
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Med. , 14 January 2022
Sec. Translational Medicine
Volume 8 - 2021 | https://doi.org/10.3389/fmed.2021.803067
This article is part of the Research Topic Skin Lesion Vitality Assessment for Forensic Science: Current Research and New perspectives View all 10 articles
 Stefania De Simone1
Stefania De Simone1 Elena Giacani1Maria Antonella Bosco1Simona Vittorio2Michela Ferrara1
Elena Giacani1Maria Antonella Bosco1Simona Vittorio2Michela Ferrara1 Giuseppe Bertozzi1
Giuseppe Bertozzi1 Luigi Cipolloni1*
Luigi Cipolloni1* Raffaele La Russa1
Raffaele La Russa1Background: The timing of wounds production is a significant issue in forensic pathology. Although various methods have been evaluated, obtaining an accurate dating of lesions is still a challenge. The pathologist uses many parameters to value wound age, such as histological and immunohistochemical. In recent years, there have been many studies regarding the use of miRNAs in wound-age estimation; indeed, miRNAs have multiple potential uses in forensic pathology.
Scope: This review aims to verify the efficacy and feasibility of miRNAs as a tool for determining the timing of lesions.
Materials and Methods: The authors conducted the systematic review according to the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guidelines. PubMed was used as a search engine to find articles published between January, 1st 2016 and October, 1st 2021, to evaluate the current state of the art regarding wound-age estimation.
Results: A total of 256 articles were collected; after screening according to PRISMA guidelines, the systematic review included 8 articles. The studies included in this review were all Original articles evaluating the use of biomarkers for wound-age determination.
Discussion and Conclusion: The literature review showed that analysis of miRNA is an innovative field of study with significant potentiality in forensic pathology. There are few studies, and almost all of them are at an early stage. The challenge is to understand how to standardize the samples' selection to obtain reliable experimental data. This observation represents a necessary prerequisite to planning further clinical trials.
The timing of wounds production is a significant issue in forensic pathology. Although various methods have been evaluated, obtaining an accurate dating of lesions is still a challenge (1).
After an injury, the pathologists must consider many vital processes (e.g., hemorrhage, inflammatory cells migration, infiltration, development of granulation tissue) for an accurate wound-age estimation (2).
Morphological and macroscopic analysis was one of the first methods used (3). But it is not of practical use because it cannot be standardized.
Histological evaluation (with Haematoxylin-Eosin staining) can detect the presence of inflammatory cells or substances secreted during the inflammatory process (4). Different types of inflammatory cells reach a peak with exact timing, allowing age estimation of the lesion [neutrophilic granulocytes reach the maximum levels after 24 h (4, 5); macrophages after 2-3 days (4); lymphocytes at least 20 h; fibroblastic cells after 6 days or more (4)].
Immunohistochemical methods can also detect different substances secreted during the inflammatory process (6) [esterase and adenosine triphosphatase increase 1 h after injury; aminopeptidase reaches a peak at about 2 h; acid phosphatase at 4 h; alkaline phosphatase at 8 h (7)] or analyze the expression of Fibronectin, CD62p, and Factor VIII, which are markers involved in coagulation and inflammation processes. These, significantly increase in wounds produced close to death (15–30 min old) (8). Also, the number of matrix metalloproteinase-2 (+) macrophages increases in accordance with wound ages (9).
Histamine is a mast cell degranulation product, used as a marker in forensics (e.g., for asphyxiation) (10). Histamine levels in the skin vary appreciably after wounding; these levels increase significantly between 5 min and 3 h after trauma and decrease until 24 h. The transient infiltration of mast cells occurs in the dermis for 3 h upon vital lesions and is followed by a protracted decrease until 24 h (11, 12).
The real-time polymerase chain reaction (PCR) measures mRNA levels (13). PCR can detect the mRNA levels of inflammatory cytokines and wound-healing factors [e.g., caspase level 3, 8, and 9 expressions in tissues (14); VEGF and TGFb1 mRNA have high grades from the early stages, reaching the peak at day 7 (5)]. As mRNA is not very stable due to the action of ribonucleases (15), it is not very useful for forensic purposes. The stability, moreover, presents significant differences depending on the tissue analyzed: skeletal muscle, heart, and brain are more stable than the pancreas (13).
According to the studies on this topic a single parameter is not sufficient for estimating wound-age; indeed, combining parameters can reduce error.
Therefore, in recent years, many studies have been performed regarding the use of miRNAs in wound-age estimation. Indeed, the researchers found microRNAs (miRNAs) expression at 0, 24, and 48 h after death. Especially high levels of miRNA-205 and miRNA-21 24 h after death were observed (16).
MiRNAs were first described in 1993 by Lee et al. (17), and the term microRNA was born in 2001 (18).
MiRNAs are small non-coding RNA molecules containing about 21–25 nucleotides (19) that interact with the 3′ untranslated region (3′ UTR) of target mRNAs to induce their degradation. They can also interact with the 5′ UTR region, coding sequence, gene promoters and activate translation or regulate transcription (20).
The interaction of miRNAs with their target genes is dynamic. It depends on many factors, such as the subcellular location of miRNAs, and both the abundance of miRNAs and target mRNAs. miRNAs can also be secreted into extracellular fluids and operate as chemical messengers to mediate intercellular communication (20).
About 1-3% of the genome encodes miRNAs, which regulate more than 30% of protein-coding genes (17). Many studies have demonstrated that miRNAs play a significant role in biological processes, including cell proliferation, differentiation, apoptosis, organ development, pathogenesis, metabolic control, and antiviral defense (21).
However, the cellular localization of miRNAs is still widely unappreciated (22).
For this reason, many studies have been carried out in recent years to identify some tissue-specific miRNAs (21, 22). MiRbase, the bioinformatic database of miRNA sequences, reports the 1917 miRNAs discovered and encoded by the human genome [www.mirbase.org].
Microarray or Next Generation Sequencing (NGS) techniques can achieve the analysis of miRNAs. After their identification, the researcher can perform the assay by Real-Time quantitative PCR (qRT-PCR) (23).
Modification in the normal miRNA expression pathway can affect normal cellular physiology and lead to different pathologies.
Chronic lymphocytic leukemia was the first human disease known to be related to miRNA deregulation. Human cancer can be associated with changes in miRNA expression by deregulation of oncogenes and oncosuppressor genes (24–26).
MiRNAs' dysregulation is also involved in other diseases like cardiac hypertrophy and heart failure (27–30), chronic kidney disease (31), obesity (32), and various neurological and neuropsychiatric disorders (33).
The aim of this review is to verify the usefulness of miRNAs as a tool for determining the age of lesions. In our opinion it is necessary to analyze the current state of the art on wound-age estimation before to plan additional experimental studies on these topics.
MiRNAs have multiple potential uses in forensic pathology. In fact, unlike mRNA, they are less sensitive to degradation (due to environmental factors, such as UV light or heat, and the action of cellular ribonucleases) (34) and more stable than mRNA. They can also be extracted together with the DNA profile, representing a double test, useful in case of shortage of biological material (35).
MiRNAs are helpful for the recognition of biological fluids found at a crime scene. Nowadays, only a few miRNAs are recognized as specific for a single biological fluid (especially for blood and seminal fluid, while miRNAs specific for venous and menstrual blood are yet to be determined) (36).
Many authors focalized their research on the study of wound vitality (37). Some authors have focused on differentiating ante- and post-mortem lesions through miRNAs (38), but studies are still few and at an early stage.
Determining the age of production of the wounds is a relevant topic to forensic purposes (39). Over the years, many authors studied how putrefaction affects the concentration of proteins, DNA and RNA (40–43). Maiese et al. (44) analyzed the progress made in recent years, showing that some miRNAs are stable and reasonably reliable as markers. Wang et al. (45) demonstrated that three miRNAs (miR-122, miR-150, miR-195) are stable in the first 24 h of PMI (Post-Mortem Interval), declining after that time. Some studies remarked whether the animal died during the day or at night, considering the modification of other miRNAs (miR-541 and miR-142-p) (46).
MiRNAs have an essential role in moderating cellular adaptations occurring in case of drug abuse and addiction (47). The alteration of regular miRNAs expression occurs even in response to substances' abuse, such as nicotine, cocaine, morphine, and alcohol (48–50).
A further field of application for miRNAs is as anti-doping marker (51, 52).
An innovative field of application for miRNAs is the estimation of wound-age, as miRNAs are variously involved in wound healing physiology and physiopathology from injury to re-epithelialization (53, 54).
Aunin et al. (55) investigated the expression of miR-200 in skin wounds of subjects of different ages, marking its role in wound repair. The study identified miR-200c as a critical determinant that inhibits cell migration during skin repair after injury and may contribute to age-associated alterations in wound repair.
Ibrahim et al. (16) studied the post-mortem expression of miR-21 and miR-205 on incisional wounds on a cohort of 18 female albino rats. The animals were injured and then sacrificed, and divided into three groups. In the first group, the samples were taken immediately after death, in the second group after 24 h, and in the third one after 48 h. The researchers analyzed the expression of miRNAs in each sample, observing higher expression in the second group than in the other two. The examined miRNAs were still expressed 48 h after death, although at a lower level. Despite these exciting results, the population studied is too restricted to be considered of meaningful impact and further studies are necessary.
The authors conducted the systematic review according to the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guideline (56). The quality assessment of this study was evaluated using the Checklist for Systematic Reviews and Research Syntheses recommended by the Joanna Briggs Institute (JBI).
PubMed and Scopus were used as a search engines to find articles published between 1 January 2016 and 1 October 2021, to evaluate the current state of the art regarding wound-age estimation. The Medical Subject Heading (MeSH) thesaurus was used for the following word: “(wound-age estimation) and (wound healing)”; “(wound-age estimation) and (post-mortem)”; “(wound-age estimation) and (miRNA).”
The inclusion criteria were: Article in English, Case report; Case series; Original Article; in vivo studies (both animals and human); Studies with searching for specific biomarkers.
The exclusion criteria were: Article not in English; Abstract; Poster; Proceedings; Review; Meta-Analysis. The researchers decided to exclude reviews and meta-analysis to give an experimental attitude to the article, analyzing only experimental research.
The methodology of the search strategy is presented in Figure 1.
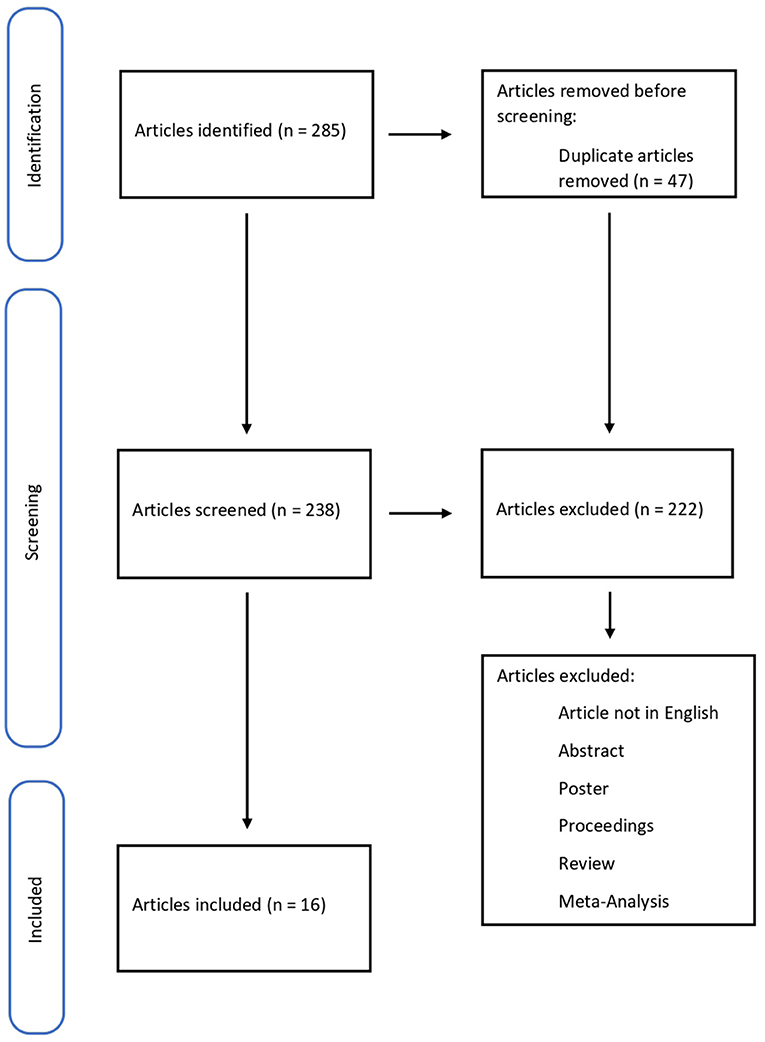
Figure 1. Flow diagram illustrating studies included in and excluded from this systematic review.
Methodological evaluation of each study was conducted according to the PRISMA standards, including assessment of bias. Data collection involved study selection and data extraction. Three researchers (S.D.S., M.A.B., E.G.) independently reviewed those documents whose title or abstract appeared to be relevant and selected those who analyzed the wound-age estimation.
This systematic review focuses on articles published in the last 5 years, doing specific research using a few keywords. This topic represents a restricted research field, in which few experiments have been carried out. The limited number of articles available can be a source of error.
A total of 285 articles were collected, removing 47 duplicates. 238 papers were screened, and 222 did not meet the inclusion criteria; 15 pertinent articles were in Chinese. In conclusion, the systematic review included 16 articles. The studies included in this review were all Original articles (n = 16) considering biomarkers for wound-age determination (Table 1).
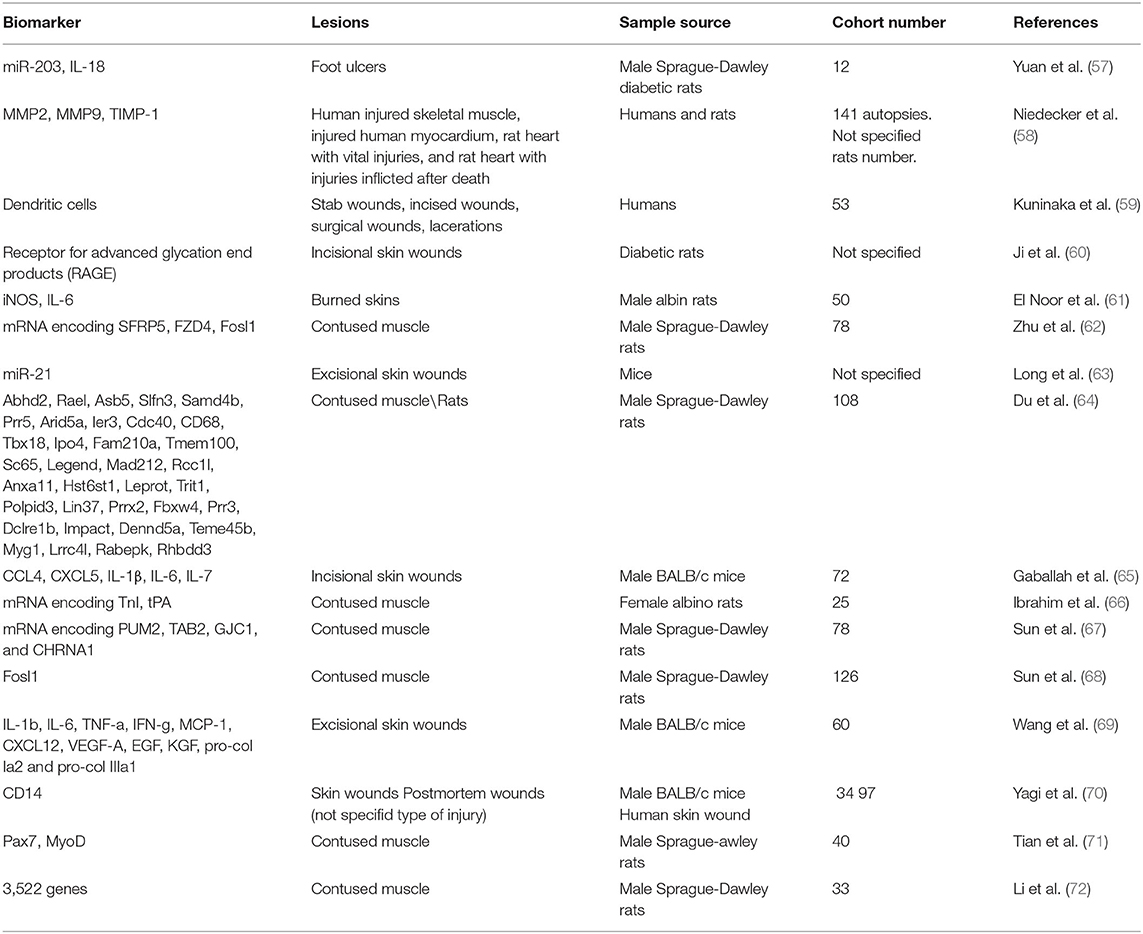
Table 1. Summary of the reviewed literature.
The wound-age estimation is still a challenge for forensic pathologists worldwide. They often attend crime scenes, in which bodies show many injuries, whose production time is only conceivable (73). The relevance of this argument encourages more studies on this topic.
In one of the latest reviews in the literature, Li et al. (2) examined the publications made in the years 2010-2016. They stated that there are no standardized and reliable biomarkers to estimate wound-age, despite the study of many molecules. The use of multiple markers could make dating more reliable.
From 2016, as emerged from our literature analysis, only a few studies have been published. All these papers specify a need for further researches.
Yuan et al. (57) investigated the role of miR-203 and IL-8 in the healing process of diabetic foot ulcers in rats. MiR-203 is involved in wound repair and promotes apoptosis, while IL-8 promotes cell proliferation and survival. MiR-203 directly downregulates mRNA expression for IL-18, inhibiting the wound healing process. However, the authors do not evaluate miR-203 or IL-18 expression timing, highlighting only greater miRNA expression in diabetic tissues.
Niedecker et al. (58) studied markers such as matrix metalloproteinases (MMP) 2 and 9 and tissue inhibitors of matrix metalloproteinases 1 (TIMP-1) in human and animal tissues. The authors collected samples of human injured skeletal muscle, injured human myocardium, rat myocardium both with vital and post-mortem injuries. Immunohistochemical investigations showed that these markers did not help wound-age estimation, as the samples did not show positivity (or weak positivity in older wounds).
The research of Kuninaka et al. (59) examined dendritic cells. Dendritic cells (DCs) are cells involved in the immune response and healing the burned skin. The authors collected 53 samples of injured human skin whose production time was known (from a few hours to 21 days). The CD11c and HLA-DRα markers detect the presence of DCs. Wounds aged 4 to 7 days showed low DCs, similar to 9 to 14 days. The samples aged 4-14 days showed the presence of more than 50 DCs per observation field. In the samples aged 17-21 days, this number gradually decreased.
This study indicates that DCs could be a good marker for wound-age estimation, even if the time interval of expression is too broad. Also, Bacci et al. (74) performed a study on DCs cells, measuring their concentration within hours of injury (a short range of time).
Ji et al. (60) made incisional injuries on diabetic mice that they killed after a defined time (6 h to 14 days), finally sampling the lesions. Subsequently, they looked for the expression of the receptor for advanced glycation end products (RAGE) expressed mainly by polymorphonuclear cells. There was an upregulation of RAGEs by 60% between 7 and 10 days after the wounds. However, the authors suggest cautiously interpreting these results, as the diabetic mice used for the study had persistent hyperglycemia from 4 weeks of age and never received treatment.
El-Noor et al. (61) studied the expression of inducible nitric oxide synthase (iNOS) and IL-6 in burned rat skin samples. The authors caused burns on the skin of the mice, sampling them after a defined time (1, 3, 5, 7, 9, 11, 13, 15, and 21 days after the burn). Six hours after the death of the rats, they inflicted another burn, also sampled. The study of markers in ante-mortem wounds revealed that iNOS becomes positive between 3 and 5 days, peaking on day 7, after which it begins to decline. The expression of IL-6 is also time-dependent, starting on day one and peaking on day 3, remaining high until day 5. The positivity begins to decrease from day 7 until it disappears by day 21. In the post-mortem wounds, both markers were weakly positive.
Zhu et al. (62) studied the expression of mRNAs encoding frizzled-related protein 5 (SFRP5), frizzled class receptor 4 (FZD4), and Fos-link antigen 1 (Fosl1) in bruised muscles of rats. These proteins are involved in the healing of injured muscle. The authors monitored their expression for up to 48 h; all mRNAs showed changes, although very different. These trends need further studies to understand if the coding proteins could be considered as useful markers.
Research by Long et al. (63) focused on studying the expression of mir-21 on the skin of female rats with excisional wounds. The authors demonstrated that miR-21 expression improved cutaneous wound repair. However, the purpose of this paper was not to use miR-21 to date the injuries production, so it cannot have forensic application.
Du et al. (64) studied the expression of 35 wound healing-related genes on contused muscle tissue samples from rats through PCR. The authors divided the samples into three groups time-based. Of the 35 genes, 14 showed particular utility in discriminating between the three age groups based on their expression time.
Gaballah et al. (65) studied the expression of five cytokines from the measurement of their RNA through qRT-PCR: chemokine ligand 4 (CCL4), chemokine ligand 5 (CXCL5), interleukin-1 beta (IL-1β), interleukin-6 (IL-6), and interleukin-7 (IL-7). The authors performed incisional wounds on the hamstring muscle of 72 mice, divided into five groups based on the time of wound collection (6, 12, 24, 36, and 48 h post-injury). CXCL5 was the most upregulated molecule with higher expression 6-36 h after injury, such as CCL4 and IL-1β. IL-6, on the other hand, peaked at 6 h post-injury, to significantly decrease at 12 h. IL-7 showed a slow and steady increase over time up to 48 h after injury. Immunohistochemical staining of the post-injury samples showed a gradual increase in intensity around the wound edges. In light of these promising results, this study points out that since post mortem RNA degrades quickly, the application of protein markers is more reliable.
Ibrahim et al. (66) analyzed skeletal troponin I mRNA (TnI) and tissue plasminogen activator (tPA) mRNA expression after inflicting a bruised wound on 25 rats. The researchers divided the rats into five groups based on the injury time. tPA expression levels significantly decreased at 1, 6, and 30 h after contusion. In contrast, TnI expression levels increased at 1 and 6 h post-traumatic, then gradually reduced to normal levels at 24 h, and they assumed significantly lower levels at 30 h after the contusion.
Sun et al. (67) developed an “up, no change or down” system to study the time-dependent expression of PUM2, TAB2, GJC1, and CHRNA1 mRNAs in bruised skeletal muscle of rats. The rats were divided into 12 groups based on the time of wound production. Subsequently, the authors combined the levels of mRNAs. The mRNA levels of PUM2, TAB2, and GJC1 decreased, while CHRNA1 mRNA levels increased. The proposed system can detect the injury time for short periods.
In another research article, Sun et al. (68) such as Zhu et al., studied Fosl1 mRNA and protein in contused skeletal muscle. The researchers divided the rats into different groups (control, contused, and postmortem). They then examined the expression of Fosl1 mRNA and the coding protein in bruised muscle, uninjured contralateral, and postmortem femoral muscle samples. Both mRNA and protein levels of Fosl1 exhibited time-dependent expression during contused skeletal muscle healing. There was no significant difference in Fosl1 protein levels among the postmortem muscle and control specimens, which means that Fosl1 protein levels remained stable in the postmortem samples within 24 h. The study suggests that the expression of Fosl1 mRNA was susceptible to the degree of injury. Fosl1 mRNA could be a promising marker to determine the force that caused the wound but is not stable enough to estimate the duration of the wound.
Wang et al. (69) studied several molecules, including IL-1b, IL-6, TNF-a, IFN-g, MCP-1, CXCL12, VEGF-A, EGF, KGF, pro-col Ia2, and pro-col IIIa1. The authors inflicted skin wounds and subsequently removed them after standardized times. The expression of each molecule was increased over a different time range, depending on their role in wound repair.
Yagi et al. (70) looked for CD14 expression in mouse wounds and postmortem humans. In particular, in humans, they evaluated CD14 in combination with other proteins, such as CD32B and CD68. CD14 appears to be a valuable marker of wound age, 1–5 days postinfliction. However, the combination of several markers increases the sensitivity of the method.
Tian et al. (71) investigated the time-dependent expression of transcription factor 7 (Pax7) and myoblast-determining protein (MyoD) during skeletal muscle wound healing. The experiment consisted of injuring the right limb of rats. Pax7 protein peaked 5 days after injury and subsequently declined, while MyoD mRNA expression peaked 3 days after injury. In conclusion, Pax7 and MyoD expression are time-dependent upregulated during skeletal muscle wound healing, suggesting that they may become potential markers for estimating wound age in skeletal muscle.
Li et al. (72) collected samples of animal injured skeletal muscle taken from the right posterior limb and investigated differentially expressed genes (DEGs) using microarray analyses. A total of 2,844 and 2,298 DEGs involved in muscle repair were identified in the mild and severe contusions, respectively. These DEGs overlap during the early stages after injury (within 48 h). During later stages (at 168 h) there is an expression of only 29 genes. The genes showed time-dependent patterns of expression, which provide a basis for further studies of wound age estimation.
There are not many recent studies on wound-age estimation, probably because of sampling difficulties. Most studies regarded animals but in a low number cohort, summarized in Table 1. Studies on human tissues are often problematic to apply because of ethical requirements (75). Most of the studies also concern protein markers and their mRNA, analyzed by immunohistochemistry, Western-Bloth, and PCR.
From a forensic point of view, it is challenging to select a homogeneous sample of patients due to the difficulty of knowing the lesion's production time. It is essential to know the timing of lesions to experimentally study the expression trend of a potential marker.
Only Kuninaka et al. (59) and Yagi et al. (70) performed studies sampling human wounds with known production time. However, in the studies on rats, the production time was established by the researchers.
The wound-age estimation is an essential issue in forensic pathology. Pathologists need sensitive and specific markers to conduct an objective assessment and to obtain scientifically and validated evidences. The literature review showed that analysis of miRNA is an innovative field of study with significant potentiality in forensic pathology. The review on the wound-age estimation revealed that research has not progressed in the past 5 years. There are few studies, and almost all of them are at an early stage. On the other hand, the field of wounds' dating in forensic pathology is essential. The main problem in obtaining sufficient data for validation is the difficulty of collecting adequate and homogeneous samples. Several studies recommend the use of multiple methods to get reliable results. In our opinion, the challenge is to understand how to standardize the samples' selection to obtain reliable experimental data. This observation represents a necessary prerequisite to planning further clinical trials.
SD, MF, and EG: analyzed the literature. SD and MB: writing. LC and RL: review and editing. GB and SV: language supervision. All authors have read and agreed to the published version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer AM declared a shared affiliation, with no collaboration, with one of the authors SV to the handling editor at the time of the review.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Thanks to all the peer reviewers and editors for their opinions and suggestions.
1. Gauchotte G, Martrille L, Plénat F, Vignaud J-M. Les marqueurs de vitalité des blessures en pathologie médicolégale. Ann Pathol. (2013) 33:93–101. doi: 10.1016/j.annpat.2013.02.006
2. Li N, Du Q, Bai R, Sun J. Vitality and wound-age estimation in forensic pathology: review and future prospects. Forensic Sci Res. (2020) 5:15–24. doi: 10.1080/20961790.2018.1445441
3. Oehmichen M. Vitality and time course of wounds. Forensic Sci Int. (2004) 144:221–31. doi: 10.1016/j.forsciint.2004.04.057
4. Betz P. Histological and enzyme histochemical parameters for the age estimation of human skin wounds. Int J Leg Med. (1994) 107:60–8. doi: 10.1007/BF01225491
5. Khalaf AA, Hassanen EI, Zaki AR, Tohamy AF, Ibrahim MA. Histopathological, immunohistochemical, and molecular studies for determination of wound age and vitality in rats. Int Wound J. (2019) 16:1416–25. doi: 10.1111/iwj.13206
6. Kondo T, Ohshima T. The dynamics of inflammatory cytokines in the healing process of mouse skin wound: a preliminary study for possible wound age determination. Int J Leg Med. (1996) 108:231–6. doi: 10.1007/BF01369816
7. Raekallio J. Determination of the age of wounds by histochemical and biochemical methods. Forensic Sci. (1972) 1:3–16. doi: 10.1016/0300-9432(72)90144-6
8. Van de Goot FRW, Korkmaz HI, Fronczek J, Witte BI, Visser R, et al. A new method to determine wound age in early vital skin injuries: a probability scoring system using expression levels of Fibronectin, CD62p and Factor VIII in wound hemorrhage. Forensic Sci Int. (2014) 244:128–35. doi: 10.1016/j.forsciint.2014.08.015
9. Ishida Y, Kuninaka Y, Nosaka M, Kimura A, Kawaguchi T, Hama M, Sakamoto S, et al. Immunohistochemical analysis on MMP-2 and MMP-9 for wound age determination. Int J Legal Med. (2015) 129:1043–8. doi: 10.1007/s00414-015-1167-5
10. Muciaccia B, Sestili C, De Grossi S, Vestri A, Cipolloni L, Cecchi R. Are mast cells implicated in asphyxia? Int J Legal Med. (2016) 130:153–61. doi: 10.1007/s00414-015-1211-5
11. Bonelli A, Bacci S, Vannelli G, Norelli G. Immunohistochemical localization of mast cells as a tool for the discrimination of vital and postmortem lesions. Int J Legal Med. (2003) 117:14–8. doi: 10.1007/s00414-002-0304-0
12. View of Commentary: Mast Cells and Dendritic Cells as a Tools to Discriminate Vital and Post Mortem Lesions. Available online at: https://esmed.org/MRA/mra/article/view/859/608 (accessed December 20, 2021).
13. Zubakov D, Hanekamp E, Kokshoorn M, van IJcken W, Kayser M. Stable RNA markers for identification of blood and saliva stains revealed from whole genome expression analysis of time-wise degraded samples. Int J Legal Med. (2008) 122:135–42. doi: 10.1007/s00414-007-0182-6
14. Akar T, Saglam ASY, Göçün PU, Alp E, Konac E, Menevşe A. Evaluation of apoptotic caspase levels in estimation of the wound age. Turkish J Biochem. (2018) 43:126–33. doi: 10.1515/tjb-2017-0131
15. Bauer M. RNA in forensic science. Forensic Sci Int Genet. (2007) 1:69–74. doi: 10.1016/j.fsigen.2006.11.002
16. Ibrahim SF, Ali MM, Basyouni H, Rashed LA, Amer EAE, Abd El-Kareem D. Histological and miRNAs postmortem changes in incisional wound. Egypt J Forensic Sci. (2019) 9:37. doi: 10.1186/s41935-019-0141-7
17. Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. (1993) 75:843–54. doi: 10.1016/0092-8674(93)90529-Y
19. Filipowicz W, Jaskiewicz L, Kolb FA, Pillai RS. Post-transcriptional gene silencing by siRNAs and miRNAs. Curr Opin Struct Biol. (2005) 15:331–41. doi: 10.1016/j.sbi.2005.05.006
20. O'Brien J, Hayder H, Zayed Y, Peng C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol. (2018) 9:402. doi: 10.3389/fendo.2018.00402
21. Gong W, Huang Y, Xie J, Wang G, Yu D, Zhang K, et al. Identification and comparative analysis of the miRNA expression profiles from four tissues of Micropterus salmoides using deep sequencing. Genomics. (2018) 110:414–22. doi: 10.1016/j.ygeno.2018.09.017
22. McCall MN, Kim M-S, Adil M, Patil AH, Lu Y, Mitchell CJ, et al. Toward the human cellular microRNAome. Genome Res. (2017) 27:1769–81. doi: 10.1101/gr.222067.117
23. Partemi S, Berne PM, Batlle M, Berruezo A, Mont L, Riuró H, et al. Analysis of mRNA from human heart tissue and putative applications in forensic molecular pathology. Forensic Sci Int. (2010) 203:99–105. doi: 10.1016/j.forsciint.2010.07.005
24. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al. MicroRNA expression profiles classify human cancers. Nature. (2005) 435:834–8. doi: 10.1038/nature03702
25. Xu X, Tao Y, Shan L, Chen R, Jiang H, Qian Z, et al. The role of microRNAs in hepatocellular carcinoma. J Cancer. (2018) 9:3557–69. doi: 10.7150/jca.26350
26. Eyking A, Reis H, Frank M, Gerken G, Schmid KW, Cario E. MiR-205 and MiR-373 are associated with aggressive human mucinous colorectal cancer. PLoS ONE. (2016) 11:e0156871. doi: 10.1371/journal.pone.0156871
27. Van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, Gerard RD, et al. A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proceed Natl Acad Sci. (2006) 103:18255–60. doi: 10.1073/pnas.0608791103
28. Tatsuguchi M, Seok HY, Callis TE, Thomson JM, Chen J-F, Newman M, et al. Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy. J Mol Cell Cardiol. (2007) 42:1137–41. doi: 10.1016/j.yjmcc.2007.04.004
29. Manetti AC, Maiese A, Paolo MD, De Matteis A, La Russa R, Turillazzi E, et al. MicroRNAs and sepsis-induced cardiac dysfunction: a systematic review. IJMS. (2020) 22:321. doi: 10.3390/ijms22010321
30. Pinchi E, Frati P, Aromatario M, Cipolloni L, Fabbri M, La Russa R, et al. miR-1, miR-499 and miR-208 are sensitive markers to diagnose sudden death due to early acute myocardial infarction. J Cell Mol Med. (2019) 23:6005–16. doi: 10.1111/jcmm.14463
31. Zhao H, Ma S-X, Shang Y-Q, Zhang H-Q, Su W. microRNAs in chronic kidney disease. Clin Chim Act. (2019) 491:59–65. doi: 10.1016/j.cca.2019.01.008
32. Landrier J-F, Derghal A, Mounien L. MicroRNAs in obesity and related metabolic disorders. Cells. (2019) 8:859. doi: 10.3390/cells8080859
33. Cao D-D, Li L, Chan W-Y. MicroRNAs: key regulators in the central nervous system and their implication in neurological diseases. IJMS. (2016) 17:842. doi: 10.3390/ijms17060842
34. An J-H, Shin K-J, Yang W-I, Lee H-Y. Body fluid identification in forensics. BMB Reports. (2012) 45:545–53. doi: 10.5483/BMBRep.2012.45.10.206
35. Van der Meer D, Uchimoto ML, Williams G. Simultaneous analysis of micro-RNA and DNA for determining the body fluid origin of DNA profiles. J Forensic Sci. (2013) 58:967–71. doi: 10.1111/1556-4029.12160
36. Rocchi A, Chiti E, Maiese A, Turillazzi E, Spinetti I. MicroRNAs: an update of applications in forensic science. Diagnostics. (2020) 11:32. doi: 10.3390/diagnostics11010032
37. Casse J-M, Martrille L, Vignaud J-M, Gauchotte G. Skin wounds vitality markers in forensic pathology: an updated review. Med Sci Law. (2016) 56:128–37. doi: 10.1177/0025802415590175
38. Neri M, Fabbri M, D'Errico S, Di Paolo M, Frati P, Gaudio RM, et al. Regulation of miRNAs as new tool for cutaneous vitality lesions demonstration in ligature marks in deaths by hanging. Sci Rep. (2019) 9:20011. doi: 10.1038/s41598-019-56682-7
39. Gelderman HT, Boer L, Naujocks T, IJzermans ACM, Duijst WLJM. The development of a post-mortem interval estimation for human remains found on land in the Netherlands. Int J Legal Med. (2018) 132:863–73. doi: 10.1007/s00414-017-1700-9
40. Larkin B, Iaschi S, Dadour I, Tay GK. Using accumulated degree-days to estimate postmortem interval from the DNA yield of porcine skeletal muscle. Forensic Sci Med Pathol. (2010) 6:83–92. doi: 10.1007/s12024-009-9109-5
41. Bauer M, Gramlich I, Polzin S, Patzelt D. Quantification of mRNA degradation as possible indicator of postmortem interval—a pilot study. Legal Med. (2003) 5:220–7. doi: 10.1016/j.legalmed.2003.08.001
42. Ali MM, Ibrahim SF, Fayed AA. Using skin gene markers for estimating early postmortem interval at different temperatures. Am J Forensic Med Pathol. (2017) 38:323–5. doi: 10.1097/PAF.0000000000000337
43. Harvey ML, Gasz NE, Voss SC. Entomology-based methods for estimation of postmortem interval. RRFMS. (2016) 6:1–9. doi: 10.2147/RRFMS.S68867
44. Maiese A, Scatena A, Costantino A, Di Paolo M, La Russa R, Turillazzi E, et al. MicroRNAs as Useful tools to estimate time since death. a systematic review of current literature. Diagnostics. (2021) 11:64. doi: 10.3390/diagnostics11010064
45. Wang H, Mao J, Li Y, Luo H, Wu J, Liao M, et al. 5 miRNA expression analyze in post-mortem interval (PMI) within 48h. Forensic Sci Int Genet Suppl Series. (2013) 4:e190–1. doi: 10.1016/j.fsigss.2013.10.098
46. Birdsill AC, Walker DG, Lue L, Sue LI, Beach TG. Postmortem interval effect on RNA and gene expression in human brain tissue. Cell Tissue Bank. (2011) 12:311–8. doi: 10.1007/s10561-010-9210-8
47. Sessa F, Salerno M, Cipolloni L, Bertozzi G, Messina G, Mizio GD, et al. Anabolic-androgenic steroids and brain injury: miRNA evaluation in users compared to cocaine abusers and elderly people. Aging. (2020) 12:15314–27. doi: 10.18632/aging.103512
48. Lewohl JM, Nunez YO, Dodd PR, Tiwari GR, Harris RA, Mayfield RD. Up-regulation of microRNAs in brain of human alcoholics: microRNAs in human alcoholics. Alcohol Clin Exp Res. (2011) 35:1928–37. doi: 10.1111/j.1530-0277.2011.01544.x
49. Chandrasekar V, Dreyer J-L. microRNAs miR-124, let-7d and miR-181a regulate Cocaine-induced Plasticity. Mol Cell Neurosci. (2009) 42:350–62. doi: 10.1016/j.mcn.2009.08.009
50. Kazemi T, Huang S, Avci NG, Waits CMK, Akay YM, Akay M. Investigating the influence of perinatal nicotine and alcohol exposure on the genetic profiles of dopaminergic neurons in the VTA using miRNA–mRNA analysis. Sci Rep. (2020) 10:15016. doi: 10.1038/s41598-020-71875-1
51. Leuenberger N, Saugy M. Circulating microRNAs: the future of biomarkers in anti-doping field. In: Santulli G, editor. MicroRNA: Medical Evidence. (2015). Cham: Springer International Publishing. p. 401–8.
52. Sessa F, Salerno M, Bertozzi G, Cipolloni L, Messina G, Aromatario M, et al. miRNAs as novel biomarkers of chronic kidney injury in anabolic-androgenic steroid users: an experimental study. Front Pharmacol. (2020) 11:1454. doi: 10.3389/fphar.2020.563756
53. Lang H, Zhao F, Zhang T, Liu X, Wang Z, Wang R, et al. MicroRNA-149 contributes to scarless wound healing by attenuating inflammatory response. Mol Med Rep. (2017) 16:2156–62. doi: 10.3892/mmr.2017.6796
54. Banerjee J, Sen CK. MicroRNA and wound healing. In: Santulli G, editor. MicroRNA: Medical Evidence. (2015). Cham: Springer International Publishing. p. 291–305.
55. Aunin E, Broadley D, Ahmed MI, Mardaryev AN, Botchkareva NV. Exploring a role for regulatory miRNAs in wound healing during ageing: involvement of miR-200c in wound repair. Sci Rep. (2017) 7:3257. doi: 10.1038/s41598-017-03331-6
56. Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. (2021) 372:n71. doi: 10.1136/bmj.n71
57. Yuan L, Sun Y, Xu M, Zeng F, Xiong X. miR-203 acts as an inhibitor for epithelial-mesenchymal transition process in diabetic foot ulcers via targeting interleukin-8. Neuroimmunomodulation. (2019) 26:239–49. doi: 10.1159/000503087
58. Niedecker A, Huhn R, Ritz-Timme St, Mayer F. Complex challenges of estimating the age and vitality of muscle wounds: a study with matrix metalloproteinases and their inhibitors on animal and human tissue samples. Int J Legal Med. (2021) 135:1843–53. doi: 10.1007/s00414-021-02563-6
59. Kuninaka Y, Ishida Y, Nosaka M, Shimada E, Kimura A, Ozaki M, et al. Forensic pathological study on temporal appearance of dendritic cells in skin wounds. Int J Legal Med. (2020) 134:597–601. doi: 10.1007/s00414-019-02185-z
60. Ji X-Y, Chen Y, Ye G-H, Dong M-W, Lin K-Z, Han J-G, et al. Detection of RAGE expression and its application to diabetic wound age estimation. Int J Legal Med. (2017) 131:691–8. doi: 10.1007/s00414-016-1529-7
61. Abo El-Noor MM, Elgazzar FM, Alshenawy HA. Role of inducible nitric oxide synthase and interleukin-6 expression in estimation of skin burn age and vitality. J Forensic Leg Med. (2017) 52:148–53. doi: 10.1016/j.jflm.2017.09.001
62. Zhu X, Du Q, Li S, Sun J. Comparison of the homogeneity of mRNAs encoding SFRP5, FZD4, and Fosl1 in post-injury intervals: Subcellular localization of markers may influence wound age estimation. J Forensic Leg Med. (2016) 43:90–6. doi: 10.1016/j.jflm.2016.07.013
63. Long S, Zhao N, Ge L, Wang G, Ran X, Wang J, Su Y, et al. MiR-21 ameliorates age-associated skin wound healing defects in mice. J Gene Med. (2018) 20:e3022. doi: 10.1002/jgm.3022
64. Du Q, Li N, Dang L, Dong T, Lu H, Shi F, et al. Temporal expression of wound healing–related genes inform wound age estimation in rats after a skeletal muscle contusion: a multivariate statistical model analysis. Int J Legal Med. (2020) 134:273–82. doi: 10.1007/s00414-018-01990-2
65. Hassan Gaballah M, Fukuta M, Maeno Y, Seko-Nakamura Y, Monma-Ohtaki J, Shibata Y, et al. Simultaneous time course analysis of multiple markers based on DNA microarray in incised wound in skeletal muscle for wound aging. Forensic Sci Int. (2016) 266:357–68. doi: 10.1016/j.forsciint.2016.06.027
66. Ibrahim SF, Issak M, Bayoumy AA, Abd El-Fatah DS. Cutaneous (tPA) and skeletal (TnI) mRNA as markers of aging in contused wound. J Forensic Sci. (2016) 61:1007–10. doi: 10.1111/1556-4029.13072
67. Sun J, Zhu X, Dong T, Zhang X, Liu Q, Li S, et al. An “up, no change, or down” system: time-dependent expression of mRNAs in contused skeletal muscle of rats used for wound age estimation. Forensic Sci Int. (2017) 272:104–10. doi: 10.1016/j.forsciint.2017.01.012
68. Sun J, Zhu X, Li S, Dong T, Du Q. Measuring temporal expression, systematic response, and post-mortem stability to assess potential markers for estimating wound age: an example of Fosl1 in contused skeletal muscle. Australian J Forensic Sci. (2019) 51:158–70. doi: 10.1080/00450618.2017.1334824
69. Wang L-L, Zhao R, Liu C-S, Liu M, Li S-S, Li J-Y, et al. A fundamental study on the dynamics of multiple biomarkers in mouse excisional wounds for wound age estimation. J Forensic Leg Med. (2016) 39:138–46. doi: 10.1016/j.jflm.2016.01.027
70. Yagi Y, Murase T, Kagawa S, Tsuruya S, Nakahara A, Yamamoto T, et al. Immunohistochemical detection of CD14 and combined assessment with CD32B and CD68 for wound age estimation. Forensic Sci Int. (2016) 262:113–20. doi: 10.1016/j.forsciint.2016.02.031
71. Tian Z-L, Jiang S-K, Zhang M, Wang M, Li J-Y, Zhao R, et al. Detection of satellite cells during skeletal muscle wound healing in rats: time-dependent expressions of Pax7 and MyoD in relation to wound age. Int J Legal Med. (2016) 130:163–72. doi: 10.1007/s00414-015-1251-x
72. Li N, Bai R, Li C, Dang L, Du Q, Jin Q, Cao J, et al. Insight into molecular profile changes after skeletal muscle contusion using microarray and bioinformatics analyses. Biosci Rep. (2021) 41:BSR20203699. doi: 10.1042/BSR20203699
73. Ferrara M, Sessa F, Rendine M, Spagnolo L, De Simone S, Riezzo I, et al. A multidisciplinary approach is mandatory to solve complex crimes: a case report. Egypt J Forensic Sci. (2019) 9:11. doi: 10.1186/s41935-019-0116-8
74. Bacci S, Defraia B, Cinci L, Calosi L, Guasti D, Pieri L, Lotti V, et al. Immunohistochemical analysis of dendritic cells in skin lesions: correlations with survival time. Forensic Sci Int. (2014) 244:179–85. doi: 10.1016/j.forsciint.2014.08.024
Keywords: wound-age estimation, wound-age determination, postmortem miRNA, miRNA, post-mortem investigation
Citation: De Simone S, Giacani E, Bosco MA, Vittorio S, Ferrara M, Bertozzi G, Cipolloni L and La Russa R (2022) The Role of miRNAs as New Molecular Biomarkers for Dating the Age of Wound Production: A Systematic Review. Front. Med. 8:803067. doi: 10.3389/fmed.2021.803067
Received: 27 October 2021; Accepted: 22 December 2021;
Published: 14 January 2022.
Edited by:
Isabel Legaz, University of Murcia, SpainReviewed by:
Aniello Maiese, Sapienza University of Rome, ItalyCopyright © 2022 De Simone, Giacani, Bosco, Vittorio, Ferrara, Bertozzi, Cipolloni and La Russa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luigi Cipolloni, bHVpZ2kuY2lwb2xsb25pQHVuaWZnLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.