
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci. , 27 February 2025
Sec. Marine Ecosystem Ecology
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1511313
This article is part of the Research Topic Bridging Knowledge Gaps in Marine Biological Invasions View all 7 articles
 Fabrizio Marcondes Machado1*†
Fabrizio Marcondes Machado1*† Cristiane Midori Suga2†Fabio Kiyoshi Camara Onodera2†Mariana Sousa Melo2†Thais Viti2†Vilma Maria Cavinatto2†Ana Carolina da Rocha Lammardo3†Carolina Yumi Yonamine3†Regina Sawaia Sáfadi3†Luis Antonio Nogueira-Junior4†Luiz Fernando Maciel Oliva4†
Cristiane Midori Suga2†Fabio Kiyoshi Camara Onodera2†Mariana Sousa Melo2†Thais Viti2†Vilma Maria Cavinatto2†Ana Carolina da Rocha Lammardo3†Carolina Yumi Yonamine3†Regina Sawaia Sáfadi3†Luis Antonio Nogueira-Junior4†Luiz Fernando Maciel Oliva4†Biological invasions are globally recognized as one of the main drivers of biodiversity loss. Over the past four decades, there has been significant progress in understanding the mechanisms of alien species introduction into marine and estuarine environments. It is currently known that although humans can spread non-native species in various ways (e.g. aquaculture, vessel biofouling and oil and gas activities), ballast water is one of the most effective being considered one of the main threats to marine biodiversity. This pathway also appears to be the main transport vector to explain the spread of the exotic bivalve Theora lubrica (or Asian Semele) since the late 1950s. In this study, we report for the first time the presence of Asian Semele in the Southwestern Atlantic, specifically along the Southeast coast of Brazil. Furthermore, we also provide new insights into shell morphology, gross anatomy and ecological aspects, as well as the updated global distribution of this invasive alien species.
Hundreds of exotic species are likely introduced annually worldwide, with ports as the primary entry point and ballast water being one of the most effective means of transport (Endresen et al., 2004; Costello et al., 2022). Some ports host more non-native species than native ones, with certain introduced species displaying massive colonization, gradually contributing to global biotic homogenization (Simpson et al., 2017).
In South American countries, unlike those in the NE Atlantic, there is a lack of ongoing projects or comprehensive studies on alien marine species. Even so, new records of exotic marine species in Brazil, Argentina, and Uruguay have increased over the past two decades (Orensanz et al., 2002; Castro et al., 2017). For Brazil, specifically, 63 non-native marine and/or estuarine species are recorded, of which 15 are categorized as invasive, 11 of which are bivalves species (>70%), the oysters Isognomon bicolor (C. B. Adams, 1845), Magallana gigas (Thunberg, 1793), Saccostrea cuccullata (Born, 1778), Crassostrea virginica (Gmelin, 1791), and the mussels Xenostrobus securis (Lamarck, 1819), Leiosolenus aristatus (Dillwyn, 1817), Mytilus galloprovincialis Lamarck, 1819, Mytilopsis cf. sallei (probably a species complex), Mytilopsis leucophaeata (Conrad, 1831), Perna perna (Linnaeus, 1758) and Perna viridis (Linnaeus, 1758) (Simone and Gonçalves, 2006; MMA, 2009; Dias et al., 2013; Queiroz et al., 2022; Fernandes et al., 2024; Beltrão et al., 2024; Machado et al., 2023, 2024).
In general, bivalves are regarded as efficient invaders and ecosystem engineers, sometimes capable of drastically altering the environment (Boltovskoy and Correa, 2015; Linares et al., 2017). This invasive potential has already been well documented in Brazilian waters, especially for larger and epifaunal species (MMA, 2009). However, studies on exotic species—whether invasive or not—that are small (<1 cm in length) and infaunal are rare. In this context, the present work provides the first record of the tiny bivalve Theora lubrica A. Gould, 1861 commonly known as the Asian Semele, in the Southwestern Atlantic. Morphological descriptions, ecological notes, and updates on the global distribution of this invasive species were also addressed here.
Ranked among the 20 ports with the highest risk of biological invasions, the Port of Santos is located within the Santos Estuary, between the municipalities of Santos, Cubatão, and Guarujá in Southeastern Brazil (Seebens et al., 2013). The port features several terminals for cargo storage and handling, boasting the largest container traffic in Brazil and connections to over 600 ports worldwide (Santos Port Authority, 2024). As expected for estuarine areas with high human impact, the water column surrounding the port exhibits low dissolved oxygen levels, elevated concentrations of nitrogen, phosphorus, and organic matter, along with harmful chemical elements, with areas of the estuary ranging from mesotrophic to eutrophic (Roversi et al., 2016; CETESB, 2023). As is typical in port areas, the Port of Santos and its surrounding regions also report annual increases in the number of exotic aquatic species (Rotundo et al., 2020). Since 2017, however, the Port has implemented and consistently maintained an on-site bioinvasion monitoring project to track and mitigate alien species (Santos Port Authority, 2024).
All 10 individuals of Theora lubrica with preserved internal tissues were provided by a consultancy project (Econsult Estudos Ambientais) aimed to evaluate the benthic fauna of the estuarine areas surrounding the Port of Santos. The specimens were collected from only three of the 50 stations covered by the port monitoring project, between December 2023 and May 2024. Samplings were performed in soft substrata at depths ranging from 4.5 to 15.6 meters, using a Petersen grab (0.058 m²), with three replicates in each of the three stations. Water parameters such as temperature, salinity, pH, conductivity, dissolved oxygen and turbidity, were measured in each station. The samples were sieved in the field and stored in 70% ethanol. The shells and soft parts of T. lubrica were photographed using a motorized Nikon SMZ25 stereomicroscope equipped with a Nikon Digital Sight 10 camera; images were stacked using Nikon NIS Elements Basic Research (v. 5.42.04) software. Some well-preserved valves were also selected for hinge characterization using a Hitachi TM4000 Plus tabletop SEM. The material is archived at Museu de Zoologia da Universidade Estadual do Norte do Paraná, under the numbers UENP 2935–2936.
Being an exotic species widely distributed around the globe, the specimens collected in Brazil were compared with all morphologically similar species of the genus Theora, as well as with Abra nitida (O. F. Müller, 1776). The latter, although never recorded in the Southwestern Atlantic, has an outline resembling that of T. lubrica. Type series of the most similar species, including Theora lata (Hinds, 1843) (Holotype - USNM 535) and Abra prismatica (Syntype - EXEMS Moll3731-5), were also examined. A redescription of the shell of T. lubrica, including the hinge teeth details, was provided due to the lack of high-resolution images in the literature; tooth numbering followed Carter et al. (2012). General anatomical features, including details of the internal tissues, were provided for the first time. It is important to note that, due to their small size and extremely fragile shells, individuals of T. lubrica can easily be broken during sampling and/or misidentified as juveniles of other species of the Tellinoidea. The authors FMM and CMS also identified specimens of T. lubrica from Northeastern Brazil (state of Bahia); however, precise collection data are not currently available for publication.
The specimens of Theora lubrica were sampled in three stations (sta. P11: Santos Port channel; sta. P20 and PD10: Largo do Canéu - inner part of the Santos Estuary) at the Port of Santos - Southeastern Brazil. All information regarding the stations and water parameters is compiled in Table 1, which indicates that these specimens inhabit a polyhaline environment (18-30 ppt) in a subtidal zone, tolerating a wide range of dissolved oxygen concentrations (3-15 mg/L).
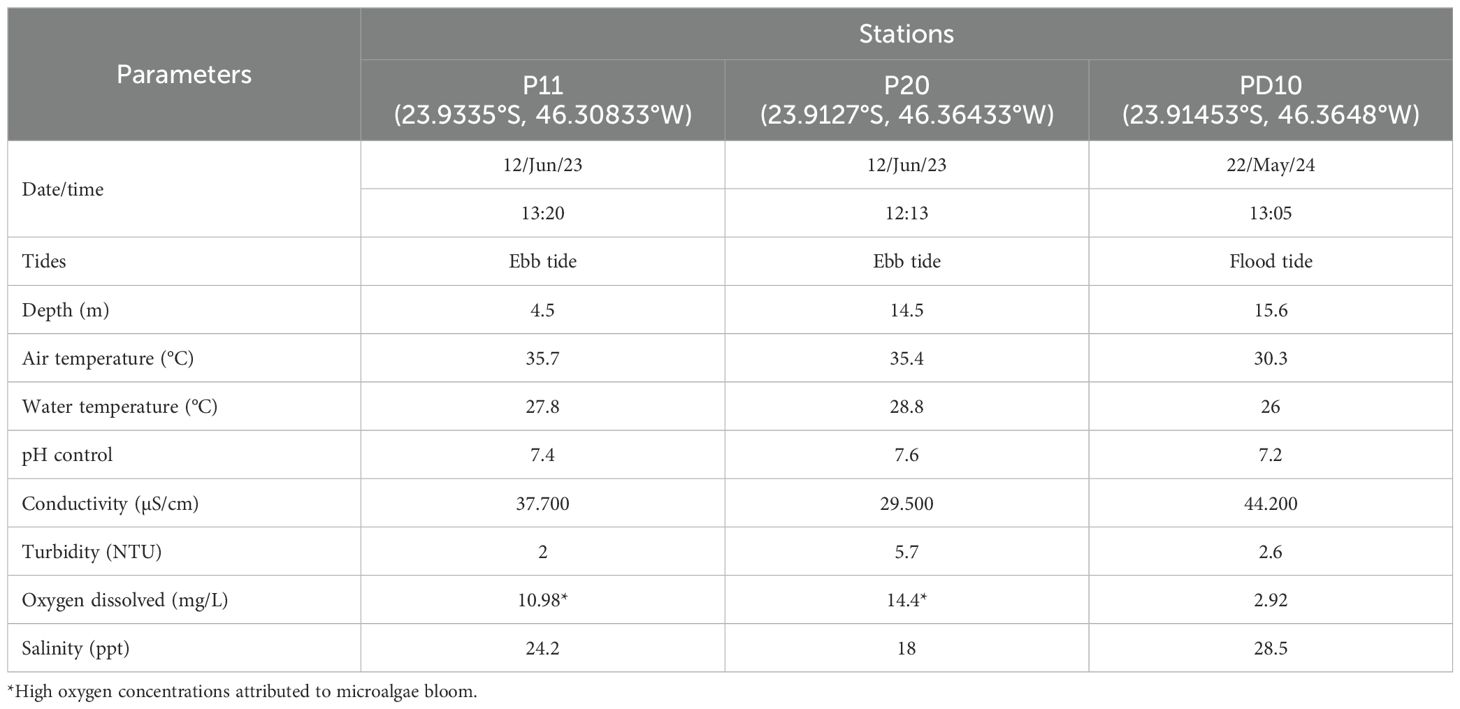
Table 1. Set of measurements of water parameters at the three sampling stations in the Port of Santos – Brazil.
Superfamily Tellinoidea Blainville, 1814
Family Semelidae Stoliczka, 1870
Genus Theora H. Adams & A. Adams, 1856
Theora lubrica Gould, 1861 (Figures 1, 2)
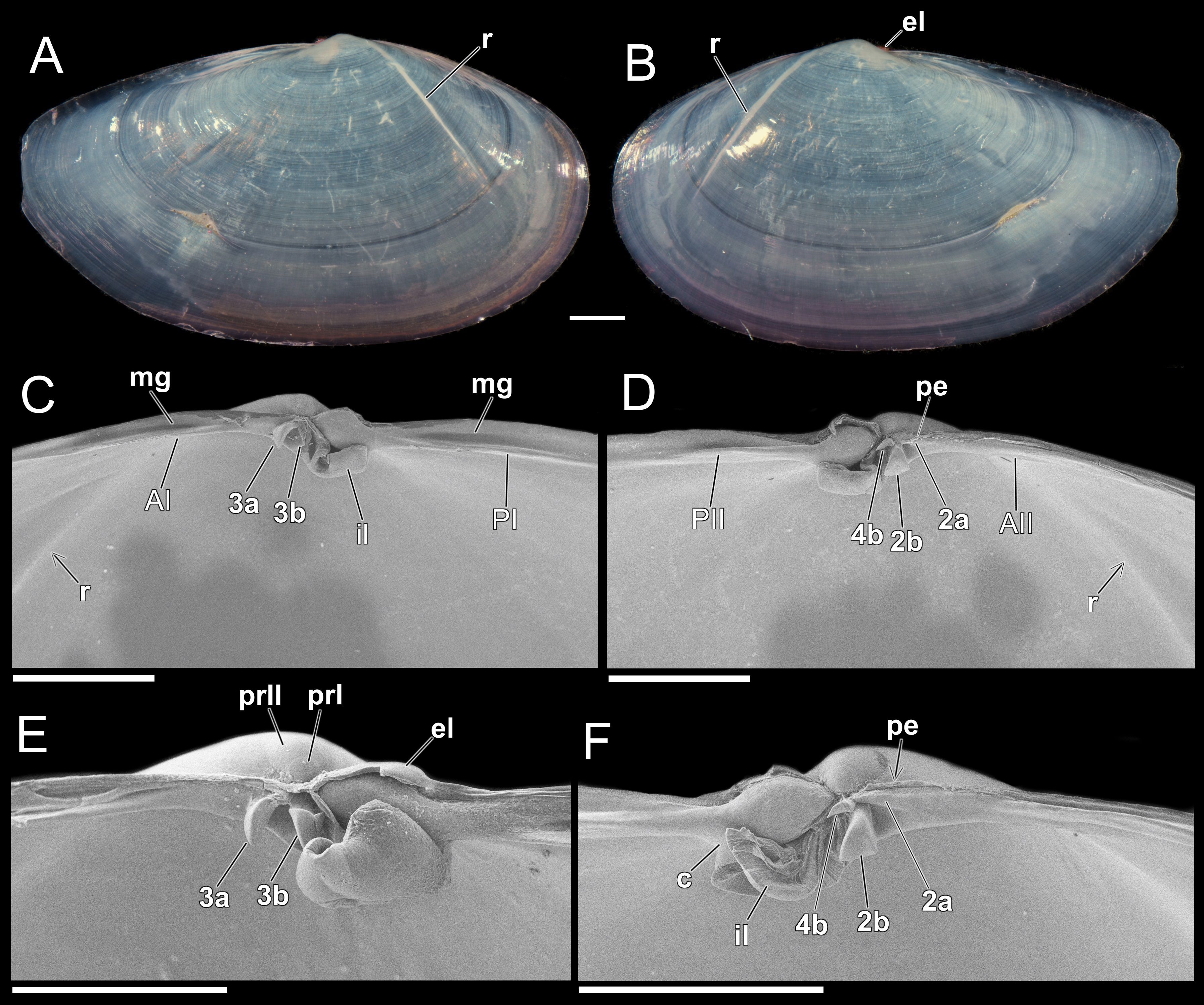
Figure 1. Theora lubrica, shell details (UENP 2935). (A, B) Photomicrography of the outer surface of the right and left valves, respectively, showing the fine concentric lines. The internal ridge (r) is seen by transparency. (C-E) SEM views. (C, D) Ligament portions (el, il), internal ridge and hinge details of the right and left valves, respectively. (C, E) Right hinge with two divergent cardinal teeth (3a, 3b) and single anterior and posterior laterals (AI, PI) – tooth 3b, partially broken in figure (C). (D, F) Left hinge with three cardinal teeth (4b, 2b, 2a) and single anterior and posterior laterals (AII, PII); (D) Tooth ‘2a’, covered by periostracum. 2a, left anterior cardinal tooth; 2b, left middle cardinal tooth; 4b, left posterior cardinal tooth; 3a, right anterior cardinal tooth; 3b, right posterior cardinal tooth; AI/AII, anterior lateral teeth; c, chondropore; el, external ligament portion; il, internal ligament portion; mg, marginal groove; PI/PII, posterior lateral teeth; pe, periostracum; prI/prII, prodissoconch I or II; r, internal ridge. Scale bars: (A, B) (1 mm), (C, D, F) (500 µm) and (E) (400 µm).
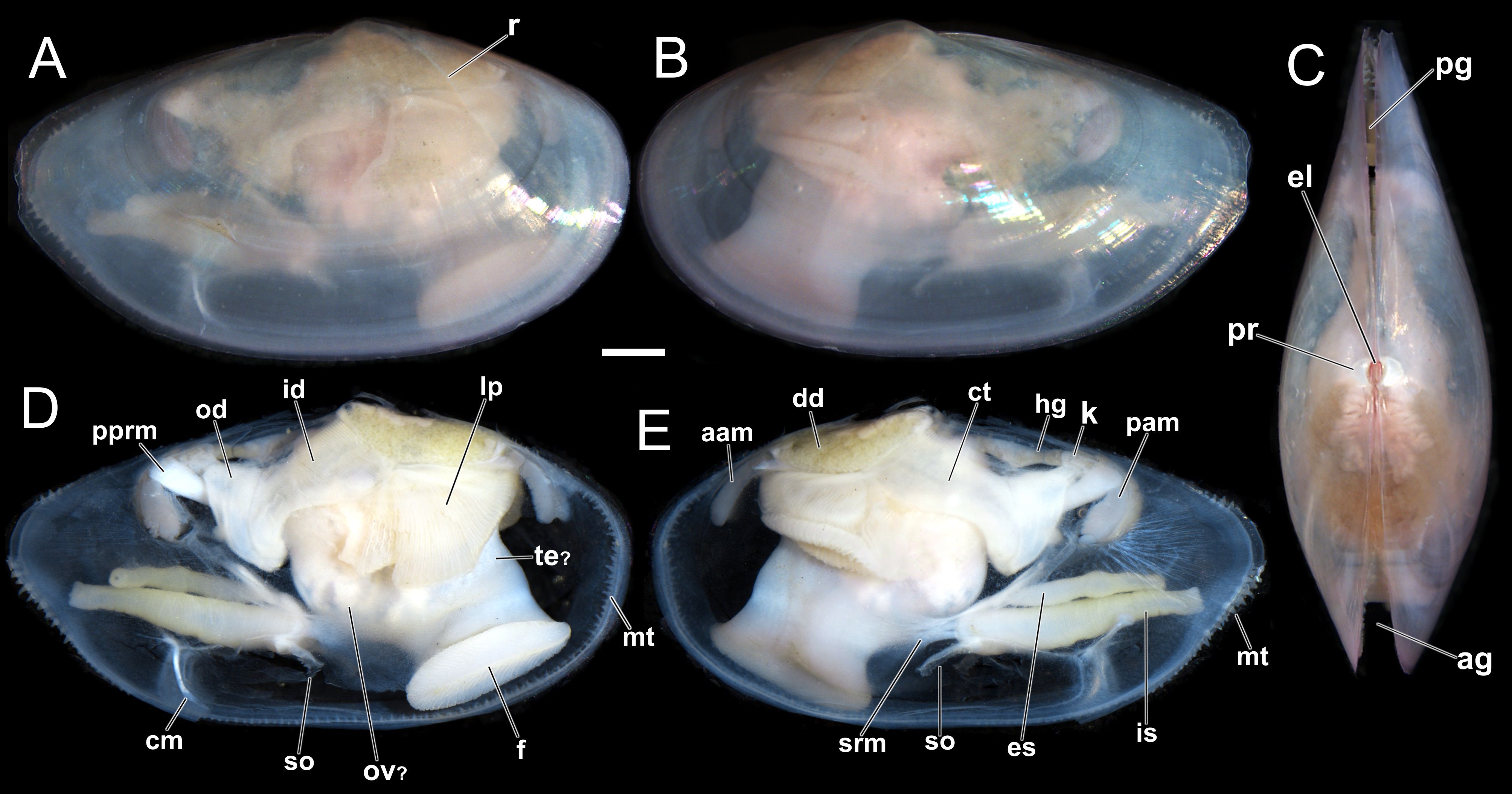
Figure 2. Theora lubrica, shell and gross anatomy (UENP 2936). (A, B) Outer views of the right and left valves with soft parts inside, respectively. (C) Frontal view showing the external ligament (el), prodissoconch (pr) and the valves with slight gapes at the anterior and posterior ends (ag, pg). (D, E) Gross anatomy, showing the topology of internal organs on the right and left sides of the same individual. aam, anterior adductor muscle; ag , anterior gape; cm, cruciform muscle; ct, ctenidia; dd, digestive diverticula; el, external ligament portion; es, exhalant siphon; f, foot; hg, hindgut; id, inner demibranch; is, inhalant siphon; lp, labial palps; mt, mantle tentacles; od, outer demibranch; ov?, ovary; pam, posterior adductor muscle; pprm, posterior pedal retractor muscle; pr, prodissoconch (=larval shell); r, internal ridge; so, siphonal organ; srm, siphonal retractor muscles; te?, testis. Scale bar: (A, B) (1 mm).
Diagnosis of Theora
Shell thin, hyaline, smooth, gaping on at least one side. Anterior end rounded; posterior end produced. Hinge with one to three cardinal teeth; lateral present in at least one valve. Chondrophore projecting. Deep pallial sinus confluent with pallial line (after Adams, 1864; Coan et al., 2000).
Shell small up to 16 mm (Brazilian specimens, 2.8 to 10.3 mm in length), thin, elongated, semi-transparent and elliptical. Equivalve, inequilateral, umbones slightly anterior, pointed and prominent; larval shell, reniform, whitish, smooth and well-preserved (290 ± 30µm n=6), prodissoconch I (80 ± 10µm n=6) and II boundary only visible in SEM (Figures 1A, B, E, 2A–C). Outer surface smooth, whitish with a shiny periostracum; shell bears fine, concentric lines; internal anterior ridge visible (r), mainly in dry shells (Figures 1A, B, 2A, B). Valves gape slightly at the anterior and posterior ends (ag, pg); lunule and escutcheon absent (Figure 2C). Inner surface whitish and shiny with a thickened internal ridge (r) extending obliquely across the shell, anterior to the umbo. Right valve with two divergent cardinal teeth (3a, 3b) and a single anterior and posterior lateral (AI, PI) (Figures 1C, E). Left valve with three cardinal teeth: a smaller posterior-dorsal one (4b), a middle-ventral, robust, trigonal (2b), and an anterior elongated and laminate (2a), sometimes covered by periostracum (pe); including a single anterior and posterior lateral (AII, PII) (Figures 1D, F). Hinge plate narrow with a projecting spoon-shaped chondrophore (c), to which is attached a robust internal ligament (il) (Figures 1E, F). External ligament (el) brownish and short (Figures 1B, 2C). Pallial sinus deep, extends beyond the mid dorso-ventral axis of the shell, confluent with pallial line.
Mantle lobes with three marginal folds (inner, middle and outer) completely unattached antero-ventrally, forming an extensive pedal opening. The middle sensorial fold bears a single row of numerous short, digitiform mantle tentacles (mt) distributed along the mantle margins, except on the dorsal margin (Figure 2). Posteriorly, two long, cylindrical and completely separated siphons probably formed by the fusion and hypertrophy of the inner folds of the mantle, i.e. Type A (Yonge, 1948, 1982). Siphonal retractor muscles (srm) present. Inhalant siphon (is) thicker, ventral and larger than the exhalant siphon (es). Cruciform muscle (cm) near the base of the inhalant siphon forming the only fusion of the ventral mantle margin. A pair of symmetric and bilateral siphonal organs (so) located close to the proximal aperture of the inhalant siphon on both sides. Ctenidia (ct) eulamellibranch, complete with outer and inner demibranchs (id/od). Labial palps (lp) trigonal, large, similar in size with gills; outer and inner palps present. Adductor muscles unequal; the anterior (aam) thin and elongated dorsoventrally and the posterior (pam) oval. Foot (f) wide, with a ventral surface (sole), posterior pedal retractor muscle (pprm) attached to the shell close to the posterior adductor muscle, insertion of the anterior pedal retractor muscle not visible; byssal thread absent. A few aspects of the visceral mass were also observed such as digestive diverticula (dd), hind gut (hg), kidney (k) and probably ovary (ov)? and testis (te)? seen only by transparency (Figures 2D, E).
The shell features of Theora lubrica have been extensively documented in the literature, including its shape, coloration, external and internal sculpture, as well as its ligament and hinge teeth (Gould, 1861; Adams, 1864; Seapy, 1974; Boyd, 1999; Coan et al., 2000). However, it seems that for over 130 years, the hinge teeth of this species have been misinterpreted, probably due to the absence of descriptions using SEM. In virtually all recent descriptions (Fasse et al., 2019; Worsfold et al., 2020), the right valve is reported to have two cardinal teeth, while the left valve is described as having only one cardinal tooth. Our description, therefore, demonstrates for the first time that the left valve actually has three cardinal teeth (2a, 2b, 4b) (Figures 1D, F). Furthermore, it hypothesizes the numbering/labeling of all hinge teeth based on their position and appearance during ontogeny, following Carter et al. (2012). It is also important to note that the tooth characterized here as ‘4b’ has been previously observed and illustrated by Faasse et al. (2019: Figure 4) but was probably interpreted as a secondary tooth. The tooth ‘2a’, observed here for the first time, may eventually become covered by periostracum (pe) in some individuals, which could make it difficult to visualize (Figure 1D). This partly explains its absence in previous descriptions. Our literature review also indicated that T. lubrica specimens collected worldwide from 1861 to the present have a shell ranging from 2 to 16 mm in length. However, shells measuring up to 20 mm were also reported in the literature by Coan et al. (2000, p. 438), but we were unable to access these specimens.
Limited information on the anatomy of T. lubrica is available in the literature, including features on the siphons (coloration and length) of specimens from the UK (Worsfold et al., 2020), a few photos showing the soft parts through the shell’s transparency (Adarraga and Martínez, 2011: Figure 2A; Fasse et al., 2019: Figure 3; Worsfold et al., 2020: Figures 2, 3), and histological sections of gonads of individuals from New Zealand (Johnston, 2005: plates 3.1-3.3 – master thesis). Here, for the first time, detailed information on the pallial cavity organs and visceral mass is provided, bringing potential new insights into the functional morphology of T. lubrica (Figures 2D, E). Among the main anatomical characteristics of T. lubrica, the following stand out: (i) the presence of numerous small tentacles along the mantle margins, (ii) a bilateral siphonal organ, and (iii) indirect evidence of hermaphroditism.
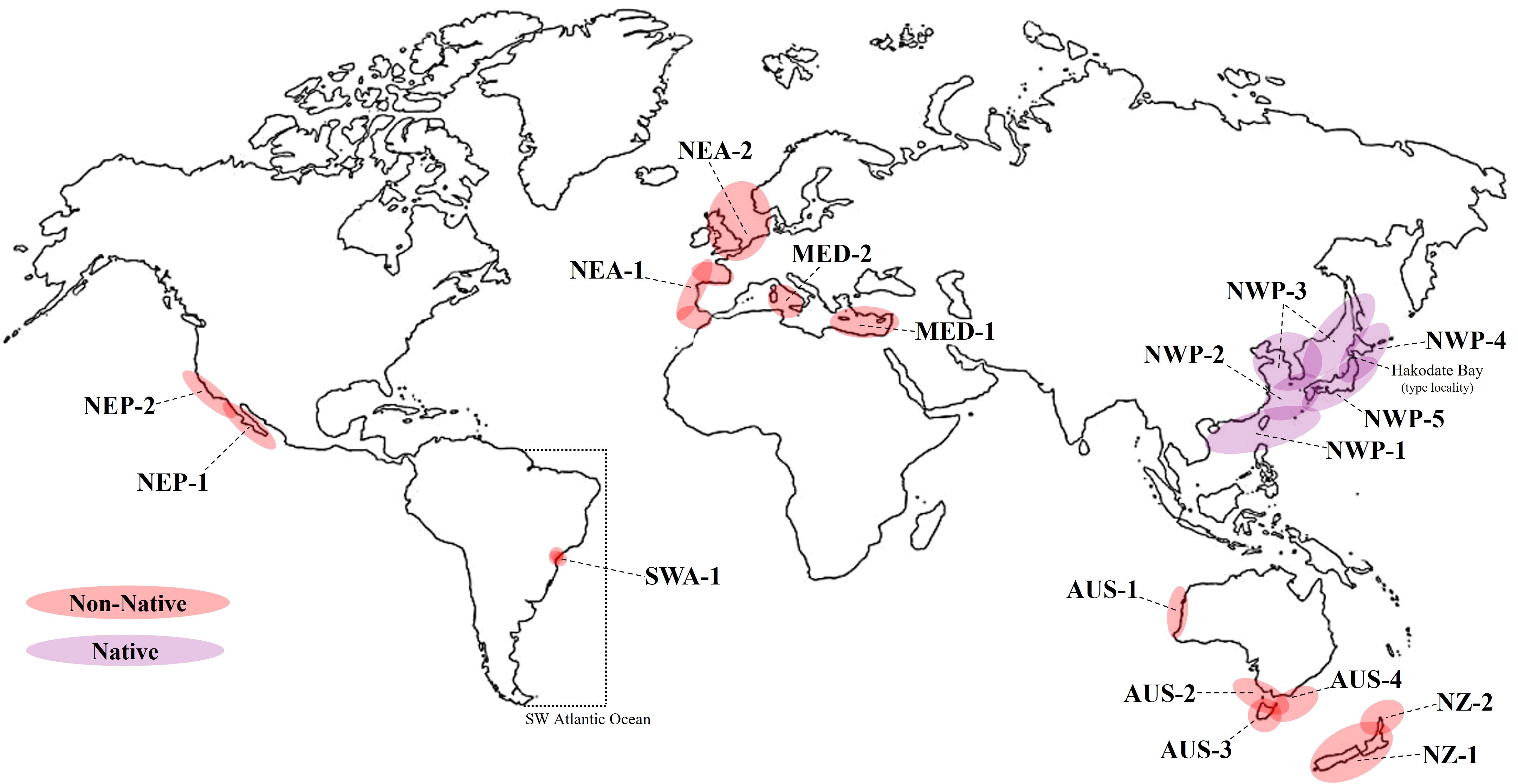
Figure 3. Updated worldwide distribution of Theora lubrica, including the first record for the Southwestern Atlantic Ocean (dotted lines). T. lubrica is considered native (purple color) to the Northwest Pacific from northern Japan and the Vladivostok area, Russia and south to the Hong Kong region. T. lubrica is considered non-native species (red color) in New Zealand (NZ-1) North Island/New Zealand/Bay of Islands (Cranfield et al., 1998; Spencer et al., 2009); NZ-2 Herekino, North Island/New Zealand/Pacific Ocean (Cranfield et al., 1998; Spencer et al., 2009); Australia (AUS-1) Perth/Australia/Cockburn Sound-Swan Estuary (Boyd, 1999); AUS-2 Victoria/Port Phillip Bay (Boyd, 1999); AUS-3 Tasmania/Georges Bay, Tasman Sea (Edgar et al., 1999); AUS-4 New South Wales/Botany Bay (Boyd, 1999); North West Pacific (NWP-1) Hong Kong/China/South China Sea; NWP-2 Amakusa, China/East China Sea, Zhoushan, Zhejiang Province (Huang, 2001) and Kyushu/Ariake Bay (Lutaenko et al., 2006); NWP-3 Hirate Hizen/Honshu/Japan/Tsukumo Bay, Sea of Japan, Honshu/Japan/Maizuru Bay Po’Haing region/South Korea/Yeongil Bay (Lutaenko et al., 2006), China/Bohai Sea (Huang, 2001), China/Yellow Sea (Huang, 2001), Russia/Possjet Bay, Sea of Japan (Lutaenko et al., 2006), Russia/Nahodka Bay, Sea of Japan (Lutaenko et al., 2006), Russia/Amursky Bay, Sea of Japan (Lutaenko et al., 2006); NWP-4 Honshu/Japan/Ofunato Bay (Lutaenko et al., 2006); NWP-5 Honshu/Japan/Tokyo Bay, Suo-Nada/Japan, Seto Inland Sea, Kochi, Shikoku/Japan/Uranouchi Bay, Japan/Tanabe Bay (Lutaenko et al., 2006); Honshu/Japan/Mikawa Bay (Lutaenko et al., 2006); Mediterranean (MED-1) Haifa/Israel/Mediterranean Sea (Bogi and Galil, 2007); MED-2 Livorno/Italy/Livorno Harbor (Tyrhennian Sea) (Zenetos et al., 2010; Occhipinti-Ambrogi et al., 2011; Crocetta, 2012); North East Atlantic (NEA-1) Bilbao, Basque Country/Spain/Nervion Estuary and Molinao/Basque Country/Spain/Pasajes Port, Gijón and Avilés/Asturias/Spain (Addaraga and Martinez, 2011; Raven and Suárez, 2024), NEA-2 Kanaal door Walcheren, West-Souburg/Netherlands, Lake Grevelingen/Netherlands/North Sea, Granton Harbour, Edinburgh-Scotland/UK and Port of Rotterdam, South Holland/Netherlands (Faasse et al., 2019; Notton, 2020; Avramidi et al., 2022); North East Pacific (NEP-1) CA/Anaheim Bay, including Sunset Bay and Huntington Harbor (Seapy, 1974, as T. fragilis); NEP-2 San Francisco/CA/Islais Creek Channel, San Francisco Bay (Cohen and Carlton, 1995) and South Western Atlantic (SWA-1) Port of Santos/Santos Estuary/municipalities of Santos, Cubatão and Guarujá in Southeastern Brazil (this study).
Generally, bivalves exhibit a great diversity of tentacles along their mantle margins, typically associated with sensory or secretory functions (Yonge, 1983; Audino and Marian, 2019). Similar to T. lubrica, other members of the superfamily Tellinoidea also have small tentacles (or papillae) along the free mantle edges such as Austromacoma biota (Arruda & Domaneschi, 2005), Tellina angrensis (Marques and Simone, 2014), Temnoconcha brasiliana (Dall, 1921) (Boss and Kenk, 1964; Piffer et al., 2011; Marques and Simone, 2014) and the confamilial species Abra alba (W. Wood, 1802), Scrobicularia plana (da Costa, 1778), Semele purpuracens (Gmelin, 1791), Semele proficua (Pulteney, 1799) and Semele trindadis Simone, 2021 (Yonge, 1949; Domaneschi, 1995; Simone, 2021). For both species, the function of these mantle tentacles is uncertain, but two hypotheses may be considered: (i) acting as sensory tentacles regulating the opening and closing of the shell during digging process or (ii) producing mucus to aid in the removal of excess and rejected particles from the pallial cavity.
For the first time a pair of siphonal organs was observed in a member of the family Semelidae. Theora lubrica is, therefore, the first semelid known to possess a bilateral siphonal organ, a condition previously observed only in Macoma biota (Tellinidae) (Piffer et al., 2011). The presence of a single asymmetric siphonal organ appears to be a more common condition among Tellinoidea, having received different names since its first description by Pelseneer (1911). The terms ‘valvulae palleale’, ‘siphonal membrane’, ‘mantle folds’, ‘parasiphonal organ’ and ‘pseudogills’ have also been used to refer to a possibly homologous structure among members of this superfamily (Pelseneer, 1911; Kellogg, 1915; Yonge, 1949; Boss and Kenk, 1964; Simone and Wilknson, 2008). The function of the siphonal organs was first elucidated by Kellogg (1915) in Rexithaerus secta (Conrad, 1837), demonstrating that it assists in the removal of waste matter. Almost 100 years later Piffer et al. (2011) in M. biota, also attributes the same function to these organs, “the siphonal organs increase the superficial area of the mantle, agglutinate a large quantity of particles eliminated by the pallial organs and convey this material to the waste channel”. This is a crucial function in deposit-feeders like Theora lubrica, where large amounts of sediment and pseudofeces often accumulate in the pallial cavity.
Theora lubrica was characterized as a dioecious species by Saito et al. (1998) and Johnston (2005); however, the individuals sampled in Brazil apparently exhibit both types of gonads (ovaries and testes) simultaneously (Figure 2D). It is important to note that this potential hermaphroditic condition is based solely on an analysis of gross anatomy and still needs future confirmation.
Although live specimens of T. lubrica were not analyzed in this study, a brief review of key aspects of its ecology is provided. Essential information on the species’ population dynamics and biology is compiled, which may support the future development and implementation of methods for the control and/or eradication of this invasive alien species in the Global South.
Theora lubrica (or Asian Semele) is a small, infaunal bivalve that typically occurs in soft, muddy subtidal or lower intertidal sediments rich in organic matter. It is buried at depths of 10 to 30 mm in the substrate and feeds on benthic algae and detritus (Hayward et al., 1999). The maximum burrowing depth suggests that in living specimens the relatively long and thin siphons of T. lubrica can extend two to three times the length of the shell. This species tolerates high variations in salinity and can therefore be classified as a marine (30-40 ppt) or estuarine (18-30 ppt) (Boyd, 1999; Saito et al., 1998; Johnston, 2005; Inglis et al., 2006; the present study). Their populations often exhibit significant fluctuations in density, ranging from 30-70 individuals/m² to as many as 2,000/m² (Imabayashi and Wakabayashi, 1992; Hayward et al., 1999). Such fluctuations, observed in New Zealand and Japan, appear to be strongly linked to water and sediment temperature, as well as low oxygen levels, with faster growth rates in summer compared to winter and significant diebacks during the warmer months. Considerable population declines have also been linked to high predation rates by a mud crab in New Zealand (Hayward et al., 1999).
T. lubrica is a fast-growing dioecious species; its individuals can mature in 1 to 3 months, at about 4-7 mm (Saito et al., 1998; Johnston, 2005). The larval prodissoconch size (PI = 80 ± 10µm) of individuals collected in Brazil indicates that T. lubrica probably has an indirect and planktotrophic development (see Ockelmann, 1965). Although there is no record of studies on the development of T. lubrica, Tamai (1996) showed that the congener Theora lata (Hinds, 1843) has planktotrophic larvae that spend about 13 days in the plankton at 24°C. The Asian Semele is also a short-lived species that can rapidly colonize disturbed and muddy habitats and is among the most pollution-tolerant mollusks found in some harbors in New Zealand, thriving even in contaminated sediments (Hayward, 1997). Due to its frequent dominance in highly polluted environments, T. lubrica is regarded as a pollution-indicator species.
Although T. lubrica has been reported as one of the two worst invasive mollusks in Europe by Adarraga and Martínez (2011), no ecological or economic impacts have yet been documented for this species. Even so, it is worth noting that species with the invasive potential of T. lubrica can cause aggressive colonization in short periods of time, gradually leading to global biotic homogenization of the environment (Shiganova, 2010). According to Hayward (1997), ‘billions’ of individuals were observed living in Waitemata Harbour – New Zealand, where T. lubrica is one of the few species capable of tolerating the polluted sediments.
Theora lubrica is native to the Northwest Pacific (NWP), ranging from northern Japan (Hakodate Bay, type locality) and the Vladivostok area in Russia, to the Hong Kong region (Gould, 1861; Huang, 2001; Lutaenko et al., 2006) but over the last 60 years it has been constantly introduced into various parts of the world (Seapy, 1974; Crooks, 1998; Ranasinghe et al., 2005; Bogi and Galil, 2007; Adarraga and Martínez, 2011; Zenetos et al., 2010; Faasse et al., 2019; Notton, 2020). Figure 3 provides an up-to-date overview of the distribution of this species, including all known localities around the globe. Currently T. lubrica is spread mainly in port regions of Australia (AUS), New Zealand (NZ), Northeast Pacific (NEP), Mediterranean (MED), Northeast Atlantic (NEA) and now in the Southwestern Atlantic (SWA). Although the new record was made for Southeast Brazil, it is worth noting that individuals of T. lubrica from Northeast Brazil were also identified by FMM and CMS, first and second author respectively. In both cases, ballast water is probably the main factor responsible for the introduction of this exotic species into Brazilian waters.
Like many other invasive species, although it does not exhibit phenotypic plasticity, T. lubrica displays other typical traits of r-strategists, such as small body size, early sexual maturity, rapid gametogenesis, an almost continuous breeding season with multiple cohorts, planktotrophic larvae that can potentially remain in the water column for up to 20 days, opportunistic and gregarious behavior, and a wide tolerance to environmental stress (Morton, 1997; Lavoie et al., 1999).
Despite being well established in the literature, the concept of invasive species (IUCN, 2021) can be difficult to apply, often due to taxonomic issues or the lack of ecological studies for some species. However, given its small shell size, which hinders identification, and its invasive potential, there is no doubt that T. lubrica should be considered an invasive species that may have already spread across much of Brazil and other countries in South America. In this way, both alien species monitoring, and ballast water treatment should be carried out continuously to prevent the spread of T. lubrica throughout the Global South, including the warmer waters of the Caribbean. Currently, among the main port countries in South America, only Brazil and Argentina are signatories to the Ballast Water Management Convention (BWMC) – a treaty adopted to help prevent the spread of potentially harmful aquatic organisms and pathogens in ships’ ballast water (IMO, 2024). Therefore, broad adherence to this treaty is essential for preserving South America’s native aquatic fauna and avoiding the potential introduction of other invasive aquatic species.
Finally, this new record of T. lubrica expands the list of invasive species in the Southwest Atlantic and helps fill part of the huge knowledge gap on alien species in the Global South.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because studies involving small, exotic, and abundant marine invertebrates in Brazil do not require ethical approval.
FM: Conceptualization, Data curation, Methodology, Supervision, Writing – original draft, Writing – review & editing. CS: Writing – review & editing, Data curation. FO: Writing – review & editing, Data curation. MM: Writing – review & editing. TV: Writing – review & editing. VC: Funding acquisition, Project administration, Writing – review & editing. AL: Writing – review & editing, Funding acquisition, Project administration. CY: Writing – review & editing. RS: Writing – review & editing. LN-J: Writing – review & editing, Funding acquisition, Project administration. LO: Writing – review & editing, Funding acquisition, Project administration.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study received funding from Port of Santos, Tetra Tech and Econsult Estudos Ambientais.
The authors are grateful to the Senckenberg Ocean Species Alliance (SOSA) for providing access to electron and optical microscopy facilities, as well as to Jan Steger for his technical advice. Special thanks are also extended to Michela Borges (MDBio/UNICAMP) for granting access to microscopy facilities during the initial stage of the project. Special thanks are due to Tetra Tech América do Sul and Porto de Santos for providing access to the samples. Thanks are due to IBAMA (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis) for supporting the ongoing monitoring of exotic species along the Brazilian coast. We also thank the entire team at the benthic invertebrate screening laboratory and the technical team at Econsult Estudos Ambientais.
Authors CS, FO, MM, TV, and VC were employed by company Econsult Estudos Ambientais. Authors AL, CY, and RS were employed by company Tetra Tech. Authors LN-J and LO were employed by company Port of Santos.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that this study received funding from Tetra Tech, Econsult Estudos Ambientais and Port of Santos. The funders had the following involvement in the study: collection, analysis and interpretation of data and decision to submit it for publication.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adams A. (1864). On the species of Neaera found in the seas of Japan. Ann. Mag. Nat. Hist. 3, 206–209. doi: 10.1080/00222936408681598
Adarraga I., Martínez J. (2011). First record of invasive Theora lubrica (Mollusca: Bivalvia: Semelidae) in the Atlantic Ocean. Mar. Biodivers. Rec. 4, 1–7. doi: 10.1017/S1755267211001011
Audino J. A., Marian J. E. A. R. (2019). Form and function of tentacles in pteriomorphian bivalves. J. Morphol. 281, 33–46. doi: 10.1002/jmor.21077
Avramidi E., Gómez S. C. G., Papaspyrou S., Louca V., Xevgenos D., Küpper F. C. (2022). Benthic biodiversity near brine discharge sites in the Port of Rotterdam. Water Resour. 27, 100173. doi: 10.1016/j.wri.2022.100173
Beltrão M., Cunha N. J. R., Laaf Y. O., dosSantos T. D., Diehl F. (2024). Molecular methods confirm the presence of the alien mussel Perna viridis Linnaeus 1758 (Bivalvia, mytilidae) in southern Brazil. SciELO. doi: 10.1590/SciELOPreprints.8889
Bogi C., Galil B. S. (2007). First record of Theora (Endopleura) lubrica Gould 1861 (Mollusca: bivalvia: semelidae) from a levantine port. Aquat. Invasions. 2 (1), 77–79. doi: 10.3391/ai.2007.2.1.11
Boltovskoy D., Correa N. (2015). Ecosystem impacts of the invasive bivalve Limnoperna fortunei (golden mussel) in South America. Hydrobiologia 746, 81–95. doi: 10.1007/s10750-014-1882-9
Boss K. J., Kenk V. C. (1964). Anatomy and relationships of Temnoconcha brasiliana. Occas. Pap. Mollusks. 2, 325–343. https://www.biodiversitylibrary.org/part/241598.
Boyd S. (1999). Introduced mollusca of port phillip bay. In: Hewitt C. L., Campbell M. L., Thresher R. E., Martin R. B. (eds) Marine biological invasions of Port Phillip Bay, Victoria. Centre for Research on Introduced Marine Pests Tech Rep No.20. CSIRO Marine Research, Hobart, pp 129–149.
Carter J. G., Harries P., Malchus N., Sartori A., Anderson L., Bieler R., et al. (2012). Treatise Online no. 48: Part N, Revised, Volume 1, Chapter 31: Illustrated Glossary of the Bivalvia. Treatise Online. 10.17161/to.v0i0.4322
Castro M. C., Fileman T. W., Hall-Spencer J. M. (2017). Invasive species in the Northeastern and Southwestern Atlantic Ocean: A review. Mar. Pollut. Bull. 116 (1-2), 41–47. 10.1016/j.marpolbul.2016.12.048
Coan E. V., Valentich-Scott P., Bernard F. R. (2000). Bivalve seashells of western north america (Santa Barbara: Santa Barbara Museum of Natural History).
Cohen A. N., Carlton J. T. (1995). Biological study of nonindigenous aquatic species in a United States estuary: A case study of the biological invasions of the san francisco bay and delta, SFEI contribution no. 185. U.S Vol. 218 (Washington, DC: Fish and Wildlife Service).
Companhia Ambiental do Estado de São Paulo - CETESB, Agência Nacional de Águas – ANA (2023). Guia nacional de coleta e preservação de amostras: água, sedimento, comunidades aquáticas e efluentes líquidos, 2a ed Vol. 326 (São Paulo: CETESB; Brasília: ANA).
Costello K. E., Lynch S. A., McAllen R., O'Riordan R. M., Culloty S. C. (2022). Assessing the potential for invasive species introductions and secondary spread using vessel movements in maritime ports. Mar. pollut. Bull. 177, 113496. doi: 10.1016/j.marpolbul.2022.113496
Cranfield H. J., Gordon D. P., Willan R. C., Marshall B. A., Battershill C. N., Francis M. P., et al. (1998). Adventive marine species in New Zealand. NIWA Tech. Rep. 34, 48.
Crocetta F. (2012). Marine alien Mollusca in Italy: a critical review and state of the knowledge. J. Mar. Biol. Assoc. U.K. 92, 1357–1365. doi: 10.1017/S002531541100186X
Crooks J. A. (1998). Habitat alteration and community-level effects of an exotic mussel, Musculista senhousia. Mar. Ecol. Prog. Ser. 162, 137–152. doi: 10.3354/meps162137
Dias T. L. P., Mota E., Almeida S., Gondim A. I., Rabelo E., Oliveira J., et al. (2013). Isognomon bicolor (C.B. Adams 1845) (Mollusca: Bivalvia): First record of this invasive species for the states of Paraíba and Alagoas and new records for other localities of Northeastern Brazil'. Check List 9, 157–161. doi: 10.15560/9.1.157
Domaneschi O. (1995). A comparative study of the functional morphology of Semele purpurascens (Gmelin 1791) and semele proficua (Pulteney 1799) (Bivalvia: semelidae). Veliger 38, 323–324.
Edgar G. J., Barrett N. S., Last P. R. (1999). The distribution of macroinvertebrates and fishes in Tasmanian estuaries. J. Biogeogr. 26, 1169–1189. doi: 10.1046/j.1365-2699.1999.00365.x
Endresen O., Behrens H. L., Brynestad S., Andersen A. B., Skjong R. (2004). Challenges in global ballast water management. Mar. pollut. Bull. 48, 615–623. doi: 10.1016/j.marpolbul.2004.01.016
Faasse M. A., Gheerardyn H., Morys C., van Haaren T., Ysebaer T., Nijland R. (2019). The non-indigenous window shell Theora lubrica Gould (1861) (Bivalvia: Cardiida: Semelidae) in the delta area of the Netherlands. Basteria 83, 53–58.
Fernandes M. R., Suga C. M., Marques M. M., Salgueiro F. (2024). From port to port: the invasive bivalve Mytilopsis cf. sallei (Dreissenidae) is confirmed in southeastern Brazil. Molluscan Research 44 (3), 229–240. doi: 10.1080/13235818.2024.2330109
Gould A. A. (1861). Description of new shells collected by the United States North Pacific Exploring Expedition. Proc. Boston Soc Nat. Hist. 7, 385–389. doi: 10.5962/bhl.part.4821
Hayward B. W. (1997). Introduced marine organisms in New Zealand and their impact in the Waitemata Harbour, Auckland. Tane 36, 197–223.
Hayward B. W., Morley M. S., Hayward J. J., Stephenson A. B., Blom W. M., Hayward K. A., et al. (1999). Monitoring studies of the benthic ecology of Waitemata Harbour, New Zealand. Rec. Auckland Inst. Mus. 36, 95–117. http://www.jstor.org/stable/42905839
Huang Z. (2001). Marine species and their distribution in China's seas Vol. 598 (Florida, USA: Vertebrata. Smithsonian Institution), 404–463.
IMO (International Maritime Organization) (2024). Implementing the ballast water management convention. Available online at: https://www.imo.org/en (Accessed October 11, 2024).
Inglis G. J., Hurren H., Oldman J., Haskew R. (2006). Using habitat suitability index and particle dispersion models for early detection of marine invaders. Ecol. Appl. 16, 1377–1390. doi: 10.1890/1051-0761(2006)016[1377:uhsiap]2.0.co;2
IUCN (International Union for Conservation of Nature) (2021). Invasive alien species and climate change. Available online at: https://www.iucn.org/resources/issues-brief/invasive-alien-species-and-climate-change (Accessed October 11, 2024).
Imabayashi H., Wakabayashi M. (1992). Production and respiration of a bivalve Theora lubrica in northern Bingo-Nada, the Seto Inland Sea. Journal of the Faculty of Applied Biological Science, Hiroshima University 31, 45–57.
Johnston O. R. (2005). Distribution and biology of the marine invasive bivalve Theora lubrica (Semelidae): 1-213. Thesis, University of Canterbury, New Zealand.
Kellogg J. L. (1915). Ciliary mechanisms of lamellibranchs with descriptions of anatomy. J. Morphol. 26, 625–701. doi: 10.1002/jmor.1050260403
Lavoie D. M., Smith L. D., Ruiz G. M. (1999). The potential for intracoastal transfer of nonindigenous species in the ballast water of ships. Estuar. Coast. Shelf. Sci. 48, 551–564. doi: 10.1006/ecss.1999.0467
Linares M. S., Callisto M., Marques J. C. (2017). Invasive bivalves increase benthic communities complexity in neotropical reservoirs. Ecol. Indic. 75, 279–285. doi: 10.1016/j.ecolind.2016.12.046
Lutaenko K. A., Je J., Shin S. (2006). Bivalve mollusks in Yeongil Bay, Korea: 2 Faunal analysis. Korean J. Malacol. 22, 61–82. doi: 10.4217/OPR.2003.25.2.155
Machado F. M., Miranda M. S., Salvador R. B., Pimenta A. D., Côrtes M. O., Gomes J. A. J., et al. (2023). How many species of Mollusca are there in Brazil? A collective taxonomic effort to reveal this still unknown diversity. Zoologia 40, e23026. doi: 10.1590/S1984-4689.v40.e23026
Machado F. M., Simone L. R. L., Miranda M. S., Salvador R. B., Pimenta A. D., Côrtes M. O., et al. (2024). Mollusca in taxonomic catalog of the Brazilian fauna (TCBF). Available online at: http://fauna.jbrj.gov.br/fauna/listaBrasil/ConsultaPublicaUC/ConsultaPublicaUC.do (Accessed October 11, 2024).
Marques R., Simone L. R. L. (2014). Eurytellina angrensis, a new species from southeastern Brazilian coast, with anatomical details (Bivalvia: Tellinoidea). Arch. Molluskenkd. 143, 39–50. doi: 10.1127/arch.moll/1869-0963/143/039-050
MMA (2009). Informe sobre as Espécies Exóticas Invasoras Marinhas no Brasil (Brasília, DF: Ministério do Meio Ambiente. Editor Científico). Rubens M. Lopes (IO-USP).
Morton B. (1997). The aquatic nuisance species problem: a global perspective and overview. D’Itri F. M. editor. Zebra Mussels and Aquatic Nuisance Species. (Chelsea, Michigan: Ann Arbor Press) 1–54.
Notton D. G. (2020). Theora lubrica (Mollusca, Semelidae) new to Scotland'. Mollusc World. 53, 23. doi: 10.3391/ai.2013.8.1.02
Occhipinti-Ambrogi A., Marchini A., Cantone G., Castelli A., Chimenz C., Cormaci M., et al. (2011). Alien species along the Italian coasts: an overview. Biol. Invasions. 13, 215–237. doi: 10.1007/s10530-010-9803-y
Ockelmann K. W. (1965). Developmental types in marine bivalves and their distribution along the Atlantic Coast of Europe. Proc. Eur. Malac. Congr. 1st London, 25–35.
Orensanz J. M. L., Schwindt E., Pastorino G., Bortolus A., Casas G., Darrigran G., et al. (2002). No longer the pristine confines of the world ocean: a survey of exotic marine species in the southwestern Atlantic. Biol. Invasions 4, 115–143.
Pelseneer P. (1911). Les lamellibranchs de l’expédition du Siboga, partie anatomique. Siboga Expéditie Monographie 53a, 1–125.
Piffer P. R., Arruda E. P., Passos F. D. (2011). The biology and functional morphology of Macoma biota (Bivalvia, Tellinidae, Macominae). Zoologia 28, 321–333. doi: 10.1590/S1984-46702011000300006
Queiroz R. N. M., Dias T. L. P., Batista R., Silva P. M. (2022). Reproduction and population dynamics of the invasive bivalves Mytilopsis sallei and Isognomon bicolor on the Northeast coast of Brazil. Zoology 153, 126028. doi: 10.1016/j.zool.2022.126028
Ranasinghe J. A., Mikel T. K., Velarde R. G., Weisberg S. B., Montagne D. E., Cadien D. B., et al. (2005). The prevalence of non-indigenous species in southern California embayments and their effects on benthic macroinvertebrate communities. Biol. Invasions. 7, 679–686. doi: 10.1007/s10530-004-5857-z
Raven J. G. M., Suárez A. A. (2024). First records of the introduced marine bivalve Theora lubrica A. Gould 1861 from Asturias, NW Spain (Semelidae). Basteria 88, 225–228.
Rotundo M. M., Gaulia L. A., Cardoso G. S., Carminatto A. A., Henrique H. S., Reigada A. L. D., et al. (2020). Ichthyofauna from Santos-São Vicente upper estuary: a study before and during fire at Santos port terminal. Res. Soc Dev. 9, 8901. doi: 10.33448/rsd-v9i10.8901
Roversi F., Rosman P. C. C., Harari J. (2016). Análise da renovação das águas do Sistema Estuarino de Santos usando modelagem computacional'. Ambiente e Água - Interdiscip. J. Appl. Science. 11, 566–585. doi: 10.4136/ambi-agua.1770
Saito H., Ueno M., Hayashi I. (1998). “Temporal fluctuation in the abundance of a semelid bivalve, Theora fragilis (A. Adams) in Maizuru Bay, Sea of Japan,” in Recruitment, colonization and physical-chemical forcing in marine biological systems, developments in hydrobiology, vol. 132 . Eds. Baden S., Phil L., Rosenberg R., Strömberg J. O., Svane I., Tiselius P. (Springer, Dordrecht). doi: 10.1007/978-94-017-2864-5_13
Santos Port Authority (2024). Connecting Brazil worldwide – maps and figures. Available online at: https://www.portodesantos.com.br/en/ (Accessed October 11, 2024).
Seapy R. R. (1974). The introduced semelid bivalve Theora (Endopleura) lubrica in bays of Southern California. Veliger. 16, 385–387.
Seebens H., Gastner M. T., Blasius B. (2013). The risk of marine bioinvasion caused by global shipping. Ecol. Lett. 16, 782–790. doi: 10.1111/ele.12111
Shiganova T. (2010). Biotic homogenization of inland seas of the Ponto-Caspian. Annu. Rev. Ecol. Evol. Syst. 41, 103–125. doi: 10.1146/annurev.ecolsys.110308.120148
Simone L. R. L. (2021). A new species of Semele from Trindade oceanic island, Brazil (Tellinoidea: Semelidae). Arch. Molluskenkd. 150, 79–86. doi: 10.1127/arch.moll/150/079-086
Simone L. R. L., Gonçalves E. P. (2006). Anatomical study on Myoforceps aristatus, an invasive boring bivalve in SE Brazilian coast (Mytilidae). Pap. Avulsos Zool. 46, 57–65. doi: 10.1590/S0031-10492006000600001
Simone L. R. L., Wilkinson S. (2008). Comparative morphological study of some Tellinidae from Thailand (Bivalvia: Tellinoidea). Raffles Bull. Zool. 18, 151–190. https://lkcnhm.nus.edu.sg/wp-content/uploads/sites/11/app/uploads/2017/06/s18rbz151-190.pdf.
Simpson T. J. S., Smale D. A., McDonald J. I., Wernberg T. (2017). Large scale variability in the structure of sessile invertebrate assemblages in artificial habitats reveals the importance of local-scale processes. J. Exp. Mar. Biol. Ecol. 494, 10–19. doi: 10.1016/j.jembe.2017.05.003
Spencer H., Marshall B., Maxwell P., Grant-Mackie J., Stilwell J. D., William R., et al. (2009). “Phylum Mollusca: Chitons, clams, tusk shells, snails, squids and kin',” in New zealand inventory of biodiversity. Ed. Gordon D. (Canterbury University Press, Christchurch), 161–254.
Tamai K. (1996). Temporal tolerance of larval Theora fragilis (Bivalvia: Semelidae) to hypoxic conditions. Fisheries Sci. 62, 996–997. doi: 10.2331/fishsci.62.996
Worsfold T. M., Pennisi N., Ashelby C. W. (2020). Theora lubrica Gould, 1861 (Bivalvia: Semelidae), new to the UK, with notes on associated non-native species, and an earlier date of introduction for Arcuatula senhousia (Bivalvia: Mytilidae) to the UK. Journal of Conchology 43, 665–674.
Yonge C. M. (1948). Formation of siphons in lamellibranchia'. Nature 161, 198–199. doi: 10.1038/161198a0
Yonge C. M. (1949). On the structure and adaptations of the Tellinacea, deposit-feeding Eulamellibranchia. Phil. Trans. R. Soc Lond. B. 234, 29–76. doi: 10.1098/rstb.1949.0006
Yonge C. M. (1982). Mantle margins with a revision of siphonal types in the Bivalvia. J. Molluscan Stud. 48, 102–103. doi: 10.1093/oxfordjournals.mollus.a065609
Yonge C. M. (1983). Symmetries and the role of the mantle margins in the Bivalve Mollusca. Natl. Widl. 16, 1–10.
Zenetos A., Gofas S., Verlaque M., Cinar M., Garcia Raso J., Bianchi C., et al. (2010). Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution'. Mediterr. Mar. Sci. 11, 381–493. doi: 10.12681/mms.87
Keywords: non-native bivalves, Asian Semele, Mollusca, shell morphology, gross anatomy, polyhaline mollusks, biomonitoring and environmental monitoring
Citation: Machado FM, Suga CM, Onodera FKC, Melo MS, Viti T, Cavinatto VM, Lammardo ACR, Yonamine CY, Sáfadi RS, Nogueira-Junior LA and Oliva LFM (2025) Spreading further: the first record of the alien bivalve Theora lubrica in the Southwestern Atlantic Ocean. Front. Mar. Sci. 12:1511313. doi: 10.3389/fmars.2025.1511313
Received: 14 October 2024; Accepted: 31 January 2025;
Published: 27 February 2025.
Edited by:
Katherine Dafforn, Macquarie University, AustraliaReviewed by:
Francesco Tiralongo, University of Catania, ItalyCopyright © 2025 Machado, Suga, Onodera, Melo, Viti, Cavinatto, Lammardo, Yonamine, Sáfadi, Nogueira-Junior and Oliva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fabrizio Marcondes Machado, ZmFicml6aW9tYXJjb25kZXNAeWFob28uY29tLmJy
†ORCID: Fabrizio Marcondes Machado, orcid.org/0000-0002-5085-865X
Cristiane Midori Suga, orcid.org/0009-0009-8598-3504
Fabio Kiyoshi Camara Onodera, orcid.org/0009-0002-7834-1027
Mariana Sousa Melo, orcid.org/0000-0002-5546-2718
Thais Viti, orcid.org/0009-0008-9362-9030
Vilma Maria Cavinatto, orcid.org/0009-0009-1982-7407
Ana Carolina da Rocha Lammardo, orcid.org/0009-0004-8963-0638
Carolina Yumi Yonamine, orcid.org/0009-0002-5826-6963
Regina Sawaia Sáfadi, orcid.org/0009-0001-6482-9796
Luis Antonio Nogueira-Junior, orcid.org/0009-0002-1876-1820
Luiz Fernando Maciel Oliva, orcid.org/0009-0003-2857-6239
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.