
A bioinformatic analysis of the FPI gene products was employed to predict the presence of secretion signals and transmembrane regions, protein localization, homology to known proteins and functional domains. However, this analysis (summarized in Table 3) only gives so much away. Most FPI components reveal no homology to any known proteins, display no functional domains and the predicted localization is primarily “unknown.” Nevertheless, five of them belong to different COGs (clusters of orthologous groups) (Tatusov et al., 2000; Table 3). COG2377, represented by AnmK, includes members of the anhydro-N-acetylmuramic acid kinase family, which contribute to the fitness of pathogenic and non-pathogenic bacteria alike but is unrelated to T6S. However, the other four COGs do indeed define proteins found in T6SS clusters, i.e., the canonical functional pairs IglA–IglB (COG3516, COG3517) and PdpB/IcmF–DotU (COG3523, COG3455) (Figure 1; Table 3). Moreover, both PdpB/IcmF and DotU display predicted transmembrane domains, which is logical if they, like the homologous protein pair of the Legionella T4SS, localize to the inner membrane. While the predicted localization for DotU is “unknown,” PdpB is predicted to localize to the outer membrane. However, fractionation experiments have revealed that PdpB is found exclusively in the bacterial inner membrane, a localization that better agrees with its homology to IcmF (Bröms et al., unpublished; Ludu et al., 2008). Together with the requirement of PdpB/IcmF and DotU for Francisella virulence (Table 2), this suggests that the functions of these core components, whatever they may be, are likely to be conserved in Francisella.
Two FPI members, PdpE and IglE, are predicted to possess Sec secretion signals (Table 3). Both localize to the bacterial outer membrane (Bröms et al., unpublished; Rodriguez, 2010) and in the case of IglE, this localization depends on the presence of a lipid attachment site suggesting that IglE is a lipoprotein (Rodriguez, 2010).
Protein alignment studies suggest that the FPI encodes a VgrG homolog, which is significantly smaller than any known VgrG. A comparison to VgrG1 from V. cholerae reveals that Francisella VgrG aligns to the central part of this protein, corresponding to the most C-terminal part of protein gp27 and to the greater part of the membrane puncturing gp5 protein of bacteriophage T4 (Figure 5). Needless to say, the diminutive FPI VgrG has no C-terminal active domain. However, it cannot be ruled out that it still possesses an effector function, since VgrG export into the host cell cytosol was recently demonstrated (Pukatzki et al., 2007; Ludu et al., 2008; Barker et al., 2009).
It has also been suggested that PdpE encodes an Hcp-like protein (Barker et al., 2009), but this homology is faint and there has been no mechanistic evidence of PdpE executing similar functions as Hcp. In addition, Hcp-like proteins are often encoded by a monocistronic operon separated from the T6SS gene cluster. In the same study, IglF was proposed to be equivalent to ClpV, but it lacks the key ATPase domain of ClpV proteins. Overall, there is some homology between FPI members and T6SS gene clusters, but mechanistic data supporting the notion that the FPI truly encodes a T6SS is scarce.
IglA–IglB – Formation of a Transmembrane Tubular Structure?
The FPI components with the greatest homology to T6SS proteins are IglA and IglB, which have been shown to interact and also depend on each other for protein stability (de Bruin et al., 2007; Bröms et al., 2009). This interaction is vital for virulence and relies on a conserved α-helix within IglA (Bröms et al., 2009). Furthermore, interactions between homologous components of pathogens like V. cholerae, P. aeruginosa, Y. pseudotuberculosis, and uropathogenic E. coli, also depend on the same α-helical domain suggesting that the mechanism of interaction is conserved (Bröms et al., 2009). Hence, it is conceivable that IglA–IglB, similar to V. cholerae VipA–VipB and B. cenocepacia BcsLB–BcsKC, form a tubular transmembrane structure that encases the internal components of a secretion conduit (Bönemann et al., 2009; Bröms et al., 2009; Aubert et al., 2010). There is, however, no existing evidence of IglA–IglB multimerization beyond dimers, and they primarily localize to the bacterial cytosol (Table 3), where they have been suggested to function as a chaperone complex (de Bruin et al., 2007). Nevertheless, in support of the notion that IglA and IglB may form a membrane spanning structure are reports of surface location of these components (Melillo et al., 2006; Ludu et al., 2008).
IglC – A Unique Effector Protein?
A comparative study of protein expression in Francisella grown in liquid culture or in J774 cells revealed significant induction of a 23-kDa protein, later named IglC, during intracellular growth (Golovliov et al., 1997; Gray et al., 2002). IglC has no homology to any characterized bacterial proteins (Table 3), suggesting that it may be a novel virulence factor. Furthermore, it possesses no recognizable functional motifs but adopts a unique beta-sandwich structure, with weak structural similarity to gp27, a cell-puncturing device component of bacteriophage T4 (Sun et al., 2007).
Although predominantly cytoplasmic (Golovliov et al., 1997), a small fraction of IglC localizes to the bacterial surface as determined by a biotinylation assay (Ludu et al., 2008). This led the authors to speculate that IglC could play a role in a secretion channel in the outer membrane of Francisella, or alternatively, in forming a pore in the host cell membrane. The biological relevance of this observation is, however, not easily assessed since IglC surface localization was demonstrated upon overexpression of PdpD (Ludu et al., 2008; Table 3). Nonetheless, IglC-export did not occur in F. novicida strains mutated for the T6SS core components pdpB, dotU, iglA, or iglB, suggesting that it was T6SS dependent. This was in contrast to PdpD-mediated export of IglA and IglB, which occurred also in the absence of pdpB or dotU (Ludu et al., 2008; Table 3). Still, if IglC depends on the outer membrane protein PdpD for export, and this export is an important feature of T6S, it is difficult to comprehend that a F. novicida pdpD mutant is able to grow as efficiently as the wild-type strain in all of 11 eukaryotic cell lines tested (Ludu et al., 2008).
Secretion of VgrG and IglI in Macrophages
The first evidence of translocation of FPI proteins was provided in a recent study by Barker et al. (2009) who demonstrated delivery of F. novicida VgrG and IglI, the latter lacking homology to known bacterial proteins, into the cytosol of infected J774 macrophages. VgrG and IglI export was demonstrated by several means: translocation of C-terminal CyaA fusions was observed based on the accumulation of cAMP in the host cell cytosol, and N-terminally FLAG-tagged proteins were visualized using immunofluorescence microscopy. Moreover, the latter proteins could also be detected in the culture supernatants of bacteria grown in broth (Barker et al., 2009).
Intriguingly, in all assays, VgrG secretion occurs in an FPI-independent manner, as seen when expressed in a ΔFPI F. novicida strain. In contrast, export of IglI within macrophages depends on the FPI and does not occur in ΔvgrG, ΔicmF, or ΔFPI backgrounds. However, IglI–FLAG is efficiently secreted to the extracellular medium of broth-grown ΔvgrG and ΔFPI mutants. Since neither VgrG nor IglI is secreted when expressed in trans in E. coli DH5α, this suggests that they can exit via an FPI-independent but Francisella-specific export mechanism during growth in liquid culture (Barker et al., 2009). It would be interesting to investigate if the rudimentary gene cluster (FTN_0040–FTN_0052), which share homology with core components of the FPI (as described above), contributes to this export. Immunofluorescence staining of VgrG and IglI during infection shows that VgrG, when expressed in a ΔvgrG background, is released into the host cell cytosol forming a plume extending from the bacterium (Barker et al., 2009). In contrast, when secreted from an ΔFPI mutant, VgrG is localized more closely to the bacterium. Similarly, staining for IglI–FLAG in ΔiglI, ΔvgrG, or ΔFPI backgrounds also reveals a close association between the protein and the bacteria. These data could be interpreted to suggest that VgrG is truly secreted while IglI is primarily exported to the bacterial surface. Combined with the data from Ludu et al. (2008) and Melillo et al. (2006) this may imply that the FPI-encoded secretion system exports proteins to form a surface located complex including IglA, IglB, IglC, IglI, and VgrG (Barker et al., 2009).
Does the FPI Encode a True T6SS?
As discussed above, there is increasing evidence that Francisella does encode a T6SS-like system that may export FPI proteins such as IglI but this system truly appears to be unique from all other systems described so far. First of all, it is evolutionary distinct (Bingle et al., 2008), and only 3 out of the 17 FPI proteins share significant homology to T6SS core components, i.e., IglA, IglB, and DotU (Nano and Schmerk, 2007; Boyer et al., 2009). So far, no convincing homolog of ClpV, the AAA + ATPase and putative energizer of T6SSs (Mougous et al., 2006), has been identified in the genome, although Barker et al suggested that FTT1348 (IglF) may be a candidate (Barker et al., 2009). Moreover, while a protein with a short IcmF-related portion (PdpB) and a potential VgrG homolog have been recognized in the FPI (Nano and Schmerk, 2007; Barker et al., 2009; Boyer et al., 2009), the Walker A box commonly present in IcmF homologs is missing from F. tularensis PdpB (Zheng and Leung, 2007; Barker et al., 2009) and the F. tularensis VgrG homolog is much smaller than VgrG homologs from other species, and it also lacks the C-terminal extensions found in, e.g., the V. cholerae VgrG-1 and VgrG-3 proteins (Pukatzki et al., 2007; Barker et al., 2009). Altogether, these differences would suggest that the Francisella FPI system is clearly different compared to other T6SS. Further supporting this notion is the observation that F. tularensis VgrG, in contrast to all VgrG proteins reported so far, is exported into the extracellular milieu in an FPI-independent fashion (Barker et al., 2009), suggesting a novel mechanism of export. While IglI-translocation into host cells is dependent on FPI core components, it can also be secreted via an FPI-independent mechanism during in vitro growth.
Together, these observations raise the question whether the FPI actually encodes a true T6SS or just a system implicated in virulence that shares some components with T6SSs and T4SSs. Much more work is needed to distinguish between these possibilities.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The work was funded in part by grant support from the Swedish Medical Research Council (K2009-5026) and the Umeå Biotech Incubator, Umeå, Sweden. The work was performed in part at the Umeå Centre for Microbial Research (UCMR).
References
Abplanalp, A. L., Morris, I. R., Parida, B. K., Teale, J. M., and Berton, M. T. (2009). TLR-dependent control of Francisella tularensis infection and host inflammatory responses. PLoS One 4, e7920. doi: 10.1371/journal.pone.0007920.
åhlund, M. K., Ryden, P., Sjöstedt, A., and Stöven, S. (2010). Directed screen of Francisella novicida virulence determinants using Drosophila melanogaster. Infect. Immun. 78, 3118–3128.
Al-Khodor, S., and Abu Kwaik, Y. (2010). Triggering Ras signalling by intracellular Francisella tularensis through recruitment of PKCalpha and betaI to the SOS2/GrB2 complex is essential for bacterial proliferation in the cytosol. Cell Microbiol. 12, 1604–1621.
Anthony, L. S., Ghadirian, E., Nestel, F. P., and Kongshavn, P. A. (1989). The requirement for gamma interferon in resistance of mice to experimental tularemia. Microb. Pathog. 7, 421–428.
Asare, R., and Abu Kwaik, Y. (2010). Molecular complexity orchestrates modulation of phagosome biogenesis and escape to the cytosol of macrophages by Francisella tularensis. Environ. Microbiol. 12, 2559–2586.
Aubert, D., Macdonald, D. K., and Valvano, M. A. (2010). BcsKC is an essential protein for the Type VI secretion system activity in Burkholderia cenocepacia that forms an outer membrane complex with BcsLB. J. Biol. Chem. 285, 35988–35998.
Ballister, E. R., Lai, A. H., Zuckermann, R. N., Cheng, Y., and Mougous, J. D. (2008). In vitro self-assembly of tailorable nanotubes from a simple protein building block. Proc. Natl. Acad. Sci. U.S.A. 105, 3733–3738.
Barbieri, M. A., Kohn, A. D., Roth, R. A., and Stahl, P. D. (1998). Protein kinase B/akt and rab5 mediate Ras activation of endocytosis. J. Biol. Chem. 273, 19367–19370.
Barker, J. R., Chong, A., Wehrly, T. D., Yu, J. J., Rodriguez, S. A., Liu, J., Celli, J., Arulanandam, B. P., and Klose, K. E. (2009). The Francisella tularensis pathogenicity island encodes a secretion system that is required for phagosome escape and virulence. Mol. Microbiol. 74, 1459–1470.
Baron, G. S., and Nano, F. E. (1998). MgIA and MgIB are required for the intramacrophage growth of Francisella novicida. Mol. Microbiol. 29, 247–259.
Bell, B. L., Mohapatra, N. P., and Gunn, J. S. (2010). Regulation of virulence gene transcripts by the Francisella novicida orphan response regulator PmrA: role of phosphorylation and evidence of MglA/SspA interaction. Infect. Immun. 78, 2189–2198.
Bingle, L. E., Bailey, C. M., and Pallen, M. J. (2008). Type VI secretion: a beginner’s guide. Curr. Opin. Microbiol. 11, 3–8.
Bönemann, G., Pietrosiuk, A., Diemand, A., Zentgraf, H., and Mogk, A. (2009). Remodelling of VipA/VipB tubules by ClpV-mediated threading is crucial for type VI protein secretion. EMBO J. 28, 315–325.
Bönemann, G., Pietrosiuk, A., and Mogk, A. (2010). Tubules and donuts: a type VI secretion story. Mol. Microbiol. 76, 815–821.
Bönquist, L., Lindgren, H., Golovliov, I., Guina, T., and Sjöstedt, A. (2008). MglA and Igl proteins contribute to the modulation of Francisella tularensis live vaccine strain-containing phagosomes in murine macrophages. Infect. Immun. 76, 3502–3510.
Bosio, C. M., Bielefeldt-Ohmann, H., and Belisle, J. T. (2007). Active suppression of the pulmonary immune response by Francisella tularensis Schu4. J. Immunol. 178, 4538–4547.
Bosio, C. M., and Dow, S. W. (2005). Francisella tularensis induces aberrant activation of pulmonary dendritic cells. J. Immunol. 175, 6792–6801.
Boyer, F., Fichant, G., Berthod, J., Vandenbrouck, Y., and Attree, I. (2009). Dissecting the bacterial type VI secretion system by a genome wide in silico analysis: what can be learned from available microbial genomic resources? BMC Genomics 10, 104.doi: 10.1186/1471-2164-10-104.
Bröms, J. E., Lavander, M., and Sjöstedt, A. (2009). A conserved α-helix essential for a type VI secretion-like system of Francisella tularensis. J. Bacteriol. 6, 6.
Brotcke, A., and Monack, D. M. (2008). Identification of fevR, a novel regulator of virulence gene expression in Francisella novicida. Infect. Immun. 76, 3473–3480.
Brotcke, A., Weiss, D. S., Kim, C. C., Chain, P., Malfatti, S., Garcia, E., and Monack, D. M. (2006). Identification of MglA-regulated genes reveals novel virulence factors in Francisella tularensis. Infect. Immun. 74, 6642–6655.
Buchan, B. W., McCaffrey, R. L., Lindemann, S. R., Allen, L. A., and Jones, B. D. (2009). Identification of migR, a regulatory element of the Francisella tularensis live vaccine strain iglABCD virulence operon required for normal replication and trafficking in macrophages. Infect. Immun. 77, 2517–2529.
Butchar, J. P., Cremer, T. J., Clay, C. D., Gavrilin, M. A., Wewers, M. D., Marsh, C. B., Schlesinger, L. S., and Tridandapani, S. (2008). Microarray analysis of human monocytes infected with Francisella tularensis identifies new targets of host response subversion. PLoS One 3, e2924. doi: 10.1371/journal.pone.0002924.
Castang, S., McManus, H. R., Turner, K. H., and Dove, S. L. (2008). H-NS family members function coordinately in an opportunistic pathogen. Proc. Natl. Acad. Sci. U.S.A. 105, 18947–18952.
Champion, M. D., Zeng, Q., Nix, E. B., Nano, F. E., Keim, P., Kodira, C. D., Borowsky, M., Young, S., Koehrsen, M., Engels, R., Pearson, M., Howarth, C., Larson, L., White, J., Alvarado, L., Forsman, M., Bearden, S. W., Sjöstedt, A., Titball, R., Michell, S. L., Birren, B., and Galagan, J. (2009). Comparative genomic characterization of Francisella tularensis strains belonging to low and high virulence subspecies. PLoS Pathog. 5, e1000459. doi: 10.1371/journal.ppat.1000459.
Charity, J. C., Blalock, L. T., Costante-Hamm, M. M., Kasper, D. L., and Dove, S. L. (2009). Small molecule control of virulence gene expression in Francisella tularensis. PLoS Pathog. 5, e1000641. doi: 10.1371/journal.ppat.1000641.
Charity, J. C., Costante-Hamm, M. M., Balon, E. L., Boyd, D. H., Rubin, E. J., and Dove, S. L. (2007). Twin RNA polymerase-associated proteins control virulence gene expression in Francisella tularensis. PLoS Pathog. 3, e84. doi: 10.1371/journal.ppat.0030084.
Chen, Q., Powell, D. W., Rane, M. J., Singh, S., Butt, W., Klein, J. B., and McLeish, K. R. (2003). Akt phosphorylates p47phox and mediates respiratory burst activity in human neutrophils. J. Immunol. 170, 5302–5308.
Chong, A., Wehrly, T. D., Nair, V., Fischer, E. R., Barker, J. R., Klose, K. E., and Celli, J. (2008). The early phagosomal stage of Francisella tularensis determines optimal phagosomal escape and Francisella pathogenicity island protein expression. Infect. Immun. 76, 5488–5499.
Chow, J., and Mazmanian, S. K. (2010). A pathobiont of the microbiota balances host colonization and intestinal inflammation. Cell Host Microbe 7, 265–276.
Chugani, S., and Greenberg, E. P. (2007). The influence of human respiratory epithelia on Pseudomonas aeruginosa gene expression. Microb. Pathog. 42, 29–35.
Clemens, D. L., Lee, B. Y., and Horwitz, M. A. (2004). Virulent and avirulent strains of Francisella tularensis prevent acidification and maturation of their phagosomes and escape into the cytoplasm in human macrophages. Infect. Immun. 72, 3204–3217.
Clemens, D. L., Lee, B. Y., and Horwitz, M. A. (2005). Francisella tularensis enters macrophages via a novel process involving pseudopod loops. Infect. Immun. 73, 5892–5902.
Clemens, D. L., Lee, B. Y., and Horwitz, M. A. (2009). Francisella tularensis phagosomal escape does not require acidification of the phagosome. Infect. Immun. 77, 1757–1773.
Cole, L. E., Elkins, K. L., Michalek, S. M., Qureshi, N., Eaton, L. J., Rallabhandi, P., Cuesta, N., and Vogel, S. N. (2006). Immunologic consequences of Francisella tularensis live vaccine strain infection: role of the innate immune response in infection and immunity. J. Immunol. 176, 6888–6899.
Cole, L. E., Santiago, A., Barry, E., Kang, T. J., Shirey, K. A., Roberts, Z. J., Elkins, K. L., Cross, A. S., and Vogel, S. N. (2008). Macrophage proinflammatory response to Francisella tularensis live vaccine strain requires coordination of multiple signaling pathways. J. Immunol. 180, 6885–6891.
Cole, L. E., Shirey, K. A., Barry, E., Santiago, A., Rallabhandi, P., Elkins, K. L., Puche, A. C., Michalek, S. M., and Vogel, S. N. (2007). Toll-like receptor 2-mediated signaling requirements for Francisella tularensis live vaccine strain infection of murine macrophages. Infect. Immun. 75, 4127–4137.
Crane, D. D., Warner, S. L., and Bosio, C. M. (2009). A novel role for plasmin-mediated degradation of opsonizing antibody in the evasion of host immunity by virulent, but not attenuated, Francisella tularensis. J. Immunol. 183, 4593–4600.
Cremer, T. J., Ravneberg, D. H., Clay, C. D., Piper-Hunter, M. G., Marsh, C. B., Elton, T. S., Gunn, J. S., Amer, A., Kanneganti, T. D., Schlesinger, L. S., Butchar, J. P., and Tridandapani, S. (2009). MiR-155 induction by F. novicida but not the virulent F. tularensis results in SHIP down-regulation and enhanced pro-inflammatory cytokine response. PLoS One 4, e8508. doi: 10.1371/journal.pone.0008508.
Das, S., Chakrabortty, A., Banerjee, R., Roychoudhury, S., and Chaudhuri, K. (2000). Comparison of global transcription responses allows identification of Vibrio cholerae genes differentially expressed following infection. FEMS Microbiol. Lett. 190, 87–91.
Das, S., and Chaudhuri, K. (2003). Identification of a unique IAHP (IcmF associated homologous proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis. In Silico Biol. 3, 287–300.
de Bruin, O. M., Ludu, J. S., and Nano, F. E. (2007). The Francisella pathogenicity island protein IglA localizes to the bacterial cytoplasm and is needed for intracellular growth. BMC Microbiol. 7, 1. doi: 10.1186/1471-2180-7-1.
Dean, R. E., Ireland, P. M., Jordan, J. E., Titball, R. W., and Oyston, P. C. (2009). RelA regulates virulence and intracellular survival of Francisella novicida. Microbiology 155, 4104–4113.
Deng, K., Blick, R. J., Liu, W., and Hansen, E. J. (2006). Identification of Francisella tularensis genes affected by iron limitation. Infect. Immun. 74, 4224–4236.
Dennis, D. T., Inglesby, T. V., Henderson, D. A., Bartlett, J. G., Ascher, M. S., Eitzen, E., Fine, A. D., Friedlander, A. M., Hauer, J., Layton, M., Lillibridge, S. R., McDade, J. E., Osterholm, M. T., O’Toole, T., Parker, G., Perl, T. M., Russell, P. K., and Tonat, K. (2001). Tularemia as a biological weapon: medical and public health management. JAMA 285, 2763–2773.
Deziel, E., Gopalan, S., Tampakaki, A. P., Lepine, F., Padfield, K. E., Saucier, M., Xiao, G., and Rahme, L. G. (2005). The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-L-homoserine lactones. Mol. Microbiol. 55, 998–1014.
Dudley, E. G., Thomson, N. R., Parkhill, J., Morin, N. P., and Nataro, J. P. (2006). Proteomic and microarray characterization of the AggR regulon identifies a pheU pathogenicity island in enteroaggregative Escherichia coli. Mol. Microbiol. 61, 1267–1282.
Edwards, J. A., Rockx-Brouwer, D., Nair, V., and Celli, J. (2009). Restricted cytosolic growth of Francisella tularensis subsp. tularensis by IFN-gamma activation of macrophages. Microbiology 156, 327–339.
Fernandes-Alnemri, T., Wu, J., Yu, J. W., Datta, P., Miller, B., Jankowski, W., Rosenberg, S., Zhang, J., and Alnemri, E. S. (2007). The pyroptosome: a supramolecular assembly of ASC dimers mediating inflammatory cell death via caspase-1 activation. Cell Death Differ. 14, 1590–1604.
Fernandes-Alnemri, T., Yu, J. W., Datta, P., Wu, J., and Alnemri, E. S. (2009). AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 458, 509–513.
Fernandes-Alnemri, T., Yu, J. W., Juliana, C., Solorzano, L., Kang, S., Wu, J., Datta, P., McCormick, M., Huang, L., McDermott, E., Eisenlohr, L., Landel, C. P., and Alnemri, E. S. (2010). The AIM2 inflammasome is critical for innate immunity to Francisella tularensis. Nat. Immunol. 11, 385–393.
Folkesson, A., Löfdahl, S., and Normark, S. (2002). The Salmonella enterica subspecies I specific centisome 7 genomic island encodes novel protein families present in bacteria living in close contact with eukaryotic cells. Res. Microbiol. 153, 537–545.
Fortier, A. H., Slayter, M. V., Ziemba, R., Meltzer, M. S., and Nacy, C. A. (1991). Live vaccine strain of Francisella tularensis: infection and immunity in mice. Infect. Immun. 59, 2922–2928.
Gavrilin, M. A., Bouakl, I. J., Knatz, N. L., Duncan, M. D., Hall, M. W., Gunn, J. S., and Wewers, M. D. (2006). Internalization and phagosome escape required for Francisella to induce human monocyte IL-1beta processing and release. Proc. Natl. Acad. Sci. U.S.A. 103, 141–146.
Golovliov, I., Baranov, V., Krocova, Z., Kovarova, H., and Sjöstedt, A. (2003a). An attenuated strain of the facultative intracellular bacterium Francisella tularensis can escape the phagosome of monocytic cells. Infect. Immun. 71, 5940–5950.
Golovliov, I., Sjöstedt, A., Mokrievich, A., and Pavlov, V. (2003b). A method for allelic replacement in Francisella tularensis. FEMS Microbiol. Lett. 222, 273–280.
Golovliov, I., Ericsson, M., Sandström, G., Tärnvik, A., and Sjöstedt, A. (1997). Identification of proteins of Francisella tularensis induced during growth in macrophages and cloning of the gene encoding a prominently induced 23-kilodalton protein. Infect. Immun. 65, 2183–2189.
Gray, C. G., Cowley, S. C., Cheung, K. K., and Nano, F. E. (2002). The identification of five genetic loci of Francisella novicida associated with intracellular growth. FEMS Microbiol. Lett. 215, 53–56.
Guina, T., Radulovic, D., Bahrami, A. J., Bolton, D. L., Rohmer, L., Jones-Isaac, K. A., Chen, J., Gallagher, L. A., Gallis, B., Ryu, S., Taylor, G. K., Brittnacher, M. J., Manoil, C., and Goodlett, D. R. (2007). MglA regulates Francisella tularensis subsp. novicida (Francisella novicida) response to starvation and oxidative stress. J. Bacteriol. 189, 6580–6586.
Henry, T., Brotcke, A., Weiss, D. S., Thompson, L. J., and Monack, D. M. (2007). Type I interferon signaling is required for activation of the inflammasome during Francisella infection. J. Exp. Med. 204, 987–994.
Hoyal, C. R., Gutierrez, A., Young, B. M., Catz, S. D., Lin, J. H., Tsichlis, P. N., and Babior, B. M. (2003). Modulation of p47PHOX activity by site-specific phosphorylation: Akt-dependent activation of the NADPH oxidase. Proc. Natl. Acad. Sci. U.S.A. 100, 5130–5135.
Hrstka, R., Stulik, J., and Vojtesek, B. (2005). The role of MAPK signal pathways during Francisella tularensis LVS infection-induced apoptosis in murine macrophages. Microbes Infect. 7, 619–625.
Ishikawa, T., Rompikuntal, P. K., Lindmark, B., Milton, D. L., and Wai, S. N. (2009). Quorum sensing regulation of the two hcp alleles in Vibrio cholerae O1 strains. PLoS One 4, e6734. doi: 10.1371/ journal.pone.0006734.
Jones, J. W., Kayagaki, N., Broz, P., Henry, T., Newton, K., O’Rourke, K., Chan, S., Dong, J., Qu, Y., Roose-Girma, M., Dixit, V. M., and Monack, D. M. (2010). Absent in melanoma 2 is required for innate immune recognition of Francisella tularensis. Proc. Natl. Acad. Sci. U.S.A. 107, 9771–9776.
Kadzhaev, K., Zingmark, C., Golovliov, I., Bolanowski, M., Shen, H., Conlan, W., and Sjostedt, A. (2009). Identification of genes contributing to the virulence of Francisella tularensis SCHU S4 in a mouse intradermal infection model. PLoS One 4, e5463. doi: 10.1371/journal.pone.0005463.
Karlsson, J., Prior, R. G., Williams, K., Lindler, L., Brown, K. A., Chatwell, N., Hjalmarsson, K., Loman, N., Mack, K. A., Pallen, M., Popek, M., Sandström, G., Sjöstedt, A., Svensson, T., Tamas, I., Andersson, S. G., Wren, B. W., Oyston, P. C., and Titball, R. W. (2000). Sequencing of the Francisella tularensis strain Schu 4 genome reveals the shikimate and purine metabolic pathways, targets for the construction of a rationally attenuated auxotrophic vaccine. Microb. Comp. Genomics 5, 25–39.
Katz, J., Zhang, P., Martin, M., Vogel, S. N., and Michalek, S. M. (2006). Toll-like receptor 2 is required for inflammatory responses to Francisella tularensis LVS. Infect. Immun. 74, 2809–2816.
Kawula, T. H., Hall, J. D., Fuller, J. R., and Craven, R. R. (2004). Use of transposon-transposase complexes to create stable insertion mutant strains of Francisella tularensis LVS. Appl. Environ. Microbiol. 70, 6901–6904.
Keim, P., Johansson, A., and Wagner, D. M. (2007). Molecular epidemiology, evolution, and ecology of Francisella. Ann. N.Y. Acad. Sci. 1105, 30–66.
Kostyuchenko, V. A., Chipman, P. R., Leiman, P. G., Arisaka, F., Mesyanzhinov, V. V., and Rossmann, M. G. (2005). The tail structure of bacteriophage T4 and its mechanism of contraction. Nat. Struct. Mol. Biol. 12, 810–813.
Kraemer, P. S., Mitchell, A., Pelletier, M. R., Gallagher, L. A., Wasnick, M., Rohmer, L., Brittnacher, M. J., Manoil, C., Skerett, S. J., and Salama, N. R. (2009). Genome-wide screen in Francisella novicida for genes required for pulmonary and systemic infection in mice. Infect. Immun. 77, 232–244.
Lai, X. H., Golovliov, I., and Sjöstedt, A. (2004). Expression of IglC is necessary for intracellular growth and induction of apoptosis in murine macrophages by Francisella tularensis. Microb. Pathog. 37, 225–230.
Lai, X. H., and Sjöstedt, A. (2003). Delineation of the molecular mechanisms of Francisella tularensis-induced apoptosis in murine macrophages. Infect. Immun. 71, 4642–4646.
Larsson, P., Elfsmark, D., Svensson, K., Wikström, P., Forsman, M., Brettin, T., Keim, P., and Johansson, A. (2009). Molecular evolutionary consequences of niche restriction in Francisella tularensis, a facultative intracellular pathogen. PLoS Pathog. 5, e1000472. doi: 10.1371/journal.ppat.1000472.
Lauriano, C. M., Barker, J. R., Nano, F. E., Arulanandam, B. P., and Klose, K. E. (2003). Allelic exchange in Francisella tularensis using PCR products. FEMS Microbiol. Lett. 229, 195–202.
Lauriano, C. M., Barker, J. R., Yoon, S. S., Nano, F. E., Arulanandam, B. P., Hassett, D. J., and Klose, K. E. (2004). MglA regulates transcription of virulence factors necessary for Francisella tularensis intraamoebae and intramacrophage survival. Proc. Natl. Acad. Sci. U.S.A. 101, 4246–4249.
Leiman, P. G., Basler, M., Ramagopal, U. A., Bonanno, J. B., Sauder, J. M., Pukatzki, S., Burley, S. K., Almo, S. C., and Mekalanos, J. J. (2009). Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl. Acad. Sci. U.S.A. 106, 4154–4159.
Lenco, J., Hubalek, M., Larsson, P., Fucikova, A., Brychta, M., Macela, A., and Stulik, J. (2007). Proteomics analysis of the Francisella tularensis LVS response to iron restriction: induction of the F. tularensis pathogenicity island proteins IglABC. FEMS Microbiol. Lett. 269, 11–21.
Lenco, J., Pavkova, I., Hubalek, M., and Stulik, J. (2005). Insights into the oxidative stress response in Francisella tularensis LVS and its mutant DeltaiglC1 + 2 by proteomics analysis. FEMS Microbiol. Lett. 246, 47–54.
Lesic, B., Starkey, M., He, J., Hazan, R., and Rahme, L. G. (2009). Quorum sensing differentially regulates Pseudomonas aeruginosa type VI secretion locus I and homologous loci II and III, which are required for pathogenesis. Microbiology 155, 2845–2855.
Li, H., Nookala, S., Bina, X. R., Bina, J. E., and Re, F. (2006). Innate immune response to Francisella tularensis is mediated by TLR2 and caspase-1 activation. J. Leukoc. Biol. 80, 766–773.
Lindgren, H., Golovliov, I., Baranov, V., Ernst, R. K., Telepnev, M., and Sjöstedt, A. (2004). Factors affecting the escape of Francisella tularensis from the phagolysosome. J. Med. Microbiol. 53, 953–958.
Lucchini, S., Rowley, G., Goldberg, M. D., Hurd, D., Harrison, M., and Hinton, J. C. (2006). H-NS mediates the silencing of laterally acquired genes in bacteria. PLoS Pathog. 2, e81. doi: 10.1371/journal.ppat.0020081.
Ludu, J. S., de Bruin, O. M., Duplantis, B. N., Schmerk, C. L., Chou, A. Y., Elkins, K. L., and Nano, F. E. (2008). The Francisella pathogenicity island protein PdpD is required for full virulence and associates with homologues of the type VI secretion system. J. Bacteriol. 190, 4584–4595.
Ma, L. S., Lin, J. S., and Lai, E. M. (2009). An IcmF family protein, ImpLM, is an integral inner membrane protein interacting with ImpKL, and its walker a motif is required for type VI secretion system-mediated Hcp secretion in Agrobacterium tumefaciens. J. Bacteriol. 191, 4316–4329.
Maier, T. M., Casey, M. S., Becker, R. H., Dorsey, C. W., Glass, E. M., Maltsev, N., Zahrt, T. C., and Frank, D. W. (2007). Identification of Francisella tularensis Himar1-based transposon mutants defective for replication in macrophages. Infect. Immun. 75, 5376–5389.
Mariathasan, S., Newton, K., Monack, D. M., Vucic, D., French, D. M., Lee, W. P., Roose-Girma, M., Erickson, S., and Dixit, V. M. (2004). Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430, 213–218.
Mariathasan, S., Weiss, D. S., Newton, K., McBride, J., O’Rourke, K., Roose-Girma, M., Lee, W. P., Weinrauch, Y., Monack, D. M., and Dixit, V. M. (2006). Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440, 228–232.
McCaffrey, R. L., and Allen, L. A. (2006). Francisella tularensis LVS evades killing by human neutrophils via inhibition of the respiratory burst and phagosome escape. J. Leukoc. Biol. 80, 1224–1230.
Meibom, K. L., and Charbit, A. (2009). The unraveling panoply of Francisella tularensis virulence attributes. Curr. Opin. Microbiol. 13, 11–17.
Meibom, K. L., Forslund, A. L., Kuoppa, K., Alkhuder, K., Dubail, I., Dupuis, M., Forsberg, å., and Charbit, A. (2009). Hfq, a novel pleiotropic regulator of virulence-associated genes in Francisella tularensis. Infect. Immun. 77, 1866–1880.
Melillo, A., Sledjeski, D. D., Lipski, S., Wooten, R. M., Basrur, V., and Lafontaine, E. R. (2006). Identification of a Francisella tularensis LVS outer membrane protein that confers adherence to A549 human lung cells. FEMS Microbiol. Lett. 263, 102–108.
Mohapatra, N. P., Soni, S., Bell, B. L., Warren, R., Ernst, R. K., Muszynski, A., Carlson, R. W., and Gunn, J. S. (2007). Identification of an orphan response regulator required for the virulence of Francisella spp. and transcription of pathogenicity island genes. Infect. Immun. 75, 3305–3314.
Mohapatra, N. P., Soni, S., Rajaram, M. V., Dang, P. M., Reilly, T. J., El-Benna, J., Clay, C. D., Schlesinger, L. S., and Gunn, J. S. (2010). Francisella acid phosphatases inactivate the NADPH oxidase in human phagocytes. J. Immunol. 184, 5141–5150.
Mörner, T., and Addison, E. (2001). “Tularemia,” in Infectious Diseases of Wild Animals, 3rd Edn., ed. B. I. Williams (Ames, Iowa: Iowa State University), 303–312.
Mougous, J. D., Cuff, M. E., Raunser, S., Shen, A., Zhou, M., Gifford, C. A., Goodman, A. L., Joachimiak, G., Ordonez, C. L., Lory, S., Walz, T., Joachimiak, A., and Mekalanos, J. J. (2006). A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 312, 1526–1530.
Nano, F. E., and Schmerk, C. (2007). The Francisella pathogenicity island. Ann. N.Y. Acad. Sci. 1105, 122–137.
Nano, F. E., Zhang, N., Cowley, S. C., Klose, K. E., Cheung, K. K., Roberts, M. J., Ludu, J. S., Letendre, G. W., Meierovics, A. I., Stephens, G., and Elkins, K. L. (2004). A Francisella tularensis pathogenicity island required for intramacrophage growth. J. Bacteriol. 186, 6430–6436.
Oyston, P. C., Sjöstedt, A., and Titball, R. W. (2004). Tularaemia: bioterrorism defence renews interest in Francisella tularensis. Nat. Rev. Microbiol. 2, 967–978.
Parsa, K. V., Butchar, J. P., Rajaram, M. V., Cremer, T. J., Gunn, J. S., Schlesinger, L. S., and Tridandapani, S. (2008). Francisella gains a survival advantage within mononuclear phagocytes by suppressing the host IFN-gamma response. Mol. Immunol. 45, 3428–3437.
Parsa, K. V., Ganesan, L. P., Rajaram, M. V., Gavrilin, M. A., Balagopal, A., Mohapatra, N. P., Wewers, M. D., Schlesinger, L. S., Gunn, J. S., and Tridandapani, S. (2006). Macrophage pro-inflammatory response to Francisella novicida infection is regulated by SHIP. PLoS Pathog. 2, e71. doi: 10.1371/journal.ppat.0020071.
Parsons, D. A., and Heffron, F. (2005). sciS, an icmF homolog in Salmonella enterica serovar Typhimurium, limits intracellular replication and decreases virulence. Infect. Immun. 73, 4338–4345.
Pell, L. G., Kanelis, V., Donaldson, L. W., Howell, P. L., and Davidson, A. R. (2009). The phage lambda major tail protein structure reveals a common evolution for long-tailed phages and the type VI bacterial secretion system. Proc. Natl. Acad. Sci. U.S.A. 106, 4160–4165.
Prior, R. G., Klasson, L., Larsson, P., Williams, K., Lindler, L., Sjöstedt, A., Svensson, T., Tamas, I., Wren, B. W., Oyston, P. C., Andersson, S. G., and Titball, R. W. (2001). Preliminary analysis and annotation of the partial genome sequence of Francisella tularensis strain Schu 4. J. Appl. Microbiol. 91, 614–620.
Pukatzki, S., Ma, A. T., Revel, A. T., Sturtevant, D., and Mekalanos, J. J. (2007). Type VI secretion system translocates a phage tail spike-like protein into target cells where it cross-links actin. Proc. Natl. Acad. Sci. U.S.A. 104, 15508–15513.
Pukatzki, S., Ma, A. T., Sturtevant, D., Krastins, B., Sarracino, D., Nelson, W. C., Heidelberg, J. F., and Mekalanos, J. J. (2006). Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl. Acad. Sci. U.S.A. 103, 1528–1533.
Pukatzki, S., McAuley, S. B., and Miyata, S. T. (2009). The type VI secretion system: translocation of effectors and effector-domains. Curr. Opin. Microbiol. 12, 11–17.
Qin, A., and Mann, B. J. (2006). Identification of transposon insertion mutants of Francisella tularensis tularensis strain Schu S4 deficient in intracellular replication in the hepatic cell line HepG2. BMC Microbiol. 6, 69. doi: 10.1186/1471-2180-6-69.
Qin, A., Scott, D. W., Thompson, J. A., and Mann, B. J. (2009). Identification of an essential Francisella tularensis subsp. tularensis virulence factor. Infect. Immun. 77, 152–161.
Rajaram, M. V., Butchar, J. P., Parsa, K. V., Cremer, T. J., Amer, A., Schlesinger, L. S., and Tridandapani, S. (2009). Akt and SHIP modulate Francisella escape from the phagosome and induction of the Fas-mediated death pathway. PLoS One 4, e7919. doi: 10.1371/journal.pone.0007919
Rajaram, M. V., Ganesan, L. P., Parsa, K. V., Butchar, J. P., Gunn, J. S., and Tridandapani, S. (2006). Akt/Protein kinase B modulates macrophage inflammatory response to Francisella infection and confers a survival advantage in mice. J. Immunol. 177, 6317–6324.
Rathinam, V. A., Jiang, Z., Waggoner, S. N., Sharma, S., Cole, L. E., Waggoner, L., Vanaja, S. K., Monks, B. G., Ganesan, S., Latz, E., Hornung, V., Vogel, S. N., Szomolanyi-Tsuda, E., and Fitzgerald, K. A. (2010). The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat. Immunol. 11, 395–402.
Ray, K., Marteyn, B., Sansonetti, P. J., and Tang, C. M. (2009). Life on the inside: the intracellular lifestyle of cytosolic bacteria. Nat. Rev. Microbiol. 7, 333–340.
Read, A., Vogl, S. J., Hueffer, K., Gallagher, L. A., and Happ, G. M. (2008). Francisella genes required for replication in mosquito cells. J. Med. Entomol. 45, 1108–1116.
Rodriguez, S. A. (2010). Genetic Regulation of Virulence in Francisella tularensis. Doctoral thesis, the University of Texas, San Antonio.
Roth, K. M., Gunn, J. S., Lafuse, W., and Satoskar, A. R. (2009). Francisella inhibits STAT1-mediated signaling in macrophages and prevents activation of antigen-specific T cells. Int. Immunol. 21, 19–28.
Sammons-Jackson, W. L., McClelland, K., Manch-Citron, J. N., Metzger, D. W., Bakshi, C. S., Garcia, E., Rasley, A., and Anderson, B. E. (2008). Generation and characterization of an attenuated mutant in a response regulator gene of Francisella tularensis live vaccine strain (LVS). DNA Cell Biol. 27, 387–403.
Santic, M., Akimana, C., Asare, R., Kouokam, J. C., Atay, S., and Kwaik, Y. A. (2009). Intracellular fate of Francisella tularensis within arthropod-derived cells. Environ. Microbiol. 11, 1473–1481.
Santic, M., Asare, R., Skrobonja, I., Jones, S., and Abu Kwaik, Y. (2008). Acquisition of the vATPase proton pump and phagosome acidification is essential for escape of Francisella tularensis into the macrophage cytosol. Infect. Immun. 7, 7.
Santic, M., Molmeret, M., and Abu Kwaik, Y. (2005a). Modulation of biogenesis of the Francisella tularensis subsp. novicida-containing phagosome in quiescent human macrophages and its maturation into a phagolysosome upon activation by IFN-gamma. Cell Microbiol. 7, 957–967.
Santic, M., Molmeret, M., Klose, K. E., Jones, S., and Kwaik, Y. A. (2005b). The Francisella tularensis pathogenicity island protein IglC and its regulator MglA are essential for modulating phagosome biogenesis and subsequent bacterial escape into the cytoplasm. Cell Microbiol. 7, 969–979.
Santic, M., Molmeret, M., Barker, J. R., Klose, K. E., Dekanic, A., Doric, M., and Abu Kwaik, Y. (2007). A Francisella tularensis pathogenicity island protein essential for bacterial proliferation within the host cell cytosol. Cell Microbiol. 9, 2391–2403.
Schell, M. A., Ulrich, R. L., Ribot, W. J., Brueggemann, E. E., Hines, H. B., Chen, D., Lipscomb, L., Kim, H. S., Mrazek, J., Nierman, W. C., and Deshazer, D. (2007). Type VI secretion is a major virulence determinant in Burkholderia mallei. Mol. Microbiol. 64, 1466–1485.
Schlieker, C., Zentgraf, H., Dersch, P., and Mogk, A. (2005). ClpV, a unique Hsp100/Clp member of pathogenic proteobacteria. Biol. Chem. 386, 1115–1127.
Schmerk, C. L., Duplantis, B. N., Howard, P. L., and Nano, F. E. (2009a). A Francisella novicida pdpA mutant exhibits limited intracellular replication and remains associated with the lysosomal marker LAMP-1. Microbiology 155, 1498–1504.
Schmerk, C. L., Duplantis, B. N., Wang, D., Burke, R. D., Chou, A. Y., Elkins, K. L., Ludu, J. S., and Nano, F. E. (2009b). Characterization of the pathogenicity island protein PdpA and its role in the virulence of Francisella novicida. Microbiology 155, 1489–1497.
Sexton, J. A., Miller, J. L., Yoneda, A., Kehl-Fie, T. E., and Vogel, J. P. (2004). Legionella pneumophila DotU and IcmF are required for stability of the Dot/Icm complex. Infect. Immun. 72, 5983–5992.
Shalom, G., Shaw, J. G., and Thomas, M. S. (2007). In vivo expression technology identifies a type VI secretion system locus in Burkholderia pseudomallei that is induced upon invasion of macrophages. Microbiology 153, 2689–2699.
Shrivastava, S., and Mande, S. S. (2008). Identification and functional characterization of gene components of Type VI Secretion system in bacterial genomes. PLoS One 3, e2955. doi: 10.1371/journal.pone.0002955
Su, J., Yang, J., Zhao, D., Kawula, T. H., Banas, J. A., and Zhang, J. R. (2007). Genome-wide identification of Francisella tularensis virulence determinants. Infect. Immun. 75, 3089–3101.
Suarez, G., Sierra, J. C., Sha, J., Wang, S., Erova, T. E., Fadl, A. A., Foltz, S. M., Horneman, A. J., and Chopra, A. K. (2008). Molecular characterization of a functional type VI secretion system from a clinical isolate of Aeromonas hydrophila. Microb. Pathog. 44, 344–361.
Sun, G. W., Chen, Y., Liu, Y., Tan, G. Y., Ong, C., Tan, P., and Gan, Y. H. (2010). Identification of a regulatory cascade controlling Type III Secretion System 3 gene expression in Burkholderia pseudomallei. Mol. Microbiol. 76, 677–689.
Sun, P., Austin, B. P., Schubot, F. D., and Waugh, D. S. (2007). New protein fold revealed by a 1.65 A resolution crystal structure of Francisella tularensis pathogenicity island protein IglC. Protein Sci. 16, 2560–2563.
Syed, K. A., Beyhan, S., Correa, N., Queen, J., Liu, J., Peng, F., Satchell, K. J., Yildiz, F., and Klose, K. E. (2009). The Vibrio cholerae flagellar regulatory hierarchy controls expression of virulence factors. J. Bacteriol. 191, 6555–6570.
Tärnvik, A. (1989). Nature of protective immunity to Francisella tularensis. Rev. Infect. Dis. 11, 440–451.
Tatusov, R. L., Galperin, M. Y., Natale, D. A., and Koonin, E. V. (2000). The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 28, 33–36.
Telepnev, M., Golovliov, I., Grundström, T., Tärnvik, A., and Sjöstedt, A. (2003). Francisella tularensis inhibits Toll-like receptor-mediated activation of intracellular signalling and secretion of TNF-alpha and IL-1 from murine macrophages. Cell Microbiol. 5, 41–51.
Telepnev, M., Golovliov, I., and Sjöstedt, A. (2005). Francisella tularensis LVS initially activates but subsequently down-regulates intracellular signaling and cytokine secretion in mouse monocytic and human peripheral blood mononuclear cells. Microb. Pathog. 38, 239–247.
Tempel, R., Lai, X. H., Crosa, L., Kozlowicz, B., and Heffron, F. (2006). Attenuated Francisella novicida transposon mutants protect mice against wild-type challenge. Infect. Immun. 74, 5095–5105.
Thakran, S., Li, H., Lavine, C. L., Miller, M. A., Bina, J. E., Bina, X. R., and Re, F. (2008). Identification of Francisella tularensis lipoproteins that stimulate the toll-like receptor (TLR) 2/TLR1 heterodimer. J. Biol. Chem. 283, 3751–3760.
Twine, S., Byström, M., Chen, W., Forsman, M., Golovliov, I., Johansson, A., Kelly, J., Lindgren, H., Svensson, K., Zingmark, C., Conlan, W., and Sjöstedt, A. (2005). A mutant of Francisella tularensis strain SCHU S4 lacking the ability to express a 58-kilodalton protein is attenuated for virulence and is an effective live vaccine. Infect. Immun. 73, 8345–8352.
Twine, S. M., Mykytczuk, N. C., Petit, M. D., Shen, H., Sjöstedt, A., Wayne Conlan, J., and Kelly, J. F. (2006). In vivo proteomic analysis of the intracellular bacterial pathogen, Francisella tularensis, isolated from mouse spleen. Biochem. Biophys. Res. Commun. 345, 1621–1633.
Ulland, T. K., Buchan, B. W., Ketterer, M. R., Fernandes-Alnemri, T., Meyerholz, D. K., Apicella, M. A., Alnemri, E. S., Jones, B. D., Nauseef, W. M., and Sutterwala, F. S. (2010). Cutting edge: mutation of Francisella tularensis mviN leads to increased macrophage absent in melanoma 2 inflammasome activation and a loss of virulence. J. Immunol. 185, 2670–2674.
Vonkavaara, M., Telepnev, M. V., Ryden, P., Sjöstedt, A., and Stöven, S. (2008). Drosophila melanogaster as a model for elucidating the pathogenicity of Francisella tularensis. Cell Microbiol. 10, 1327–1338.
Wehrly, T. D., Chong, A., Virtaneva, K., Sturdevant, D. E., Child, R., Edwards, J. A., Brouwer, D., Nair, V., Fischer, E. R., Wicke, L., Curda, A. J., Kupko, J. J., 3rd, Martens, C., Crane, D. D., Bosio, C. M., Porcella, S. F., and Celli, J. (2009). Intracellular biology and virulence determinants of Francisella tularensis revealed by transcriptional profiling inside macrophages. Cell Microbiol. 11, 1128–1150.
Weiss, D. S., Brotcke, A., Henry, T., Margolis, J. J., Chan, K., and Monack, D. M. (2007). In vivo negative selection screen identifies genes required for Francisella virulence. Proc. Natl. Acad. Sci. U.S.A. 104, 6037–6042.
Wickstrum, J. R., Bokhari, S. M., Fischer, J. L., Pinson, D. M., Yeh, H. W., Horvat, R. T., and Parmely, M. J. (2009). Francisella tularensis induces extensive caspase-3 activation and apoptotic cell death in the tissues of infected mice. Infect. Immun. 77, 4827–4836.
Woolard, M. D., Hensley, L. L., Kawula, T. H., and Frelinger, J. A. (2008). Respiratory Francisella tularensis live vaccine strain infection induces Th17 cells and prostaglandin E2, which inhibits generation of gamma interferon-positive T cells. Infect. Immun. 76, 2651–2659.
Wu, H. Y., Chung, P. C., Shih, H. W., Wen, S. R., and Lai, E. M. (2008). Secretome analysis uncovers an Hcp-family protein secreted via a type VI secretion system in Agrobacterium tumefaciens. J. Bacteriol. 190, 2841–2850.
Keywords: Francisella tularensis, phagosomal escape, intracellular growth, FPI, T6SS
Citation: Bröms JE, Sjöstedt A, and Lavander M (2010) The role of the Francisella tularensis pathogenicity island in type VI secretion, intracellular survival, and modulation of host cell signaling. Front. Microbio. 1:136. doi: 10.3389/fmicb.2010.00136
Received: 09 November 2010;
Accepted: 02 December 2010;
Published online: 21 December 2010.
Edited by:
John S. Gunn, The Ohio State University, USAReviewed by:
Thomas C. Zahrt, Medical College of Wisconsin, USATonyia Eaves-Pyles, University of Texas Medical Branch, USA
Copyright: © 2010 Bröms, Sjöstedt, and Lavander. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Anders Sjöstedt, Department of Clinical Microbiology, Clinical Bacteriology, Umeå University, SE-901 85 Umeå, Sweden. e-mail:YW5kZXJzLnNqb3N0ZWR0QGNsaW1pLnVtdS5zZQ==