 Catarina Sousa Guerreiro
Catarina Sousa Guerreiro Ângelo Calado
Ângelo Calado Joana Sousa
Joana Sousa João Eurico Fonseca
João Eurico Fonseca
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Med., 14 December 2018
Sec. Rheumatology
Volume 5 - 2018 | https://doi.org/10.3389/fmed.2018.00349
This article is part of the Research TopicNutrition and Metabolism in Rheumatic DiseasesView all 6 articles
Growing experimental and clinical evidence suggests that a chronic inflammatory response induced by gut dysbiosis can critically contribute to the development of rheumatic diseases, including rheumatoid arthritis (RA). Of interest, an adherence to a Mediterranean diet has been linked to a reduction in mortality and morbidity in patients with inflammatory diseases. Diet and intestinal microbiota are modifying factors that may influence intestinal barrier strength, functional integrity, and permeability regulation. Intestinal microbiota may play a crucial role in RA pathogenesis, but up to now no solid data has clarified a mechanistic relationship between gut microbiota and the development of RA. Nonetheless, microbiota composition in subjects with RA differs from that of controls and this altered microbiome can be partially restored after prescribing disease modifying antirheumatic drugs. High levels of Prevotella copri and similar species are correlated with low levels of microbiota previously associated with immune regulating properties. In addition, some nutrients can alter intestinal permeability and thereby influence the immune response without a known impact on the microbiota. However, critical questions remain to be elucidated, such as the way microbiome fluctuates in relation to diet, and how disease activity may be influenced by changes in diet, microbiota or diet-intestinal microbiota equilibrium.
A growing body of experimental and clinical evidence suggests that a chronic inflammatory response induced by gut dysbiosis can critically contribute to the development of a number of rheumatic diseases, including RA (1–3). Of interest to this discussion, some RA patients suffer from clinical or subclinical gut disturbances (4). It has been hypothesized that at some point of the preclinical phase of RA, at the level of the mucosal surface, interactions between microbes, and other potential environmental factors (e.g., diet, physical, and emotional stress) as well as host factors lead to mucosal inflammation and to the breaking of immune tolerance. This mucosal inflammation may enhance local—and then systemic—immune disturbances, through mechanisms that may include molecular mimicry or facilitation of direct autoimmunity to self-antigens (5, 6). Breaching this single layer of epithelium can lead to pathological exposure of the highly immunoreactive subepithelium to foreign antigens in the lumen (4). The equilibrium between tolerance and immunity to non-self-antigens can be ruptured by this increased intestinal permeability, which in many cases may facilitate absorption of antigens and contribute to the persistence and exacerbation of some immune mediated diseases, including RA (7).
Two major altering factors that may influence barrier strength and functional integrity, with an effect in intestinal permeability regulation, are diet and intestinal microbiota. These factors may allow the entry of external antigens from the gut lumen into the host (6, 8, 9). Both are life style related, which suggests that environmental factors might influence the function of the intestinal barrier and thus, influence immune health and the onset and activity of diseases such as RA (9).
The intestinal tract harbors the largest bacterial community associated with the human body. Everyone carries up to a few hundred species of intestinal bacteria (9). Over 90% of intestinal bacteria belong to the Bacteroidetes and Firmicutes phyla, but other phyla like Proteobacteria, Actinobacteria, Fusobacteria, Verrucomicrobia, and Cyanobacteria also play a crucial role in the maintenance and regulation of homeostasis in intestinal microflora (3, 7). Among other functions, the intestinal microbiota is a critical factor for the homeostasis of the host immune system and as such, any alteration on the gut microbiota may impact on the host immune response (10, 11).
Human studies revealed that patients with RA display significant differences of the intestinal microbiota and a decreased gut microbial diversity in comparison to healthy controls (3), both related with disease duration and autoantibody levels (3, 12, 13). Patients with RA, particularly erosive patients, carry a distinctive enterotype of gut microbiota characterized by a lower abundance of bacteria belonging to the family Bifidobacterium and Bacteroides (12, 14, 15) and, at least at early stages of the disease, an abundance of Prevotella copri (14, 16).
The association between an imbalance of the intestinal microbiota and RA has been suggested to happen due to different mechanisms that may impact the host immune system and its function, including:
(i) the activation of antigen-presenting cells (APCs), such as the dendritic cells, which can impact the cytokine production and antigen presentation. Such disturbances can further modulate the host immune response by, for instance, impinging on T cells differentiation and function. These microbiota-induced effects can be mediated through pathogen recognition receptors, key innate immune receptors, capable of perceiving pathogen-associated molecular patterns, such as the toll-like receptors (TLRs) (10);
(ii) the ability to promote the citrullination of peptides via the enzymatic action of peptidyl-arginine deiminases (PADs). Concerning this, it is of interest that the intestinal epithelium is a major producer of citrulline in the human body (4). Furthermore, PAD is active in the human intestine and the intestinal microbiome may also encode active microbial PADs (4). Thus, the intestine may stand as a source of citrullinated peptides, along with other mucosal surfaces. In this respect, peptide citrullination by the bacterial PAD enzyme expressed by P. gingivalis has been suggested to strongly contribute for the close association between periodontitis, an inflammatory disease of the oral mucosa, with an increased susceptibility to RA (10, 17);
(iii) antigenic mimicry, which can result from similarities existing between foreign antigens and self-antigens, that then evoke the activation of pathogen-derived autoreactive T and B cells and thus lead to autoimmunity (10);
(iv) impact on the permeability of the intestinal mucosal, by modulating the expression of tight junction (TJ) proteins;
(v) and control of the host immune system. This effect can be exerted, for instance, by modulating T cells differentiation and unbalancing the homeostasis between T helper type 17 (Th17) cells and T regulatory (Treg) cells. In RA mice models, it has been shown that specific alterations in the intestinal microbiota may favor the pathophysiological action of Th17 cells in detriment of the suppressive action of Treg cells, consequently, promoting Th17-mediated mucosal inflammation (10).
Overall, the elicited immune responses may be dependent on the presence of certain genera/species. Bacteroides fragilis, for example, can stimulate Th1-mediated immune responses in the initial stages of colonization by producing polysaccharide A. Collinsella sp and may contribute to RA pathogenesis by increasing gut permeability, lowering the expression of TJ proteins and influencing the epithelial production of IL-17A (10, 12). Butyrate producing microbes, such as Clostridia, Faecalibacteria, and some species of Lachnospiraceae may also play a crucial role in keeping the integrity of intestinal epithelia, having a documented anti-inflammatory effect in the context of rheumatic diseases, including RA (3, 14).
In addition, the idea that the onset of autoimmunity may be related to gastrointestinal tract is supported not only by the fact that microbiota composition in subjects with RA differs from controls, but also by the observation that altered microbiome can be partially restored after prescribing disease modifying antirheumatic drugs (3).
Adding an additional level of complexity, it is becoming increasingly clear that the close interplay between variations in microbiota composition and RA may not be so simple. As a paradigmatic example, several conflicting associations have been observed regarding Prevotella species and RA pathogenesis. In fact, although some studies in RA patients display an association of Prevotella species as a contributive risk factor to the onset of the disease, contrarily, others have suggested that it may be protective (10). The existing data sustain that the gut microbiota of established RA patients exhibits a lower abundance of Prevotella species (15). However, Scher et al. (18) reported that individuals with early RA were more likely to harbor Prevotella copri (PC) compared to controls. Also, Maeda et al. (19) observed PC in abundance within gut microbiota in Japanese patients with early RA. According to these authors this species has a relevant impact on the inflammatory response, by inducing Th17 related cytokines, such as IL-6 and IL-23, and an increase on the intestinal permeability as well, with influence on bacteria penetration throughout the body. In addition, high levels of PC and similar species are correlated with low levels of protective microbiota, which are believed to regulate the immune system (20). An abundance of PC in early RA stages may very likely stand as an important mechanism that links dysbiosis with arthritis pathogenesis (10).
Marietta et al. (16) also focused their attention at the Prevotella species. In an experimental study they showed that Prevotella histicola can lead to a lower gut permeability by increasing the expression of enzymes required to produce antimicrobial peptides, as well as, of TJ proteins (zonula occludens 1 and occludin). These authors suggest this species to have immunomodulating properties, by generating Treg cells and increasing the transcription of interleukin-10 (16).
It is worth noticing that more than 40 different Prevotella species exist, with a vast and different array of genome repertoires among strains and between Prevotella species and hosts. This genetic high diversity found within Prevotella species might explain the highly different behavior observed when considering the genus-level identification (18), which possibly explain part of inconsistences observed when relationship between diet and health/disease is tested (10, 21). It's still missing, however, a question that so far any study has placed, which is of whether of the same genus are consistently linked to dietary patterns or equally responsive to diet variations and at a more complex level, in what manner such relationship may influence the disease behavior (22).
Discussion about possible mechanisms by which diet influence RA, its effect on intestinal microbiota and gut permeability are also a current hot topic. Literature has been showing that individual diet content may have an important impact on microbiota and metabolome expression, thereby influencing intestinal integrity and permeability (11, 23, 24).
Clustering of the human gut microbiota, designated enterotypes, was first described in 2011 (25, 26). One of the largest bacterial groups that can be found in the gut are the Bacteroidetes. Amongst this phylum, there are a lot of genera which are diet-responsive. Particularly, the increase in Prevotella and Bacteroidetes can be associated to a high-fiber diet and to the consumption of a diet rich in both fat and animal protein, respectively. (8, 19).
The Bacteroides-driven enterotype is reported to be predominant in individuals consuming more animal protein and saturated fats (western diet), becoming clearly that some kind of nutrients/food may enhance this type of enterotype. In contrast, the Prevotella-driven enterotype is poorly represented in these individuals but present at high values in individuals consuming carbohydrates, simple sugars and fiber, suggesting an association with a carbohydrate-based diet (8, 27).
The food patterns of the populations who lived around the Mediterranean sea during the 60's, inspire what is, nowadays, commonly known as The Mediterranean Diet (MD), which is, proven, one of the healthiest dietary pattern existing (28–30). The essence of this diet consists in the high consumption of products like fruits, vegetables, legumes, unrefined cereals and nuts, the moderate consumption of fish, poultry and dairy products as cheese and yogurt, and the low consumption of red meat products. Olive oil is used as the main edible-fat source and wine is consumed in a regular, but moderate, basis (29, 30). The reduction in overall mortality and morbidity has been linked to a greater adherence to the MD and this food pattern is highly proposed as a beneficial dietary approach for patients with inflammatory diseases (30, 31).
According to some authors, important RA disease characteristics namely, inflammatory activity disease, physical function and vitality, may be clinically related to MD pattern, particularly when compared to a western diet (10, 32). No association has been proved regarding the susceptibility to disease, nonetheless. On the Skoldstam's study (33), after 12 weeks of intervention, patients in the MD group showed a significant improvement in DAS-28 (Disease Activity Score-−28 joints), HAQ (Health Assessment Questionnaire), whereas subjects in the control group displayed no significant changes in these parameters. In another study, developed by Mckellar et al. (34) and Tedeschi and Costenbader (35), after 6 months, patient global pain VAS (Visual Analog Scale), and morning stiffness were significantly improved in the intervention arm when compared to the control arm (35). It is important to note that RA disease activity was significantly decrease in patients with adequate level of typical MD antioxidant as vitamin C, retinol, and uric acid (32).
An association between MD and short chain fatty acids (SCFAs) production, is well known and theoretically plausible. Rich fiber foods such as fruit, vegetables, and legumes, all regularly consumed by individuals who practice a MD pattern, have the ability to become degraded by Firmicutes and Bacteroidetes bacteria, leading to high feacal SCFA (30, 36). Fiber is the most well-known nutrient with an important impact on microbiota. Diets rich in this nutrient increase SCFA producing bacteria, with benefits on intestinal barrier structure (11). Particularly the butyrate strengthens the barrier by increasing TJ protein expression and transepithelial electrical resistance (TER), and by consequently decreasing intestinal permeability and bacterial translocation. This has been shown to prevent the activation of effector T cells and to abrogate the manifestation of undesirable local and systemic inflammatory responses (9, 11, 37). The changes in gut microbial ecology and associated SCFA driven immune modulation can, in a logical way, explain the mechanisms behind clinical amelioration of RA in individuals exposed to MD. Despite of that, the validation in humans it's still needed (36).
Overall, additional research is necessary to elucidate the links between the health-promoting food pattern of the MD with gut microbiota characteristics (30).
Four small RCT have also studied the potential benefits of probiotics in RA. Probiotics are living organisms with a well-documented range of health benefits, associated not only with antimicrobial effects, but also to immune system, or with gut permeability (9, 10, 38, 39). In two small RCTs, Lactobacillus casei was shown to significantly reduce the expression of pro-inflammatory cytokines, namely IL-6 and TNF-α, and COX-2 activity, alleviating the manifestations of RA (3, 40, 41). Another study (42), with an association of probiotics (Lactobacillus acidophilus, Lactobacillus casei, and Bifidobacterium bifidum) resulted in improved DAS28 and a lower high sensitivity C-reactive protein concentrations when compared with placebo. Another RCT, testing a different Lactobacillus strain (L. rhamnosus), did not demonstrate any efficacy after one year of use (32, 43). In addition, several in vivo studies have assayed the use of probiotics as a therapeutic intervention. Lactobacillus casei was observed to attenuate symptoms in the mouse collagen-induced arthritis model by changing Th17 cells production, via no expression of IL-6, IL-17 and IL-23 cytokines (14, 15, 44). Same results were reported when using the Lactobacillus GG strain in the antigen-induced arthritis model in Lewis rats (14, 15, 45). Moreover, the use of E.coli Nissle 1917, Bifidobacterium infantis and Lactobacillus plantarum appears to have a positive impact on barrier strength with benefits on expression and production tight junction proteins (6, 9). By contrast, Lactobacillus bifidus showed a probable ability to promote joint swelling in germ free mice, by TLR2/TLR4 activation (3, 34, 46). Although these studies have globally suggested the potential benefits of probiotic supplementation in RA, until now the evidence is not considered strong enough to propose the inclusion of probiotic supplements in the diet of RA patients as part of their disease management strategy (47).
There are also some nutrients that can alter intestinal permeability and thereby influence the immune response without a known impact on the microbiota (Table 1). The depletion of some amino degrade carbohydrates acids like glutamine (55) or tryptophan (11) has been associated with a lower barrier function, translated in increased gut permeability. Vitamin D (7, 9, 11, 18) or polyphenols, as quercetin, myricetin, kaempferol, or curcumin have been shown to participate in the regulation of the intestinal barrier too, once they promote TER enhancement and a higher expression of TJ proteins, likeZO-1 and claudin-1. Also, a depletion of zinc, a micronutrient known to be essential for cell survival and function, has been shown to increase intestinal permeability (59). Interestingly most of these nutrients are commonly abundant in a MD pattern, which as referred before, is known as a potentially beneficial diet in RA. Apart from these nutrients, other food compounds, mostly present at western diets such as fatty acids [specially from high fat diets; (48)], alcohol (11), additives used in food industry (53), gliadin [protein present in wheat and several other cereals; (50, 51)], chitosan (52) or even some food processing methods using different microbial and fungal strains [that promotes eventual horizontal gene exchange; (60)], are known to negatively regulate barrier function via compromising its integrity as they alter TJ proteins expression and distribution, decrease TER and favor the growth of pathogenic or opportunistic bacteria (9, 61).
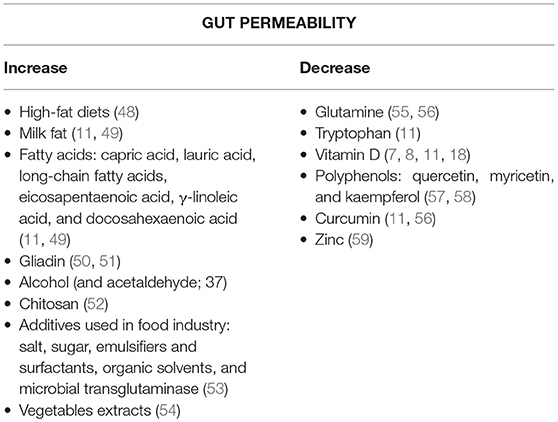
Table 1. Nutrients/Compounds that increase or decrease the intestinal permeability.
However, the real-life impact of all these nutrients on the intestinal permeability and the immune response in RA patients is still undisclosed (62).
Overall, it is now known that RA patients display a distinct gut microbiota composition at the moment of diagnosis in comparison to healthy individuals. Moreover, the microbiota composition of RA patients suffers further changes as the disease progresses. On the other hand, diet, especially the MD pattern, seems to influence the physiopathology of RA, potentially lowering disease activity, and improving the outcome (Figure 1).
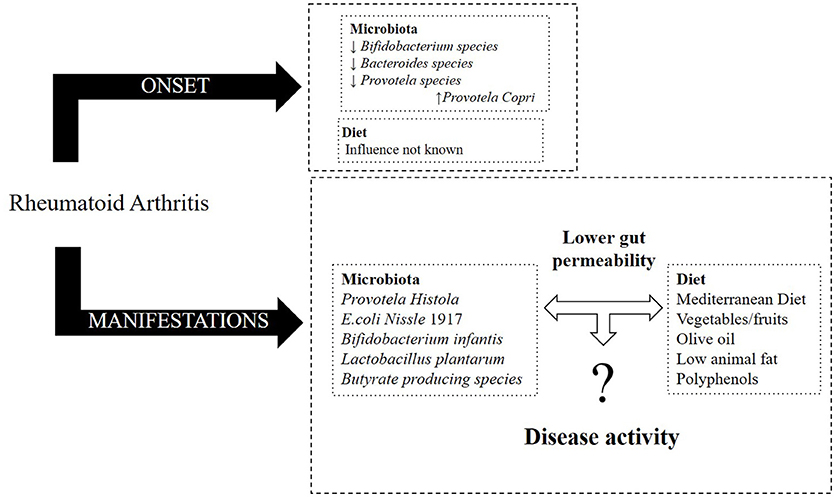
Figure 1. Effect of microbiota and diet in RA (onset and its manifestations).
However, critical questions remain to be elucidated, keeping the research agenda active in this field:
1) Are the clinical benefits observed with MD associated with the selective growth of a “healthy gut microbiota”? This possible association remains to be documented.
2) What is the relevance of intestinal permeability in this equation since both diet and microbiota seem to change its functionality? It is still unclear if intestinal permeability is directly affected independently by both variables or if the mechanism is sequential, depending on the influence of diet on microbioma.
3) How the microbiome fluctuates in relation to diet, and how disease activity may be influenced by changes in diet, microbiota or diet-intestinal microbiota equilibrium.
All authors listed have made substantial, direct, and intellectual contribution to the work and approved it for publication. CG, ÂC, JS, and JF specifically in concept. CG design and literature search. CG, ÂC, and JS data processing and writing manuscript. AC and JF critical review and in supervision.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med. (2011) 365:2205–19. doi: 10.1056/NEJMra1004965
2. Ciccia F, Ferrante A, Guggino G, Triolo G. The role of the gastrointestinal tract in the pathogenesis of rheumatic diseases. Best Pract Res Clin Rheumatol. (2016) 30:889–900. doi: 10.1016/j.berh.2016.10.003
3. Zhong D, Wu C, Zeng X, and Wang Q. The role of gut microbiota in the pathogenesis of rheumatic diseases. Clin Rheumatol. (2018) 37:25–34. doi: 10.1007/s10067-017-3821-4
4. Lerner A, Matthias T. Rheumatoid arthritis–celiac disease relationship: joints get that gut feeling. Autoimmun Rev. (2015) 14:1038–47. doi: 10.1016/j.autrev.2015.07.007
5. Van Spaendonk H, Ceuleers H, Witters L, Patteet E, Joossens J, Augustyns K, et al. Regulation of intestinal permeability: the role of proteases. World J Gastroenterol. (2017) 23:2106–23. doi: 10.3748/wjg.v23.i12.2106
6. Deane KD, Demoruelle MK, Kelmenson LB, Kuhn KA, Norris JM, Holers VM. Genetic and environmental risk factors for rheumatoid arthritis. Best Pract Res Clin Rheumatol. (2017) 31:3–18. doi: 10.1016/j.berh.2017.08.003
7. Mu Q, Kirby J, Reilly CM, Luo XM. Leaky gut as a danger signal for autoimmune diseases. Front Immunol. (2017) 8:598. doi: 10.3389/fimmu.2017.00598
8. Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, Keilbaugh SA, et al. Linking long-term dietary patterns with gut microbial enterotypes. Science (2011) 334:105–8. doi: 10.1126/science.1208344
9. Bischoff SC, Barbara G, Buurman W, Ockhuizen T, Schulzke JD, Serino M, et al. Intestinal permeability–a new target for disease prevention and therapy. BMC Gastroenterol. (2014) 14:189. doi: 10.1186/s12876-014-0189-7
10. Horta-Baas G, Romero-Figueroa MDS, Montiel-Jarquín AJ, Pizano-Zárate ML, García-Mena J, Ramírez-Durán N. Intestinal dysbiosis and rheumatoid arthritis: a link between gut microbiota and the pathogenesis of rheumatoid arthritis. J Immunol Res. (2017) 2017:1–13. doi: 10.1155/2017/4835189
11. De Santis S, Cavalcanti E, Mastronardi M, Jirillo E, Chieppa M. Nutritional keys for intestinal barrier modulation. Front Immunol. (2015) 6:612. doi: 10.3389/fimmu.2015.00612
12. Chen J, Wright K, Davis JM, Jeraldo P, Marietta EV, Murray J, et al. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis.Genome Med. (2016) 8:43. doi: 10.1186/s13073-016-0299-7
13. Diamanti AP, Rosado MM, Laganà B, D'Amelio R. Microbiota and chronic inflammatory arthritis: an interwoven link. J Transl Med. (2016) 14:233. doi: 10.1186/s12967-016-0989-3
14. Kim D, Yoo SA, Kim WU. Gut microbiota in autoimmunity: potential for clinical applications. Arch Pharm Res. (2016) 39:1565–76. doi: 10.1007/s12272-016-0796-7
15. Vaahtovuo J, Munukka E, Korkeamaki M, Luukkainen R, Toivanen P. Fecal microbiota in early rheumatoid arthritis. J Rheumatol. (2008) 35:1500–5.
16. Marietta EV, Murray JA, Luckey DH, Jeraldo PR, Lamba A, Patel R, et al. Human gut-derived Prevotella histicola suppresses inflammatory arthritis in humanized mice. Arthritis Rheumatol. (2016) 68:2878–88. doi: 10.1002/art.39785
17. Lerner A, Aminov R, Matthias T. Dysbiosis may trigger autoimmune diseases via inappropriate post-translational modification of host proteins. Front Microbiol. (2016) 7:84. doi: 10.3389/fmicb.2016.00084
18. Scher JU, Sczesnak A, Longman RS, Segata N, Ubeda C, Bielski C, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. ELife (2013) 2:e01202. doi: 10.7554/eLife.01202
19. Maeda Y, Kurakawa T, Umemoto E, Motooka D, Ito Y, Gotoh K, et al. Dysbiosis contributes to arthritis development via activation of autoreactive T cells in the intestine. Arthritis Rhematol. (2016) 68:2646–61. doi: 10.1002/art.39783
20. Bernard NJ. Rheumatoid arthritis: prevotella copri associated with new-onset untreated RA. Nat Rev Rheumatol. (2014) 10:2. doi: 10.1038/nrrheum.2013.187
21. Ley RE. Gut microbiota in 2015: prevotella in the gut: choose carefully. Nat Rev Gastroenterol Hepatol. (2016 13:69–70. doi: 10.1038/nrgastro.2016.4
22. De Filippis F, Pellegrini N, Laghi L, Gobbetti M, Ercolini D. Unusual sub-genus associations of faecal Prevotella and Bacteroides with specific dietary patterns. Microbiome (2016) 4:2–6. doi: 10.1186/s40168-016-0202-1
23. Kau AL, Ahern PP, Griffin NW, Goodman AL, Gordon JI. Human nutrition, the gut microbiome and the immune system. Nature (2011) 474:327–36. doi: 10.1038/nature10213
24. Lerner A, Neidhöfer S, Matthias T. The gut microbiome feelings of the brain: a perspective for non-microbiologists. Microorganisms (2017) 12:E66. doi: 10.3390/microorganisms5040066
25. Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature (2011) 473:174–80. doi: 10.1038/nature09944
26. Hjorth MF, Roager HM, Larsen TM, Poulsen SK, Licht TR, et al. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int J Obes. (2018) 42:580–3. doi: 10.1038/ijo.2017.220
27. Gorvitovskaia A, Holmes SP, Huse SM. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome (2016) 4:1–12. doi: 10.1186/s40168-016-0160-7
28. Sofi F, Cesari F, Abbate R, Gensini GF, Casini A. Adherence to Mediterranean diet and health status: meta-analysis. BMJ (2008) 337:2–7. doi: 10.1136/bmj.a1344
29. Willett WC, Sacks F, Trichopoulou A, Drescher G, Ferro-Luzzi A, Helsing E, et al. Mediterranean diet pyramid: a cultural model for healthy eating. Am J Clin Nutr. (1995) 61:1402S−6S. doi: 10.1093/ajcn/61.6.1402S
30. Mitsou EK, Kakali A, Antonopoulou S, Mountzouris KC, Yannakoulia M, Panagiotakos DB, et al. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br J Nutr. (2017) 117:1645–55. doi: 10.1017/S0007114517001593
31. Bifulco M. Mediterranean diet: the missing link between gut microbiota and inflammatory diseases. Eur J Clin Nutr. (2015) 69:1078. doi: 10.1038/ejcn.2015.81
32. Clemente JC, Manasson J, Scher JU. The role of the gut microbiome in systemic inflammatory disease. BMJ (2018) 2018:5145. doi: 10.1136/bmj.j5145
33. Skoldstam L, Hagfors L, Johansson G. An experimental study of a Mediterranean diet intervention for patients with rheumatoid arthritis. Ann Rheum Dis. (2003) 62:208–14. doi: 10.1136/ard.62.3.208
34. McKellar G, Morrison E, McEntegart A, Hampson R, Tierney A, Mackle G, et al. A pilot study of a Mediterranean-type diet intervention in female patients with rheumatoid arthritis living in areas of social deprivation in Glasgow. Ann Rheum Dis. (2007) 66:1239–43. doi: 10.1136/ard.2006.065151
35. Tedeschi SK, Costenbader KH. Is there a role for diet in the therapy of rheumatoid arthritis? Curr Rheumatol Rep. (2016) 18:23. doi: 10.1007/s11926-016-0575-y
36. De Filippis F, Pellegrini N, Vannini L, Jeffery IB, La Storia A, Laghi L, et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut (2016) 65:1812–21. doi: 10.1136/gutjnl-2015-309957
37. Lerner A, Patricia J, Matthias T. Nutrients, bugs and us: the short-chain fatty acids story in celiac disease. Int J Celiac Dis. (2016) 4:92–4. doi: 10.12691/ijcd-4-3-12
38. Stoidis CN, Misiakos EP, Patapis P, Fotiadis CI, Spyropoulos BG. Potential benïits of pro- and prebiotics on intestinal mucosal immunity and intestinal barrier in short bowel syndrome. Nutr Res Rev. (2011) 24:21–30. doi: 10.1017/S0954422410000260
39. Foschi C, Laghi L, Parolin C, Giordani B, Compri M, Cevenini R, et al. Novel approaches for the taxonomic and metabolic characterization of lactobacilli: integration of 16S rRNA gene sequencing with MALDI-TOF MS and 1H-NMR. PLoS ONE (2017) 12:e0172483. doi: 10.1371/journal.pone.0172483
40. Alipour B, Homayouni-Rad A, Vaghef-Mehrabany E, Sharif SK, Vaghef-Mehraany L, Asghari-Jafarabadi M, et al. Effects of Lactobacillus casei supplementation on disease activity and inflammatory cytokines in rheumatoid arthritis patients: a randomized double-blind clinical trial. Int J Rheum Dis. (2014) 17:519–27. doi: 10.1111/1756-185X.12333
41. Hatakka K, Martio J, Korpela M, Herranen M, Poussa T, Laasanen T, et al. Effects of probiotic therapy on the activity and activation of mild rheumatoid arthritis-a pilot study. Scand J Rheumatol. (2003) 32:211–5. doi: 10.1080/03009740310003695
42. Zamani B, Golkar HR, Farshbaf S, Emadi-Baygi M, Tajabadi-Ebrahimi M, Jafari P, et al. Clinical and metabolic response to probiotic supplementation in patients with rheumatoid arthritis: a randomized, double-blind, placebo-controlled trial. Int J Rheum Dis. (2016) 19:869–79. doi: 10.1111/1756-185X.12888
43. Pineda MA, Thompson SF, Summers K, de Leon F, Pope J, Reid G. A randomized, double-blinded, placebo-controlled pilot study of probiotics in active rheumatoid arthritis. Med Sci Monitor (2011) 17:CR347–54. doi: 10.12659/MSM.881808
44. Amdekar S, Singh V, Singh R, Sharma P, Keshav P, Kumar A. Lactobacillus casei reduces the inflammatory joint damage associated with collagen-induced arthritis (CIA) by reducingg the pro-inflammatory cytokines. J Clin Immunol. (2011) 31:147–54. doi: 10.1007/s10875-010-9457-7
45. Baharav E, Mor F, Halpern M, Weinberger A. Lactobacillus GG bacteria ameliorate arthritis in Lewis rats. J Nutr. (2004) 134:1964–9. doi: 10.1093/jn/134.8.1964
46. Bdollahi-Roodsaz S, Joosten LA, Koenders MI, Devesa I, Roelofs MF, Radstake TR, et al. Stimulation of TLR2 and TLR4 differentially skews the balance of T cells in a mouse model of arthritis. J Clin Invest. (2008) 118:205–16. doi: 10.1172/JCI32639
47. Rudbane SMA, Rahmdel S, Abdollahzadeh SM, Zare M, Bazrafshan A, Mazloomi SM. The efficacy of probiotic supplementation in rheumatoid arthritis: a meta-analysis of randomized, controlled trials. Inflammopharmacology (2018) 26:67–76. doi: 10.1007/s10787-017-0436-y
48. Graf D, Di Cagno R, Fåk F, Flint HJ, Nyman M, Saarela M, et al. Contribution of diet to the composition of the human gut microbiota. Microb Ecol Health Dis. (2015) 26:26164. doi: 10.3402/mehd.v26.26164
49. Guzman JR, Conlin VS, Jobin C. Diet, microbiome, and the intestinal epithelium: an essential triumvirate? Biomed Res Int. (2013) 2013:425146. doi: 10.1155/2013/425146
50. Lammers KM, Lu R, Brownley J, Lu B, Gerard C, Thomas K, et al. Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3. Gastroenterology (2008) 135:194–204.e3. doi: 10.1053/j.gastro.2008.03.023
51. Lerner A, Shoenfeld Y, Matthias T. Adverse effects of gluten ingestion and advantages of gluten withdrawal in nonceliac autoimmune disease. Nutr Rev. (2017) 75:1046–58. doi: 10.1093/nutrit/nux054
52. Schipper NG, Olsson S, Hoogstraate JA, deBoer AG, Vårum KM, Artursson P. Chitosans as absorption enhancers for poorly absorbable drugs 2: mechanism of absorption enhancement. Pharm Res. (1997) 14:923–9. doi: 10.1023/A:1012160102740
53. Lerner A, Matthias T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun Rev. (2015) 14:479–89. doi: 10.1016/j.autrev.2015.01.009
54. Ulluwishewa D, Anderson RC, McNabb WC, Moughan PJ, Wells JM, Roy NC. Regulation of tight junction permeability by intestinal bacteria and dietary components. J Nutr. (2011) 141:769–76. doi: 10.3945/jn.110.135657
55. Lima AA, Brito LF, Ribeiro HB, Martins MC, Lustosa AP, Rocha EM, et al. Intestinal barrier function and weight gain in malnourished children taking glutamine supplemented enteral formula. J Pediatr Gastroenterol Nutr. (2005) 40:28–35. doi: 10.1097/00005176-200501000-00006
56. Rapin JR, Wiernsperger N. Possible links between intestinal permeablity and food processing: a potential therapeutic niche for glutamine. Clinics (2010) 65:635–43. doi: 10.1590/S1807-59322010000600012
57. Rao RK, Basuroy S, Rao VU, Karnaky KJ Jr, Gupta A. Tyrosine phosphor-ylation and dissociation of occludin-ZO-1 and E-cadherin-beta-catenin complexes from the cytoskeleton by oxidative stress. Biochem J. (2002) 368:471–81. doi: 10.1042/BJ20011804
58. Atkinson KJ, Rao RK. Role of protein tyrosine phosphorylation in acet- aldehyde-induced disruption of epithelial tight junctions. Am J Physiol Gastrointest Liver Physiol. (2001) 280:G1280–8. doi: 10.1152/ajpgi.2001.280.6.G1280
59. Finamore A, Massimi M, Conti Devirgiliis L, Mengheri E. Zinc deficiency induces membrane barrier damage and increases neutrophil transmigration in Caco-2 cells. J Nutr. (2008) 138:1664–70. doi: 10.1093/jn/138.9.1664
60. Lerner A, Matthias T, Aminov R. Potential effects of Horizontal Gene exchange in the Human Gut. Front Immunol. (2017) 8:1630. doi: 10.3389/fimmu.2017.01630
61. Suzuki T. Regulation of intestinal epithelial permeability by tight junctions. Cell Mol Life Sci (2013) 70:631–59. doi: 10.1007/s00018-012-1070-x
Keywords: diet, Mediterranean Diet, gut microbiota, gut permeability, rheumatoid arthritis
Citation: Guerreiro CS, Calado Â, Sousa J and Fonseca JE (2018) Diet, Microbiota, and Gut Permeability—The Unknown Triad in Rheumatoid Arthritis. Front. Med. 5:349. doi: 10.3389/fmed.2018.00349
Received: 26 April 2018; Accepted: 27 November 2018;
Published: 14 December 2018.
Edited by:
Kayo Masuko, Sanno Medical Center, JapanReviewed by:
Aaron Lerner, AESKU.KIPP Institute, GermanyCopyright © 2018 Guerreiro, Calado, Sousa and Fonseca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Catarina Sousa Guerreiro, Y2ZndWVycmVpcm9AbWVkaWNpbmEudWxpc2JvYS5wdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.