 Lennon Ramos Pereira1
Lennon Ramos Pereira1 Rúbens Prince dos Santos Alves1
Rúbens Prince dos Santos Alves1 Natiely Silva Sales1
Natiely Silva Sales1 Robert Andreata-Santos1
Robert Andreata-Santos1 Aléxia Adrianne Venceslau-Carvalho1
Aléxia Adrianne Venceslau-Carvalho1 Samuel Santos Pereira1
Samuel Santos Pereira1 Maria Fernanda Castro-Amarante1
Maria Fernanda Castro-Amarante1 Mônica Josiane Rodrigues-Jesus1
Mônica Josiane Rodrigues-Jesus1 Marianna Teixeira de Pinho Favaro1
Marianna Teixeira de Pinho Favaro1 Rosa Maria Chura-Chambi2
Rosa Maria Chura-Chambi2 Ligia Morganti2
Ligia Morganti2 Luís Carlos de Souza Ferreira1*
Luís Carlos de Souza Ferreira1*- 1Laboratory of Vaccine Development, Department of Microbiology, Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil
- 2Biotechnology Center, Institute of Energy and Nuclear Research (IPEN), São Paulo, Brazil
Zika virus (ZIKV) is a globally-distributed flavivirus transmitted to humans by Aedes mosquitoes, usually causing mild symptoms that may evolve to severe conditions, including neurological alterations, such as neonatal microcephaly and Guillain-Barré syndrome. Due to the absence of specific and effective preventive methods, we designed a new subunit vaccine based on a DNA vector (pgDNS1-ZIKV) encoding the non-structural protein 1 (NS1) genetically fused to the Herpes Simplex Virus (HSV) glycoprotein D (gD) protein. Recombinant plasmids were replicated in Escherichia coli and the expression of the target protein was confirmed in transfected HEK293 cells. C57BL/6 and AB6 (IFNAR1–/–) mice were i.m. immunized by electroporation in order to evaluate pgDNS1-ZIKV immunogenicity. After two doses, high NS1-specific IgG antibody titers were measured in serum samples collected from pgDNS1-ZIKV-immunized mice. The NS1-specific antibodies were capable to bind the native protein expressed in infected mammalian cells. Immunization with pgDNS1-ZIKV increased both humoral and cellular immune responses regarding mice immunized with a ZIKV NS1 encoding vaccine. Immunization with pgDNS1-ZIKV reduced viremia and morbidity scores leading to enhanced survival of immunodeficient AB6 mice challenged with a lethal virus load. These results give support to the use of ZIKV NS1 as a target antigen and further demonstrate the relevant adjuvant effects of HSV-1 gD.
Introduction
Zika virus (ZIKV) is an arthropod-borne virus with a positive single-stranded RNA that codes for three structural and seven non-structural proteins (1). While the ZIKV classical transmission cycle mainly involves Aedes mosquitos (1, 2), intrauterine and sexual transmission routes have also been demonstrated (3). According to the WHO, ZIKV has been reported in more than 80 countries. Infections related to this virus causes a plethora of symptoms ranging from flu-like symptoms such as fever, rash, conjunctivitis, headache and eye pain to severe forms such as Guillan-Barré Syndrome (GBS) and Zika Congenital Syndrome (CZS) (4). The latter is a set of malformations that deeply impact the development of newborns and may lead to microcephaly (5–7). Despite the burden of the disease and the urge to develop approaches to prevent virus dissemination, there are no licensed therapies or vaccines to ZIKV infection so far.
Despite the absence of a clear protection correlate against ZIKV infection, recent evidences indicate that an optimal ZIKV vaccine should induce both cellular and humoral immune responses (8–10). The induction of ZIKV-specific neutralizing antibodies (nAb) by vaccines was capable to confer protection in mice and non-human primates (NHP). In addition, passive immunization with sera isolated from vaccinated or infected NHP or humans conferred protection under experimental conditions (11–15). On the other hand, T cell mediated immune responses also contribute to the control of ZIKV infection. Increased viral loads and mortality rates are observed in CD8+ T cell-deficient mice while transfer of ZIKV-specific CD8+T cells to T cell deficient mice reduced infection and conferred protection (16–18). Recently, the role of type I CD4+ T helper cells has also shown to contribute to the control of ZIKV infection (19–21). Further published evidences repeatedly demonstrated that both nAbs and T cells responses contribute to protective immunity to ZIKV infection (14, 19, 22–24).
The abnormally high incidence of GBS and CZS in certain locations raised the question whether the pre-existing immunity to other flavivirus, such as DENV, could be implicated in such a phenomenon, likely through Antibody-Dependent Enhancement (ADE). It is well-known that ZIKV and DENV share antigen cross-reactivity at both antibody (Ab) and T cell levels, and since Abs can play a dual role in protection and in DENV pathogenesis, the same might be true for ZIKV (17, 25). In fact, studies using mouse models and, more recently, human data have already shown a direct role of anti-structural protein Abs in both DENV and ZIKV pathogenesis (26–28). Despite these evidences, most ZIKV vaccines tested under experimental conditions target generation of neutralizing antibodies against structural antigens (13, 14, 29–33). An alternative would be the use of non-structural proteins in vaccine formulations, since these antigens may trigger both B and T cell protective responses without the undesirable risks of ADE (9).
The flavivirus non-structural proteins 1 (NS1) are glycoproteins, with molecular weight ranging from 46 to 55 kDa, implicated in several mechanisms, such as replication, negative RNA strand synthesis and evasion from the host's immune response (34). NS1 is highly immunogenic and may be associated to the cytoplasmic membrane as dimers or hexamers that are secreted to extracellular medium. It has been reported that anti-NS1 Abs target infected cells and induce virus clearance by Antibody-dependent Cytotoxicity (ADCC) and deposition of complement system proteins. In addition, ZIKV-infected cells may also be targeted by T cells through MHC presentation of NS1-derived epitopes. In fact, both immunological responses generate protective immunity to ZIKV. Anti-ZIKV vaccine strategies based on recombinant vesicular stomatitis virus (rVSV) (35), DNA vaccines (23, 36) or Modified Vaccinia Ankara virus (37) showed different protection levels by inducing either one or both humoral and cellular responses. Furthermore, mice and human isolated NS1-specific monoclonal antibodies (mAbs) showed Fc-dependent protection to ZIKV challenges, including non-pregnant and pregnant mice (38, 39). These results show that ZIKV NS1 protein is a suitable antigen for vaccines based on different delivery platforms.
Previous studies showed that the genetic fusion of antigens at the C-terminal region of the herpes simplex virus type I (HSV-1) glycoprotein D (gD) may improve the induction of antigen-specific humoral and cellular immune responses, either based on the DNA vaccine platform or as purified recombinant protein-based vaccines (40–42). The HSV-1 gD is capable to bind with high affinity to several antigen presenting cell receptors, including nectin-1, and the herpes virus entry mediator (HVEM) (43), which is a member of the tumor necrosis factor receptor (TNFR) family. Interaction of gD and HVEM induces NF-κB RelA expression in a TRAF2-dependent-way, resulting in pro-survival signals within activated T cells (44, 45). The binding of gD to HVEM induces strong immunomodulatory effects on T cells as previously demonstrated by our group and others (40, 41, 46–49).
In this study we designed and tested a DNA vaccine based on expression of a chimeric ZIKV NS1 protein genetically fused to the HSV-1 gD protein in order to enhance the anti-NS1 immune responses and protective immunity to ZIKV. Immunization of immunocompetent mice with the DNA-vaccine induced increased serum NS1-specific IgG titers as well as NS1-specific IFNγ-secreting cells, in comparison with mice immunized with a vector encoding only NS1. The presence of gD was also correlated to the reduced morbidity and mortality induced in immunodeficient AB6 mice challenged with ZIKV. These results support the use of the NS1 antigen in anti-ZIKV vaccine strategies and validate the adjuvant effects associated with co-expression of HSV-1 gD protein.
Materials and Methods
Ethics Statement
Male or female C57BL/6 (6–8 weeks-old) or AB6 (IFNAR1–/–) (4 weeks-old) mice were bred under specific pathogen-free conditions at the Isogenic Mouse Facility of the Microbiology Department, University of São Paulo, Brazil. The protocols were approved by the Institutional Animal Care and Use Committee (CEUA) of the University of São Paulo (protocol number 96/2016) and conducted according to the Ethical Principles of Animal Experimentation established by the Brazilian College of Animal Experimentation. Animal group size was determined to achieve 80% resolution power using the Experimental Design Assistant (EDA) platform (https://eda.nc3rs.org.uk/).
Plasmids and Recombinant Proteins
The DNA vaccines encoding the full-length ZIKV NS1 (GenBank: ALU33341) isolated (pNS1-ZIKV) or as a chimeric protein genetically fused the HSV-1 gD protein (pgDNS1-ZIKV) were synthetized at GenScript, USA (Figure 1A). The synthetic genes were optimized to the human codon usage and designed to contain the restriction sites PstI and BglII at the N‘ and C‘ terminus, respectively. The sequences were cloned into a pUMVC3 vector (Aldevron, ND, USA), as previously described (47). For the pgDNS1-ZIKV construction the sequence of full-length ZIKV NS1 was flanked by ApaI restriction sites (Supplementary Figure 1). The Escherichia coli DH5α strain was transformed with the recombinant plasmids separately and the plasmid DNA was purified using the EndoFree Plasmid Mega kit (Cat.: 12183, QIAGEN), according to the manufacturer's instructions. The constructions were confirmed by restriction analyses and gene sequencing.
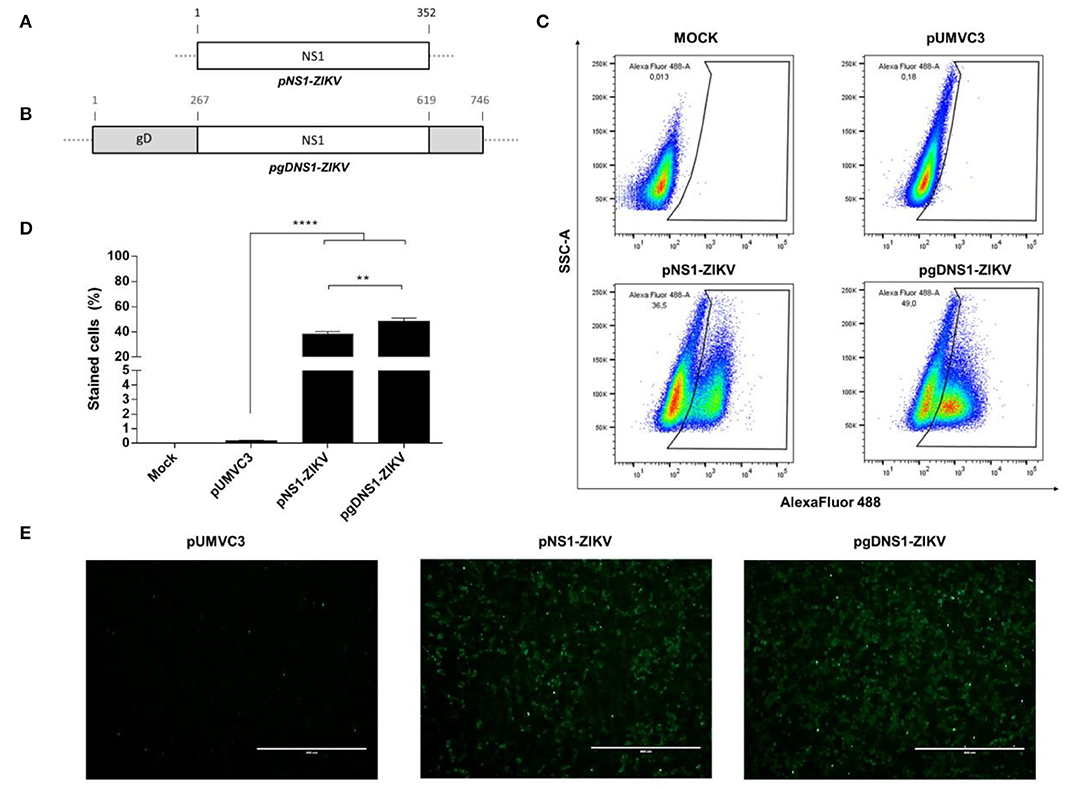
Figure 1. Expression of the recombinant ZIKV NS1 and the chimeric ZIKV NS1 and HSV-1 gD. (A,B) Schematic representations of pNS1-ZIKV, encoding the full length ZIKV NS1 protein (A) and pgDNS1-ZIKV, encoding the genetic fusion of NS1 and gD (B). (C–E) Detection of the ZIKV NS1 expressed in HEK293-T cells 24 h after transfection with the tested plasmids. Cells were labeled with anti-ZIKV NS1 mAb followed by anti-mouse IgG antibodies conjugated to Alexa Fluor 488 and analyzed by flow cytometry (C,D) and immunofluorescence (E). Data are expressed as mean ± SEM of two independent experiments performed in duplicate. Scale bar = 400 μm. Statistical significance was calculated using One-way ANOVA with post-test Bonferroni (**p < 0.01, ****p < 0.0001, NS = not significant).
The ZIKV NS1 protein was expressed on the recombinant E. coli BL21-CodonPlus (DE3)-RIL cells and purified by affinity chromatography after in vitro refolding of the protein under high pressure conditions, as previously described (50).
Virus and Cells Lines
The Brazilian clinical ZIKV isolate (ZIKVBR) (GenBank: KU729217.2) was obtained from Evandro Chagas Institute in Belém (Pará, Brazil). ZIKVBR propagation was carried out in Aedes albopictus clone C6/36 cells cultured in Leibovitz L-15 medium (Vitrocell, Brazil) supplemented with 2% fetal bovine serum (FBS) (Life Technologies, USA), followed by a concentration protocol as previously described (5, 51). Vero CCL-81 cells were cultured in Minimum Essential Medium Eagle (MEM, Vitrocell) with 10% FBS. Human embryonic kidney cells (HEK-293 cells, ATCC No. CRL-11268) were cultured in Dulbecco's modified Eagle's medium (DMEM, Life Technologies) supplemented with 10% FBS.
Detection of ZIKV NS1
Expression of ZIKV NS1 protein was detected either through immunofluorescence or flow cytometry. In both cases, HEK-293 cells were equally transfected with the plasmids according to a previously described protocol (52). Briefly, HEK-293 cells were seeded (105 cell/well) in 24-wells plates (Corning, USA) and cultured at 37°C (5% CO2) until reaching 70–80% confluence. One hour before the transfection, the cell culture supernatant was removed and 200 μl/well of DMEM (supplemented with 2% FBS) was added to the cells. For transfection, MIX solutions containing 1 μg of the plasmids to 3 μg of polyethyleneimine (PEI) (Cat.: 408727, Sigma Aldrich, USA) were prepared in 150 mM NaCl solution. After incubation for 5 min at room temperature (RT), the mixtures were distributed evenly over the plates and the cells were incubated for up to 24 h (37°C, 5% CO2).
For detection of ZIKV NS1, transfected cell monolayers were washed (2×) with PBS and fixed with 4% formaldehyde diluted in PBS for 15 min (RT). After incubation, cells were permeabilized with 0.1% Triton X-100 in PBS (15 min at RT) and blocked with a blocking solution (BS) (2% BSA diluted in PBS) for 30 min at RT. After a new washing cycle (3×), the cells were stained with anti-ZIKV NS1 4H2 mAb (20 μg/ml) diluted in BS and kept for 1 h at room temperature (RT) under agitation. After washing (3×) with 0.05% Tween 20 in PBS (PBS-T), cells were incubated (45 min, at RT) with anti-mouse IgG antibody conjugated to AlexaFluor 488 (Cat.: A11001, Invitrogen, USA). Finally, cells were washed with PBS-T and observed under an immunofluorescence microscope (Evos FL Thermo Scientific, USA) with images captured at 100x magnification.
For flow cytometry assays, cell monolayers were washed 2× with PBS, trypsinized, and fixed/permeabilized with a Cytofix/Cytoperm kit (BD Bioscience, USA), according to the manufacturer's instructions. Cells were, then, stained with the primary antibody 4H2 mAb (5 μg/ml) on ice for 30 min. After washing (2×), cells were incubated (30 min on ice) with the anti-mouse conjugated to Alexa Fluor 488 (Cat.: A11001, Thermo Fisher Scientific) diluted 1:800. Finally, after washings (2×), the cells were suspended in 200 μl of PBS-2% FBS solution and analyzed by an LSR FortessaTM analyzer (BD, Franklin Lakes, NJ, USA). The data obtained were analyzed using FlowJo software (version 10, Tree Star, San Carlo, CA) to determine the percentage of stained cells.
Mice and Immunization Regimen
C57BL/6 (5–7 animals/group) or AB6 (4–6 animals/group) mice were immunized with 2 doses of 50 μg of pUMVC3, pNS1-ZIKV, or pgDNS1-ZIKV formulated in apyrogenic saline (0.9%). For immunization, the animals were previously anesthetized with a mixture of Ketamine and Xylazine (100 and 10 mg/kg, respectively) administered intraperitoneally (i.p.). Then, the animals received intramuscular (i.m.) injections with the formulations followed by electroporation (two 130 V pulses with 1 ms duration and four 70 V pulses with 50 ms duration, with an interval of 450 ms between each pulse), which was carried out with the CUY560-5-0.5 electrode using the NEPA21 Super Electroporator (Nepa Gene Co, Japan.), at an interval of 2 weeks. Blood samples were obtained by submandibular plexus puncture 14 days after the administration of each dose and centrifuged at 3,000 g for 30 min to separate the sera. The obtained serum samples were stored at −20°C for future analysis.
Enzyme-Linked Immunosorbent Assay (ELISA)
High-binding ELISA plates (Corning) were coated with purified ZIKV NS1 protein (200 ng/well) diluted in coating buffer (32.5 mM NaHCO3, 14.9 mM Na2CO3, pH 9.6) for 18 h at 4°C. The plates were washed three times with PBS containing 0.05% Tween-20 (PBS-T) and added with 200 μl/well of a blocking buffer (BB) containing 5% non-fat milk solution with 1% bovine serum albumin (BSA) in PBS-T. After incubation (2 h at 37°C), plates were washed (3×) and 100 μl of the individual serum samples, previously diluted (starting 1:25) in BB, were added to the wells and incubated for 1 h at 37°C. After a new wash cycle (3×), anti-mouse IgG antibody conjugated to horseradish peroxidase (HRP) (Sigma Aldrich) was added to the wells at 1:4,000 dilution in BB. For the determination of IgG subclasses, anti-mouse IgG1 or IgG2c HRP-conjugated antibodies (Southern Biotech, USA) were used. After incubation (1 h, 37°C), plates were washed (3×) and 100 μl of developing solution containing o-phenylenediamine dihydrochloride (OPD) (Sigma Aldrich) and H2O2 were added to each well and incubated for 15 min at RT in the dark. The reaction was stopped with 50 μl/well of 1 M sulfuric acid and the absorbance was measured at 492 nm (Ab492nm) with a plate reader (BioTek, Winooski, VT, USA). Antibody titers were determined as the reciprocal end dilution value of the serum sample at which the absorbance obtained was at least 0.1 units above of the cut-off value (mean + 2 SD of Ab492nm obtained on wells containing no serum).
Antibody Binding to Native NS1 Protein
Vero cells were seeded (5 × 104 cell/well) in flat 96-well plates (Corning) and incubated for 24 h at 37°C (5% CO2). Established cell monolayers were infected with ZIKVBR at multiplicity of infection (MOI) 1.0 for 24 h (37°C, 5% CO2). After infection, the monolayers were washed with PBS (2×), trypsinized (Vitrocell) and fixed/permeabilized with a Cytofix/Cytoperm kit (BD Bioscience) according to the manufacturer's instructions. The cells were labeled with pooled serum samples from immunized mice diluted 1:1,000 or 4H2 mAb (5 μg/ml) for 30 min on ice. After washing (2×), cells were stained for 30 min (on ice) with a goat anti-mouse IgG antibody conjugated to Alexa Fluor 488, diluted 1:800 (Cat.: A11001, Thermo Fisher Scientific). After new wash cycles (2×), the cells were suspended and analyzed in the LSR FortessaTM analyzer (BD, Franklin Lakes, NJ, USA). The data were analyzed using FlowJo software (version 10, Tree Star, San Carlo, CA) to determine the percentage of stained Vero cells.
Peptide Synthesis
The peptides used in this work were synthesized at GenScript USA Inc. The prediction of immunogenic regions was carried out with algorithm (53, 54) hosted at the Immune Epitope Database (IEDB) Tools (http://tools.iedb.org/mhci/), using the amino acid sequence corresponding to the C-terminal portion of the native ZIKV NS1 protein (GenBank–ALU33341). Binding affinities were obtained for all 8–14 mer peptides for the H2-Kb and H2-Db alleles. The selected alleles had the classification of the consensus percentage restricted to 1.7% with a score ≤1. Two high-score-predicted peptides were selected, custom-made by GenScript (Piscataway, NJ, USA), suspended in DMSO and stored at −80°C before use.
Analysis of Cellular Immune Response
Measurement of induced cellular immune responses was carried out according to a previously described adapted protocol (16). Briefly, 2 weeks after the last vaccine dose, the immunized mice were intravenously (i.v.) infected with 106 plaque-forming units (PFU) of ZIKVBR. Three days after infection (d.p.i) the animals were euthanized, the spleens were harvested surgically under aseptic conditions and the splenocytes were isolated, as previously described (52). Then, the isolated spleen cells were stimulated in vitro with peptides derived from ZIKV NS1 and the number of IFNγ-producing cells was determined by enzyme-linked immune absorbent spot (ELISpot).
ELISpot was carried out according to a previously described protocol (55). Briefly, splenocytes from immunized and infected mice were seeded (2 × 105 cells/well) in 96-well plates (Millipore, USA), previously coated with capture antibody to IFNγ (BD Biosciences, USA). Then, the cells were cultured for 48 h (37°C, 5% CO2) in the presence or absence of NS1-derived peptides (100 ng of each peptide/well). After stimulation, the plates were washed (3 × with PBS and 5 × with PBS-T) and stained (2 h at RT) with a biotinylated mouse anti-IFNγ mAb (BD Biosciences, USA) at 2 μg/ml. After washing, the plates were incubated with peroxidase-labeled streptavidin (Sigma, USA) for 2 h at RT. Plates were washed again and developed with TrueBlue™ Substrate (KPL, Milford, MA, USA) for 20 min at RT. The spots were counted and expressed as IFNγ-producing cells/106 splenocytes.
Lethal Challenge With the ZIKVBR
Two weeks after administration of the second vaccine dose, immunized AB6 mice were i.v. infected with 106 PFU of ZIKVBR. The animals were monitored for up to 15 days for the appearance of clinical signs according to an arbitrary score scale (Healthy, score 0; ruffled fur, score 1; paralysis, score 2; deformed spinal column, score 3; moribund, score 4), measurement of body weight and mortality. Serum samples were collected on alternate days and stored at −80°C for analysis of viremia.
Virus Titration
The number of infectious ZIKV particles was determined by plaque assay. Briefly, 10-fold serial dilutions of virus samples were prepared in MEM medium, which were added to the Vero cells monolayers previously established in 24-wells plates (Corning) and incubated for 1 h at 37°C. After incubation, the cell supernatants were removed by aspiration, an overlay solution containing MEM plus carboxymethylcellulose (1%) and SFB (2%) was added to the cells and incubated (37°C, 5% CO2) for 4 days. The cells monolayers were fixed with formaldehyde (4%) diluted in PBS for 15 min (RT). After washing with water, cells were stained with 1% violet crystal solution (Laborclin) for 10 min (RT). Viral lysis plates were counted and expressed as plaque form units per mL (PFU/ml).
Statistical Analysis
Statistical analyses were performed using the Prism 6 software (GraphPad Software Inc, LA Jolla, CA). T-test was used to compare only two groups. One-way ANOVA was applied with Bonferroni's post-hoc test to compare results involving several (≥3) groups. Two-way ANOVA followed by Bonferroni's correction was used when the data involved several groups and more than one variable (time points). Log-rank test (Mantel-Cox) was used to analyze the survival and morbidity data. Differences were considered significant when the p-value (p) was ≤0.05.
Results
The DNA Vectors Encoding the ZIKV NS1 Protein
Two plasmids encoding the full-length ZIKV NS1 protein, either isolated (pNS1-ZIKV) (Figure 1A) or genetically fused to HSV-1 gD protein (pgDNS1-ZIKV) (Figure 1B), were constructed. Plasmids were submitted to restriction analyses with BglII and PstI and the released fragments had the expected electrophoretic mobilities (NS1–1,056 pb; gDNS1–2,262 pb) (Supplementary Figure 1). The expression of the encoded proteins in HEK 293 cells transfected with pNS1-ZIKV or pgDNS1-ZIKV was confirmed with NS1-specific antibodies by flow cytometry and immunofluorescence (Figures 1C,E). Interestingly, cells transfected with pgDNS1-ZIKV showed enhanced expression of ZIKV NS1 with regard to cells transfected with pNS1-ZIKV (Figure 1D). The expression of the HSV-1 gD in pgDNS1-ZIKV-transfected cells was also confirmed by immune blots (Supplementary Figure 2). Taken together, these results demonstrate that the recombinant proteins encoded by the DNA vaccines were properly expressed in mammalian cells.
Fusion of ZIKV NS1 to HSV-1 gD Enhances the Induced Humoral and Cellular Immune Responses
Next, we accessed the immunogenicity of the DNA vaccines in wild-type (WT) C57BL/6 mice. The animals were immunized with two i.m. doses by electroporation (Figure 2A). As observed in Figure 2B, similar serum NS1-specific IgG titers were detected 14 days after the second vaccine dose in mice immunized with pNS1-ZIKV or pgDNS1-ZIKV. Analyses of the NS1-specific serum IgG subclass responses indicated that vaccinated mice elicited similar IgG1 and IgG2c subclass responses (Figure 2D). We also investigated the binding of pooled anti-NS1 serum samples to native NS1 expressed in the ZIKV-infected Vero cells. As shown in Figure 2C and Supplementary Figure 3, anti-NS1 antibodies raised in the mice immunized with pgDNS1-ZIKV showed higher cell binding activity with regard to serum samples collected from mice immunized with pNS1-ZIKV. As expected, mice immunized with the pUMVC3 vector did not elicit anti-NS1 antibody responses (Figures 2B,C).
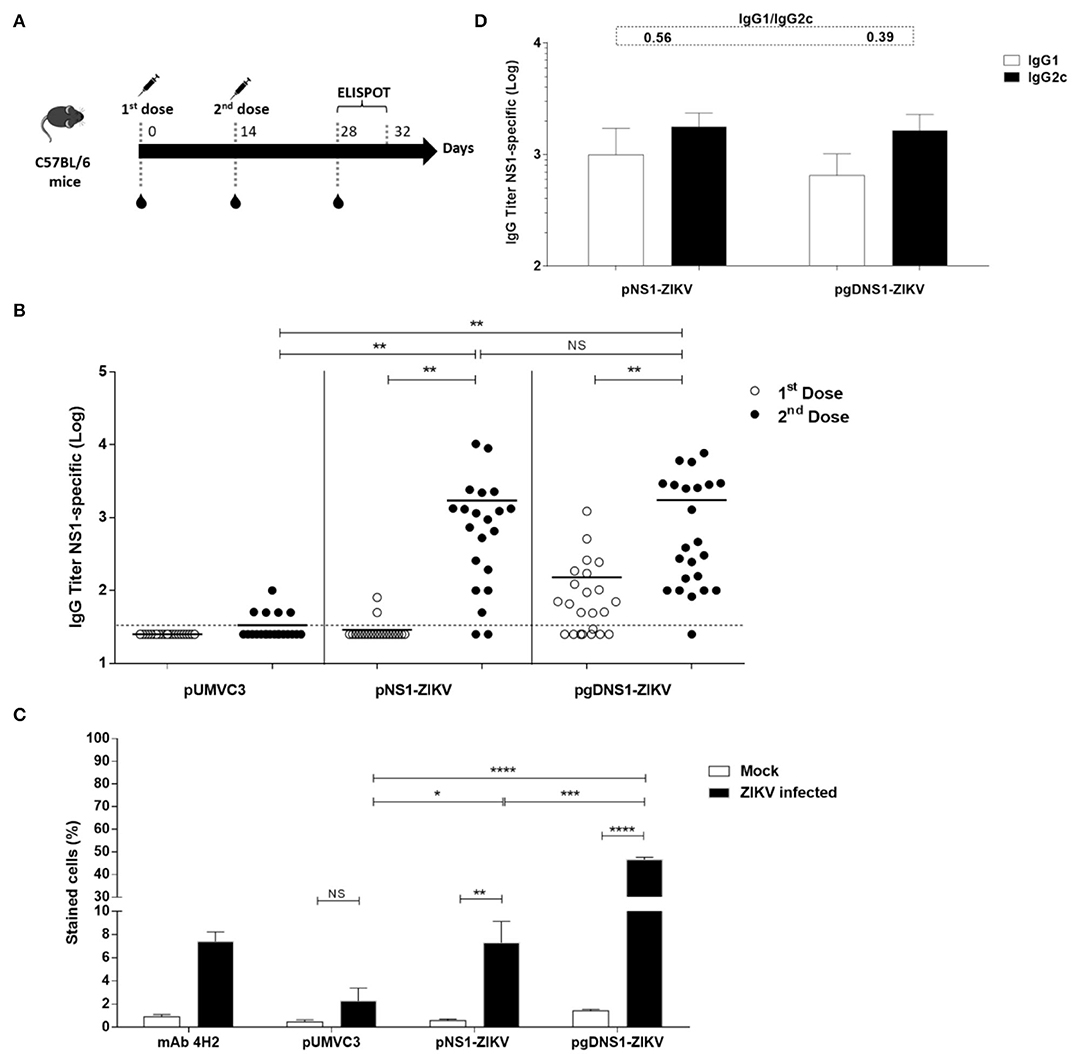
Figure 2. Immune responses elicited in vaccinated immunocompetent mice. (A) Schematic representation of the vaccination regimen. C57BL/6 mice were i.m. immunized with two doses (50 μg/animal) of DNA vaccines administered with electroporation. Serum samples were collected 14 days after each vaccine dose and the specific T cell responses were measured 2 weeks after the last vaccine dose by ELISpot. (B) Anti-NS1 IgG dose-response profiles measured by ELISA with individual serum samples collected from vaccinated mice (pUMVC3 and NS1-ZIKV, n = 21/each group; pgDNS1-ZIKV, n = 23). Data are expressed as means (bars) and individual (symbols) IgG titers (Log10) obtained from three independent experiments. (C) Binding of anti-NS1 serum antibodies to the native ZIKV NS1 protein. Diluted pooled serum samples collected from vaccinated mice (n = 10) were reacted with ZIKV-infected VERO cells and analyzed by flow cytometry. The 4H2 mAb (specific to NS1) was used as a control. (D) IgG subclass responses measured 14 days after the second vaccine dose in the individual serum samples from immunized mice (n = 10). The IgG1/IgG2c ratios are indicated at the top of the figure. Statistical significance was calculated using Two/One-way ANOVA with Bonferroni correction. T-test was used to analyze the IgG subclass responses (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, NS = not significant).
Fusion of antigens to HSV-1 gD has been shown to increase cellular immune responses to passenger antigens in vaccinated mice (40, 46–48). Thus, we also evaluated the antigen-specific cellular responses induced in mice immunized with the vaccine formulations. For that purpose, in silico predicted MHC-I restrict peptides of ZIKV NS1 were validated with splenocytes of ZIKV-infected mice (Supplementary Figure 4). The peptide prediction was performed using the C-terminal region of NS1 protein based on a recent report describing the presence of immunodominant CD8+ T cell epitopes in mice (23). Moreover, in order to maximize the detection of NS1-specific T cells, immunized mice were infected with ZIKV and, 3 days later, the numbers of IFNγ-producing cells were determined. This rationale was based on previous evidences demonstrating that expansion of antigen-specific CD8+ T cells may be accessed at this time-point after virus infection (16). Thus, the selected peptides (Supplementary Table 1) were applied in IFNγ-ELISPOT assays carried out with splenocytes harvested at 3 d.p.i from immunized mice (Figure 2A). As shown in Figure 3, only mice immunized with pgDNS1-ZIKV elicited statistically significant enhancement in the number of IFN-γ secreting spleen cells after in vitro stimulation with two different MHC-I restricted peptides. Taken together, these results indicate that immunization with pgDNS1-ZIKV simultaneously improves NS1-specific IgG response and T cell responses.
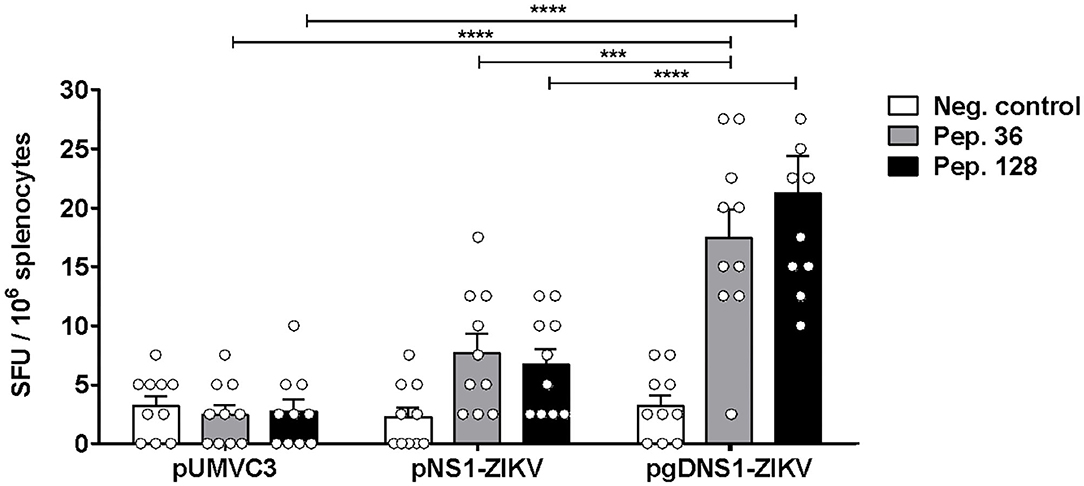
Figure 3. Analysis of cellular immune responses elicited in vaccinated immunocompetent mice. Immunized mice (n = 10) were infected with ZIKVBR and the splenocytes were harvested 3 d.p.i. Isolated cells were stimulated in vitro with ZIKV NS1 peptides and analyzed regarding cytokine production via IFNγ-ELISpot. Data are presented as mean ± SEM (bars) or individual (symbols) amounts of spots. Statistical significance was calculated using Two-way ANOVA with Bonferroni's correction (***p < 0.001, ****p < 0.0001).
ZIKV-Infected Immunodefient Mice Showed Reduced Viremia After Immunization With NS1-Encoding DNA Vaccines
Since wild type mice are not susceptible to ZIKV, we tested the protective immunity induced by the NS1-encoding DNA vaccines in the AB6 mouse strain, deficient in the expression of type I IFN receptor (IFNAR1–/–) (56, 57). AB6 mice were immunized following the same regimen with 2 i.m. doses 2 weeks apart, and, 2 weeks after the last dose, animals were challenged with i.v. inoculation of 106 PFU of ZIKVBR (Figure 4A). Before the virus challenge, higher anti-NS1 serum IgG antibodies were detected in mice immunized with pgDNS1-ZIKV when compared to mice immunized with pNS1-ZIKV (Figure 4B). Notably, mice immunized with pNS1-ZIKV or pgDNS1-ZIKV showed prevailing IgG2c subclass responses (IgG1/IgG2c ratios of 0.074 and 0.161, respectively) (Figure 4C). Viremia was followed in vaccinated and challenged mice up to 7 days. As shown in the Figures 4D–F, mice immunized with the NS1-encoding DNA vaccines presented reduction of viremia with regard to mice immunized with pUMVC3 group. Similarly, mice immunized with either pNS1-ZIKV or pgDNS1-ZIKV showed a shorter viremia period (Figures 4D–F). These results clearly demonstrate that immunization with DNA vaccines encoding ZIKV NS1 has a direct impact on the length of the period and intensity of the viremic state, as measured in vaccinated AB6 mice.
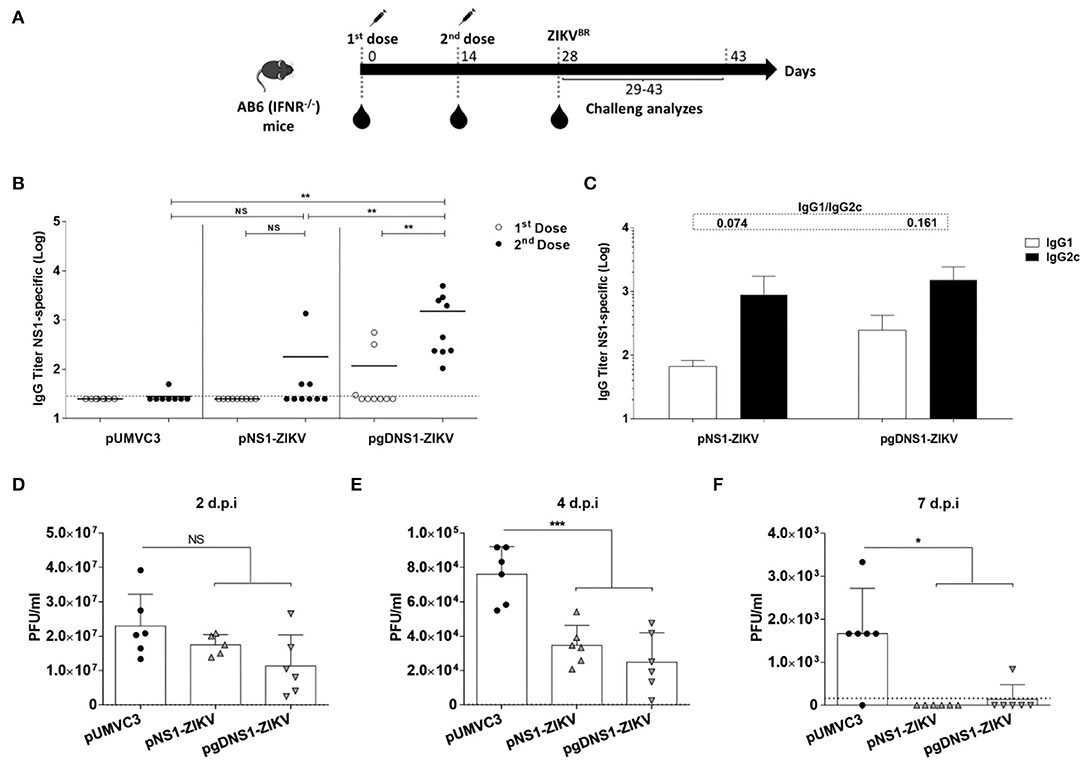
Figure 4. Immune responses and protective immunity induced by the DNA vaccines in immunodeficient mice. (A) Schematic representation of the vaccination regimen. AB6 mice were immunized twice by i.m. route in association with electroporation. Serum samples were collected 14 days after each vaccine dose. Mice were i.v. infected with 106 PFU of ZIKVBR 2 weeks after the last vaccine dose and monitored for viremia, morbidity and mortality for 15 days. (B) Total and (C) IgG subclass anti-NS1 responses were measured by ELISA with serum samples collected from immunized mice (n = 9). Data were obtained from three independent experiments and are expressed as mean (bars) ± SEM or individual values (symbols) of antibodies titers (Log10). The IgG1 and IgG2c ratios are indicated at the top of the figure. (D–F) 2 weeks after the last dose, immunized mice (n = 5) were i.v. challenged with ZIKV (106 PFU/animal). Viremia is expressed in mean ± SD of PFU/ml, measured individually until day seven after infection. The results are representative of three independent experiments. Statistical significance was calculated using Two/One-way ANOVA with Bonferroni correction. T-test was used to analyze the IgG subclass results (*p < 0.05, **p < 0.01, ***p < 0.001, NS = not significant).
Immunization With pgDNS1-ZIKV Increased Anti-ZIKV Protective Immunity in AB6 Mice
Immunized AB6 mice were monitored for vaccine-induced protection to ZIKV-induced morbidity and mortality. Using a symptom scale classification, mice immunized with pgDNS1-ZIKV showed reduced morbidity compared to non-vaccinated mice than animals immunized with pNS1-ZIKV (Figures 5A–D). pgDNS1-ZIKV vaccinated mice recovered body weight in ~10 days post infection, while those immunized with pNS1-ZIKV recovered their body weight 2 weeks after the challenge (Figure 5E). We also measured the vaccine induced anti-ZIKV protection in AB6 mice challenged with a lethal i.v. virus dose. As indicated in the Figure 5F, immunization with pgDNS1-ZIKV conferred higher survival (46%) to lethal ZIKV infection compared to mice immunized with pNS1-ZIKV (27%). No protection was observed in mice immunized with the control vector. Taken together, these results indicated that DNA vaccines encoding ZIKV NS1 confer partial protection to ZIKV infection in immunodeficient AB6 mice. In addition, the present results demonstrated that genetic fusion of NS1 to gD protein improves the induction of antigen-specific immune responses and the protective immunity to ZIKV.
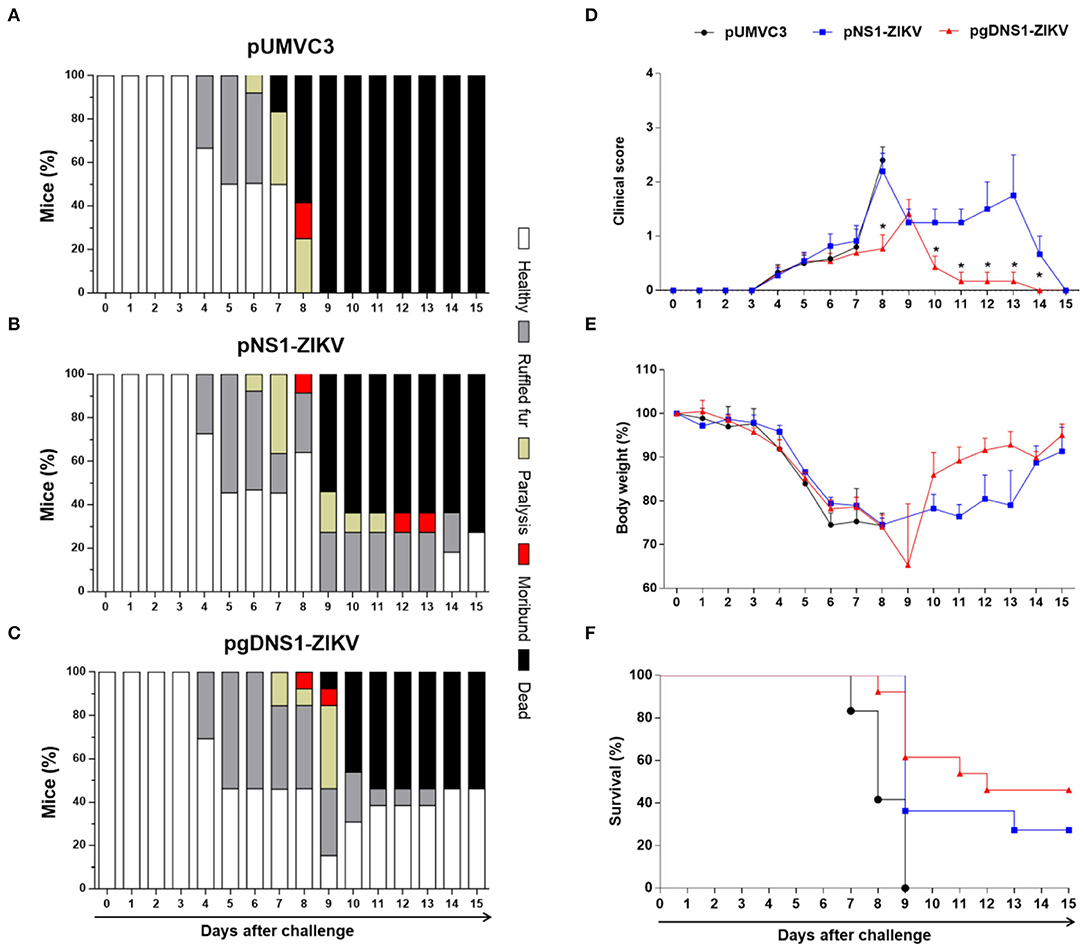
Figure 5. Protective immunity induced by ZIKV NS1-encoding DNA vaccines. AB6 mice were immunized with pUMVC3 (n = 12) (A), pNS1-ZIKV (n = 11) (B) or pgDNS1-ZIKV (n = 13) (C) and monitored daily for up to 15 days after the ZIKV challenge. The results are represented as the percentage of animals classified with the clinical score condition (Healthy, ruffled fur, paralysis, moribund, or dead), in a color scale, as indicated in the figure. Clinical score (D), body weight change (E) and mortality (F) profiles were evaluated in the challenged animals. The data represent results of three independent experiments and are presented as means ± SEM of percentage of the respective parameters followed in the different immunization groups. Statistical significance was evaluated using Two-way ANOVA with Bonferroni's correction. Mantel-Cox test was used to analyze the survival results. (*p < 0.001).
Discussion
Recent advances demonstrate that both cellular and humoral immune responses play important roles in the development of protective immunity to flavivirus under experimental and clinical conditions. Therefore, ideal vaccines for diseases associated with DENV and ZIKV infections shall preferentially activate both arms of the immune system (19, 58, 59). Here, we describe the use of a novel DNA vaccine strategy based on the expression of ZIKV NS1, either alone or genetically fused to the HSV-1 gD protein, as an approach to evaluate the protective role of anti-NS1 specific immune responses under experimental conditions. The results clearly demonstrated that immunization with NS1-encoding DNA vaccines induce NS1-specific immune responses in immunocompetent and immunodeficient mice leading to partial protection against lethal systemic ZIKV challenge in immunodeficient AB6 mice. The use of electroporation as a mean to deliver the DNA vaccines resulted in similar serum anti-NS1 IgG titers in mice immunized with pgDNS1-ZIKV or pNS1-ZIKV. Nonetheless, fusion of ZIKV NS1 to gD enhanced the binding of antibodies to the native NS1 antigen expressed in infected mammalian cells. Moreover, using splenocytes from pgDNS1-ZIKV-vaccinated mice we detected higher numbers of IFN-γ-secreting cells after in vitro stimulation with MHC-I-restricted NS1-derived peptides. More relevantly, immunization of immune deficient mice with the gD-NS1-encoding DNA vaccine reduced with more efficiency the morbidity and mortality rates detected after challenge with ZIKV. Collectively, these results support the use of genetic fusion of antigens to HSV-1 gD as promising platform for the development of ZIKV vaccine strategies and validate the adjuvant effects of the gD protein on the immune responses induced by ZIKV NS1-based DNA vaccines.
The potential of NS1 as a target antigen has been investigated regarding induction of both humoral and cellular immune responses (23, 35, 37, 60). This protein is targeted to circulating anti-NS1 antibodies either in plasma, as hexamers, or at cytoplasmic membrane of infected cells, as dimers, which may lead to cell lysis (38). Since NS1 protein is not present on the virus particle, anti-NS1 antibodies are not capable to induce the ADE phenomenon (36, 38, 39, 61). Although anti-DENV-NS1 antibodies have been implicated to side effects associated with cross-reactivity with host proteins (62–64), such effects seems to be virus specific, since no similar effects have been reported with Japanese encephalitis virus (JEV) anti-NS1 antibodies (64). Moreover, there are conflicting evidences regarding the protective and deleterious effects associated with NS1-specific antibodies both in DENV and ZIKV infections (65, 66). On the other hand, passive immunization of mice with polyclonal or monoclonal anti-NS1 antibodies promoted a clear protective effect to virus infection (36, 38, 39). Recently, a vaccinia ankara virus (MVA) modified to express the ZIKV NS1 induced protection after an intracranial challenge by inducing NS1-specific IgG2a and polyfunctional NS1-specific CD8+ T cells (35, 37). Human-derived anti-NS1 mAbs were also found to confer partial protection to immunodeficient ZIKV-challenged mice through passive immunity (38, 39). Furthermore, T cell–mediated immunity plays an important role in the protection induced by NS1-based DNA vaccines against ZIKV infection (23). Thus, robust induction of anti-NS1 antibodies associated with activation of T cell responses represent key features of a NS1-based vaccine capable to generate protective immunity to ZIKV.
The protective role of anti-NS1 antibodies has been correlated to the binding activity to surface–associated NS1 in ZIKV-infected cells, which promotes the clearance of the infected cells via ADCC or FcR-mediated complement (FC) cytotoxicity (36–39). Our data indicate that although similar serum anti-NS1 IgG responses were induced with the tested NS1-based DNA vaccines in immunocompetent mice, pgDNS1-ZIKV-vaccined mice generated anti-NS1 antibodies with higher cell binding activity compared to those immunized with pNS1-ZIKV. Since these antibodies are capable to recognize membrane-associated NS1 dimers in ZIKV-infected cells, we expect that, under in vivo conditions, this would favor the clearance of infected cells. The presence of higher IgG2c levels in pgDNS1-ZIKV immunized AB6 mice also supports this hypothesis, since IgG2c antibodies are known to strongly bind to ADCC-mediator FcyRIV (67, 68). Thus, the enhanced immunity observed in this group may be related to the role of gD on the modulation of the induced antigen-specific responses. Moreover, cell transfection with pgDNS1-ZIKV increases the in vitro expression of NS1 compared to pNS1-ZIKV-transfected cells, a characteristic that may also impact the immunogenicity of the encoded proteins. These findings highlight the adjuvant effects of gD on the immunogenicity of antigens encoded by DNA vaccines.
The relevance of T cells in the control of ZIKV infection has been highlighted in different studies. This type of immune response was recently linked to protection to ZIKV challenge at experimental conditions, but also in the context of protection to secondary heterologous flavivirus infections (58, 69, 70). Moreover, T cell mediated immune responses induced by NS1-vaccination have being described as a key feature in the control of ZIKV infections under preclinical conditions (23, 37). Similar findings have also been reported in clinical studies demonstrating that NS1 is capable to induce robust IFN-γ production by T cells during ZIKV infection (71, 72). Since gD antigen-fusion platform has been extensively explored to increase T cell mediated protective immune responses (40, 41, 46–48), here we tested whether, under a different vaccine context, such approach would lead to enhancement of NS1-specific IFN-γ producing T cells. Using MHC-I restrict peptides of ZIKV NS1 to activate in vitro spleen cells of immunocompetent C57BL/6 immunized mice, we observed that only expression of NS1 fused to gD protein promoted significant enhancement of IFN-γ secreting cell responses. Since we used MHC-I restrict peptides to stimulate the cells in vitro, these results also suggest activation of CD8+ T cell responses in vaccinated mice and open perspectives for the use of the identified MHC-I-restricted peptides in future studies dealing with induction of NS1-specific T cell responses under experimental conditions. Taken together, these findings indicated that gD fusion strategy is a promising alternative to enhance NS1-specific cellular response induced by vaccination.
Our data also corroborate with a recent study reporting the use of DNA vaccines encoding the ZIKV NS1 protein (23). Despite the differences in the tested vaccine regimen, including number of doses (total of three), immunization route (intradermal) and mouse genetic background (BALB/c), only expression of a modified form of NS1 (pVAX-tpaNS1) was able to induce high titers of antibodies capable of recognizing NS1 on infected cells and significant activation of T cell responses. Moreover, the authors reported the presence of immunodominant CD4+ and CD8+ T cell epitopes at the C-terminal region of NS1 (aa 172 to 352) (23). Similarly, the epitopes restricted to MHC-I predicted and validated in our work are mapped in the same region of the protein. In addition, the genetic fusion with HSV-1 gD protein led to enhanced activation of NS1-specific IFN-γ secreting cells in vaccinated mice. The differences in the CD8+ T cell epitopes used in the two studies are reflect the distinct genetic backgrounds of the tested vaccinated mouse strains (BALB/c × C57BL/6). On the other hand, since the validation of the peptides described in our study was performed after infection with ZIKV, these findings may reflect part of repertoire of immunodominant epitopes detected in more natural conditions. Nonetheless, since we did not evaluate the induced immune responses with the full length NS1, other epitopes may also be targeted after immunization with pgDNS1-ZIKV. Similarly, further studies shall address the positive impacts of gD antigen fusion with the induction of NS1-specific CD4+ T cell responses.
Wild type (WT) mice, such as C57BL/6, have preserved type I IFN responses that provide defense against flaviviruses infections and, consequently, do not permit efficient virus replication, generation of morbidity and lethality effects related to ZIKV infection (73, 74). To access the protective capacity of the formulations tested here we used a type I IFN receptor–deficient (IFNAR1–/–) mouse strain, which is susceptible to ZIKV infection (57). Adopting the same vaccination protocol, we detected higher serum anti-NS1 IgG titers in mice immunized with pgDNS1-ZIKV when compared to mice immunized with pNS1-ZIKV. After challenge, pgDNS1-ZIKV-vaccinated mice showed reduced morbidity and lethality scores compared to animals immunized with pNS1-ZIKV. In addition, both NS1-based DNA vaccines induced predominant serum IgG2c subclass response and reduced viremia after infection. Taken together, these observations confirm the adjuvant effects of the gD-fusion strategy applied to DNA vaccines, leading to enhanced NS1-specific humoral and cellular immune responses in immunocompetent mice and protective immunity in AB6 mice. The observed NS1-mediated protective profiles are aligned with previous studies dealing with ZIKV NS1 based in immunodeficient mice (35), as well as in immunocompetent BALB/c (23, 60) or CD-1/ICR (37) mice. Moreover, in contrast to the findings described by a study using NS1-based DNA vaccine (23), our formulations was capable to confer protection in IFNAR1−/− mice even after challenge with a high infection dose (106 PFU).
Despite the advantages of DNA vaccines with regard to other vaccine approaches, which includes manufacturing and costs, DNA vaccines usually show reduced immunogenicity when tested at clinical conditions (75). Electroporation represent one of the most effective strategy to enhance the immunogenicity of DNA vaccines both in mice and humans (76). Previous evidences demonstrated that electroporation enhance activation of T-cell mediated protective responses in a murine tumor challenge model (77). In the present study, administration of the DNA vaccines via electroporation contributed to the induction of immune responses with reduction of vaccine doses administered in the animals with regard to other study based on DNA vaccine encoding ZIKV NS1 (23). Altogether, the present evidences further support the relevance of electroporation as a preferred delivery method for administration of DNA vaccines either at experimental or clinical conditions.
Although the adjuvant role of gD is evident in all strategies explored so far, the mechanisms related to the observed adjuvant effects remain not totally elucidated. The gD immunomodulatory effects are mainly based on the binding capacity to specific receptors located on the surface of antigen presenting cells, such as the HVEM receptor in dendritic cells (DC) (48). Binding of gD interferes with interaction of HVEM to its native immunosuppressive ligands leading to final enhancement of B and T cells activation (48). gD binding to HVEM also triggers the activation of NF-kappa B pathway promoting pro-survival signals in T cells (44, 45, 78). Additionally, as previously described, purified chimeric gD-based proteins are capable to activate a DC subset specialized in antigen cross-presentation and leading to enhance generation of activated antigen-specific CD8+ T lymphocytes (41). These effects are in line with the results reported here in which generation of a chimeric NS1/gD protein promoted enhanced NS1-specific cellular responses.
Altogether, the findings reported here described for the first time the strategy based on the use of DNA vaccines encoding ZIKV NS1 genetically fused with the HSV-1 gD protein. The adjuvant effects observed with the chimeric NS1/gD, regarding induction of NS1-specific cellular and humoral immune responses, support the use of such approach in further attempts to enhance immunity to ZIKV induced by DNA vaccines and open perspectives for the development of effective anti-ZIKV vaccines as well as other flaviruses.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by Institutional Animal Care and Use Committee (CEUA) of the University of São Paulo (protocol number 96/2016).
Author Contributions
LP and LF: conceived and designed the experiments. LP, RA, NS, RA-S, AV-C, SP, MC-A, MR-J, MF, and RC-C: performed the experiments. LP and RA: analyzed the data. MR-J and LM: contributed reagents, materials, and analysis tools. LP: prepared the figures. LP, RA, RA-S, MC-A, and LF: wrote the paper. NS, AV-C, SP, MR-J, and MF: revised the manuscript. All authors read and approved the final version of the manuscript.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). FAPESP grants numbers: 2016/20045-7 (LF), 2016/05570-8 (LP), 2014/17595-0 (RA), 2016/14344-1 (NS), 2016/23560-0 (RA-S), 2017/09661-0 (SP), 2018/14459-9 (MC-A) and 2018/08199-4 (MF). CAPES grant numbers: 88887.473724/2020-00 (AV-C) and 88887.185337/2018-00 (MR-J).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Eduardo Gimenes Martins for the invaluable technical support. We also thanked Dr. Roxane Maria Fontes Piazza (Butantan Institute, Brazil) for donating the 4H2 mAb.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmedt.2020.604160/full#supplementary-material
References
1. Faye O, Freire CCM, Iamarino A, Faye O, de Oliveira JVC, Diallo M, et al. Molecular evolution of zika virus during its emergence in the 20th century. PLoS Negl Trop Dis. (2014) 8:36. doi: 10.1371/journal.pntd.0002636
2. Hayes EB. Zika virus outside Africa. Emerg Infect Dis. (2009) 15:1347–50. doi: 10.3201/eid1509.090442
3. Hennessey M, Fischer M, Staples JE. Zika virus spreads to new areas - region of the Americas, May 2015-January 2016. Am J Transplant. (2016) 16:1031–4. doi: 10.1111/ajt.13743
4. Tognarelli J, Ulloa S, Villagra E, Lagos J, Aguayo C, Fasce R, et al. A report on the outbreak of Zika virus on Easter Island, South Pacific, 2014. Arch Virol. (2016) 161:665–8. doi: 10.1007/s00705-015-2695-5
5. Cugola FR, Fernandes IR, Russo FB, Freitas BC, Dias JLM, Guimarães KP, et al. The Brazilian Zika virus strain causes birth defects in experimental models. Nature. (2016) 534:267–71. doi: 10.1038/nature18296
6. Driggers RW, Ho C-Y, Korhonen EM, Kuivanen S, Jääskeläinen AJ, Smura T, et al. Zika virus infection with prolonged maternal viremia and fetal brain abnormalities. N Engl J Med. (2016) 374:2142–51. doi: 10.1056/NEJMoa1601824
7. Oliveira DBL, Almeida FJ, Durigon EL, Mendes ÉA, Braconi CT, Marchetti I, et al. Prolonged shedding of zika virus associated with congenital infection. N Engl J Med. (2016) 375:1202–4. doi: 10.1056/NEJMc1607583
8. Scott JM, Lebratti TJ, Richner JM, Jiang X, Fernandez E, Zhao H, et al. Cellular and humoral immunity protect against vaginal zika virus infection in mice. Pfeiffer JK, editor. J Virol. (2018) 92:e00038-18. doi: 10.1128/JVI.00038-18
9. Richner JM, Diamond MS. Zika virus vaccines: immune response, current status, and future challenges. Curr Opin Immunol. (2018) 53:130–6. doi: 10.1016/j.coi.2018.04.024
10. Elong Ngono A, Shresta S. Cross-reactive t cell immunity to dengue and zika viruses: new insights into vaccine development. Front Immunol. (2019) 10:1316. doi: 10.3389/fimmu.2019.01316
11. Tebas P, Roberts CC, Muthumani K, Reuschel EL, Kudchodkar SB, Zaidi FI, et al. Safety and immunogenicity of an anti–zika virus DNA vaccine — preliminary report. N Engl J Med. (2017). doi: 10.1056/NEJMoa1708120. [Epub ahead of print].
12. Abbink P, Larocca RA, Visitsunthorn K, Boyd M, De La Barrera RA, Gromowski GD, et al. Durability and correlates of vaccine protection against Zika virus in rhesus monkeys. Sci Transl Med. (2017) 9:eaao4163. doi: 10.1126/scitranslmed.aao4163
13. Larocca RA, Abbink P, Peron JPS, Zanotto PM de A, Iampietro MJ, Badamchi-Zadeh A, et al. Vaccine protection against Zika virus from Brazil. Nature. (2016) 536:474–8. doi: 10.1038/nature18952
14. Abbink P, Larocca RA, De La Barrera RA, Bricault CA, Moseley ET, Boyd M, et al. Protective efficacy of multiple vaccine platforms against Zika virus challenge in rhesus monkeys. Science. (2016) 353:1129–32. doi: 10.1126/science.aah6157
15. Berry N, Kempster S, Ham C, Jenkins A, Hall J, Page M, et al. Passive immunisation of convalescent human anti-Zika plasma protects against challenge with New World Zika virus in cynomolgus macaques. npj Vaccines. (2020) 5:86. doi: 10.1038/s41541-020-00234-y
16. Elong Ngono A, Vizcarra EA, Tang WW, Sheets N, Joo Y, Kim K, et al. Mapping and role of the CD8 + T cell response during primary zika virus infection in mice. Cell Host Microbe. (2017) 21:35–46. doi: 10.1016/j.chom.2016.12.010
17. Regla-Nava JA, Elong Ngono A, Viramontes KM, Huynh A-T, Wang Y-T, Nguyen A-VT, et al. Cross-reactive Dengue virus-specific CD8+ T cells protect against Zika virus during pregnancy. Nat Commun. (2018) 9:3042. doi: 10.1038/s41467-018-05458-0
18. Huang H, Li S, Zhang Y, Han X, Jia B, Liu H, et al. CD8 +T cell immune response in immunocompetent mice during zika virus infection. Diamond MS, editor. J Virol. (2017) 91:e00900-17. doi: 10.1128/JVI.00900-17
19. Wen J, Wang Y-T, Valentine KM, dos Santos Alves RP, Xu Z, Regla-Nava JA, et al. CD4+ T Cells Cross-Reactive with Dengue and Zika Viruses Protect against Zika Virus Infection. Cell Rep. (2020) 31:107566. doi: 10.1016/j.celrep.2020.107566
20. Hassert M, Wolf KJ, Schwetye KE, DiPaolo RJ, Brien JD, Pinto AK. CD4+T cells mediate protection against Zika associated severe disease in a mouse model of infection. Fernandez-Sesma A, editor. PLoS Pathog. (2018) 14:e1007237. doi: 10.1371/journal.ppat.1007237
21. Lucas CGOO, Kitoko JZ, Ferreira FM, Suzart VG, Papa MP, Coelho SVAA, et al. Critical role of CD4+ T cells and IFNγ signaling in antibody-mediated resistance to Zika virus infection. Nat Commun. (2018) 9:3136. doi: 10.1038/s41467-018-05519-4
22. Pardi N, Hogan MJ, Pelc RS, Muramatsu H, Andersen H, DeMaso CR, et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature. (2017) 543:248–51. doi: 10.1038/nature21428
23. Grubor-Bauk B, Wijesundara DK, Masavuli M, Abbink P, Peterson RL, Prow NA, et al. NS1 DNA vaccination protects against Zika infection through T cell–mediated immunity in immunocompetent mice. Sci Adv. (2019) 5:eaax2388. doi: 10.1126/sciadv.aax2388
24. Zhan Y, Pang Z, Du Y, Wang W, Yang Y, Wang W, et al. NS1-based DNA vaccination confers mouse protective immunity against ZIKV challenge. Infect Genet Evol. (2020) 85:104521. doi: 10.1016/j.meegid.2020.104521
25. McCracken MK, Gromowski GD, Friberg HL, Lin X, Abbink P, De La Barrera R, et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. Heise MT, editor. PLoS Pathog. (2017) 13:e1006487. doi: 10.1371/journal.ppat.1006487
26. Zellweger RM, Prestwood TR, Shresta S. Enhanced infection of liver sinusoidal endothelial cells in a mouse model of antibody-induced severe dengue disease. Cell Host Microbe. (2010) 7:128–39. doi: 10.1016/j.chom.2010.01.004
27. Dejnirattisai W, Supasa P, Wongwiwat W, Rouvinski A, Barba-Spaeth G, Duangchinda T, et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat Immunol. (2016) 17:1102–8. doi: 10.1038/ni.3515
28. Katzelnick LC, Narvaez C, Arguello S, Lopez Mercado B, Collado D, Ampie O, et al. Zika virus infection enhances future risk of severe dengue disease. Science. (2020) 369:1123–8. doi: 10.1126/science.abb6143
29. Richner JM, Himansu S, Dowd KA, Butler SL, Salazar V, Fox JM, et al. Modified mRNA vaccines protect against zika virus infection. Cell. (2017) 168:1114–25. doi: 10.1016/j.cell.2017.02.017
30. Dowd KA, Ko S-Y, Morabito KM, Yang ES, Pelc RS, DeMaso CR, et al. Rapid development of a DNA vaccine for Zika virus. Science. (2016) 354:237–40. doi: 10.1126/science.aai9137
31. Han J-F, Qiu Y, Yu J-Y, Wang H-J, Deng Y-Q, Li X-F, et al. Immunization with truncated envelope protein of Zika virus induces protective immune response in mice. Sci Rep. (2017) 7:10047. doi: 10.1038/s41598-017-10595-5
32. Araujo SC, Pereira LR, Alves RPS, Andreata-Santos R, Kanno AI, Ferreira LCS, et al. Anti-Flavivirus vaccines: review of the present situation and perspectives of subunit vaccines produced in Escherichia coli. Vaccines. (2020) 8:492. doi: 10.3390/vaccines8030492
33. Poland GA, Ovsyannikova IG, Kennedy RB. Zika vaccine development: current status. Mayo Clin Proc. (2019) 94:2572–86. doi: 10.1016/j.mayocp.2019.05.016
34. Muller D, Young PR. The flavivirus NS1 protein: molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antiviral Res. (2013) 98:192–208. doi: 10.1016/j.antiviral.2013.03.008
35. Li A, Yu J, Lu M, Ma Y, Attia Z, Shan C, et al. A Zika virus vaccine expressing premembrane-envelope-NS1 polyprotein. Nat Commun. (2018) 9:3067. doi: 10.1038/s41467-018-05276-4
36. Bailey MJ, Broecker F, Duehr J, Arumemi F, Krammer F, Palese P, et al. Antibodies elicited by an NS1-based vaccine protect mice against Zika virus. Griffin DE, editor. MBio. (2019) 10:e02861-18. doi: 10.1128/mBio.02861-18
37. Brault AC, Domi A, McDonald EM, Talmi-Frank D, McCurley N, Basu R, et al. A Zika vaccine targeting NS1 protein protects immunocompetent adult mice in a lethal challenge model. Sci Rep. (2017) 7:14769. doi: 10.1038/s41598-017-15039-8
38. Bailey MJ, Duehr J, Dulin H, Broecker F, Brown JA, Arumemi FO, et al. Human antibodies targeting Zika virus NS1 provide protection against disease in a mouse model. Nat Commun. (2018) 9:4560. doi: 10.1038/s41467-018-07008-0
39. Wessel AW, Kose N, Bombardi RG, Roy V, Chantima W, Mongkolsapaya J, et al. Antibodies targeting epitopes on the cell-surface form of NS1 protect against Zika virus infection during pregnancy. Nat Commun. (2020) 11:5278. doi: 10.1038/s41467-020-19096-y
40. Lasaro MO, Diniz MO, Reyes-Sandoval A, Ertl HC, Ferreira LCS. Anti-tumor DNA vaccines based on the expression of human papillomavirus-16 E6/E7 oncoproteins genetically fused with the glycoprotein D from herpes simplex virus-1. Microbes Infect. (2005) 7:1541–50. doi: 10.1016/j.micinf.2005.05.024
41. Porchia BFMM, Moreno ACR, Ramos RN, Diniz MO, de Andrade LHTM, Rosa DS, et al. Herpes simplex virus glycoprotein D targets a specific dendritic cell subset and improves the performance of vaccines to human papillomavirus-associated tumors. Mol Cancer Ther. (2017) 16:1922–33. doi: 10.1158/1535-7163.MCT-17-0071
42. Connolly SA, Jackson JO, Jardetzky TS, Longnecker R. Fusing structure and function: a structural view of the herpesvirus entry machinery. Nat Rev Microbiol. (2011) 9:369–81. doi: 10.1038/nrmicro2548
43. Murphy TL, Murphy KM. Slow down and survive: enigmatic immunoregulation by BTLA and HVEM. Annu Rev Immunol. (2010) 28:389–411. doi: 10.1146/annurev-immunol-030409-101202
44. Steinberg MW, Cheung TC, Ware CF. The signaling networks of the herpesvirus entry mediator (TNFRSF14) in immune regulation. Immunol Rev. (2011) 244:169–87. doi: 10.1111/j.1600-065X.2011.01064.x
45. Cheung TC, Steinberg MW, Oborne LM, Macauley MG, Fukuyama S, Sanjo H, et al. Unconventional ligand activation of herpesvirus entry mediator signals cell survival. Proc Natl Acad Sci USA. (2009) 106:6244–9. doi: 10.1073/pnas.0902115106
46. Porchia BFMM, Diniz MO, Cariri FAMO, Santana VC, Amorim JH, Balan A, et al. Purified herpes simplex type 1 glycoprotein D (gD) genetically fused with the type 16 human papillomavirus E7 oncoprotein enhances antigen-specific CD8+ T cell responses and confers protective antitumor immunity. Mol Pharm. (2011) 8:2320–30. doi: 10.1021/mp200194s
47. Diniz MO, Cariri FAMO, Aps LRMM, Ferreira LCS. Enhanced therapeutic effects conferred by an experimental DNA vaccine targeting human papillomavirus-induced tumors. Hum Gene Ther. (2013) 24:861–70. doi: 10.1089/hum.2013.102
48. Lasaro MO, Tatsis N, Hensley SE, Whitbeck JC, Lin S-W, Rux JJ, et al. Targeting of antigen to the herpesvirus entry mediator augments primary adaptive immune responses. Nat Med. (2008) 14:205–12. doi: 10.1038/nm1704
49. Lásaro MO, Alves AM, Botosso VF, Durigon EL, Ferreira LC. Antibody-inducing properties of a prototype bivalent herpes simplex virus/enterotoxigenic Escherichia coli DNA vaccine. FEMS Immunol Med Microbiol. (2003) 35:25–31. doi: 10.1111/j.1574-695X.2003.tb00645.x
50. Rosa da Silva CM, Chura-Chambi RM, Ramos Pereira L, Cordeiro Y, de Souza Ferreira LC, Morganti L. Association of high pressure and alkaline condition for solubilization of inclusion bodies and refolding of the NS1 protein from zika virus. BMC Biotechnol. (2018) 18:78. doi: 10.1186/s12896-018-0486-2
51. Andreata-Santos R, Alves RP dos S, Pereira SA, Pereira LR, Freitas CL de, Pereira SS, et al. Transcutaneous administration of dengue vaccines. Viruses. (2020) 12:514. doi: 10.3390/v12050514
52. Henriques HR, Rampazo EV, Gonçalves AJS, Vicentin ECM, Amorim JH, Panatieri RH, et al. Targeting the non-structural protein 1 from dengue virus to a dendritic cell population confers protective immunity to lethal virus challenge. PLoS Negl Trop Dis. (2013) 7:233008648–903079. doi: 10.1371/journal.pntd.0002330
53. Lundegaard C, Lamberth K, Harndahl M, Buus S, Lund O, Nielsen M. NetMHC-3.0: accurate web accessible predictions of human, mouse and monkey MHC class I affinities for peptides of length 8–11. Nucleic Acids Res. (2008) 36:W509–12. doi: 10.1093/nar/gkn202
54. Moutaftsi M, Peters B, Pasquetto V, Tscharke DC, Sidney J, Bui H-H, et al. A consensus epitope prediction approach identifies the breadth of murine T(CD8+)-cell responses to vaccinia virus. Nat Biotechnol. (2006) 24:817–9. doi: 10.1038/nbt1215
55. Alves RP dos S, Pereira L, Fabris DLN, Salvador FS, Santos RA, Zanotto PM de A, et al. Production of a recombinant Dengue virus 2 NS5 protein and the potential use as a vaccine antigen. Clin Vaccine Immunol. (2016) 23:460–9. doi: 10.1128/CVI.00081-16
56. Caine E, Jagger B, Diamond M. Animal models of zika virus infection during pregnancy. Viruses. (2018) 10:598. doi: 10.3390/v10110598
57. Miner JJ, Cao B, Govero J, Smith AM, Fernandez E, Cabrera OH, et al. Zika virus infection during pregnancy in mice causes placental damage and fetal demise. Cell. (2016) 165:1081–91. doi: 10.1016/j.cell.2016.05.008
58. Subramaniam KS, Lant S, Goodwin L, Grifoni A, Weiskopf D, Turtle L. Two is better than one: evidence for T-cell cross-protection between dengue and zika and implications on vaccine design. Front Immunol. (2020) 11:517. doi: 10.3389/fimmu.2020.00517
59. Pinheiro-Michelsen JR, Souza R da SO, Santana IVR, da Silva P de S, Mendez EC, Luiz WB, et al. Anti-dengue vaccines: from development to clinical trials. Front Immunol. (2020) 11:1252. doi: 10.3389/fimmu.2020.01252
60. Liu X, Qu L, Ye X, Yi C, Zheng X, Hao M, et al. Incorporation of NS1 and prM/M are important to confer effective protection of adenovirus-vectored Zika virus vaccine carrying E protein. npj Vaccines. (2018) 3:29. doi: 10.1038/s41541-018-0072-6
61. Stettler K, Beltramello M, Espinosa DA, Graham V, Cassotta A, Bianchi S, et al. Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science. (2016) 353:823–6. doi: 10.1126/science.aaf8505
62. Wan S-W, Lin C-F, Chen M-C, Lei H-Y, Liu H-S, Yeh T-M, et al. C-terminal region of dengue virus nonstructural protein 1 is involved in endothelial cell cross-reactivity via molecular mimicry. Am J Infect Dis. (2008) 4:85–91. doi: 10.3844/ajidsp.2008.85.91
63. Lin C-F, Lei H-Y, Shiau A-L, Liu H-S, Yeh T-M, Chen S-H, et al. Endothelial cell apoptosis induced by antibodies against dengue virus nonstructural protein 1 via production of nitric oxide. J Immunol. (2002) 169:657–64. doi: 10.4049/jimmunol.169.2.657
64. Chen M-C, Lin C-F, Lei H-Y, Lin S-C, Liu H-S, Yeh T-M, et al. Deletion of the C-terminal region of dengue virus nonstructural protein 1 (NS1) abolishes anti-NS1-mediated platelet dysfunction and bleeding tendency. J Immunol. (2009) 183:1797–803. doi: 10.4049/jimmunol.0800672
65. Reyes-Sandoval A, Ludert JE. The dual role of the antibody response against the flavivirus non-structural protein 1 (NS1) in protection and immuno-pathogenesis. Front Immunol. (2019) 10:1651. doi: 10.3389/fimmu.2019.01651
66. Amorim JH, Alves RPDS, Boscardin SB, Ferreira LCDS. The dengue virus non-structural 1 protein: risks and benefits. Virus Res. (2014) 181:53–60. doi: 10.1016/j.virusres.2014.01.001
67. Nimmerjahn F, Bruhns P, Horiuchi K, Ravetch JV. FcγRIV: a novel FcR with distinct IgG subclass specificity. Immunity. (2005) 23:41–51. doi: 10.1016/j.immuni.2005.05.010
68. Falconer DJ, Barb AW. Mouse IgG2c Fc loop residues promote greater receptor-binding affinity than mouse IgG2b or human IgG1. Permyakov EA, editor. PLoS ONE. (2018) 13:e0192123. doi: 10.1371/journal.pone.0192123
69. Saron WAA, Rathore APS, Ting L, Ooi EE, Low J, Abraham SN, et al. Flavivirus serocomplex cross-reactive immunity is protective by activating heterologous memory CD4 T cells. Sci Adv. (2018) 4:eaar4297. doi: 10.1126/sciadv.aar4297
70. Singh R, Rothman AL, Potts J, Guirakhoo F, Ennis FA, Green S. Sequential immunization with heterologous chimeric flaviviruses induces broad-spectrum cross-reactive CD8 + T cell responses. J Infect Dis. (2010) 202:223–33. doi: 10.1086/653486
71. El Sahly HM, Gorchakov R, Lai L, Natrajan MS, Patel SM, Atmar RL, et al. Clinical, virologic, and immunologic characteristics of zika virus infection in a cohort of US patients: prolonged RNA detection in whole blood. Open Forum Infect Dis. (2019) 6:ofy352. doi: 10.1093/ofid/ofy352
72. Waggoner JJ, Rouphael N, Xu Y, Natrajan M, Lai L, Patel SM, et al. Pericarditis associated with acute zika virus infection in a returning traveler. Open Forum Infect Dis. (2017) 4:ofx103. doi: 10.1093/ofid/ofx103
73. Alves dos Santos E, Fink K. Animal models for dengue and zika vaccine development. Adv Exp Med Biol. (2018) 1062:215–39. doi: 10.1007/978-981-10-8727-1_16
74. Lazear HM, Govero J, Smith AM, Platt DJ, Fernandez E, Miner JJ, et al. A mouse model of zika virus pathogenesis. Cell Host Microbe. (2016) 19:720–30. doi: 10.1016/j.chom.2016.03.010
75. MacGregor RR, Boyer JD, Ugen KE, Lacy KE, Gluckman SJ, Bagarazzi ML, et al. First human trial of a DNA-based vaccine for treatment of human immunodeficiency virus type 1 infection: safety and host response. J Infect Dis. (1998) 178:92–100. doi: 10.1086/515613
76. Low L, Mander A, McCann K, Dearnaley D, Tjelle T, Mathiesen I, et al. DNA vaccination with electroporation induces increased antibody responses in patients with prostate cancer. Hum Gene Ther. (2009) 20:1269–78. doi: 10.1089/hum.2009.067
77. Sales NS, Silva JR, Aps LRMM, Silva MO, Porchia BFMM, Ferreira LCS, et al. In vivo electroporation enhances vaccine-mediated therapeutic control of human papilloma virus-associated tumors by the activation of multifunctional and effector memory CD8+ T cells. Vaccine. (2017) 35:7240–9. doi: 10.1016/j.vaccine.2017.11.011
Keywords: Zika virus, NS1 protein, gD protein, DNA vaccine, HSV-1, flavivirus
Citation: Pereira LR, Alves RPdS, Sales NS, Andreata-Santos R, Venceslau-Carvalho AA, Pereira SS, Castro-Amarante MF, Rodrigues-Jesus MJ, Favaro MTdP, Chura-Chambi RM, Morganti L and Ferreira LCdS (2020) Enhanced Immune Responses and Protective Immunity to Zika Virus Induced by a DNA Vaccine Encoding a Chimeric NS1 Fused With Type 1 Herpes Virus gD Protein. Front. Med. Technol. 2:604160. doi: 10.3389/fmedt.2020.604160
Received: 08 September 2020; Accepted: 11 November 2020;
Published: 03 December 2020.
Edited by:
Jorge Fuentealba, University of Concepcion, ChileReviewed by:
Roberto Lins, Aggeu Magalhães Institute (IAM), BrazilAngel Alejandro Oñate, University of Concepcion, Chile
Copyright © 2020 Pereira, Alves, Sales, Andreata-Santos, Venceslau-Carvalho, Pereira, Castro-Amarante, Rodrigues-Jesus, Favaro, Chura-Chambi, Morganti and Ferreira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luís Carlos de Souza Ferreira, bGNzZkB1c3AuYnI=