 Sandra Freitag-Wolf1†
Sandra Freitag-Wolf1† Jonas C. Schupp2,3,4†
Jonas C. Schupp2,3,4† Björn C. Frye2Annegret Fischer5Raihanatul Anwar1Robert Kieszko6Violeta Mihailović-Vučinić7Janusz Milanowski6
Björn C. Frye2Annegret Fischer5Raihanatul Anwar1Robert Kieszko6Violeta Mihailović-Vučinić7Janusz Milanowski6 Dragana Jovanovic7
Dragana Jovanovic7 Gernot Zissel2
Gernot Zissel2 Elena Bargagli8
Elena Bargagli8 Paola Rottoli8
Paola Rottoli8 Dragos Bumbacea9René Jonkers10Ling-Pei Ho11
Dragos Bumbacea9René Jonkers10Ling-Pei Ho11 Karoline I. Gaede12Anna Dubaniewicz13Ben G. Marshall14
Karoline I. Gaede12Anna Dubaniewicz13Ben G. Marshall14 Andreas Günther15
Andreas Günther15 Martin Petrek16
Martin Petrek16 Michael P. Keane17Sigridur O. Haraldsdottir18
Michael P. Keane17Sigridur O. Haraldsdottir18 Francesco Bonella19Christian Grah20Tatjana Peroš-Golubičić21Zamir Kadija22Stefan Pabst23Christian Grohé24János Strausz25Martina Safrankova26Ann Millar27Jiří Homolka28Wim A. Wuyts29Lisa G. Spencer30Michael Pfeifer31Dominique Valeyre32Venerino Poletti33Hubertus Wirtz34
Francesco Bonella19Christian Grah20Tatjana Peroš-Golubičić21Zamir Kadija22Stefan Pabst23Christian Grohé24János Strausz25Martina Safrankova26Ann Millar27Jiří Homolka28Wim A. Wuyts29Lisa G. Spencer30Michael Pfeifer31Dominique Valeyre32Venerino Poletti33Hubertus Wirtz34 Antje Prasse2,3Stefan Schreiber5,35Astrid Dempfle1‡
Antje Prasse2,3Stefan Schreiber5,35Astrid Dempfle1‡ Joachim Müller-Quernheim2*‡
Joachim Müller-Quernheim2*‡- 1Institute of Medical Informatics and Statistics, Kiel University, Kiel, Germany
- 2Department of Pneumology, Faculty of Medicine, University Medical Centre, Freiburg, Germany
- 3Department of Respiratory Medicine, Hannover Medical School, German Center for Lung Research (DZL), Hannover, Germany
- 4Biomedical Research in End-Stage and Obstructive Lung Disease (BREATH), Hannover Medical School (MHH), German Center for Lung Research (DZL), Hannover, Germany
- 5Institute of Clinical Molecular Biology, Kiel University, Kiel, Germany
- 6Department of Pneumonology, Oncology and Allergology, Medical University of Lublin, Lublin, Poland
- 7Department of Pneumology, University Hospital, Belgrade, Serbia
- 8Respiratory Diseases and Lung Transplant Unit, University Hospital, Siena, Italy
- 9Department of Cardio-Thoracic Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania
- 10Pulmonology Department, Academic Medical Center Amsterdam, Amsterdam, Netherlands
- 11Oxford Sarcoidosis Service, Oxford University Hospitals NHS Foundation Trust, Churchill Hospital, Oxford, United Kingdom
- 12Medical Hospital, Research Center Borstel, Borstel, Germany
- 13Department of Pulmonology, Medical University of Gdansk, Gdansk, Poland
- 14Department of Respiratory Medicine, University Hospital, Southampton, United Kingdom
- 15Department of Pneumology and Intensive Care, University Hospital, Giessen, Germany
- 16Faculty of Medicine and Dentistry, Palacký University and University Hospital Olomouc, Olomouc, Czechia
- 17Division of Pulmonary and Critical Care Medicine, University College Dublin and St Vincent’s University Hospital, Dublin, Ireland
- 18Landspitali, University Hospital, Reykjavik, Iceland
- 19Ruhrlandklinik, Westdeutsches Lungenzentrum am Universitätsklinikum Essen, Universitätsklinik Essen, Essen, Germany
- 20Hospital Berlin-Havelhöhe, Berlin, Germany
- 21Pulmonary Department, University Hospital Jordanovac, Zagreb, Croatia
- 22Foundation IRCCS Policlinico San Matteo - Pulmonology Unit, Pavia, Italy
- 23Department of Pneumology, University Hospital, Bonn, Germany
- 24Evangelische Lungenklinik Berlin, Berlin, Germany
- 25National Koranyi Institute, Budapest, Hungary
- 26Thomayer Hospital and 1st Faculty of Medicine, Charles University, Praha, Czechia
- 27Pulmonary Department, University Hospital, Bristol, United Kingdom
- 28Prague General Hospital, Charles University, Prague, Czechia
- 29Laboratory of Respiratory Diseases and Thoracic Surgery (BREATHE), University Hospital, Leuven, Belgium
- 30Liverpool Interstitial Lung Disease Service, Aintree Chest Centre, Liverpool University Hospitals NHS FT, Liverpool, United Kingdom
- 31Department of Pneumology, University Hospital Regensburg, Regensburg, Germany
- 32Groupe Hospitalier Avicenne-Jean Verdier-René Muret, Service de Pneumologie, Bobigny, France
- 33Pulmonary Unit, Department of Thoracic Diseases, Azienda USL Romagna, GB Morgagni-L-Pierantoni Hospital, Forlì, Italy
- 34Department of Pneumology, University Hospital Leipzig, Leipzig, Germany
- 35Department of Internal Medicine I, University Hospital Schleswig-Holstein, Kiel, Germany
Introduction: Sarcoidosis is a highly variable disease in terms of organ involvement, type of onset and course. Associations of genetic polymorphisms with sarcoidosis phenotypes have been observed and suggest genetic signatures.
Methods: After obtaining a positive vote of the competent ethics committee we genotyped 1909 patients of the deeply phenotyped Genetic-Phenotype Relationship in Sarcoidosis (GenPhenReSa) cohort of 31 European centers in 12 countries with 116 potentially disease-relevant single-nucleotide polymorphisms (SNPs). Using a meta-analysis, we investigated the association of relevant phenotypes (acute vs. sub-acute onset, phenotypes of organ involvement, specific organ involvements, and specific symptoms) with genetic markers. Subgroups were built on the basis of geographical, clinical and hospital provision considerations.
Results: In the meta-analysis of the full cohort, there was no significant genetic association with any considered phenotype after correcting for multiple testing. In the largest sub-cohort (Serbia), we confirmed the known association of acute onset with TNF and reported a new association of acute onset an HLA polymorphism. Multi-locus models with sets of three SNPs in different genes showed strong associations with the acute onset phenotype in Serbia and Lublin (Poland) demonstrating potential region-specific genetic links with clinical features, including recently described phenotypes of organ involvement.
Discussion: The observed associations between genetic variants and sarcoidosis phenotypes in subgroups suggest that gene–environment-interactions may influence the clinical phenotype. In addition, we show that two different sets of genetic variants are permissive for the same phenotype of acute disease only in two geographic subcohorts pointing to interactions of genetic signatures with different local environmental factors. Our results represent an important step towards understanding the genetic architecture of sarcoidosis.
Introduction
Sarcoidosis is a clinically heterogeneous disease, particularly regarding the affected organs, the type of onset and the course of the disease (1) and this clinical heterogeneity needs to be addressed therapeutically (2). Sarcoidosis is a complex disorder with a strong genetic background (3–5), for which a number of genetic risk factors have been identified by genome-wide association and candidate gene studies, e.g., various HLA-haplotypes (6), BTNL2 (7), ANXA11 (8), and IL23R (9), some of which confer susceptibility to certain sarcoidosis phenotypes (10). Sarcoidosis is, therefore, pathogenetically regarded as a disease resulting from a complex interplay of an unknown environmental causative agent or agents with a genetically permissive subject. Thus, genetic factors might have a potential to distinguish sarcoidosis phenotypes and should, in general, be most suitable as prospective biomarkers.
Many of the reported genetic associations with sarcoidosis or with a specific clinical phenotype of sarcoidosis could not be replicated in independent studies or showed contradictory results in a replication cohort (4). As an example, the combination of HLA-DRB1*01 and TNFA2 predisposes Europeans to favorable prognosis, but is associated with poor prognosis and cardiac manifestation of sarcoidosis in a Japanese cohort (11, 12). Similarly, two ANXA11 variants are associated with sarcoidosis in African Americans, but not in European Americans (13). In some studies, insufficient statistical power due to small sample size could be a reason for this lack of replication. However, in other cases, this might reflect the fact that different causative agents encounter a different genetic architecture to cause sarcoidosis.
The aim of this study was to investigate whether genetic variants known or suggested to be associated with sarcoidosis or implicated in sarcoidosis pathogenesis, are associated with clinical phenotypes of sarcoidosis, including type of onset, phenotypes of organ involvement, specific organ involvements itself, or symptoms and to evaluate whether these associations are consistent throughout multiple European subcohorts or subcohort-specific.
Patients and methods
Patient recruitment and phenotyping
The Genetic-Phenotype Relationship in Sarcoidosis (GenPhenReSa) cohort includes a total of 2,163 patients from 31 European centers from 12 countries that were deeply phenotyped at three different time points over a course of 4 years (14). A positive vote by the Ethics Committee of the University of Freiburg was obtained prior to the initiation of the study, which was also registered in the German Clinical Trials Register (identifier DRKS00000045). In short, sarcoidosis patients were recruited to the study who had been diagnosed with sarcoidosis according to the ATS/ERS/WASOG consensus statement (15) with a documented course of disease over at least 2 years prior to and 2 years following recruitment. As a prevention of migration bias only patients from families living for at least 3 generations in the same country were recruited. The 31 centers were grouped into 13 geographic regions based on geographic (16), clinical, hospital provision and sample size considerations. In general, centers were merged to form presumably homogenous regions with sample sizes of at least 24 patients per region. Thus, some countries had two or three geographically distinct regions (Poland with Lublin (n = 111) and Gdansk (n = 59), and Germany with North (n = 161), Central (n = 152), and South (n = 70)), while small centers from different countries were considered one region based on geographic proximity, e.g., Louven (n = 17) and Bobigny (n = 8) were merged with Amsterdam (n = 67) and Lille (n = 31) to form a Netherlands-Belgium-France region. A full list of recruitment centers and regional groups is given in the Supplementary material, and a detailed description of the GenPhenReSa cohort is given elsewhere (14). For this study, data from 1909 patients with complete information on basic variables (age, sex and region of origin) and documentation of the clinical course over 4 years were analyzed. Moreover, clinical and specific organ involvements such as central nervous system (CNS), skin or ocular involvement, and specific symptoms like night sweat, subfebrile temperature, and dyspnoea were collected using the definition of extrapulmonary organ involvement of the ACCESS study (17). Based on the course of disease in the 2 years prior to recruitment, patients were classified as acute or sub-acute onset phenotype (18). Except for this, all phenotypes used for analysis were defined based on the presentation at baseline, i.e., time of recruitment Patients were grouped into five previously described phenotypes of organ involvement (an abdominal, an ocular–cardio–cutaneous–CNS, musculoskeletal–cutaneous, a pulmonary–lymphonodal and an extrapulmonary phenotype) (14, 19).
Sample and data preparation
DNA from blood samples was isolated using the Qiagen Kit following the manufacturer’s instructions. The amount and quality of all DNA samples was checked by agarose gel electrophoresis, and the DNA was amplified using the REPLI-g whole genome amplification kit prior to genotyping, which was performed using Sequenom Mass-ARRAY iPlex (Sequenom, Inc.) or Taqman (Thermo Fisher Scientific Inc.) technology. Initially, 137 SNPs were selected for investigation based on (i) a known or suggested association with sarcoidosis, (ii) an implication of the gene product in sarcoidosis pathogenesis or (iii) an association of the variant with a clinical phenotype. Allele calling was checked by visual inspection of cluster plots. SNPs or variants with a call rate (CR) < 90%, a minor allele frequency (MAF) < 1% (over all samples from all centers) or an extreme deviation from Hardy–Weinberg equilibrium (p < 10−5) were excluded from analysis, leaving 116 SNPs for further statistical analysis. A full list of SNPs that were included in the analysis before filtering for these quality control parameters together with references is given in the Supplementary material.
Statistical methods
All evaluated phenotype variables were dichotomous (present or not), and their frequencies between study regions were compared by chi-square tests. As the main analysis, the overall association between geno- and phenotypes was investigated in the full cohort. Thus, sarcoidosis patients with a specific phenotype were compared to those without the specific phenotype. Taking the nested data structure into account, a meta-analysis using individual participant data (IPD) approach (20, 21) was conducted, where the regional SNP-wise impact upon each phenotype was estimated by a logistic regression model and combined afterwards across all regions to an overall odds ratio (OR). Such a meta-analysis is necessary due to both clinical and genetic heterogeneity between regions (see results section) which could lead to spurious associations in a “mega-analysis” of simple pooled data (Simpson’s paradox). In detail, for each region i (i = 1, …, 13), all SNPs were modelled separately as an independent variable using an additive genetic model (which has good power even under different true genetic models)In all these region-specific logistic regression models, sex, age and smoking status were included as possible confounders such that the resulting ORi are adjusted for these variables. For the meta-analysis, these individually calculated ORi, by phenotype and by region, were combined across the geographical regions in a weighted random effects model to get an overall OR, respectively for all phenotypes (22). Low phenotype prevalence and low MAF lead to very imprecise OR estimates and extremely large confidence intervals which cannot sensibly be included in a meta-analysis. However, this depends on sample size and in order to include as much information as possible in the meta-analysis, the region-specific ORs were only excluded from the meta-analysis (and further analyses) if the region-specific combination of phenotype prevalence, SNP MAF and sample size was insufficient to allow a reliable estimation of the ORi, i.e., lead to a width of the confidence interval more than 100. Reported ORs always refer to the minor allele: in case of an OR > 1, the minor allele increases the risk of the respective phenotype, while an OR < 1 indicates a reduced risk of the phenotype for the minor allele. Using Bonferroni correction for multiple testing, the significance level of 0.05 was divided by the number of tested SNPs which results in α* = 0.00043 for each phenotype and each SNP in the meta-analysis. Since the considered phenotypes are correlated, we do not correct for the number of phenotypes here.
In an explorative manner, the larger regional sub-cohorts (n > 100) were investigated individually in multi-locus models. In particular, for the acute onset phenotype, SNPs with univariate OR which had a p value smaller than 0.01 were included in a multiple model. The subsequent model selection was done by likelihood ratio criteria to identify a multi-locus model, which allowed interpretation as a genetic signature. In the explorative analyses of the sub-cohorts, all results with a nominal p < 0.01 are presented to balance power and false positive results. Point estimates of the OR are provided with 95% confidence intervals (given in square brackets). All statistical analyses were conducted in R version 3.62 (particularly library metafor (22)).
Results
Patient characteristics
In total, 1909 patients from 13 European regions with a minimum number of 24 patients (in Romania) were included in this analysis. Table 1 lists the phenotypes used for the association analysis together with their prevalences in the largest cohorts. The other 323 patients from in total seven regions with a sample size of less than 100 patients each (Romania, n = 24, Czech Republic, n = 60, Dublin, n = 29, England, n = 29, Gdansk (Poland), n = 59, Iceland, n = 52, South Germany, n = 70) are here summarized in one separate category and are given in (Supplementary Table 2, see https://www.uni-kiel.de/medinfo/mitarbeiter/freitag-wolf/download/). Frequencies of clinical manifestations (i.e., phenotypes) were significantly heterogeneous between regions (p < 0.006 for all characteristics), e.g., in total 755 of 1866 patients suffered from an acute disease onset (40.5%), which had the highest prevalence in the Serbian cohort with 53.6% (480/913) and the lowest prevalence in North Germany with only 18.4% (25/161). In addition to this, the occurrence of missing values differed depending on phenotype and region (23). Genetic heterogeneity, i.e., different minor allele frequencies (see (Supplementary Table 1, for details), between regions was also large, e.g., for rs4143332 at HLA-B the minor allele frequency was 7% in the Netherland-Belgian-France region (n = 125) and 16% in Serbia (n = 913), with even more extreme values in regions with smaller sample sizes, which required the statistical meta-analysis strategy as described above.

Table 1. Frequencies (%) of phenotypes together with the proportion of missing values (%M) in total and in the largest subgroups. Neth-Bel-France: the Netherlands-Belgium-France.
Individual participant data meta-analyses
In a meta-analysis approach all region-wise effects of each SNP upon the phenotypes were combined for the main analysis. After correction for multiple testing, this analysis did not yield any significant association of any of the investigated genetic variants to any phenotype (Supplementary Table 3, see https://www.uni-kiel.de/medinfo/mitarbeiter/freitag-wolf/download/). The largest effect was found for rs4143332 at HLA-B associated with acute onset adjusted for sex, age and smoking status (OR = 1.79, p = 0.013), followed by rs1800629 at TNF (OR = 1.65, p = 0.017), Figures 1 2.
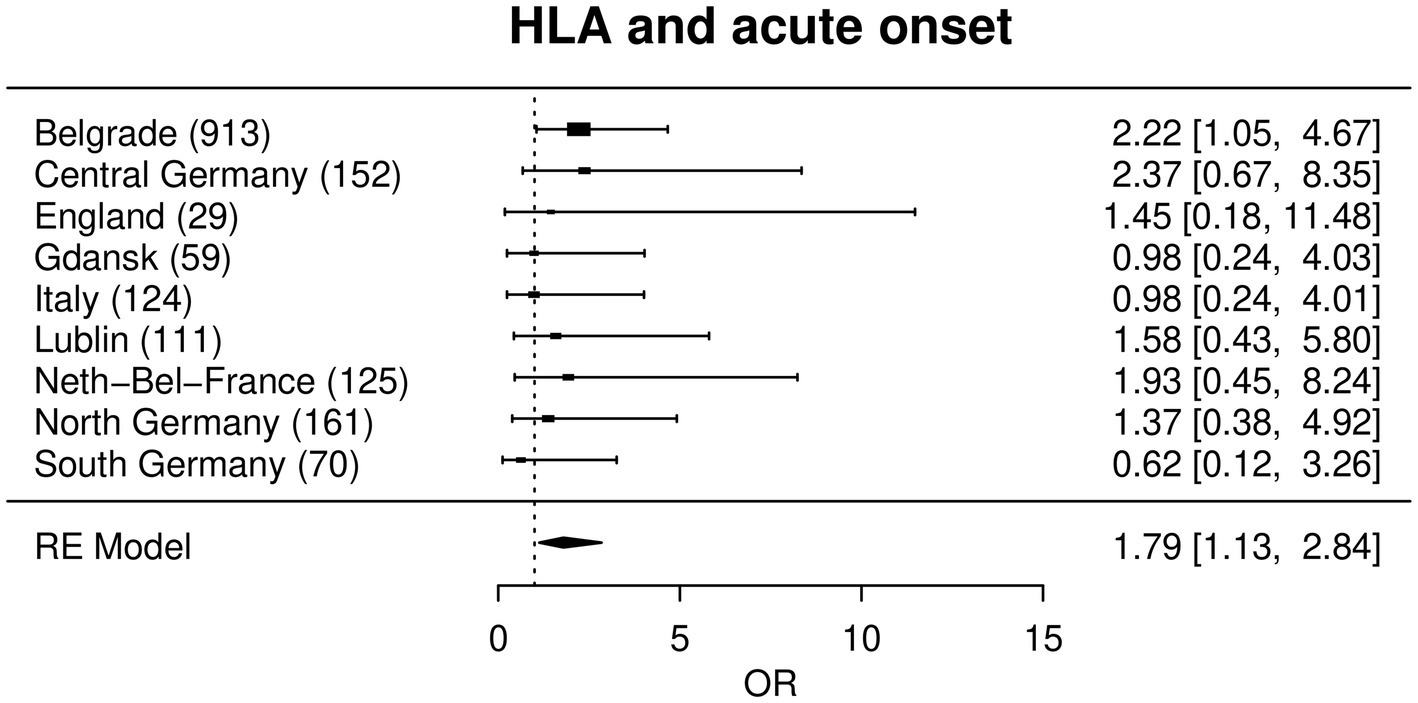
Figure 1. Forest plot with a random effects meta-analysis model for the association between rs4143332 (HLA) and acute onset. Not included are Bucharest, Dublin, Czech Republic and Iceland due to a combination of low sample size, low MAF and low prevalence of acute onset (see statistical methods). Odds ratios (OR) are given with corresponding 95% confidence intervals in square brackets.

Figure 2. Forest plot with a random effects meta-analysis model for the association between rs1800629 (=TNFA -308G/A) and acute onset. Not included are Bucharest and Iceland due to a combination of low sample size, low MAF and low prevalence of acute onset (see statistical methods). Odds ratios (OR) are given with corresponding 95% confidence intervals in square brackets.
Focusing on and summarizing the Central Europe regions (Germany and Netherlands-Belgium-France), the estimated effects were slightly smaller for the above mentioned two loci (OR = 1.58, p = 0.19 and OR = 1.42, p = 0.27, respectively). As an aside, the association with the smallest p-value in the pooled meta-analysis of the Central Europe regions was night sweat and rs2075800, a missense variant near HSPA1L (OR = 3.95, p = 0.0075).
Region-specific analyses
Because of the strong phenotypic and genetic heterogeneity, we also report results of the largest individual sub-cohorts as exploratory analyses. We investigated potential genetic associations with type of onset, phenotypes of organ involvement, specific organ involvements and symptoms within the defined regional sub-cohorts. Apart from acute onset, none of the tested phenotypes in the regions reached the required level of significance after correcting for multiple testing and all reported p values in the subsequent paragraphs are nominal p values. Figure 3 gives a map of the 12 included European countries with their most relevant phenotype–genotype relation in terms of their smallest p-values for each of the analyzed 13 regions. All potentially meaningful (p < 0.01) ORs, CI and p-values of all regions are given in Table 2 and all results with p < 0.05 are given in Supplementary Table 3. In the following, we report details on associations with p < 0.01.
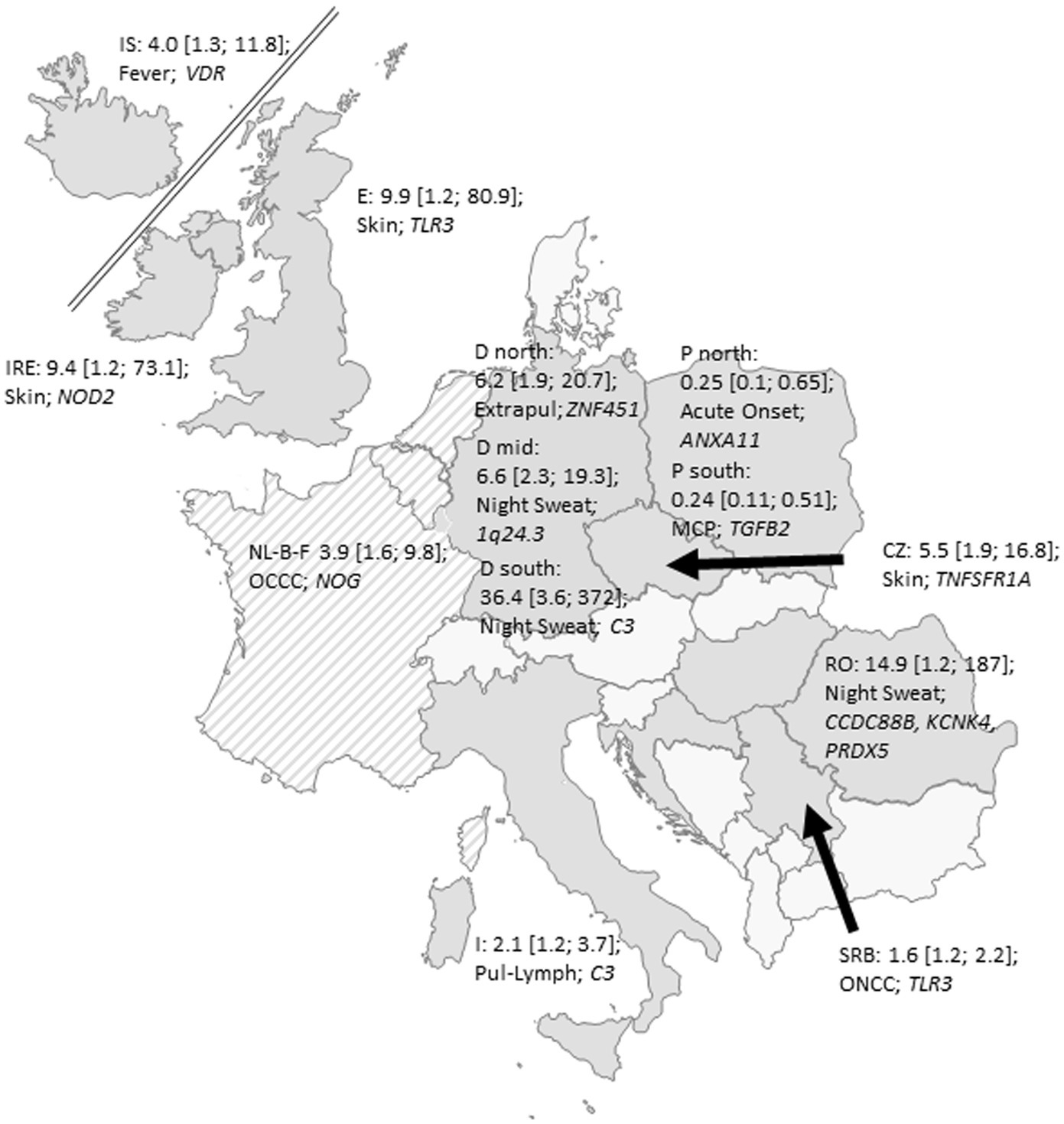
Figure 3. Map of the recruited European countries with their most relevant phenotype–genotype relation (smallest p-values) for each of the analyzed 13 regions expressed as OR with 95% confidence intervals; IS: Iceland; I: Italy; IRE: Ireland; E: England; D north: Northern Germany; D mid: Central Germany; D south: Southern Germany; NL-B-F: the Netherlands-Belgium-France Neth-Bel-France; I: Italy; SRB: Serbia; RO: Romania: Pl1: Gdansk, Poland: Pl2; Lublin, Poland CZ: Czech Republic; Pul-Lymph: Pulmonary–Lymphonodal Phenotype; ONCC: Ocular–Cardio–Cutaneous–CNS Phenotype; Extrapul: Extrapulmonary Phenotype; MCP: Musculoskeletal–Cutaneous Phenotype. Grey: participating countries; hatched: pooled cohorts.
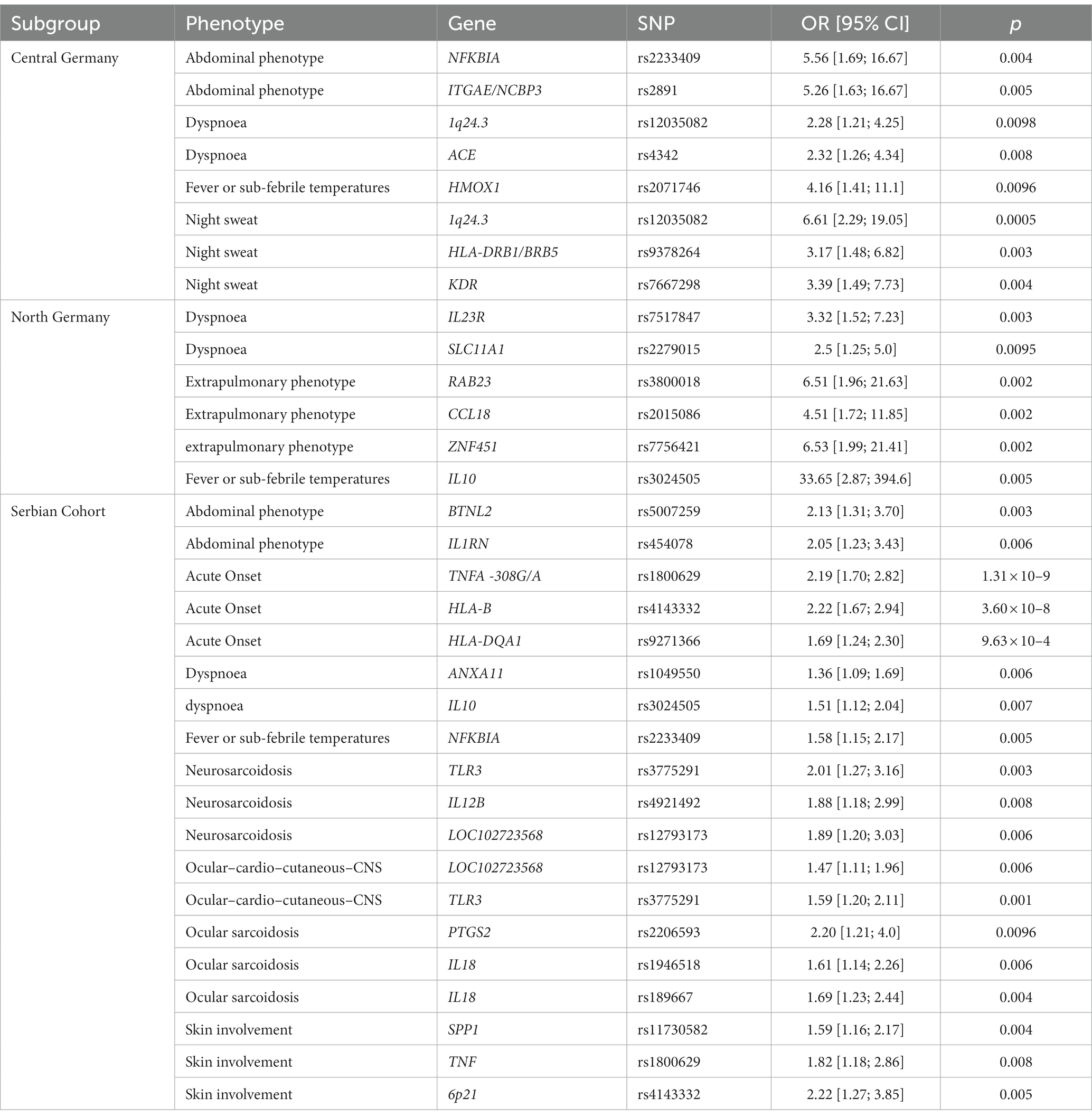
Table 2. Single locus region-specific ORs (sex-, age, and smoking-status adjusted) for phenotypes with p values < 0.01. OR: Odds Ratio; CI: 95% confidence interval.
Acute vs. sub-acute onset
In the Serbian sub-cohort (Table 2), the strongest association of acute onset in the single locus model was found for rs1800629 (=TNFA-308G/A), a well-known sarcoidosis risk SNP that is located in the TNF promoter region: the A-allele was significantly associated with acute onset of the disease with an OR of 2.19 and displayed the smallest p value in the regression analysis (p = 1.31 × 10−9). Moreover, one marker in the HLA-region showed a significant association, namely rs4143332 in the HLA-B gene region (p = 3.60 × 10−8; OR = 2.22). The next strongest signal was found with rs9271366 near HLA-DQA1 (p = 9.63 × 10−4; OR = 1.69; (Supplementary Table 3).
Phenotypes, organ involvement and symptoms
We recently defined novel phenotypes of organ involvement: an abdominal, an ocular–cardio–cutaneous–CNS, a musculoskeletal–cutaneous, a pulmonary–lymphonodal and an extrapulmonary phenotype (14). Here, we analyzed whether these phenotypes are associated with genetic variants and even though none of these reached statistical significance after adjustment for multiple testing, several are still noteworthy with large effect sizes in individual sub-cohorts (Supplementary Table 3). The extrapulmonary phenotype was associated with rs2015086 near CCL18 (p = 0.002; OR = 4.51), rs3800018 near RAB23 (p = 0.002; OR = 6.51) and rs7756421 near ZNF451 (p = 0.002; OR = 6.53) in the North German sub-cohort. The abdominal phenotype was associated with rs5007259 at BTNL2 (p = 0.003; OR = 0.47) and rs454078 IL1RN (p = 0.006; OR = 2.05) in the Serbian sub-cohort, and with rs2233409 near NFKBIA (p = 0.004; OR = 0.18) and rs2891 near ITGAE/NCBP3 (p = 0.005; OR = 0.19) in the Central German sub-cohort. The ocular–cardio–cutaneous–CNS phenotype showed associations with rs12793173 near LOC102723568 (p = 0.006; OR = 0.68) and rs3775291 in the TLR3 gene – resulting in a missense variant - (p = 0.001; OR = 1.59) in the Serbian sub-cohort.
Specific organ involvements
Sarcoidosis is a highly variable disease, especially variable in the distribution of affected organs. We hypothesized that specific genetic variants are associated with distinct organ involvements in sarcoidosis. In the Serbian sub-cohort we found noteworthy risk loci for neurosarcoidosis and ocular sarcoidosis. The minor alleles at three genetic variants are associated with increased risk of neurosarcoidosis: rs3775291 near TLR3 (p = 0.003; OR = 2.01), rs4921492 near IL12B (p = 0.008; OR = 1.88), while the minor allele at rs12793173 near LOC102723568 (p = 0.006; OR = 0.53) is protective. The following risk loci were observed for ocular sarcoidosis: rs2206593 near PTGS2 (p = 0.0096; OR = 2.20), and two variants near IL18, however with opposite directionality: rs1946518 (p = 0.006; OR = 1.61) and rs189667 (p = 0.004; OR = 0.59). In the Serbian cohort, skin involvement was associated with rs11730582 near SPP1 (p = 0.004; OR = 1.59), the aforementioned TNF variant rs1800629 (p = 0.008; OR = 0.55) and rs4143332 at 6p21 (p = 0.005; OR = 0.45). Again, borderline significance of these associations prevents any definite conclusions without further confirmation in other cohorts.
Symptoms
Next, we analyzed whether specific symptoms in patients with sarcoidosis are influenced by distinct genetic variants. This analysis might give valuable hints towards the underlying, genetically influenced bodily response to the disease. In the Central German sub-cohort, night sweat was associated with rs12035082 at 1q24.3 (p = 0.0005; OR = 6.61), rs9378264 near HLA-DRB1/BRB5 (p = 0.003; OR = 3.17) and rs7667298 near KDR (p = 0.004; OR = 3.39). We identified novel genetic associations for fever or sub-febrile temperatures with rs3024505 near IL10 (p = 0.005; OR = 33.65) in the North German sarcoidosis patients, rs2233409 near NFKBIA (p = 0.005; OR = 1.58) in the Serbian patients, and rs2071746 near HMOX1 (p = 0.0096; OR = 0.24) in patients from Central Germany. Dyspnoea in sarcoidosis patients was associated with rs1049550 near ANXA11 (p = 0.006; OR = 1.36) and rs3024505 near IL10 (p = 0.007; OR = 0.66) in the Serbian cohort, rs12035082 at 1q24.3 (p = 0.0098; OR = 2.28) and rs4342 near ACE (p = 0.008; OR = 0.43) in the Central German cohort, and rs7517847 near IL23R (p = 0.003; OR = 3.32) and rs2279015 near SLC11A1 (p = 0.0095; OR = 0.4) in the North German cohort. Taken together, these results suggest that the emergence of some symptoms in patients with sarcoidosis might indeed be associated with specific genetic loci, however, no association reached statistical significance when adjusted for multiple testing.
Multi-locus models
In the Serbian and Lublin (Poland) cohorts, two separate sets of three SNPs in six different genes showed association to the acute onset phenotype in multi-locus models (Table 3). In the Serbian cohort, after backward selection of all loci with p < 0.01, the individual impact of the analyzed SNPs on acute onset decreased for rs1800629 from OR = 2.19 in the single-locus model to 1.95 in the multi-locus model and for rs9271366 from OR = 1.69 to OR = 1.43, whereas the OR of rs4143332 increased slightly from 2.22 to OR = 2.34. A combination of the risk alleles at all three SNPs conferred an OR of 6.6 when considered as a genetic signature. In Lublin, all three SNPs had substantially stronger effects in a multi-locus model and the combined effect of a genetic signature comprising all three risk alleles corresponded to an OR of 37.9. Two of the SNPs in this genetic signature, namely rs1891467 near TGFB2 and rs7667298 near KDR, had been reported to be associated with an acute course of sarcoidosis before in a German cohort (4).
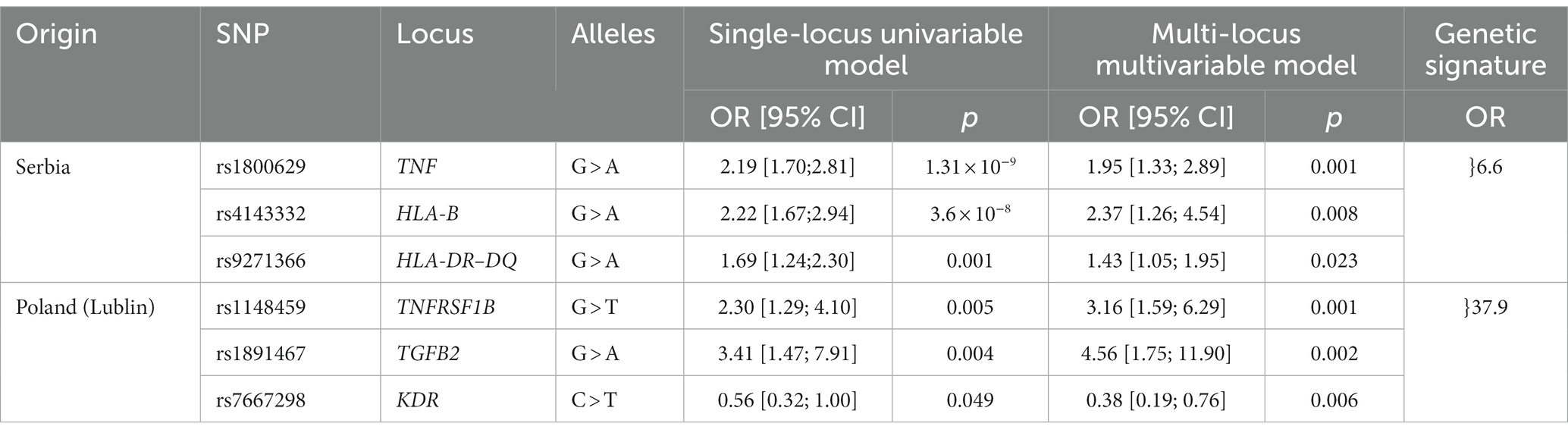
Table 3. Single and multi-locus models of acute onset in the Serbian and Lublin (Poland) sub-cohort. The ORs are sex-, age, and smoking-status adjusted. OR: Odds Ratio; CI: confidence interval.
In amalgamate, two different sets of genetic variants are permissive for the same phenotype of acute disease onset resulting in combined ORs of 6.6 for the Serbian and of 37.9 for the Polish subcohort which points at their likely interactions with different local environmental factors resulting in the same phenotype.
Discussion
Sarcoidosis is a highly variable and heterogeneous disease with a need for understanding the genetic background of this heterogeneity. In total 116 selected candidate SNPs were analyzed for their association with clinical traits in sarcoidosis patients of the GenPhenReSa cohort (14). The GenPhenReSa patient collection represents a large sarcoidosis cohort of almost 2,000 patients of European origin that was deeply phenotyped over 4 years at three points in time, i.e., every 2 years (14). In line with the known heterogeneity, we did not observe any genotype–phenotype association that was uniformly present in the whole European cohort, when analyzed with an appropriate meta-analysis. However, different unique associations were present in different geographic sub-cohorts. With our approach, we were able to identify specific SNPs that are associated with course of the disease (acute onset) in the Serbian and Lublin (Poland) sub-cohorts which result in increased ORs if used as a set. In addition, we found some genetic associations with the phenotypes of organ involvement and specific symptoms, mostly restricted to individual geographic cohorts (Figure 3). In these smaller samples, effect sizes were large but did not reach the predefined, stringent significance levels.
Most strikingly and of potential prognostic value, marker rs1800629 (=TNF-308G/A), a SNP in the TNF promoter region, was associated with acute onset. The A-allele of this SNP is associated with acute onset with an OR of 2.18 in the Serbian sub-cohort, which is in line with previous reports that describe the A-allele being associated with a favorable prognosis (24). Likewise, associations with acute onset were observed for the HLA variants rs4143332 (OR = 2.22) and rs9271366 (OR = 1.69). The latter variant is located between HLA-DRB1 and HLA-DQA1 and is associated with inflammatory bowel disease, multiple sclerosis, systemic lupus erythematosus and others (25–27), but has not yet been reported with regards to sarcoidosis. Whether or not this finding might be dependent on HLA haplotypes that are known to be associated with certain sarcoidosis subtypes has to be analyzed in future studies. Noteworthy, used in combination, these variants are associated with acute onset sarcoidosis with an OR of 6.6 (Table 3) which could be used as a genetic signature.
Interestingly, the estimated effects of SNPs near TNFRSF1B, TGFB2, and KDR on acute onset in the Polish sub-cohort from Lublin increased in the multi-locus models and result in an OR of 37.9 (Table 3) when considered as a genetic signature. TGFB2 and KDR variants were reported to be associated in a German cohort with an acute course of sarcoidosis (4) and variants of TNFRSF1B are known to influence the response to anti-TNF therapy in sarcoidosis (28). Which leads to the concept of a possible regulation of acute sarcoid inflammation by these genetic variants.
The effects of SNPs near TNFSFR1B, TGFB2 and KDR on acute onset are increased in the multi-locus models. The latter two SNPs were reported to be associated with an acute course of sarcoidosis before (4). Different roles of the variant gene products in the regulation of inflammation can be assumed on the basis of ex-vivo data. The SNP rs1148459 is located at the position −9,298 of the 5′-flanking region of TNFRSF1B and it is speculated that this SNP may regulate the expression of the receptor (29). TNFSFR1B is associated with the generation of Treg (30). Lack of TNFSFR1B expression of Treg may lead to non-functional Treg which suggests a functional influence on the effect of anti-TNF therapy of sarcoidosis (28). The SNP rs1891467 is located in the beginning of the second intron of TGFB2, however, the consequences of the SNP are unclear. Members of the TGFβ family are anti-inflammatory mediators (31) and in bronchoalveolar lavage cell culture supernatants from patients suffering from active sarcoidosis with spontaneous remission we found increased levels of TGFβ indicating an anti-inflammatory activity of TGFβ also in sarcoidosis. However, it is currently not clear whether or not this is related with SNP rs1891467. The SNP rs7667298 is located in the 5’UTR of the gene KDR which codes for the VEGF-R2. It has been demonstrated that rs7667298 is related with diminished expression of VEGF-R2 (32). In contrast, in sarcoidosis increased levels of VEGF have been detected (33). VEGF-R2 is a strong inducer of vascularization whereas VEGF-R1 exerts pro-inflammatory stimulation. Thus, lack of VEGF-R2 may lead to abnormal vascularization (33) but fosters inflammation. Thus, relevance of these genetic variants on the up- and down-regulation of sarcoid alveolitis is most likely.
The number of significant associations between SNPs and clinical variables in the entire cohort of 1909 Caucasians fell short of expectations, which may suggest that permissive genetic signatures depend strongly on specific environmental or occupational events in a particular geographic region or cohort to cause a particular phenotype as suggested by the ACCESS-study (19, 34). This conclusion seems likely since all patients came from families that lived for at least three generations in the same region. Nevertheless, the application of genetic markers as predictors for certain phenotypes has turned out to be challenging for the majority of complex diseases and various methods have been applied to identify risk loci sets [reviewed in (35)]. A critical point in our study may have been the choice of markers that were genotyped which had been selected because of previous reports in sarcoidosis or other granulomatous diseases. As this was not a genome-wide association study, we might have missed important genetic associations. Furthermore, an association with the investigated phenotypes had not been established before for the majority of the analyzed SNPs and therefore replication in independent cohorts of diverse ethnicities and with different study designs is warranted.
Within the entire cohort, which comprised a large, genetically and phenotypically heterogeneous population there were no clearly significant genetic associations of the phenotypes of organ involvement; however, the observed effects were large (ORs >2 or <0.5) within specific and more homogeneous geographic areas.
In this multicentre study, the number of patients and prevalences of specific phenotypes for the clinical variables were not large enough to detect significant genetic effects on most phenotypes. Indeed, in terms of statistical considerations, with the overall sample size of more than 1900 patients, an OR larger 1.3 could have been found with a power above 90% (additive model, alpha set to 0.00043, allele frequency assumed to be 0.25), but the genetic and phenotypic heterogeneity of the included patients turned out to be most challenging. While the genotype frequencies of investigated markers show at least a slight gradient within Europe (16), which was addressed by the meta-analysis, the phenotypic heterogeneity was so large that it could not be fully compensated by statistical methods. This large phenotypic heterogeneity between regions might be due to differences in health care systems and recruitment settings (different age distributions of cases, different commonly used treatment) and different frequencies of individual non-genetic risk factors such as smoking which has been reported to increase, e.g., the susceptibility in Sweden by a gene–environment interaction (36). Moreover, about 30% of the sarcoidosis incidence is attributed to occupational exposures (37) and many associations of sarcoidosis with environmental, infectious or occupational exposures have been reported (19, 38, 39). Unfortunately, individual exposure information is not available for the GenPhenReSa cohort. Chronic Beryllium Disease (CBD) is a perfect phenocopy of sarcoidosis and SNPs associated with CBD exert janiform functions depending on disease stage which might also be the case in sarcoidosis which demands more detailed phenotyping in future studies (40). Thus, gene - environment / occupation interactions have to be considered which cause a complex setting of exposures to be analyzed. An example would be the specific T-cell response to a peptide of Aspergillus nidulans documented in Swedish HLA-DR3 positive Löfgren’s syndrome patients (41). Further genetic studies are needed and our data presented in the (Supplementary Tables will be very helpful in designing future studies.
Taken together, we confirmed the association of rs4143332 at HLA-B and rs1800629 at TNF locus with acute onset of sarcoidosis in the Serbian sub-cohort and describe potential genetic links with several clinical features, including the recently described phenotypes of organ involvement (14, 19). Furthermore, we demonstrate that the concept of genetic signatures needs to be complemented with environmental and occupational data. The observed associations of genetic and clinical features suggest that the clinical phenotype is highly dependent on subcohort-specific environmental interactions with genetics and is more complex than anticipated. This view is supported by the fact that in the Polish and German geographic subcohorts different gene variants exhibited highest ORs (Figure 3). Most likely different genetic backgrounds in combination with local exposures are responsible for identical sarcoid phenotypes in different environmental and regional settings of Europe (14, 19). Our finding represents an important and necessary step towards understanding the genetic architecture of the heterogeneous and complex disease sarcoidosis. Local signatures comprising genetic, occupational, environmental and infectious parameters will be necessary and seem achievable to predict courses of the disease.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.uni-kiel.de/medinfo/mitarbeiter/freitag-wolf/download/.
Ethics statement
The studies involving human participants were reviewed and approved by Ethik-Kommission der Albert-Ludwigs-Universität Freiburg, email: ekfr@uniklinik-freiburg.de. The patients/participants provided their written informed consent to participate in this study.
Author contributions
JM-Q, SS, GZ, and AP: design and planning of study, application for funding, directing performance, obtaining ethics committee vote, and clinical trials registration. JCS, RK, VM-V, JM, DJ, EB, PR, DB, RJ, L-PH, KG, AnD, BM, AG, MiP, MK, SH, FB, CGrah, TP-G, ZK, SP, CGrohé, JS, MS, AM, JH, WW, LS, MaP, DV, VP, HW, and AP: patient recruitment, phenotyping, physical examination, radiograph reading, checking inclusion criteria, and documentation. AF and GZ: biobanking and DNA quality control. AP, JM-Q, JCS, EB, and PR: phenotyping quality control. JM-Q, JS, and BF: clinical data quality control. SS, AF, and RA: genotyping. SF-W, SS, and AsD: genotyping quality control. SF-W, AsD, JCS, SS, RA, JM-Q, and BF: data analysis. SF-W and AsD: statistics. SF-W, AsD, JCS, JM-Q, BF, and GZ: manuscript writing. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by German Federal Ministry for Education and Research grant 01EY1103 and German Research Foundation grant MU 692/8-1 and FI 1935/1-1, DFG Exzellenzcluster PMI EXC 2/67 and Clusterlab XI EXC 306/2. JCS is supported by the CORE100Pilot Advanced Clinician Scientist Program of Hannover Medical School funded by Else Kröner-Fresenius Foundation (2020_EKSP.78) and the Ministry of Science and Culture of Lower Saxony, and the Fritz Thyssen-Foundation.
Acknowledgments
We thank all patients for participating in this study. Late V. Kolek (Dept. Respiratory Medicine, University Hospital Olomouc) is acknowledged for supporting this study.
Conflict of interest
FB received from Boehringer Ingelheim, Fujirebio, Galapagos NV and Roche; personal travel support from Boehringer Ingelheim and Roche, board membership fees from Boehringer Ingelheim, Bristol Myers Squibb, Fujirebio, Galapagos, GlaxoSmithKline, Roche and Takeda. DB grants paid to the institution from Synairgen and Sanofi; personal honoraria received from Chiesi, Norvatis and Sanofi; personal honoraria for lectures received from Astra Zeneca, Eli Lilly and Norvatis; not for profit activities for ministry of health, Romania. AF received a grant from the German Research Foundation. BF: received grants from Bristol Myers Squibb and AdVita Lifescience paid to the institution; personal honoraria for lectures from Boehringer Ingelheim, Astra Zeneca and Roche, personal travel support from Boehringer Ingelheim; reports a pending patient WO2020225246A1 and stocks from Relief Therapeutics. CG received lecture fees from Astra Zeneca paid to the institution. MK received consulting fees from Boehringer Ingelheim. JM-Q grants from German Research Council, Bristol Myers Squibb and AdVita Lifesciences paid to the institution, received personal honoria from AdVita Lifesciences and Roche, received lecture fees from Astra Zeneca and Roche, received payments for expert testimony from AdVita Lifescience; received travel support from AdVita Lifescience and Grifols, reports a pending patent WO2020225246A1 and stocks from Relief Therapeutics. MaP received grants from Palacky University paid to the institution. MiP reports fiduciary activities for Deutsche Gesellschaft für Pneumologie und Beatmungsmedizin. VP received personal consulting fees from Ambu and Erbe; lecture honoraria from Boehringer Ingelheim and Roche and participated in a data safety monitoring board for Boehringer Ingelheim. AP received personal consulting fees from Boehringer Ingelheim, Roche, AstraZeneca and Galapagos, lecture fees from Boehringer Ingelheim, Norvatis and Gilead, participated in data safety monitoring boards for Boehringer Ingelheim, Roche and AstraZeneca and reports fiduciary activities for the European Rare Disease Network Interstitial Lung Disease Group and the steering committee of the World Association of Sarcoidosis and Other Granulomtous Disorders. SS reports grants from the German Research Council paid to the institution, personal consulting fees from Abbvie, Arenal, BMS, Biogen, Celltrion, Celgen, IMAB, Gilead, MSD, Mylan, Pfizer, Fresinius, Janssen, Takeda, Theravance, Provention Bio, Protagonist, Falk, Ferring, UCB, Amgen, Sandoz Hexal, Lilly, Galapagos. JCS grants from the German Research Council, Else Kröner-Fresenius Foundation, and Fritz Thyssen-Foundation paid to the institution; received personal lecture honoraria from Boehringer Ingelheim. LS received personal honoraria from Boehringer Ingelheim for educational events. D received personal honoraria from Boehringer Ingelheim for activities in advisory boards and lecture fees from Boehringer Ingelheim and AstraZeneca and reports travel support from Boehringer Ingelheim. WW grants from Roche, Boehringer Ingelheim, and Galapagos, consulting fees from Sanofi, Boehringer Ingelheim and Roche, honoraria for lectures from Roche and Beohdringer Ingelheim and payments for activities in data safety monitoring boards from Boehringer Ingelheim and Roche all paid to the institution. GZ reports a pending patent (EP 2585089 A1) and stock options for Moderna, Biontech and Relief Therapeutics.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2023.1218106/full#supplementary-material
References
1. Grunewald, J, Grutters, JC, Arkema, EV, Saketkoo, LA, Moller, DR, and Müller-Quernheim, J. Sarcoidosis. Nat Rev Dis Primers. (2019) 5:45. doi: 10.1038/s41572-019-0096-x
2. Schupp, JC, Frye, BC, Zissel, G, and Müller-Quernheim, J. Sarcoidosis: drugs under investigation. Semin Respir Crit Care Med. (2017) 38:532–7. doi: 10.1055/s-0037-1603768
3. Sverrild, A, Backer, V, Kyvik, KO, Kaprio, J, Milman, N, Svendsen, CB, et al. Heredity in sarcoidosis: a registry-based twin study. Thorax. (2008) 63:894–6. doi: 10.1136/thx.2007.094060
4. Fischer, A, Grunewald, J, Spagnolo, P, Nebel, A, Schreiber, S, and Müller-Quernheim, J. Genetics of sarcoidosis. Semin Respir Crit Care Med. (2014) 35:296–306. doi: 10.1055/s-0034-1376860
5. Crouser, ED, Fingerlin, TE, Yang, IV, Maier, LA, Nana-Sinkam, P, Collman, RG, et al. Application of "omics" and systems biology to sarcoidosis research. Ann Am Thorac Soc. (2017) 14:S445–51. doi: 10.1513/AnnalsATS.201707-567OT
6. Rivera, NV, Ronninger, M, Shchetynsky, K, Franke, A, Nöthen, MM, Müller-Quernheim, J, et al. High-density genetic mapping identifies new susceptibility variants in sarcoidosis phenotypes and shows genomic-driven phenotypic differences. Am J Respir Crit Care Med. (2016) 193:1008–22. doi: 10.1164/rccm.201507-1372OC
7. Valentonyte, R, Hampe, J, Huse, K, Rosenstiel, P, Albrecht, M, Stenzel, A, et al. Sarcoidosis is associated with a truncating splice site mutation in BTNL2. Nat Genet. (2005) 37:357–64. doi: 10.1038/ng1519
8. Hofmann, S, Franke, A, Fischer, A, Jacobs, G, Nothnagel, M, Gaede, KI, et al. Genome-wide association study identifies ANXA11 as a new susceptibility locus for sarcoidosis. Nat Genet. (2008) 40:1103–6. doi: 10.1038/ng.198
9. Fischer, A, Nothnagel, M, Franke, A, Jacobs, G, Saadati, HR, Gaede, KI, et al. Association of inflammatory bowel disease risk loci with sarcoidosis, and its acute and chronic subphenotypes. Eur Respir J. (2011) 37:610–6. doi: 10.1183/09031936.00049410
10. Grunewald, J, Brynedal, B, Darlington, P, Nisell, M, Cederlund, K, Hillert, J, et al. Different HLA-DRB1 allele distributions in distinct clinical subgroups of sarcoidosis patients. Respir Res. (2010) 11:25. doi: 10.1186/1465-9921-11-25
11. Song, GG, Kim, JH, and Lee, YH. Associations between TNF-α −308 A/G and lymphotoxin-α +252 A/G polymorphisms and susceptibility to sarcoidosis: a meta-analysis. Mol Biol Rep. (2014) 41:259–67. doi: 10.1007/s11033-013-2859-x
12. Takashige, N, Naruse, TK, Matsumori, A, Hara, M, Nagai, S, Morimoto, S, et al. Genetic polymorphisms at the tumour necrosis factor loci (TNFA and TNFB) in cardiac sarcoidosis. Tissue Antigens. (1999) 54:191–3. doi: 10.1034/j.1399-0039.1999.540211.x
13. Levin, AM, Iannuzzi, MC, Montgomery, CG, Trudeau, S, Datta, I, McKeigue, P, et al. Association of ANXA11 genetic variation with sarcoidosis in African Americans and European Americans. Genes Immun. (2013) 14:13–8. doi: 10.1038/gene.2012.48
14. Schupp, JC, Freitag-Wolf, S, Bargagli, E, Mihailović-Vučinić, V, Rottoli, P, Grubanovic, A, et al. Phenotypes of organ involvement in sarcoidosis. Eur Respir J. (2018) 51:1700991. doi: 10.1183/13993003.00991-2017
15. Statement on sarcoidosis. Joint statement of the American Thoracic Society (ATS), the European Respiratory Society (ERS) and the world Association of Sarcoidosis and Other Granulomatous Disorders (WASOG) adopted by the ATS Board of directors and by the ERS executive committee, February 1999. Am J Respir Crit Care Med. (1999) 160:736–55.
16. Lao, O, Lu, TT, Nothnagel, M, Junge, O, Freitag-Wolf, S, Caliebe, A, et al. Correlation between genetic and geographic structure in Europe. Curr Biol. (2008) 18:1241–8. doi: 10.1016/j.cub.2008.07.049
17. Judson, MA, Baughman, RP, Teirstein, AS, Terrin, ML, and Yeager, H Jr. Defining organ involvement in sarcoidosis: the ACCESS proposed instrument. ACCESS research group. A case control etiologic study of sarcoidosis. Sarcoidosis Vasc Diffuse Lung Dis. (1999) 16:75–86.
18. Prasse, A, Katic, C, Germann, M, Buchwald, A, Zissel, G, and Müller-Quernheim, J. Phenotyping sarcoidosis from a pulmonary perspective. Am J Respir Crit Care Med. (2008) 177:330–6. doi: 10.1164/rccm.200705-742OC
19. Lhote, R, Annesi-Maesano, I, Nunes, H, Launay, D, Borie, R, Sacré, K, et al. Clinical phenotypes of extrapulmonary sarcoidosis: an analysis of a French, multi-ethnic, multicentre cohort. Eur Respir J. (2021) 57:2001160. doi: 10.1183/13993003.01160-2020
20. Stewart, GB, Altman, DG, Askie, LM, Duley, L, Simmonds, MC, and Stewart, LA. Statistical analysis of individual participant data meta-analyses: a comparison of methods and recommendations for practice. PLoS One. (2012) 7:e46042. doi: 10.1371/journal.pone.0046042
21. Debray, TP, Moons, KG, van Valkenhoef, G, Efthimiou, O, Hummel, N, Groenwold, RHH, et al. Get real in individual participant data (IPD) meta-analysis: a review of the methodology. Res Synth Methods. (2015) 6:293–309. doi: 10.1002/jrsm.1160
22. Viechtbauer, W. Conducting Meta-analyses inRwith themetaforPackage. J Stat Softw. (2010) 36:1–48. doi: 10.18637/jss.v036.i03
23. Choi, SC, and Lu, IL. Effect of non-random missing data mechanisms in clinical trials. Stat Med. (1995) 14:2675–84. doi: 10.1002/sim.4780142407
24. Wijnen, PA, Nelemans, PJ, Verschakelen, JA, Bekers, O, Voorter, CE, and Drent, M. The role of tumor necrosis factor alpha G-308A polymorphisms in the course of pulmonary sarcoidosis. Tissue Antigens. (2010) 75:262–8. doi: 10.1111/j.1399-0039.2009.01437.x
25. Yang, SK, Hong, M, Zhao, W, Jung, Y, Baek, J, Tayebi, N, et al. Genome-wide association study of Crohn's disease in Koreans revealed three new susceptibility loci and common attributes of genetic susceptibility across ethnic populations. Gut. (2014) 63:80–7. doi: 10.1136/gutjnl-2013-305193
26. Australia, New Zealand Multiple Sclerosis Genetics C. Genome-wide association study identifies new multiple sclerosis susceptibility loci on chromosomes 12 and 20. Nat Genet. (2009) 41:824–8. doi: 10.1038/ng.396
27. Ruiz-Narvaez, EA, Fraser, PA, Palmer, JR, Cupples, LA, Reich, D, Wang, YA, et al. MHC region and risk of systemic lupus erythematosus in African American women. Hum Genet. (2011) 130:807–15. doi: 10.1007/s00439-011-1045-2
28. Crommelin, H, Vorselaars, A, van der Vis, J, Deneer, V, and van Moorsel, CHM. Pharmacogenetics of antitumor necrosis factor therapy in severe sarcoidosis. Curr Opin Pulm Med. (2020) 26:267–76. doi: 10.1097/MCP.0000000000000681
29. Chadha, S, Miller, K, Farwell, L, Sacks, S, Daly, MJ, Rioux, JD, et al. Haplotype analysis of tumour necrosis factor receptor genes in 1p36: no evidence for association with systemic lupus erythematosus. Eur J Hum Genet. (2006) 14:69–78. doi: 10.1038/sj.ejhg.5201527
30. Mohamud, R, LeMasurier, JS, Boer, JC, Sieow, JL, Rolland, JM, O’Hehir, RE, et al. Synthetic nanoparticles that promote tumor necrosis factor receptor 2 expressing regulatory T cells in the lung and resistance to allergic airways inflammation. Front Immunol. (2017) 8:1812. doi: 10.3389/fimmu.2017.01812
31. Zissel, G, Schlaak, J, Schlaak, M, and Müller-Quernheim, J. Regulation of cytokine release by alveolar macrophages treated with interleukin-4, interleukin-10, or transforming growth factor beta. Eur Cytokine Netw. (1996) 7:59–66.
32. An, SJ, Chen, ZH, Lin, QX, Su, J, Chen, HJ, Lin, JY, et al. The −271 G>a polymorphism of kinase insert domain-containing receptor gene regulates its transcription level in patients with non-small cell lung cancer. BMC Cancer. (2009) 9:144. doi: 10.1186/1471-2407-9-144
33. Yamashita, M, Mouri, T, Niisato, M, Kowada, K, Kobayashi, H, Chiba, R, et al. Heterogeneous characteristics of lymphatic microvasculatures associated with pulmonary sarcoid granulomas. Ann Am Thorac Soc. (2013) 10:90–7. doi: 10.1513/AnnalsATS.201209-078OC
34. Newman, LS, Rose, CS, Bresnitz, EA, Rossman, MD, Barnard, J, Frederick, M, et al. A case control etiologic study of sarcoidosis: environmental and occupational risk factors. Am J Respir Crit Care Med. (2004) 170:1324–30. doi: 10.1164/rccm.200402-249OC
35. Jostins, L, and Barrett, JC. Genetic risk prediction in complex disease. Hum Mol Genet. (2011) 20:R182–8. doi: 10.1093/hmg/ddr378
36. Rivera, NV, Patasova, K, Kullberg, S, Diaz-Gallo, LM, Iseda, T, Bengtsson, C, et al. A gene-environment interaction between smoking and gene polymorphisms provides a high risk of two subgroups of sarcoidosis. Sci Rep. (2019) 9:18633. doi: 10.1038/s41598-019-54612-1
37. Blanc, PD, Annesi-Maesano, I, Balmes, JR, Cummings, KJ, Fishwick, D, Miedinger, D, et al. The occupational burden of nonmalignant respiratory diseases. An official American Thoracic Society and European Respiratory Society statement. Am J Respir Crit Care Med. (2019) 199:1312–34. doi: 10.1164/rccm.201904-0717ST
38. Judson, MA. Environmental risk factors for sarcoidosis. Front Immunol. (2020) 11:1340. doi: 10.3389/fimmu.2020.01340
39. Oliver, LC, and Zarnke, AM. Sarcoidosis: An occupational disease? Chest. (2021) 160:1360–7. doi: 10.1016/j.chest.2021.06.003
40. Frye, BC, Gaede, KI, Saltini, C, Rossman, MD, Monos, DS, Rosenman, KD, et al. Analysis of single nucleotide polymorphisms in chronic beryllium disease. Respir Res. (2021) 22:107. doi: 10.1186/s12931-021-01691-2
Keywords: genetic polymorphism, genetic risk factors, sarcoidosis, genotype–phenotype-relationship, region-specific genetic links
Citation: Freitag-Wolf S, Schupp JC, Frye BC, Fischer A, Anwar R, Kieszko R, Mihailović-Vučinić V, Milanowski J, Jovanovic D, Zissel G, Bargagli E, Rottoli P, Bumbacea D, Jonkers R, Ho L-P, Gaede KI, Dubaniewicz A, Marshall BG, Günther A, Petrek M, Keane MP, Haraldsdottir SO, Bonella F, Grah C, Peroš-Golubičić T, Kadija Z, Pabst S, Grohé C, Strausz J, Safrankova M, Millar A, Homolka J, Wuyts WA, Spencer LG, Pfeifer M, Valeyre D, Poletti V, Wirtz H, Prasse A, Schreiber S, Dempfle A and Müller-Quernheim J (2023) Genetic and geographic influence on phenotypic variation in European sarcoidosis patients. Front. Med. 10:1218106. doi: 10.3389/fmed.2023.1218106
Edited by:
Ivan Castellví, Hospital Universitari de la Santa Creu i Sant Pau, SpainReviewed by:
Elzbieta Radzikowska, National Institute of Tuberculosis and Lung Diseases, PolandShu-Yi Liao, National Jewish Health, United States
Eliott Crouser, The Ohio State University, United States
Copyright © 2023 Freitag-Wolf, Schupp, Frye, Fischer, Anwar, Kieszko, Mihailović-Vučinić, Milanowski, Jovanovic, Zissel, Bargagli, Rottoli, Bumbacea, Jonkers, Ho, Gaede, Dubaniewicz, Marshall, Günther, Petrek, Keane, Haraldsdottir, Bonella, Grah, Peroš-Golubičić, Kadija, Pabst, Grohé, Strausz, Safrankova, Millar, Homolka, Wuyts, Spencer, Pfeifer, Valeyre, Poletti, Wirtz, Prasse, Schreiber, Dempfle and Müller-Quernheim. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joachim Müller-Quernheim, am9hY2hpbS5tdWVsbGVyLXF1ZXJuaGVpbUB1bmlrbGluaWstZnJlaWJ1cmcuZGU=
†These authors share first authorship
‡These authors share senior authorship