 Kinga Borsodi1
Kinga Borsodi1 Helga Balla1
Helga Balla1 Péter József Molnár1,2
Péter József Molnár1,2 Ádám Lénárt1
Ádám Lénárt1 István Kenessey3
István Kenessey3 András Horváth2Attila Keszthelyi2Miklós Romics2Attila Majoros2Péter Nyirády2Stefan Offermanns4
András Horváth2Attila Keszthelyi2Miklós Romics2Attila Majoros2Péter Nyirády2Stefan Offermanns4 Zoltán Benyó1*
Zoltán Benyó1*- 1Institute of Translational Medicine, Semmelweis University, Budapest, Hungary
- 2Department of Urology, Semmelweis University, Budapest, Hungary
- 32ndDepartment of Pathology, Semmelweis University, Budapest, Hungary
- 4Department of Pharmacology, Max Planck Institute for Heart and Lung Research, Bad Nauheim, Germany
Bradykinin (BK) has been proposed to modulate urinary bladder functions and implicated in the pathophysiology of detrusor overactivity. The present study aims to elucidate the signaling pathways of BK-induced detrusor muscle contraction, with the goal of better understanding the molecular regulation of micturition and identifying potential novel therapeutic targets of its disorders. Experiments have been carried out on bladders isolated from wild-type or genetically modified [smooth muscle-specific knockout (KO): Gαq/11-KO, Gα12/13-KO and constitutive KO: thromboxane prostanoid (TP) receptor-KO, cyclooxygenase-1 (COX-1)-KO] mice and on human bladder samples. Contractions of detrusor strips were measured by myography. Bradykinin induced concentration-dependent contractions in both murine and human bladders, which were independent of secondary release of acetylcholine, ATP, or prostanoid mediators. B2 receptor antagonist HOE-140 markedly diminished contractile responses in both species, whereas B1 receptor antagonist R-715 did not alter BK's effect. Consistently with these findings, pharmacological stimulation of B2 but not B1 receptors resembled the effect of BK. Interestingly, both Gαq/11- and Gα12/13-KO murine bladders showed reduced response to BK, indicating that simultaneous activation of both pathways is required for the contraction. Furthermore, the Rho-kinase (ROCK) inhibitor Y-27632 markedly decreased contractions in both murine and human bladders. Our results indicate that BK evokes contractions in murine and human bladders, acting primarily on B2 receptors. Gαq/11-coupled and Gα12/13-RhoA-ROCK signaling appear to mediate these contractions simultaneously. Inhibition of ROCK enzyme reduces the contractions in both species, identifying this enzyme, together with B2 receptor, as potential targets for treating voiding disorders.
Introduction
Bradykinin (BK) is a nonapeptide pro-inflammatory mediator with a diverse set of functions, including regulation of smooth muscle tone. It has been reported that BK induces smooth muscle contraction in the vascular system, airways (1, 2), ureter (3), uterus (4), colon (5), and prostate (6).
Bradykinin exerts its effects in smooth muscle tissues via two major subtypes of BK receptors: B1 and B2 (7). Despite their structural similarity, B1 and B2 receptors differ greatly regarding their expression profiles in tissues and their function as well (8). The amount of B1 receptors in healthy human tissues is negligible, however, as a consequence of inflammatory stimuli (e.g., IL-1β), tissue injury, and endogenous factors, their expression may increase rapidly (9). In contrast, B2 receptors' presence is constitutive and predominant under physiological conditions (8, 10). Expression of both B1 and B2 receptors has been reported in urinary bladder smooth muscle (UBSM), however, their role and signaling pathways remain to be elucidated (11). B1 and B2 receptors are members of the seven-transmembrane-domain, type 1 G protein-coupled receptor (GPCR) family, representing a major class of drug targets (10, 12).
Bradykinin is a multi-faceted mediator of urinary bladder functions as its receptors are expressed in practically all cell types of the bladder, including the mucosa, UBSM, afferent nerve fibers as well as cells of innate and acquired immunity (11–14). Interestingly, however, no voiding abnormalities have been reported in mice lacking both the B1 and B2 receptor genes (15) or in control rat bladders pretreated with the B2 receptor antagonist HOE-140 (16), indicating a limited role of BK in the micturition reflex under physiological conditions. In contrast, BK has been reported to increase bladder activity (17); furthermore, the expression of its receptors is increased markedly in association with voiding disorders (16, 18). Bradykinin has been proposed as a mediator of augmented bladder contractions in a rat model of overactive bladder (OAB) syndrome (19). According to this study, B1 and B2 receptor agonists induced higher contractile responses in bladder strips of the OAB as compared to the sham-operated group. In addition, under the above-mentioned conditions, non-resident inflammatory cells capable of facilitating BK production and sensitivity to BK may appear in the bladder, further enhancing the possibility of BK-related functional changes.
Although it has been known for a long time that BK is present in the urine, assumingly, it is not the primary source of BK's effect on bladder smooth muscle, as the intact urothelium forms a tight barrier highly impermeable for compounds in the urine. However, the mucosa layer of the bladder wall exhibits diverse functions in regulating bladder tone, including releasing numerous mediator molecules. Saban et al. reported that BK is released from the bladder mucosa under physiological conditions, moreover, the mucosa layer also plays a major role in degrading the peptide (20). Under pathological conditions, the balance of peptide release and degradation may be disturbed, and either overproduction or decreased degradation of the peptide may lead to symptoms of bladder smooth muscle overactivity. In addition, BK may facilitate the release of other inflammatory mediators, like prostanoids, which can induce detrusor muscle contraction (21, 22).
The general principles of smooth muscle contraction apply to the urinary bladder, and in order to understand better the actions of BK we focused on the signaling pathways of UBSM contraction. Generally, smooth muscle contraction is initiated by the elevation of intracellular Ca2+ concentration entering the cytoplasm either via cell membrane channels or from the sarcoplasmic reticulum (SR), resulting in the activation of myosin light chain (MLC) kinase that phosphorylates MLC, eventually leading to cross-bridge cycling between actin and myosin. A common stimulus for the elevation of intracellular Ca2+ concentration is GPCR activation. Gαq/11-coupled receptors stimulate phospholipase C-ß (PLC-ß) activity resulting in formation of inositol triphosphate and diacylglycerol (DAG). Inositol triphosphate binding to its receptors on the SR is the stimulus for intracellular Ca2+ release, which in turn may induce the opening of store-operated Ca2+ channels of the plasma membrane resulting in Ca2+-influx and further activation of MLC kinase.
Myosin light chain phosphorylation is also regulated by MLC phosphatase, which removes the phosphate from MLC and promotes relaxation. Gα12/13-coupled receptor signaling involves activation of the small G protein RhoA and consequently Rho-kinase (ROCK), which inactivates MLC phosphatase leading to a sustained contraction (23). This pathway is often referred to as the Ca2+-sensitizing pathway, as an increase in intracellular Ca2+ concentration is not needed for the contraction. The ROCK enzyme has gained attention recently in disorders associated with lower urinary tract smooth muscle contractility (24, 25). In addition, ROCK activity may be enhanced under pathological conditions, for instance, in cystitis (26) and during aging (27) highlighting its possible role in bladder dysfunctions.
This paper focuses on signal transduction pathways contributing to BK-induced detrusor smooth muscle contraction and their potential role in regulating bladder tone. Intracellular signaling of BK-induced contraction has been examined in several smooth muscle tissues (2, 4, 12). We aimed to elucidate whether the same pathways mediate its effect in the detrusor muscle as well and to find potential targets within the signaling cascade of BK-evoked contractions for intervention in UBSM dysfunctions.
Materials and Methods
Animals
All procedures were carried out according to the guidelines of the Hungarian Law of Animal Protection (28/1998) and were approved by the Government Office of Pest County (Permission number: PEI/001/2709-13/2014). Urinary bladders were obtained from adult (90-120-day-old, 30-35 g) male wild-type (WT) mice (C57BL/6N mice from Charles River Laboratories, Isaszeg, Hungary) and from animals deficient for the cyclooxygenase (COX)-1 enzyme (COX-1-KO) or thromboxane prostanoid (TP) receptor (TP-KO) or from mice with induced smooth muscle-specific deficiency of Gαq/11 or Gα12/13 proteins as described previously (Gαq/11-KO and Gα12/13-KO) (28, 29).
Cyclooxygenase-1 enzyme mice were kindly provided by Ingvar Bjarnason (Department of Medicine, Guy's, King's College, St. Thomas' School of Medicine, London, United Kingdom), and prostanoid receptor-deficient mice were from Shuh Narumiya (Kyoto University, Kyoto, Japan). In the case of the COX-1-KO studies, littermate COX-1+/+ animals, whereas in TP-KO experiments, WT C57Bl/6N mice served as controls, as the TP-KO strain has been previously backcrossed with C57Bl/6N mice for more than 10 generations.
The mouse lines with smooth muscle-specific inducible deletion of the Gαq/11 or Gα12/13 proteins were generated on Gα11-deficient (G) or Gα12-deficient (G) background (30, 31) with floxed alleles of the genes coding Gαq (Gnaqflox/flox) or Gα13 (Gnα13flox/flox), and expressing a fusion protein of the Cre-recombinase with a modified estrogen receptor-binding domain (Cre-ERT2) (32) under the control of the smooth muscle myosin heavy chain (SMMHC) promoter. Deletion of Gnaq or Gna13 was induced by intraperitoneal tamoxifen treatment (1 mg/day for five consecutive days) in SMMHC-CreERT2+/−; Gnaqflox/flox; Gna11−/− and SMMHC-CreERT2+/−; Gna12−/−; Gna13flox/flox mice, respectively, as described previously elsewhere (28, 29). Mice with floxed alleles but without Cre expression served as controls. More precisely, tamoxifen-treated SMMHC-CreERT2−/−; Gnaqflox/flox; Gna11+/+ and SMMHC-CreERT2−/−; Gna12+/+; Gna13flox/flox mice served as controls and are referred to as Gαq/11-CTRL and Gα12/13-CTRL.
Human Tissues
All procedures involving human urinary bladder tissues have been approved by the Scientific and Research Committee of the Medical Research Council of Hungary (License No.: 21545-2/2019/EKU). Human urinary bladder tissues were obtained from 19 patients (15 males, 4 females; mean age of 65.5 ± 9.3 years, range between 44 and 78 years) undergoing open radical cystectomy due to muscle-invasive bladder malignancy after having obtained written patient consent. None of the patients had any urodynamic disorders, symptoms of OAB syndrome, or was taking drugs for OAB.
Following the surgical removal of the bladders, they were immediately placed in physiological saline solution and transported to the 2nd Department of Pathology of the Semmelweis University, Budapest. Here, the healthy, tumor-free whole bladder wall tissue was provided by uro-pathologists within approximately 15–20 min following removal of the bladders from patients as described previously (33). The healthy bladder tissue was immediately placed into room temperature Hank's Balanced Salt Solution (HBSS) and transported to our myograph laboratory, where preparation of the smooth muscle strips was performed without delay. Overall, myographic experiments started within 45–60 min following bladder removal from the patients.
Preparation of Smooth Muscle Strips
Mice were euthanized by cervical dislocation under general anesthesia [i.p., Ketamine (300 mg/kg) + Xylazine (30 mg/kg)], the urinary bladders were removed from a lower midline incision and were placed into Krebs solution (119 mM NaCl, 4.7 mM KCl, 1.2 mM KH2PO4, 2.5 mM CaCl2·H2O, 1.2 mM MgSO4·7H2O, 20 mM NaHCO3, 0.03 mM EDTA, and 10 mM glucose, pH 7.4) at room temperature. Under a dissection microscope (M3Z; Wild Heerbrugg AG, Gais, Switzerland), adipose and connective tissues were removed from the serosal surface. Bladders were cut into four strips of equal lengths and, the mucosa layer was also gently removed to prevent the potential release of paracrine factors from the mucosal epithelium or submucosa and avoid tension changes related to myofibroblasts.
Human urinary bladder specimens were also placed into Krebs solution (same as described before, at room temperature) during the preparation. Under a dissection microscope, the serosal tissue and the mucosal layer were removed. The isolated detrusor muscle specimens were cut into equal, approximately 3 × 2 × 1 mm strips for myography.
Measurement of Bladder Contractility
Both murine and human detrusor muscle strips were mounted on two parallel, horizontal stainless-steel tissue-holding needles of a myograph (needle diameter 200 μm, 610 M Multi Wire Myograph System, Danish Myo Technology A/S, Aarhus, Denmark). Chambers were filled with 6 ml of Krebs solution aerated with carbogen (mixture of 5% CO2 and 95% O2) at 37°C. Detrusor muscle contractions were recorded under isometric conditions. Every experiment started with a 60-min resting period while the strips were stretched to and stabilized at a passive tension of 5 mN (murine) or 3 mN (human). After the resting period, UBSMs were challenged twice with 124 mM K+-containing Krebs solution to examine the viability of the tissues. The contractile effect of 124 mM K+ was comparable in the detrusor strips obtained from the WT and the genetically modified mouse lines (Supplementary Figure 1 at https://doi.org/10.6084/m9.figshare.16815061.v5). After several washes with normal Krebs solution, the contractile effects of BK (10−10-10−4 M), Lys-[Des-Arg9]-bradykinin (B1 receptor agonist, 10−5 M), [Phe8Ψ(CH-NH)-Arg9]-bradykinin (B2 receptor agonist, 10−5 M), carbamoylcholine chloride [carbachol (CCh, 10−6 M)], α,β-methyleneadenosine 5′-triphosphate [α,β-meATP, ATP-analog (10−5 M)] was measured. Some of the strips were preincubated with one of the following inhibitors without washing out: R-715 (Ac-Lys-Arg-Pro-Pro-Gly-Phe-Ser-DβNal-Ile, B1 receptor antagonist, 10−6 M, 20 min), HOE-140 (icatibant) (D-Arg-Arg-Pro-Hyp-Gly-Thi-Ser-D-Tic-Oic-Arg, B2 receptor antagonist, 10−6 M, 20 min), atropine [muscarinic-acetylcholine (ACh)-receptor antagonist, 10−6 M, 20 min], pyridoxalphosphate-6-azophenyl-2′,4′-disulfonate (PPADS; P2 purinergic receptor antagonist, 10−5 M, 20 min), Y-27632 [ROCK inhibitor, 10−5 M, 20 min], indomethacin [non-isoform-selective COX inhibitor, 10−5 M, 20 min], N-[2-Cyclohexyloxy-4-nitrophenyl]methanesulfonamide [NS-398, COX-2 inhibitor 10−5 M, 20 min]. When acetic acid, dimethyl sulfoxide (DMSO), or saline was the solvent of the inhibitor, they were applied in matched concentrations as vehicle control. The final concentration of acetic acid in the tissue bath was 0.1 mM, while that of DMSO was 0.1%. Since repeated use of BK or its analogs in the same specimen is problematic due to the rapid desensitization and the viability of the tissues may decline in a prolonged experiment, we performed unpaired, time-control experiments in this study. Finally, bladder strips were exposed to 124 mM K+-containing Krebs solution to retest the viability of the detrusor strips. Agonist-induced tension changes were normalized to the reference contraction induced by 124 mM K+-containing Krebs solution (second administration). A schematic diagram of our experimental protocol is demonstrated in Supplementary Figure 2 (at https://doi.org/10.6084/m9.figshare.16815097.v2).
MP100 system and AcqKnowledge 3.9.2 software from Biopac System (Goleta, CA) were used for the acquisition and analysis of myographic measurements. The moving average smoothening function of the software was applied on recordings solely in order to eliminate the noises arising from the bubbling of the medium and to reduce the high frequency—low amplitude spontaneous tension oscillations. The parameters of the smoothening filter were carefully chosen in order to eliminate only the noises but not to alter the amplitude of the BK-induced responses—the baseline and peak values were always compared before and after the smoothing. The sample rate of the recordings was 10 samples/s (10 Hz), the smoothing factor was between 10 and 40 samples. Spontaneous micro-contractions of the bladder strips were observed occasionally, however, they were not reproducible, thus these data were not considered for demonstration or evaluation in the present study.
Drugs and Solutions
Bradykinin was purchased from Bachem (Bubendorf, Switzerland) and dissolved in acetic acid (0.1 M) to stock solutions of 10−2 M. Lys-[Des-Arg9]-bradykinin, [Phe8Ψ(CH-NH)-Arg9]-bradykinin, HOE-140, and R-715 were purchased from Tocris (Bristol, UK) and were dissolved in saline. Stock solutions of Lys-[Des-Arg9]-bradykinin, [Phe8Ψ(CH-NH)-Arg9]-bradykinin, and R-715 were 10−3 M, whereas due to its poor solubility in water, stock solutions of HOE-140 were 5 × 10−4 M. Carbachol was from Sigma-Aldrich (St. Louis, MO) and dissolved in saline to a stock solution of 2 × 10−1 M. Atropine (atropinum sulfuricum) was purchased from Egis Pharmaceutical PLC (Budapest, Hungary) and was diluted in water to a stock solution of 1.44 × 10−4 M. α,β-Methyleneadenosine 5′-triphosphate, PPADS, and Y-27632 were purchased from Cayman Chemical (Ann Arbor, MI) and all substances were dissolved in saline (α,β-meATP: 10−2 M, PPADS: 10−2 M, and Y-27632: 10−3 M). Indomethacin was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in DMSO (10−2 M stock concentration), as its aqueous solutions are quite unstable (34). NS-398 was also purchased from Sigma-Aldrich (St. Louis, MO), and DMSO was applied as solvent for preparing a 10−2 M stock solution.
Data Analysis and Statistics
The maximum contraction was defined as the peak value of tension developed after the addition of agonists. Average curves of individual contraction responses were also determined and presented on the left side of the figures, where they were plotted as mean values. All data are presented with the median values except concentration-response curves, in which cases mean ± SEM were used. For mouse concentration-response curve analysis, curves were fitted for data from each experiment, thus Emax and EC50-values were determined for each curve, and the average values were calculated thereafter. In the case of human concentration-response correlation, curves were fitted on data gained from numerous experiments, as human tissues exhibit more variable responses which made curve-fitting from each individual experiment difficult.
For statistical analysis, data sets were subjected to non-parametric testing, as in the case of small sample sizes and skewed data, parametric testing might not be appropriate. In the case of comparing two data sets the Mann-Whitney test, while in the case of comparing several data sets, the Kruskal-Wallis test was performed for determining the corresponding p-values. The following formula was used for demonstrating case numbers: n = x/y, where x represents the number of the bladder strips and y indicates the number of bladders. Statistical analysis and graph plotting were performed with GraphPad Prism software (v.6.07; GraphPad Software Inc., La Jolla, CA, USA), and p < 0.05 was considered a statistically significant difference.
Results
Bradykinin Induces Concentration-Dependent Contractions in Mouse and Human Bladders With Similar Characteristics
First, we aimed to evaluate the effect of BK in murine and human urinary bladders. Bradykinin induced marked, transient contraction in the mouse bladder, although it did not reach the level of the response evoked by CCh, a stable analog of ACh, the main physiological mediator of detrusor muscle contraction (Figure 1A). The effect of BK was dose-dependent, with the EC50 of 1.24 μM and Emax of 52.4%, expressed as the percentage of the reference contraction induced by 124 mM KCl (Figure 1B). In human bladder strips, BK induced comparable contractions with a similar ratio to the CCh's effect to that observed in mice (Figure 1C). Bradykinin-induced contractions were also dose-dependent in human bladders with an EC50 of 5.1 μM and Emax at 42.4% (Figure 1D). As repeated administration of BK appeared to desensitize BK receptors both in murine and human bladders, the dose-response curves have been obtained by applying only one single concentration of BK to each muscle strip and also in the further experiments, we avoided repeated administration of BK. Based on the dose-response relationship presented in Figures 1B,D, we decided to apply BK in subsequent experiments in a concentration of 10 μM, which induces a submaximal contractile effect enabling the determination of the signaling pathways involved.
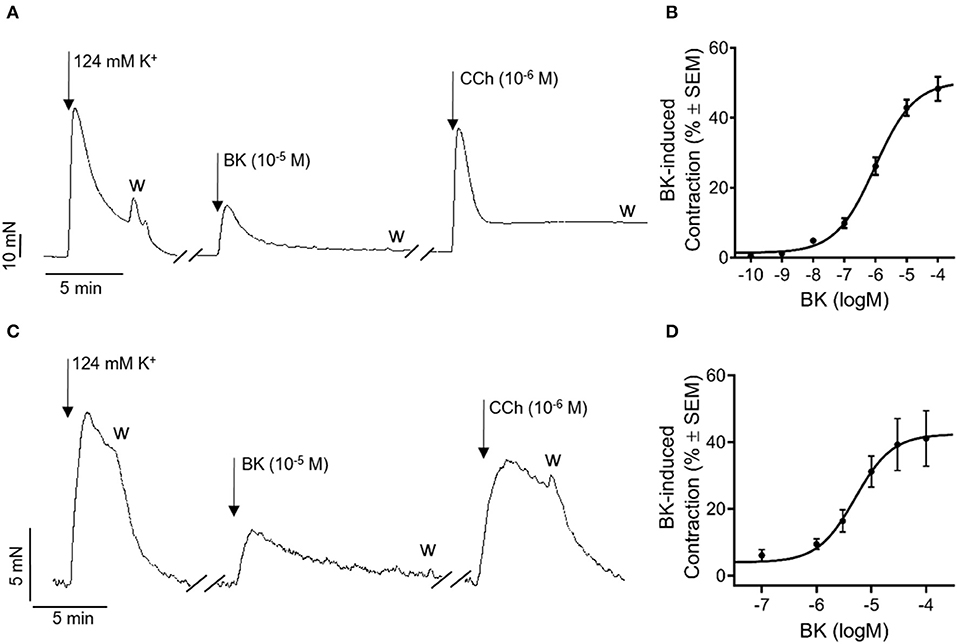
Figure 1. Bradykinin induces concentration-dependent contractions in mouse and human bladders with similar characteristics. Original trace: BK (10−5 M) evoked contractions in isolated murine detrusor smooth muscle strips which were comparable to the responses evoked by the muscarinic ACh receptor agonist CCh (10−6 M). (A) Concentration-response curve of BK in murine urinary bladder strips (Emax: 52.4% EC50: 1.2 μM) (B) Case numbers: n (10−10) = 2/2, n (10−9) = 3/3, n (10−8) = 3/3, n (10−7) = 6/6, n (10−6) = 9/7, n (10−5) = 4/4. n (10−4) = 10/10. Original trace: In accordance with our results gained from murine bladder strips, BK (10−5 M) evoked contraction in human detrusor smooth muscle as well. Moreover, the amplitude of the contractile effect was comparable to that induced by the muscarinic-acetylcholine-receptor agonist CCh (10−6 M). (C) Concentration-response curve representing BK's contractile effect in human detrusor smooth muscle. (Emax: 42.4%; EC50: 5.1 μM), (D) Case numbers: n (10−7) = 7/3, n (10−6) = 7/3, n (3 × 10−6) = 11/4, n (10−5) = 13/6, n (3 × 10−5) = 15/5, n (10−4) = 11/4.
Signaling Pathways of Bradykinin-Induced Detrusor Muscle Contractions
First, in order to test whether secondary ACh or ATP release from parasympathetic nerve fibers may mediate the effects of BK, we applied the muscarinic ACh receptor antagonist atropine (10−6 M, 20 min incubation) (Figure 2A) or the P2 purinergic receptor antagonist PPADS (10−5 M, 20 min) (Figure 2B). Neither atropine nor PPADS altered the effect of BK in mouse detrusor muscle, suggesting that the BK-induced contractile responses are independent of secondary ACh or ATP release (Figures 2A,B). To verify the effectiveness of atropine and PPADS, we applied the specific purinergic agonist α,ß-meATP, and the muscarinergic agonist CCh following preincubation with either PPADS or atropine, respectively (Supplementary Figures 3A,B at https://doi.org/10.6084/m9.figshare.14718657.v4). Pyridoxalphosphate-6-azophenyl-2′,4′-disulfonate abolished the contractile effect of α,ß-meATP, likewise, atropine inhibited CCh-induced contractions, which confirmed the effectiveness of the two inhibitors.
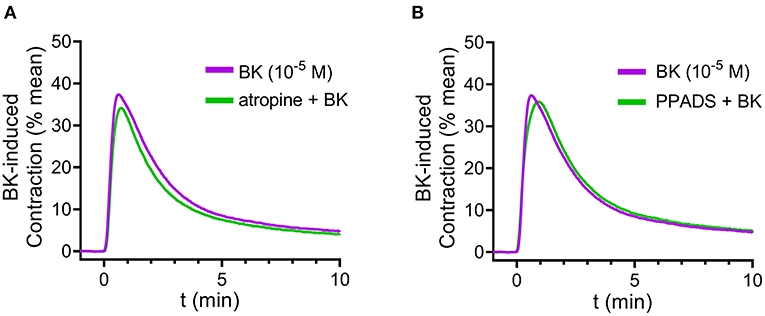
Figure 2. Bradykinin-induced detrusor muscle contraction is independent of purinergic or cholinergic neurotransmission. Neither inhibition of purinergic receptors with PPADS (10−5 M, 20 min incubation) nor the muscarinic receptor antagonist atropine (10−6 M, 20 min incubation) altered detrusor contraction induced by BK (10−5 M). (A,B) Case numbers: (A): BK: n = 6/6, atropine + BK: n = 7/7, (B): BK: n = 6/6, PPADS + BK: n = 8/8.
Data gained from human airway smooth muscle tissues suggested that BK may constrict bronchial smooth muscle through thromboxane A2 release resulting in TP receptor activation (1, 35). Thus, we also performed experiments with TP-KO mouse bladders, however, their contractile responses to BK were similar to those of the control bladder strips (Figure 3A).
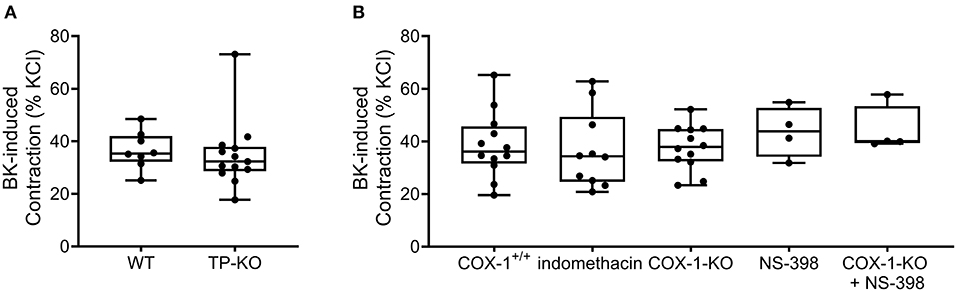
Figure 3. Bradykinin induces detrusor muscle contractions independently of COX-derived prostanoids. Contractile responses evoked by BK (10−5 M) were not altered in the bladder strips from mice deficient for TP receptors compared to those from WT mice. (A) The presence of the non-specific COX inhibitor indomethacin (10−5 M, 20 min) did not change BK-induced contractions. Furthermore, deficiency for COX-1 enzymes or treatment with the specific COX-2 inhibitor (NS-398, 10−5 M, 20 min) as well as their combination (COX-1-KO+NS-398) failed to influence the contractile effects elicited by BK. (B) (A: Mann-Whitey test, B: Kruskal-Wallis test). Case numbers: (A): WT: n = 8/4, TP-KO: n = 13/4, (B): COX-1+/+: n = 12/4 indomethacin: n = 10/5, COX-1-KO: n = 12/4, NS-398: n = 4/2, COX-1-KO + NS-398: n = 4/2.
Next, we aimed to analyze the potential involvement of other prostanoids in mediating the effect of BK. However, incubation of the smooth muscle strips with either the nonselective COX enzyme inhibitor indomethacin (10−5 M, 20 min) or with the selective COX-2 inhibitor NS-398 (10−5 M, 20 min) failed to alter the contractile responses induced by BK (Figure 3B). For studying the function of the COX-1 enzyme selectively, COX-1-KO mouse detrusor muscle tissues were subjected to BK, however, the contractions were unaffected in the absence of COX-1 enzyme as well. As COX-1 enzyme deletion may be compensated by COX-2 enzyme upregulation (36–38), we treated COX-1-KO bladders with NS-398 to address the possibility of such compensatory mechanism. Again, we found that the contractile effect of BK was unaltered, indicating that neither COX-1 nor COX-2 appears to be involved in mediating the response (Figure 3B).
Following verification of the direct contractile effect of BK in detrusor muscle, we investigated the role of B1 and B2 receptors in the contraction by applying the specific B1 receptor antagonist R-715 (10−6 M, 20 min incubation) and B2 receptor antagonist HOE-140 (10−6 M, 20 min incubation). Bradykinin-induced contractions were strongly inhibited by HOE-140, whereas R-715 failed to reduce them, implying that B2 receptors play the main role in mediating the effect of BK in UBSM (Figure 4A). Furthermore, simultaneous application of the two inhibitors abolished the contractile responses. Smooth muscle strips were also treated with specific agonists of B1 and B2 receptors. B1 agonist Lys-[Des-Arg9]-bradykinin (10−5 M) elicited only minor smooth muscle tone elevation, whereas the B2 agonist [Phe8Ψ(CH-NH)-Arg9]-bradykinin (10−5 M) had a potent constrictor effect in murine UBSM comparable to that of BK (Figure 4B) verifying the predominant role of B2.
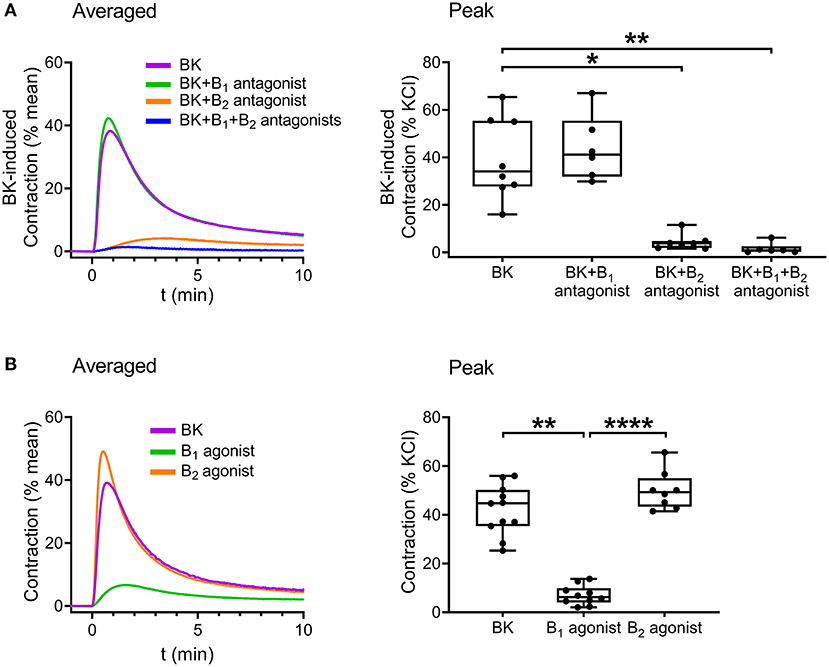
Figure 4. Role of B2 receptors in mediating BK-induced detrusor muscle contraction in murine urinary bladder strips. The BK (10−5 M)-induced contraction was abolished by the B2 receptor-specific antagonist HOE-140 (10−6 M, 20 min), whereas it failed to be reduced by the B1 receptor antagonist R-715 (10−6 M, 20 min). (A) The B2 receptor agonist (10−5 M) induced contractions of the same magnitude as BK, whereas the B1 receptor agonist (10−5 M) evoked only minor bladder contractions. (B) (A,B: Kruskal-Wallis test, *p < 0.05, **p < 0.01, ****p < 0.0001). Case numbers: (A): BK: n = 8/8, B1 antagonist: n = 6/6, B2 antagonist: n = 7/7, B1 + B2 antagonist: n = 6/6, (B): BK: n = 11/11, B1 agonist: n = 10/8, B2 agonist: n = 8/7.
Next, we intended to examine the contribution of Gαq/11- and Gα12/13-mediated pathways to the constriction evoked by B2 receptor activation. Contractile responses induced by BK (10−5 M) were reduced in Gαq/11-deficient UBSM compared to those of Gαq/11-CTRL murine bladders (Figure 5A). Interestingly, BK-induced contractions were also decreased in Gα12/13-KO mouse bladder (Figure 5B). Therefore, we aimed to investigate further the downstream signaling of Gα12/13-activation via application of the ROCK inhibitor Y-27632 (10−5 M, 20 min), which also reduced BK-evoked contractions in murine bladders (Figure 6A). In addition, the presence of Y-27632 (10−5 M, 20 min) completely abolished BK-induced contractions in Gαq/11-KO UBSM (Figure 6B).
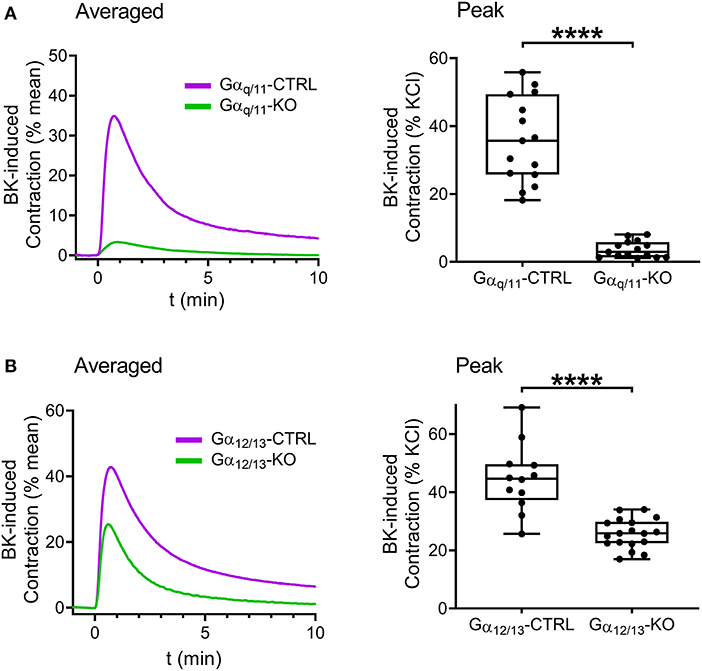
Figure 5. Gαq/11 and the Gα12/13 G proteins mediate the effects of BK in murine urinary bladder detrusor muscle. Contractile responses evoked by BK (10−5 M) were markedly reduced in the UBSM from Gαq/11-KO mice compared to bladder strips from Gαq/11-CTRL. (A) The detrusor contractions elicited by BK were also diminished in the UBSMs from Gα12/13-KO compared to the strips from Gα12/13-CTRL animals. (B) (A,B: Mann-Whitney test, ****p < 0.0001). Case numbers: (A): Gαq/11-CTRL: n = 15/4, Gαq/11-KO: n = 15/4, (B): Gα12/13-CTRL n = 12/3, Gα12/13-KO: n = 18/4.
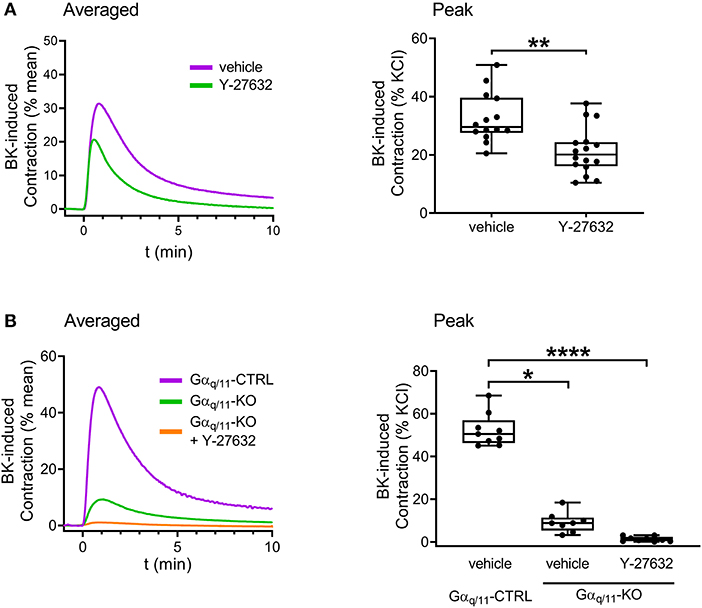
Figure 6. Role of Gα12/13-RhoA-ROCK pathway in mediating BK-induced contractions in murine UBSM. The contractile responses induced by BK (10−5 M) were decreased in the presence of the ROCK inhibitor (Y-27632, 10−5 M, 20 min). (A) The addition of Y-27632 (10−5 M, 20 min) completely suppressed the remaining BK-induced contractile responses in UBSM strips from Gαq/11-KO mice. (B) (A: Mann-Whitney test, B: Kruskal-Wallis test, *p < 0.05, **p < 0.01, and ****p < 0.0001). Case numbers: (A): vehicle: n = 14/8, Y-27632: n = 16/8, (B): vehicle: n = 9/4, Gαq/11-KO + vehicle: n = 8/4, Gαq/11-KO + Y-27632: n = 9/4.
Signaling Pathways of Bradykinin-Induced Contractions in the Human Urinary Bladder
Finally, the signaling pathways of BK in the human bladder were investigated. The contribution of BK receptor subtypes to the evoked contractions was examined by applying specific B1 and B2 receptor antagonists. The presence of the B2 antagonist HOE-140 (10−6 M, 20 min) almost completely abolished BK-induced contractions, whereas the B1 antagonist R-715 (10−6 M, 20 min) failed to alter contractions, indicating that B2 receptors play a prominent role in mediating BK-induced contractile responses of human UBSM as well (Figure 7A). Furthermore, the same selective B1 and B2 receptor agonists were applied to the human bladder strips as in the case of the murine experiments. The B2 agonist (10−5 M) evoked contractions approximately of the same magnitude as BK, however, the B1 agonist (10−5 M) had only a minor constricting activity compared to them (Figure 7B). Bladder strips were also treated with Y-27632 (10−5 M, 20 min) to examine the involvement of ROCK in mediating the effect of BK, and similarly to our murine results, the ROCK inhibitor decreased BK-induced contractile responses of the human detrusor muscle as well (Figure 8).
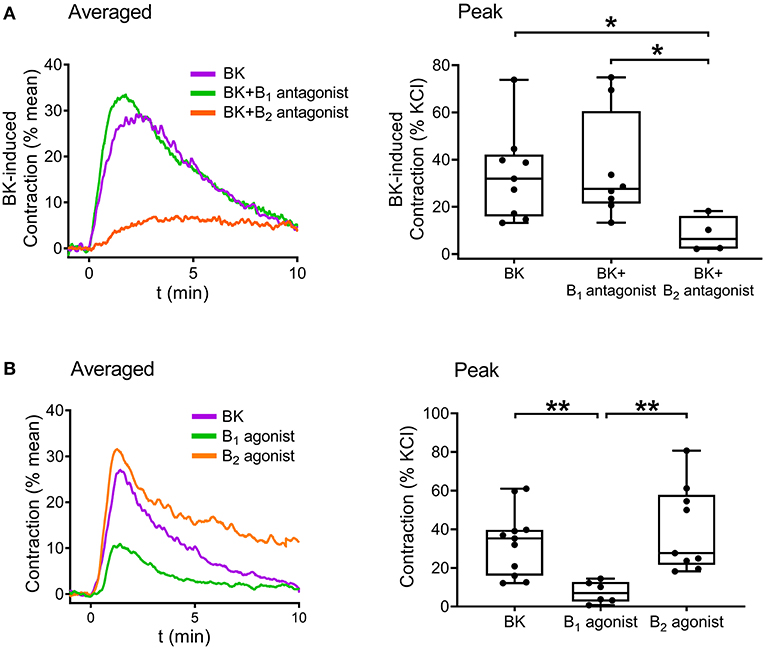
Figure 7. Bradykinin evokes concentration-dependent smooth muscle contraction in human urinary bladder mediated mostly by B2 receptors. Contractile responses evoked by BK (10−5 M) in human detrusor muscle were almost completely abolished in the presence of the B2 receptor antagonist HOE-140 (10−6 M, 20 min), whereas the B1 receptor antagonist R-715 (10−6 M, 20 min) failed to reduce BK-induced contractions. (A) The B2 receptor agonist (10−5 M) induced contractions similarly to BK, whereas the B1 receptor agonist (10−5 M) had only minor contractile activity in human detrusor muscle strips, similarly to our murine results. (B) (A,B: Kruskal-Wallis test, *p < 0.05, **p < 0.01). Case numbers: (A): BK: n = 9/3, B1 antagonist: n = 8/3, B2 antagonist: n = 4/3, (B): BK: n = 11/4, B1 agonist: n = 6/3, B2 agonist: n = 9/3.
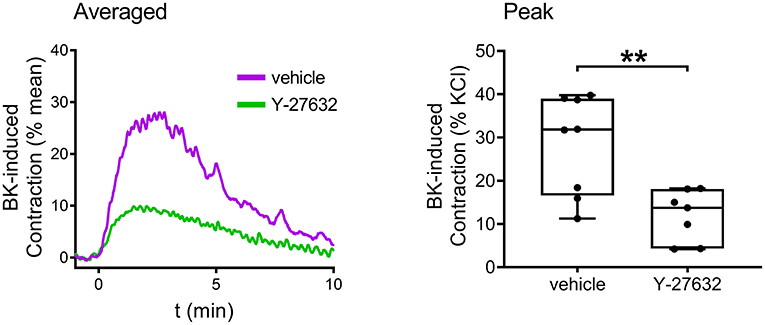
Figure 8. Role of the RhoA-ROCK pathway in mediating human detrusor muscle contraction evoked by BK. The ROCK inhibitor Y-27632 (10−5 M, 20 min) markedly reduced the contractions elicited by BK (10−5 M) in human UBSM, similarly to its effect in mouse bladder (Mann-Whitney test, **p < 0.01). Case numbers: vehicle: n = 8/2, Y-27632: n = 7/2.
Discussion
Bradykinin has been suggested as a potential mediator of disorders affecting the lower urinary tract, especially the urinary bladder (39, 40). Hence, we intended to outline the effect of BK in murine and human detrusor muscle, focusing on signal transduction of BK's actions in UBSM.
The present study demonstrates that BK evokes a concentration-dependent contraction in murine as well as in human detrusor muscle, although slight differences were observed in the contractile responses in the two species. Bradykinin contracts the mouse bladder smooth muscle with higher potency compared to that of humans (EC50: 1.2 × 10−6 M and 5.1 × 10−6 M, respectively), and the maximum of the contractile responses was also slightly higher in the case of mouse than in human bladders (Emax: 52 and 42%, respectively), although these differences were not statistically significant. Though, the differing values could perhaps be attributed to interspecies variance of BK receptor expression. Another notable difference between the two species regards the characteristics of contractions elicited by the receptor agonists and the high-concentration potassium solution. The decay of contractions evoked by either CCh, BK, α,ß-meATP, or KCl in human bladder required a longer time period compared to contractions evoked in mouse detrusor strips.
Although BK has been reported to induce changes in bladder functions via activation of neuronal circuits, our results obtained by using either the muscarinic receptor antagonist atropine or the purinergic receptor antagonist PPADS indicate that neither cholinergic nor purinergic neurotransmission contributes to the contractile effect of BK in our experimental settings. Accordingly, tetrodotoxin also failed to inhibit BK-induced contractions in rabbit detrusor muscle (41). Thus, BK's contractile effect appears to be independent of releasing neurotransmitters via activating BK receptors on nerve endings.
As it has been proposed previously that BK's effects may be mediated via prostaglandin (PG)-release, we were intrigued to know whether these mediators contribute to the BK-induced contractions in murine UBSM. Treatment of the bladder strips with the non-isoform-selective COX inhibitor indomethacin failed to change contractions in response to BK. According to previous studies, BK is more prone to exert its contractile effect via COX-1 enzyme activation and resultant PG release (21). However, under our experimental conditions, BK's contractile activity remained unaltered in COX-1-KO mouse UBSM strips. Moreover, neither the presence of COX-2 inhibitor NS-398 nor simultaneously abolishing the signaling of COX-1 and COX-2 isoenzymes via the addition of NS-398 to COX-1-KO bladder strips affected BK-induced contractions. Although a role of thromboxane A2 has also been suggested in BK-evoked smooth muscle contraction (1, 42), our results gained from TP-KO mouse bladders indicate that TP receptors are not involved in UBSM responses induced by BK. Based on the experiments summarized above, we have concluded that neither COX-1-nor COX-2-derived prostanoids play a significant role in BK-induced contractile effects in the mouse bladder. This is rather surprising, as several studies reported that BK's smooth muscle contractile effect may be indirect and is a result of secondary PG release (21, 43). However, it has also been proposed that BK-induced prostanoid release originates from the urothelium (44), which could not be evaluated in our present study as the urothelium had been intentionally removed during the preparation of the UBSM strips.
B2 receptor antagonist HOE-140 diminished the contractions almost completely, whereas the presence of B1 antagonist R-715 left BK's effect unchanged, indicating that B2 receptors mediate the BK-induced contractions in the murine detrusor muscle. Our conclusion is in line with expression data implying that B2 is the predominant BK receptor subtype in the UBSM under physiological conditions (45). Interestingly, using the B1 and B2 inhibitors concurrently completely abolished BK-induced contractions, indicating that in the absence of B2 receptors, the weak contractile effect mediated by B1 receptors was unmasked. However, this weak contractile effect may increase significantly when B1 receptors are upregulated under pathological conditions (9, 19).
The predominant role of B2 receptors in mediating BK-evoked contractions has also been proposed by Fabiyi and Brading in whole murine bladders (17). Interestingly, the EC50 value calculated for BK's concentration-response curves is different in our study compared to that published by Fabiyi and Brading (1.2 × 10−6 and 9 × 10−8 M, respectively), which, perhaps, is the result of the differences in the experimental setups (isolated detrusor strips vs. whole bladders).
Verification of the B2 receptor-mediated effect of BK in mouse UBSM was followed by exploring the intracellular signaling involved. Our experiments with Gαq/11-KO or Gα12/13-KO mouse bladder strips proved that both Gαq/11 and Gα12/13 protein deficiency results in reduced contractile effect of BK in murine UBSM. These findings indicate that both Gαq/11- and Gα12/13-coupled signaling pathways mediate contractions induced by BK via B2 receptors, which appears to be a specific feature of the UBSM, as most functions of B2 receptors are mediated exclusively by Gαq/11 proteins.
It has been demonstrated that the expression of ROCK enzyme in the urinary bladder is relatively high (46), and it may contribute to elevated bladder contractions in pathological conditions as mentioned previously in the introduction, thus we also investigated the role of ROCK inhibitor Y-27632 in BK-induced mouse detrusor contractions. As the inhibitor reduced BK-evoked contractions, we concluded that the ROCK enzyme plays an important role in the intracellular signaling of BK's contractile effect in the mouse bladder. Interestingly, however, Y-27632 also inhibited contractions elicited by CCh, α,ß-meATP, and 124 mM KCl solution, whereas it did not affect the resting bladder tone (Supplementary Figure 4 at https://doi.org/10.6084/m9.figshare.16815082.v5). These results show that the signaling pathways of the contractions evoked by various stimuli share a common effector with that of BK, the ROCK enzyme. An alternative interpretation of these findings could be that Y-27632 non-selectively decreases all contractions in the urinary bladder, and therefore our experimental data does not prove the involvement of ROCK in mediating the effects of BK. However, it is noteworthy that the involvement of the Rho-ROCK pathway has also been demonstrated in BK-induced contractions of the vascular and the airway smooth muscles (47–49). On the other hand, Ribeiro et al. reported recently that inhibition of ROCK by Y-27632 failed to alter the effect of BK in the pig intravesical ureter, indicating that not all BK-induced contractile effects in the lower urinary tract are sensitive to Y-27632 (3).
Finally, we simultaneously inhibited the two G-protein mediated signaling pathways by administration of Y-27632 to Gαq/11-KO bladders. As BK failed to elicit contraction under these conditions, we concluded that the Ca2+-dependent Gαq/11- and the Ca2+-sensitizing Gα12/13-pathways are the exclusive mediators of BK-evoked contractions in mouse UBSM, which we believe is a major conclusion of this paper.
Our observations regarding BK's contractile effect in murine UBSM and the associated pathways established a solid base for studying BK's role in human bladder smooth muscle. Although a few studies are available reporting the contractile effect of BK in human urinary bladder (50, 51), to our knowledge, no data have been presented regarding details of BK's contractile mechanism in human UBSM.
Our findings upon application of the selective BK receptor antagonists indicate that BK evokes its contractile response predominantly via B2 receptors in human bladder, which is in line with our murine results. This conclusion is affirmed by data gained via BK receptor agonist administration, as the B2 receptor agonist evoked contractions of the same magnitude as BK, in contrast to the B1 receptor agonist, which had only moderate contractile activity in human detrusor muscle. The prominent role of B2 receptors in BK-induced human UBSM contraction is also supported by data published by Belucci et al. (11), who demonstrated that mostly B2 receptors are expressed in human cultured detrusor smooth muscle cells. Nevertheless, one should note that our findings rely on experiments performed on tumor-free, healthy bladder tissues without exhibiting any signs of obvious pathological alterations. As mentioned previously, while B2 receptors are expressed constitutively in healthy tissues, B1 receptors' expression increases rather as a response to tissue injury or inflammation. Thus, the ratio of the two BK receptor's contribution to detrusor contraction evoked by BK may be altered under pathological conditions with increased B1 receptor expression in UBSM.
Based on our results in murine bladder, we predicted the role of ROCK within the intracellular signaling of B2 receptor-mediated detrusor muscle contraction also in humans. Indeed, our experiments with the ROCK-inhibitor Y-27632 demonstrate that ROCK plays a major role in BK-evoked contraction in human UBSM, as the inhibitor decreased contractile responses induced by BK. This is in accordance with our corresponding murine experiments, furthermore, we believe that this is the first study demonstrating that ROCK is a key mediator of BK's contractile activity in human UBSM. Similar to mouse experiments, the ROCK inhibitor reduced contractions induced by CCh, α,ß-meATP, and 124 mM KCl as well (Supplementary Figure 5 at https://doi.org/10.6084/m9.figshare.16815088.v5).
Taken together, this report presents for the first time a thorough investigation of signaling pathways contributing to BK's contractile effect in mouse and human detrusor muscle. It has been clarified that this effect results from BK acting predominantly on UBSM B2 receptors. The contractions are independent of cholinergic or purinergic neurotransmission as well as prostanoids. The role of the simultaneous contribution of Gq/11 and G12/13 protein-coupled pathways to the contraction was also demonstrated. Furthermore, the ROCK enzyme within the G12/13 protein signaling pathway also proved to be an important mediator, as its inhibition markedly decreased BK-induced contractions of both the murine and the human bladder. Interestingly, inhibition of ROCK has been shown to increase bladder compliance under both resting and stretched conditions, and ROCK inhibitors induce a relaxation of the bladder smooth muscle (52). Therefore, ROCK inhibitors represent a promising new tool for the treatment of detrusor overactivity. However, the appropriate dosage would definitely be an important issue and potentially a limitation of the use of ROCK inhibitors for the treatment of voiding disorders, as the normal micturition also appears to involve ROCK activation.
In conclusion, research on the role of the B2 receptor as well as ROCK in UBSM contraction may become a promising field for further studies and contribute to a better understanding of urinary bladder physiology and pathophysiology, which may lead to the development of novel drugs for the treatment of voiding disorders.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Scientific and Research Committee of the Medical Research Council of Hungary. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by Government Office of Pest County.
Author Contributions
KB, HB, PJM, PN, SO, and ZB conceived and designed research. AH and AK performed radical cystectomies and helped in patient selection. IK performed the pathological examination of the human urinary bladder samples. KB, HB, PJM, ÁL, and ZB performed experiments. KB and HB analyzed data, prepared figures, and drafted manuscript. KB, HB, PJM, IK, AH, AK, AM, PN, SO, and ZB interpreted the results of experiments. KB, HB, PJM, AM, PN, SO, and ZB edited and revised manuscript. KB, HB, PJM, ÁL, IK, AH, AK, MR, AM, PN, SO, and ZB approved the final version of manuscript.
Funding
This research was funded by the Hungarian National Research, Development and Innovation Office (K-125174, K-135683, K-139230, 2020-1.1.6-JÖVÖ-2021-00010, TKP2021-EGA-25, and NVKP_16-1-2016-0042 grants) as well as by the Higher Education Institutional Excellence Program of the Ministry of Human Capacities in Hungary, within the framework of the Molecular Biology thematic program of the Semmelweis University, and supported by the EFOP-3.6.3-VEKOP-16-2017-00009 and ÚNKP-19-2-I-SE-48 grants as well as by Gedeon Richter Plc Talent Foundation. The Central Library of Semmelweis University also kindly provided fund for open access publication fees.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are grateful to Ildikó Murányi for expert technical help and to Dr. Erzsébet Fejes for critically reading the manuscript.
Abbreviations
α, β-meATP, α, β-methyleneadenosine 5′-triphosphate; ACh, acetylcholine; ATP, adenosine triphosphate; BK, bradykinin; CCh, carbachol; COX, cyclooxygenase; Cre-ERT2, cre-recombinase with a modified estrogen receptor-binding domain; CTRL, control; DAG, diacylglycerol; DMSO, dimethyl sulfoxide; GPCR, G protein-coupled receptor; HBSS, Hank's Balanced Salt Solution; KO, knockout; MLC, myosin light chain; OAB, overactive bladder; PG, prostaglandin; PLC-β, phospholipase C-β; PPADS, pyridoxalphosphate-6-azophenyl-2′, 4′-disulfonate; ROCK, Rho-kinase; SMMHC, smooth muscle myosin heavy chain; SR, sarcoplasmic reticulum; TP, thromboxane prostanoid; UBSM, urinary bladder smooth muscle; WT, wild-type.
References
1. Molimard M, Martin CA, Naline E, Hirsch A, Advenier C. Role of thromboxane A2 in bradykinin-induced human isolated small bronchi contraction. Eur J Pharmacol. (1995) 278:49–54. doi: 10.1016/0014-2999(95)00103-r
2. Schlemper V, Calixto JB. Mechanisms underlying the contraction induced by bradykinin in the guinea pig epithelium-denuded trachea. Can J Physiol Pharmacol. (2002) 80:360–7. doi: 10.1139/y02-061
3. Ribeiro AS, Fernandes VS, Martinez MP, Lopez-Oliva ME, Barahona MV, Recio P, et al. Pre- and post-junctional bradykinin B2 receptors regulate smooth muscle tension to the pig intravesical ureter. Neurourol Urodyn. (2016) 35:115–21. doi: 10.1002/nau.22685
4. Wassdal I, Nicolaysen G, Iversen JG. Bradykinin causes contraction in rat uterus through the same signal pathway as oxytocin. Acta Physiol Scand. (1998) 164:47–52. doi: 10.1046/j.1365-201X.1998.00394.x
5. Zagorodnyuk V, Santicioli P, Maggi CA. Evidence for the involvement of multiple mechanisms in the excitatory action of bradykinin in the circular muscle of guinea-pig colon. Naunyn Schmiedebergs Arch Pharmacol. (1998) 357:197–204. doi: 10.1007/pl00005158
6. Steidle CP, Cohen ML, Neubauer BL. Bradykinin-induced contractions of canine prostate and bladder: effect of angiotensin-converting enzyme inhibition. J Urol. (1990) 144(2 Pt 1):390–2. doi: 10.1016/s0022-5347(17)39467-3
7. Bhoola KD, Figueroa CD, Worthy K. Bioregulation of kinins: kallikreins, kininogens, and kininases. Pharmacol Rev. (1992) 44:1–80.
8. Sharma J, Al-Sherif G. The Kinin system: present and future pharmacological targets. Am J Biomed Sci. (2011) 3:156–69. doi: 10.5099/aj110200156
9. Marceau F, Larrivée JF, Saint-Jacques E, Bachvarov DR. The kinin B1 receptor: an inducible G protein coupled receptor. Can J Physiol Pharmacol. (1997) 75:725–30. doi: 10.1139/y97-049
10. Howl J, Payne SJ. Bradykinin receptors as a therapeutic target. Expert Opin Ther Targets. (2003) 7:277–85. doi: 10.1517/14728222.7.2.277
11. Bellucci F, Cucchi P, Santicioli P, Lazzeri M, Turini D, Meini S. Characterization of kinin receptors in human cultured detrusor smooth muscle cells. Br J Pharmacol. (2007) 150:192–9. doi: 10.1038/sj.bjp.0706976
12. Leeb-Lundberg LM, Marceau F, Muller-Esterl W, Pettibone DJ, Zuraw BL. International union of pharmacology. XLV Classification of the kinin receptor family: from molecular mechanisms to pathophysiological consequences. Pharmacol Rev. (2005) 57:27–77. doi: 10.1124/pr.57.1.2
13. de Groat WC. Highlights in basic autonomic neuroscience: contribution of the urothelium to sensory mechanisms in the urinary bladder. Auton Neurosci. (2013) 177:67–71. doi: 10.1016/j.autneu.2013.03.010
14. Scharfstein J, Ramos PIP, Barral-Netto M. G Protein-Coupled Kinin Receptors and Immunity Against Pathogens. Adv Immunol. (2017) 136:29–84. doi: 10.1016/bs.ai.2017.05.007
15. Kakoki M, McGarrah RW, Kim HS, Smithies O. Bradykinin B1 and B2 receptors both have protective roles in renal ischemia/reperfusion injury. Proc Natl Acad Sci U S A. (2007) 104:7576–81. doi: 10.1073/pnas.0701617104
16. Chopra B, Barrick SR, Meyers S, Beckel JM, Zeidel ML, Ford AP, et al. Expression and function of bradykinin B1 and B2 receptors in normal and inflamed rat urinary bladder urothelium. J Physiol. (2005) 562(Pt 3):859–71. doi: 10.1113/jphysiol.2004.071159
17. Fabiyi AC, Brading AF. The use of the isolated mouse whole bladder for investigating bladder overactivity. J Pharmacol Exp Ther. (2006) 319:1386–94. doi: 10.1124/jpet.106.108902
18. Busser BW, Hammond TG, Bjorling DE, Saban R. Lipopolysaccharide upregulates bradykinin 1 receptors in the isolated mouse bladder. J Urol. (1998) 160:2267–73. doi: 10.1097/00005392-199812010-00098
19. Forner S, Andrade EL, Martini AC, Bento AF, Medeiros R, Koepp J, et al. Effects of kinin B(1) and B(2) receptor antagonists on overactive urinary bladder syndrome induced by spinal cord injury in rats. Br J Pharmacol. (2012) 167:1737–52. doi: 10.1111/j.1476-5381.2012.02127.x
20. Saban R, Franz J, Bjorling DE. Spontaneously released substance P and bradykinin from isolated guinea-pig bladder. BJU Int. (1997) 79:516–24. doi: 10.1046/j.1464-410x.1997.00092.x
21. Meini S, Lecci A, Cucchi P, Catalioto RM, Criscuoli M, Maggi CA. Inflammation modifies the role of cyclooxygenases in the contractile responses of rat detrusor smooth muscle to kinin agonists. J Pharmacol Exp Ther. (1998) 287:137–43.
22. Sjuve R, Boels PJ, Uvelius B, Arner A. Up-regulation of bradykinin response in rat and human bladder smooth muscle. J Urol. (2000) 164:1757–63. doi: 10.1016/s0022-5347(05)67102-9
23. Kao CY, Carsten ME. Cellular Aspects of Smooth Muscle Function. Cambridge; New York, NY: Cambridge University Press (2005).
24. Peters SL, Schmidt M, Michel MC. Rho kinase: a target for treating urinary bladder dysfunction? Trends Pharmacol Sci. (2006) 27:492–7. doi: 10.1016/j.tips.2006.07.002
25. Anjum I. (2018). Calcium sensitization mechanisms in detrusor smooth muscles. J Basic Clin Physiol Pharmacol. 29:227–35. doi: 10.1515/jbcpp-2017-0071
26. Shimizu N, De Velasco MA, Umekawa T, Uemura H, Yoshikawa K. Effects of the Rho kinase inhibitor, hydroxyfasudil, on bladder dysfunction and inflammation in rats with HCl-induced cystitis. Int J Urol. (2013) 20:1136–43. doi: 10.1111/iju.12119
27. Kirschstein T, Protzel C, Porath K, Sellmann T, Kohling R, Hakenberg OW. Age-dependent contribution of Rho kinase in carbachol-induced contraction of human detrusor smooth muscle in vitro. Acta Pharmacol Sin. (2014) 35:74–81. doi: 10.1038/aps.2013.126
28. Wirth A, Benyo Z, Lukasova M, Leutgeb B, Wettschureck N, Gorbey S, et al. G12-G13-LARG-mediated signaling in vascular smooth muscle is required for salt-induced hypertension. Nat Med. (2008) 14:64–8. doi: 10.1038/nm1666
29. Dér B, Molnár PJ, Ruisanchez É, Orsy P, Kerék M, Faragó B, et al. NK2 receptor-mediated detrusor muscle contraction involves G(q/11)-dependent activation of voltage-dependent Ca(2+) channels and the RhoA-Rho kinase pathway. Am J Physiol Renal Physiol. (2019) 317:F1154–63. doi: 10.1152/ajprenal.00106.2019
30. Wettschureck N, Rutten H, Zywietz A, Gehring D, Wilkie TM, Chen J, et al. Absence of pressure overload induced myocardial hypertrophy after conditional inactivation of Galphaq/Galpha11 in cardiomyocytes. Nat Med. (2001) 7:1236–40. doi: 10.1038/nm1101-1236
31. Moers A, Nieswandt B, Massberg S, Wettschureck N, Grüner S, Konrad I, et al. G13 is an essential mediator of platelet activation in hemostasis and thrombosis. Nat Med. (2003) 9:1418–22. doi: 10.1038/nm943
32. Indra AK, Warot X, Brocard J, Bornert JM, Xiao JH, Chambon P, et al. Temporally-controlled site-specific mutagenesis in the basal layer of the epidermis: comparison of the recombinase activity of the tamoxifen-inducible Cre-ER(T) and Cre-ER(T2) recombinases. Nucleic Acids Res. (1999) 27:4324–7. doi: 10.1093/nar/27.22.4324
33. Molnár PJ, Dér B, Borsodi K, Balla H, Borbás Z, Molnár K, et al. Isoprostanes evoke contraction of the murine and human detrusor muscle via activation of the thromboxane prostanoid TP receptor and Rho kinase. Am J Physiol Renal Physiol. (2021) 320:F537–47. doi: 10.1152/ajprenal.00400.2020
34. Curry SH, Brown EA, Kuck H, Cassin S. Preparation and stability of indomethacin solutions. Can J Physiol Pharmacol. (1982) 60:988–92. doi: 10.1139/y82-139
35. Hulsmann AR, Raatgeep HR, Saxena PR, Kerrebijn KF, de Jongste JC. Bradykinin-induced contraction of human peripheral airways mediated by both bradykinin beta 2 and thromboxane prostanoid receptors. (1994) 150:1012–8. doi: 10.1164/ajrccm.150.4.7921430
36. Li X, Ballantyne LL, Crawford MC, Fitzgerald GA, Funk CD. Isoform-specific compensation of cyclooxygenase (Ptgs) genes during implantation and late-stage pregnancy. Sci Rep. (2018) 8:12097. doi: 10.1038/s41598-018-30636-x
37. Li X, Mazaleuskaya LL, Ballantyne LL, Meng H, Fitzgerald GA, Funk CD. Genomic and lipidomic analyses differentiate the compensatory roles of two COX isoforms during systemic inflammation in mice. J Lipid Res. (2018) 59:102–12. doi: 10.1194/jlr.m080028
38. Li X, Mazaleuskaya LL, Yuan C, Ballantyne LL, Meng H, Smith WL, et al. Flipping the cyclooxygenase (Ptgs) genes reveals isoform-specific compensatory functions. J Lipid Res. (2018) 59:89–101. doi: 10.1194/jlr.m079996
39. Watts SW, Cohen ML. Effect of bombesin, bradykinin, substance P and CGRP in prostate, bladder body and neck. Peptides. (1991) 12:1057–62. doi: 10.1016/0196-9781(91)90060-3
40. Yoshimura N, Kaiho Y, Miyazato M, Yunoki T, Tai C, Chancellor MB, et al. Therapeutic receptor targets for lower urinary tract dysfunction. Naunyn Schmiedebergs Arch Pharmacol. (2008) 377:437–48. doi: 10.1007/s00210-007-0209-z
41. Nakahata N, Ono T, Nakanishi H. Contribution of prostaglandin E2 to bradykinin-induced contraction in rabbit urinary detrusor. Japan J Pharmacol. (1987) 43:351–9. doi: 10.1254/jjp.43.351
42. Weinberg ML, Moreira E, Weinberg J. Arachidonic acid products-mediated contraction induced by bradykinin in relaxed mesenteric arterial rings from Holtzman rats. Eur J Pharmacol. (1997) 320:145–50. doi: 10.1016/s0014-2999(96)00896-5
43. Tsuda Y, Nakahara T, Mori A, Sakamoto K, Ishii K. Resveratrol prevents bradykinin-induced contraction of rat urinary bladders by decreasing prostaglandin production and calcium influx. Eur J Pharmacol. (2011) 666:189–95. doi: 10.1016/j.ejphar.2011.05.019
44. Pinna C, Zanardo R, Puglisi L. Prostaglandin-release impairment in the bladder epithelium of streptozotocin-induced diabetic rats. Eur J Pharmacol. (2000) 388:267–73. doi: 10.1016/s0014-2999(99)00833-x
45. Marceau F, Bachvarov DR. Kinin receptors. Clin Rev Allergy Immunol. (1998) 16:385–401. doi: 10.1007/bf02737658
46. Wibberley A, Chen Z, Hu E, Hieble JP, Westfall TD. Expression and functional role of Rho-kinase in rat urinary bladder smooth muscle. Br J Pharmacol. (2003) 138:757–66. doi: 10.1038/sj.bjp.0705109
47. Cazzola M, Calzetta L, Rogliani P, Lauro D, Novelli L, Page CP, et al. (2012). High glucose enhances responsiveness of human airways smooth muscle via the Rho/ROCK pathway. Am J Respir Cell Mol Biol. 47:509–16. doi: 10.1165/rcmb.2011-0449OC
48. Zhang J, Yang GM, Zhu Y, Peng XY, Liu LM, Li T. Bradykinin induces vascular contraction after hemorrhagic shock in rats. J Surg Res. (2015) 193:334–43. doi: 10.1016/j.jss.2014.06.033
49. Shaifta Y, Mackay CE, Irechukwu N, O'Brien KA, Wright DB, Ward JPT, et al. Transforming growth factor-? enhances Rho-kinase activity and contraction in airway smooth muscle via the nucleotide exchange factor ARHGEF1. J Physiol. (2018) 596:47–66. doi: 10.1113/jp275033
50. Andersson KE, Hedlund H, Stahl M. Contractions induced by angiotensin I, angiotensin II and bradykinin in isolated smooth muscle from the human detrusor. Acta Physiol Scand. (1992) 145:253–9. doi: 10.1111/j.1748-1716.1992.tb09362.x
51. Meini S, Patacchini R, Giuliani S, Lazzeri M, Turini D, Maggi CA, et al. Characterization of bradykinin B2 receptor antagonists in human and rat urinary bladder. Eur J Pharmacol. (2000) 388:177–82. doi: 10.1016/S0014-2999(99)00882-1
Keywords: urinary bladder smooth muscle, bradykinin, signal transduction, Rho-kinase, detrusor reactivity
Citation: Borsodi K, Balla H, Molnár PJ, Lénárt Á, Kenessey I, Horváth A, Keszthelyi A, Romics M, Majoros A, Nyirády P, Offermanns S and Benyó Z (2022) Signaling Pathways Mediating Bradykinin-Induced Contraction in Murine and Human Detrusor Muscle. Front. Med. 8:745638. doi: 10.3389/fmed.2021.745638
Received: 22 July 2021; Accepted: 23 December 2021;
Published: 20 January 2022.
Edited by:
Guiming Liu, Case Western Reserve University, United StatesReviewed by:
Timo Kirschstein, University of Rostock, GermanyMartin C. Michel, Johannes Gutenberg University Mainz, Germany
Copyright © 2022 Borsodi, Balla, Molnár, Lénárt, Kenessey, Horváth, Keszthelyi, Romics, Majoros, Nyirády, Offermanns and Benyó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zoltán Benyó, YmVueW8uem9sdGFuQG1lZC5zZW1tZWx3ZWlzLXVuaXYuaHU=