
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 15 April 2025
Sec. Global Change and the Future Ocean
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1586402
 Jessica Lombardo1
Jessica Lombardo1 Silvia Tejada2,3,4
Silvia Tejada2,3,4 Montserrat Compa1
Montserrat Compa1 Vicent Forteza5Lorenzo Gil6
Vicent Forteza5Lorenzo Gil6 Samuel Pinya3,6
Samuel Pinya3,6 Antoni Sureda1,3,4*
Antoni Sureda1,3,4*Algae represent more than 41% of marine biological invasions worldwide. If non-native algae become an invasive and dominant species, they will inevitably affect the characteristics of the ecosystem. The study aimed to evaluate the potential effect of the recently arrived Batophora occidentalis growing over three native algae: Acetabularia acetabulum, Dasycladus vermicularis and Caulerpa prolifera in S’Estany des Peix (Formentera Island, Spain), a shallow saltwater lagoon environment, by means of antioxidant biomarkers. The environmental conditions of the lagoon favoured the extensive progression of B. occidentalis, which since 2020 has progressively been spreading to the entire perimeter of the lagoon where it has found a substrate to attach itself to, even epiphytize native algae. The activities of the antioxidant enzymes (catalase, superoxide dismutase, and glutathione reductase), the phase II detoxification enzyme glutathione S-transferase and levels of malondialdehyde (as an indicator of lipid peroxidation) were evaluated. The results showed significant increases in all monitored biomarkers in the three native algae affected by B. occidentalis, with notable elevations in antioxidant enzyme activities and detoxification response. However, MDA levels increased only in A. acetabulum and D. vermicularis, but not in C. prolifera, suggesting that the latter species exhibits greater resistance to lipid peroxidation. These findings indicate that native algae experience biochemical stress, likely induced by competition or environmental changes caused by B. occidentalis. In conclusion, the presence of the invasive alga B. occidentalis can induce a stressful situation in epiphytized native algae, as evidenced by an increase in oxidative stress biomarkers. Further studies are needed to assess its potential effects on the growth and survival of the affected algae, as well as its ecological impact on the ecosystem.
Global change is the set of large-scale environmental alterations and transformations that result from anthropogenic activities and impact our planet. Over the past fifty years, humans have altered the structure and functioning of the world’s ecosystems more rapidly and extensively than in any other period in human history, causing a substantial and largely irreversible loss of biodiversity. For example, the rise in ocean temperature has facilitated changes in the distributional ranges of species, faster than those observed in terrestrial ecosystems, due to the open nature of the oceanic system and the high mobility of marine species (Poloczanska et al., 2016). One of these main drivers is climate change, which can accelerate the introduction and spread of alien species (Walther et al., 2002; Thuiller, 2007). These long-term shifts in increasing temperatures can contribute to their spread by providing favourable conditions for their growth in addition to increasing resource competition with native fauna. These alien species can become invasive if they negatively impact native biodiversity, ecosystem services or human well-being Invasive Alien Species and Climate Change International union for conservation of nature (IUCIN) (2021). The combination of invasive species and climate change can reduce ecosystem resilience and negatively impact biodiversity. The arrival of a new invasive species could also result in physiological stress for native species due to direct competition for substrate and nutrients, the release of toxins or allelochemical compounds or a reduction in light availability (Ballesteros et al., 2007; Pergent et al., 2008; Gniazdowska et al., 2015). Invasive algae are a major global concern, as they are known to deeply modify marine ecosystems, having strong ecological and economic impacts (Schaffelke et al., 2006; Williams and Smith, 2007; Tomas et al., 2011). Anthropogenic impacts can contribute to the widespread proliferation of new algae, particularly in highly disturbed areas. The introduction of alien marine species has accelerated in recent decades due to rapid globalization and increasing trends in trade, travel, and transportation (Katsanevakis et al., 2013, Katsanevakis et al., 2014; Gollasch et al., 2019).
The Mediterranean Sea, due to its characteristics as a semi-enclosed sea, densely populated in its coastal areas and subject to high maritime traffic, is highly susceptible to suffering the consequences of human action. It is a hotspot for introduced marine species, many of them potentially invasive (Galil, 2000; Boudouresque, 2008), and on a global scale, it harbours the highest number of introduced species (Boudouresque and Verlaque, 2002; Fortič et al., 2023). In this sense, the Mediterranean Sea is the most affected region in the world in terms of exotic algae (Williams and Smith, 2007). The most common source of introduction into the Mediterranean is directly associated with ships, since introduced species can travel attached to the hulls or with ballast water (Gollasch et al., 2019). Moreover, a recent study highlighted that most non-native macrophyte species are not limited by lower temperatures due to their thermal plasticity and, consequently, those algae that originate from areas with warmer temperatures can spread throughout the Mediterranean (Wesselmann et al., 2024). Regarding the potential effects of the arrival of new species, those with invasive character have led in the Mediterranean to significant biogeographic changes, displacement of native species, permanent shifts in the structure and function of colonised benthic habitats, and, in some cases, local extinctions (Mancuso et al., 2022). In fact, the Mediterranean is one of the regions where the ecological impacts of this phenomenon are predicted to be particularly damaging, due to its unique biogeographical features, and due to the increase in its surface water temperatures at a rate greater than that of the oceans, favouring the settlement of macroalgae of tropical origin (Taylor et al., 2021). Among the established invasive macroalgae with described ecological effects, the chlorophyte Caulerpa cylindracea and the rhodophytes Womersleyella setacea, Asparagopsis taxiformis and Lophocladia lallemandii stand out, especially in the western Mediterranean ecoregion (Tsirintanis et al., 2022).
Formentera Island (Balearic Islands, Western Mediterranean) is a hotspot for luxury yachting, which has been listed as a pathway for species introductions (Hewitt et al., 2007; Brine et al., 2013; Cecere et al., 2016). S’Estany des Peix is a small and shallow lagoon environment on Formentera Island with a narrow opening to the sea, which allows the entry and mooring of small boats. The lagoon is characterised by a community of photophilous algae with a predominance of species such as Acetabularia acetabulum (Linnaeus, 1952), Dasycladus vermicularis (Scopoli, 1898) Caulerpa prolifera (Forsskal, 1809) and Halimeda tuna (J. Ellis & Solander) J.V. Lamouroux 1816. The calm and protected waters of the lagoon, together with rising temperatures, create suitable conditions for species with higher thermal tolerances, such as those from the Caribbean and the eastern tropical Atlantic. Batophora occidentalis (Harvey, 1998), a green alga native to the Atlantic coasts of the Caribbean and Gulf of Mexico, was first detected at the entrance of S’Estany des Peix in May 2020, blooming on the northern side of the lagoon. Since then, it has rapidly spread, colonizing the lagoon’s rocky substrate and growing on native species like algae and sponges (Ballesteros et al., 2008) (Terradas-Fernández et al., 2022). This rapid expansion poses an ecological threat due to increased competition with native fauna and potential alterations to ecosystem dynamics. Furthermore, members of the Dasycladaceae family, to which B. occidentalis belongs, such as the genus Dasycladus, are known to produce phenolic acids and coumarins, which may act as biofilm inhibitors or have allelopathic effects. Thus, B. occidentalis may not only cause epiphytism but also engage in allelopathic interactions (Hartmann et al., 2018). The most likely pathway of introduction of this alga into the lagoon is yachting, probably via fouling of adult individuals or via ballast water transporting gametophores or gametangia (Ballesteros, 2020). In S’Estany des Peix, B. occidentalis has been observed covering small algae, such as A. acetabulum, D. vermicularis and C. prolifera as shown in Figure 1. According to the classification by Leonardi et al. (2006), the epiphytism of B. occidentalis on the studied species falls between type I and II infections, as it is moderately attached to the surface of the host without causing any evidence of host tissue damage.

Figure 1. Representative pictures of Acetabularia acetabulum, Dasycladus vermicularis and Caulerpa prolifera under normal growth conditions (A, C, E) and, respectively, under epiphytism conditions by Batophora occidentalis (B, D, F).
The arrival and spread of invasive species, such as B. occidentalis, not only affect biodiversity and ecosystem dynamics but also induce physiological stress in native species. Thus, the invasion of alien species triggers a series of biochemical responses that compromise the cellular health of native organisms, further exacerbating the negative ecological effects of invasion. Specifically, when organisms are faced with a potentially stressful situation, they often respond with metabolic activation, resulting in an increase in reactive oxygen species (ROS) production (Ross et al., 2006), which includes hydrogen peroxide (H2O2), superoxide anion (O−2), and hydroxyl radical (OH•) (Bartoskova et al., 2013; Faggio et al., 2016). Under physiological conditions, ROS act as secondary messengers in numerous cellular processes but, when overproduced, can oxidise biomolecules and induce oxidative damage (Rezayian et al., 2019). To remove excess ROS and prevent oxidative damage, cells contain a complex network of antioxidant defences that remove or prevent the generation of ROS, which includes both enzymatic and non-enzymatic mechanisms (Tejada and Sureda, 2014; Rezayian et al., 2019). Furthermore, the presence of invasive species can induce oxidative stress conditions for native species (Guzzetti et al., 2019; Quetglas-Llabrés et al., 2020). For example, previous studies have shown the induction of oxidative stress in different species such as the phanerogam Posidonia oceanica and the bryozoan Reteporella grimaldii epiphytized by the invasive red alga L. lallemandi or in the algae D. vermicularis and Padina Pavonica affected by the tropical green alga Halimeda incrassata (Sureda et al., 2008, Sureda et al., 2017; Deudero et al., 2010; Cerrato et al., 2023).
Considering all this information, the aim of the present study was to evaluate the potential effect of the recently arrived B. occidentalis growing over three native and abundant algae: A. acetabulum (Linnaeus, 1952), D. vermicularis (Scopoli, 1898) and C. prolifera (Forsskal, 1809), in S’Estany des Peix. This was achieved by applying antioxidant-related biomarkers to compare the communities of each species affected by the invasion of B. occidentalis with those that have not been impacted by its growth. The selected biomarkers—antioxidant enzymes (catalase, superoxide dismutase, glutathione reductase), the detoxification enzyme glutathione S-transferase, and lipid peroxidation marker malondialdehyde—are widely used in oxidative stress studies. These biomarkers provide insight into the cellular response to environmental stressors, allowing us to assess the impact of B. occidentalis on native algae.
S’Estany des Peix (38° 43′ 33″ N, 1° 24′ 45″ E) is a small saltwater lagoon located within the Ses Salines d’Eivissa i Formentera Natural Park on the northern coast of Formentera Island, which is part of the Balearic Islands in the western Mediterranean Sea (Figure 2). The lagoon is designated as a Ramsar site (Site no. 641) and is also part of the Natura 2000 Network, classified as both a Special Protection Area for Birds (ZEPA) and a Special Conservation Area (ZEC) under the code ES0000084 Ses Salines d’Eivissa i Formentera. Its waters host several priority habitats recognised by the European Union, including 1110 (Posidonia oceanica meadows) and 1150 (coastal lagoons), as outlined in Directive 92/43/EEC of 21 May 1992. It has a roughly circular shape, with a length of about 1040 m (north-south axis) and a width of 1070 m (east-west axis). It occupies a total area of about 105 hectares, and the maximum bathymetry does not exceed 5 meters in depth in the central area. The central and northern part is separated from the sea by a dune coastal bar that is open to a small shoreline (50 m wide). Caulerpa prolifera meadows predominate in the bottoms with an occupancy of approximately 49.9%, followed by sandy bottoms (28.8%) and communities of photophilous algae (15.1%) (Ballesteros et al., 2008). According to E.U. Copernicus Marine Service Information (https://doi.org/10.48670/moi-00170), sea surface temperatures for s’Estany des Peix ranged from ~14°C during winter months to above 29°C during summer months. The temperature fluctuations were slightly higher than those of the open sea, due to the shallow depth of the lagoon. This increase in seasonal temperature can cause significant ecological implications, especially for species that are sensitive to specific temperatures in addition to shifts in species distribution and community structure. The lagoon is euhaline, becoming slightly hypersaline further from the open sea, reaching 39 PSU. In terms of ecological importance, S’Estany des Peix harbours a diverse range of algal and seagrass species, including Posidonia oceanica (L.) Delile, Cymodocea nodosa (Ucria) Asch., Stuckenia pectinata (L.) Börner, Ruppia cirrhosa (Petagna) Grande, and Zostera noltii Hornem.
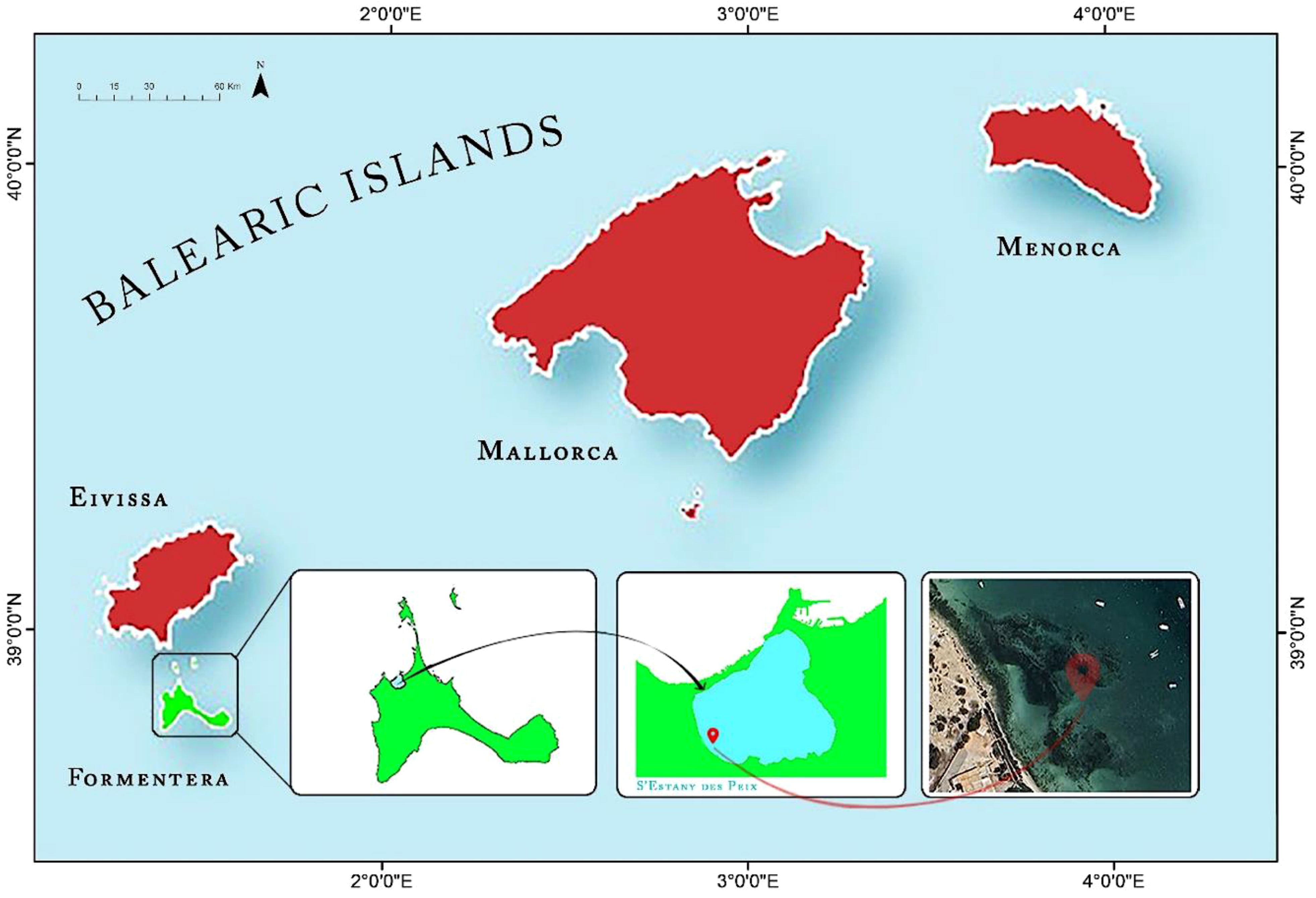
Figure 2. Map of the geographic location of the Balearic Islands with the names of the four principal islands. Formentera Island is indicated in green and “S’Estany des Peix” in light blue. The sampling area is indicated with the red map marker.
The monitoring of the coverage and evolution of the occupation of B. occidentalis was carried out by members of the Natural Park, inspecting the entire perimeter of the lagoon from the shore to a depth of 50 cm. The survey was conducted annually by walking in a zig-zag pattern along the lagoon, through systematic georeferencing using GPS devices to mark all the areas where B. occidentalis was present. The first survey, in 2020, showed a coverage of 18%, concentrated in two areas: the northern area near the exit to the sea and the northeastern area (Figure 3). By the end of 2021, the coverage had increased, reaching approximately 47% of the perimeter. By 2022, an increased expansion in the coverage of B. occidentalis is observed, reaching close to 100% of the perimeter of the lagoon. This high level of coverage continued into 2023 and 2024 (Forteza et al., 2024), extending to the outer zone of the connection with the open sea.

Figure 3. Progression of the expansion of Batophora occidentalis in S’Estany des Peix (Formentera Island) from its detection in 2020 (A) to 2024 (B), where it has practically occupied the entire perimeter of the lagoon.
All samples were collected by free diving close to shore, in the upper infralittoral zone, at an average depth of 40-60 cm in the south-west coast of the lagoon. The selection of individuals was done randomly, although specimens of similar size were chosen to avoid potential biases related to biomass or metabolic differences. Eight biologically independent replicates for the three species in each situation (affected and not affected by B. occidentalis) were obtained, for a total of 48 samples. The sampling of Acetabularia acetabulum, Dasycladus vermicularis and Caulerpa prolifera was carried out on April 10 and 11, 2023. The samples were rapidly introduced into hermetic recipients containing seawater and transported to the laboratory and immediately frozen at −80°C until biochemical analysis. In addition, five different temperature and salinity measurements were taken throughout the sampling area using a multiparameter probe (Hanna HI9828) to characterise the environmental conditions.
The three algae species chosen were the three most common algae found within S’Estany des Peix and, in addition, they have been observed to be encroached by the invasive B. occidentalis. A. acetabulum is unicellular green alga native to the Mediterranean Sea and grows in the upper infralittoral zone in calm and well-lit areas. D. vermicularis is a green macroalga commonly found in similar infralittoral conditions. Previous studies have reported that this species, along with others, is particularly susceptible to the impacts of non-native species, underscoring its potential sensitivity to invasions (Sureda et al., 2017). C. prolifera is another native green alga species that is found in shallow sandy or muddy bottoms of the Balearic Islands and is often found coexisting with Posidonia oceanica, providing essential stabilizations of the coastal seafloor. Given the unique conditions of s’Estany des Peix—shallow waters, ample illumination, and low hydrodynamics—the infralittoral zone offers an ideal habitat for all the selected species.
First, algae samples (0.2 g per sample) were rinsed with distilled water and homogenised under ice-cold conditions, on ice, in a 1:5 (w/v) ratio of 50 mM Tris–HCl buffer, 1 mM EDTA, pH 7.5 using a small sample dispersing system (ULTRA-TURRAX® Disperser, IKA) and centrifuged at 9000×g, 10 min, 4°C (Sureda et al., 2006). After centrifugation, the supernatants were collected and used for all biochemical analyses. All biochemical assays were performed in duplicate to ensure reproducibility
CAT, SOD and GRd activities as antioxidant defences and GST activity as an enzyme implicated in detoxification processes were determined. MDA levels as a marker of oxidative damage to lipids and ROS production as indicative of metabolic stress were also evaluated. CAT activity (mK (s-1)/mg protein) was measured as previously described by Aebi (1984) at 240 nm. SOD activity (pKat/mg protein) was determined at 550 nm according to Flohé and Ötting (1984). GRd activity (nmol/min/mg protein) was measured by a modification of the Goldberg and Spooner (1983) method at 339 nm. GST activity (nKat/mg protein) was determined at 340 nm using the technique of Habig et al. (1974). All enzymatic activities were determined using a Shimadzu UV-2100 spectrophotometer at 25°C. MDA levels (nM/mg protein) were quantified by a colorimetric assay kit (Merk Life Science S.L.U., Madrid, Spain), following the manufacturer’s instructions. The assay was based on the reaction of MDA with a chromogenic reagent to form a coloured product measured at 586 nm in a microplate reader (Bio-Tek®, PowerWaveXS). ROS production was measured in the homogenates using the fluorescent probe 2,7-dichlorofluorescein-diacetate (DCFH-DA). Briefly, samples were introduced in a 96-well microplate containing phosphate-buffered saline, pH 7.4. Then, DCFH-DA (61.6 μM in Hanks’ Balanced Salts Medium) was added to each well and fluorescence (Ex, 480 nm; Em, 530 nm) was monitored for 1 hour in FLx800 Microplate Fluorescence Reader (Biotek Instruments, Inc., Winuschi, VT, USA) at 25°C.
All biochemical analyses were referred to the total protein content of the samples determined with the colorimetric Bradford method (Biorad® Protein Assay, Alcobendas, Spain) using bovine serum albumin as a standard.
Statistical analyses were performed using the statistical package for social sciences (IBM PSS® v27.0, Armonk, NY, USA). The normality of the data was assessed by the Shapiro–Wilk test and homogeneity of variances by the Levene’s test. The statistical significance was compared by t-test for unpaired data, with the absence or presence of the invasive species as a comparison factor. Results were expressed as mean ± standard error of the mean (S.E.M.) and p < 0.05 was considered statistically significant.
Water temperature reported an average value of 17.1 ± 0.1°C, and salinity had an average value of 38.5 ± 0.2 PSU during the sampling period.
B. occidentalis was detected for the first time in 2020 in the S’Estany des Peix covering rocky substrates in the northern area (Figure 3). Since then, the alga has progressively expanded to cover the entire perimeter area of the lagoon between approximately 20 and 100 cm deep, except for the sandiest areas where it cannot settle. Furthermore, as seen in Figure 1, B. occidentalis is capable of epiphytize other algae species, covering them almost completely.
Beyond its distribution and expansion, the presence of B. occidentalis also has biochemical implications. The different biomarkers analysed in the three native algae are reported in Figures 4–7. In addition, the numerical values of these parameters are presented as Supplementary Table 1.

Figure 4. Antioxidant enzyme activities of A. acetabulum, D. vermicularis and C. prolifera samples, alone (control situation) and in competition with the invasive alga B. occidentalis. (A) catalase (CAT); (B) superoxide dismutase (SOD); (C) glutathione reductase (GRd). * indicates statistically significant differences (p < 0.05) compared to control. Values represent mean ± S.E.M.
The presence of the invasive alga B. occidentalis significantly increased the enzymatic activities in all three native algae compared to unaffected conditions. The activities of antioxidant enzymes, CAT, SOD and GRd significantly increased in the presence of the invasive alga (Figure 4).
Similarly, GST activity significantly increased in all three algae species affected by B. occidentalis compared to unaffected specimens (Figure 5).
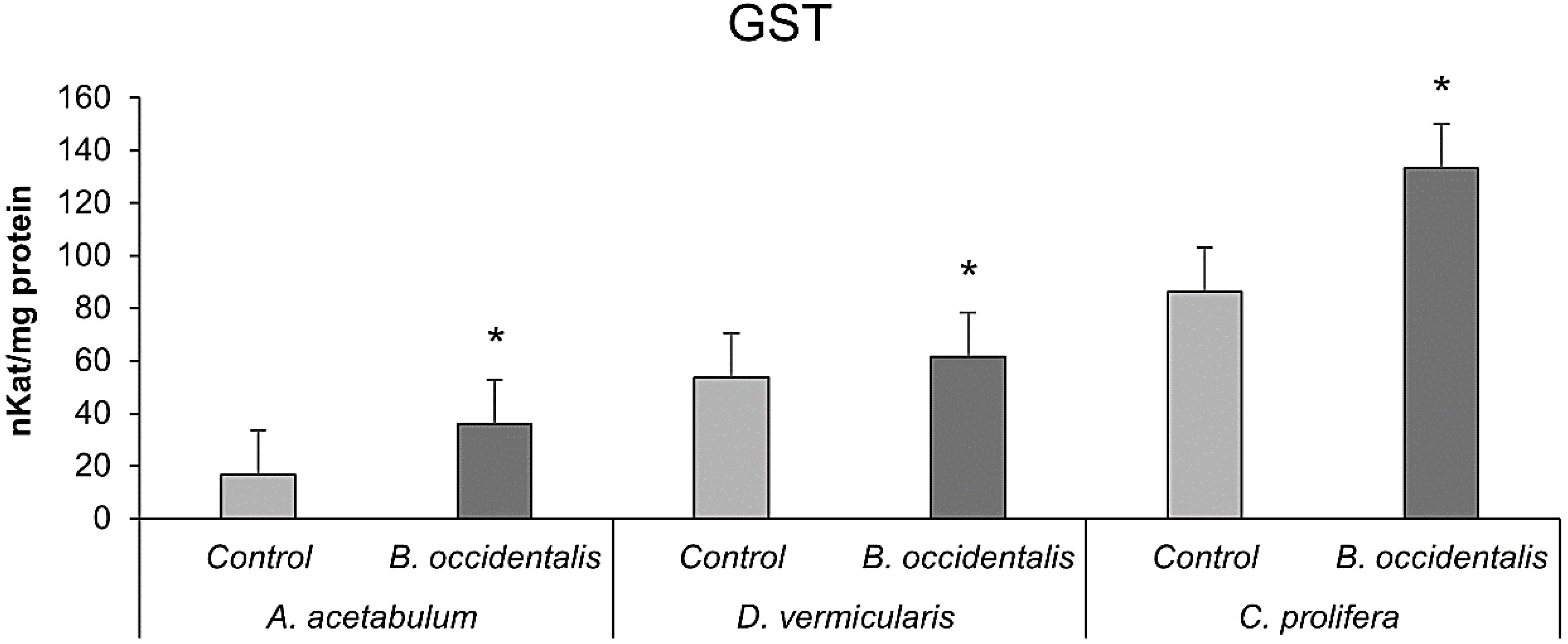
Figure 5. Activity of glutathione S-transferase (GST) of A. acetabulum, D. vermicularis and C. prolifera samples, alone (control situation) and in competition with the invasive alga B. occidentalis. * indicates statistically significant differences (p < 0.05) compared to the control. Values represent mean ± S.E.M.
The MDA levels in the three algae affected and unaffected by B. occidentalis are shown in Figure 6. MDA levels increased significantly in A. acetabulum and D. vermicularis in the presence of the invasive alga, but not in C. prolifera which maintains similar values.
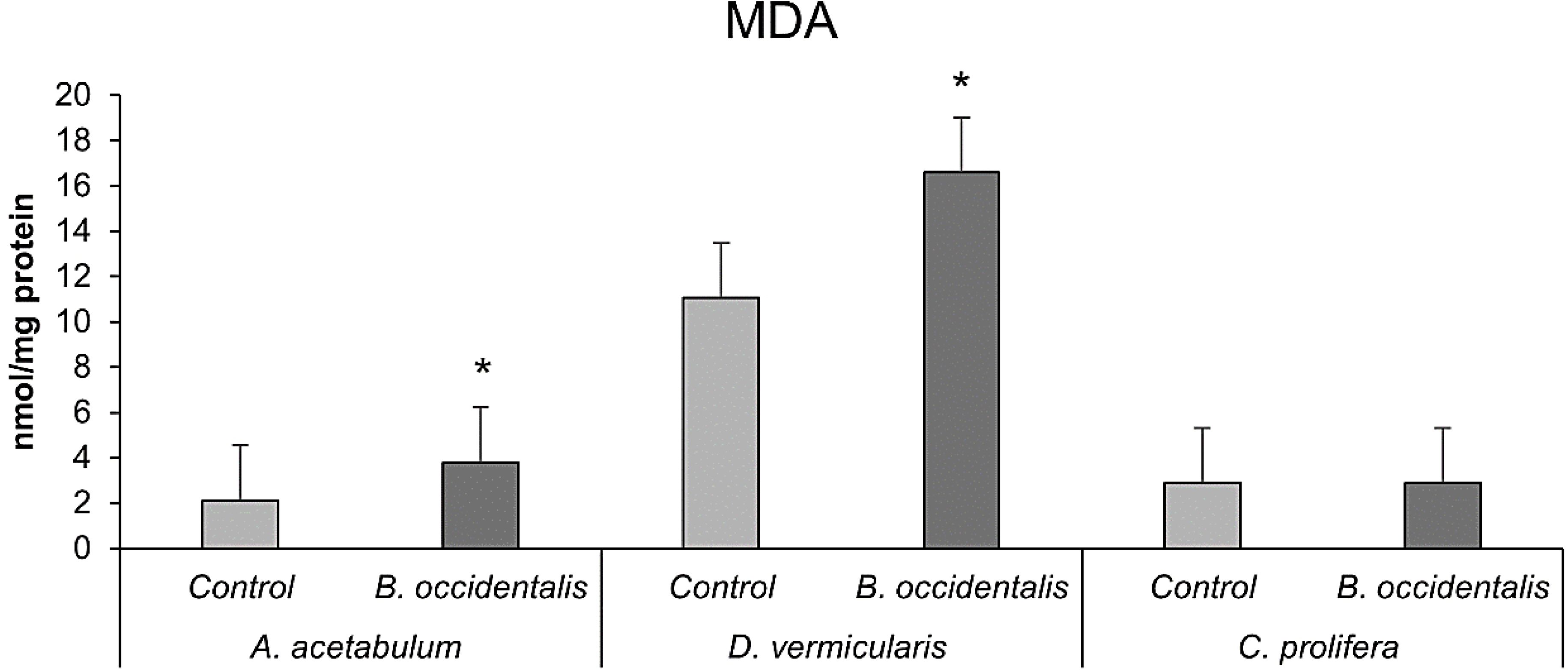
Figure 6. Malondialdehyde (MDA) levels of A. acetabulum, D. vermicularis and C. prolifera samples, alone (control situation) and in competition with the invasive alga B. occidentalis. * indicates statistically significant differences (p < 0.05) compared to the control. Values represent mean ± S.E.M. No significant differences were found in C. prolifera samples.
ROS production in the three algae species studied was higher in those specimens in direct contact with B. occidentalis (Figure 7). When analysing the percentage of increase between algae unaffected and affected by B. occidentalis, in the case of A. acetabulum and D. vermicularis it was 28.6 and 25.2%, respectively, while for C. prolifera the increase was 12.9%.
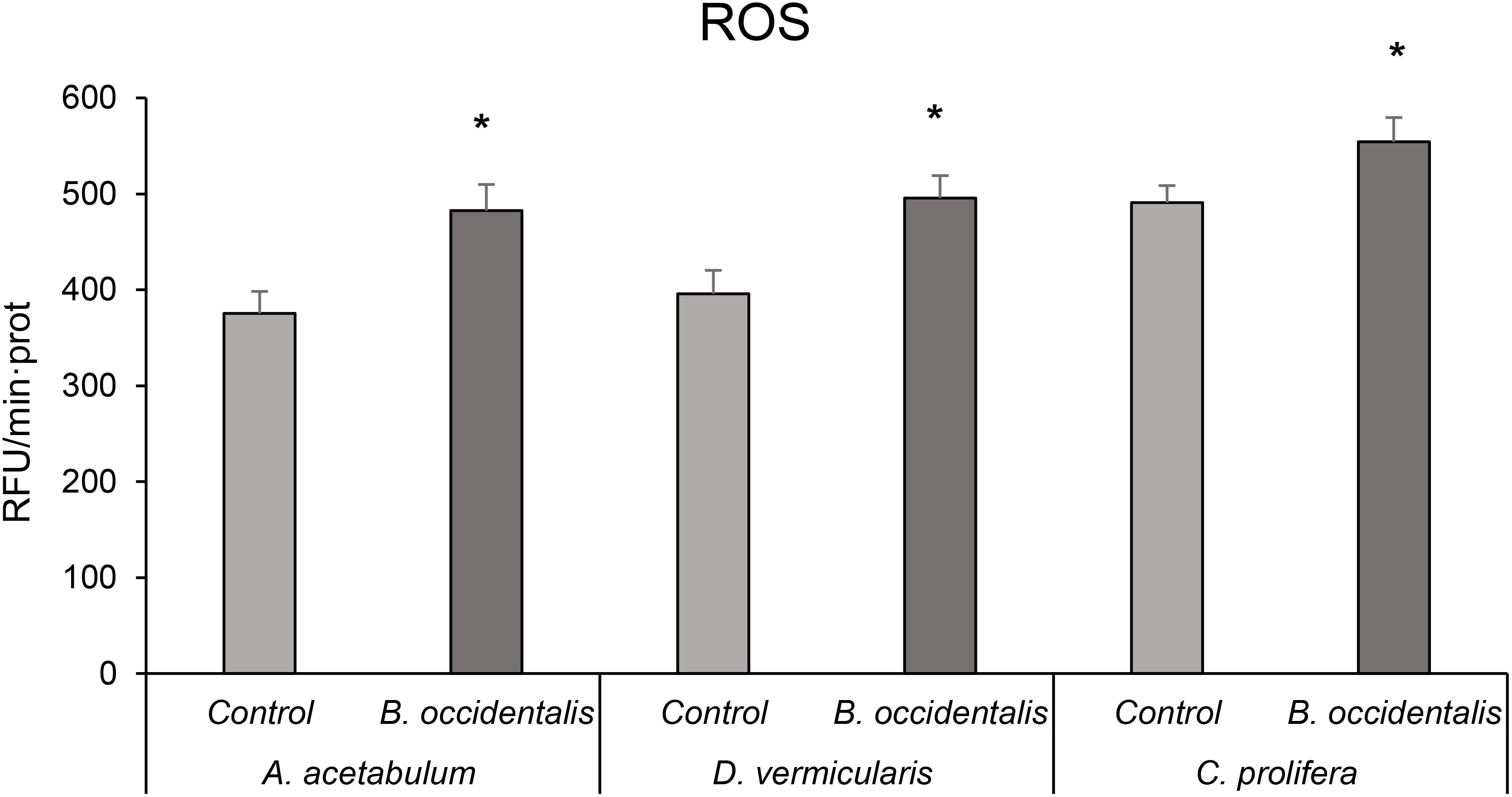
Figure 7. Reactive oxygen species (ROS) production in A. acetabulum, D. vermicularis and C. prolifera samples, alone (control situation) and in competition with the invasive alga B. occidentalis. * indicates statistically significant differences (p < 0.05) compared to the control. Values represent mean ± S.E.M.
Coastal lagoons are highly productive environments that support diverse habitats and biological communities. However, despite their environmental significance, their ecological balance is increasingly threatened by anthropogenic pressures, including tourism, agricultural practices, or the arrival of invasive species, which threaten the sustainability of their delicate equilibrium (Lacoste et al., 2023). The results of this work show the rapid expansion that B. occidentalis has undergone since its arrival in 2020, occupying practically the entire perimeter of the S’Estany des Peix lagoon and covering various native algae. Furthermore, in the three species of native algae studied (A. acetabulum, D. vermicularis and C. prolifera), the presence of B. occidentalis induced a situation of oxidative stress that was evidenced by an increase in antioxidant and detoxification enzymes.
B. occidentalis, native to the Gulf of Mexico, was observed for the first time at S’Estany des Peix in May 2020. However, this species had already been reported in other areas of the Mediterranean Sea such as in the Mar Menor lagoon (Spain) (Terradas-Fernández et al., 2022) and near Taranto (Ionian Sea, Italy) (Bottalico and Delle Foglie, 2003). In addition, B. occidentalis has also been reported in the western tropical and subtropical Atlantic (Dawes and Mathieson, 2008; Suárez et al., 2014; Wynne, 2017), the Canary Islands (Reyes et al., 1993) and the Mexican Pacific (Pedroche et al., 2005). The work by (Terradas-Fernández et al., 2022) suggests that the specimens from Mar Menor lagoon and those from Formentera present very similar morphological characters, so both locations could be related to the same introduction event. In fact, in the Mar Menor, clumps of Batophora were found floating and dispersed by waves and currents, so it could not be ruled out that some of them could have reached Formentera. In its native distribution area B. occidentalis grows on submerged roots of marine-coastal lagoons, inlets and mangroves, being highly resistant to desiccation and inhabits waters with a wide range of salinity and nutrients (Littler and Litter, 2000; Marcano et al., 2014). It can also grow in different habitats and substrates, such as pebbles, molluscs and artificial substrates (plates of PVC, ropes, etc.). Moreover, as in the present study, specimens form aggregates of 5-40 cm in diameter and are found in sandy shallow bottoms along with Cymodocea nodosa and C. prolifera meadows (Terradas-Fernández et al., 2022). In this sense, S’Estany des Peix is characterised as a semi-enclosed lagoon with high water renewal, minimal wave activity, and salinity levels similar to those of the sea, providing suitable conditions for this species to settle and expand. In 2022, there was a great expansion in the surface area occupied by B. occidentalis, probably favoured by the increase in water temperature that the surface waters of the Balearic Islands suffered that year. This anomaly temperature, in addition to being intense, was prolonged and persistent over time, as it lasted between the first days of May and the end of November, reaching a maximum of 29.2°C (on August 13, 2022, Agencia Estatal de Meteorología) (Informe sobre el estado del clima de España, 2023). On 94% of the days of the year, the Balearic Sea had an average temperature above the normal average.
Any environmental stress can affect the energy balance of an organism as a result of the additional energy needed to recover and maintain homeostasis (Sokolova et al., 2012). Consequently, the activation of the metabolism is associated in parallel with an increase in the generation of ROS. In this sense, competition between algae communities and other sessile organisms is known to be mainly related to physical competitions for light, substrate, and nutrients (Gennaro et al., 2015). Mechanisms of chemical competition among algae, such as allelopathy, have also been reported, involving in some cases ROS production and the activation of an antioxidant response (Gniazdowska et al., 2015; Del Monaco et al., 2017; Cerrato et al., 2023). The activation of antioxidant defences, in addition to being related to the increased metabolic activity, has been widely observed in algae subjected to several stressful abiotic factors such as pollutants, salinity, temperature, or physical damage (Dring, 2005), but also under biotic stress such as herbivory or competition (Sureda et al., 2008; Tejada and Sureda, 2014; Guzzetti et al., 2019).
In the present study, the presence of the invasive alga B. occidentalis was associated with a significant increase in ROS production and the activation of antioxidant mechanisms in the three species of epiphyted algae analysed- A. acetabulum, D. vermicularis and C. prolifera-. This response was evidenced by the increased activity of the antioxidant enzymes CAT, SOD and GRd. Furthermore, an increase in the activity of the phase II detoxification enzyme, GST, was observed, suggesting the potential presence of allelochemical compounds generated by the invader. When faced with a stressful environment, affected organisms activate their metabolism to respond and adapt to this new situation. This metabolic increase is associated with an increase in the production of ROS, mainly derived from mitochondrial activity to meet heightened energy demands. These reactive species, in addition to contributing to organismal defence through their oxidative action against pathogens and function as cellular redox messengers, can induce oxidative damage in the organism itself if not efficiently neutralised. In this sense, ROS have been reported to activate some transcription factors such as Nrf2, which regulates the synthesis of antioxidants and detoxification enzymes (Potin et al., 2002). Consistent with these results, the activation of antioxidant enzymes has been documented in various algae competing with non-native and potentially invasive species (Sureda et al., 2008; Tejada and Sureda, 2014; Sureda et al., 2017). For example, Cerrato et al. (2023) demonstrated that native P. pavonica assemblages exhibited increased antioxidant activities and ROS production when coexisting with the non-native algae Penicillus capitatus and H. incrassata. A similar pattern was previously observed in D. vermicularis when cohabiting with H. incrassata, showing an elevated antioxidant response with respect to isolated specimens (Sureda et al., 2017). This response has also been reported in invasive alga such as Caulerpa cylindracea Sonder, with the activation of antioxidant defences when interacting with native species, probably, contributing to its invasion success (Cavas and Yurdakoc, 2005; Sureda et al., 2017). Furthermore, the impact of invasive algae is not limited to plant organisms, since they have also been shown to induce biological stress and oxidative damage in animal species. A notable example is the bivalve Pinna nobilis, which, when colonised by the invasive red macroalga Lophocladia lallemandii (Box et al., 2009), exhibited increased GST activity, possibly related to the detoxification of ingested lophocladines produced by the invasive alga. In the present study, the significantly higher GST activity observed in the three species investigated and could be a response to toxic secondary metabolites released by the invasive B. occidentalis, consistent with previous studies, particularly in Caulerpa species (Sureda et al., 2006). The significant increase in GST activity observed in the three affected algal species could be linked to allelochemical interactions. However, since no specific allelochemicals have been identified in B. occidentalis to date, this remains speculative. However, members of the Dasycladales order, such as Ulva sp., have allelopathic compounds such as phenolic acids and coumarins that can alter competitive interactions by inhibiting the germination or development of algal and animal competitors (Green-Gavrielidis et al., 2018). Given this, future metabolomic and chemical ecology studies are necessary to confirm whether B. occidentalis produces bioactive compounds that contribute to its competitive success. Regarding MDA levels as an indicator of oxidative damage, higher values were observed in A. acetabulum and D. vermicularis, but not in C. prolifera. This difference could be due to C. prolifera being larger than the other two species, preventing complete coverage by B. occidentalis, thereby limiting the intensity of the disturbance. Moreover, it cannot be ruled out that C. prolifera physiologically presents a greater tolerance to oxidative stress, possibly due to more efficient antioxidant mechanisms or differences in its metabolic plasticity compared to the other two species of algae. Indeed, the increase in ROS production in C. prolifera was lower (about 13%) than in the other two algae (more than 25%), which could partially explain the absence of variations in MDA, and a lower degree of metabolic stress. For example, C. prolifera has been shown to significantly enhance its photosynthetic efficiency and production with increasing temperature while maintaining stable respiration rates, suggesting a great capacity to adjust to environmental stressors (Olivé et al., 2021). Similar results were observed in Posidonia oceanica, a large marine phanerogam, interacting with the invasive alga H. incrassata, where no significant oxidative damage was evident (Sureda et al., 2017), probably also because the invader only covers the lower areas of the plant. On the contrary, higher MDA values were observed in D. vermicularis when coexisting with H. incrassata.
While our biochemical data indicates significant increases in oxidative stress markers in native algae epiphytized by B. occidentalis, the ecological implications of these responses remain unclear. Sustained oxidative stress could lead to impaired photosynthetic efficiency, reduced growth rates, or even reproductive inhibition, ultimately affecting population dynamics and competitive interactions. Previous studies have shown that chronic oxidative stress in marine macroalgae can drive shifts in community structure, favouring species with higher resilience to environmental stressors (Cerrato et al., 2023; Duarte et al., 2018). Further research incorporating physiological parameters such as photosynthetic performance and growth rates will be crucial to determine whether B. occidentalis represents a long-term ecological threat to native algal communities
In conclusion, in the present study, we have reported the competitive interaction between the invasive Batophora occidentalis and three native green algae Acetabularia acetabulum, Dasycladus vermicularis and Caulerpa prolifera, in the lagoon S’Estany des Peix (Formentera Island) by means of antioxidant/oxidative biomarkers. Since its first observation in 2020, B. occidentalis has shown a strong expansion capacity, acting as a potential invader and inducing significant increases in all analysed enzymatic activities in native algae compared to non-affected specimens. Given the rapid spread of B. occidentalis, continuous monitoring of its invasion both inside and outside the lagoon is highly recommended. Regular monitoring would help track the spread and potential impacts to native species, in addition to public awareness campaigns to inform local boaters about the potential ecological harm further spread may cause. This monitoring, together with the evolution of the native green algae and the measurement of physiological parameters such as photosynthesis or growth rate, to determine whether B. occidentalis can become an introduced species of great concern in Formentera and other Balearic Islands. Although the current study did not include a quantitative assessment of B. occidentalis density at each sampling site, an interesting aspect for future research would be to examine the potential correlation between the extent of B. occidentalis coverage and the intensity of physiological stress responses in native algae.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
JL: Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. ST: Conceptualization, Investigation, Methodology, Writing – review & editing. MC: Investigation, Methodology, Writing – original draft, Writing – review & editing. VF: Investigation, Methodology, Writing – review & editing. LG: Conceptualization, Investigation, Methodology, Writing – review & editing. SP: Conceptualization, Funding acquisition, Investigation, Methodology, Writing – review & editing. AS: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This work has been funded by the Government of the Balearic Islands through the Direcció General d’Universitats, Recerca i Ensenyaments Artístics Superiors and the European Union -Next Generation EU (BIOPRO2024-004). Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency. Neither the European Union nor the granting authority can be held responsible for them. MC holds a postdoctoral contract from Juan de la Cierva-Formación (FJC2021-047606-I) financed by the Spanish State Agency (Agencia Estatal de Investigacion) and the Spanish Ministry of Science and Innovation (Ministero de Ciencia e Innovación, AEI/10.13039/501100011033) and the European Union (NextGenerationEU/PRTR). The present work was also supported by the Programme of Promotion of Biomedical Research and Health Sciences, Instituto de Salud Carlos III (CIBEROBN CB12/03/30038) and by the Spanish Ministry of Science, Innovation and Universities (MICIU), the Spanish State Research Agency (AEI/10.13039/501100011033) and the European Union (NextGenerationEU/PRTR) through the projects PID2020-117686RB-C33 and TED2021-131303B-I00.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1586402/full#supplementary-material
Aebi H. (1984). Catalase in vitro. Methods Enzymol. 105, 121–126. doi: 10.1016/S0076-6879(84)05016-3
Ballesteros E. (2020). On the presence of a species of Batophora J. Agardh 1854 (Chlorophyta: Dasycladales) in Formentera, Balearic Islands. Boll. Soc Hist. Nat. Balears. 63, 109–117.
Ballesteros E., Cebrian E., Alcoverro T. (2007). Mortality of shoots of Posidonia oceanica following meadow invasion by the red alga Lophocladia lallemandii. Bot. Mar. 50, 8–13. doi: 10.1515/BOT.2007.002
Ballesteros E., Pinedo S., Torras X. (2008). Cartografia detallada dels fons marins de s’Estany des Peix. Palma: Centre d’Estudis Avançats de Blanes i Govern de les Illes Balears 36.
Bartoskova M., Dobsikova R., Stancova V., Zivna D., Blahova J., Marsalek P., et al. (2013). Evaluation of ibuprofen toxicity for zebrafish (Danio rerio) targeting on selected biomarkers of oxidative stress. Neuroendocrinol. Let. 34, 102–108.
Bottalico A., Delle Foglie C. I. (2003). Contribution to the knowledge of the benthic marine flora along the eastern coastline of the Gulf of Taranto (Ionian Sea). Flora Mediterr. 13, 261–272.
Boudouresque C. F. (2008). Les espèces introduites et invasives en milieu marin. Troisième édition (Marseille: GIS Posidonie Publications), 1–201.
Boudouresque C. F., Verlaque M. (2002). Biological pollution in the Mediterranean Sea: Invasive versus introduced macrophytes. Mar. pollut. Bull. 44, 32–38. doi: 10.1016/S0025-326X(01)00150-3
Box A., Sureda A., Deudero S. (2009). Antioxidant response of the bivalve Pinna nobilis colonised by invasive red macroalgae Lophocladia lallemandii. Comp. Biochem. Physiol. Toxicol. Pharmacol. 149, 456–460. doi: 10.1016/j.cbpc.2008.10.107
Brine O., Hunt L., Costello M. J. (2013). Marine biofouling on recreational boats on swing moorings and berths. Manage. Biol. Invasion. 4, 327–341. doi: 10.3391/mbi.2013.4.4.07
Cavas L., Yurdakoc K. (2005). A comparative study: Assessment of the antioxidant system in the invasive green alga Caulerpa racemosa and some macrophytes from the Mediterranean. J. Exp. Mar. Biol. Ecol. 321, 35–41. doi: 10.1016/j.jembe.2004.12.035
Cecere E., Petrocelli A., Belmonte M., Portacci G., Rubino F. (2016). Activities and vectors responsible for the biological pollution in the Taranto Seas (Mediterranean Sea, southern Italy): a review. Environ. Sci. pollut. Res. 23, 12797–12810. doi: 10.1007/s11356-015-5056-8
Cerrato M., Mir-Rosselló P. M., Ferriol P., Gil L., Monserrat-Mesquida M., Tejada S., et al. (2023). Oxidative stress response in the seaweed Padina pavonica associated with the invasive Halimeda incrassata and Penicillus capitatus. Water. 15, 557. doi: 10.3390/w15030557
Del Monaco C., Hay M. E., Gartrell P., Mumby P. J., Diaz-Pulido G. (2017). ). Effects of ocean acidification on the potency of macroalgal allelopathy to a common coral. Sci. Rep. 7, 41053. doi: 10.1038/srep41053
Deudero S., Blanco A., Box A., Mateu-Vicens G., Cabanellas-Reboredo M., Sureda A. (2010). Interaction between the invasive macroalga Lophocladia lallemandii and the bryozoan Reteporella grimaldii at seagrass meadows: Density and physiological responses. Biol. Inv. 12, 41–52. doi: 10.1007/s10530-009-9428-1
Dring M. J. (2005). Stress resistance and disease resistance in seaweeds: the role of reactive oxygen metabolism. Adv. Bot. Res. 43, 175–207. doi: 10.1016/S0065-2296(05)43004-9
Duarte B., Martins I., Rosa R., Matos A. R., Roleda M. Y., Reusch T. B. H., et al. (2018). Climate change impacts on seagrass meadows and macroalgal forests: An integrative perspective on acclimation and adaptation potential. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00190
Faggio C., Pagano M., Alampi R., Vazzana I., Felice M. R. (2016). Cytotoxicity, haemolymphatic parameters, and oxidative stress following exposure to sub-lethal concentrations of quaternium-15 in Mytilus galloprovincialis. Aquat. Toxicol. 180, 258–265. doi: 10.1016/j.aquatox.2016.10.010
Flohé L., Ötting F. (1984). Superoxide Dismutase Assays. Superoxide Dismutase Assays. Methods Enzymol. 105, 93–104. doi: 10.1016/S0076-6879(84)05013-8
Forteza V., Castelló M., Ballesteros E. (2024). Invasion of batophora J. Agard(Chlorophyta: dasycladales) in formentera, balearic islands. Boll. Soc Hist. Nat. Balears. 67, 67–74.
Fortič A., Rasheed R. A. S., Almajid Z., Badreddine A., Báez J. C., Belmonte-Gallegos Á., et al. (2023). New records of introduced species in the Mediterranean Sea (April 2023). Mediterr. Mar. Sci. 24, 182–202. doi: 10.12681/MMS.34016
Galil B. S. (2000). A sea under siege - Alien species in the Mediterranean. Biol. Inv. 2, 177–186. doi: 10.1023/A:1010057010476
Gennaro P., Piazzi L., Persia E., Porrello S. (2015). Nutrient exploitation and competition strategies of the invasive seaweed Caulerpa cylindracea. Eur. J. Phycol. 50, 384–394. doi: 10.1080/09670262.2015.1055591
Gniazdowska A., Krasuska U., Andrzejczak O., Soltys D. (2015)Allelopathic compounds as oxidative stress agents: yes or NO, in Reactive oxygen and nitrogen species signaling and communication in plants. Eds. Gupta K., Igamberdiev A. (Springer, Cham), 155–176.
Goldberg D. M., Spooner R. J. (1983). Glutathione reductase”, in: Methods of Enzymatic Analysis. 3rd ed. Eds. Bergmeyer H. U., Bergmeyer J., GraBI M. (Weinheim: Verlag Chemie), 258–265.
Gollasch S., Hewitt C. L., Bailey S., David M. (2019). Introductions and transfers of species by ballast water in the Adriatic Sea. Mar. pollut. Bull. 147, 8–15. doi: 10.1016/j.marpolbul.2018.08.054
Green-Gavrielidis L. A., MacKechnie F., Thornber C. S., Gomez-Chiarri M. (2018). Bloom-forming macroalgae (Ulva spp.) inhibit the growth of co-occurring macroalgae and decrease eastern oyster larval survival. Mar. Ecol. Prog. Ser. 595, 27–37. doi: 10.3354/meps12556
Guzzetti E., Salabery E., Ferriol P., Díaz J. A., Tejada S., Faggio C., et al. (2019). Oxidative stress induction by the invasive sponge Paraleucilla magna growing on Peyssonnelia squamaria algae. Mar. Environ. Res. 150, 104763. doi: 10.1016/j.marenvres.2019.104763
Habig W. H., Pabst M. J., Jakoby W. B. (1974). Glutathione S transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139. doi: 10.1016/S0021-9258(19)42083-8
Hartmann A., Ganzera M., Karsten U., Skhirtladze A., Stuppner H. (2018). Phytochemical and analytical characterization of novel sulfated coumarins in the marine green macroalga dasycladus vermicularis (Scopoli) krasser. Molecules 23, 2735. doi: 10.3390/molecules23112735
Hewitt C. L., Campbell M. L., Schaffelke B. (2007). Introductions seaweeds: Accidental transfer pathways mechanisms. Introductions seaweeds: Accidental transfer pathways mechanisms. Bot. Mar. 50, 326–337. doi: 10.1515/BOT.2007.038
Informe sobre el estado del clima de España (2023).Evolución de la temperatura superficial del agua del mar)(Madrid, Spain: Agencia Estatal de Meteorología). Available at: https://aemetblog.es/2023/06/15/informe-sobre-el-estado-del-clima-de-espana-2022evolucion-de-la-temperatura-superficial-del-agua-del-mar/. (Accessed February 25, 2025)
Invasive Alien Species and Climate ChangeInternational union for conservation of nature (IUCIN). (2021). Available online at: https://www.iucn.org/resources/annual-reports/iucn-2021-international-union-conservation-nature-annual-report. (Accessed February 25, 2025)
Katsanevakis S., Wallentinus I., Zenetos A., Leppäkoski E., Çinar M. E., Oztürk B., et al. (2014). Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Inv. 9, 391–423. doi: 10.3391/ai.2014.9.4.01
Katsanevakis S., Zenetos A., Belchior C., Cardoso A. C. (2013). Invading European Seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manage. 76, 64–74. doi: 10.1016/j.ocecoaman.2013.02.024
Lacoste É., Jones A., Callier M., Klein J., Lagarde F., Derolez V. (2023). A review of knowledge on the impacts of multiple anthropogenic pressures on the soft-bottom benthic ecosystem in Mediterranean coastal lagoons. Estuaries Coasts. 46, 2190–2207. doi: 10.1007/s12237-023-01188-9
Leonardi P. I., Miravalles A. B., Faugeron S., Flores V., Beltrán J., Correa J. A. (2006). Diversity, phenomenology and epidemiology of epiphytism in farmed Gracilaria Chilensis (Rhodophyta) in northern Chile. Eur. J. Phycol. 41, 247–257. doi: 10.1080/09670260600645659
Littler D. M., Litter M. M. (2000). Caribbean reef plants: An identification guide to the reef plants of the Caribbean, Bahamas, Florida and Gulf of México (Washington, DC: Offshore graphics, Inc).
Mancuso F. P., D’Agostaro R., Milazzo M., Badalamenti F., Musco L., Mikac B., et al. (2022). The invasive seaweed Asparagopsis taxiformis erodes the habitat structure and biodiversity of native algal forests in the Mediterranean Sea. Mar. Environ. Res. 173, 105515. doi: 10.1016/j.marenvres.2021.105515
Marcano A., Rodríguez-Reyes J. C., López D. (2014). Composición y estructura de la vegetación en un humedal dulceacuícola del monumento natural Laguna de las Marites, estado Nueva Esparta, Venezuela. Ciencia. 22, 90–104.
Olivé I., Varela-Álvarez E., Silva J., Serrão E. A., Santos R. (2021). Physiological potential of the chlorophyte Caulerpa prolifera for proliferation across the Mediterranean-Atlantic basins in a warmer ocean. Mar. Ecol. Prog. Ser. 668, 73–84. doi: 10.3354/meps13719
Pedroche F. F., Silva P. C., Aguilar Rosas L. E., Dreckmann K. M., Aguilar Rosas R., Universidad Metropolitana, et al. (2005). Catálogo de las algas marinas bentónicas del Pacífico de México I. Chlorophycota Vol. Vol. 129) (Mexicali, México: Universidad Autónoma de Baja California).
Pergent G., Boudouresque C. F., Dumay O., Pergent-Martini C., Wyllie-Echeverria S. (2008). Competition between the invasive macrophyte Caulerpa taxifolia and the seagrass Posidonia oceanica: Contrasting strategies. BMC Ecol. 8, 20. doi: 10.1186/1472-6785-8-20
Poloczanska E. S., Burrows M. T., Brown C. J., García Molinos J., Halpern B. S., Hoegh-Guldberg O., et al. (2016). Responses of marine organisms to climate change across oceans. Front. Mar. Sci. 3,62. doi: 10.3389/fmars.2016.00062
Potin P., Bouarab K., Salaün J. P., Pohnert G., Kloareg B. (2002). Biotic interactions of marine algae. Curr. Opin. Plant Biol. 5, 308–317. doi: 10.1016/S1369-5266(02)00273-X
Quetglas-Llabrés M. M., Tejada S., Capó X., Langley E., Sureda A., Box A. (2020). Antioxidant response of the sea urchin Paracentrotus lividus to pollution and the invasive algae Lophocladia lallemandii. Chemosphere. 261, 127773. doi: 10.1016/j.chemosphere.2020.127773
Reyes J., Sanson M., Afonso-Carrillo J. (1993). Notes on some interesting marine algae new from the Canary Islands. Cryptogamic Botany. 4, 50–59.
Rezayian M., Niknam V., Ebrahimzadeh H. (2019). Oxidative damage and antioxidative system in algae. Toxicol. Rep. 6, 1309–1313. doi: 10.1016/j.toxrep.2019.10.001
Ross C., Küpper F. C., Jacobs R. S. (2006). Involvement of reactive oxygen species and reactive nitrogen species in the wound response of Dasycladus vermicularis. Chem. Biol. 13, 353–364. doi: 10.1016/j.chembiol.2006.01.009
Schaffelke B., Smith J. E., Hewitt C. L. (2006). Introduced macroalgae - A growing concern. J. Appl. Phycol. 18, 529–541. doi: 10.1007/s10811-006-9074-2
Sokolova I. M., Frederich M., Bagwe R., Lannig G., Sukhotin A. A. (2012). Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 79, 1–15. doi: 10.1016/j.marenvres.2012.04.003
Suárez A. M., Martínez-Daranas B., Alfonso Y. (2014). Macroalgas marinas de Cuba (La Habana, Cuba: Editorial Universidad de la Habana).
Sureda A., Box A., Enseñat M., Alou E., Tauler P., Deudero S., et al. (2006). ). Enzymatic antioxidant response of a labrid fish (Coris julis) liver to environmental caulerpenyne. Comp. Biochem. Physiol. Toxicol. Pharmacol. 144, 191–196. doi: 10.1016/j.cbpc.2006.08.001
Sureda A., Box A., Terrados J., Deudero S., Pons A. (2008). Antioxidant response of the seagrass Posidonia oceanica when epiphytized by the invasive macroalgae Lophocladia lallemandii. Mar. Environ. Res. 66, 359–363. doi: 10.1016/j.marenvres.2008.05.009
Sureda A., Tejada S., Capó X., Melià C., Ferriol P., Pinya S., et al. (2017). Oxidative stress response in the seagrass Posidonia oceanica and the seaweed Dasycladus vermicularis associated to the invasive tropical green seaweed Halimeda incrassata. Sci. Total Environ. 601-602, 918–925. doi: 10.1016/j.scitotenv.2017.05.261
Taylor A., Grillas P., Al Hreisha H., Balkız Ö., Borie M., Boutron O., et al. (2021). The future for Mediterranean wetlands: 50 key issues and 50 important conservation research questions. Reg. Environ. Change 21, 33. doi: 10.1007/s10113-020-01743-1
Tejada S., Sureda A. (2014). Antioxidant response of the brown algae Dictyota dichotoma epiphyted by the invasive red macroalgae Lophocladia lallemandii. J. Coast. Life Med. 2, 362–366. doi: 10.12980/JCLM.2.201414J5
Terradas-Fernández M., Valverde-Urrea M., López-Moya F., Fernández-Torquemada Y. (2022). On the presence and ubiquity of the exotic Batophora (J. Agardh) in the Mar Menor Lagoon (SE Spain). Water. 14, 2909. doi: 10.3390/w14182909
Tomas F., Box A., Terrados J. (2011). Effects of invasive seaweeds on feeding preference and performance of a keystone Mediterranean herbivore. Biol. Inv. 13, 1559–1570. doi: 10.1007/s10530-010-9913-6
Tsirintanis K., Azzurro E., Crocetta F., Dimiza M., Froglia C., Gerovasileiou V., et al. (2022). Bioinvasion impacts on biodiversity, ecosystem services, and human health in the Mediterranean Sea. Aquat. Inv. 17, 308–352. doi: 10.3391/ai.2022.17.3.01
Walther G. R., Post E., Convey P., Menzel A., Parmesan C., Beebee T. J. C., et al. (2002). ). Ecological responses to recent climate change. Nature. 416, 389–395. doi: 10.1038/416389a
Wesselmann M., Hendriks I. E., Johnson M., Jordà G., Mineur F., Marbà N. (2024). Increasing spread rates of tropical non-native macrophytes in the Mediterranean Sea. Global Change Biol. 30, e17249. doi: 10.1111/gcb.17249
Williams S. L., Smith J. E. (2007). ). A global review of the distribution, taxonomy, and impacts of introduced seaweeds. Annu. Rev. Ecol. Evol. Syst. 38, 327–359. doi: 10.1146/annurev.ecolsys.38.091206.095543
Keywords: Batophora occidentalis, invasive species, algae, oxidative stress, antioxidant enzymes, Formentera Island
Citation: Lombardo J, Tejada S, Compa M, Forteza V, Gil L, Pinya S and Sureda A (2025) Oxidative stress response in native algae exposed to the invasive species Batophora occidentalis in S’Estany des Peix, Formentera (Balearic Islands). Front. Mar. Sci. 12:1586402. doi: 10.3389/fmars.2025.1586402
Received: 02 March 2025; Accepted: 25 March 2025;
Published: 15 April 2025.
Edited by:
Jorge L. Gutiérrez, National Scientific and Technical Research Council (CONICET), ArgentinaReviewed by:
Sara Usandizaga Solana, Pontificia Universidad Católica de Chile, ChileCopyright © 2025 Lombardo, Tejada, Compa, Forteza, Gil, Pinya and Sureda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antoni Sureda, YW50b25pLnN1cmVkYUB1aWIuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.