 Yunhua Fu1†
Yunhua Fu1† Hao Yue
Hao Yue Yulin Dai
Yulin Dai
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Med., 19 August 2022
Sec. Gastroenterology
Volume 9 - 2022 | https://doi.org/10.3389/fmed.2022.988507
This article is part of the Research TopicRecent Advances in Enhancing Chemotherapeutic Efficacy of Colorectal CancerView all 10 articles
Seaweeds are classified as Chlorophyta, Rhodophyta, and Phaeophyta. They constitute a number of the most significant repositories of new therapeutic compounds for human use. Seaweed has been proven to possess diverse bioactive properties, which include anticancer properties. The present review focuses on colorectal cancer, which is a primary cause of cancer-related mortality in humans. In addition, it discusses various compounds derived from a series of seaweeds that have been shown to eradicate or slow the progression of cancer. Therapeutic compounds extracted from seaweed have shown activity against colorectal cancer. Furthermore, the mechanisms through which these compounds can induce apoptosis in vitro and in vivo were reviewed. This review emphasizes the potential utility of seaweeds as anticancer agents through the consideration of the capability of compounds present in seaweeds to fight against colorectal cancer.
Colorectal cancer (CRC) is the most common type of cancer throughout the world, accounting for approximately 10% of all new cancer cases and mortality, as projected in GLOBOCAN 2020 (1). The prevalence rates of CRC are increasing among nations with a medium human development index, such as Brazil, Russia, and countries of Latin America (2). The pathology of CRC includes carcinogenesis of the rectum, colon, appendix, and anus (3). Familial and environmental factors contribute to the risk of CRC from two well-defined causes particularly amenable to dietary influence (4). In the clinical situation, chemotherapy is a common treatment modality for CRC (5). Nevertheless, the majority of current chemotherapeutic drugs for the therapy of advanced-stage CRC, for instance cisplatin, have been repeatedly reported to elicit adverse side effects and are comparatively less effective (6). Several lines of scientific evidence, from molecular mechanisms to clinical trials, show that herbal medicines have anti-CRC potential and have been used for therapy and recovery (7).
Seaweeds have been utilized for food and medicinal herbs since ancient times in Asia (8). It has been consumed as a food for over 1,700 years, which can be dated back to Japan in the fourth century and China during sixth century. Particularly, people living long in coastal areas frequently used seaweed as a main dish, side dish, or soup (9). Consumption of seaweed supplies sufficient macro and micronutrients, which are essential to maintaining human health (10). Besides nutritional effects, seaweed has long been adopted as a drug in Traditional East Asian Medicine to alleviate the progression of multiple cancers (11). Seaweeds as large multicellular marine organisms are classified into three major groups based on their pigments and the origin of sulfated polysaccharides: green (Chlorophyta), red (Rhodophyta), and brown (Phaeophyta) (12, 13). They represent a main source of bioactive compounds, yielding primary metabolites essential for natural growth and many secondary metabolites, which include polysaccharides, polyunsaturated fatty acids, phenolics, vitamins, pigments, minerals, terpenes, and phytosterols (14). Due to their various constituents, seaweeds have shown diverse biological activities, including anticancer activity (15).
Seaweeds have long been recognized as a therapeutic option of cancer (16). Accumulating evidence advocates that the anticancer effects of bioactive ingredients extracted from seaweed are produced via multiple mechanisms of action, including inhibition of growth, invasion, and metastasis of cancer cells, and through the stimulation of apoptosis in cancer cells (17). Among the East Asian population, people who regularly consumed seaweed reduced their risk of CRC development by half (10). Several researches have suggested that CRC can be effectively treated with marine natural products (18). According to one such report, brown seaweed Turbinaria decurrens has the potential as an anti-CRC agent (19). The highly cytotoxic and antiproliferative activities of seaweeds from the Portuguese coast have been proven in a model of Caco-2 CRC cells in vitro (20). Sargassum oligocystom significantly decreased cell viability in SW742, HT-29, WiDr, and CT-26 CRC cells through activation of the APC gene (21). Although several studies have reported the therapeutic properties of seaweed in CRC, its mechanism of action and active ingredients are still unclear and unclassified. In this review, we summarize the various effects of diverse compounds derived from seaweed on CRC.
The polysaccharides present in seaweeds are many and diverse (22). They are hydrophilic molecules with high solubility in water and a repeating structure (23). The polysaccharides in seaweed are divided into sulfated (fucoidan, carrageenan, and ulvan) and non-sulfated (agarose and laminarin) (24). Chlorophyta, Phaeophyta, and Rhodophyta contain polysaccharides of varied chemical composition and structure (25).
Previous studies suggested that polysaccharides from seaweed showed strong anti-CRC and preventive activities. They can either directly inhibit cancer cells or affect various phases of carcinogenesis and the progression of tumor through the regulation of the balance between proliferation and programmed cell death and can also be potentially used for cancer prophylaxis (26). Three polysaccharide fractions isolated from Porphyra haitanensis exerted inhibitory effects on growth in the HT-29, LoVo, and SW-480 colon cancer cell lines (27). Other active components contained in seaweed that exhibit similar effects against CRC are shown in Table 1. Another study reported on an evaluation of the anti-CRC activity of sulfated glucuronorhamnoxylan polysaccharides from Capsosiphon fulvescens (28). Polysaccharides from Jania rubens upregulated the gene expression of Bax, caspase 8, and P53 in human colon cancer Caco-2 cells (29). A summary of mechanisms for other active components from seaweed on anti-CRC is shown in Table 2.
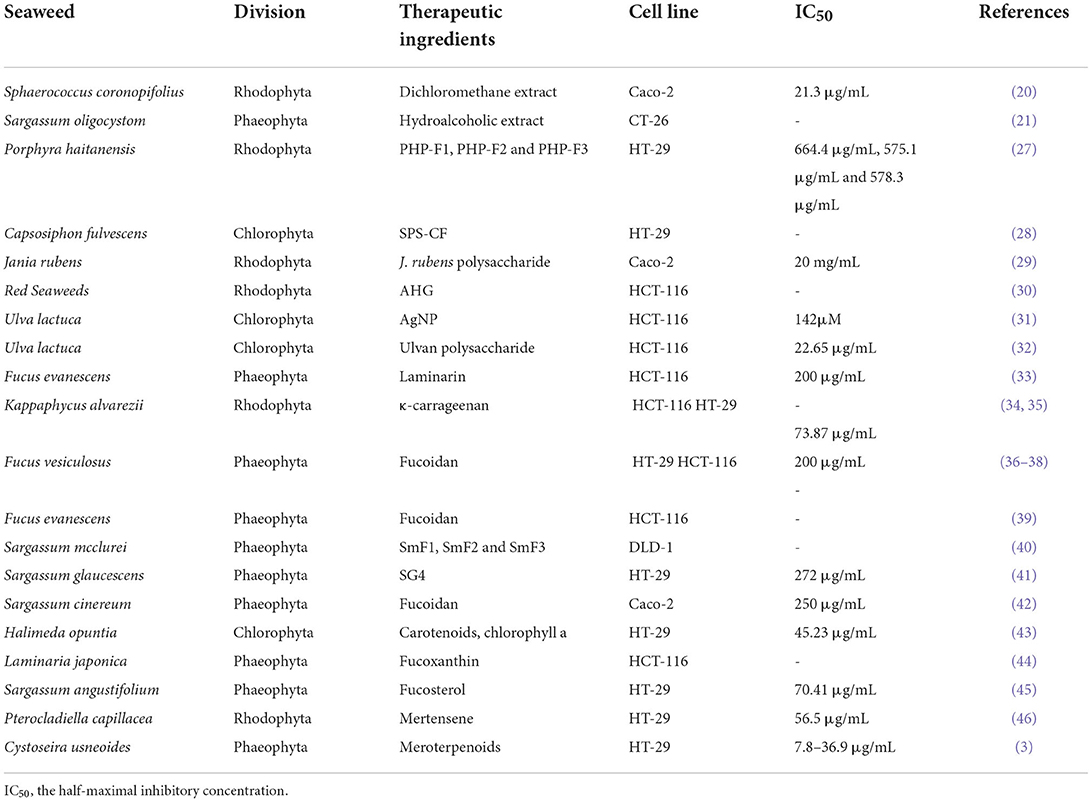
Table 1. The effects of active components isolated from seaweeds on colorectal cancer.
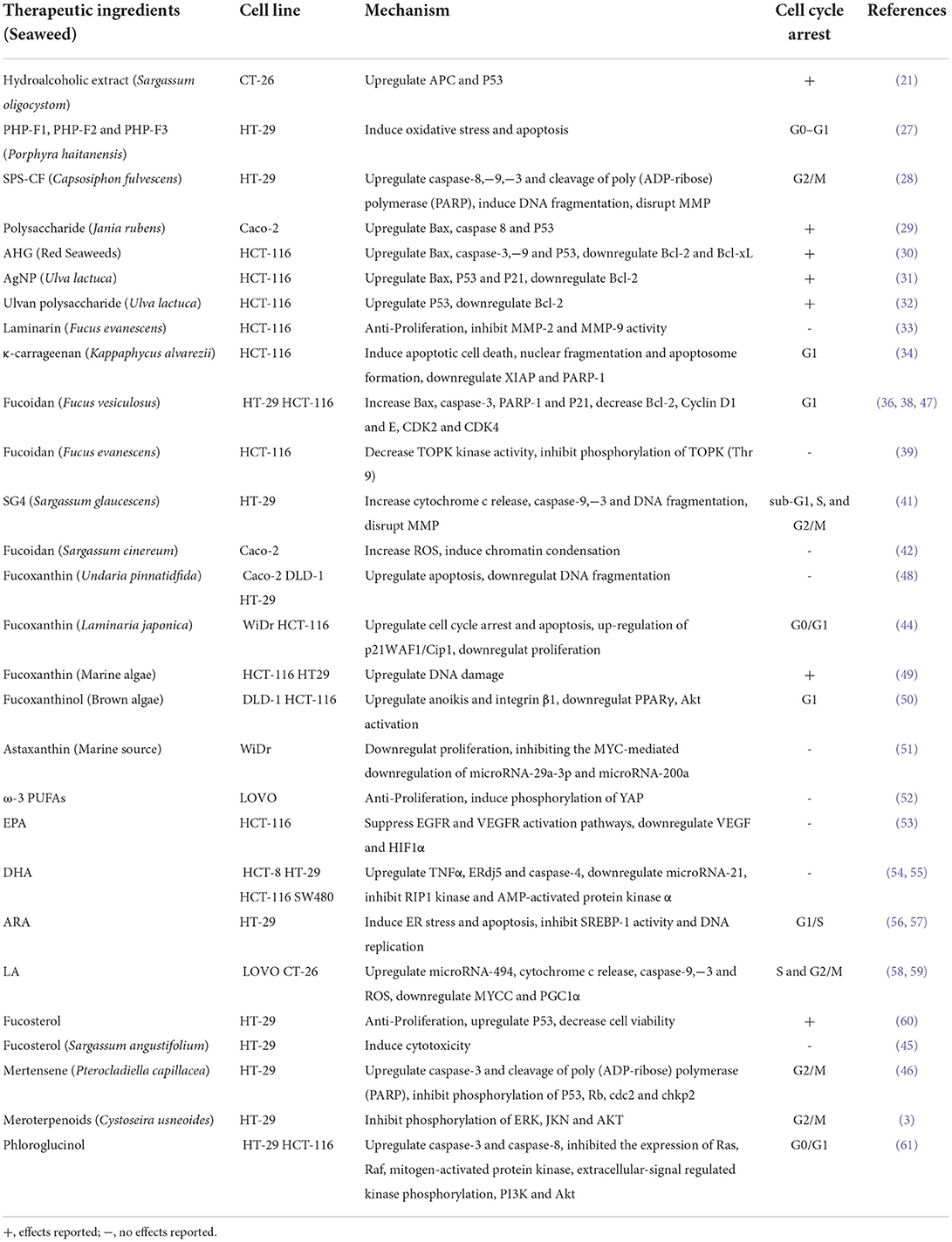
Table 2. Properties of active components isolated from seaweed against colorectal cancer.
Red algae cell walls mainly consist of agarose which is composed of alternative units of D-galactose and 3,6-anhydro-L-galactose (AHG) linked by alternating α-1,3- and β-1,4-glycosidic bonds (62). After being consumed, agarose is digested, fermented, and metabolized by intestinal microbiota in the human large intestine, which makes it unique among red algal polysaccharides (63). Clinical trials have suggested that people in Asia who regularly consume red seaweeds are at a lower risk of CRC, which is relevant to their daily intake of seaweeds (64). It is speculated that this effect may be related to biologically active agarose components enriched from red seaweed. Upregulation of caspase-3, Bax, and caspase-9 expression and downregulation of Bcl-2 and Bcl-xL were observed in HCT-116 cells after AHG treatment (30). Therefore, the growth of human colon cancer HCT-116 cells was effectively suppressed by AHG, indicating that AHG is a potential alternative as an anti-CRC agent.
The sulfated polysaccharide known as “ulvan” is extracted from green algae of the ulva species (35). Ulvan mainly consists of cellulose, xyloglucan, and glucuronan with various other types of sugars (65). It is reported to possess diverse physiological and bioactive activities, including anticancer activity (66). Ulvan has been demonstrated to decrease viability in cancer cells while leaving healthy cells unaffected (65). The current study has categorically proven that biogenic silver nanoparticles (AgNP), which were generated via an extract of the marine alga Ulva lactuca, can induce p53-dependent apoptosis in colon cancer HCT-116 cells (31, 32).
Laminarin, sometimes referred to as laminaran, which is an essential biodegradable and non-toxic polysaccharide isolated from the cell wall reservoirs of brown algae, has caught the interest of researchers (67). Laminarins are essentially a group of low-molecular-weight storage β-glucans consisting of (1,3)-β-D-glucan (68). (1,3)-β-D-glucopyranose residues with a few 6-O-branching on the main chain, and also several β-(1,6)-intra-chain links, which are abundant in their structures (69). β-glucans can promote cell apoptosis of colon cancer, and they may be beneficial natural agents for colon cancer treatment and chemoprevention (70). Additionally, certain studies have suggested that the biological activity of laminarin can be strengthened with particular chemical modifications (71). For example, Ji et al. (71) demonstrated that laminarin treated with sulfated provided a stronger antitumor effect compared with unmodified laminarin in human colorectal adenocarcinoma cells. The cell survival rate was significantly decreased after culturing with sulfated laminarin in LoVo cells. Apparently, peculiarities of the polysaccharide structure and sulfation contribute to the anticancer activity of laminarins. Malyarenko et al. (33) found that the antiproliferative activity of laminarins from Fucus evanescens was comparable to that of their sulfated derivatives. The anticancer effect of laminarin isolated from F. evanescens was stronger than that of its sulfated derivatives in HCT-116 colon carcinoma cells. Ji et al. (72) proved that laminarin increased the intracellular reactive oxygen species (ROS) level, increased intracellular Ca2+, decreased intracellular pH, and induced LoVo apoptosis through a mitochondrial pathway. A further study revealed that the expression of procaspase-8 and−3 was downregulated and the activity of caspase-8,−3,−6, and−7 was increased in human colon cancer LoVo cells through the TRAIL/DR pathway after treatment with laminarin (73). Thus, laminarin induces apoptosis in human colon cancer via the mitochondrial and DR pathways, indicating that laminarin is a potent anticancer agent.
A set of sulfated polysaccharides generically described as carrageenan is present in red algae, which is the major ingredient of cell walls and interstitial spaces, acting as structural compounds and supplying intercellular adhesion and signaling. The structural units of these natural polysaccharides are a mixture of sulfated linear galactans, which consist of disaccharides of α-(1,4)-linked D-galactopyranose (D) residues or 3,6-anhydrogalactopyranose (DA) and β-(1,3)-linked D-galactopyranose (G) residues (74). According to the concentration, position, and sulfation of 3,6-anhydrogalactose, they are categorized into κ, λ, ι, ν, m, and θ types (75). Polysaccharides with a molecular weight ranging from 500 to 1,000 kDa are present in most of them; however, up to 25% of them may contain polysaccharides at a molecular weight of <100 kDa (76). The significant anticancer and antitumor activities were found in the low-molecular-weight κ- and λ-carrageenan, probably attributed to their antiviral and antioxidant effects as well as the stimulation of immunity against tumors (77). Some studies have reported that the risk of colon cancer appears to be minimized with low-molecular-weight carrageenan, a type of functional food ingredient (34). Carrageenans from Gigartina pistillata (78), Apostichopus japonicus (79) and Kappaphycus alvarezii (80) have an anti-CRC effect on the colon cancer HT-29 cell line. Native carrageenan exerted high cell suppressive activity in colon cancer cells compared with commercial carrageenan. Raman et al. (34) examined the role of the κ-carrageenan-containing soluble dietary fiber fraction of red algae in human colon cancer HCT-116 cells.
However, conflicting studies have suggested that colitis and inflammation may be induced by carrageenan (81). Wei et al. (82) suggested that the existing intestinal inflammation was magnified and TNBS-induced intestinal inflammation was aggravated by κ-carrageenan via activating the TLR4-NF-κB and MAPK/ERK1/2 pathways, which indicates it might act as a potential pro-inflammatory factor. In addition, further studies from their group revealed that the LPS-induced inflammation can be synergistically activated by κ-carrageenan through the Bcl10-NF-κB pathway, as illustrated by the aggravation of Citrobacter freundii DBS100-induced colitis in mice treated with it (83). Mi et al. (84) investigated the effectiveness of the carrageenan intake form and host intestinal microecology on toxicity in C57BL/6J mice. The severity of colitis in high-fat diet-fed mice could be increased by native carrageenan from drinking water via decreasing the abundance of the anti-inflammatory bacterium Akkermansia muciniphila and increasing that of harmful bacteria. The inflammatory effect and secretion of proinflammatory cytokines in HT-29 cells can be increased and promoted by using the fermentation supernatants of κ-carrageenan oligosaccharides (85). The inflammatory property of κ-carrageenan oligosaccharides in the context of gut microbiota was evidenced by these results.
Sulfated L-fucose present in the fibrous cell walls and intercellular spaces of brown seaweeds is a major component of fucoidan, which belongs to a large family of marine sulfated polysaccharides (86, 87). Fucoidan is a heparin-like molecule with a simple chemical structure composed of a repeating unit of disaccharides containing α-1,3-linked fucose and α-1,4-linked fucose with branches linked at the C2 positions (86). Generally, L-fucose polymerized with sulfated ester groups is present substantially in fucoidan, while galactose, glucose, mannose, xylose, and glucuronic acid residues are found in only a small proportion (88). Derivatives of fucoidan with a molecular weight varying from 40 to 330 kDa have been examined for their anticancer activities (89). Numerous experiments have shown that tumor cell proliferation and its growth or metastasis can be counteracted by fucoidan via eliciting cell apoptosis and suppressing angiogenesis (90). Health can be promoted and gut dysbiosis can be treated by fucoidan, a potential intestinal microbiota modulator. In addition, the effects of induced apoptosis in CRC cells by fucoidan have been evaluated (91).
In HT-29 colon cancer cells, cell viability was reduced by fucoidan in a dose- and time-dependent manner through reducing the expression of CDK2, CDK4, and Cyclin D1 (36). Apoptosis was also mediated by fucoidan via inhibition of the IGF-I/IGF-IR/IRS-1/PI3K/AKT (92) and PI3K-Akt-mTOR signaling pathways (37). A further study found that cell cycle arrest at G1-phase was induced by fucoidan via the upregulation of p21WAF1 and downregulation of Cyclin D1/CDK4 and Cyclin E/CDK2 expression (47).
In HCT-116 human colon cancer cells, CDK activity was suppressed by fucoidan via combining the CDK inhibitor proteins p21 and p27 with the Cyclin/CDK complexes (38). EGF-induced neoplastic cell transformation was significantly inhibited by fucoidan from F. evanescens via suppressing the TOPK/ERK1/2/MSK 1 signaling axis (39). In colon cancer cells DLD-1, fucoidan from Sargassum mcclurei was observed to be less cytotoxic and inhibited colony formation (40).
Fucoidan strongly regulated the mitochondrial membrane in cancer cells. The evidence found that apoptosis was caused by fucoidan through MMP loss, an increase in cytochrome c release and DNA fragmentation, activated caspase-9 and−3, and an increasing percentage of early and late apoptotic cells in HT-29 cells. Other biological studies indicated that apoptosis was induced by SG4 via involvement of the Akt/mTOR/S6 pathway in HT-29 cells (41). Another report showed that fucoidan from Sargassum cinereum suppressed the proliferation of Caco-2 cells in a dose-dependent manner, increased ROS production, and augmented mitochondrial membrane permeability (42).
In an animal model, tumor morbidity and average tumor weight were reduced and cellular apoptosis was increased by the treatment of dietary fucoidan in 1,2-dimethylhydrazine (DMH)-induced colorectal carcinogenesis in rats. The expression of β-catenin, C-Myc, Cyclin D1, and Survivin was reduced by treatment with fucoidan, whereas the Hippo pathway was highly activated and the phosphorylation levels of Mst1, Mst2, LATS1, LATS2, and YAP were significantly upregulated (91).
Generally, marine seaweed and animals are rich in pigments, which are widely used in functional food and pharmaceutical industries (93). There are mainly three types of pigments in seaweed involving chlorophylls, carotenoids, and phycobiliproteins (94). The seaweed color was determined by the content and type of pigments (95). For instance, chlorophylls a and b contribute to the green color in green seaweed, whereas allophycocyanin and c-phycoerythrin are responsible for the red color in red seaweed. β-carotene and fucoxanthin (Fx) are commonly observed in brown seaweed (96).
Tumor cells from CRC patients grow uninhibited in the body and enter the blood vessels to spread systemically (97). The apoptosis mechanism is strongly associated with antioxidant properties. Hence, anticancer compounds generally have antioxidant, antiangiogenic, and anti-inflammatory effects to regulate tumor development (98). A recent report showed that the strong antioxidant activity of carotenoids and chlorophyll a from green seaweed Halimeda opuntia against HT-29 human colorectal adenocarcinoma was investigated. The finding suggests that natural pigments are potential anticancer ingredients (43).
Carotenoids give seaweed colors from yellow to orangish (99), which of them possess strong activities involving Fx, astaxanthin (AXT), and violaxanthin (100). The evidence showed that carotenoids inhibited the PI3K/Akt apoptosis pathway, eventually integrating the mitochondrial membrane (101).
Fx is one of the most famous lipophilic carotenoids in brown algae, which is responsible for the strong antitumor property. Clinical trials reported that Fx decreased the causing risk of CRC, which has been found to Fx possess potential anti-CRC activity via downregulation of tumor-related proteins (102).
In cell experiments, Fx caused a markably decrease of the survival rate in Caco-2, HT-29, DLD-1 (48), and HCT-116 colorectal carcinoma cells (103). It was found that the activation of apoptosis and fragmentation of DNA contributes to the anticancer effect of Fx. Fx showed a significant antiproliferative effect by controlling the level of signaling proteins such as MAPK, NF-κB, and caspase family (99, 104). The disruption of cell cycle arrest causes cell apoptosis. Researchers found that Fx regulated sub-G1 cell cycle arrest in WiDr colon cancer cells (44). Furthermore, Fx stimulated cell cycle arrest at the G0/G1 or G2/M phases and caused programmed cell death (104). It is deduced that Fx induced cell cycle arrest and caused programmed cell death through enhancing intercellular communication between tumor cells.
The derivative compound of Fx was found that exists stronger anticancer activity than Fx. Chemical structural factors significantly influence the antiproliferative properties of Fx. The anti-CRC effect of Fx-degrading compounds was evaluated in Caco-2 cells and its activity may result in partial structures (105). The 13-cis and 13′-cis isomers of Fx showed a markably cancer-preventive effect compared to other derivatives (106). In another study, the antiproliferative effects of 5-fluorouracil (5-Fu) and Fx were determined and compared in HCT-116 and HT-29 cells (49). Fucoxanthinol (FxOH) is the deacetylated type of Fx, which can induce DLD-1 cell apoptosis into anoikis-like changes through the distribution of FAK and integrin β1 expression (50). The mechanism of the anti-CRC effect of FxOH was evaluated via regulation of MAPK and STAT apoptosis pathways in HT-29 and HCT-116 cell lines (107), and through inhibiting NF-κB activation in CRC cells (108). According to the previous reports, we found that the anti-CRC activity of FxOH is stronger than Fx, and FxOH induced cancer cells through downregulation of MAPK, STAT, and NF-κB apoptosis pathways. The anticancer effects of Fx and FxOH on six types of CRC cell lines and twenty kinds of tissues from surgically resected clinical CRC specimens were determined as well (109).
In a CRC model animal experiment, a continuous 5-week oral administration of Fx-rich fraction strongly inhibited the number of colorectal adenocarcinomas in DSS-treated male mice (110). Additionally, Fx significantly suppressed colon cancer in azoxymethane-dextrane sodium sulfate (AOM/DSS) carcinogenic model mice (111). In an inflammation-associated CRC mouse model, after a 4-month period of Fx administration, the multiplicity of colorectal adenocarcinoma was strongly decreased via upregulated anoikis-like integrin β1low/−/cleaved caspase-3high cells in colonic mucosal crypts (112). Additionally, Fx also markedly decreased HSP70 protein in colorectal mucosal crypts for 15-week administration (113). In a 14-week administration in a CRC mouse model, Fx markedly decreased CCR1, pAKT(Ser473), Cyclin D1, and pSmad2 compared with untreated mice (114). Therefore, Fx had chemopreventive potency and therapy ability in the progression of colorectal carcinogenesis in mice.
AXT is mainly contained in seaweed, and showed anti-metastatic activity through inhibiting microRNA-29a-3p and microRNA-200a, thereby downregulating MMP2 and ZEB1 (51). Natural AXT isolated from Haematococcus pluvialis showed a significantly anti-CRC effect in a dose-dependent manner in HCT-116 cells by regulating the ratio of Bax/Bcl-2 and upregulating the phosphorylation of p38, JNK, and ERK1/2 (115). AXT induced programmed cell death in DMH-induced rat colon carcinogenesis by modifying NF-κB, COX-2, MMPs-2/9, Akt, and ERK-2 expressions (116). A finding observed that AXT inhibits the progression of colonic premalignant lesions in an obesity-associated colorectal carcinogenesis animal model by attenuating oxidative stress, reducing inflammation and NF-κB activation in the colonic mucosa (117). Hence, AXT is a potential cancer-preventive compound in the therapy of CRC.
Seaweeds are known as low-energy food. Despite their low lipid content, seaweeds contain ω-3 and ω-6 polyunsaturated fatty acids (PUFAs) as a significant portion of their lipids (118). PUFAs are the precursors of eicosanoids and all cell membrane components, which can effectively reduce the risk of cancer (119). Several studies have demonstrated that the colorectal tissue distribution of PUFAs is associated with CRC prognosis (120). There are studies suggesting the potential use of some oxidized metabolites of PUFAs as biomarkers of CRC (121).
PUFAs in seaweeds contain a substantial amount of ω-3 fatty acids as major components. Eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) in seaweed are important ω-3 fatty acids in the marine environment (122). At present, more and more epidemiology and experiments have verified the antitumor activity of ω-3 PUFAs (123). Consumption of diets rich in ω-3 PUFAs not only inhibits the proliferation of CRC cells, but also can be combined with chemotherapy to enhance their sensitivity (124).
PUFAs have pro-apoptotic and growth-inhibitory effects on cancer cells. In LoVo and RKO colon cancer cells, PUFAs can reduce the synthesis of PGE2 and LTB4, inhibit the expression of ALOX5, LTB4, mPGES, COX-2, and PGE2, and increase the expression of LXA4, thereby promoting apoptosis and inhibiting the growth of LoVo and RKO colon cancer cells (125).
In an animal model, dietary supplementation of ω-3 PUFAs increased CRC cell apoptosis and decreased the tumor incidence in AOM/DSS-induced CRC in mice. ω-3 PUFAs treatment activated the hippo pathway, with increased cytoplasmic retention and phosphorylation of YAP (mediated by LATS1 and MST1/2) and the levels of epoxydocosapentaenoic acids (52). Notably, dietary ω-3 PUFAs treatment suppressed the growth of MC38 colorectal tumors. In C57BL/6 mice, ω-3 PUFAs modulate eicosanoid and fatty acid metabolite profiles (126). Huang et al. (127) demonstrated that ω-3 PUFAs reduced tumor incidence in rats by regulation of the DNA methylation process. Together, these results support the notion that ω-3 PUFAs might contribute to the anti-CRC effects of seaweed.
There is evidence of the utility of ω-3 PUFA EPA in the treatment of CRC (128). In a clinical study, patients with familial adenomatous polyposis (FAP) were randomized to receive free fatty acid (FFA). Experimental data proved that EPA 2 g daily in the form of FFA has chemo-preventive efficacy in FAP patients (129). Recently, it has been shown that EPA can prevent FAP-related CRC by acting on several molecular mechanisms (129, 130).
The increased risk of colitis-associated colorectal cancer (CAC) is strongly associated with inflammatory bowel disease, but the effectiveness of dietary EPA-FFA in anti-inflammatory and anticancer activities is unclear. In EPA-FFA-treated AOM-DSS mice, Piazzi et al. (131) found an enrichment of Lactobacillus species in the gut microbiota, as well as restored Notch signaling and decreased nuclear β-catenin expression, while tumor cell apoptosis increased. Morin et al. (53) reported that the eicosapentaenoic acid monoglyceride (MAG-EPA) treatment increased HCT-116 cell apoptosis and decreased the tumor of a mouse xenograft model of HCT-116 via activating the vascular endothelial growth factor (VEGF) receptor pathway and decreasing the epidermal growth factor receptor (EGFR).
As an ω-3 PUFA, DHA has various biological properties, including anticancer activity (132). The anticancer effect of DHA might be a consequence of its ability to regulate the production of proinflammatory mediators in cancer cells and/or host cells, changing the inflammatory status of the systemic or tumor microenvironment (54, 133). Numerous studies have demonstrated that in CRC cells, the activity of DHA-triggered caspase family members is associated with apoptosis (134). Mechanisms including DHA-induced cellular protein expression explain the antitumor activity of DHA (55).
Fluckiger et al. (54) reported that the TNFα-dependent manner triggers apoptosis in HCT-116, HCT-8, and human CRC cells in DHA-mediated, and cells induced with DHA increased TNFα mRNA content by downregulating microRNA-21 expression, stating that the effect of TNFα on DHA-mediated apoptosis of colon cancer cells. Sarabi et al. (135) demonstrated that DHA could specifically attenuate promoter DNA methylation and VEGF protein levels of microRNA-126 in HCT-116, Caco-2, and CRC cells. Fasano et al. (55) reported that DHA treatment induced apoptosis in HT-29, HCT-116, and SW480 colon cancer cell lines and inhibited their total and surface GRP78 expression, suggesting that pERK1/2 could be the first upstream target of DHA. These studies offer insight into the epigenetic mechanisms by which DHA influences gene expression regulation in CRC cells.
The ω-6 PUFAs contained in seaweeds include mostly linoleic acid (LA) and arachidonic acid (ARA) (136). Previous studies have suggested that, unlike ω-3 PUFAs, the ω-6 PUFAs, especially ARA, are generally associated with many adverse effects on the human body, including the promotion of multiple cancer types, such as CRC (137, 138). However, there is little in vitro evidence to show that ARA exerts anticancer activity in CRC cells. Bae et al. (56) and González-Fernández et al. (57) evidenced that ARA may inhibit DNA replication and G1/S cell cycle transition and induce endoplasmic reticulum stress in HT-29 CRC cells, thereby suppressing cell viability and inducing apoptosis.
Research evidence shows that LA may be involved in both pro- and anticancer activities (137). Lu et al. (139) and Ohmori et al. (140) reported that LA at low concentrations (100-200 μM) reduced caspase-3 activation in CRC cells and promoted tumor cell proliferation. LA at high concentrations (above 300μM) enhanced ROS generation, caused mitochondrial dysfunction and inhibited tumor cell growth (58). A recent report from Ogata et al. (59) suggested that LA induced quiescence by promoting microRNA-494 expression, resulting in the dormancy of CT-26 CRC cells. Therefore, it is speculated that LA can inhibit the growth of CRC cells.
Phytosterols are the major nutritional components of seaweed and the most important chemical constituents of algae (141). Phytosterols are the major lipid components of plant cell biofilms. In the marine environment, brown algae are a major source of phytosterols, including brassicosterols with a small amount of plant cholesterol and fucosanols (142). Not only can phytosterols lower cholesterol, they also have strong anticancer activity, and several studies have shown that foods rich in phytosterols may help control the growth of many types of tumors (143).
β-Sitosterol, isolated from seaweed, is the most common dietary phytosterol and has a proven potential role in the treatment of CRC (144, 145). Shathviha et al. (146) reported on the evaluation of AgNP synthesized using β-sitosterol and its cytotoxic potential in HT-29 human colon cancer cells. β-Sitosterol-mediated AgNP treatments induced p53 expression and early apoptosis in HT-29 cells. Arul et al. (147) investigated the β-Sitosterol significantly reduces fecal bacteria and colonic bioconverting enzymes in mice with DMH-induced colon cancer, thereby preventing colon cancer development. Amplified activities of colonic biotransformation enzymes are considered hallmarks of colon carcinogenesis. Hence, β-sitosterol is a potential chemopreventive agent in colon carcinogenesis.
The major phytosterol in brown seaweed is fucosterol, which has various biological activities, including anticancer activity (148). A previous study indicated that oxygenated fucosterol inhibited HCT-116 human colon cancer cell growth with higher cytotoxicity than commercial cytotoxic drugs (149). Ramos et al. (60) reported that the combination of fucosterol with 5-Fu can enhance the toxic effect in HT-29 cells. Furthermore, fucosterol is not toxic to normal cells, indicating specificity for cancer cells. The hexane fraction of fucosterol produced by Sargassum angustifolium confirmed its cytotoxic activity against HT-29 (45).
Within the marine environment, terpenes synthesized by algae and secreted to the outside of cells to resist environmental insults are major secondary metabolites from seaweeds (150). It is a chemical compound that contains one or more isoprene units with strong anticancer activity (15). Epidemiological and experimental studies suggest that terpenes may be helpful in curbing the growth of a variety of cancer cells, including colon cells, and provide additional opportunities for cancer therapy (151). Terpenes can modulate pathophysiological processes such as the cell cycle, invasion, migration, proliferation, and apoptosis in different types of tumor cells (152), exhibiting a wide spectrum of antitumor activities (153).
Previous studies have reported that a pentahalogenated monoterpene, halomon, isolated from the red seaweed Portieria hornemannii, exhibited strong anticancer activity (154). The halogenated monoterpene mertensene from the red seaweed Pterocladiella capillacea (S.G. Gmelin) Santelices & Hommersand inhibited LS174 and HT-29 human colorectal adenocarcinoma cell lines by activating caspase-3 and NF-κB, Akt, and MAPK ERK-1/-2 pathways vitality (46). Terpenes from the brown seaweed Cystoseira usneoides have anticancer effects on HT-29 colon cancer cells by reducing the phosphorylation levels of JNK and ERK and inhibiting the ERK/JNK/AKT signaling pathway (3). These reports demonstrate the potential of terpenes as drug candidates for the treatment of colon cancer.
Phenolic agents are one of the most active compounds in seaweed. Polyphenols with their high molecular weight, such as phlorotannins, specifically exist in brown algae (155). Phlorotannins are composed of a number of phloroglucinol (Ph) monomeric units (156). Those active compounds play a pivotal role in anti-CRC effects, for instance, as apoptotic, anti-metastatic, and antiangiogenic proprieties. They inhibited CRC cell growth directly or indirectly through attenuated inflammatory cytokines and oxidative stress (157). In other reports, the anticancer effects of Ph on insulin-like growth factor-1 receptor signaling in HT-29 human colon cancer cells have been investigated. In addition, Ph inhibited the levels of Ras, mitogen-activated protein kinase, and mTOR (158). The polyphenol-rich agent showed a lower survival rate in CRC cells than the non-polyphenol-rich agent from seaweeds involving Laminaria japonica, U. lactuca, and Porphyra tenera. Additionally, the polyphenol-rich agent caused G0/G1 cell cycle arrest in HCT-116 cells (159). Phlorofucofuroeckol A (PFF-A) (160) and Ph (161), isolated from brown seaweed, decreased survival rates via activating the apoptosis pathway in CRC cells. Further, Ph decreased the survival rate dose-dependently and induced apoptosis in HT-29 cells, altering Bcl-2 and caspase family proteins (61). The evidence proved that phenolic agents play an important role in understanding the development of colon CRC.
Seaweed is an important source of various vitamins, among which vitamin C and vitamin D have strong anticancer activity (162). A previous study indicated a potential interplay of vitamin D and immune cells in the tumor microenvironment reduces CRC risk (163). Moreover, some research groups have reported an inverse association between vitamin D3 levels and CRC incidence, and that higher vitamin D3 levels reduce polyp recurrence and improve overall survival in CRC patients (164, 165). Tumor migration and proliferation were inhibited by vitamin D and its analogs in the colon of C57Bl/6 mice (166). Rawson et al. (167) suggested that vitamin D may alter CRC risk by mediating extracellular inhibition. Ferrer-Mayorga et al. (168) indicated that the vitamin D metabolite calcitriol promotes vitamin D receptor expression and inhibits fibroblasts (associated with colon cancer), resulting in anti-CRC effects. There is also a study on vitamin C from Yun et al. (169), who found that cultured CRC cells harboring KRAS or BRAF mutations were selectively killed when exposed to high levels of vitamin C.
The minerals in seaweed are 10–20 times those of land plants and are easily bio-accumulated from seawater (12, 170). The macronutrients (e.g., magnesium, potassium, calcium, and sodium) and microelements (e.g., selenium, iodine, iron, manganese, and zinc) contained in seaweed are important for normal physiological functioning of the body and have potential relevance in cancer treatment (171, 172). Numerous clinical and epidemiological studies indicated that the risk of CRC may be reduced with a higher intake of calcium and magnesium (173). Ali et al. (174) evaluated the antitumor effect of selenium against DMH-induced CRC in BALB/C mice and its effect on apoptosis and angiogenesis. The group treated with DMH plus selenium exhibited significantly lower expression of cloned caudal-type homeobox gene-2 and VEGF but a higher caspase-3 expression level than the DMH-treated group.
Many studies have demonstrated the effect and mechanism of red, green, and brown seaweeds in CRC-prevention and therapy. Various therapeutic compounds from seaweed involving large molecular polysaccharides and small molecular pigments, fatty acids, phytosterols, terpenes, phenolics, vitamins and minerals induce programmed cell death via various signaling pathways. Such treatments can alter the protein expression of Bax, caspases, Bcl-2, MAPK, NF-κB, VEGF, DNA methylation, and CDK inhibitor and induce changes in the cell cycle and the CRC cellular functions of adhesion, migration, and invasion. Important anti-CRC agents such as polysaccharides and fatty acids existed for their antiproliferative and anti-metastatic effects in vivo. Furthermore, terpenes and pigments inhibited CRC cell survival rate and induced programmed cell death via regulation of the Akt signaling pathway.
A few molecular alterations in human CRC cells were correspondingly observed in CRC animal models treated with seaweed. Consecutive studies in vitro will be important as the basis for clarifying the molecular mechanisms underlying cancer prevention in humans with CRC and CRC animal models. CRC animal models, AOM-DSS, BALB/C, and C57BL/6 mice, have been the vehicles for many discoveries concerning the anti-CRC effects of seaweed. The oxidation, inflammation and gut microbiota, which are significant factors associated with colorectal carcinogenesis, have been reported to be prime targets of various therapeutic compounds from seaweed. In addition, the administration of Fx induced anoikis in CRC animal models. However, the detailed molecular mechanisms underlying the cancer chemopreventive effect in animals remain poorly understood.
Finally, this review highlights the importance of seaweed as a potential agent candidate for preventing CRC. However, the underlying mechanisms remain elusive. Further clinical investigations are needed to assess the anticancer effect of seaweed in humans.
YD and ZP conceptualized the idea and scope of the review. YZ, XZ, HY, and KZ both performed a literature review. YF and DX wrote the manuscript and created the figures. All authors critically revised and edited the manuscript.
This research was funded by the Scientific and Technological Development Program of Jilin Province of China, grant numbers (20210401065YY and 20210204046YY).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. (2021) 71:209–49. doi: 10.3322/caac.21660
2. Rawla P, Sunkara T, Barsouk A. Epidemiology of colorectal cancer: incidence, mortality, survival, and risk factors. Prz Gastroenterol. (2019) 14:89–103. doi: 10.5114/pg.2018.81072
3. Zbakh H, Zubia E, Reyes CL, Calderon-Montano JM, Motilva V. Anticancer activities of meroterpenoids isolated from the brown alga cystoseira usneoides against the human colon cancer cells Ht-29. Foods. (2020) 9:300. doi: 10.3390/foods9030300
4. Davis CD, Milner JA. Molecular targets for nutritional preemption of cancer. Curr Cancer Drug Targets. (2007) 7:410–5. doi: 10.2174/156800907781386560
5. Huang G, Liu Z, He L, Luk KH, Cheung ST, Wong KH, et al. Autophagy is an important action mode for functionalized selenium nanoparticles to exhibit anti-colorectal cancer activity. Biomater Sci. (2018) 6:2508–17. doi: 10.1039/C8BM00670A
6. Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol. (2014) 740:364–78. doi: 10.1016/j.ejphar.2014.07.025
7. Feng Y, Wang N, Zhu M, Feng Y, Li H, Tsao S. Recent progress on anticancer candidates in patents of herbal medicinal products. Recent Pat Food Nutr Agric. (2011) 3:30–48. doi: 10.2174/2212798411103010030
8. Desideri D, Cantaluppi C, Ceccotto F, Meli MA, Roselli C, Feduzi L. Essential and toxic elements in seaweeds for human consumption. J Toxicol Environ Health A. (2016) 79:112–22. doi: 10.1080/15287394.2015.1113598
9. Kilinç B, Cirik S, Turan G, Tekogul H, Koru E. Seaweeds for food and industrial applications. In: Muzzalupo, editor. Food Industry IntechOpen. Izmir (2013). p. 735–48. doi: 10.5772/53172
10. Cotas J, Pacheco D, Gonçalves AMM, Silva P, Carvalho LG, Pereira L. Seaweeds' nutraceutical and biomedical potential in cancer therapy: a concise review. J Cancer Metast Treat. (2021) 7:13. doi: 10.20517/2394-4722.2020.134
11. Brown EM, Allsopp PJ, Magee PJ, Gill CI, Nitecki S, Strain CR, et al. Seaweed and Human Health. Nutr Rev. (2014) 72:205–16. doi: 10.1111/nure.12091
12. Makkar HP, Tran G, Heuzé V, Giger-Reverdin S, Lessire M, Lebas F, et al. Seaweeds for livestock diets: a review. Anim Feed Sci Technol. (2016) 212:1–17. doi: 10.1016/j.anifeedsci.2015.09.018
13. Arokiarajan MS, Thirunavukkarasu R, Joseph J, Ekaterina O, Aruni W. Advance research in biomedical applications on marine sulfated polysaccharide. Int J Biol Macromol. (2022) 194:870–81. doi: 10.1016/j.ijbiomac.2021.11.142
14. Murphy C, Hotchkiss S, Worthington J, McKeown SR. The potential of seaweed as a source of drugs for use in cancer chemotherapy. J Appl Phycol. (2014) 26:2211–64. doi: 10.1007/s10811-014-0245-2
15. Gutierrez-Rodriguez AG, Juarez-Portilla C, Olivares-Banuelos T, Zepeda RC. Anticancer activity of seaweeds. Drug Discov Today. (2018) 23:434–47. doi: 10.1016/j.drudis.2017.10.019
16. Namvar F, Baharara J, Mahdi AA. Antioxidant and anticancer activities of selected persian gulf algae. Indian J Clin Biochem. (2014) 29:13–20. doi: 10.1007/s12291-013-0313-4
17. Farooqi AA, Butt G, Razzaq Z. Algae extracts and methyl jasmonate anti-cancer activities in prostate cancer: choreographers of ‘the dance macabre'. Cancer Cell Int. (2012) 12:1–6. doi: 10.1186/1475-2867-12-50
18. Ryu MJ, Kim AD, Kang KA, Chung HS, Kim HS, Suh IS, et al. The green algae ulva fasciata delile extract induces apoptotic cell death in human colon cancer cells. In Vitro Cell Dev Biol Anim. (2013) 49:74–81. doi: 10.1007/s11626-012-9547-3
19. Zakaria AD, Basah K, Bahtiar A, Deviyani Z, Basah K, Bahtiar A. Cytotoxic activity of extract and active fraction of turbinaria decurrens bory on colon cancer cell Line Hct-116. Int J Morphol. (2018) 36:979–83. doi: 10.4067/S0717-95022018000300979
20. Alves C, Pinteus S, Rodrigues A, Horta A, Pedrosa R. Algae from portuguese coast presented high cytotoxicity and antiproliferative effects on an in vitro model of human colorectal cancer. Pharmacognosy Res. (2018) 10:24-30. doi: 10.4103/pr.pr_151_16
21. Ghazy E, Fadil AG, Hussen BM, Al-Marzoqi AH. Anticancer Effect of Sargassum Oligocystom Hydroalcoholic Extract against Sw742, Ht-29, Widr and Ct-26 Colorectal Cancer Cell Lines and Expression of P53 and Apc Genes. (2021). doi: 10.21203/rs.3.rs-906543/v1
22. Sakthivel R, Devi KP. Antioxidant, anti-inflammatory and anticancer potential of natural bioactive compounds from seaweeds. Stud Nat Prod Chem. (2019) 63:113–60. doi: 10.1016/B978-0-12-817901-7.00005-8
23. Pereira L. Seaweeds as source of bioactive substances and skin care therapy—cosmeceuticals, algotheraphy, and thalassotherapy. Cosmetics. (2018) 5:68. doi: 10.3390/cosmetics5040068
24. Hentati F, Tounsi L, Djomdi D, Pierre G, Delattre C, Ursu AV, et al. Bioactive polysaccharides from seaweeds. Molecules. (2020) 25:3152. doi: 10.3390/molecules25143152
25. Wang Z, Xie J, Shen M, Nie S, Xie M. Sulfated modification of polysaccharides: synthesis, characterization and bioactivities. Trends Food Sci Technol. (2018) 74:147–57. doi: 10.1016/j.tifs.2018.02.010
26. Fedorov SN, Ermakova SP, Zvyagintseva TN, Stonik VA. Anticancer and cancer preventive properties of marine polysaccharides: some results and prospects. Mar Drugs. (2013) 11:4876–901. doi: 10.3390/md11124876
27. Yao W-Z, Veeraperumal S, Qiu H-M, Chen X-Q, Cheong K-L. Anti-cancer effects of porphyra haitanensis polysaccharides on human colon cancer cells via cell cycle arrest and apoptosis without causing adverse effects in vitro. 3 Biotech. (2020) 10:1–11. doi: 10.1007/s13205-020-02379-y
28. Choi JW, Lee J, Kim SC, You S, Lee CW, Shin J, et al. Glucuronorhamnoxylan from capsosiphon fulvescens inhibits the growth of Ht-29 human colon cancer cells in vitro and in vivo via induction of apoptotic cell death. Int J Biol Macromol. (2019) 124:1060–8. doi: 10.1016/j.ijbiomac.2018.12.001
29. Gheda S, El-Sheekh M, Abou-Zeid A. In vitro anticancer activity of polysaccharide extracted from red alga jania rubens against breast and colon cancer cell lines. Asian Pac J Trop Med. (2018) 11:583. doi: 10.4103/1995-7645.244523
30. Yun EJ, Yu S, Kim YA, Liu JJ, Kang NJ, Jin YS, et al. In vitro prebiotic and anti-colon cancer activities of agar-derived sugars from red seaweeds. Mar Drugs. (2021) 19:213. doi: 10.3390/md19040213
31. Acharya D, Satapathy S, Yadav KK, Somu P, Mishra G. Systemic evaluation of mechanism of cytotoxicity in human colon cancer Hct-116 cells of silver nanoparticles synthesized using marine algae ulva lactuca extract. J Inorg Organomet Polym Mater. (2022) 32:596–605. doi: 10.1007/s10904-021-02133-8
32. Ahmed O, Ahmed R. Anti-proliferative and apoptotic efficacies of ulvan polysaccharides against different types of carcinoma cells. vitro and in vivo. J Cancer Sci Ther. (2014) 6:202–8. doi: 10.4172/1948-5956.1000272
33. Malyarenko OS, Usoltseva RV, Shevchenko NM, Isakov VV, Zvyagintseva TN, Ermakova SP. In vitro anticancer activity of the laminarans from far eastern brown seaweeds and their sulfated derivatives. J Appl Phycol. (2017) 29:543–53. doi: 10.1007/s10811-016-0915-3
34. Raman M, Doble M. κ-Carrageenan from marine red algae, kappaphycus alvarezii–a functional food to prevent colon carcinogenesis. J Funct Foods. (2015) 15:354–64. doi: 10.1016/j.jff.2015.03.037
35. Morelli A, Puppi D, Chiellini F. Perspectives on Biomedical Applications of Ulvan. Seaweed Polysaccharides. Elsevier (2017). p. 305–30. doi: 10.1016/B978-0-12-809816-5.00016-5
36. Yun CW, Yun S, Lee JH, Han Y-S, Yoon YM, An D, et al. Silencing prion protein in Ht29 human colorectal cancer cells enhances anticancer response to fucoidan. Anticancer Res. (2016) 36:4449–58. doi: 10.21873/anticanres.10989
37. Han YS, Lee JH, Lee SH. Fucoidan inhibits the migration and proliferation of Ht-29 human colon cancer cells via the phosphoinositide-3 Kinase/Akt/Mechanistic target of rapamycin pathways. Mol Med Rep. (2015) 12:3446–52. doi: 10.3892/mmr.2015.3804
38. Park HY, Park S-H, Jeong J-W, Yoon D, Han MH, Lee D-S, et al. Induction of P53-independent apoptosis and G1 cell cycle arrest by fucoidan in Hct116 human colorectal carcinoma cells. Mar Drugs. (2017) 15:154. doi: 10.3390/md15060154
39. Vishchuk OS, Sun H, Wang Z, Ermakova SP, Xiao J, Lu T, et al. Pdz-binding kinase/T-Lak cell-originated protein kinase is a target of the fucoidan from brown alga fucus evanescens in the prevention of Egf-induced neoplastic cell transformation and colon cancer growth. Oncotarget. (2016) 7:18763. doi: 10.18632/oncotarget.7708
40. Thinh PD, Menshova RV, Ermakova SP, Anastyuk SD, Ly BM, Zvyagintseva TN. Structural characteristics and anticancer activity of fucoidan from the brown alga sargassum mcclurei. Mar Drugs. (2013) 11:1456–76. doi: 10.3390/md11051456
41. Shiao W-C, Kuo C-H, Tsai Y-H, Hsieh S-L, Kuan A-W, Hong Y-H, et al. In vitro evaluation of anti-colon cancer potential of crude extracts of fucoidan obtained from sargassum glaucescens pretreated by compressional-puffing. Applied Sciences. (2020) 10:3058. doi: 10.3390/app10093058
42. Narayani SS, Saravanan S, Ravindran J, Ramasamy M, Chitra J. In vitro anticancer activity of fucoidan extracted from sargassum cinereum against Caco-2 cells. Int J Biol Macromol. (2019) 138:618–28. doi: 10.1016/j.ijbiomac.2019.07.127
43. Nazarudin M, Yasin I, Mazli N, Saadi A, Azizee M, Nooraini M, et al. Preliminary screening of antioxidant and cytotoxic potential of green seaweed, halimeda opuntia (linnaeus) lamouroux. Saudi J Biol Sci. (2022) 29:2698–705. doi: 10.1016/j.sjbs.2021.12.066
44. Das SK, Hashimoto T, Shimizu K, Yoshida T, Sakai T, Sowa Y, et al. Fucoxanthin induces cell cycle arrest at G0/G1 phase in human colon carcinoma cells through up-regulation of P21waf1/Cip1. Biochimica et Biophysica Acta. (2005) 1726:328–35. doi: 10.1016/j.bbagen.2005.09.007
45. Khanavi M, Gheidarloo R, Sadati N, Ardekani MRS, Nabavi SMB, Tavajohi S, et al. Cytotoxicity of fucosterol containing fraction of marine algae against breast and colon carcinoma cell line. Pharmacogn Mag. (2012) 8:60. doi: 10.4103/0973-1296.93327
46. Tarhouni-Jabberi S, Zakraoui O, Ioannou E, Riahi-Chebbi I, Haoues M, Roussis V, et al. Mertensene, a halogenated monoterpene, induces G2/M cell cycle arrest and caspase dependent apoptosis of human colon adenocarcinoma Ht29 cell line through the modulation of Erk-1/-2, Akt and Nf-κb signaling. Mar Drugs. (2017) 15:221. doi: 10.3390/md15070221
47. Han Y-s, Lee JH, Lee SH. Antitumor effects of fucoidan on human colon cancer cells via activation of Akt signaling. Biomol Ther. (2015) 23:225. doi: 10.4062/biomolther.2014.136
48. Hosokawa M, Kudo M, Maeda H, Kohno H, Tanaka T, Miyashita K. Fucoxanthin induces apoptosis and enhances the antiproliferative effect of the Pparγ ligand, troglitazone, on colon cancer cells. Biochimica et Biophysica Acta. (2004) 1675:113–9. doi: 10.1016/j.bbagen.2004.08.012
49. Lopes-Costa E, Abreu M, Gargiulo D, Rocha E, Ramos AA. Anticancer effects of seaweed compounds fucoxanthin and phloroglucinol, alone and in combination with 5-fluorouracil in colon cells. J Toxicol Environ Health A. (2017) 80:776–87. doi: 10.1080/15287394.2017.1357297
50. Terasaki M, Maeda H, Miyashita K, Mutoh M. Induction of anoikis in human colorectal cancer cells by fucoxanthinol. Nutr Cancer. (2017) 69:1043–52. doi: 10.1080/01635581.2017.1339814
51. Kim H-Y, Kim Y-M, Hong S. Astaxanthin suppresses the metastasis of colon cancer by inhibiting the Myc-mediated downregulation of microrna-29a-3p and microrna-200a. Sci Rep. (2019) 9:1–10. doi: 10.1038/s41598-019-45924-3
52. Zhang K, Hu Z, Qi H, Shi Z, Chang Y, Yao Q, et al. G-protein-coupled receptors mediate Ω-3 pufas-inhibited colorectal cancer by activating the hippo pathway. Oncotarget. (2016) 7:58315. doi: 10.18632/oncotarget.11089
53. Morin C, Rodríguez E, Blier PU, Fortin S. Potential application of eicosapentaenoic acid monoacylglyceride in the management of colorectal cancer. Mar Drugs. (2017) 15:283. doi: 10.3390/md15090283
54. Fluckiger A, Dumont A, Derangere V, Rebe C, De Rosny C, Causse S, et al. Inhibition of colon cancer growth by docosahexaenoic acid involves autocrine production of Tnfα. Oncogene. (2016) 35:4611–22. doi: 10.1038/onc.2015.523
55. Fasano E, Serini S, Piccioni E, Toesca A, Monego G, Cittadini AR, et al. Dha induces apoptosis by altering the expression and cellular location of Grp78 in colon cancer cell lines. Biochimica et Biophysica Acta. (2012) 1822:1762–72. doi: 10.1016/j.bbadis.2012.08.003
56. Bae S, Kim M-K, Kim HS, Moon Y-A. Arachidonic acid induces er stress and apoptosis in Ht-29 human colon cancer cells. Animal Cells Syst. (2020) 24:260–6. doi: 10.1080/19768354.2020.1813805
57. González-Fernández MJ, Fabrikov D, Ramos-Bueno RP, Guil-Guerrero JL, Ortea I. Swath differential abundance proteomics and cellular assays show in vitro anticancer activity of arachidonic acid-and docosahexaenoic acid-based monoacylglycerols in Ht-29 colorectal cancer cells. Nutrients. (2019) 11:2984. doi: 10.3390/nu11122984
58. Lu X, Yu H, Ma Q, Shen S, Das UN. Linoleic acid suppresses colorectal cancer cell growth by inducing oxidant stress and mitochondrial dysfunction. Lipids Health Dis. (2010) 9:1–11. doi: 10.1186/1476-511X-9-106
59. Ogata R, Mori S, Kishi S, Sasaki R, Iwata N, Ohmori H, et al. Linoleic acid upregulates microrna-494 to induce quiescence in colorectal cancer. Int J Mol Sci. (2021) 23:225. doi: 10.3390/ijms23010225
60. Ramos AA, Almeida T, Lima B, Rocha E. Cytotoxic activity of the seaweed compound fucosterol, alone and in combination with 5-fluorouracil, in colon cells using 2d and 3d culturing. J Toxicol Environ Health A. (2019) 82:537–49. doi: 10.1080/15287394.2019.1634378
61. Kang M-H, Kim I-H, Nam T-J. Phloroglucinol induces apoptosis via apoptotic signaling pathways in Ht-29 colon cancer cells. Oncol Rep. (2014) 32:1341–6. doi: 10.3892/or.2014.3355
62. Knutsen S, Myslabodski D, Larsen B, Usov A. A Modified System of Nomenclature for Red Algal Galactans. (1994). doi: 10.1515/botm.1994.37.2.163
63. Shang Q, Jiang H, Cai C, Hao J, Li G, Yu G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: an overview. Carbohydr Polym. (2018) 179:173–85. doi: 10.1016/j.carbpol.2017.09.059
64. Kim J, Lee J, Oh JH, Chang HJ, Sohn DK, Shin A, et al. Associations among dietary seaweed intake, C-Myc Rs6983267 polymorphism, and risk of colorectal cancer in a korean population: a case–control study. Eur J Nutr. (2020) 59:1963–74. doi: 10.1007/s00394-019-02046-w
65. Cindana Mo'o FR, Wilar G, Devkota HP, Wathoni N. Ulvan, a polysaccharide from macroalga ulva sp.: a review of chemistry, biological activities and potential for food and biomedical applications. Appl Sci. (2020) 10:5488. doi: 10.3390/app10165488
66. Thanh TTT, Quach TMT, Nguyen TN, Luong DV, Bui ML, Van Tran TT. Structure and cytotoxic activity of ulvan extracted from green seaweed ulva lactuca. Int J Biol Macromol. (2016) 93:695–702. doi: 10.1016/j.ijbiomac.2016.09.040
67. Zargarzadeh M, Amaral AJ, Custódio CA, Mano JF. Biomedical applications of laminarin. Carbohydr Polym. (2020) 232:115774. doi: 10.1016/j.carbpol.2019.115774
68. Rioux L-E, Turgeon SL, Beaulieu M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr Polym. (2007) 69:530–7. doi: 10.1016/j.carbpol.2007.01.009
69. Kadam SU, Tiwari BK, O'Donnell CP. Extraction, structure and biofunctional activities of laminarin from brown algae. Int J Food Sci Technol. (2015) 50:24–31. doi: 10.1111/ijfs.12692
70. Kim M-J, Hong S-Y, Kim S-K, Cheong C, Park H-J, Chun H-K, et al. β-Glucan enhanced apoptosis in human colon cancer cells Snu-C4. Nutr Res Pract. (2009) 3:180–4. doi: 10.4162/nrp.2009.3.3.180
71. Ji C-F, Ji Y-B, Meng D-Y. Sulfated modification and anti-tumor activity of laminarin. Exp Ther Med. (2013) 6:1259–64. doi: 10.3892/etm.2013.1277
72. Ji YB, Ji CF, Zhang H. Laminarin induces apoptosis of human colon cancer lovo cells through a mitochondrial pathway. Molecules. (2012) 17:9947–60. doi: 10.3390/molecules17089947
73. Ji CF, Ji YB. Laminarin-induced apoptosis in human colon cancer lovo cells. Oncol Lett. (2014) 7:1728–32. doi: 10.3892/ol.2014.1952
74. Khotimchenko M, Tiasto V, Kalitnik A, Begun M, Khotimchenko R, Leonteva E, et al. Antitumor potential of carrageenans from marine red algae. Carbohydr Polym. (2020) 246:116568. doi: 10.1016/j.carbpol.2020.116568
75. Necas J, Bartosikova L. Carrageenan: a review. Vet Med. (2013) 58:187–205. doi: 10.17221/6758-VETMED
76. Lascombes C, Agoda-Tandjawa G, Boulenguer P, Le Garnec C, Gilles M, Mauduit S, et al. Starch-carrageenan interactions in aqueous media: role of each polysaccharide chemical and macromolecular characteristics. Food Hydrocoll. (2017) 66:176–89. doi: 10.1016/j.foodhyd.2016.11.025
77. Zhou G, Sun Y, Xin H, Zhang Y, Li Z, Xu Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from chondrus ocellatus. Pharmacol Res. (2004) 50:47–53. doi: 10.1016/j.phrs.2003.12.002
78. Cotas J, Marques V, Afonso MB, Rodrigues CM, Pereira L. Carrageenans Extracted from Gigartina Pistillata Demonstrate Potential against Colorectal Cancer Stem Cell-Enriched Tumorspheres. In: Conference: Conferência APAA 2019 - Associação Portuguesa de Algologia Aplicada (APAA). Coimbra; Lisbon (2019). doi: 10.13140/RG.2.2.17470.02889
79. Yu-lin D, Dong-yue Z, Yun-Fei J, Fei Z, Hao Y, You-Jin J. 6-Bromohypaphorine isolated from red sea cucumbers apostichopus japonicus exhibits potent anticancer activity in A549 cancer cell line. Chin J Analyt Chemist. (2021) 49:37–42. doi: 10.1016/j.cjac.2021.05.001
80. Suganya AM, Sanjivkumar M, Chandran MN, Palavesam A, Immanuel G. Pharmacological importance of sulphated polysaccharide carrageenan from red seaweed kappaphycus alvarezii in comparison with commercial carrageenan. Biomed Pharmacother. (2016) 84:1300–12. doi: 10.1016/j.biopha.2016.10.067
81. Benard C, Cultrone A, Michel C, Rosales C, Segain J-P, Lahaye M, et al. Degraded carrageenan causing colitis in rats induces tnf secretion and Icam-1 upregulation in monocytes through Nf-κb activation. PLoS ONE. (2010) 5:e8666. doi: 10.1371/journal.pone.0008666
82. Wei W, Feng W, Xin G, Tingting N, Zhanghe Z, Haimin C, et al. Enhanced effect of κ-Carrageenan on Tnbs-induced inflammation in mice. Int Immunopharmacol. (2016) 39:218–28. doi: 10.1016/j.intimp.2016.07.031
83. Wu W, Zhen Z, Niu T, Zhu X, Gao Y, Yan J, et al. κ-Carrageenan enhances lipopolysaccharide-induced interleukin-8 secretion by stimulating the Bcl10-Nf-κb pathway in Ht-29 cells and aggravates C. freundii-induced inflammation in mice. Mediat Inflamm. (2017) 2017:8634865. doi: 10.1155/2017/8634865
84. Mi Y, Chin YX, Cao WX, Chang YG, Lim PE, Xue CH, et al. Native κ-Carrageenan induced-colitis is related to host intestinal microecology. Int J Biol Macromol. (2020) 147:284–94. doi: 10.1016/j.ijbiomac.2020.01.072
85. Sun Y, Cui X, Duan M, Ai C, Song S, Chen X. In vitro fermentation of κ-carrageenan oligosaccharides by human gut microbiota and its inflammatory effect on ht29 cells. J Funct Foods. (2019) 59:80–91. doi: 10.1016/j.jff.2019.05.036
86. Hsu H-Y, Hwang P-A. Clinical applications of fucoidan in translational medicine for adjuvant cancer therapy. Clin Transl Med. (2019) 8:1–18. doi: 10.1186/s40169-019-0234-9
87. Dai Y-L, Jiang Y-F, Lu Y-A, Kang M-C, Jeon Y-J. Fucoidan from acid-processed hizikia fusiforme attenuates oxidative damage and regulate apoptosis. Int J Biol Macromol. (2020) 160:390–7. doi: 10.1016/j.ijbiomac.2020.05.143
88. Chollet L, Saboural P, Chauvierre C, Villemin J-N, Letourneur D, Chaubet F. Fucoidans in nanomedicine. Mar Drugs. (2016) 14:145. doi: 10.3390/md14080145
89. Takahashi H, Kawaguchi M, Kitamura K, Narumiya S, Kawamura M, Tengan I, et al. An exploratory study on the anti-inflammatory effects of fucoidan in relation to quality of life in advanced cancer patients. Integr Cancer Ther. (2018) 17:282–91. doi: 10.1177/1534735417692097
90. Fitton JH, Stringer DN, Karpiniec SS. Therapies from fucoidan: an update. Mar Drugs. (2015) 13:5920–46. doi: 10.3390/md13095920
91. Xue M, Liang H, Ji X, Zhou Z, Liu Y, Sun T, et al. Effects of fucoidan on gut flora and tumor prevention in 1, 2-dimethylhydrazine-induced colorectal carcinogenesis. J Nutr Biochem. (2020) 82:108396. doi: 10.1016/j.jnutbio.2020.108396
92. Kim I-H, Nam T-J. Fucoidan downregulates insulin-like growth factor-I receptor levels in Ht-29 human colon cancer cells. Oncol Rep. (2018) 39:1516–22. doi: 10.3892/or.2018.6193
93. Pangestuti R, Kim S-K. Biological activities and health benefit effects of natural pigments derived from marine algae. J Funct Foods. (2011) 3:255–66. doi: 10.1016/j.jff.2011.07.001
94. Osório C, Machado S, Peixoto J, Bessada S, Pimentel FBC, Alves R, et al. Pigments content (chlorophylls, fucoxanthin and phycobiliproteins) of different commercial dried algae. Separations. (2020) 7:33. doi: 10.3390/separations7020033
95. Pérez MJ, Falqué E, Domínguez H. Antimicrobial action of compounds from marine seaweed. Mar Drugs. (2016) 14:52. doi: 10.3390/md14030052
96. Wang H-MD, Li X-C, Lee D-J, Chang J-S. Potential biomedical applications of marine algae. Bioresour Technol. (2017) 244:1407–15. doi: 10.1016/j.biortech.2017.05.198
97. Martínez Andrade KA, Lauritano C, Romano G, Ianora A. Marine microalgae with anti-cancer properties. Mar Drugs. (2018) 16:165. doi: 10.3390/md16050165
98. Smyrniotopoulos V, Vagias C, Bruyère C, Lamoral-Theys D, Kiss R, Roussis V. Structure and in vitro antitumor activity evaluation of brominated diterpenes from the red alga sphaerococcus coronopifolius. Bioorg Med Chem. (2010) 18:1321–30. doi: 10.1016/j.bmc.2009.12.025
99. Bandaranayake WM. The nature and role of pigments of marine invertebrates. Nat Prod Rep. (2006) 23:223–55. doi: 10.1039/b307612c
100. Manivasagan P, Bharathiraja S, Santha Moorthy M, Mondal S, Seo H, Dae Lee K, et al. Marine natural pigments as potential sources for therapeutic applications. Crit Rev Biotechnol. (2018) 38:745–61. doi: 10.1080/07388551.2017.1398713
101. Sathasivam R, Ki J-S. A Review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar Drugs. (2018) 16:26. doi: 10.3390/md16010026
102. Martin LJ. Fucoxanthin and its metabolite fucoxanthinol in cancer prevention and treatment. Mar Drugs. (2015) 13:4784–98. doi: 10.3390/md13084784
103. Chuyen HV, Eun J-B. Marine carotenoids: bioactivities and potential benefits to human health. Crit Rev Food Sci Nutr. (2017) 57:2600–10. doi: 10.1080/10408398.2015.1063477
104. Mondal A, Bose S, Banerjee S, Patra JK, Malik J, Mandal SK, et al. Marine cyanobacteria and microalgae metabolites—a rich source of potential anticancer drugs. Mar Drugs. (2020) 18:476. doi: 10.3390/md18090476
105. Komba S, Kotake-Nara E, Tsuzuki W. Degradation of fucoxanthin to elucidate the relationship between the fucoxanthin molecular structure and its antiproliferative effect on Caco-2 cells. Mar Drugs. (2018) 16:275. doi: 10.3390/md16080275
106. Nakazawa Y, Sashima T, Hosokawa M, Miyashita K. Comparative evaluation of growth inhibitory effect of stereoisomers of fucoxanthin in human cancer cell lines. J Funct Foods. (2009) 1:88–97. doi: 10.1016/j.jff.2008.09.015
107. Terasaki M, Mima M, Kudoh S, Endo T, Maeda H, Hamada J, et al. Glycine and succinic acid are effective indicators of the suppression of epithelial-mesenchymal transition by fucoxanthinol in colorectal cancer stem-like cells. Oncol Rep. (2018) 40:414–24. doi: 10.3892/or.2018.6398
108. Tamura S, Narita T, Fujii G, Miyamoto S, Hamoya T, Kurokawa Y, et al. Inhibition of Nf-Kappab transcriptional activity enhances fucoxanthinol-induced apoptosis in colorectal cancer cells. Genes Environ. (2019) 41:1–8. doi: 10.1186/s41021-018-0116-1
109. Takahashi K, Hosokawa M, Kasajima H, Hatanaka K, Kudo K, Shimoyama N, et al. Anticancer effects of fucoxanthin and fucoxanthinol on colorectal cancer cell lines and colorectal cancer tissues. Oncol Lett. (2015) 10:1463–7. doi: 10.3892/ol.2015.3380
110. Terasaki M, Hamoya T, Kubota A, Kojima H, Tanaka T, Maeda H, et al. Fucoxanthin prevents colorectal cancer development in dextran sodium sulfate-treated Apcmin/+ Mice. Anticancer Res. (2021) 41:1299–305. doi: 10.21873/anticanres.14887
111. Terasaki M, Uehara O, Ogasa S, Sano T, Kubota A, Kojima H, et al. Alteration of fecal microbiota by fucoxanthin results in prevention of colorectal cancer in Aom/Dss mice. Carcinogenesis. (2021) 42:210–9. doi: 10.1093/carcin/bgaa100
112. Terasaki M, Ikuta M, Kojima H, Tanaka T, Maeda H, Miyashita K, et al. Dietary fucoxanthin induces anoikis in colorectal adenocarcinoma by suppressing integrin signaling in a murine colorectal cancer model. J Clin Med. (2019) 9:90. doi: 10.3390/jcm9010090
113. Terasaki M, Murase W, Kamakura Y, Kawakami S, Kubota A, Kojima H, et al. A biscuit containing fucoxanthin prevents colorectal carcinogenesis in mice. Nutr Cancer. (2022) 2022:1–11. doi: 10.1080/01635581.2022.2086703
114. Terasaki M, Ono S, Hashimoto S, Kubota A, Kojima H, Ohta T, et al. Suppression of Cc chemokine receptor 1 is a key regulation for colon cancer chemoprevention in Aom/Dss mice by fucoxanthin. J Nutr Biochem. (2022) 99:108871. doi: 10.1016/j.jnutbio.2021.108871
115. Palozza P, Torelli C, Boninsegna A, Simone R, Catalano A, Mele MC, et al. Growth-inhibitory effects of the astaxanthin-rich alga haematococcus pluvialis in human colon cancer cells. Cancer Lett. (2009) 283:108–17. doi: 10.1016/j.canlet.2009.03.031
116. Nagendraprabhu P, Sudhandiran G. Astaxanthin inhibits tumor invasion by decreasing extracellular matrix production and induces apoptosis in experimental rat colon carcinogenesis by modulating the expressions of Erk-2, Nfκb and Cox-2. Invest New Drugs. (2011) 29:207–24. doi: 10.1007/s10637-009-9342-5
117. Kochi T, Shimizu M, Sumi T, Kubota M, Shirakami Y, Tanaka T, et al. Inhibitory effects of astaxanthin on azoxymethane-induced colonic preneoplastic lesions in C57/Bl/Ksj-Db/Dbmice. BMC Gastroenterol. (2014) 14:1–10. doi: 10.1186/s12876-014-0212-z
118. Mišurcová L, AmbroŽová J, Samek D. Seaweed lipids as nutraceuticals. Adv Food Nutr Res. (2011) 64:339–55. doi: 10.1016/B978-0-12-387669-0.00027-2
119. Zárate R, el Jaber-Vazdekis N, Tejera N, Pérez JA, Rodríguez C. Significance of long chain polyunsaturated fatty acids in human health. Clin Translat Med. (2017) 6:1–19. doi: 10.1186/s40169-017-0153-6
120. Zhang J, Zhang L, Ye X, Chen L, Zhang L, Gao Y, et al. Characteristics of fatty acid distribution is associated with colorectal cancer prognosis. Prostaglandins Leukot Essent Fatty Acids. (2013) 88:355–60. doi: 10.1016/j.plefa.2013.02.005
121. Zhang L-j, Chen B, Zhang J-j, Li J, Yang Q, Zhong Q-s, et al. Serum polyunsaturated fatty acid metabolites as useful tool for screening potential biomarker of colorectal cancer. Prostaglandins Leukot Essent Fatty Acids. (2017) 120:25–31. doi: 10.1016/j.plefa.2017.04.003
122. Rajapakse N, Kim S-K. Nutritional and digestive health benefits of seaweed. Adv Food Nutr Res. (2011) 64:17–28. doi: 10.1016/B978-0-12-387669-0.00002-8
123. Cockbain A, Toogood G, Hull MA. Omega-3 polyunsaturated fatty acids for the treatment and prevention of colorectal cancer. Gut. (2012) 61:135–49. doi: 10.1136/gut.2010.233718
124. Granci V, Cai F, Lecumberri E, Clerc A, Dupertuis YM, Pichard C. Colon cancer cell chemosensitisation by fish oil emulsion involves apoptotic mitochondria pathway. Br J Nutr. (2013) 109:1188–95. doi: 10.1017/S000711451200308X
125. Zhang C, Yu H, Ni X, Shen S, Das UN. Growth inhibitory effect of polyunsaturated fatty acids (Pufas) on colon cancer cells via their growth inhibitory metabolites and fatty acid composition changes. PLoS ONE. (2015) 10:e0123256. doi: 10.1371/journal.pone.0123256
126. Wang W, Yang J, Nimiya Y, Lee KSS, Sanidad K, Qi W, et al. Ω-3 Polyunsaturated fatty acids and their cytochrome P450-derived metabolites suppress colorectal tumor development in mice. J Nutr Biochem. (2017) 48:29–35. doi: 10.1016/j.jnutbio.2017.06.006
127. Huang Q, Wen J, Chen G, Ge M, Gao Y, Ye X, et al. Omega-3 polyunsaturated fatty acids inhibited tumor growth via preventing the decrease of genomic dna methylation in colorectal cancer rats. Nutr Cancer. (2016) 68:113–9. doi: 10.1080/01635581.2016.1115526
128. Hawcroft G, Volpato M, Marston G, Ingram N, Perry SL, Cockbain AJ, et al. The omega-3 polyunsaturated fatty acid eicosapentaenoic acid inhibits mouse mc-26 colorectal cancer cell liver metastasis via inhibition of Pge2-dependent cell motility. Br J Pharmacol. (2012) 166:1724–37. doi: 10.1111/j.1476-5381.2012.01882.x
129. West NJ, Clark SK, Phillips RK, Hutchinson JM, Leicester RJ, Belluzzi A, et al. Eicosapentaenoic acid reduces rectal polyp number and size in familial adenomatous polyposis. Gut. (2010) 59:918–25. doi: 10.1136/gut.2009.200642
130. Cockbain AJ, Volpato M, Race AD, Munarini A, Fazio C, Belluzzi A, et al. Anticolorectal cancer activity of the omega-3 polyunsaturated fatty acid eicosapentaenoic acid. Gut. (2014) 63:1760–8. doi: 10.1136/gutjnl-2013-306445
131. Piazzi G, D'Argenio G, Prossomariti A, Lembo V, Mazzone G, Candela M, et al. Eicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on notch signaling and gut microbiota. Int J Cancer. (2014) 135:2004–13. doi: 10.1002/ijc.28853
132. D'Eliseo D, Velotti F. Omega-3 fatty acids and cancer cell cytotoxicity: implications for multi-targeted cancer therapy. J Clin Med. (2016) 5:15. doi: 10.3390/jcm5020015
133. Calviello G, Di Nicuolo F, Gragnoli S, Piccioni E, Serini S, Maggiano N, et al. N-3 Pufas reduce vegf expression in human colon cancer cells modulating the Cox-2/Pge 2 induced Erk-1 and-2 and Hif-1α induction pathway. Carcinogenesis. (2004) 25:2303–10. doi: 10.1093/carcin/bgh265
134. Giros A, Grzybowski M, Sohn VR, Pons E, Fernandez-Morales J, Xicola RM, et al. Regulation of colorectal cancer cell apoptosis by the n-3 polyunsaturated fatty acids docosahexaenoic and eicosapentaenoic. Cancer Prev Res. (2009) 2:732–42. doi: 10.1158/1940-6207.CAPR-08-0197
135. Moradi Sarabi M, Zahedi SA, Pajouhi N, Khosravi P, Bagheri S, Ahmadvand H, et al. The effects of dietary polyunsaturated fatty acids on Mir-126 promoter DNA methylation status and vegf protein expression in the colorectal cancer cells. Genes Nutr. (2018) 13:1–9. doi: 10.1186/s12263-018-0623-5
136. Rodríguez-González I, Díaz-Reinoso B, Domínguez H. Intensification strategies for the extraction of polyunsaturated fatty acids and other lipophilic fractions from seaweeds. Food Bioproc Technol. (2022) 15:1–20. doi: 10.1007/s11947-021-02757-1
137. Xu Y, Qian SY. Anti-cancer activities of Ω-6 polyunsaturated fatty acids. Biomed J. (2014) 37:112-9. doi: 10.4103/2319-4170.131378
138. Jones R, Adel-Alvarez L-A, Alvarez OR, Broaddus R, Das S. Arachidonic acid and colorectal carcinogenesis. Mol Cell Biochem. (2003) 253:141–9. doi: 10.1023/A:1026060426569
139. Lu X-f, He G-q, Yu H-n, Ma Q, Shen S-r, Das UN. Colorectal cancer cell growth inhibition by linoleic acid is related to fatty acid composition changes. J Zhejiang Univ Sci B. (2010) 11:923–30. doi: 10.1631/jzus.B1000125
140. Ohmori H, Luo Y, Fujii K, Sasahira T, Shimomoto T, Denda A, et al. Dietary linoleic acid and glucose enhances azoxymethane-induced colon cancer and metastases via the expression of high-mobility group box 1. Pathobiology. (2010) 77:210–7. doi: 10.1159/000296305
141. Pal A, Kamthania MC, Kumar A. Bioactive compounds and properties of seaweeds—a review. Open Access Library J. (2014) 1:1–17. doi: 10.4236/oalib.1100752
142. Sohn S-I, Rathinapriya P, Balaji S, Jaya Balan D, Swetha TK, Durgadevi R, et al. Phytosterols in seaweeds: an overview on biosynthesis to biomedical applications. Int J Mol Sci. (2021) 22:12691. doi: 10.3390/ijms222312691
143. Jiang L, Zhao X, Xu J, Li C, Yu Y, Wang W, et al. The protective effect of dietary phytosterols on cancer risk: a systematic meta-analysis. J Oncol. (2019) 2019:7479518. doi: 10.1155/2019/7479518
144. Sánchez-Machado D, López-Hernández J, Paseiro-Losada P, López-Cervantes J. An Hplc method for the quantification of sterols in edible seaweeds. Biomed Chromatogr. (2004) 18:183–90. doi: 10.1002/bmc.316
145. Wang Z, Zhan Y, Xu J, Wang Y, Sun M, Chen J, et al. β-Sitosterol reverses multidrug resistance via bcrp suppression by inhibiting the P53–Mdm2 interaction in colorectal cancer. J Agric Food Chem. (2020) 68:3850–8. doi: 10.1021/acs.jafc.0c00107
146. Shathviha PC, Ezhilarasan D, Rajeshkumar S, Selvaraj J. β-Sitosterol mediated silver nanoparticles induce cytotoxicity in human colon cancer Ht-29 cells. Avicenna J Med Biotechnol. (2021) 13:42-6. doi: 10.18502/ajmb.v13i1.4577
147. Arul AB, Al Numair K, Al Saif M, Savarimuthu I. Effect of Dietary β-Sitosterol on Fecal Bacterial and Colonic Biotransformation Enzymes in 1, 2-Dimethylhydrazine–Induced Colon Carcinogenesis. Turkish J Med Sci. (2012) 42:1307–13. doi: 10.3906/sag-1106-16
148. Abdul QA, Choi RJ, Jung HA, Choi JS. Health benefit of fucosterol from marine algae: a review. J Sci Food Agric. (2016) 96:1856–66. doi: 10.1002/jsfa.7489
149. Lee D-G, Park J-H, Yoo K-H, Chung I-S, Lee Y-H, Lee J-K, et al. 24-Ethylcholesta-4, 24 (28)-Dien-3, 6-Dione from osmanthus fragrans var. aurantiacus flowers inhibits the growth of human colon cancer cell line, Hct-116. J Korean Soc Appl Biol Chemist. (2011) 54:206–10. doi: 10.3839/jksabc.2011.034
150. Leal MC, Munro MH, Blunt JW, Puga J, Jesus B, Calado R, et al. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat Prod Rep. (2013) 30:1380–90. doi: 10.1039/c3np70057g
151. Rabi T, Bishayee A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res Treat. (2009) 115:223–39. doi: 10.1007/s10549-008-0118-y
152. Rocha DH, Seca AM, Pinto DC. Seaweed secondary metabolites in vitro and in vivo anticancer activity. Mar Drugs. (2018) 16:410. doi: 10.3390/md16110410
153. Huang M, Lu J-J, Huang M-Q, Bao J-L, Chen X-P, Wang Y-T. Terpenoids: natural products for cancer therapy. Expert Opin Investig Drugs. (2012) 21:1801–18. doi: 10.1517/13543784.2012.727395
154. Andrianasolo EH, France D, Cornell-Kennon S, Gerwick WH. DNA Methyl transferase inhibiting halogenated monoterpenes from the madagascar red marine alga portieria h ornemannii. J Nat Prod. (2006) 69:576–9. doi: 10.1021/np0503956
155. Cotas J, Leandro A, Monteiro P, Pacheco D, Figueirinha A, Gonçalves AM, et al. Seaweed phenolics: from extraction to applications. Mar Drugs. (2020) 18:384. doi: 10.3390/md18080384
156. Isaza Martínez JH, Torres Castañeda HG. Preparation and chromatographic analysis of phlorotannins. J Chromatogr Sci. (2013) 51:825–38. doi: 10.1093/chromsci/bmt045
157. Catarino MD, Silva A, Cruz MT, Mateus N, Silva AM, Cardoso SM. Phlorotannins from fucus vesiculosus: modulation of inflammatory response by blocking Nf-κb signaling pathway. Int J Mol Sci. (2020) 21:6897. doi: 10.3390/ijms21186897
158. Kang M-H, Kim I-H, Nam T-J. Phloroglucinol induces apoptosis through the regulation of insulin-like growth factor 1 receptor signaling pathways in human colon cancer Ht-29 cells. Int J Oncol. (2014) 45:1036–42. doi: 10.3892/ijo.2014.2521
159. Yi L, Wang Q, Luo H, Lei D, Tang Z, Lei S, et al. Inhibitory effects of polyphenols-rich components from three edible seaweeds on inflammation and colon cancer in vitro. Front Nutr. (2022) 9:856273. doi: 10.3389/fnut.2022.856273
160. Eo HJ, Kwon T-H, Park GH, Song HM, Lee S-J, Park N-H, et al. In vitro anticancer activity of phlorofucofuroeckol a via upregulation of activating transcription factor 3 against human colorectal cancer cells. Mar Drugs. (2016) 14:69. doi: 10.3390/md14040069
161. Karthik R, Manigandan V, Sheeba R, Saravanan R, Rajesh PR. Structural characterization and comparative biomedical properties of phloroglucinol from indian brown seaweeds. J Appl Phycol. (2016) 28:3561–73. doi: 10.1007/s10811-016-0851-2
162. Pereira L. A review of the nutrient composition of selected edible seaweeds. Seaweed Ecol Nutr Composit Med Use. (2011) 7:15–47.
163. Song M, Nishihara R, Wang M, Chan AT, Qian ZR, Inamura K, et al. Plasma 25-Hydroxyvitamin D and colorectal cancer risk according to tumour immunity status. Gut. (2016) 65:296–304. doi: 10.1136/gutjnl-2014-308852
164. Ng K, Meyerhardt JA, Wu K, Feskanich D, Hollis BW, Giovannucci EL, et al. Circulating 25-Hydroxyvitamin D levels and survival in patients with colorectal cancer. J Clin Oncol. (2008) 26:2984–91. doi: 10.1200/JCO.2007.15.1027
165. Gorham ED, Garland CF, Garland FC, Grant WB, Mohr SB, Lipkin M, et al. Vitamin D and prevention of colorectal cancer. J Steroid Biochem Mol Biol. (2005) 97:179–94. doi: 10.1016/j.jsbmb.2005.06.018
166. Newmark HL, Yang K, Kurihara N, Fan K, Augenlicht LH, Lipkin M. Western-style diet-induced colonic tumors and their modulation by calcium and vitamin D in C57bl/6 mice: a preclinical model for human sporadic colon cancer. Carcinogenesis. (2009) 30:88–92. doi: 10.1093/carcin/bgn229
167. Rawson JB, Sun Z, Dicks E, Daftary D, Parfrey PS, Green RC, et al. Vitamin D intake is negatively associated with promoter methylation of the wnt antagonist gene Dkk1 in a large group of colorectal cancer patients. Nutr Cancer. (2012) 64:919–28. doi: 10.1080/01635581.2012.711418
168. Ferrer-Mayorga G, Larriba MJ, Crespo P, Muñoz A. Mechanisms of action of vitamin D in colon cancer. J Steroid Biochem Mol Biol. (2019) 185:1–6. doi: 10.1016/j.jsbmb.2018.07.002
169. Yun J, Mullarky E, Lu C, Bosch KN, Kavalier A, Rivera K, et al. Vitamin C selectively kills kras and braf mutant colorectal cancer cells by targeting gapdh. Science. (2015) 350:1391–6. doi: 10.1126/science.aaa5004
170. Matos GS, Pereira SG, Genisheva ZA, Gomes AM, Teixeira JA, Rocha CM. Advances in extraction methods to recover added-value compounds from seaweeds: sustainability and functionality. Foods. (2021) 10:516. doi: 10.3390/foods10030516
171. Michalak I, Tiwari R, Dhawan M, Alagawany M, Farag MR, Sharun K, et al. Antioxidant effects of seaweeds and their active compounds on animal health and production–a review. Vet Quarter. (2022) 42:48–67. doi: 10.1080/01652176.2022.2061744
172. Venturelli S, Leischner C, Helling T, Renner O, Burkard M, Marongiu L. Minerals and cancer: overview of the possible diagnostic value. Cancers. (2022) 14:1256. doi: 10.3390/cancers14051256
173. Lee J, Shin A, Choi J-Y, Kang D, Lee J-K. Adherence to the Recommended Intake of Calcium and Colorectal Cancer Risk in the Hexa Study. Cancer Res Treat. (2021) 53:140. doi: 10.4143/crt.2020.480
Keywords: colorectal cancer, therapeutic compounds, Chlorophyta, Rhodophyta, Phaeophyta
Citation: Fu Y, Xie D, Zhu Y, Zhang X, Yue H, Zhu K, Pi Z and Dai Y (2022) Anti-colorectal cancer effects of seaweed-derived bioactive compounds. Front. Med. 9:988507. doi: 10.3389/fmed.2022.988507
Received: 07 July 2022; Accepted: 01 August 2022;
Published: 19 August 2022.
Edited by:
Jiannan Li, Jilin University, ChinaReviewed by:
Zhiming Ma, Jilin University, ChinaCopyright © 2022 Fu, Xie, Zhu, Zhang, Yue, Zhu, Pi and Dai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yulin Dai, ZGFpeWxAY2N1Y20uZWR1LmNu; Zifeng Pi, bXNsYWIyMUBjaWFjLmFjLmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.