 Aratrika Ray
Aratrika Ray Sandipan Mondal1,2
Sandipan Mondal1,2 Ming-An Lee
Ming-An Lee Yi-Chen Wang
Yi-Chen Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 05 March 2025
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1526762
Introduction: Prior research emphasizes the beneficiaries and detractors in fisheries due to climate change, focusing on alterations in biomass, species mix, and potential yields. Comprehending the potential impact of climate change on the fisheries income of maritime nations is an essential subsequent step in formulating effective socio-economic policies and food sustainability plans to mitigate and adapt to climate change. Globally, our understanding of small-scale fisheries remains inadequate, despite their significance for food security and livelihoods.
Methods: This study demonstrates by using global circulation model (GCM: IPSL) that moonfish catches react favorably to climate alterations in southwest Taiwan, through the analysis of primary fishery data from 2014-2020 spanning two seasons – northeast and southwest monsoons by using generalized additive models. The anticipated habitat shifts were utilized to examine the impact of future environmental changes on moonfish catch rates.
Result: By the 2060s, moonfish emerged as a winner of climate change in this study region. The habitats of moonfish are projected to shift westward during southwest monsoon, with a mean habitat centroid displacement of about 50-150 Km between RCP 2.6 and 8.5 during NE monsoon while 20-40 Km during the SW monsoon. A possible explanation could be that the Taiwan Strait (TS) may serve as a geographical barrier to the northward migration of south-dwelling moonfish. The study further demonstrates that moonfish fisheries catch rates may increase by 22.5% and 17.2% beyond the present catch rates by the 2060s under elevated CO2 emission scenarios during northeast and southwest monsoons respectively.
Discussion: Our findings indicate the necessity for comprehensive economic evaluations regarding the potential impacts of climate change on regional small-scale marine fisheries, contributing to the adaptive conservation and management of fish habitats.
Climate change has significantly and adversely impacted the global environment, biodiversity, and sustainable human development, primarily by modifying global temperature patterns, the hydrological cycle, and inducing acidification (Habib et al., 2025). The primary response variables (e.g., physical, chemical, and biological) in oceans can serve as sentinel indicators of climate change impacts. The anticipated extinction rates of aquatic biodiversity are typically greater than those of terrestrial species in both contemporary and forthcoming climate change scenarios (Huang et al., 2021).
Small-scale fisheries (SSFs) significantly contribute to food security, poverty alleviation, employment, and the maintenance of healthy marine ecosystems (Gatta, 2022), hence facilitating the attainment of certain sustainable development objectives. Despite being a primary livelihood option for millions globally, SSFs encounter escalating uncertainty and variability associated with globalization, climate change, and overfishing (Nilsson et al., 2019). Climate variability poses a significant danger to SSFs by impacting fishing resources, the livelihoods of fishermen, and altering fish populations and production value per fisherman (Mbaye et al., 2023). Coastal regions are especially vulnerable to the detrimental impacts of global warming, primarily seen in the convergence of terrestrial and marine factors. The repercussions may be oceanic, ecological, or socio-economic. Oceanographic implications include alterations in fishing seasons, shifts in fishing locations, and heightened dangers associated with maritime activities due to elevated wave heights and turbulent winds (N’Souvi et al., 2024). Simultaneously, the unpredictability of fishermen’s earnings and the potential biodiversity losses resulting from impending climate change (Pörtner et al., 2023) exemplify socio-economic and ecological repercussions, respectively.
Additional consequences of climate change include alterations in coastal water temperature, precipitation patterns, sea level rise, coastal flooding, and erosion, which significantly influence fish diversity, distribution, and abundance, subsequently affecting marine biota and ecosystems, as well as fluctuations in fish populations (N’Souvi et al., 2024). For example, sea level rise impacts the coastal landscape and the livelihoods of communities by diminishing the productivity and value of fisheries (N’Souvi et al., 2024), hence compromising the safety and efficiency of fishing operations (Bertrand et al., 2019). Moreover, alterations in precipitation, storm occurrences, and drought patterns influence water flow rates, thereby affecting species movement and recruitment patterns as well as salinity levels in coastal regions (Trégarot et al., 2024). Consequently, the accelerated rise in sea temperature (Cheng et al., 2019), salinity (Cheng et al., 2020), sea level (Kulp and Strauss, 2019), acidification (Cattano et al., 2018), and deoxygenation (Kwiatkowski et al., 2020), along with alterations in species distribution, a general poleward migration of both marine and terrestrial species (Venegas et al., 2023), a decrease in abundance (McCauley et al., 2015), and shifts in productivity (Venegas et al., 2023), result in socioeconomic repercussions on fisheries by modifying seasonal patterns and diminishing fishing yields and fishers’ income (Franco et al., 2020). Ocean warming has predominantly intensified extraction patterns, resulting in diminishing fisheries, with rare examples where elevated temperatures have enhanced fishing yields (Free et al., 2019). The intensity and duration of these responses are frequently species-specific and influenced by habitat preferences (Champion and Coleman, 2021) and the nutritional adaptability of marine species (Monaco et al., 2020). A recent study (Hayashi et al., 2021) demonstrated that the unprecedented sea surface temperature (SST) in the northern Pacific in 2020 was unlikely to have occurred in the absence of anthropogenic climate change. The exceedingly heated conditions are expected to establish a new climatic norm by the mid-21st century, necessitating the immediate execution of adaption strategies for anthropogenic global warming. Moreover, the worldwide sea level has risen simultaneously (Nerem et al., 2018), and global ocean kinetic energy has markedly grown since the early 1990s (Hu et al., 2020). Various long-term ocean reanalysis agree that the upper 700-meter ocean heat content has risen in the Northwest Pacific (NWP), North Atlantic, and Southern Oceans since 1970 (Palmer et al., 2017). Consequently, long-term alterations in the world’s oceans up to the current climate are evident in observations. The effects of climate change on marine species in the NWP are still inadequately documented. The information deficit is concerning, as this region is among the most rapidly warming areas of the world ocean (Palmer et al., 2017) and harbors remarkable biodiversity of marine species endangered by climate change (Asch et al., 2018). Moreover, prolonged periods of intensive fishing have led to the depletion of numerous stocks to unsustainable biological levels in different parts of the Pacific. Furthermore, due to the absence of systematic stock assessments or regionally coordinated management for most exploited species, allegations of unsustainable fishing may be understated (FAO, 2022). The interconnected issues of climate change and unsustainable fisheries are concerning, as they may compromise species’ adaptations to present and forthcoming environmental alterations, notwithstanding management initiatives aimed at stock restoration. Therefore, it is imperative to identify which fishery resources exhibit more resilience or sensitivity to climate change for effective management in this region. The Southwestern waters off Taiwan are acknowledged as significantly susceptible to the effects of climate change (Lee et al., 2023), however limited research has documented the understanding of climate change and its perceived effects on moonfish in this region. As the marine environment becomes warmer, more acidic, and oxygen-depleted, the effects on fish populations are significant, with certain stocks relocating to maintain optimal temperature conditions, while others that are less mobile or adaptable are experiencing decline (Barange et al., 2018).
Moonfish (Mene maculata), a coastal forage fish in the Indo-Pacific, is a significant commercial fish in Taiwan, inhabiting depths of 20-100 m concentrated in the Southwestern waters, principally caught by Taiwanese purse-seine fisheries. Moonfish, as a small forage fish, is essential in the energy transmission to apex predators such as marlins, dolphinfish, and tunas (Ray et al., 2024). Coastal fish species such as moonfish has a high sensitivity to environmental changes, exacerbated by climate change, that can trigger a cascade of effects throughout the food web. Their biomass at intermediate trophic levels implement wasp-waist control (Albo Puigserver et al., 2018) that contributes to their catchability, which is influenced by ecosystem components like food availability, feeding behavior, habitat, and climatic changes. The apex of moonfish production was attained during the 1980s, wherein the annual production exceeded 16000 Kg (Supplementary Figure S1), but further experiencing a decline to mere 1000 Kg in 2020 as a consequence of World War II. This decline in moonfish production may have been due to the emergence of illicit fishing vessels and alterations in the marine ecosystem (Liao et al., 2019). Between 1991 and 2008, approximately 30% of large-scale longline fishing vessels and 18% of coastal fishing vessels underwent scrappage. Moreover, there has been a notable shift in the assortment of fish species. Nevertheless, the management of forage fish stocks like moonfish is challenged by their propensity to react significantly to environmental variability, resulting in fast changes in distribution or abundance (Murray et al., 2019) however, comparisons are limited. To improve their stock management, it is crucial to acknowledge the probable relationship between regional climate change and moonfish landings.
This study seeks to investigate how the catch rates of moonfish respond to climate change scenarios in short, medium, and long term (2034-2040, 2044-2050, and 2054-2060 respectively) under RCP 2.6, 4.5, and 8.5; addressing existing gaps in the current understanding by offering a seasonally explicit, predictive, and spatially resolved examination of moonfish in Southwestern Taiwan (SWT). This investigation focused on the understanding of climate-related and oceanic effects between two seasons – the northeast (NE) and southwest (SW) monsoon to unravel the impact of changing oceanographic features on the overall annual moonfish catch rate by incorporation of generalized additive models (GAMs) (Figure 1) in contrast to prior research that generically analyzes climate-induced changes in fisheries. Furthermore, it discusses the function of TS as a possible impediment to northward migration, a variable seldom addressed in prior moonfish research. The research reveals important environmental determinants impacting moonfish distribution and abundance by combining several oceanographic variables, including sea surface temperature (SST), salinity, sea surface height (SSH), mixed layer depth (MLD), and chlorophyll content. The objectives of this study were threefold: (i) to evaluate potential shifts in moonfish distribution resulting from changes in environmental conditions; (ii) What alterations will occur in habitat suitability over the SWT shelf for moonfish? and (iii) Will moonfish be ‘winners’ (experiencing an increase in acceptable habitat or catch rates) or ‘losers’ (experiencing a reduction in suitable habitat or catch rates) due to forthcoming climate change? The results of this study are anticipated to furnish stakeholders with a foundation for formulating adaptation strategies that correspond to the needs, capabilities, and interests of SSFs targeting moonfish in Taiwan to promote sustainable development targeting the FAO Blue Growth Initiative (BGI) and the United Nations Sustainable Development Goals (SDG). Since Taiwanese purse-seiners form the major fishing gear of this species in Taiwan contributing to 98% of fish catch (Ray et al., 2024), only the fishery data from this gear has been used in the present study from January 2014 to December 2020 focusing on the monsoons. Considering this fact, it is crucial to comprehend their susceptibility to climate variability in order to formulate adaptation strategies that conform to the FAO’s advocacy for climate-smart fisheries (FAO, 2022).
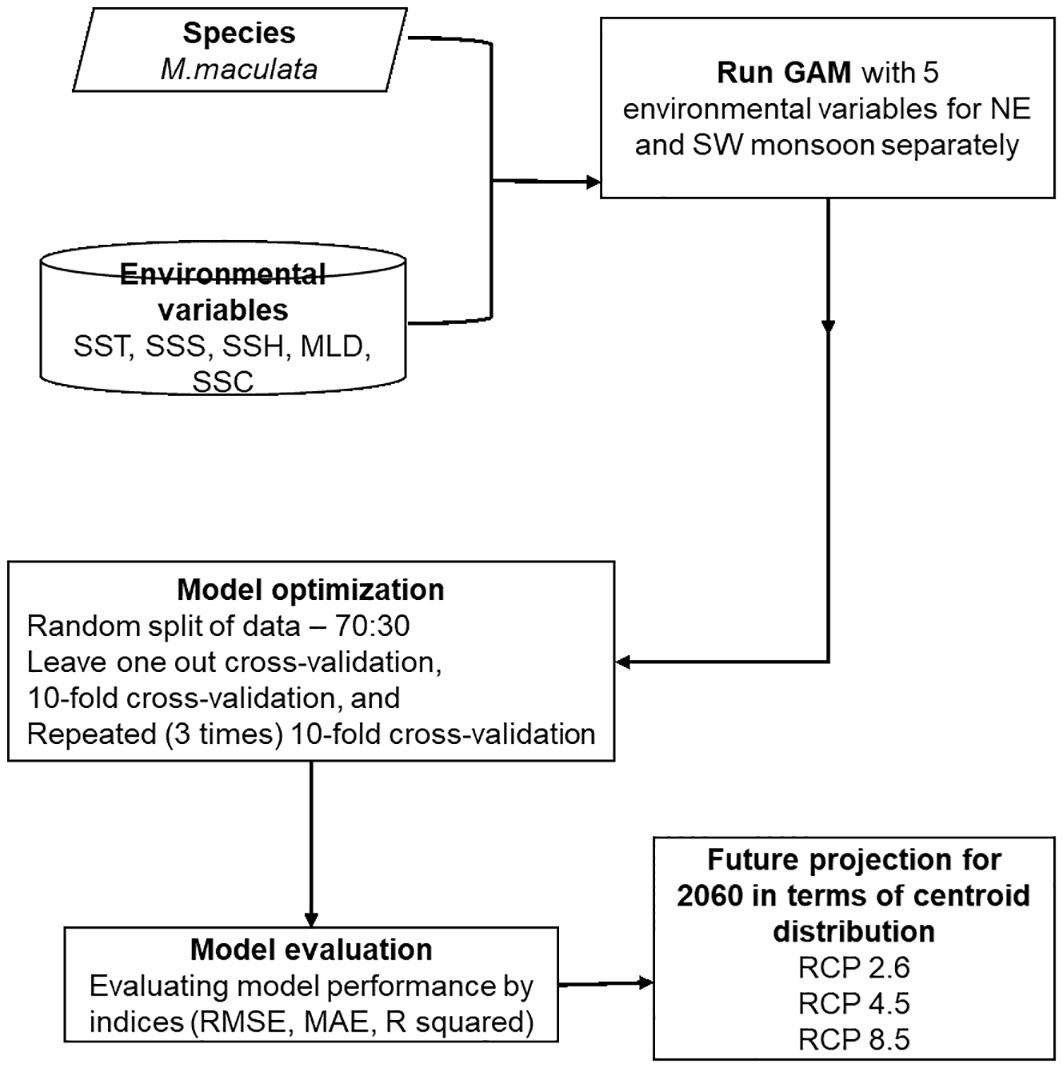
Figure 1. The flowchart of methodology used in this study.
A combined data on the moonfish fishery were gathered from both purse seine fishing operations, specifically those involving coastal water fishing with boats under 100 tons in gross register tonnage and less than 24 meters in length, and voyage data recorders (VDRs) from January to December between 2014 and 2020; sourced from the Taiwan Fishery Agency. The collected data covered a spatial range from 21°NS to 25°N and from 117°E to 121°E, with a spatial resolution of 0.1° × 0.1°. The data gathered encompassed the area coordinates, fishing effort (number of fishing hours), fishing dates, total catch weight (kg), and vessel tonnage. The fishery data was separated into NE monsoon data (October – April) and SW monsoon data (May – September) for the entire analysis. The total catch was calculated using the subsequent formula according to Mondal et al. (2024):
The monthly nominal catch per unit effort (N-CPUE) was subsequently calculated as follows (Mondal et al., 2024):
Since 1978, multi-satellite remote sensing data have been utilized to acquire images of ocean surface temperature, thermal and chlorophyll-a fronts, and phytoplankton pigment concentration, all of which have proven beneficial in fisheries management, fisheries oceanography, and operational fisheries oceanography (Vayghan et al., 2020) to obtain knowledge of fish habitats (Lan et al., 2018). Environmental parameters, including SST, dissolved oxygen concentration, salinity, and chlorophyll are frequently utilized in predictive models for marine fish populations (Zhang et al., 2020a). In this study, data concerning five oceanographic parameters were collected: SST, SSH, sea surface salinity (SSS), MLD, and sea surface chlorophyll (SSC) (Table 1). The spatial resolution of the data related to fishing activities conducted between 21°NS to 25°N and 117°E to 121°E exhibited variability. The data collected relates to fishing activities conducted from January to December during the years 2014 to 2020. Due to the spatial resolution of the fishery data being 0.1° × 0.1°, Matlab version 2019a was utilized to interpolate the collected oceanographic data to a 0.1° × 0.1° grid, ensuring alignment with the spatial coverage of the fishery data.
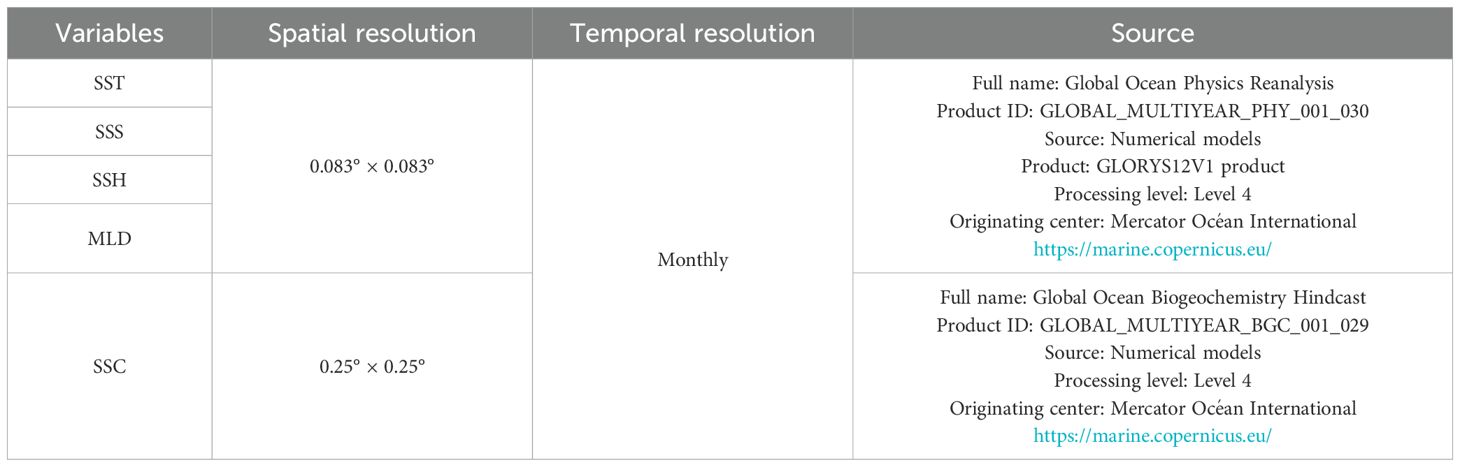
Table 1. Oceanographic parameters analysed in the study.
Future scenarios for the 2060s involve four representative concentration pathways (RCPs) predicted in the Coupled Model Intercomparison Project Phase 6 (CMIP6), which illustrate potential future emission trajectories related to the greenhouse effect (Basheer et al., 2016). CMIP6 seeks to assess the climate system and forecast regional climate changes under future scenarios through the collection and comparison of simulation results from multiple global circulation models (GCMs). The RCPs were established based on the projected radiative forcing target level for 2100. Researchers collaborated globally to predict future climate and environmental conditions based on varying radiative levels, resulting in a comprehensive data set. We selected the most optimistic (2.6), moderate (4.5) and pessimistic (8.5) RCPs to predict and compare moonfish dispersal in the 2060s between the two monsoons. Long-term environmental changes were predicted from GCM, specifically IPSL (Institute Pierre Simon Laplace) (Schickele et al., 2021). The data were subsequently resampled to a resolution of 0.1° × 0.1°, aligning with the current environmental dataset, through spatial interpolation. This study analyzed projection data from January to December for the years 2034 - 2040, 2044 - 2050, and 2054 - 2060, respectively focusing on the short-, medium-, and long-term impacts of climate-related changes on the distribution patterns of moonfish during NE and SW monsoon.
To reduce the impact of spatial (latitude, longitude) and temporal (year, month) variables, the N-CPUE data were standardized using generalized additive models (GAMs), a method commonly employed to derive filtered and unbiased standardized catch per unit effort (S-CPUE) values. GAMs, which are progressively used in ecology to discern nonlinear functional linkages (Pedersen et al., 2019), provide a method for achieving this decomposition of temporal variation into components that describe trend, seasonality, and other cyclical variations. GAMs may be succinctly characterized as modified generalized linear models (GLMs) where the linear predictor incorporates a summation of smooth functions that depict the functional connections between variables and the response variable:
Where E(Y) is the conditional expectation of a response derived from an exponential family distribution, β0 is the unknown intercept, the si’s are a collection of smooth functions pertaining to one or many predictor variables (x’s), and the link function (g) is monotonic. Every smooth function si is constituted of basis functions, whose coefficients, requiring estimation, serve as weights to regulate the form of the function. The overall quantity of basis functions constrains the possible complexity of the smooth function, with an increased collection of basis functions permitting more flexibility. Beyond their capacity to depict intricate and nonlinear ecological relationships, GAMs offer several additional benefits, including the ability to model various response families that address ecological characteristics (such as zero inflation) and the option to incorporate hierarchical smoothing for multivariate responses (Pedersen et al., 2019). Following the above equation, the standardization model was structured as below:
The S.CPUE results obtained were utilized in subsequent analyses. Predictor variables included year, month, vessel identifier (CT), latitude, longitude, and NPGO, while N-CPUE served as the targeting variable. The interactions employed included latitude × longitude, year × longitude, and month × longitude. NPGO served as an environmental predictor for moonfish CPUE, as indicated by the findings of Ray et al. (2024), where NPGO was seen to have the highest correlation with moonfish catch rate among all other climatic oscillations employed. Additionally, random splitting strategies, particularly at a ratio of 70:30, were employed to train and evaluate data for the purpose of establishing a standardization model (Vrigazova, 2021).
Elevated multicollinearity across predictive variables may negatively impact the precision of model predictions and the assessment of variable effects (Gregorich et al., 2021). To mitigate the problem of elevated multicollinearity, we evaluated five potential parameters using variance inflation factor (VIF) test. A score approaching 5 or more denotes a more robust positive collineairity, while a value < 5 implies a more no collinearity between the two variables (Kim, 2019). A VIF threshold < 5 was therefore used to identify potential components for inclusion in the final model iterations.
GAMs were utilized for species distribution modeling with the chosen environmental variables. GAMs provide the versatility to ascertain if a certain parameter is a significant element of the habitat connection and to identify its functional form. Habitat preferences, both regional and temporal, were examined throughout two monsoon seasons. The S-CPUE data for moonfish from the NE and SW monsoons were aggregated separately. The “gam” function from the mgcv package was used to create models in R software (version 4.2.3; R Foundation for Statistical Computing, Vienna, Austria). S-CPUE served as the response variable, whereas location (longitude and latitude) and environmental parameters (SST, SSD, MLD, SSS, and SSC) functioned as predictor variables. All data were used to create foundational models, and no interactions were included. The following steps were used to develop GAMs: assessing the relevance of each individual component, optimizing the model structure and evaluating residuals, and fitting the final response curves. The primary objective of the first stage was to validate the importance of the essential oceanographic variables identified by GAM. The last phase included constructing GAMs including numerous important components while accounting for the impact of interaction terms. We computed the effective degrees of freedom for each principal factor, supposing that all covariates were continuous. A statistically significant association was identified between S-CPUE and each predictor at p < 0.05. The model with the optimal conformation for goodness of fit (GoF) was chosen using a systematic procedure that prioritized the minimal root mean square error (RMSE), mean absolute error (MAE), and generalized cross-validation (GCV). The last phase included fitting the response curves for each smoothing term of the GAMs to ascertain the link between S-CPUE and the essential oceanographic parameters, as well as to identify the suitable range of these factors.
Smoothing spline regression was employed to analyze the relationships between the relative abundance of moonfish and their oceanographic preferences (Haghi Vayghan et al., 2013) as well as the projected scenarios under the RCPs 2.6 and 8.5. Abundance of moonfish served as the dependent variable in the regression analysis, while all chosen oceanographic parameters functioned as explanatory variables. The suitability index (SI) curve for moonfish was derived by normalization using the specified formula (Lee et al., 2020):
where Ymax and Ymin represent the maximum and minimum values of S-CPUE or predicted CPUE (P-CPUE) or oceanographic variable observations, respectively, and Y denotes the simulated (predicted) value within the range of Ymax to Ymin; SI values are constrained between 0 and 1.
SI values were derived from the summed frequency distribution of the S-CPUE or P-CPUE for each class, with the assumption that SI values range from 0 to 1. The midpoint of each class interval for the environmental variables served as the observed value in the fitting of the SI model. Finally, the relationships between the SI and oceanographic variable results were established using the following formula (Lee et al., 2019):
In this equation, m represents the response variable (oceanographic variables), while α and β are determined by applying the nonlinear least squares estimate to minimize the residual between a SI observation and the SI function.
A suitable habitat is an environment that fulfills the essential requirements for an organism or species to survive, reproduce, and develop. The suitability of a habitat is assessed based on its temperature, humidity, food availability, shelter availability, and other essential resources necessary for an organism’s survival. All variable combinations were evaluated, with the optimal model identified by the lowest Akaike’s Information Criterion (AIC), root mean square error (RMSE), mean absolute error (MAE), and generalized cross-validation (GCV) scores; individually for NE and SW monsoon under the three timespans representing the RCPs.
In the absence of an optimal species distribution model (SDM) for a specific species, geographic area, and spatial resolution, making decisions based solely on the output of a single SDM, while neglecting the uncertainty associated with alternative models, may lead to ineffective management planning decisions (Davies et al., 2023). Uncertainty analysis is crucial for two primary reasons (Douglas-Smith et al., 2020). Firstly, comprehending the extent of variability linked to model types may assist engineers and decision-makers in identifying potential areas for improvement; secondly, this comprehension can signify the degree of confidence related to SDM predictions. If simulated results derived from numerous models with acceptable performance exhibit considerable discrepancies, the total outcome might be considered to possess high variability (low confidence). Therefore, to check uncertainty, we randomly split the fishing data set in half to utilize 70% for training and 30% for testing in order to validate the chosen model. For both the training and testing datasets, the RMSE and MAE were computed. To further minimize the model’s bias, we used a threefold cross-validation technique between the training and testing sets of the data, using the same performance metrics.
Combining the future and present prediction data with GAM, it allowed us to investigate the patterns in habitat alteration. Latitudinal and longitudinal centers of gravity were calculated to indicate habitat range expansion, both current and future habitats, for moonfish distribution under each climate change scenario. NE monsoon and SW monsoon were the two seasons that the study’s months were categorized under. The following were the latitudinal and longitudinal centers of gravity of the moonfish P-CPUE from the Taiwanese small-scale purse seiners that were estimated from the RCP scenarios and the present P-CPUE in order to have a better understanding of the seasonal geographical distribution of fishing and the accompanying variations:
Using Matlab, we mapped out potential habitats for moonfish and compared them to the ones that would be appropriate in each future climatic scenario in terms of percentage change of P-CPUE. This allowed us to see how the range of viable habitats would change and to ascertain whether appropriate habitats were losing or gaining ground for moonfish in the study region. This allowed us to delve deeper into the pattern of habitat change.
With all covariates incorporated, the final GAM yielded a deviance explained value of 39.2%, RMSE of 2.39, MAE of 1.85, and GCV score of 5.8. We utilized the standardization model to standardize the moonfish N-CPUE because its histogram and quantile-quantile plot (QQ plot) (Figure 2) showed a distribution that was approximately normal. Supplementary Figure S2 shows that the monthly CPUE varied between 7 and 40 individuals after standardization. Minor differences in the coefficient between two randomly split data sets (70:30) indicate no considerable bias in the predictive model, as seen in the final GAM, which did not show any significant difference (Table 2).
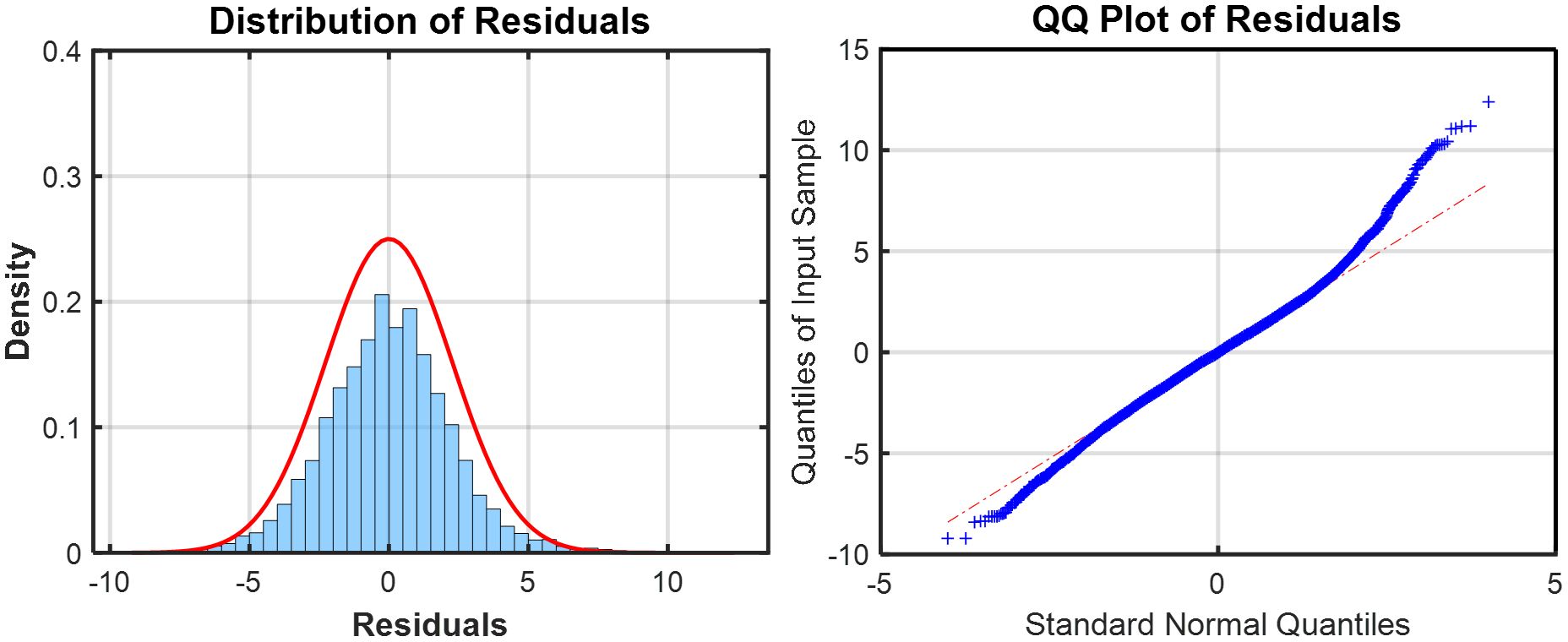
Figure 2. Distribution of residuals and quantile-quantile (Q-Q) plot for chosen generalized additive model (GAM) in moonfish.
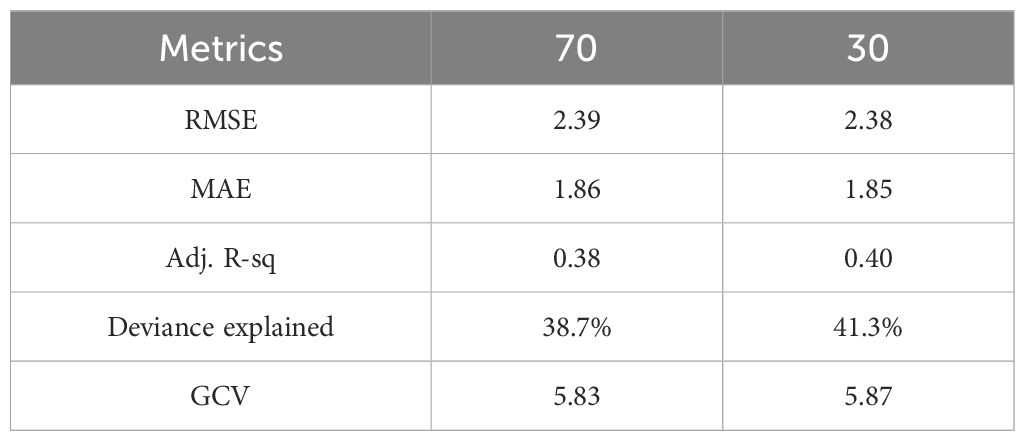
Table 2. Random splitting for verification of the standardization model.
The GAM prediction assessment revealed that various environmental conditions differently influence the distribution of moonfish in the two different monsoon seasons. The distribution of moonfish is primarily influenced by SSH during the NE monsoon, but the distribution during SW monsoon is more significantly impacted by SSC (Figure 3) as evident by the lowest RMSE, MAE, and GCV values. The correlation relevance of environmental variables by GAM indicated that SSH, SSS, and MLD from NE monsoon were the most significant factors influencing the models while SST had a moderate influence. On the other hand, the GAM revealed that SSC and SSH during the SW monsoon were the most critical components affecting the models, with MLD having a moderate impact.
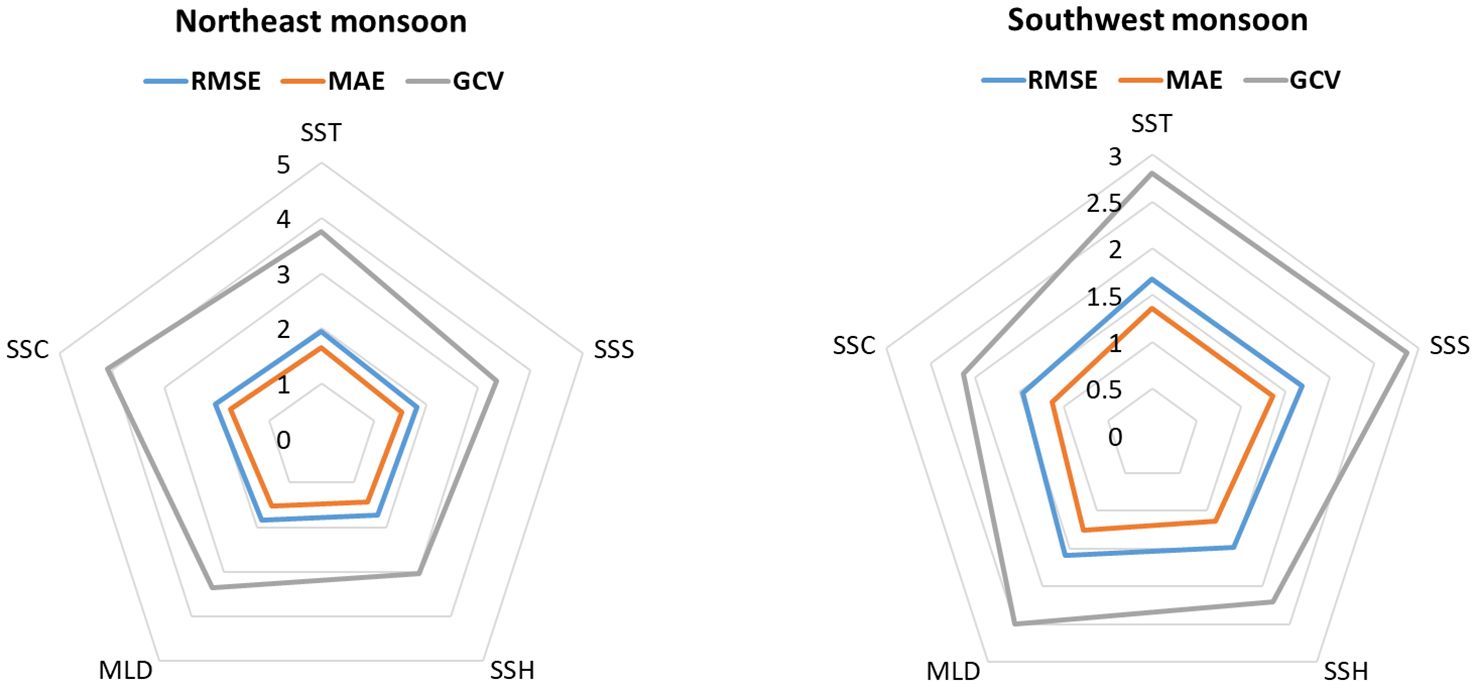
Figure 3. GAM evaluations for environmental conditions.
Figure 4 illustrates the relative contribution of each variable across two RCPs for moonfish in the two monsoons. The values indicate the significance of environmental predictors in forecasting the future distribution of moonfish under each RCP. During NE monsoon, MLD functions as the main predictor, with RI values of 0.35 (RCP 2.6), 0.39 (RCP 4.5), and 0.33 (RCP 8.5), highlighting its importance in nutrient mixing and prey availability for foraging. SSC remains significant, however its importance declines with higher RCPs, dropping from 0.21 (RCP 2.6) to 0.15 (RCP 8.5), indicating changes in primary production. SST (0.24–0.25) and SSH (0.16–0.22) have modest significance, with SSH’s relevance increasing under RCP 8.5 owing to potential changes in stratification, although SSS has a negligible influence (0.04–0.11). During the SW monsoon, SSC functions as the key predictor, with its importance rising from 0.43 (RCP 2.6) to 0.48 (RCP 8.5), highlighting its crucial role in spawning success via primary production. MLD has a secondary position (0.14–0.18), indicating its influence on prey dynamics during spawning. SST is less important in the SW monsoon than the NE monsoon, being constant at 0.12 throughout all RCPs, although SSH (0.20–0.22) has moderate relevance, and SSS contributes little (0.04–0.09). SSC is more influential during SW monsoon, perhaps aiding in spawning, whereas the MLD is more critical in NE monsoon owing to its role in nutrient mixing for foraging.
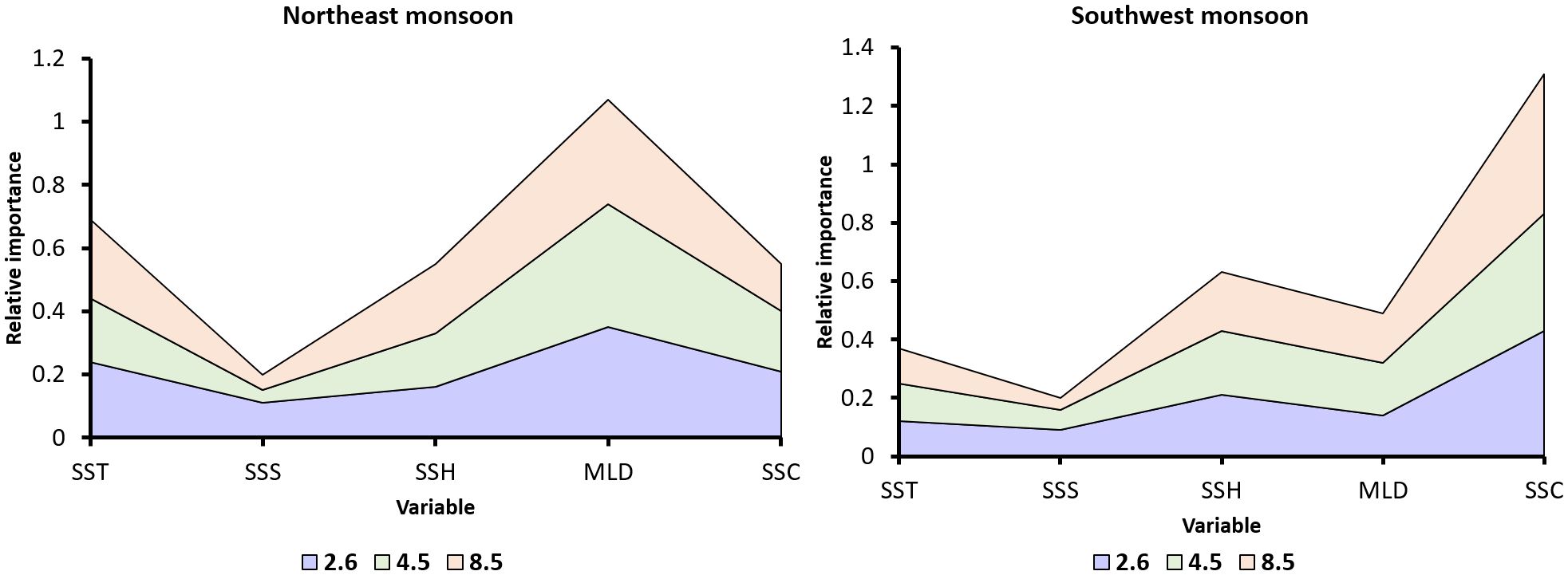
Figure 4. The relative importance of environmental variables in forecasting future distributions of moonfish species: across various RCP scenarios in northeast (NE) and southwest (SW) monsoon.
SI curves for moonfish were developed based on a range of the selected parameters. Following this, habitat suitability index (HSI) scores were computed for both the seasons separately for timescales 2014 – 2020, followed by the RCPs 2.6 and 8.5 for 2040, 2050, and 2060. RCP 4.5 has not been showed here since the ranges were quite similar to that of 8.5. During the NE monsoon, the ideal ranges for moonfish’s SST, SSS, SSH, MLD, and SSC were found to be 22.5-23.5°C, 34.5 PSU, 0.66-0.7 m, 21-41 m, and 0.15–0.45 mg/m3, respectively, when the SI is greater than 0.6. on the other hand, during SW monsoon, the optimal values for moonfish’s SST, SSS, SSH, MLD, and SSC were determined to be 29.5°C, 33.2 PSU, 0.7-0.78 m, 10-12 m, and 0.12 mg/m3, respectively, when SI exceeds 0.6. When the SI value exceeded 0.6, the abundance for moonfish in the study region from exhibited a strong correlation with all the parameters (Figures 5, 6). SI curves were subsequently plotted utilizing the smoothing spline technique consequently for the RCP scenarios showing the suitable ranges of environmental parameters for the years 2034-2040, 2044-2050, and 2054-2060 (Supplementary Figures S3A–S3D).
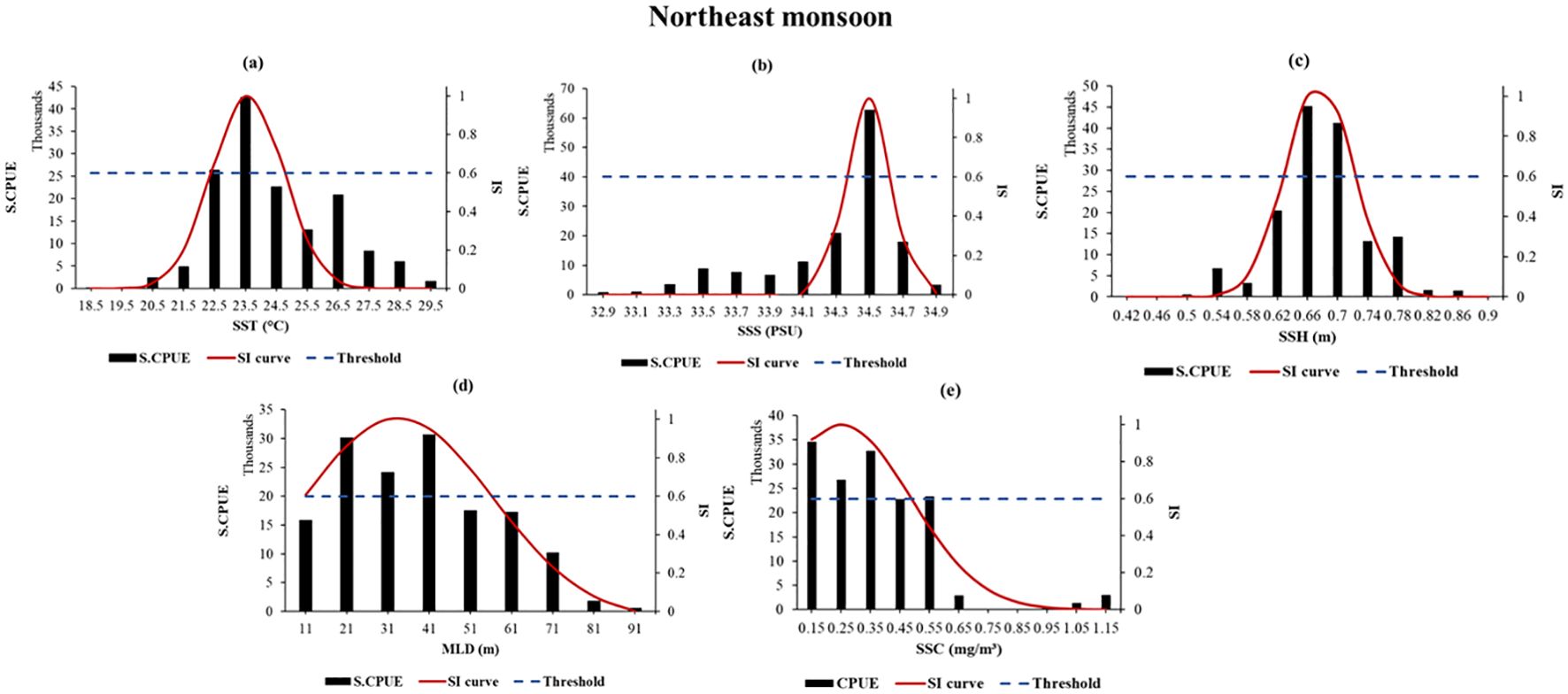
Figure 5. (a–e) SI curves for selected environmental variables related to moonfish from NE monsoon generated using smoothing spline regression. The black bars, blue dotted line, and red solid line represent the summed S-CPUE, SI scores with a cut-off value exceeding 0.6, and SI curves, respectively. The points of intersection between the horizontal dotted lines and SI curves denote the optimal environmental range for each parameter.
The VIF scores among the variables for both the seasons were notably low and < 5 (Supplementary Figure S4). Therefore, all the five variables were selected as the definitive environmental predictors for the model’s inputs. This option seeks to mitigate the possibility of multicollinearity in the model, guaranteeing that each variable distinctly and substantially contributes to the prediction of moonfish distribution in the study region.
Table 3 displays the results derived from the application of GAMs utilizing all the selected parameters. The final GAM for NE monsoon demonstrated enhanced performance with 82% deviance explained, 0.87 RMSE, 0.65 MAE, and 0.78 GCV scores compared to the other models. Table 3 further indicated that the model for SW monsoon exhibited a deviance explained of 74.4%, 0.86 RMSE, 0.6 MAE, and 0.77 GCV scores.
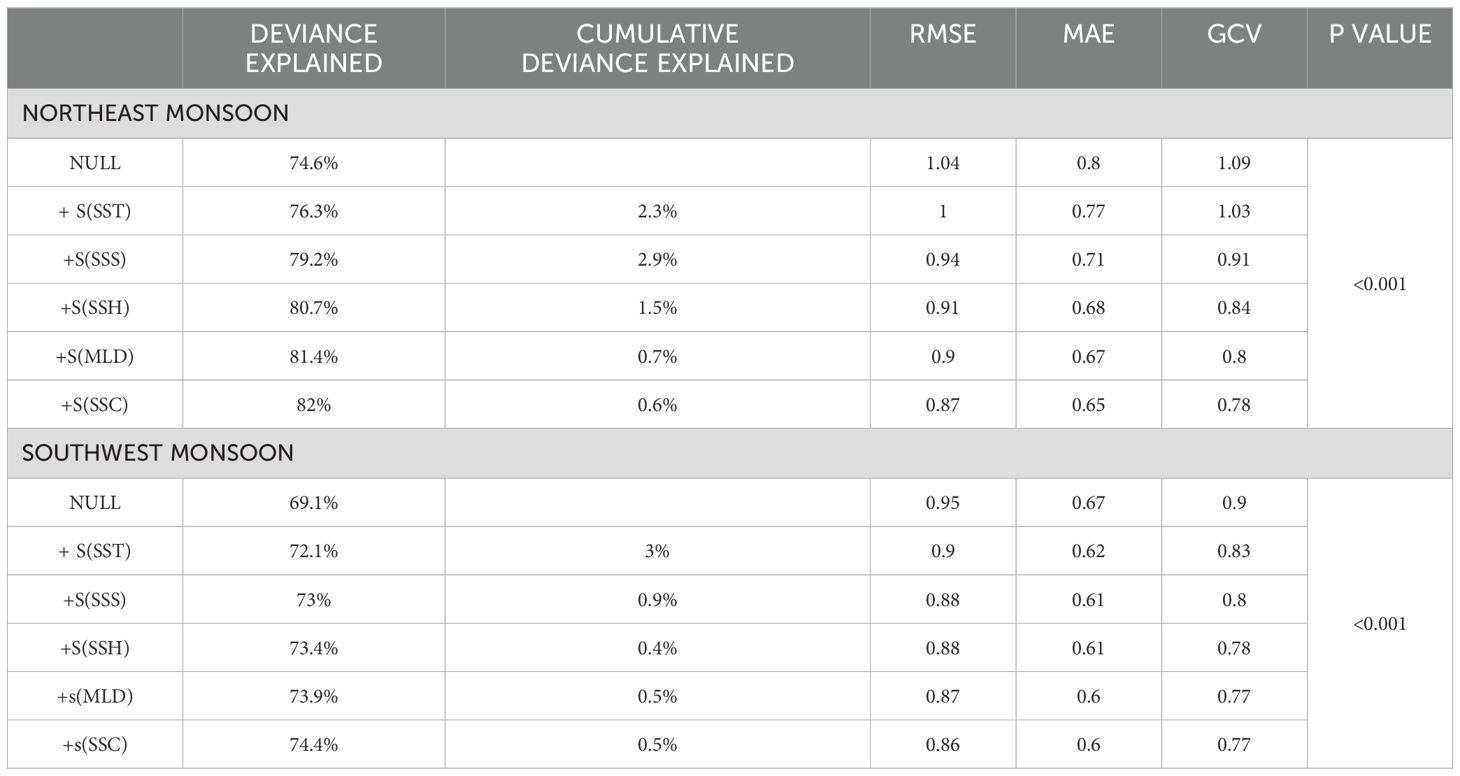
Table 3. Evaluation of GAMs for moonfish during the period of NE and SW monsoons.
GAMs were assessed through two validation techniques, all demonstrating negligible variations in the coefficient values (RMSE and MAE) across random splitting (70:30) and three-fold cross validation approaches. The findings suggest that the predictive performance of the models remained unaffected by significant bias (Table 4). Consequently, these models were utilized to produce a comprehensive climate forecast for all sampling locations within the study area under RCPs 2.6, 4.5, and 8.5 in three timescales – 2034 – 2040, 2044 – 2050, and 2054 – 2060, separated between two monsoons.
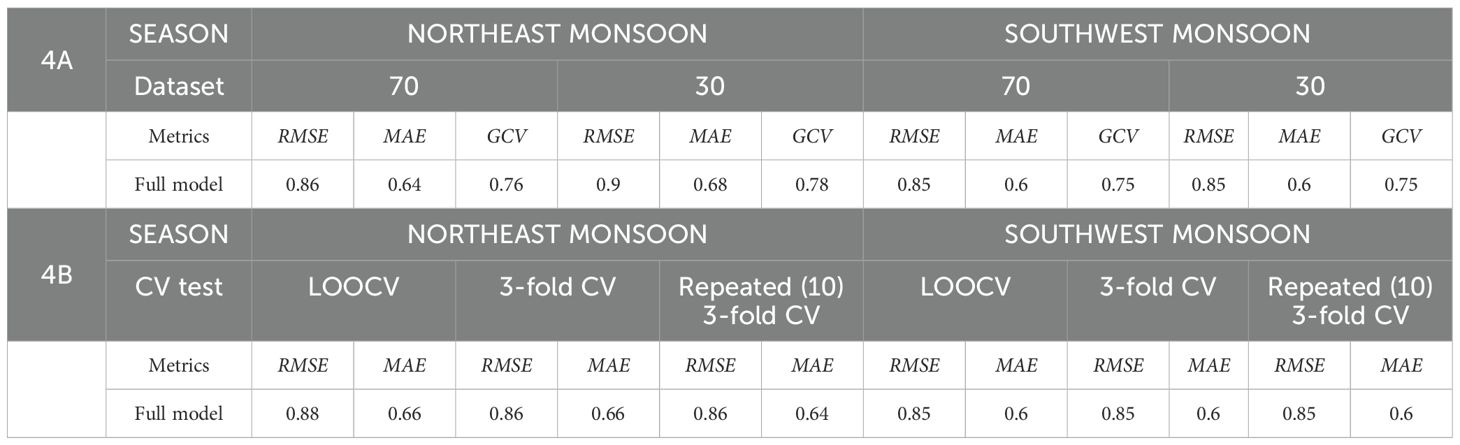
Table 4. Results of validation for GAM prior to habitat prediction.
The P-CPUE exhibited notable variability between the two monsoons; however, the primary fishing ground remained consistently unchanged across the seasons (Figure 7). The projected CPUE values derived from the habitat GAMs and environmental data were compared with the S-CPUE values across different seasons. The findings revealed variations in the size and intensity of moonfish habitats between the NE and SW monsoons. No significant shift in moonfish habitats was observed in the SWT region. During the NE monsoon season, moonfish distributions were observed between 119° and 121°E, while in the SW monsoon, their distributions extended in the Taiwan Bank near 118°E. Throughout the two seasons, the anticipated CPUE regions demonstrated elevated S-CPUE values. Furthermore, low forecast and S-CPUE values were noted, suggesting heightened fishing activity in the coastal regions of northern Taiwan.
During the NE monsoon, moonfish habitats were predominantly observed in the coastal waters off central and western Taiwan, specifically between 21.5°N and 24°N, indicating a broader distribution. Throughout the two seasons, a small habitat region was consistently monitored between 119°E and 120°E. The findings indicate that our vessels demonstrated a high cumulative CPUE, correlated with latitude and longitude. During the NE monsoon, the observed peak of S-CPUE gradually shifted eastward, ultimately reaching 118°E during the Southwest monsoon. GAM indicated a positive correlation between S-CPUE growth and P-CPUE growth during the two monsoons, despite seasonal volatility in S-CPUE. The average S-CPUE values were 17.48 kg/h during the NE monsoon and 14.75 kg/h during the SW monsoon. The mean S-CPUE was lower during the SW monsoon compared to the NE monsoon, corroborating the findings of our previous study by Ray et al. (2024). The S-CPUE values recorded during the NE and SW monsoons were 977.7 kg/h and 856.41 kg/h, respectively. The P-CPUE was greater during the NE monsoon compared to the SW monsoon season.
Figure 8 depicts the fluctuations in the geographical distribution of moonfish during the NE and SW monsoon across several RCP scenarios and temporal markers. The centroid for the NE monsoon demonstrates a consistent southern shift throughout all RCPs, indicating a decrease in appropriate habitat approaching the equator. Under RCP 2.6, the shift is mild, progressing around 50–60 km southward by 2060, but under RCP 4.5, the change is more pronounced, reaching almost 100 km to the south. RCP 8.5 denotes the greatest significant southerly displacement, exceeding 150 km by 2060, demonstrating the impact of extreme warming on habitat sustainability. The southern movements during NE monsoon indicate that the population often relocates to warmer tropical waters. In contrast, during SW monsoon, the centroid migrates southward and westward, highlighting the synergistic impacts of warming and oceanographic changes on spawning habitats. The westward shift is particularly pronounced under RCP 8.5, with the center moving around 40 km west and 20–30 km south by 2060. Under RCP 2.6 and 4.5, the displacements are minimal, with the centroid shifting less than 30 km westward and 10–20 km southward. The westward expansion during SW monsoon may be attributed to changes in ocean productivity and stratification patterns that promote spawning activities in western regions such as the Taiwan Bank. Unlike the two seasons, the NE monsoon demonstrates a more direct southerly movement, indicating habitat contraction due to increasing temperatures and changes in prey availability. In contrast, the SW monsoon has a more complex movement, with westward expansion likely affected by fluctuations in production and spawning-related dynamics. The magnitude of these modifications increases with the severity of the RCP scenario, with RCP 8.5 demonstrating the most pronounced and varied changes.
The percentage change analysis of moonfish P-CPUE indicates that it exhibits the greatest percentage of P-CPUE change (~25%) during the NE monsoon under 2060_8.5 scenario in comparison to SW monsoon 2060_8.5 (~18%) (Figure 9). In contrast, the only negative % change of the moonfish P-CPUE was exhibited in 2050_2.6 scenario. Under three RCPs, the two minimum % changes of P-CPUE were seen during 2040_2.6 in NE (2%) and (4%) SW monsoon respectively. The percentages of P-CPUE change showed a constant positive increasing pattern from 2.6 to 8.5 emission scenarios between the two seasons.
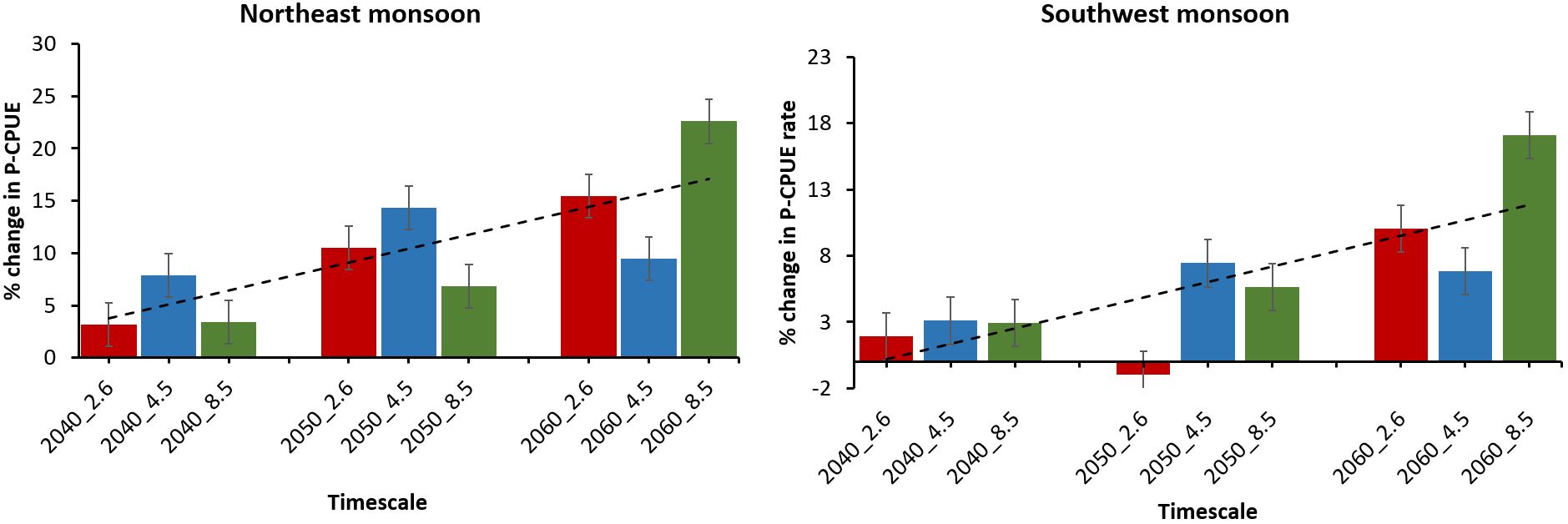
Figure 9. The proportion of P-CPUE experiencing % change from the current to future simulation periods for moonfish habitats in the 2040s, 2050s and 2060s is presented under RCP 2.6, RCP 4.5, and RCP 8.5 climate scenarios.
This study presents the development of a species distribution model aimed at predicting the potential distribution of moonfish in the southwest region of Taiwanese waters, considering both current and projected future climate conditions for the first time that elucidates complex ecological relationships, establishing a robust basis for subsequent ecological research. Model predictions indicate that moonfish predominantly inhabits the coastal and offshore regions west and south of Taiwan during both NE and SW monsoons, showing a preference for areas with varying oceanographic characteristics in the two seasons. The oceanographic parameters exhibited significantly different ranges across the two seasons, suggesting that moonfish inhabit distinct environments during the NE and SW monsoons.
The present study found that although SSC concentration is typically higher during the NE monsoon, its contribution to CPUE in the study area was more significant during the SW monsoon, suggesting that this period might be more suitable for moonfish spawning. This increased SSC aligns with greater food availability, attracting moonfish for reproduction. Prior studies have shown that tropical species, like siphonophores and euphausiid shrimp, increase in summer due to the South China Sea Warm Current (SCSWC), enhancing moonfish prey concentrations (Hsieh et al., 2016). In Taiwan’s tropical and subtropical waters, seasonal planktonic variation differs from other regions, likely influencing SSC thresholds for moonfish during the SW monsoon. During the NE monsoon, chlorophyll levels are high but range narrowly due to nutrient influx from river runoff, promoting primary productivity via eutrophication of nitrogen and phosphorus, which supports higher trophic levels through a bottom-up effect (Sinnickson et al., 2021). Previous studies indicate primary productivity is higher in Taiwan during the NE monsoon (Hsu et al., 2020), and increased turbidity during this period is observed due to light intensity, promoting photosynthesis (Zainuri et al., 2018). The Kuroshio Branch Current (KBC) and China Coastal Current (CCC) contribute to upwelling, with nutrient-rich cold water rising and boosting productivity in TS (Mondal et al., 2023). This enhanced light during the NE monsoon promotes efficient photosynthesis and increased SSC, creating ideal conditions for fishing and moonfish fry refuge.
We concentrated on a selection of key factors to enhance our understanding of the elements affecting the distribution of moonfish. The significance of each predictor variable and the identified relationships indicate that our models possess biological plausibility. Numerous studies have identified temperature as the primary environmental factor influencing the growth, development, abundance, and recruitment of marine fish (Deutsch et al., 2015). A separate study indicated that SST and chlorophyll-a concentration were the primary factors influencing the distribution of marine pelagic fish in the northwest Atlantic Shelf area, consistent with the findings of the present study (Wang et al., 2018), where the relative importance of SST was found to be the second highest after MLD in both NE and SW monsoons under all the RCPs.
MLD’s thickness plays a crucial role in the entrapment of sediment, nutrients, or suspended matter (de Oliveira et al., 2024). In areas characterized by a deeper MLD, vertical convective mixing is more significant, facilitating the upward movement of nutrients from the depths to the surface, thereby enhancing the food supply for moonfish, as evident in Table 3 as well as from its highest relative importance under the RCPs (Figure 4). During the transitional period of the spring monsoon (SW monsoon), reduced wind speed and variable wind direction result in inadequate energy to sustain a deep MLD in the South China Sea (SCS) (Wu and Cheng, 2013). Thus, although ocean depth plays a significant role in the distribution of moonfish in this study, the extent of variations in its SI values under different timeframes in different RCPs is comparatively to a great extent.
We propose that variations in SSH induce alterations in upwelling, water vortex dynamics, and storm surges, resulting in differing levels of mixing between surface and bottom waters, alongside changes in freshwater inputs, which ultimately influence the stability of pH levels (Li et al., 2022). SSH may indicate different warm and cold water masses as well as eddy structures. It is utilized very frequently in distribution models for coastal to pelagic fish and is acknowledged as a significant factor affecting their distribution (Liu et al., 2022). This study found that SSH significantly influences the distribution of moonfish, ranking the first (Figure 3) for moonfish during NE monsoon in impact, as well as having the second-highest relative importance under the RCPs during the SW monsoon, since plays a key role during this season. SSH variations impact the SCSWC, bringing nutrient-rich waters northward during summer, promoting plankton productivity crucial for fish larvae survival (Hsieh et al., 2016). Eddies, created by SSH anomalies, provide retention zones with stable, food-rich environments for spawning fish. Seasonal SSH changes also serve as cues for spawning migration, guiding fish to productive spawning grounds, enhancing larval survival and recruitment (Asch and Checkley, 2013). These factors collectively create optimal spawning conditions through increased nutrient availability and stable habitats.
This study emphasizes the significance of sea temperature in influencing habitat selection by moonfish in a tropical-latitude marine ecosystem, since it had a dominant relative importance for the species in both the seasons under the RCPs (Figure 4). Species with a preference for warmer water, such as moonfish, moved towards further south or southeast during winter or NE monsoon when surface temperatures drop (RCP 4.5 and 8.5), transitioning to more south-westward areas as surface waters warmed in summer.
Moonfish utilized the warm water layers accessible throughout the year. The warm-water preference aligns with its southern distribution range and optimal temperature ranges exceeding 20°C during NE and 29°C during SW monsoon (spawning time for moonfish) (Figure 5a). The SI curves for the oceanographic parameters under the RCPs for both the seasons show us that moonfish has a high range of tolerance of SST (Supplementary Figures S2A–S2D). Moonfish exhibited a preference for southwestern region of Taiwan, around the Taiwan Bank area in all the scenarios under the RCPs for SW monsoon (Figures 6, 7). The abundance of prey in these habitats, combined with enhanced physiological performance in warm waters, likely creates optimal growth conditions for this species during the summer months. Moonfish preferentially chose to move towards the lower southern part of Taiwan during winter or NE monsoon, which may offer lower food availability but provide more favorable thermal conditions. Overall, moonfish showed a higher degree of longitudinal centroid shift than in the latitude.
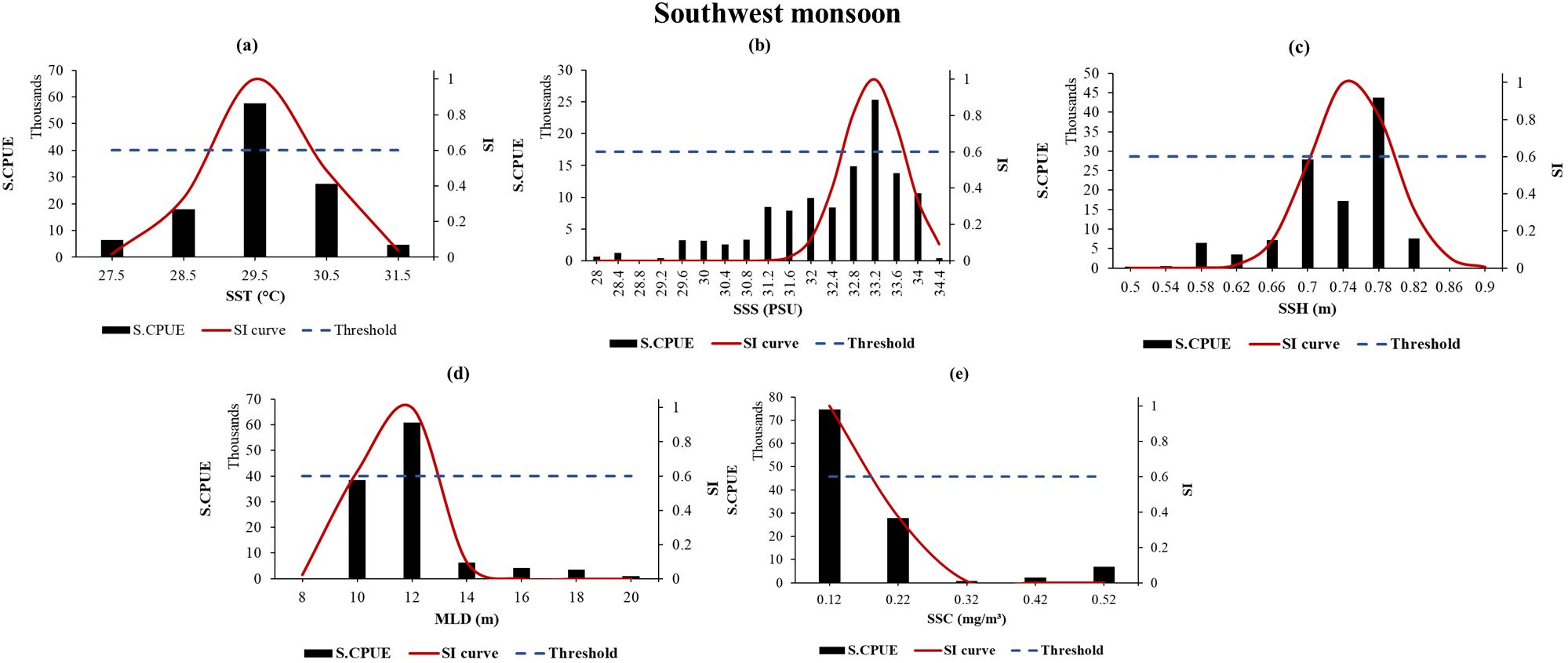
Figure 6. (a–e) SI curves for selected environmental variables related to moonfish from SW monsoon generated using smoothing spline regression. The black bars, blue dotted line, and red solid line represent the summed S-CPUE, SI scores with a cut-off value exceeding 0.6, and SI curves, respectively. The points of intersection between the horizontal dotted lines and SI curves denote the optimal environmental range for each parameter.
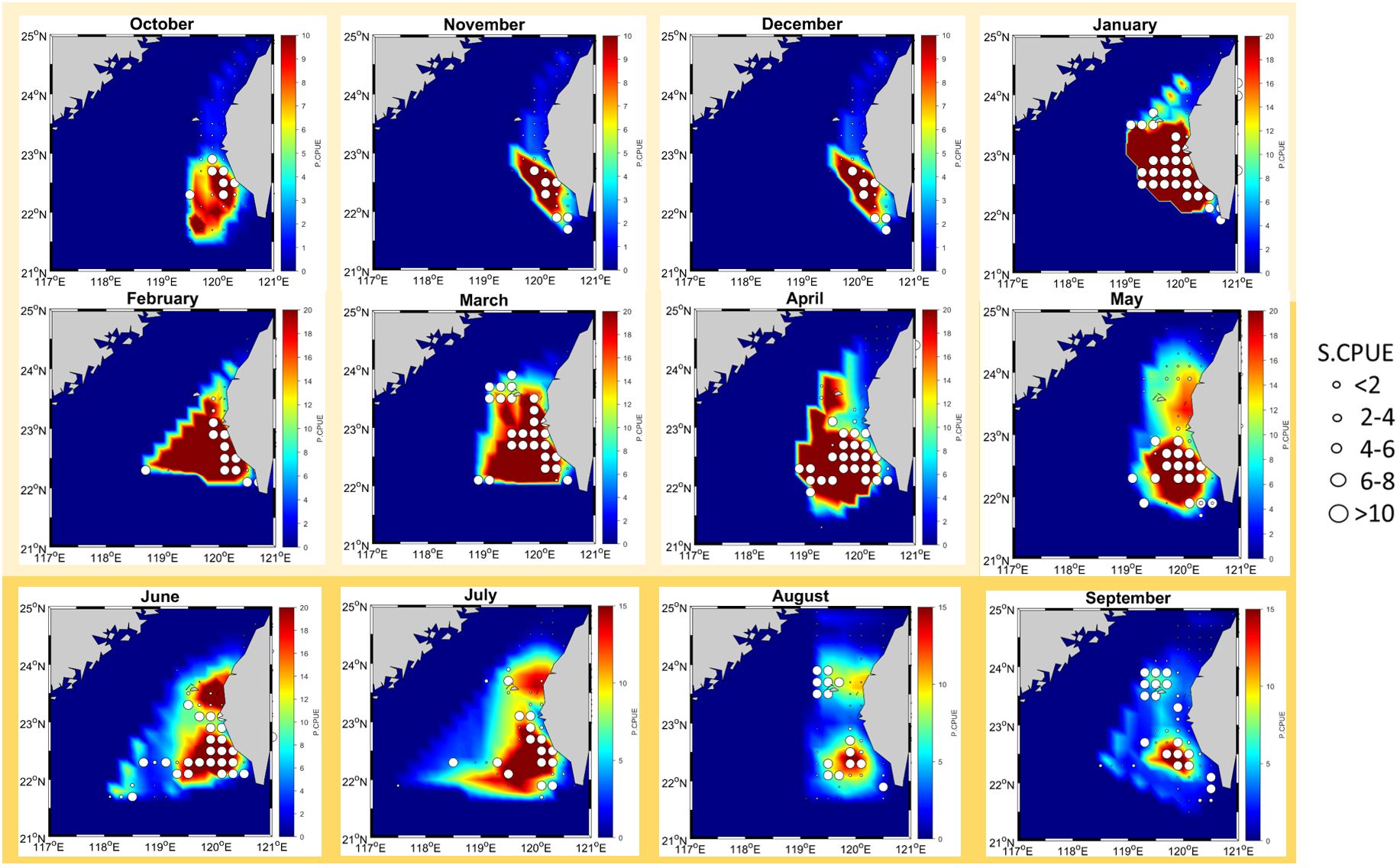
Figure 7. Monthly spatial distribution from NE monsoon (October – April, in light yellow) and SW monsoon (May – September, in darker yellow) of average S-CPUE for moonfish in the waters off southwest Taiwan, mapped onto monthly habitat prediction for the years 2014 – 2020.
The modeling research indicates that the distribution range of moonfish exhibited stability, characterized by a moderate southward shift in the centroid during the NE monsoon while with a southwestward movement during the SW monsoon. The catch potential of moonfish has shown a significant increase under the RCP scenarios (Figure 8). TS may serve as a natural geographical barrier to the northward migration of southern moonfish species in response to climate change as stated by Tzeng et al. (2016). This study demonstrates that moonfish may be a warm-water affinity species that can modify their behavior spatially and temporally to adapt to the seasonal oceanographic variability of the ecosystem. This may enable these fish species to adapt to future temperature changes, including earlier summer warming or extended durations of cold or warm periods. On the other hand, since they are not fast-swimmers, it may also be difficult for them to migrate northward through the Kuroshio region due to strong influence of currents, as well as TS being a natural geographic barrier. The findings indicate a consensus that behavioral traits significantly influence the resilience of animal species to a rapidly changing global climate. Furthermore, it is essential to incorporate the behavioral responses of animals to climate change into predictive models assessing species susceptibility to this biodiversity threat (Buchholz et al., 2019). The utilization of thermal refuges by individuals may, however, deprive fish of other essential resources, such as optimal feeding habitats.
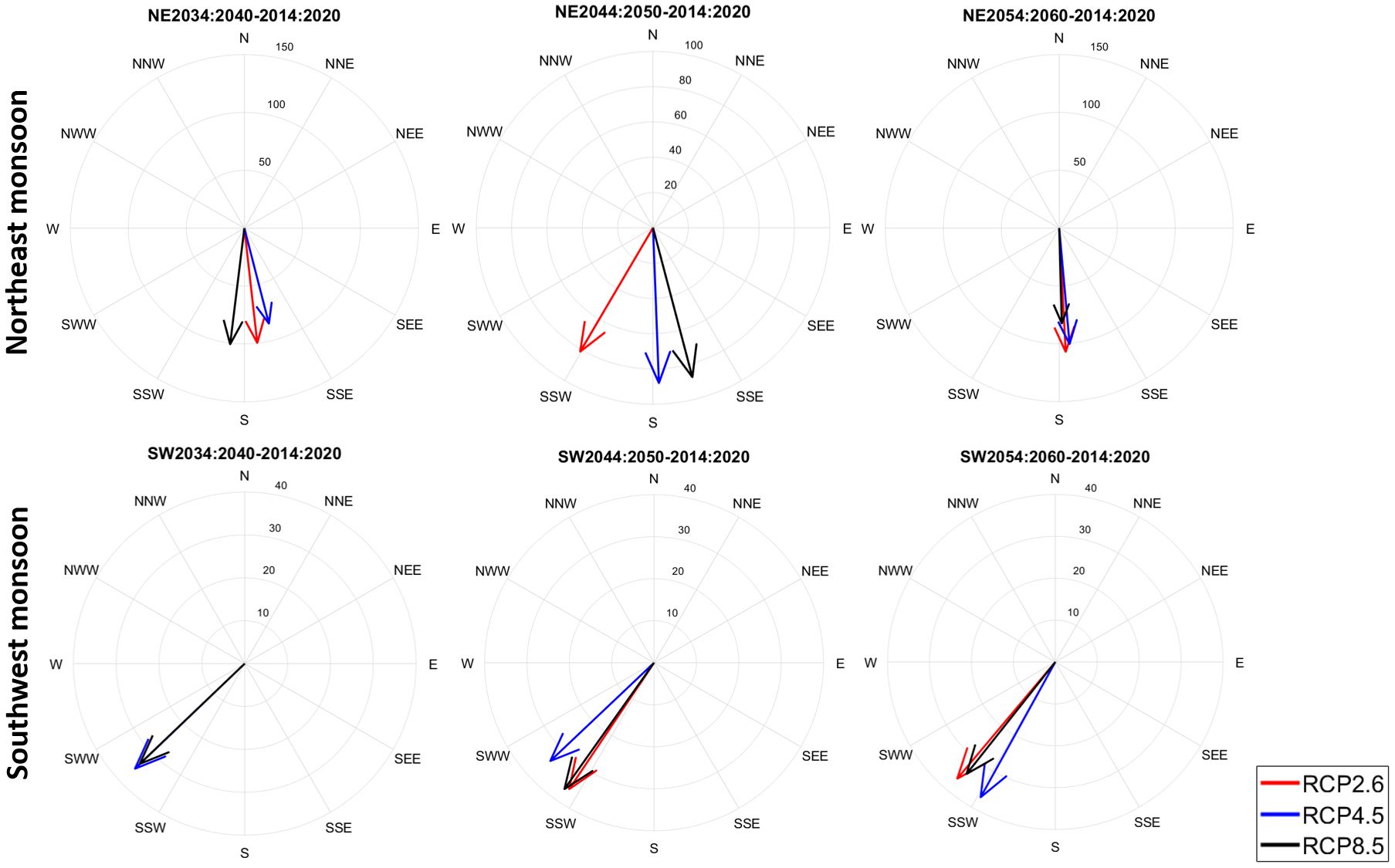
Figure 8. Analysis of the geographical shift for moonfish (in Km) during NE and SW monsoons across different RCP scenarios and time points. Each rose diagram represents the change from the time period 20140 – 2020 to 2034 – 2040, 2044 – 2050, and 2054 – 2060.
Projected rises in summer temperatures in Taiwanese waters may exacerbate the trade-offs between optimal temperature and feeding areas for marine species, potentially impacting their growth rates and overall condition. Concurrently, species adapted to warmer waters, such as moonfish, are anticipated to flourish that could later result in novel assemblages dominated by warm-water species, correlated with rising sea temperatures (Barceló et al., 2016). Buchholz et al. (2019) indicate that additional research is necessary to enhance understanding of the fitness trade-offs associated with behavioral responses to environmental change. Future research should also examine individual-level responses to environmental change, as these may be crucial for understanding individual fitness and, ultimately, how populations adapt to new environments.
Species-specific reactions to climate-induced changes are modifying the geographical overlap and phenological synchronization between predator and prey populations (Carroll et al., 2019). This is expected to impact encounter rates between them, eventually affecting the intensity of predation interactions. Fundamental physiological principles and extensive laboratory research demonstrate that the intensity of predator–prey interactions is influenced by temperature and other abiotic factors, both directly through kinetic impacts on consumption rates (Pepi et al., 2018) and indirectly through effects on predator foraging (Holsman et al., 2019) and prey escape behaviors (Domenici et al., 2019). Consequently, climate change may modify the geographical overlap between predators and prey, as well as the intensity of trophic interactions in their coexistence, thereby affecting predator-prey dynamics in intricate, nonlinear manners (Boukal et al., 2019). Considering that moonfish are a crucial forage species for apex predators, these predators may be required to modify their movement patterns to pursue their prey. If predators do not adeptly track the fluctuating moonfish populations, geographical mismatches may arise, thereby diminishing their feeding efficiency and resulting in cascade impacts on predator populations. An anticipated rise in moonfish abundance may temporarily enhance predator numbers by providing a more plentiful food source leading to increased predation pressure, possibly affecting moonfish behavior, including modified schooling patterns or changes in habitat use to evade predation. Furthermore, a significant population surge might destabilize the current ‘wasp-waist’ regulation within the ecosystem that may result in imbalances, with some prey species of moonfish facing overgrazing, while others may thrive owing to less competition. In addition to predator-prey dynamics, the overall stability of the ecosystem may also be influenced by these changes. The relocation of moonfish habitats and the corresponding variations in their biomass may affect nitrogen cycle and primary production patterns, especially by modifying grazing pressure on planktonic communities. The research indicates that chlorophyll levels and MLD substantially affect moonfish distribution, implying that changes in oceanographic circumstances would also effect primary production. A drop or change in primary production resulting from modified oceanographic processes may induce feedback effects across the food chain, possibly disrupting regional marine ecosystems. Moreover, if moonfish persist under elevated temperatures while other species diminish, the area may see unprecedented species assemblages, potentially impacting biodiversity and the viability of fisheries.
The combined impacts of climate variability and change on species fitness mediate the habitat suitability of small pelagic organisms like moonfish. The importance of acknowledging the effects of long-term environmental changes on small pelagic fish habitats must be emphasized. Gradual environmental modifications can influence the overall habitat suitability of these species, independently or in conjunction with the effects of overfishing. It is important to recognize that using catch rates per vessel as an index of fish abundance may be subject to debate. This is especially applicable in the waters of southwest Taiwan, where the small-scale purse seine fishery is predominantly industrial. Catch rates are contingent upon fish abundance and are affected by catchability and socio-economic factors (Ba et al., 2017).
It is essential to comprehend the habitat preferences of a forage fish like moonfish, and the factors influencing a high CPUE in specific regions require clarification. This study identified latitudinal and longitudinal shifts in the center of gravity of suitable habitats to determine habitat shift patterns of moonfish. This study found that longitudinal changes in the center of gravity of these habitats are more pronounced, offering valuable insights into the observed patterns. A significant challenge in examining these parameters to detect potential distribution shifts is the availability of data on oceanographic and biological variables suitable for integration into climate models.
Alterations in community composition can result in significant ecological and social implications (Asch et al., 2018). The likelihood of species invasion is greater at high latitudes, while species extirpation predominates at low latitudes (Hu et al., 2022). Species richness is anticipated to decline more frequently in the Indo-Pacific region and semi-enclosed seas, such as the southwest Taiwan, compared to other regions (Huang et al., 2016). The transmission of this effect to the catch indicates that cold temperate countries may benefit from these changes, whereas tropical countries could experience adverse impacts (Asch et al., 2018). The current study in Taiwan indicates that catch rates of moonfish may rise in the future as projected from the RCPs (Figure 8) pointing towards an increased abundance of moonfish which may happen if larger predatory fishes migrate to other areas due to changing ocean conditions. Projecting habitat suitability for fish species aids in elucidating their distribution patterns in the context of climate change, which is critical for informing conservation policy (Zhang et al., 2020b).
Our principal results are obtained from SDMs for 7 years of fishery data that analyze relationships between fish species occurrences and factors linked to oceanographic variables. This method neglects the possible species interactions or other human pressures which can also affect the niche, as well as a better interpretation possible from a longer dataset. Furthermore, this analysis presumes that all fish may travel freely and equitably, irrespective of body size consequently stressing the consideration of fish dispersion in future research. Future studies should also investigate the impact of alterations in the growth, reproduction, and survival rates of moonfish on their distribution.
The variation in model outcome demonstrates that although these methodologies are instrumental in comprehending the potential impacts of environmental changes on species distributions, the results are significantly influenced by the specific model selected, and there is no universally ‘best’ or ‘correct’ model. Utilizing a diverse ensemble of models, varying in their conservatism about species estimates, will allow us to illustrate the robustness and consistency of specific projections relative to the underlying assumptions.
The models presented include just certain variables (temperature, salinity, MLD, SSH, SSC), but other factors influencing the appropriate habitat for fish include seabed substrate, currents, pH, and prey availability. The variables were unavailable for the future periods analyzed, hence they were excluded from this analysis. Nevertheless, when further data become accessible, they may be included into future modeling. Subsequent research might use the model outputs to examine predator-prey dynamics, species turnover, functional groupings or attributes, nursery habitats, recruitment, and the economic significance of the species. These evaluations would garner significant attention and may be integrated with estimates about invertebrates, biogenic habitat engineers, or apex predators.
Although the majority of fish habitats are expected to shift northward, the presence of TS as a migration barriers and the ‘cul-de-sac’ effect may significantly impact moonfish movement as seen in the current study. All RCP scenarios indicated a uniform southerly shift in moonfish habitats during the NE monsoon, with the extent of displacement escalating at higher RCP (~150 km). This movement, prompted by rising temperatures and altered oceanic conditions, signifies a reduction of appropriate foraging areas towards equatorial regions, anticipated to exhibit improved habitat suitability as seen by elevated P-CPUE rates. During the SW monsoon, the habitat centroid exhibits southward and westward (~20-40 km) movements, especially under elevated RCPs. These shifts align with the dynamic nature of spawning habitats, influenced by localized productivity hotspots and thermal conditions. The P-CPUE rates during the SW monsoon exhibit slower initial rises (~18%) than the NE monsoon (~25%), underscoring the difficulties in maintaining adequate spawning habitats under constantly evolving oceanographic circumstances. Environmental factors like as SSC and MLD show the highest relative importance under RCP 2.6 to 8.5, with SSC and MLD being predominant in the SW and NE monsoon respectively. The anticipated modifications in SSC under elevated RCPs indicate substantial changes in primary production, directly affecting habitat quality and prey accessibility. Simultaneously, the fluctuation of MLD in future scenarios highlights the significance of ocean mixing in sustaining nutrient-dense foraging areas. These findings underscore the necessity of integrating adaptive management techniques, such as safeguarding equatorial and spawning habitats, to alleviate climate-induced disturbances to moonfish populations. The study highlights the importance of tracking environmental changes and their ecological consequences to guarantee sustainable fisheries management in a warming ocean.
Effective spatial planning and management in moonfish fisheries need governmental assistance and collaboration from key sea-users to guarantee their sustained growth. While marine spatial planning should be integrated into policy development at all levels, our research corroborates that such case studies may assist authorities in gradual modifications of management methods. Cooperation from stakeholders whose livelihoods will be affected by spatial planning should be the first emphasis. Moreover, the integration of stakeholder concerns and the assessment of the effectiveness of current methodologies are essential to enhance the proposed system. Regular communication during workshops by scientists and NGO personnel helps clarify spatial planning objectives and strategies for stakeholders, hence enhancing the efficacy of management activities, since they play a crucial role in community representation, conservation efforts, and mediating advocacy positions, establishing a knowledge base and amplifying overlooked perspectives.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethical approval was not required for the studies on animals in accordance with the local legislation and institutional requirements because the study used an existing dataset.
AR: Conceptualization, Formal analysis, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. SM: Conceptualization, Investigation, Supervision, Validation, Writing – review & editing. M-AL: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – review & editing. Q-HL: Software, Visualization, Writing – review & editing. RS: Writing – review & editing. Y-CW: Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. We would like to express our sincere gratitude to Taiwan Fisheries Agency for supplying data and funding (Grant no:113AS-6.4.2-FA-01) and the National Science and Technology Council (Grant no: NSTC 113-2611-M-019-007) for funding contribution to the completion of this research paper (Grant no: NSTC 113-2611-M-019-007).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1526762/full#supplementary-material
Albo Puigserver M., Giráldez A., Hidalgo M., Ramirez J., Torres P., Massaro A., et al. (2018). Report on historical reproductive pattern: Size and age at first maturity and reproductive period. doi: 10.20350/digitalCSIC/12669
Asch R. G., Checkley D. M. Jr. (2013). Dynamic height: A key variable for identifying the spawning habitat of small pelagic fishes. Deep Sea Res. Part I: Oceanogr. Res. Pap. 71, 79–91. doi: 10.1016/j.dsr.2012.08.006
Asch R. G., Cheung W. W., Reygondeau G. (2018). Future marine ecosystem drivers, biodiversity, and fisheries maximum catch potential in Pacific Island countries and territories under climate change. Mar. Policy 88, 285–229. doi: 10.1016/j.marpol.2017.08.015
Ba A., Schmidt J., Dème M., Lancker K., Chaboud C., Cury P., et al. (2017). Profitability and economic drivers of small pelagic fisheries in West Africa: A twenty year perspective. Mar. Policy 76, 152–158. doi: 10.1016/j.marpol.2016.11.008
Barange M., Bahri T., Beveridge M. C., Cochrane K. L., Funge-Smith S., Poulain F. (2018). Impacts of climate change on fisheries and aquaculture. United Nations’ Food Agric. Organ. 12 (4), 628–635. Available online at: https://www.researchgate.net/profile/Manuel-Barange/publication/325871167_Impacts_of_Climate_Change_on_Fisheries_and_Aquaculture_Synthesis_of_Current_Knowledge_Adaptation_and_Mitigation_Options/links/5b5f1e48aca272a2d6754247/Impacts-of-Climate-Change-on-Fisheries-and-Aquaculture-Synthesis-of-Current-Knowledge-Adaptation-and-Mitigation-Options.pdf.
Barceló C., Ciannelli L., Olsen E. M., Johannessen T., Knutsen H. (2016). Eight decades of sampling reveal a contemporary novel fish assemblage in coastal nursery habitats. Global Change Biol. 22, 1155–1167. doi: 10.1111/gcb.13047
Basheer A. K., Lu H., Omer A., Ali A. B., Abdelgader A. M. (2016). Impacts of climate change under CMIP5 RCP scenarios on the streamflow in the Dinder River and ecosystem habitats in Dinder National Park, Sudan. Hydrol. Earth Sys. Sci. 20, 1331–1353. doi: 10.5194/hess-20-1331-2016
Bertrand A., Vögler R., Defeo O. (2019). Climate change impacts, vulnerabilities and adaptations: Southwest Atlantic and Southeast Pacific marine fisheries1. Impacts Climate Change fisheries aquaculture, 325.
Boukal D. S., Bideault A., Carreira B. M., Sentis A. (2019). Species interactions under climate change: connecting kinetic effects of temperature on individuals to community dynamics. Curr. Opin. Insect Sci. 35, 88–95. doi: 10.1016/j.cois.2019.06.014
Buchholz R., Banusiewicz J. D., Burgess S., Crocker-Buta S., Eveland L., Fuller L. (2019). Behavioural research priorities for the study of animal response to climate change. Anim. Behav. 150, 127–137. doi: 10.1016/j.anbehav.2019.02.005
Carroll G., Holsman K. K., Brodie S., Thorson J. T., Hazen E. L., Bograd S. J., et al. (2019). A review of methods for quantifying spatial predator–prey overlap. Global Ecol. Biogeogr. 28, 1561–1577. doi: 10.1111/geb.v28.11
Cattano C., Claudet J., Domenici P., Milazzo M. (2018). Living in a high CO2 world: A global meta-analysis shows multiple trait-mediated fish responses to ocean acidification. Ecol. Monogr. 88, 320–335. doi: 10.1002/ecm.1297
Champion C., Coleman M. A. (2021). Seascape topography slows predicted range shifts in fish under climate change. Limnol. Oceanogr. Lett. 6, 143–153. doi: 10.1002/lol2.10185
Cheng L., Abraham J., Hausfather Z., Trenberth K. E. (2019). How fast are the oceans warming? Science 363 (6423), 128–129. doi: 10.1126/science.aav7619
Cheng L., Trenberth K. E., Gruber N., Abraham J. P., Fasullo J. T., Li G., et al. (2020). Improved estimates of changes in upper ocean salinity and the hydrological cycle. J. Climate 33, 10357–10381. doi: 10.1175/JCLI-D-20-0366.1
Davies S. C., Thompson P. L., Gomez C., Nephin J., Knudby A., Park A. E., et al. (2023). Addressing uncertainty when projecting marine species’ distributions under climate change. Ecography 2023, e06731. doi: 10.1111/ecog.06731
de Oliveira L. E., Yamasaki T. N., Janzen J. G., Gualtieri C. (2024). Effects of vegetation density on flow, mass exchange and sediment transport in lateral cavities. Journal of Hydrology 632, 130910. doi: 10.1016/j.jhydrol.2024.130910
Deutsch C., Ferrel A., Seibel B., Pörtner H. O., Huey R. B. (2015). Climate change tightens a metabolic constraint on marine habitats. Science 348, 1132–1135. doi: 10.1126/science.aaa1605
Domenici P., Allan B. J., Lefrançois C., McCormick M. I. (2019). The effect of climate change on the escape kinematics and performance of fishes: implications for future predator–prey interactions. Conserv. Physiol. 7, coz078. doi: 10.1093/conphys/coz078
Douglas-Smith D., Iwanaga T., Croke B. F., Jakeman A. J. (2020). Certain trends in uncertainty and sensitivity analysis: An overview of software tools and techniques. Environ. Model. Softw. 124, 104588. doi: 10.1016/j.envsoft.2019.104588
FAO, R (2022). The state of world fisheries and aquaculture 2022. Towards blue transformation (Rome: The State of World Fisheries and Aquaculture (SOFIA), 266.
Franco B. C., Defeo O., Piola A. R., Barreiro M., Yang H., Ortega L., et al. (2020). Climate change impacts on the atmospheric circulation, ocean, and fisheries in the southwest South Atlantic Ocean: a review. Climatic Change 162, 2359–2377. doi: 10.1007/s10584-020-02783-6
Free C. M., Thorson J. T., Pinsky M. L., Oken K. L., Wiedenmann J., Jensen O. P. (2019). Impacts of historical warming on marine fisheries production. Science 363, 979–983. doi: 10.1126/science.aau1758
Gatta P. P. (2022). The state of world fisheries and aquaculture 2022 (FAO eBooks). doi: 10.4060/cc0461en
Gregorich M., Strohmaier S., Dunkler D., Heinze G. (2021). Regression with highly correlated predictors: variable omission is not the solution. Int. J. Environ. Res. Public Health 18, 4259. doi: 10.3390/ijerph18084259
Habib A., Borazon E. Q., Nallos I. M., Macusi E. D. (2025). Climate change vulnerability, adaptation and ecosystem services in different fisheries and aquaculture in Asia: a review: English. Mar. Fish. Sci. (MAFIS) 38, 1. doi: 10.47193/mafis.3822025010101
Haghi Vayghan A., Poorbagher H., Taheri Shahraiyni H., Fazli H., Nasrollahzadeh Saravi H. (2013). Suitability indices and habitat suitability index model of Caspian kutum (Rutilus frisii kutum) in the southern Caspian Sea. Aquat. Ecol. 47, 441–451. doi: 10.1007/s10452-013-9457-9
Hayashi M., Shiogama H., Emori S., Ogura T., Hirota N. (2021). The northwestern Pacific warming record in August 2020 occurred under anthropogenic forcing. Geophys. Res. Lett. 48, e2020GL090956. doi: 10.1029/2020GL090956
Holsman K. K., Aydin K., Sullivan J., Hurst T., Kruse G. H. (2019). Climate effects and bottom-up controls on growth and size-at-age of Pacific halibut (Hippoglossus stenolepis) in Alaska (USA). Fish. Oceanogr. 28, 345–358. doi: 10.1111/fog.12416
Hsieh R. J., Hsieh H. Y., Lo W. T. (2016). Succession of monsoons and water mass influences on Euphausiid assemblages in the waters around Taiwan, western North Pacific Ocean. Zool. Stud. 55, p. e46. doi: 10.6620/2FZS.2016.55-46
Hsu P. C., Lu C. Y., Hsu T. W., Ho C. R. (2020). Diurnal to seasonal variations in ocean chlorophyll and ocean currents in the north of Taiwan observed by Geostationary Ocean Color Imager and coastal radar. Remote Sens. 12, 2853. doi: 10.3390/rs12172853
Hu W., Du J., Su S., Tan H., Yang W., Ding L., et al. (2022). Effects of climate change in the seas of China: Predicted changes in the distribution of fish species and diversity. Ecol. Indic. 134, 108489. doi: 10.1016/j.ecolind.2021.108489
Hu S., Sprintall J., Guan C., McPhaden M. J., Wang F., Hu D., et al. (2020). Deep-reaching acceleration of global mean ocean circulation over the past two decades. Sci. Adv. 6, eaax7727. doi: 10.1126/sciadv.aax7727
Huang M., Ding L., Wang J., Ding C., Tao J. (2021). The impacts of climate change on fish growth: A summary of conducted studies and current knowledge. Ecol. Indic. 121, 106976. doi: 10.1016/j.ecolind.2020.106976
Huang D., Hoeksema B. W., Affendi Y. A., Ang P. O., Chen C. A., Huang H., et al. (2016). Conservation of reef corals in the South China Sea based on species and evolutionary diversity. Biodivers. Conserv. 25, 331–344. doi: 10.1007/s10531-016-1052-7
Kim J. H. (2019). Multicollinearity and misleading statistical results. Korean J. anesthesiol. 72, 558–569. doi: 10.4097/kja.19087
Kulp S. A., Strauss B. H. (2019). New elevation data triple estimates of global vulnerability to sea-level rise and coastal flooding. Nat. Commun. 10 (1), 1–12. doi: 10.1038/s41467-019-12808-z
Kwiatkowski L., Torres O., Bopp L., Aumont O., Chamberlain M., Christian J. R., et al. (2020). Twenty-first century ocean warming, acidification, deoxygenation, and upper-ocean nutrient and primary production decline from CMIP6 model projections. Biogeosciences 17, 3439–3470. doi: 10.5194/bg-17-3439-2020
Lan K. W., Lee M. A., Chou C. P., Vayghan A. H. (2018). Association between the interannual variation in the oceanic environment and catch rates of bigeye tuna (Thunnus obesus) in the Atlantic Ocean. Fish. Oceanogr. 27, 395–407. doi: 10.1111/fog.12259
Lee T. Y., Chiu C. C., Chen C. J., Lin C. Y., Shiah F. K. (2023). Assessing future availability of water resources in Taiwan based on the Budyko framework. Ecol. Indic. 146, 109808. doi: 10.1016/j.ecolind.2022.109808
Lee D., Son S. H., Lee C. I., Kang C. K., Lee S. H. (2019). Spatio-temporal variability of the habitat suitability index for the Todarodes pacificus (Japanese common squid) around South Korea. Remote Sens. 11, 2720. doi: 10.3390/rs11232720
Lee M. A., Weng J. S., Lan K. W., Vayghan A. H., Wang Y. C., Chan J. W. (2020). Empirical habitat suitability model for immature albacore tuna in the North Pacific Ocean obtained using multisatellite remote sensing data. Int. J. Remote Sens. 41, 5819–5837. doi: 10.1080/01431161.2019.1666317
Li Y., Yang H., Dang J., Yang X., Xue L., Zhang L. (2022). Seasonal variation of sea surface pH and its controls in the Jiaozhou Bay, China. Continent. Shelf Res. 232, 104613. doi: 10.1016/j.csr.2021.104613
Liao C. P., Huang H. W., Lu H. J. (2019). Fishermen's perceptions of coastal fisheries management regulations: Key factors to rebuilding coastal fishery resources in Taiwan. Ocean Coast. Manage. 172, 1–13. doi: 10.1016/j.ocecoaman.2019.01.015
Liu S., Liu Y., Li J., Cao C., Tian H., Li W., et al. (2022). Effects of oceanographic environment on the distribution and migration of Pacific saury (Cololabis saira) during main fishing season. Sci. Rep. 12, 13585. doi: 10.1038/s41598-022-17786-9
Mbaye A., Schmidt J., Cormier-Salem M. C. (2023). Social construction of climate change and adaptation strategies among Senegalese artisanal fishers: Between empirical knowledge, magico-religious practices and sciences. Soc. Sci. Human. Open 7, 100360. doi: 10.1016/j.ssaho.2022.100360
McCauley D. J., Pinsky M. L., Palumbi S. R., Estes J. A., Joyce F. H., Warner R. R. (2015). Marine defaunation: animal loss in the global ocean. Science 347, 1255641. doi: 10.1126/science.1255641
Monaco C. J., Bradshaw C. J., Booth D. J., Gillanders B. M., Schoeman D. S., Nagelkerken I. (2020). Dietary generalism accelerates arrival and persistence of coral-reef fishes in their novel ranges under climate change. Global Change Biol. 26, 5564–5573. doi: 10.1111/gcb.15221
Mondal S., Lee M. A., Weng J. S., Osuka K. E., Chen Y. K., Ray A. (2023). Seasonal distribution patterns of Scomberomorus commerson in the Taiwan Strait in relation to oceanographic conditions: An ensemble modeling approach. Mar. pollut. Bull. 197, 115733. doi: 10.1016/j.marpolbul.2023.115733
Mondal S., Ray A., Ito S. I., Osuka K. E., Lee M. A., Lu Q. H. (2024). Nonunidirectional habitat changes associated with global climate change: the example of the indo-pacific king mackerel (Scomberomorus guttatus) in the Taiwan strait. Fish. Oceanogr., e12718. doi: 10.1111/fog.12718
Murray C. S., Wiley D., Baumann H. (2019). High sensitivity of a keystone forage fish to elevated CO2 and temperature. Conserv. Physiol. 7, coz084. doi: 10.1093/conphys/coz084
N’Souvi K., Adjakpenou A., Sun C., Ayisi C. L. (2024). Climate change perceptions, impacts on the catches, and adaptation practices of the small-scale fishermen in Togo’s coastal area. Environ. Dev. 49, 100957. doi: 10.1016/j.envdev.2023.100957
Nerem R. S., Beckley B. D., Fasullo J. T., Hamlington B. D., Masters D., Mitchum G. T. (2018). Climate-change–driven accelerated sea-level rise detected in the altimeter era. Proc. Natl. Acad. Sci. 115, 2022–2025. doi: 10.1073/pnas.1717312115
Nilsson J. A., Johnson C. R., Fulton E. A., Haward M. (2019). Fisheries sustainability relies on biological understanding, evidence-based management, and conducive industry conditions. ICES J. Mar. Sci. 76 (6), 1436–1452. doi: 10.1093/icesjms/fsz065
Palmer M. D., Roberts C. D., Balmaseda M., Chang Y. S., Chepurin G., Ferry N., et al. (2017). Ocean heat content variability and change in an ensemble of ocean reanalyses. Climate Dynam. 49, 909–930. doi: 10.1007/s00382-015-2801-0
Pedersen E. J., Miller D. L., Simpson G. L., Ross N. (2019). Hierarchical generalized additive models in ecology: an introduction with mgcv. PeerJ 7, e6876. doi: 10.1111/2041-210X.13974
Pepi A., Grof-Tisza P., Holyoak M., Karban R. (2018). As temperature increases, predator attack rate is more important to survival than a smaller window of prey vulnerability. Ecology 99, 1584–1590. doi: 10.1002/ecy.2356
Pörtner H. O., Scholes R. J., Arneth A., Barnes D. K. A., Burrows M. T., Diamond S. E., et al. (2023). Overcoming the coupled climate and biodiversity crises and their societal impacts. Science 380, eabl4881. doi: 10.1126/science.abl4881
Ray A., Mondal S., Osuka K. E., Sihombing R. I., Lee M. A., Wang Y. C., et al. (2024). Impact of climate variability on moonfish (Mene maculata) catch rate in the waters off southwestern Taiwan. Fish. Oceanogr. 33, e12667. doi: 10.1111/fog.12667
Schickele A., Goberville E., Leroy B., Beaugrand G., Hattab T., Francour P., et al. (2021). European small pelagic fish distribution under global change scenarios. Fish Fish. 22, 212–225. doi: 10.1111/faf.12515
Sinnickson D., Chagaris D., Allen M. (2021). Exploring impacts of river discharge on forage fish and predators using ecopath with ecosim. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.689950
Trégarot E., D’Olivo J. P., Botelho A. Z., Cabrito A., Cardoso G. O., Casal G., et al. (2024). Effects of climate change on marine coastal ecosystems–A review to guide research and management. Biol. Conserv. 289, 110394. doi: 10.1016/j.biocon.2023.110394
Tzeng C. H., Chiu TsanYu C. T., Chen ChihShin C. C., Wang HuiYu W. H., Chiu TaiSheng C. T. (2016). The current population structure of the demersal hairtail (Trichiurus japonicus) in the western North Pacific was shaped by the Taiwan Strait, as revealed by mitochondrial DNA. 0372-333X, 61 (4), 279–287. doi: 10.6165/tai.2016.61.279
Vayghan A. H., Lee M. A., Weng J. S., Mondal S., Lin C. T., Wang Y. C. (2020). Multisatellite-based feeding habitat suitability modeling of albacore tuna in the Southern Atlantic Ocean. Remote Sens. 12, 2515. doi: 10.3390/rs12162515
Venegas R. M., Acevedo J., Treml E. A. (2023). Three decades of ocean warming impacts on marine ecosystems: A review and perspective. Deep Sea Res. Part II: Topical Stud. Oceanogr. 212, 105318. doi: 10.1016/j.dsr2.2023.105318
Vrigazova B. (2021). The proportion for splitting data into training and test set for the bootstrap in classification problems. Business Syst. Res.: Int. J. Soc. Adv. Innovation Res. Econ. 12, 228–242. doi: 10.2478/bsrj-2021-0015
Wang L., Kerr L. A., Record N. R., Bridger E., Tupper B., Mills K. E., et al. (2018). Modeling marine pelagic fish species spatiotemporal distributions utilizing a maximum entropy approach. Fish. Oceanogr. 27, 571–586. doi: 10.1111/fog.12279
Wu Y., Cheng G. S. (2013). Seasonal and inter-annual variations of the mixed layer depth in the South China Sea. Mar. Forcasts 30, 9–17. doi: 10.11737/j.issn.1003-0239.2013.03.002
Zainuri M., Ario R., Munandar B., Prayogi H. (2018). “February. Impact of monsoon to aquatic productivity and fish landing at Pesawaran Regency Waters,” in IOP Conference Series: Earth and Environmental Science, 116 (1), 012021 (Indonesia: IOP Publishing). doi: 10.1088/1755-1315/116/1/012021
Zhang Z., Mammola S., Xian W., Zhang H. (2020a). Modelling the potential impacts of climate change on the distribution of ichthyoplankton in the Yangtze Estuary, China. Diversity Distrib. 26, 126–137. doi: 10.1111/ddi.13002
Keywords: climate change projections, small-scale fisheries, moonfish, catch increase, Taiwan, catch potential, RCP
Citation: Ray A, Mondal S, Lee M-A, Lu Q-H, Sihombing RI and Wang Y-C (2025) Impact of climate change on the waters off southwest Taiwan: predicted alterations in moonfish distribution and catch rates. Front. Mar. Sci. 12:1526762. doi: 10.3389/fmars.2025.1526762
Received: 12 November 2024; Accepted: 10 February 2025;
Published: 05 March 2025.
Edited by:
Edison D. Macusi, Davao Oriental State University (DOrSU), PhilippinesReviewed by:
Ahasan Habib, University of Malaysia Terengganu, MalaysiaCopyright © 2025 Ray, Mondal, Lee, Lu, Sihombing and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ming-An Lee, bWFsZWVAbWFpbC5udG91LmVkdS50dw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.