
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Med. , 27 January 2021
Sec. Infectious Diseases – Surveillance, Prevention and Treatment
Volume 7 - 2020 | https://doi.org/10.3389/fmed.2020.603961
This article is part of the Research Topic Coronavirus Disease (COVID-19): Pathophysiology, Epidemiology, Clinical Management and Public Health Response, Volume II View all 165 articles
 Caroline Ruetsch1,2†
Caroline Ruetsch1,2† Vesna Brglez3†
Vesna Brglez3† Marion Crémoni3Kévin Zorzi3Céline Fernandez3
Marion Crémoni3Kévin Zorzi3Céline Fernandez3 Sonia Boyer-Suavet3
Sonia Boyer-Suavet3 Sylvia Benzaken1Elisa Demonchy4Karine Risso4
Sylvia Benzaken1Elisa Demonchy4Karine Risso4 Johan Courjon4Eric Cua4Carole Ichai5Jean Dellamonica3,5Thierry Passeron2,6
Johan Courjon4Eric Cua4Carole Ichai5Jean Dellamonica3,5Thierry Passeron2,6 Barbara Seitz-Polski1,3*
Barbara Seitz-Polski1,3*Coronavirus disease 2019 (COVID-19) caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has emerged in Wuhan in December 2019 and has since spread across the world. Even though the majority of patients remain completely asymptomatic, some develop severe systemic complications. In this prospective study we compared the immunological profile of 101 COVID-19 patients with either mild, moderate or severe form of the disease according to the WHO classification, as well as of 50 healthy subjects, in order to identify functional immune factors independently associated with severe forms of COVID-19. Plasma cytokine levels, and cytokine levels upon in vitro non-specific stimulation of innate and adaptive immune cells, were measured at several time points during the course of the disease. As described previously, inflammatory cytokines IL1β, IL6, IL8, and TNFα associated with cytokine storm were significantly increased in the plasma of moderate and severe COVID-19 patients (p < 0.0001 for all cytokines). During follow-up, plasma IL6 levels decreased between the moment of admission to the hospital and at the last observation carried forward for patients with favorable outcome (p = 0.02148). After in vitro stimulation of immune cells from COVID-19 patients, reduced levels of both type I and type II interferons (IFNs) upon in vitro stimulation were correlated with increased disease severity [type I IFN (IFNα): p > 0.0001 mild vs. moderate and severe; type II IFN (IFNγ): p = 0.0002 mild vs. moderate and p < 0.0001 mild vs. severe] suggesting a functional exhaustion of IFNs production. Stimulated IFNα levels lower than 2.1 pg/ml and IFNγ levels lower than 15 IU/mL at admission to the hospital were associated with more complications during hospitalization (p = 0.0098 and p =0.0002, respectively). A low IFNγ level was also confirmed by multivariable analysis [p = 0.0349 OR = 0.98 (0.962; 0.999)] as an independent factor of complications. In vitro treatment with type IFNα restored type IFNγ secretion in COVID-19 patients while the secretion of pro-inflammatory cytokines IL6 and IL1β remained stable or decreased, respectively. These results (a) demonstrate a functional exhaustion of both innate and adaptive immune response in severe forms of COVID-19; (b) identify IFNα and IFNγ as new potential biomarkers of severity; and (c) highlight the importance of targeting IFNs when considering COVID-19 treatment in order to re-establish a normal balance between inflammatory and Th1 effector cytokines.
In the beginning of December 2019 the first cases of a viral pneumonia of unknown origin were identified in Wuhan, the capital of the Hubei province in China (1, 2). The virus responsible has been identified as a new beta coronavirus now called severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) from the same family as SARS-CoV responsible for the SARS outbreak in 2003. This coronavirus which causes the new coronavirus disease (COVID-19) has since spread across the world and caused a pandemic (3, 4).
Common symptoms in patients with COVID-19 include fever, dry cough, anosmia, shortness of breath and other flu-like symptoms (3–5). Even though the majority of patients may remain completely asymptomatic or may present with only mild symptoms, 10–20% of patients progress to severe disease characterized by severe pneumonia, acute respiratory distress and multiple organ failure, requiring immediate hospitalization in intensive care units, and often leading to death (3, 6, 7). Severe clinical symptoms such as diffuse alveolar damage, thrombosis, haemophagocytosis, and immune cell depletion have been described in the subset of patients with severe COVID-19 (8). Patients suffering from diabetes, cancer or other chronic diseases are most at risk of developing a severe form (9).
To better stratify patients who might be at risk for complications, numerous studies identified biological markers of worse prognosis, such as lymphopenia, and inflammatory markers such as C-reactive protein (CRP), lactate dehydrogenase (LDH), and cytokine levels (3, 10–15).
Several authors investigated the role of different cytokines in COVID-19 patients. As a part of the immunological response to a SARS-CoV-2 infection, patients often display an aggressive and uncontrolled inflammatory response with a secretion of large amounts of pro-inflammatory cytokines such as interleukin (IL) 6, IL10 and tumor necrosis factor α (TNFα), in an event known as cytokine storm (3, 16–22). Cytokine storm is directly correlated with lung injury, multiple organ failure, and unfavorable prognosis (19).
The interplay between the innate and adaptive immune response seems to be crucial in determining the patient's evolution, characterized by an imbalance of pro- and anti-inflammatory cytokines and the subsequent dysregulation of patient's immune response (23). Interferons (IFNs) act as a key link between the innate and the adaptive immune response. Type I IFNs (IFN-α/β) are secreted by plasmacytoid dendritic cells (pDCs), while type II IFNs (IFN-γ) are predominantly produced by natural killer cells and in minor proportion by T cells and macrophages (24–26). Both type I and type II IFNs have a plethora of antiviral effects such as inducing apoptosis of infected cells and activating macrophages, natural killer (NK) cells and T lymphocytes (24–26). In COVID-19 patients, several studies have shown a dysregulation of IFNs production (14, 27, 28).
In search of new and powerful biomarkers of unfavorable outcome in COVID-19 patients, we analyzed the capability of the immune system response by the means of in vitro stimulation of both adaptive and innate immune cells, thus effectively mimicking a viral infection. Indeed, in vitro stimulation of innate and adaptive immunity cells has previously been shown to be predictive of worse outcome in other immune-related diseases (29, 30), but has to our knowledge not yet been investigated in COVID-19 patients. In this prospective, single-center study we compared the function of innate and adaptive immune cells of COVID-19 patients with either mild, moderate or severe form of the disease, aiming to underline the mechanism responsible for the dysregulation of immune response.
We performed a prospective cohort study at Nice University Hospital. The inclusion criteria were: (1) all adult patients admitted for COVID-19 in consultation unit (dermatology or infectious diseases unit), in infectious diseases units or in intensive care unit, in Nice University Hospital, from March to April 2020; (2) not having received immunosuppressive therapy in the 6 months prior to inclusion; (3) ability to sign an informed consent. Exclusion criteria were: (1) all patients under 18; (2) patients under custody, in prison or with a mental illness; (3) pregnant or breastfeeding; (4) with a known immunodeficiency or having received previous immunosuppressive therapy. Fifty healthy donors not infected with SARS-CoV-2 were also recruited (confirmed by negative serological test).
According to the severity of infection with SARS-CoV-2, the patients were divided in three groups: (a) patients with a severe form of COVID-19 were those hospitalized or transferred in the intensive care unit with respiratory distress or respiratory failure requiring mechanical ventilation or multiple organ failure; (b) patients with a moderate from of COVID-19 were patients hospitalized in the infectious diseases units, defined by clinical symptoms associated with dyspnea and radiological findings of pneumonia on thoracic CT scan; (c) COVID-19 patients with mild symptoms of COVID such as chilblains in fingers and toes or flu-like symptoms not requiring hospital supervision. All patients presented a COVID-19 symptomatology according to WHO classification with a CT scan characteristic of COVID-19 (31) or chilblains (32) or two consecutive positive RT-PCR tests for SARS-CoV-2 on upper and lower respiratory tract specimens (nasopharyngeal swab or invasive respiratory sample) or positive serological test (Euroimmun® ELISA).
Epidemiological, biological and clinical data at day 0 (D0) are reported in Table 1 and Supplementary Table 1. Treatment(s) received after D0 are summarized in Supplementary Table 2. Complications were defined as all adverse events such as admission in intensive care unit after worsening of the symptoms, mechanical ventilation, deep vein thrombosis, secondary bacterial infection, kidney failure, hepatitis, heart failure and death.
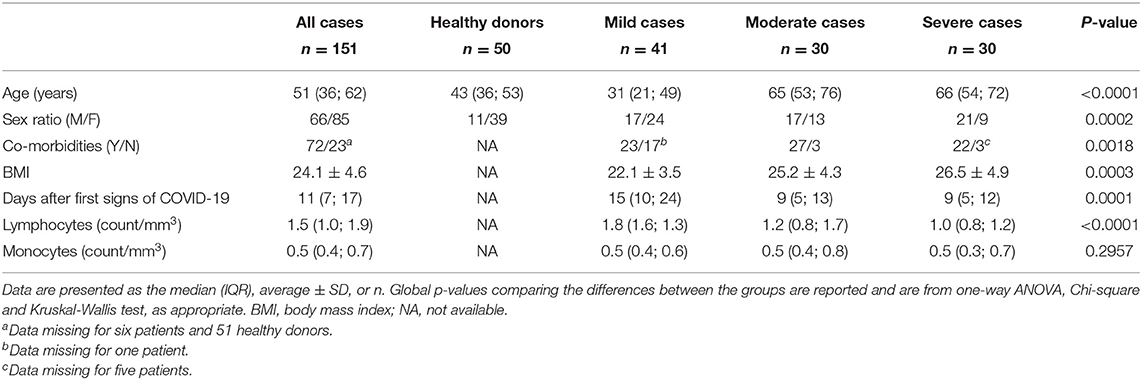
Table 1. Demographics and baseline characteristics of healthy donors and of patients with COVID-19.
An informed consent was obtained for all patients. The study protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki and was approved by the appropriate institutional review committee (NCT04355351).
Blood samples were collected at D0 and several follow-up time points up to 2 months after admission to the hospital. One milliliter of whole blood was stimulated with immune ligands (anti-CD3 as T-cells stimulant, and R848 as Toll-like receptors 7/8 (TLR 7/8) agonist) on single lyophilized spheres (LyoSphereTM, Qiagen) within 8 h from blood collection. Stimulated blood samples were incubated for 16–24 h at 37°C and then centrifuged at 2,000 to 3,000 × g for 15 min to harvest the stimulated serum. Non-stimulated serum and plasma and stimulated serum were stored at −20°C until the analysis and freeze-thaw cycles were minimized to preserve the quality of the samples. Serum and plasma levels of cytokines with or without non-specific stimulation were measured using either QuantiFERON-Monitor test for the detection of IFN-γ, or custom-designed cartridges Ella (ProteinSimple) for the detection of IL-1β, IL-6, IL-8, IL-10, IL-17A, TNF-α, and IFN-α, following the manufacturers' instructions.
For 18 COVID-19 patients, one milliliter of whole blood taken at D0 was pretreated with different molecules for 6 h at 37°C, followed by stimulation with immune ligands on single lyophilized spheres (LyoSphereTM, Qiagen), as described under Blood collection and cytokine assay. The molecules used were those commonly administered to COVID-19 patients (33–43): hydroxychloroquine (100 μM, Inresa), anti-IL6 Tocilizumab (100 μg/mL, RoActemra, Roche), methylprednisolone (20 μg/mL, Mylan), anti-TNFα Adalimumab (10 μg/mL, Humira, AbbVie), recombinant human IL-2 (6 ng/mL, Sigma), recombinant human IFN-alpha (100 ng/mL, Sigma) and Nivolumab (1 μg/mL, Opdivo, Bristol Myers Squibb).
For descriptive statistics, data are presented as mean and standard deviation for continuous values with Gaussian distribution, as median and range for continuous values with non-Gaussian distribution, and as counts and percentages for categorical variables. The D'Agostino & Pearson normality test was used to determine if a variable had a Gaussian distribution or not. Groups of continuous values were compared by the Mann-Whitney test, one-way ANOVA (>2 groups), or Kruskal-Wallis test (>2 groups). Multiple comparison tests were performed with Kruskal-Wallis test using Dunn's post hoc test. Categorical variables were compared using Chi-square test. AUC (Area Under the Curve) ROC (Receiver Operating Characteristic) curve was used to define an IFN-γ threshold that best discriminates patients with or without complications. Log-rank test was used to compare survival data. A Wilcoxon matched pairs signed rank test was used to compare two measurements of a continuous variable performed on the same subjects (paired data). Logistic regression were performed to determine ODDS ratios and 95% confidence intervals (CI). In the mulitivariable model, we adjusted for age, sex and BMI.
Statistical analyses were performed using GraphPad Prism 7.0 (GraphPad Software, Inc., San Diego, CA) or SAS 9.4. All comparisons were two-tailed, and the differences were considered significant when P-value < 0.05.
A total of 101 patients with a symptomatology of COVID-19 infection (Table 1) were included and divided in three groups based on the severity of their symptoms into mild (n = 41), moderate (n = 30) and severe cases (n = 30), as described in Methods. Fifty healthy donors were also recruited. As described previously (1, 3, 4), there was a significant difference in age, gender, BMI and number of comorbidities among the three groups of patients (p < 0.0001, p = 0.0002, p = 0.0003, and p = 0.0018 respectively). Most common symptoms of COVID-19 infection included cough, dyspnea and fever in 52, 52, and 42% of patients, respectively (Supplementary Table 1).
As expected, higher plasma levels of pro-inflammatory cytokines IL1β, IL6, IL8, and TNFα at admission and before specific treatment were positively correlated with the severity of COVID-19 symptoms (p < 0.0001 for all cytokines) (Table 2), confirming the results from previous studies (3, 16–18).
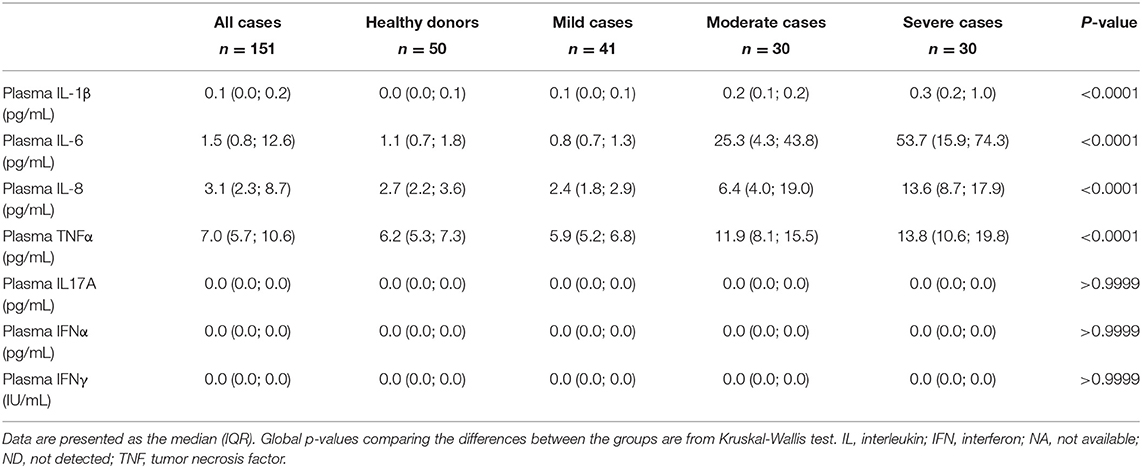
Table 2. Non-stimulated plasma cytokine levels of healthy donors and of patients with COVID-19, at baseline.
While the current state of inflammatory response to SARS-CoV-2 infection, as evidenced by the plasma cytokine levels, reflects the ongoing interplay between innate and adaptive immunity, it tells us little about immune function. To this end, we stimulated innate cells and T lymphocytes of COVID-19 patients at admission and before specific treatment with Toll-like receptor 7/8 (TLR 7/8) agonist and anti-CD3, respectively, and we measured the cytokines secreted. TLR7 is predominantly expressed in plasmacytoid dendritic cells (pDC) (44) and TLR8 is more strongly expressed in myeloid dendritic cells, monocytes and to a lesser extent in pDC (45). Th17 cytokine IL17A, as well as type I and type II IFN were not detectable in non-stimulated plasma of COVID-19 patients (Table 2). However, after in vitro stimulation of immune cells significant differences in cytokine levels emerged between patients with various severity of COVID-19 (Table 3, Figure 1), reflecting the fitness of their immune system. On the innate immunity side, DCs and NK cells of moderate and severe patients were functionally exhausted as illustrated by lower IFNα (and IFNγ from NK cells) levels upon in vitro stimulation (p < 0.0001) (Figure 1A), as previously suggested (46), and the differences remained significant after correction for monocyte count (p < 0.0001 mild vs. moderate and mild vs. severe) (Figure 1B). Levels of IL6, which is secreted by cells of both innate and adaptive immunity, remained unchanged between groups upon in vitro stimulation (p = 0.1247) (Figure 1C). On the adaptive immunity side, functional exhaustion was observed for Th17 lymphocytes producing IL17A in severe COVID-19 patients in comparison to mild forms and healthy subjects (p = 0.0004 and p = 0.002 respectively) (Figure 1D). Strikingly, lower secretion of IFNγ correlated with increased severity of COVID-19 (p < 0.0001) (Figure 1E). This lower production of IFNγ remained significant even when corrected for lymphocyte count (p = 0.0183 mild vs. moderate and p = 0.0009 mild vs. severe) (Figure 1F).
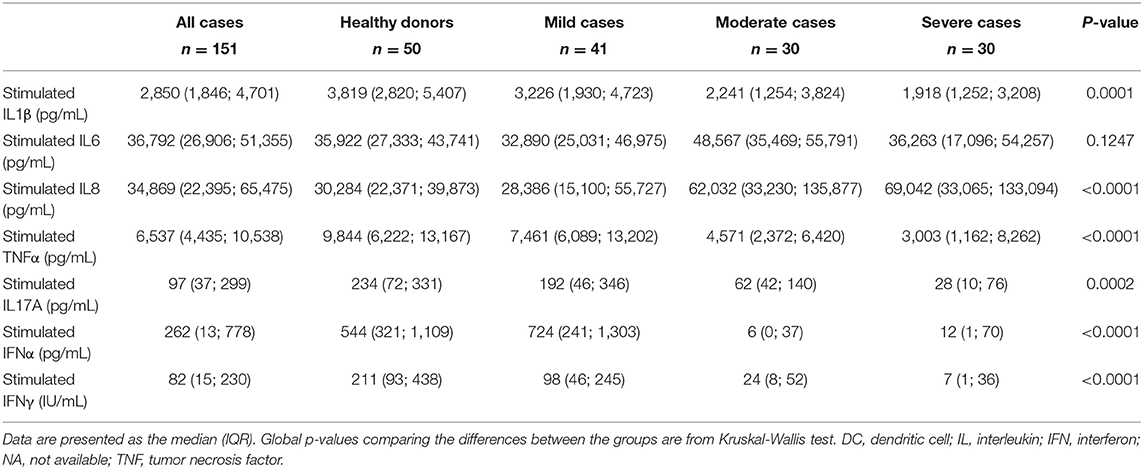
Table 3. Serum cytokine levels after non-specific stimulation of T lymphocytes and DCs, in healthy donors and in patients with COVID-19, at baseline.
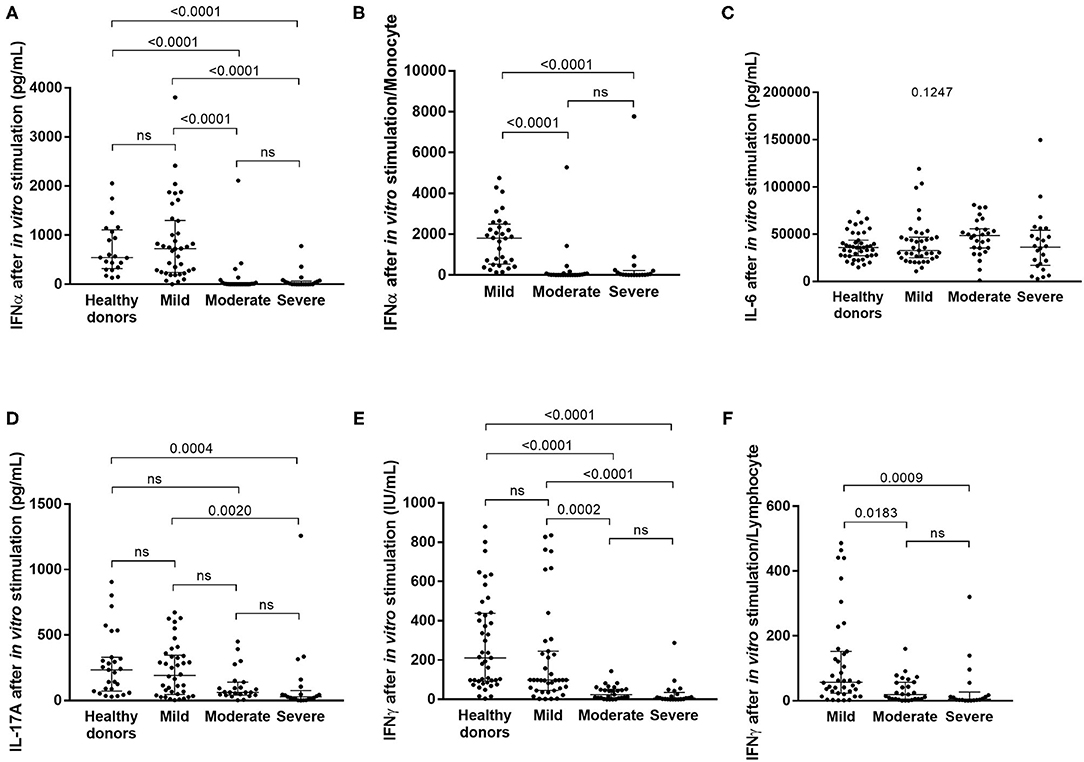
Figure 1. Serum cytokine levels after in vitro stimulation of innate and adaptive immune cells in healthy subjects and in COVID-19 patients with mild, moderate or severe symptoms. The levels of IFNα (A: in mild COVID-19 3 points are missing, in moderate COVID-19 3 points are missing and in severe COVID-19 patients 6 points are missing), IL6 (C: 3, 3, and 6 points, respectively), IL17 (D; 3, 7, and 9 points, respectively) and IFNγ (E: 2, 3 and 9 points, respectively) were measured, and the levels of IFNα and IFNγ were corrected for monocyte (B) and lymphocyte (F) count, respectively. Differences between groups were compared with Kruskal-Wallis test using Dunn's post hoc test.
The level of IFNα and IFNγ production upon in vitro stimulation of innate and adaptive immunity cells at admission and before specific treatment was predictive of the risk of complications (p = 0.003 and p < 0.0001, respectively) (Figures 2A,C). Indeed, patients with a level of IFNα and IFNγ lower than 2.1 pg/mL and 15 IU/mL, respectively, as defined by a ROC curve (data not shown), were more likely to develop complications during hospitalization (p = 0.0098 and p = 0.0002, respectively) (Figures 2B,D). As confirmed also by multivariable analysis (Table 4), stimulated IFNγ levels are an independent predictor of complications in patients with COVID-19 [p = 0.0349 OR = 0.98 (0.962; 0.999)].
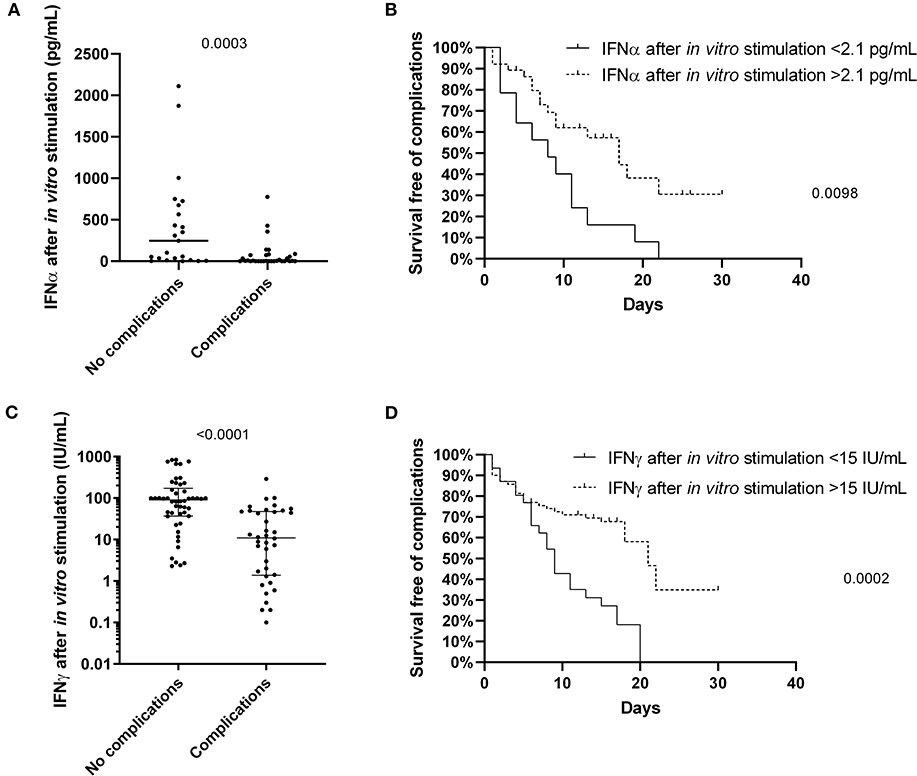
Figure 2. The predictive value of IFNα and IFNγ after non-specific stimulation of innate and adaptive immune cells for the patients with COVID-19. The level of IFNα (A; n = 55) and IFNγ (C; n = 89) differs between the patients with and without complications, and predicts the survival free of complications for patients with COVID-19 (B,D). Differences between groups were compared with Mann-Whitney test, and log-rank test was used to compare survival data.
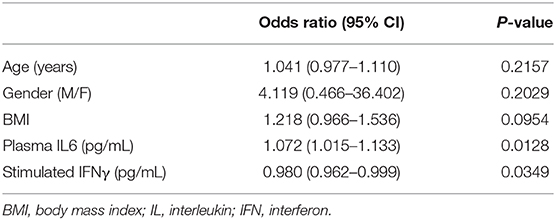
Table 4. Multivariable analysis for the evaluation of the relationship between considered variables at baseline and complications.
We further assessed the evolution of cytokine production and IFN response during hospitalization in moderate and severe cases. During follow-up, non-stimulated plasma IL6 levels decreased between the moment of admission to the hospital and at the last observation carried forward for patients with favorable outcome (p = 0.02148) (Figure 3A and Supplementary Figure 1A), while they remained high in deceased patients (p = 0.5625) (Figure 3B and Supplementary Figure 1B). The level of IFNγ after in vitro stimulation, however, did not significantly differ between the time of admission to the hospital and the last observed time point (Figures 3C,D), which was likely due to a small number of patients per group. Two individual cases were chosen to better demonstrate the evolution of cytokine production during the course of the disease. The first case resulted in recovery with an increased stimulated IFNγ levels at the last point (Supplementary Figure 1A), while the second case resulted in death with stable low stimulated IFNγ levels throughout hospitalization (Supplementary Figure 1B).
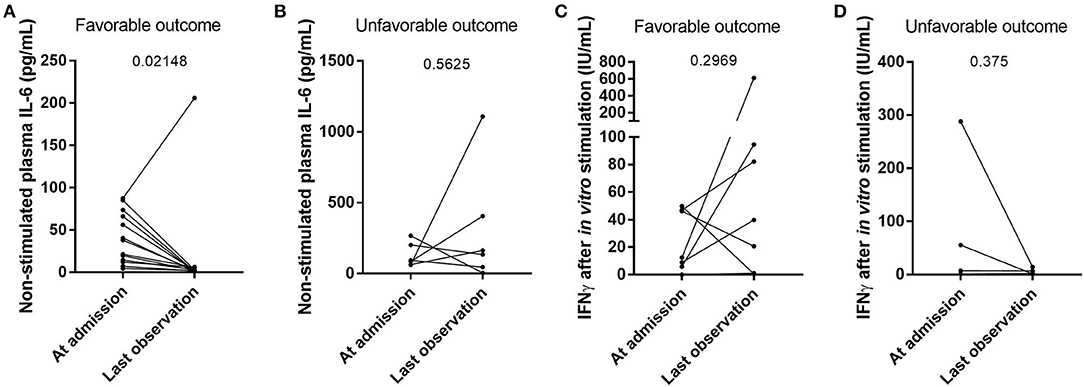
Figure 3. The evolution of cytokine levels in individual patients following their clinical outcome. The levels of IL6 in non-stimulated plasma (A,B; n = 13 and n = 6, respectively) and the levels of IFNγ after in vitro non-specific stimulation of innate and adaptive immune cells (C,D; n = 6 and n = 3, respectively) were compared between the patients who recovered from their SARS-CoV-2 infection (A,C) and the deceased patients (B,D). Differences between groups were compared with a Wilcoxon matched pairs signed rank test.
Several drugs commonly used to treat COVID-19 patients were tested for their potential to restore cytokine balance in vitro, notably to increase IFNγ production and decrease the production of inflammatory cytokines, while keeping the secretion of regulatory cytokines constant. Chloroquine and methylprednisolone proved efficient in reducing secretion of all cytokines (Figure 4) while Adalimumab reduced only IL6 and IL10 secretion. Interestingly, IFNα had a more balanced effect with a strong stimulation of IFNγ and a decrease of inflammatory cytokine IL1β, while the secretion of T regulatory cytokine IL10 and pro-inflammatory cytokine (IL6) remained unchanged. The individual results are detailed in Supplementary Table 3.
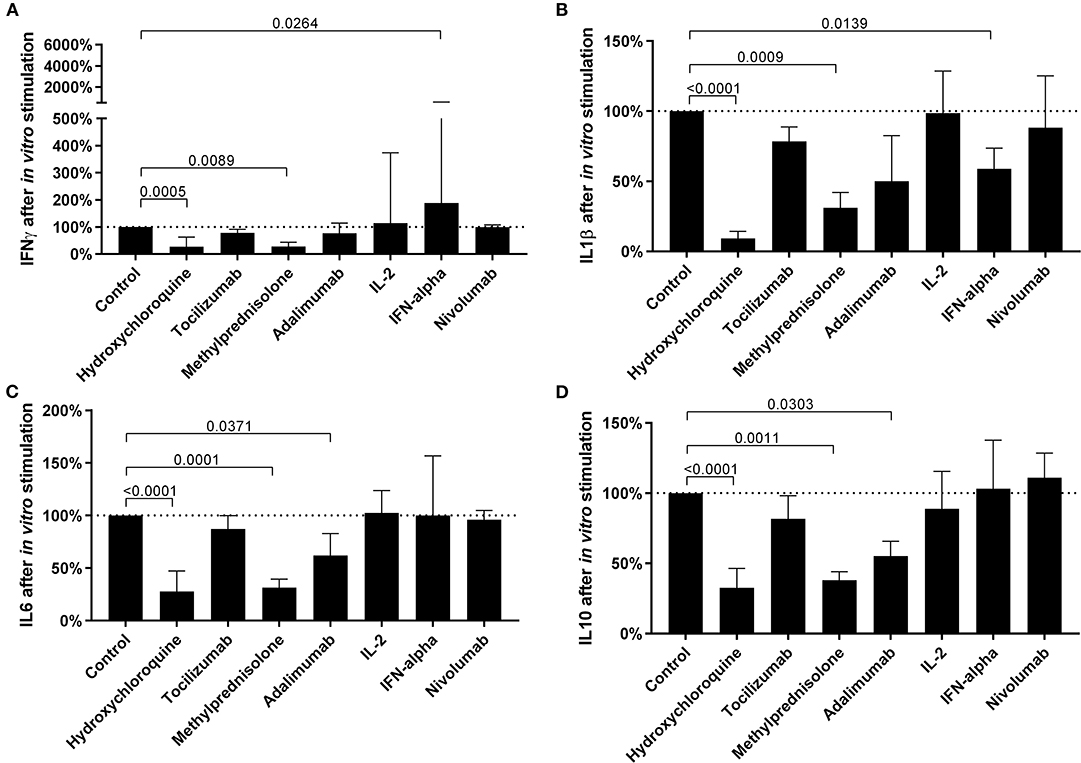
Figure 4. The efficacy of in vitro treatment with different drugs commonly used in COVID-19 to modulate cytokine expression. The levels of IFNγ (A), IL1β (B), IL6 (C), and IL10 (D) after in vitro pretreatment with drugs, followed by non-specific stimulation of innate and adaptive immune cells, in 18 COVID-19 patients. Differences between groups were compared with Kruskal-Wallis test using Dunn's post hoc test.
We report here a cohort of 101 patients with a symptomatology of COVID-19 infection. We aimed to reveal their specific immunological profiles and to correlate them to the evolution and the extent of symptoms in individual patients. Our results confirmed a functional exhaustion of type I (NK cells and DCs) and type II IFN (T cells) production in moderate and severe patients traducing an evasion of both innate and adaptive immune response and in accordance with recent studies (14, 16, 18, 21, 47–49).
It is well-known that the innate immune response is triggered by virally infected cells which can be recognized by host pattern-recognition receptors (PRRs) expressed by DCs that produce a variety of cytokines (50) such as type I IFN, which in turn recruit lymphocytes and monocytes to inflamed sites (51–54). Type I IFN primarily activates epithelial cells and reduces the mononuclear macrophage-mediated proinflammatory activity (55). Type II IFNs have different functions, eliciting T helper 1 (Th1)-driven immune responses, and also enabling induced regulatory T (Treg) cells to control and regulate immune responses (56). Consequently, SARS-CoV-2 has evolved several mechanisms to inhibit type I IFN induction and signaling (57). During SARS-CoV-2 infection, both innate and adaptive immune response are required for successful virus clearance and must be adequately controlled to minimize immunopathological damage (57). By assessing the response of immune cells of infected patients after stimulation, we demonstrate here a marked decrease in type I and type II IFN response from mild to severe patients. The molecular mechanism(s) of this IFN evasion remain to be confirmed, however, several studies have suggested different pathways that could contribute to the decreased amount of IFNs in severe COVID-19 patients, from concealed viral production invisible to PPARs to direct synthesis of structural and nonstructural viral proteins that antagonize IFN signaling (47–49). Indeed, SARS-CoV-2 induced an aberrant type-I IFN response in cultured cells, characterized by a delayed antiviral response which may provide a window for virus replication and an improper recruitment of inflammatory monocyte macrophage populations (21).
The originality of our work lies in the stimulation of TLR7 and TLR8 which reproduce in vitro a viral infection by the activation of innate immune system and produce type I IFN (58). On the other hand, the stimulation of T lymphocytes by an anti-CD3 allowed us to quantify the production of type II IFN and to evaluate the adaptive immune response. The innate immune recognition of virus infection triggers antiviral immune responses by residual genomic RNA recognized by PRR expressed mainly by DCs (59). In moderate and severe COVID-19 patients we observed that innate cells produce less type I IFN, and consequently NK cells produce less type II IFN. In accordance with previous studies (21, 60), our results suggest that an uncontrolled infection maintains monocyte and macrophagic activation, and that the regulatory T lymphocytes remain inactivated due to a weak production of type II IFN, thus reinforcing the cytokine storm and leading to severe complications in patients with COVID-19.
This longitudinal study allowed us to conclude that a functional analysis of IFN production at the beginning of the hospitalization is a powerful tool to predict the clinical evolution of patients infected with SARS-CoV-2. While previous studies demonstrated opposing results with either impaired (14, 28) or increased (61) type I IFN response in severe COVID-19 patients, our results tip the balance toward impaired IFN signaling. According to our study on stimulated IFN production as well as in other studies (14, 21), type I IFN plays a major role in the activation of type II IFN and represents a strategic target for early treatment of COVID-19 patients, in order to destroy the immune evasion caused by SARS-CoV-2 and to treat the specific immune dysfunction.
However, there is increasing evidence that patients with severe COVID-19 may have a robust type I IFN response, which contrasts the delayed, possibly suppressed, IFN response seen early in infection (27, 62). While limited by a small sample size of eight and seven patients, respectively, Zhou et al. and Wilk et al. demonstrated that many IFN-stimulated genes are overexpressed in COVID-19 patients (62, 63).
Since more studies are needed to further illuminate the role of IFNs in COVID-19, both from a clinical and a molecular perspective, IFN treatment remains controversial as well. Nevertheless, an in vitro study on cultured cells has shown a potential benefit of IFNβ treatment (48), as well as a recent clinical study NCT04276688 with favorable outcome for IFNβ (33), while others are still in progress. Two recent retrospective studies found that IFNα treatment may be beneficial for COVID-19 patients (37, 38), however it seems that adequate timing in IFN administrating is crucial for its efficacy since early administration decreased mortality, while late administration had an opposite effect (38).
Our study presents several limitations. First, this is an observational study showing an association between IFN I and II levels and COVID-19 severity and outcome. Randomized clinical trials using functional interferon assays at admission to predict outcome are needed to clearly evaluate the efficacy and utility of IFN measurement in clinical practice. Second, while our in vitro tests and several recently published studies (33, 37, 38, 48) show the potential of IFNα treatment in order to restore the cytokine balance, the results nevertheless need to be confirmed in large controlled clinical trials. Third, this is a study on a relatively small number of patients that needs to be confirmed in larger cohorts. Notably, there were only six deaths in our cohort of COVID-19 patients, severely limiting the power of statistical analyses. Fourth, while male gender, older age and obesity have been shown to be strongly associated with increased mortality in COVID-19 patients (10, 11), low number of deaths prevented us from identifying these factors in our cohort. Instead, we tested the predictive power of age, gender and BMI on COVID-19-related complications, but apart from plasma IL6 levels and stimulated IFNγ levels the other variables remained non-significant at multivariable analysis.
The variability of symptomatology lies at the heart of our cells, among the immune responses involved in fighting infection by the COVID-19. IFNγ represents a predictive biomarker of the evolution of SARS-CoV-2 which can be safely and routinely measured in laboratory by QuantiFERON Monitor. It could allow clinicians to provide adjusted treatment and medical care in this epidemic context. Based on our results, our functional test could be an important tool to predict severe COVID-19 and guide personalized therapy targeting the immune restoration of NK and T-cells (inhibiting check-point inhibitor) and IFN production.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by CPP SUD-OUEST et OUTRE-MER I. The patients/participants provided their written informed consent to participate in this study.
CR, VB, MC, SB, and BS-P designed the study. VB and MC carried out experiments. KZ, CF, CR, and MC collected clinical data. BS-P, CR, VB, and MC analyzed and interpreted the data. BS-P, CR, ED, KR, JC, EC, CI, JD, and TP provided medical oversight. BS-P, CR, and VB drafted and revised the manuscript. All authors contributed to the article and approved the submitted version.
This research was supported by a grant from Conseil Départemental des Alpes-Maritimes and from the Agence Nationale pour la Recherche (ANR).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all patients involved in this study, as well as Elodie Mallet, Géraldine Dalmasso, and Natalia Chauzenoux for their help within the laboratory.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2020.603961/full#supplementary-material
1. Li Q, Guan X, Wu P, Wang X, Zhou L, Tong Y, et al. Early transmission dynamics in Wuhan, China, of novel coronavirus-infected pneumonia. N Engl J Med. (2020) 382:1199–207. doi: 10.1056/NEJMoa2001316
2. Zhou P, Yang X Lou, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. (2020) 579:270–3. doi: 10.1038/s41586-020-2951-z
3. Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. (2020) 395:497–506. doi: 10.1016/S0140-6736(20)30183-5
4. Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, He JX, et al. Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med. (2020) 382:1708–20. doi: 10.1056/NEJMoa2002032
5. Jin YH, Cai L, Cheng ZS, Cheng H, Deng T, Fan YP, et al. A rapid advice guideline for the diagnosis and treatment of 2019 novel coronavirus (2019-nCoV) infected pneumonia (standard version). Mil Med Res. (2020) 7:1–23. doi: 10.1186/s40779-020-0233-6
6. Wu Z, McGoogan JM. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: summary of a report of 72314 cases from the Chinese center for disease control and prevention. J Am Med Assoc. (2020) 323:1239–42. doi: 10.1001/jama.2020.2648
7. Zaim S, Chong JH, Sankaranarayanany V, Harky A. COVID-19 and multiorgan response. J Am Med Assoc. (2020) 323:1239. doi: 10.1016/j.cpcardiol.2020.100618
8. Hanley B, Naresh KN, Roufosse C, Nicholson AG, Weir J, Cooke GS, et al. Histopathological findings and viral tropism in UK patients with severe fatal COVID-19: a post-mortem study. Lancet Microbe. (2020) 1:e245–53. doi: 10.1016/S2666-5247(20)30115-4
9. Liu K, Fang YY, Deng Y, Liu W, Wang MF, Ma JP, et al. Clinical characteristics of novel coronavirus cases in tertiary hospitals in Hubei Province. Chin Med J. (2020) 133:1025–31. doi: 10.1097/CM9.0000000000000744
10. Gong J, Ou J, Qiu X, Jie Y, Chen Y, Yuan L, et al. A tool to early predict severe corona virus disease 2019 (COVID-19): a multicenter study using the risk nomogram in Wuhan and Guangdong, China. Clin Infect Dis. (2020) 71:833–40. doi: 10.1093/cid/ciaa443
11. Chen X, Liu Z. Early prediction of mortality risk among severe COVID-19 patients using machine learning. Int J Epidemiol. (2020) 42:dyaa171. doi: 10.1093/ije/dyaa171
12. Carvelli J, Demaria O, Vely F, Batista L, Benmansour NC, Fares J, et al. Association of COVID-19 inflammation with activation of the C5a–C5aR1 axis. Nature. (2020) 588:146–50. doi: 10.1038/s41586-020-2600-6
13. Vabret N, Britton GJ, Gruber C, Hegde S, Kim J, Kuksin M, et al. Immunology of COVID-19: current state of the science. Immunity. (2020) 52:910–41.
14. Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, Pere H, et al. Impaired type I interferon activity and exacerbated inflammatory responses in severe Covid-19 patients. Science (80-). (2020) 369:718–24. doi: 10.1126/science.abc6027
15. Fei J, Fu L, Li Y, Xiang H-X, Xiang Y, Li M-D, et al. Reduction of lymphocyte count at early stage elevates severity and death risk of COVID-19 patients: a hospital-based case-cohort study. MedRxiv. (2020). doi: 10.1101/2020.04.02.20050955
16. Moore JB, June CH. Cytokine release syndrome in severe COVID-19. Science (80-). (2020) 368:473 LP−4. doi: 10.1126/science.abb8925
17. Johannessen LE, Pedersen NM, Pedersen KW, Madshus IH, Stang E. Activation of the epidermal growth factor (EGF) receptor induces formation of EGF receptor- and Grb2-containing clathrin-coated pits. Mol Cell Biol. (2006) 26:389–401. doi: 10.1128/MCB.26.2.389-401.2006
18. Pedersen SF, Ho YC. SARS-CoV-2: a storm is raging. J Clin Invest. (2020) 130:2202–5. doi: 10.1172/JCI137647
19. Ragab D, Salah Eldin H, Taeimah M, Khattab R, Salem R. The COVID-19 cytokine storm: what we know so far. Front Immunol. (2020) 11:1–4. doi: 10.3389/fimmu.2020.01446
20. Chen G, Wu D, Guo W, Cao Y, Huang D, Wang H, et al. Clinical and immunologic features in severe and moderate Coronavirus Disease 2019. J Clin Invest. (2020) 130:2620–9. doi: 10.1101/2020.02.16.20023903
21. Blanco-Melo D, Nilsson-Payant BE, Liu W-C, Uhl S, Hoagland D, Møller R, et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell. (2020) 181:1036–45. doi: 10.1016/j.cell.2020.04.026
22. Cao X. COVID-19: immunopathology and its implications for therapy. Nat Rev Immunol. (2020) 20:269–70. doi: 10.1038/s41577-020-0308-3
23. Du SQ, Yuan W. Mathematical modeling of interaction between innate and adaptive immune responses in COVID-19 and implications for viral pathogenesis. J Med Virol. (2020) 92:1615–28. doi: 10.1002/jmv.25866
24. Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol. (2008) 9:503–10. doi: 10.1038/ni1582
25. Bonjardim CA. Interferons (IFNs) are key cytokines in both innate and adaptive antiviral immune responses - and viruses counteract IFN action. Microbes Infect. (2005) 7:569–78. doi: 10.1016/j.micinf.2005.02.001
26. Tough DF. Type I interferon as a link between innate and adaptive immunity through dendritic cell stimulation. Leuk Lymphoma. (2004) 45:257–64. doi: 10.1080/1042819031000149368
27. Lee JS, Shin E-C. The type I interferon response in COVID-19: implications for treatment. Nat Rev Immunol. (2020) 20:19–20. doi: 10.1038/s41577-020-00429-3
28. Meffre E, Iwasaki A. Interferon deficiency can lead to severe COVID. Nat. (2020) 587:374–6. doi: 10.1038/d41586-020-03070-1
29. Boyer-Suavet S, Cremoni M, Dupeyrat T, Zorzi K, Brglez V, Benzaken S, et al. Functional immune assay using interferon-gamma could predict infectious events in end-stage kidney disease. Clin Chim Acta. (2019) 502:287–92. doi: 10.1016/j.cca.2019.11.018
30. Sood S. Immune monitoring post liver transplant. World J Transplant. (2014) 4:30. doi: 10.5500/wjt.v4.i1.30
31. Hornuss D, Laubner K, Monasterio C, Thimme R, Wagner D. [COVID-19 associated pneumonia despite repeatedly negative PCR-analysis from oropharyngeal swabs]. Dtsch Med Wochenschr. (2020) 145:844–9. doi: 10.1055/a-1170-6061
32. Bouaziz JD, Duong T, Jachiet M, Velter C, Lestang P, Cassius C, et al. Vascular skin symptoms in COVID-19: a french observational study. J Eur Acad Dermatol Venereol. (2020) 34:e451–2. doi: 10.1111/jdv.16544
33. Hung IFN, Lung KC, Tso EYK, Liu R, Chung TWH, Chu MY, et al. Triple combination of interferon beta-1b, lopinavir–ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID-19: an open-label, randomised, phase 2 trial. Lancet. (2020) 395:1695–704. doi: 10.1016/S0140-6736(20)31042-4
34. Vijayvargiya P, Esquer Garrigos Z, Castillo Almeida NE, Gurram PR, Stevens RW, Razonable RR. Treatment considerations for COVID-19: a critical review of the evidence (or lack thereof). Mayo Clin Proc. (2020) 95:1454–66. doi: 10.1016/j.mayocp.2020.04.027
35. The RECOVERY Collaborative Group. Dexamethasone in hospitalized Patients with Covid-19 — preliminary report. N Engl J Med. (2020). [Epub ahead of print].
36. The RECOVERY Collaborative Group. Effect of hydroxychloroquine in hospitalized patients with Covid-19. N Engl J Med. (2020) 383:2030–40. doi: 10.1056/NEJMoa2022926
37. Zhou Q, Chen V, Shannon CP, Wei XS, Xiang X, Wang X, et al. Interferon-α2b treatment for COVID-19. Front Immunol. (2020) 11:1–6. doi: 10.3389/fimmu.2020.615275
38. Wang N, Zhan Y, Zhu L, Hou Z, Liu F, Song P, et al. Retrospective multicenter cohort study shows early interferon therapy is associated with favorable clinical responses in COVID-19 patients. Cell Host Microbe. (2020) 28:455–64.e2. doi: 10.1016/j.chom.2020.07.005
39. Wiersinga WJ, Rhodes A, Cheng AC, Peacock SJ, Prescott HC. Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): a review. JAMA - J Am Med Assoc. (2020) 324:782–93. doi: 10.1001/jama.2020.12839
40. Rizk JG, Kalantar-Zadeh K, Mehra MR, Lavie CJ, Rizk Y, Forthal DN. Pharmaco-immunomodulatory therapy in COVID-19. Drugs. (2020) 80:1267–92. doi: 10.1007/s40265-020-01367-z
41. Szabados B, Abu-Ghanem Y, Grant M, Choy J, Bex A, Powles T. Clinical characteristics and outcome for four SARS-CoV-2-infected cancer patients treated with immune checkpoint inhibitors. Eur Urol. (2020) 78:276–80. doi: 10.1016/j.eururo.2020.05.024
42. Gomes F, Serra-Bellver P, Lorigan P. The role of nivolumab in melanoma. Futur Oncol. (2018) 14:1241–52. doi: 10.2217/fon-2017-0484
43. Saadoun D, Rosenzwajg M, Joly F, Six A, Carrat F, Thibault V, et al. Regulatory T-cell responses to low-dose interleukin-2 in HCV-induced vasculitis. N Engl J Med. (2011) 365:2067–77. doi: 10.1056/NEJMoa1105143
44. Patinote C, Karroum NB, Moarbess G, Cirnat N, Kassab I, Bonnet PA, et al. Agonist and antagonist ligands of toll-like receptors 7 and 8: ingenious tools for therapeutic purposes. Eur J Med Chem. (2020) 193:112238. doi: 10.1016/j.ejmech.2020.112238
45. Crozat K, Vivier E, Dalod M. Crosstalk between components of the innate immune system: promoting anti-microbial defenses and avoiding immunopathologies. Immunol Rev. (2009) 227:129–49. doi: 10.1111/j.1600-065X.2008.00736.x
46. Zheng M, Gao Y, Wang G, Song G, Liu S, Sun D, et al. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell Mol Immunol. (2020) 17:533–5. doi: 10.1038/s41423-020-0402-2
47. Xia H, Cao Z, Xie X, Zhang X, Chen JYC, Wang H, et al. Evasion of type I interferon by SARS-CoV-2. Cell Rep. (2020) 33:108234. doi: 10.1016/j.celrep.2020.108234
48. Lei X, Dong X, Ma R, Wang W, Xiao X, Tian Z, et al. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat Commun. (2020) 11:1–12. doi: 10.1038/s41467-020-17665-9
49. Acharya D, Liu GQ, Gack MU. Dysregulation of type I interferon responses in COVID-19. Nat Rev Immunol. (2020) 20:397–8. doi: 10.1038/s41577-020-0346-x
50. Kawai T, Akira S. Innate immune recognition of viral infection. Nat Immunol. (2006) 7:131–7. doi: 10.1038/ni1303
51. Akira S, Hemmi H. Recognition of pathogen-associated molecular patterns by TLR family. Immunol Lett. (2003) 85:85–95. doi: 10.1016/S0165-2478(02)00228-6
52. Ito T, Wang YH, Liu YJ. Plasmacytoid dendritic cell precursors/type I interferon-producing cells sense viral infection by Toll-like receptor (TLR) 7 and TLR9. Springer Semin Immunopathol. (2005) 26:221–9. doi: 10.1007/s00281-004-0180-4
53. Theofilopoulos AN, Baccala R, Beutler B, Kono DH. Type I interferons (A/B) in immunity and autoimmunity. Annu Rev Immunol. (2005) 23:307–35. doi: 10.1146/annurev.immunol.23.021704.115843
54. Faure E, Poissy J, Goffard A, Fournier C, Kipnis E, Titecat M, et al. Distinct immune response in two MERS-CoV-infected patients: Can we go from bench to bedside? PLoS ONE. (2014) 9:e88716. doi: 10.1371/journal.pone.0088716
55. Davidson S, McCabe TM, Crotta S, Gad HH, Hessel EM, Beinke S, et al. IFN λ is a potent anti-influenza therapeutic without the inflammatory side effects of IFN α treatment. EMBO Mol Med. (2016) 8:1099–112. doi: 10.15252/emmm.201606413
56. Wood KJ, Sawitzki B. Interferon γ: a crucial role in the function of induced regulatory T cells in vivo. Trends Immunol. (2006) 27:183–7. doi: 10.1016/j.it.2006.02.008
57. Lagunas-Rangel FA, Chávez-Valencia V. High IL-6/IFN-γ ratio could be associated with severe disease in COVID-19 patients. J Med Virol. (2020) 92:1789–90. doi: 10.1002/jmv.25900
58. Sood S, Cundall D, Yu L, Miyamasu M, Boyle JS, Ong SY, et al. A novel biomarker of immune function and initial experience in a transplant population. Transplantation. (2014) 97:50–1. doi: 10.1097/TP.0000000000000078
59. Tatematsu M, Funami K, Seya T, Matsumoto M. Extracellular RNA sensing by pattern recognition receptors. J Innate Immun. (2018) 10(5–6):398–406. doi: 10.1159/000494034
60. Moreira-Teixeira L, Mayer-Barber K, Sher A, O'Garra A. Type I interferons in tuberculosis: foe and occasionally friend. J Exp Med. (2018) 215:1273–85. doi: 10.1084/jem.20180325
61. Lucas C, Wong P, Klein J, Castro TBR, Silva J, Sundaram M, et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature. (2020) 584:463–9. doi: 10.1038/s41586-020-2588-y
62. Zhou Z, Ren L, Zhang L, Zhong J, Xiao Y, Jia Z, et al. Heightened innate immune responses in the respiratory tract of COVID-19 patients. Cell Host Microbe. (2020) 27:883–90.e2. doi: 10.1016/j.chom.2020.04.017
Keywords: immunology, infectious diseases, COVID-19, interferon, personalized medicine
Citation: Ruetsch C, Brglez V, Crémoni M, Zorzi K, Fernandez C, Boyer-Suavet S, Benzaken S, Demonchy E, Risso K, Courjon J, Cua E, Ichai C, Dellamonica J, Passeron T and Seitz-Polski B (2021) Functional Exhaustion of Type I and II Interferons Production in Severe COVID-19 Patients. Front. Med. 7:603961. doi: 10.3389/fmed.2020.603961
Received: 08 September 2020; Accepted: 18 December 2020;
Published: 27 January 2021.
Edited by:
Zisis Kozlakidis, International Agency For Research On Cancer (IARC), FranceReviewed by:
Renee W. Y. Chan, The Chinese University of Hong Kong, ChinaCopyright © 2021 Ruetsch, Brglez, Crémoni, Zorzi, Fernandez, Boyer-Suavet, Benzaken, Demonchy, Risso, Courjon, Cua, Ichai, Dellamonica, Passeron and Seitz-Polski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Barbara Seitz-Polski, c2VpdHotcG9sc2tpLmJAY2h1LW5pY2UuZnI=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.