
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Med. , 03 March 2025
Sec. Dermatology
Volume 12 - 2025 | https://doi.org/10.3389/fmed.2025.1531777
This article is part of the Research Topic Advancing Atopic Dermatitis Management: Translational Research and Emerging Therapies View all 4 articles
 Demet Kartal1*
Demet Kartal1* Muhammed Hanefi Dirican2
Muhammed Hanefi Dirican2 Serpil Taheri3
Serpil Taheri3 Mehmet Memiş3,4Eda Öksüm Solak1Salih Levent Cinar1Murat Borlu1
Mehmet Memiş3,4Eda Öksüm Solak1Salih Levent Cinar1Murat Borlu1Background: Genetic and epigenetic mechanisms have been shown to play a role in the pathogenesis of atopic dermatitis (AD). However, the role of genes involved in the establishment of DNA methylation has not yet been demonstrated.
Methods: A total of 15 pediatric patients with AD and 15 healthy volunteers were included in this study. The mRNA gene expression levels of eight different genes involved in the regulation of DNA methylation were examined in the blood and tissue samples.
Results: The mRNA expression levels of DNMT3A genes were significantly increased, while the mRNA expression levels of DNMT3B, TET1, and TET2 genes were statistically significantly reduced in the lesional tissue of patients compared to the control group. It was observed that the mRNA expression levels of DNMT1, DNMT3A, and TET3 genes were increased, while the mRNA expression levels of DNMT3L and TET1 genes were found to be decreased in the blood of the patients.
Conclusion: The results indicated that the DNA methylation pattern in the patients was hypermethylated, especially in the lesional tissue. The data obtained may contribute to the understanding of the epigenetic regulation of AD and aid in the development of new diagnostic and treatment options.

Graphical Abstract
Atopic dermatitis (AD), also known as atopic eczema, is a common inflammatory skin condition, and it affects 15–20% of children and 7–10% of adults in developed countries (1, 2). AD is typically developed during childhood and is characterized by intensely pruritic, recurrent eczematous lesions (1). Patients with AD also have an increased risk of developing allergic rhinitis, food allergies, and asthma, making AD the first step in the atopic march (3).
AD is a multifactorial disease, and its pathophysiology is not yet fully understood. However, existing evidence suggest that AD is associated with impaired epidermal barrier function, activation of different immune cell types, and changes in the skin microbiome due to genetic and environmental factors. These mechanisms are thought to contribute to the clinical presentation of the disease (4). A correlation between AD and over 70 different genes has been demonstrated in previous studies (5). However, these genetic correlations only account for a small portion of the disease’s variability. Mutations in these genes have also been detected in healthy individuals. Familial inheritance and the increasing frequency of AD indicate that the disease may be an acquired epigenetic regulation disorder. Although epigenetic mechanisms are significant risk factors that play a role in familial allergic diseases, there are only a few studies on this topic in the literature (5).
Epigenetics is the study of heritable changes in gene expression without changes in the DNA sequence and is influenced by environmental factors. Key mechanisms of epigenetics include CpG methylation, histone modifications, and regulatory non-coding RNAs (ncRNAs). The deregulation of these epigenetic mechanisms has been associated with various human diseases. In dermatology, skin disorders, such as AD and psoriasis, significantly affect skin integrity and disrupt epidermal differentiation. It has been shown that the epigenetic mechanisms play a role in the disruption of epidermal differentiation. Recent studies have demonstrated that the dysregulation of epigenetic mechanisms occur in many inflammatory skin diseases, such as psoriasis (6). In particular, it has been shown that this disruption occurs locally in the skin tissue. The dysregulation of multiple epigenetic mechanisms, including abnormal DNA methylation, changes in histone modifications, and miRNA expression, has been shown to play a crucial role in the pathogenic processes of these skin conditions (7). Notably, differences in epigenetic signatures have been observed not only between the skin of healthy controls and the lesional skin of psoriatic patients but also between the lesional, perilesional, and unaffected skin of individual patients (8).
DNA methylation within these epigenetic mechanisms may occur in the promoter, exon, or intergenic regions of a gene. In the literature, studies on AD have examined the methylation profiles of numerous genes, demonstrating that these profiles in patients with AD differ from those in healthy individuals. However, changes in the methylation patterns of various genes, similar to mutations in several genes linked to Alzheimer’s disease (AD), may show comparable profiles. This raises the question of whether there is an issue with the establishment of methylation profiles in the pathogenesis of AD. DNA methylation occurs through the transfer of a methyl group to the 5th carbon atom of the cytosine base by DNA methyl transferase enzymes (DNMTs) that changes the transcriptional regulation of the genes. Ten-eleven translocation (TET) factors function in the removal of the methyl group from methylated cytosine, in contrast to DNMT enzymes. In this study, we aimed to investigate the roles of DNMT and TET genes responsible for the regulation of the DNA methylation profile in the pathogenesis of AD.
After receiving approval from the Erciyes University Human Research Ethics Committee and obtaining consent from the participants, this study included patients diagnosed with AD who presented to the Faculty of Medicine, Department of Dermatology and Venereal Diseases, Erciyes University. In addition, healthy individuals matched for age and sex were also included in the study (Decision No. 2020/416). The blood and skin biopsy samples collected from the patients and healthy participants were sent to the Betül-Ziya Eren Genome and Stem Cell Center, Department of Genome, Erciyes University, for molecular studies.
The recruitment method for the patients included in the study was prospective. Patients aged 2–18 years who presented to the Faculty of Medicine, Department of Dermatology and Venereal Diseases Outpatient Clinic, Erciyes University and diagnosed with AD were included in the study. To be eligible, these patients should not have any chronic diseases other than AD and must not have received topical treatments in the previous 2 weeks or systemic treatments in the previous 4 weeks for the management of AD. The SCORAD scores of the patients were calculated, and 15 patients with scores higher than 25 were included in the study.
The control group consisted of 15 healthy individuals aged 2–18 years, without chronic or dermatological diseases. These participants were scheduled for fracture reduction or arthroscopic knee procedures at the Faculty of Medicine, Department of Orthopedics and Traumatology, Erciyes University.
In this study, tissue samples were collected from both lesional (affected) and non-lesional (unaffected) skin under standardized conditions. For lesional samples, 15 patients with atopic dermatitis (AD) underwent 3 mm punch biopsies of the skin in the popliteal fossa (the area behind the knee) after receiving local anesthesia. The procedure was conducted under sterile conditions to ensure sample integrity and to prevent infection. For non-lesional samples, 15 healthy volunteers had normal skin biopsies taken from the same anatomical region using the same method. This consistent approach allowed for a direct comparison between the lesional skin of the AD patients and the normal skin of the healthy controls. Also, 3 mm punch biopsies of the skin were taken from the popliteal fossa of 15 healthy volunteers.
Blood samples were collected from the blood vessels into EDTA tubes from both patients and healthy volunteers on the same day as when the tissue samples were obtained. This simultaneous collection ensured consistency in sample timing and minimized potential variability.
These tissue and blood samples were stored at −80°C until the study began, following the completion of RNA isolation.
Total RNA was isolated from skin biopsies and blood samples using Trizol (Thermo Fisher Scientific, MA, USA). The concentration and quality of the RNA were determined using a Nanodrop device (Shimadzu, Japan). cDNA was synthesized from the RNA samples using an Evo Script cDNA synthesis kit (Roche, Mannheim, Germany). The cDNA synthesis procedure was conducted according to the manufacturer’s protocol. A total of 3 μL of RNA was added to the reaction mix, resulting in a final concentration of 2.5 μg. Quantitative PCR (q-PCR) was then performed using the high-throughput LightCycler 480 II Real-Time PCR system (Roche, Germany, Mannheim). The cDNA was diluted with nuclease-free water at a ratio of 1:5. Syber Green Master (Roche, Germany, Mannheim, Cat No: 04707516001) was used for detection. The reaction mix was prepared according to the manufacturer’s instructions, with 5 μL of the cDNA added to the mix. The primers used in the study are listed in Supplementary Table 1. The ACTB gene was used as a housekeeping gene. For all samples, each gene was run in duplicate on the same plate. Ct values were normalized using the 2−ΔΔCt method (9, 10).
The conformity of the data to a normal distribution was evaluated using histograms, q–q plots, and the Shapiro–Wilk test, while the homogeneity of variance was evaluated using Levene’s test. Student’s t-test was performed to compare the differences in the mRNA expression levels of eight different genes found in the tissue and blood samples collected from the patient and control groups.
The relationship between quantitative data was evaluated using the Spearman correlation analysis. These data were evaluated using GraphPad Prism (version 8.0.1), and a p-value of <5% was considered statistically significant.
A total of 15 patients with AD were included in the study. Among these patients, eight (53.33%) were girls and seven (46.67%) were boys. The control group consisted of 15 healthy individuals, of which 8 (53.33%) were girls and seven (46.67%) were boys. The mean age of the patients was 8.8 years, and the mean age of the healthy individuals was 9.8 years. There was no statistically significant difference in terms of age and sex between the groups (p > 0.05). Six (40%) patients had only facial and neck involvement, while four (27%) patients had neck, facial, and flexural involvement. Five (33%) patients had only flexural involvement. Among the 15 patients, 3 (20%) were diagnosed with asthma accompanying atopic dermatitis, 1 (6.66%) had a food allergy (peanut), and 2 (13.33%) were diagnosed with allergic rhinitis. Three (20%) patients had a family history of asthma (mother and/or father), and two (13.33%) had a diagnosis of atopic dermatitis. The serum IgE levels were elevated in nine (60%) participants in the patient group, according to their age range. The mean SCORAD index of the patient group was calculated to be 36.9 (Table 1).
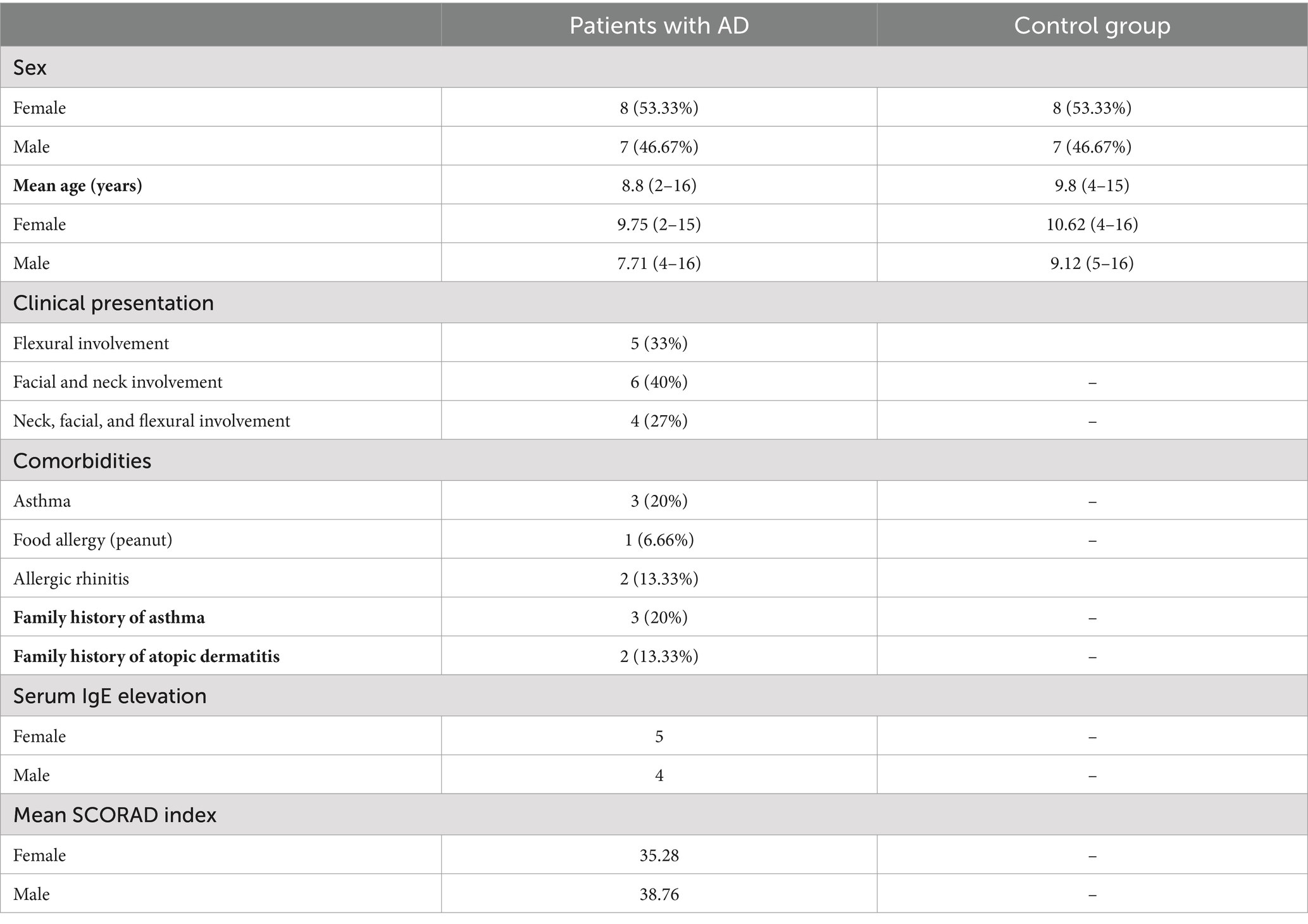
Table 1. Demographic characteristics of the patients with AD and the control group.
The mRNA expression levels of eight different genes found in the tissue and venous blood samples obtained from the AD patients and the healthy control group are summarized in Supplementary Table 1.
When DNMT1 transcript levels in the tissue and blood samples from the AD patients were compared with those from the control group, it was found that the DNMT1 levels in the tissue samples were similar between AD patients and the control group, with no significant difference. However, the DNMT1 levels were lower in the tissue samples of AD patients compared to the control samples. In contrast, a significant two-fold increase in the DNMT1 transcript levels was observed in the blood samples of AD patients compared to the control group (p = 0.008) (Figure 1A). Although there was an observed increase in the DNMT2 transcript levels in both tissue and blood samples from AD patients, the difference was not statistically significant (Figure 1B). The transcript levels of the DNMT3A gene were significantly elevated, showing a two-fold increase in both blood (p = 0.001) and tissue samples (p = 0.001) of AD patients compared to the control group (Figure 1C). In contrast, the DNMT3B transcript levels in the tissue samples of AD patients exhibited a four-fold reduction compared to the control group (p = 0.0030); however, no significant difference was observed in the blood samples (Figure 1D). The DNMT3L gene showed a three-fold increase in the transcript levels exclusively in the blood samples of AD patients compared to the control group (p = 0.0231) (Figure 1E).
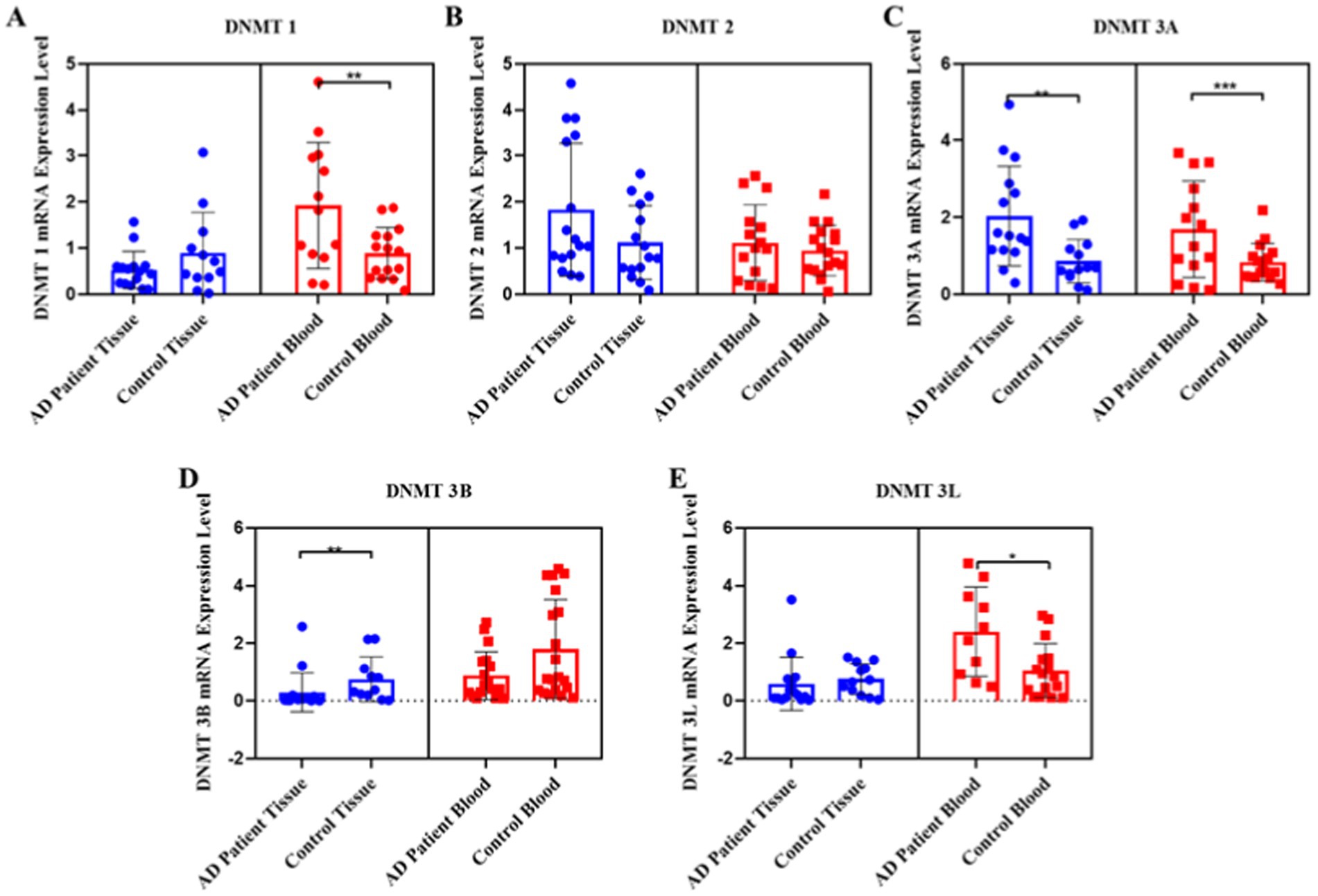
Figure 1. Pooled demonstration of the transcript levels of DNMT1, DNMT2, DNMT3A, DNMT3B, and DNMT3L genes in the blood and tissue samples obtained from AD patients and the healthy control group. Graphs for DNMT1, DNMT2, DNMT3A, DNMT3B, and DNMT3L are shown in panels (A–E), respectively.
Regarding the TET gene family, the TET1 transcript levels decreased two-fold in both blood (p = 0.015) and tissue samples (p = 0.013) of AD patients compared to the control group (Figure 2A). A two-fold decrease in the TET2 transcript levels was specifically observed in the skin tissue samples of AD patients compared to the control group (p = 0.008) (Figure 2B). In contrast, the TET3 transcript levels increased approximately two-fold in the blood samples of AD patients compared to the control group (p = 0.0093), with no significant difference observed in the tissue samples (Figure 2C).
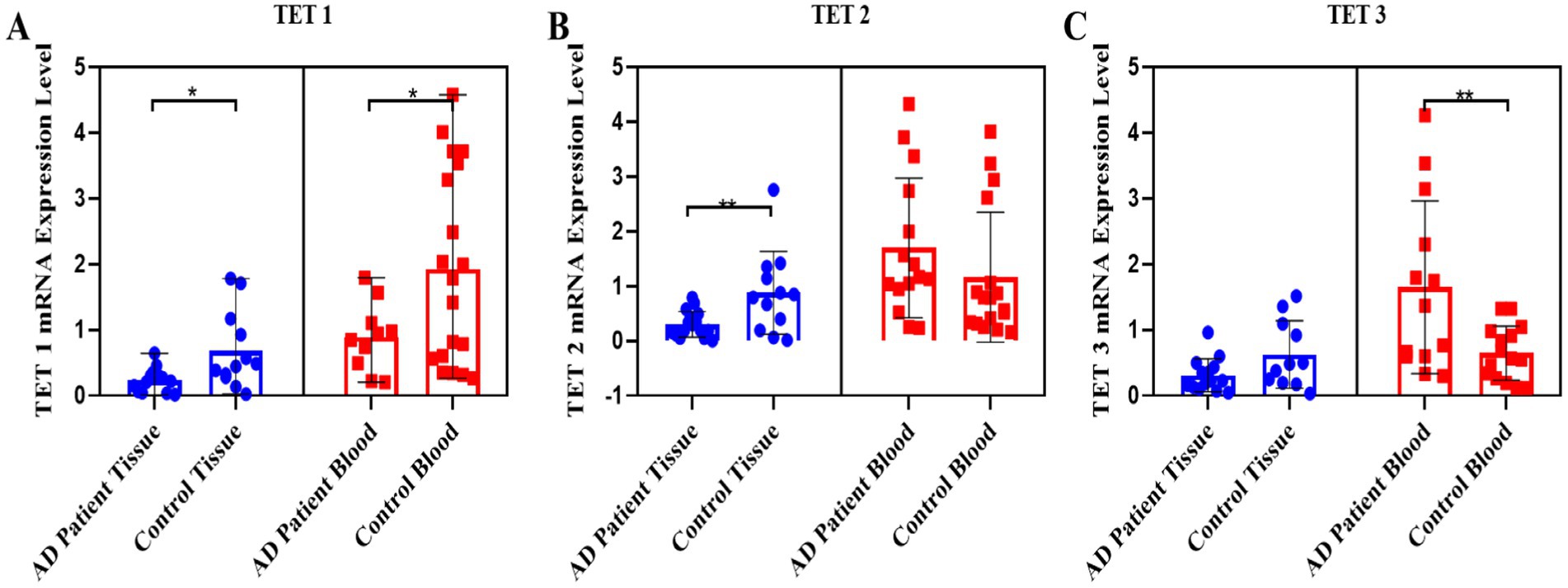
Figure 2. Pooled demonstration of the transcript levels of the TET1, TET2, and TET3 genes in the blood and tissue samples obtained from AD patients and the healthy control group. Graphs for TET1, TET2, and TET3 are shown in panels (A–C), respectively.
In addition, the potential relationship between the SCORAD scores of the patients and the expression levels of DNMT1, DNMT2, DNMT3A, DNMT3B, DNMT3L, TET1, TET2, and TET3 genes in the blood samples and lesional tissues was evaluated using the correlation analysis. However, no significant positive or negative relationship was found.
Atopic dermatitis (AD) is a chronic inflammatory skin disease that affects people globally. This condition not only has a significant impact on the quality of life for those affected but also places a considerable burden on healthcare systems and economies (11). Understanding the prevalence of AD in both developed and developing countries is essential for effective public health strategies and healthcare planning. While higher prevalence rates have been reported in developed countries, developing countries face challenges including limited access to healthcare services in managing AD. The increased prevalence in developed countries can be attributed to better healthcare infrastructure and advanced diagnostic capabilities (12–13). Environmental factors play a significant role in the development and exacerbation of AD. Factors such as air pollution, household dampness, mold, proximity to green spaces, and dietary habits can influence the development of AD. The high prevalence of AD in developing countries is associated with the impact of these environmental and lifestyle factors (14). The complex pathogenesis of AD may be influenced by interactions between genetic predisposition, immune dysregulation, and environmental stress factors (15). In addition, exposure to allergens, especially inhaled allergens such as house dust mites, can increase the incidence of AD (16–18). Stress also has been identified as an important factor in both the development and exacerbation of AD (19).
Epigenetic modifications refer to reversible changes in gene expression without alterations to the DNA sequence and occur at all stages of development or in response to environmental factors (7). Studies have previously shown that AD has a hereditary component and that epigenetic mechanisms play a role in its pathogenesis (20).
There are three main epigenetic mechanisms: DNA methylation at the DNA level, histone modifications affecting histone tails, and small RNAs (smRNAs) and long non-coding RNAs (lncRNAs) at the RNA levels. “Epigenome” refers to the global profile of gene expression, and epigenetic mechanisms can alter the epigenome.
DNA methylation, one of the epigenetic mechanisms, involves the transfer of a methyl group to the 5th carbon atom of cytosine by DNMTs and generally mediates the suppression of gene mRNA expression. When studies in the literature on AD-related DNA methylation were reviewed, it was found that there were changes in the methylation profiles of many genes associated with AD. However, changes in the methylation profiles of different genes alone cannot account for the cause of this disease, just as mutations alone cannot provide a complete explanation (5, 21–23). Our hypothesis suggests that these changes stem from a global shift in the genome’s methylation profile. The literature has yet to clarify how changes in the expression of genes that establish and remove the global DNA methylation profile play a role in the pathogenesis of AD. DNMT and TET enzymes play a role in the establishment of the methylation profile of the genome. Consequently, changes in the expression of these genes could lead to changes in the methylation profiles of many genes controlled by DNA methylation, which in turn may affect their expression levels (5, 21–23).
Our primary aim in this study was to analyze the transcript profiles of DNMT and TET genes and investigate how these genes contribute to AD pathogenesis. These genes are crucial in regulating global DNA methylation changes in the skin and blood of AD patients. Previous research has examined the methylation profiles of various individual genes, but these studies have not provided a comprehensive view of hyper- or hypo-methylation states in the context of AD. This suggests that focusing solely on the methylation status of individual genes or global methylation changes may not fully elucidate the complex etiology of AD.
We hypothesized that the discrepancy arises because global DNA methylation patterns are influenced by environmental factors, which can lead to alterations in the methylation pattern. DNMT and TET genes that are responsible for maintaining and modifying this global methylation map may therefore play a significant role in the development of AD. Fundamental changes in epigenetic mechanisms often begin with alterations in the transcript levels of these genes. Although changes in transcript levels may not always correspond directly to protein levels, they can indicate shifts in underlying epigenetic regulation. Understanding these changes could provide valuable insights into how epigenetic mechanisms contribute to AD pathogenesis and highlight potential targets for therapeutic intervention.
In our study, we found that the expression levels of DNMT1 in the peripheral blood samples of AD patients were higher compared to those of the control group. This finding contrasts with a study performed by Nakamura et al. (24), which served as the starting point for our research. This study focused solely on blood samples of AD patients and demonstrated that DNMT1 transcript levels in peripheral blood mononuclear cells were significantly lower in AD patients compared to the control group (24).
When the literature was reviewed, various studies showed changes in the DNA methylation profile of different genes, generally in the promoter regions, in AD patients. However, the relationship between changes in the expression levels of genes involved in DNA methylation and the pathogenesis of AD has not yet been studied. DNMT1 is a general DNA methyltransferase that, after establishing DNA methylation patterns, transfers methylation to the next generation by copying the methylation onto the newly formed strand based on the old DNA strand. DNMT3A and DNMT3B are responsible for establishing new DNA methylation patterns through de novo DNA methylation activities by methylating unmethylated CpG dinucleotides during early embryonic stages and germ cell differentiation (25, 26).
In our study, we found that the DNMT1 transcript level increased two-fold in the blood samples of AD patients compared to the control group. DNMT1 is generally responsible for maintaining DNA methylation, and its increase in blood may indicate a shift toward hypermethylation in the genome of AD patients. DNMT3A and DNMT3B are responsible for de novo methylation profile changes after mitosis or in response to environmental factors. It was found that the DNMT3A transcript level increased two-fold both in the skin and blood samples of AD patients, whereas the transcript level of DNMT3B decreased four-fold in both skin tissue and blood samples of AD patients compared to the control group. There is limited information in the literature regarding how DNMT3A and DNMT3B genes establish a de novo methylation pattern. One study suggested that DNMT3A and DNMT3B function differently. It was shown that an increased DNMT3A transcript level is associated with triggering cellular aging, while a decreased DNMT3B transcript level indicates increased DNA damage and disruption of DNA repair mechanisms (27). In light of these data available in the literature, our results indicate that cellular aging may be triggered by increased expression of the DNMT3A gene in AD patients. Additionally, the decrease in DNMT3B expression could indicate the presence of increased DNA damage and a possible disruption in the repair mechanisms of the damage and the DNA methylation pattern is changed. However, other studies, especially those that assess the protein levels, are required to confirm these data completely. When the data we obtained were evaluated alongside the findings in the literature, the main point we aimed to highlight in this study was that changes in epigenetic mechanisms in AD patients are influenced by environmental factors (28), and these altered epigenetic mechanisms affect the transcript levels of genes that establish the global DNA methylation map. As a result, DNMTs and TETs, whose transcript levels change, may affect the global methylation map from the outset.
The DNMT2 enzyme is an RNA methyltransferase that plays a crucial role in tRNA and mRNA methylation. It has been shown that the DNMT2 enzyme is essential for the recognition of DNA damage, DNA recombination, and mutation repair. Furthermore, global DNA methylation cannot occur in embryonic stem cells in the absence of this enzyme (29, 30). In our study, no significant differences in the DNMT2 levels in blood or skin tissue samples were observed between AD patients and the control group. DNMT1 and DNMT2 perform similar roles on different nucleic acids. DNMT1 is directly involved in DNA methylation, whereas DNMT2 is more closely associated with methylating tRNA and other small RNA molecules and does not directly impact DNA methylation. In our study, while DNMT1 exhibited high expression levels in the blood of AD patients, no significant differences were observed in DNMT2. This lack of differentiation may suggest that DNA methylation, rather than RNA methylation, plays a more significant role in AD pathogenesis. To confirm this hypothesis, more detailed methylation screenings should be conducted to provide further insights. In our study, we measured the transcript levels of the TET genes, which are responsible for the removal of DNA methylation, in the skin tissue and blood samples of AD patients and compared them those of healthy controls. The TET1 transcript levels were detected to be decreased two-fold in the blood and skin tissue of AD patients compared to the control group. The TET2 transcript levels were also decreased two-fold in the skin tissue of AD patients compared to the control group, although the difference was not significant; however, they were increased in the blood samples. It was shown in the literature that a decrease in TET2 expression led to reduced potential for Treg cell activation (31). The TET3 transcript levels were found to be increased by approximately two-fold in the blood of AD patients.
There are no other studies in the literature that have evaluated the roles of DNMT and TET transcript levels in the pathogenesis of AD. However, the roles of DNMTs and TETs transcript levels in disease pathogenesis have been shown in other conditions, particularly in the context of the atopic march (32–41).
It was demonstrated in a study that particulate matter (PM) accelerates the aging process in human keratinocytes and mouse skin tissue by affecting epigenetic inheritance (42). This finding supports the data that the altered DNMT3A and 3B transcript profiles in AD patients accelerate the aging process.
It was also demonstrated that cells treated with PM 2.5 show cellular aging characteristics, leading to a decrease in DNMT1 and DNMT3B expression levels and an increase in TET1 transcript levels, and the genome shifts to a hypomethylated state with allergen exposure (42).
When we compared our data with the findings in the literature, we observed that changes in the transcript levels of DNMT and TET genes, especially DNMT3A and DNMT3B, could trigger DNA damage and accelerate cellular aging (26). However, we believe that epigenetic destabilization plays an important role in the onset of AD, and our main point in the study is that changes in the transcript levels of DNMT and TET genes can change the global methylation map as well.
The increase in DNMT3A expression levels observed in the lesional tissues of the patients with AD and the elevated levels of DNMT1, DNMT3A, DNMT3L, and TET3 in the blood, compared to the controls, may be attributed to cellular stress resulting from inflammation associated with AD. This inflammatory process could potentially trigger epigenetic modifications, thereby inducing changes in various transcription factors and modulating the expression of genes encoding DNMT and TET enzymes. To better understand these findings, it would be beneficial to assess the expression levels of these genes in non-lesional biopsy samples, which would provide insights into the effects of the inflammatory process. This represents a significant limitation of our study.
Consistent with the data found in the literature, the totality of our available data indicates that changes in DNMT and TET transcript levels, especially DNMT3A and DNMT3B, trigger DNA damage, accelerate cellular aging, and lead to de novo methylation profile changes in AD. According to the results we obtained, we believe that epigenetic destabilization plays a role in the onset of AD. The changes in DNMT and TET transcript levels in AD patients were significantly higher than in the control group. Our findings indicate that in AD disease both the methylation profile of individual genes and the global genome methylation pattern are altered. Changes in the global DNA methylation pattern can activate some genes and repress others. In other words, it alters the epigenome and may lead to genomic instability. The data we obtained are preliminary and indicate that the DNA methylation pattern changes in AD, especially under the influence of skin tissue. These findings may contribute to the diagnosis of AD patients and the development of new treatment strategies. This should be supported by future studies involving comprehensive transcriptome and protein analyses. The key point at this stage is that revealing the genome’s tendency toward a hypermethylated or hypomethylated state can provide opportunities for either negative or positive modulation of methyl group donor pathways, which may help in controlling AD symptoms.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving humans were approved by the Erciyes University Ethics Committee. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
DK: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. MD: Conceptualization, Data curation, Investigation, Resources, Writing – review & editing. ST: Formal analysis, Investigation, Methodology, Resources, Writing – review & editing. MM: Data curation, Formal analysis, Investigation, Software, Writing – original draft. ES: Supervision, Writing – review & editing. SC: Supervision, Writing – review & editing. MB: Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Erciyes University Scientific Research Projects Unit (TTU-2021-11212).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2025.1531777/full#supplementary-material
1. Weidinger, S, Beck, LA, Bieber, T, Kabashima, K, and Irvine, AD. Atopic dermatitis. Nat Rev Dis Prim. (2018) 4:1–20. doi: 10.1038/s41572-018-0001-z
2. Silverberg, JI. Atopic dermatitis in adults. Med Clin North Am. (2020) 104:157–76. doi: 10.1016/j.mcna.2019.08.009
3. Dharmage, SC, Lowe, AJ, Matheson, MC, Burgess, JA, Allen, KJ, and Abramson, MJ. Atopic dermatitis and the atopic march revisited. Allergy. (2014) 69:17–27. doi: 10.1111/all.12268
4. Kim, J, Kim, BE, and Leung, DYM. Pathophysiology of atopic dermatitis: clinical implications. Allergy Asthma Proc. (2019) 40:84–92. doi: 10.2500/aap.2019.40.4202
5. Nedoszytko, B, Reszka, E, Gutowska-Owsiak, D, Trzeciak, M, Lange, M, Jarczak, J, et al. Genetic and epigenetic aspects of atopic dermatitis. Int J Mol Sci. (2020) 21:1–18. doi: 10.3390/ijms21186484
6. Leśniak, W. Epigenetic regulation of epidermal differentiation. Epigenomes. (2021) 5:1. doi: 10.3390/epigenomes5010001
7. Moltrasio, C, Romagnuolo, M, and Marzano, AV. Epigenetic mechanisms of epidermal differentiation. Int J Mol Sci. (2022) 23:4874. doi: 10.3390/IJMS23094874
8. Mehmetbeyoglu, E, Kianmehr, L, Borlu, M, Yilmaz, Z, Kılıc, SB, Rajabi-Maham, H, et al. Decrease in RNase HII and accumulation of lncRNAs/DNA hybrids: a causal implication in psoriasis? Biomol Ther. (2022) 12:368. doi: 10.3390/biom12030368
9. Karaca, Z, Taheri, S, Firat, ST, Borlu, M, Zararsiz, G, Mehmetbeyoglu, E, et al. Molecular skin changes in Cushing syndrome and the effects of treatment. J Endocrinol Investig. (2020) 44:153–63. doi: 10.1007/s40618-020-01285-7
10. Livak, KJ, and Schmittgen, TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. (2001) 25:402–8. doi: 10.1006/meth.2001.1262
11. Laughter, MR, Maymone, MBC, Mashayekhi, S, Arents, BWM, Karimkhani, C, Langan, SM, et al. The global burden of atopic dermatitis: lessons from the global burden of disease study 1990-2017. Br J Dermatol. (2021) 184:304–9. doi: 10.1111/bjd.19580
12. Ilic, I, Stojkovic, A, Velickovic, V, Zivanovic Macuzic, I, and Ilic, M. Atopic dermatitis in children under 5: prevalence trends in central, eastern, and western Europe. Children. (2023) 10:1275. doi: 10.3390/children10081275
13. Silverberg, JI, Barbarot, S, Gadkari, A, Simpson, EL, Weidinger, S, Mina-Osorio, P, et al. Atopic dermatitis in the pediatric population. Ann Allergy Asthma Immunol. (2021) 126:417–428.e2. doi: 10.1016/j.anai.2020.12.020
14. Cho, Y-T, Hsieh, W, Chan, TS, Tang, C, and Chu, C. Prevalence of baseline comorbidities in patients with atopic dermatitis: a population-based cohort study in Taiwan. Jaad Int. (2020) 1:50–8. doi: 10.1016/j.jdin.2020.05.002
15. Son, C-G. Demand survey for application of environmental therapy for atopic dermatitis. J Korean Med. (2014) 35:34–40. doi: 10.13048/jkm.14020
16. Al, S, Asilsoy, S, Anal, Ö, Tezcan, D, Şirin, S, Atakul, G, et al. Unexpected cat allergy in infants with persistent atopic dermatitis. J Dr Behçet Uz Child Hosp. (2021) 11:210–4. doi: 10.5222/buchd.2021.26213
17. Tubalawony, SL, Tuasela, D, and Siahaya, GC. Factors associated with the incidence of atopic dermatitis in school-age children in the working area of the Waai health center in 2022. J Ilm Ilmu Keperawatan Indones. (2022) 12:59–67. doi: 10.33221/jiiki.v12i02.1909
18. Baek, J-H, Sung-Hwa, H, Son, D, Lee, J-R, Roh, JY, and Kwon, H-J. Analysis of the prevalence of and risk factors for atopic dermatitis using an ISAAC questionnaire in 8,750 Korean children. Int Arch Allergy Immunol. (2013) 162:79–85. doi: 10.1159/000351403
19. Suna, S. Pollutants and climatic conditions related to the childhood asthma and atopic dermatitis. World J Biol Pharm Heal Sci. (2022) 9:027–38. doi: 10.30574/wjbphs.2022.9.1.0025
20. Girolomoni, G, de Bruin-Weller, M, Aoki, V, Kabashima, K, Deleuran, M, Puig, L, et al. Nomenclature and clinical phenotypes of atopic dermatitis. Ther Adv Chronic Dis. (2021) 12:204062232110029. doi: 10.1177/20406223211002979
21. Barnes, KC. An update on the genetics of atopic dermatitis: scratching the surface in 2009. J Allergy Clin Immunol. (2010) 125:16–29.e11. doi: 10.1016/j.jaci.2009.11.008
22. Mervis, JS, and McGee, JS. DNA methylation and inflammatory skin diseases. Arch Dermatol Res. (2020) 312:461–6. doi: 10.1007/s00403-019-02005-9
23. Feng, Y, Arvey, A, Chinen, T, Van Der Veeken, J, Gasteiger, G, and Rudensky, AY. Control of the inheritance of regulatory T cell identity by a cis element in the Foxp3 locus. Cell. (2014) 158:749–63. doi: 10.1016/j.cell.2014.07.031
24. Nakamura, T, Sekigawa, I, Ogasawara, H, Mitsuishi, K, Hira, K, Ikeda, S, et al. Expression of DNMT-1 in patients with atopic dermatitis. Arch Dermatol Res. (2006) 298:253–6. doi: 10.1007/s00403-006-0682-0
25. Jeltsch, A, and Jurkowska, RZ. New concepts in DNA methylation. Trends Biochem Sci. (2014) 39:310–8. doi: 10.1016/j.tibs.2014.05.002
26. Hamidi, T, Singh, AK, and Chen, T. Genetic alterations of DNA methylation machinery in human diseases. Epigenomics. (2015) 7:247–65. doi: 10.2217/epi.14.80
27. Camero, S, Vitali, G, Pontecorvi, P, Ceccarelli, S, Anastasiadou, E, Cicchetti, F, et al. DNMT3A and DNMT3B targeting as an effective radiosensitizing strategy in embryonal rhabdomyosarcoma. Cells. (2021) 10:2956. doi: 10.3390/cells10112956
28. Asher, MI, Montefort, S, Björkstén, B, Lai, CK, Strachan, DP, Weiland, SK, et al. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC phases one and three repeat multicountry cross-sectional surveys. Lancet (London, England). (2006) 368:733–43. doi: 10.1016/S0140-6736(06)69283-0
29. Hermann, A, Schmitt, S, and Jeltsch, A. The human Dnmt2 has residual DNA-(cytosine-C5) methyltransferase activity. J Biol Chem. (2003) 278:31717–21. doi: 10.1074/jbc.M305448200
30. Tajima, S, Suetake, I, Takeshita, K, Nakagawa, A, and Kimura, H. Domain structure of the Dnmt1, Dnmt3a, and Dnmt3b DNA methyltransferases. Adv Exp Med Biol. (2016) 945:63–86. doi: 10.1007/978-3-319-43624-1_4
31. Tan, L, Qiu, T, Xiang, R, Cao, C, Deng, Y, Tao, Z, et al. Down-regulation of Tet2 is associated with Foxp3 TSDR hypermethylation in regulatory T cell of allergic rhinitis. Life Sci. (2020) 241:117101. doi: 10.1016/J.LFS.2019.117101
32. Li, H, Lu, T, Sun, W, Ma, R, Zhong, H, Wei, Y, et al. Ten-eleven translocation (TET) enzymes modulate the activation of dendritic cells in allergic rhinitis. Front Immunol. (2019) 10:2271. doi: 10.3389/fimmu.2019.02271
33. Hashimoto, H, Liu, Y, Upadhyay, AK, Chang, Y, Howerton, SB, Vertino, PM, et al. Recognition and potential mechanisms for replication and erasure of cytosine hydroxymethylation. Nucleic Acids Res. (2012) 40:4841–9. doi: 10.1093/nar/gks155
34. Vercelli, D. Discovering susceptibility genes for asthma and allergy. Nat Rev Immunol. (2008) 8:169–82. doi: 10.1038/nri2257
35. Brand, S, Kesper, DA, Teich, R, Kilic-Niebergall, E, Pinkenburg, O, Bothur, E, et al. DNA methylation of TH1/TH2 cytokine genes affects sensitization and progress of experimental asthma. J Allergy Clin Immunol. (2012) 129:1602–1610.e6. doi: 10.1016/j.jaci.2011.12.963
36. Akinbami, LJ, Moorman, JE, Garbe, PL, and Sondik, EJ. Status of childhood asthma in the United States, 1980-2007. Pediatrics. (2009) 123:S131–45. doi: 10.1542/PEDS.2008-2233C
37. Cheng, RY, Shang, Y, Limjunyawong, N, Dao, T, Das, S, Rabold, R, et al. Alterations of the lung methylome in allergic airway hyper-responsiveness. Environ Mol Mutagen. (2014) 55:244–55. doi: 10.1002/em.21851
38. Burleson, JD, Siniard, D, Yadagiri, VK, Chen, X, Weirauch, MT, Ruff, BP, et al. TET1 contributes to allergic airway inflammation and regulates interferon and aryl hydrocarbon receptor signaling pathways in bronchial epithelial cells. Sci Rep. (2019) 9:7361. doi: 10.1038/s41598-019-43767-6
39. Somineni, HK, Zhang, X, Biagini Myers, JM, Kovacic, MB, Ulm, A, Jurcak, N, et al. TET1 methylation is associated with childhood asthma and traffic-related air pollution. J Allergy Clin Immunol. (2016) 137:797–805.e5. doi: 10.1016/j.jaci.2015.10.021
40. Yeung, BHY, Huang, J, An, SS, Solway, J, Mitzner, W, and Tang, WY. Role of isocitrate dehydrogenase 2 on DNA hydroxymethylation in human airway smooth muscle cells. Am J Respir Cell Mol Biol. (2020) 63:36–45. doi: 10.1165/rcmb.2019-0323OC
41. Li, H, Ryu, MH, Rider, CF, Tse, W, Clifford, RL, Aristizabal, MJ, et al. Predominant DNMT and TET mediate effects of allergen on the human bronchial epithelium in a controlled air pollution exposure study. J Allergy Clin Immunol. (2021) 147:1671–82. doi: 10.1016/j.jaci.2020.08.044
Keywords: atopic dermatitis, DNA methylation, TET1-2 and DNMT gene expression, epigenetic, pediatric atopic dermatitis
Citation: Kartal D, Dirican MH, Taheri S, Memiş M, Solak EÖ, Cinar SL and Borlu M (2025) Regulation of DNA methylation in lesional tissue of children with atopic dermatitis. Front. Med. 12:1531777. doi: 10.3389/fmed.2025.1531777
Edited by:
Luis F. Santamaria-Babí, University of Barcelona, SpainReviewed by:
Wayne Robert Thomas, University of Western Australia, AustraliaCopyright © 2025 Kartal, Dirican, Taheri, Memiş, Solak, Cinar and Borlu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Demet Kartal, ZGVtZXRrYXJ0YWxAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.