 Johann D. Lankes1,2*†
Johann D. Lankes1,2*† Heather N. Page3†
Heather N. Page3† Amanda Quasunella4Jaclyn F. Torkelson5Clöe Lemaire6
Amanda Quasunella4Jaclyn F. Torkelson5Clöe Lemaire6 Robert J. Nowicki7Lillia Blasius8
Robert J. Nowicki7Lillia Blasius8 Emily R. Hall1,9
Emily R. Hall1,9- 1Elizabeth Moore International Center for Coral Reef Research and Restoration, Mote Marine Laboratory, Summerland Key, FL, United States
- 2Isometric, London, United Kingdom
- 3Sea Education Association, Woods Hole, MA, United States
- 4Southern Cross University, Lismore, NSW, Australia
- 5Department of Chemistry and Biochemistry, Northern Arizona University, Flagstaff, AZ, United States
- 6Department of Marine and Environmental Science, Savannah State University, Savannah, GA, United States
- 7Storm Anchor Insights, Key West, FL, United States
- 8Department of Biological Science, Purdue University, West Lafayette, IN, United States
- 9Ocean Acidification Research Program, Mote Marine Laboratory, Sarasota, FL, United States
Increasingly frequent large-scale pelagic Sargassum algae blooms in the Atlantic have become a problem for coastal ecosystems. The mass decay of these blooms reduces water quality for coastal flora and fauna. However, the effects of living Sargassum blooms on seawater quality and consequently coral reef ecosystems that rely on delicately balanced carbonate chemistry are more ambiguous. Future oceans are predicted to be more acidic as additional anthropogenic CO2 emissions are absorbed, potentially tipping the balance in favor of algal blooms at the cost of coral survival. This study aimed to simulate the indirect interaction between pelagic Sargassum spp. and Acropora cervicornis coral fragments from the Florida Reef in current-day and future ocean pH conditions over the course of 70 days in a mesocosm experimental system. Measurements of coral growth and health via buoyant weight and Pulse Amplitude Modulated (PAM) fluorescence measurements reveal an unexpected coral-algal interaction. After 1 month, coral growth was significantly reduced under ocean acidification conditions and exposure to Sargassum; at the same time quantum yield and maximum electron transport rate of photosynthesis were increased relative to control counterparts in ambient and future pH scenarios by up to 14% and 18% respectively. These improvements in photosynthetic efficiency did not translate to significant differences in growth by the final measurement time point. In addition, the presence of Sargassum spp. did not raise seawater pH in the system, raising questions about how it benefited photosynthetic efficiency in exposed corals. Heterotrophy of detrital algal matter is suspected to compensate for impaired photosynthesis of pH stressed corals. Therefore, despite their current negative reputation, Sargassum blooms could provide short term localized benefits to corals in present and future ocean conditions.
1 Introduction
Caribbean coral reefs, including the Florida Reef, are increasingly affected by local events and global climate change. One of these regional impacts is the growing prevalence of pelagic Sargassum algal species (S. natans and S. fluitans, simply referred to as Sargassum from here on) which are only found in the Atlantic Ocean basin and are carried by currents into the Caribbean Sea, Gulf of Mexico, and Florida Straits (Wang et al., 2019). Harmful benthic and pelagic algal blooms, such as these, are occurring with increasing frequency, duration and scale globally with other similar events recorded in the Southern Atlantic and Northwestern Pacific regions (Sissini et al., 2017; Xiao et al., 2021). For example, pelagic Sargassum was sighted in 46% of shipboard observations in 2011-2020 across the Western Atlantic with higher prevalence in the Gulf of Mexico/Florida Straits region (Goodwin et al., 2022). The interannual variation and increasing density of Sargassum in recent decades (Johnson et al., 2013; Goodwin et al., 2022) may be primarily due to higher nutrient concentrations (i.e., from agricultural runoff, changes in mixed layer depth, and/or Saharan dust) and warmer temperatures (Gower and King, 2011; Wang et al., 2019).
While studies on live pelagic Sargassum highlight potential ecosystem benefits of blooms, such as providing habitats and nurseries for marine species (Wells and Rooker, 2004) and feeding particulate organic matter (POM) into the biological carbon pump through detrital shedding (Baker et al., 2018; Zhao et al., 2023), many studies have investigated the negative impacts of decaying Sargassum algae on coastal communities and shallow-water marine ecosystems. In 2011, high densities of beached pelagic Sargassum led to decreased tourism which in turn harmed local economies (Louime et al., 2017). Inorganic heavy metals and organic pollutants have been known to contaminate Sargassum through bioaccumulation, and when stressed, Sargassum may leach micropollutants such as chlordecone and arsenic into surrounding environments (Devault et al., 2022). Decaying Sargassum also reduces water quality by lowering pH and dissolved oxygen to hypoxic conditions as well as increasing concentrations of hydrogen sulfide and organic matter (van Tussenbroek et al., 2017). This reduction of water quality can lead to mass mortality events of coastal marine fishes, crustaceans, echinoderms, molluscs, and polychaetes (Rodríguez-Martínez et al., 2019). Larval coral swimming behavior (Antonio-Martínez et al., 2020) and adult survivorship (van Tussenbroek et al., 2017) also are impacted by decaying Sargassum which may lead to reduced reef resilience in the future. However, prior to decay, Sargassum may provide benefits to coral reefs by acting as a buffer against global climate change impacts.
Climate change (ocean warming and acidification) will have widespread negative impacts on coral physiology which can scale up to influence reef ecosystem health. Caribbean reefs have experienced warming along with increased frequency and duration of marine heat waves over the past century (Orr et al., 2005; Muñiz-Castillo et al., 2019; Bove et al., 2022). Heat stress has caused widespread coral bleaching and mortality (Hughes et al., 2018; Sully et al., 2019; Virgen-Urcelay and Donner, 2023) and may be linked to the increased prevalence of coral diseases (Miller and Richardson 2015; van Woesik and Randall, 2017). Ocean acidification (OA) shifts seawater carbonate chemistry equilibrium to favor net calcium carbonate (CaCO3) dissolution rather than precipitation. Given that reef-building corals deposit CaCO3 in the form of aragonite to build their skeletons (Farfan et al., 2022), decreased carbonate availability and seawater pH under OA decreases coral biomineralization (Erez et al., 2011). The reduction in coral calcification rates and increased CaCO3 dissolution rates can result in loss of reef accretion and flattening (Alvarez-Filip et al., 2011; Perry et al., 2013; Toth et al., 2015). The combined impacts of future ocean warming and OA on coral reefs are projected to cause declines in global reef carbonate production (i.e., reef accretion) by 2100 under every future CO2 emission scenario (Cornwall et al., 2021).
Macroalgae, such as Sargassum, may buffer against these impacts by locally increasing seawater pH and shading, or accelerate reef health decline through altered competitive interactions. Pelagic Sargassum algae can provide localized shading which has been shown to reduce the prevalence of coral bleaching and disease (Muller and van Woesik, 2009; van Woesik and McCaffrey, 2017). Benthic macroalgae can buffer against acidification through daytime net community production (i.e., photosynthesis) (Jompa and McCook, 2003; Page et al., 2016; Doo et al., 2020). Fleshy macroalgae also tend to maintain or increase growth and photosynthesis under acidification depending on their carbon affinity and uptake mechanism (Cornwall et al., 2017). Pelagic and benthic Sargassum has been shown to adapt to and even benefit from future ocean scenarios (Xu et al., 2017; Kumar et al., 2017; Gao et al., 2019). Additionally, the outcome of coral-algal interactions may be modified under acidification to favor macroalgae (McManus and Polsenberg, 2004; Diaz-Pulido et al., 2011; del Monaco et al., 2017), though this phenomenon is not consistent or well understood (Brown et al., 2018; Rölfer et al., 2021; Page et al., 2021). More research is needed to fully understand how coral-algal interactions may be modified under changing environmental conditions across species and regions.
The Florida Reef was once dominated by Acroporid corals which supported a variety of socio-ecological services (Cramer et al., 2020). However, over 80% of the Acroporid population has disappeared due to human impacts, historical disease outbreaks, hurricane damage, and bleaching events (Aronson & Precht, 2016; Brandt, 2009; Walton et al., 2018; Cramer et al., 2020) with Acropora cervicornis in particular taking on great losses (Muller et al., 2021; Dilworth et al., 2024). Consequently, these species are now listed as threatened under the United States Endangered Species Act (Miller and Chiappone, 2013) and critically endangered under the International Union for Conservation of Nature Red List of Threatened Species (Crabbe et al, 2022). Many coral restoration programs throughout the Caribbean are focused on recovering these declining populations to restore historical ecosystem functions of these reefs (Young et al., 2012; Schopmeyer et al., 2017), including at the location of this study, in the lower Florida Keys. Therefore, it is of particular interest to investigate how future ocean conditions will influence local and global coral restoration programs. We investigated the interaction between pelagic Sargassum and OA (reduced seawater pH) on growth and physiology of A. cervicornis over two months to determine how this coral-algal relationship may interact with future ocean conditions.
2 Materials and methods
2.1 Experimental system
All experiments took place at Mote Marine Laboratory’s Elizabeth Moore International Center for Coral Reef Restoration and Research (IC2R3) on Summerland Key, Florida. Live corals and Sargassum algae were maintained in the Climate and Acidification Ocean Simulator (CAOS), an outdoor experimental system which supplies CO2-treated water from header tanks (to control pH) to flow-through aquaria situated within temperature-controlled raceways. Prior to treatment, bulk seawater is pumped into the CAOS system from the Atlantic side of the Florida Keys, filtered through sand and particulate (20 µm membrane) filters, then treated and mixed in one of six 3,800 L header tanks. The header tanks are monitored daily for pHNBS (pH National Bureau of Standards scale) and temperature (Walchem W900, United States). The ambient pH treatment over the course of the study was 8.04 ± 0.01 (Ambient pH). Seawater for the low pH treatment was manipulated by CO2 injection through a Venturi pump and maintained at 7.83 ± 0.01 (ocean acidification [OA]) over the course of the study. The ocean OA pH is based on the value predicted to occur by the year 2100 under moderate shared socio-economic pathways for atmospheric greenhouse gas concentrations (SSP2-4.5 to SSP3-7.0; IPCC, 2021).
Raceway water baths were set to maintain a target temperature of 28°C inside 20 L aquaria using dual temperature controllers (Walchem Series W900 controllers, Iwaki America, Inc., Holliston, MA). This temperature represents average regional sea surface temperatures between May and July (NOAA National Data Buoy Center: https://www.ndbc.noaa.gov/. Last accessed July 2024). Flow rates into each aquarium were set to 300 mL min-1, allowing the water in individual tanks a residence time of one hour.
2.2 Collection and acclimation of corals and Sargassum algae
Fragments of A. cervicornis were grown at the Mote Marine Laboratory offshore nursery on “tree” style PVC rigs where individual branches are suspended by monofilament lines (Supplementary Figure 1). Fragments (n = 180) of the same genotype (genet ML-38) were collected in May 2020. Coral fragment size and form were variable, so larger fragments were cut to bring individual branch lengths to approximately 10 cm. On the same day, each coral fragment was glued (cyanoacrylate glue, Bulk Reef Supply, MN, United States) onto a ceramic puck, individually labeled, and placed in aquaria (n = 36; 5 fragments per aquarium) for acclimation to the experimental system ambient conditions for 24 hours.
After the one day acclimation period, each tank with coral fragments were randomly assigned to four treatments: 1) Amb pH, Absent, 2) Amb pH, Present, 3) OA pH, Absent, and 4) OA pH, Present (where Amb = Ambient pH [~8.02], OA = Acidified pH [~7.80], Absent = no Sargassum present, Present = Sargassum present). The aquaria order was randomized by raceway, and the order was shuffled weekly to avoid any potential light effects (Supplementary Figure 2). Prior to initial measurements (T0), coral fragments were acclimated to pH treatment conditions over 8 days where pH was decreased by 0.1 each day until the treatment pH values were met.
Every week throughout the experiment, bundles of fresh Sargassum were collected, cleaned, and added to appropriate treatment aquaria. Two species of pelagic Sargassum were identified across all collections during the study (S. natans and S. fluitans) but were not separated as it was not possible to do so with 100% accuracy and Sargassum mats found in the field consist of an assemblage of both species. Hence, we refer to them by Sargassum rather than by single species name. Sargassum were collected via dip nets from nearshore environments and boats 1-5 miles offshore on the Atlantic coast of Summerland Key, FL. They were immediately rinsed with filtered seawater and manually cleaned of epibionts using soft brushes and tweezers. Health was assessed visually (e.g., all thalli appeared intact with no damage, rot, or disease, and the color was a uniform yellow/brown) before addition to the treatment aquaria. Healthy Sargassum were bundled (100 g) into mesh bags and added to the aquaria away from coral fragments (i.e., not touching or covering coral fragments) each week (Supplementary Figures 3, 4).
2.3 Monitoring seawater conditions
A handheld multiparameter meter (YSI Professional Plus multiprobe; Texas Instruments Inc.) was used to measure seawater temperature (± 0.2°C), salinity (± 1% of the reading), and pHNBS (± 0.2) of the header tanks and experimental aquaria. The pH probe was calibrated daily using a three-point NBS buffer calibration (pH 4, 7, 10) to a minimum of 99.7% accuracy. Measurements were taken twice a day (0900 and 1500 EST) during the week. Additionally, measurements were taken at midnight three times a week to characterize daily mean ± SD and diurnal variability of each tank and aquaria. In addition to the handheld instrument, electrodes (WALCHEM pH WEL series United States) within the header tanks and subset of experimental aquaria were used to constantly monitor seawater temperature and pHNBS. A handheld light meter (LI-COR, LI-1500 DLI, United States) was used to measure PAR (µmol s-1m-2) under water in the center of each experimental aquaria daily.
Seawater samples were taken from each header tank and a random subset of experimental aquaria (n = 3 per treatment) each week for laboratory analysis of carbonate chemistry (dissolved inorganic carbon [DIC] and total seawater alkalinity [TA]). Samples were collected from aquaria in 125 mL borosilicate bottles, immediately inoculated with 60 µL of a saturated mercuric chloride (HgCl2) solution and placed in a refrigerator at 5°C until analysis (Dickson et al., 2007). Samples were analyzed for DIC and TA using a DIC analyzer (Apollo SciTech, United States) and an automatic titrator (Metrohm, Model 905 Titrando, Switzerland), respectively. Certified reference materials from the Dickson Laboratory (Scripps Institution of Oceanography) were used to ascertain measurement accuracy and precision of each instrument (Dickson et al., 2003). The accuracy and precision of DIC and TA was less than four µmol kg-1. Along with in situ temperature and salinity, seawater carbonate chemistry was constrained using CO2SYS (Version 3.2.0) (Lewis et al., 1998) with K1 and K2 constants from Lueker et al., 2000.
2.4 Coral color
Coral color was measured after the initial 9-day pH acclimation period at the beginning (T0), midpoint (T1; 1 month), and end (T2; 2 months) of the experiment. A visual health assessment was performed on each individual coral fragment three times a week using a Coral Watch Coral Health Chart (CHW Chart: Supplementary Figure 5). The CHW is a six-color shade scale representing zooxanthellae density which is proportional to the shade of the coral tissue; 1 is the palest and least healthy rating and 6 is the darkest and healthiest. Coral fragments were individually compared to the shades on the color scale to determine relative health. The average rating between all coral fragments in all treatments based on the CWH visual health rating scale of 1-6 was 5 (1 = bleached, poor health; 6 = dense zooxanthellae, healthy). Throughout the duration of the experiment, there was no visible difference in the health of any coral fragments between or within treatment groups, nor did any corals leave the 5-6 range.
2.5 Coral growth
All corals underwent a 3D scan (Metron 3D scanner, Flexscan 3D analysis software) over three time points to determine surface area (mm2). The scanning process takes 12-24 images of each subject at 15-30° rotations. Each image is then manually aligned to form the full 3D image. Scans are then “meshed” together and “smoothed” automatically. Any remaining holes in the image are manually filled, following Koch and Piadyk (2022). Images with excessively large holes (< 10% of the total area) at the initial (T0) measurements were not used.
All coral fragments were also weighed at three time points using the buoyant weight technique to measure net calcification since coral tissues are neutrally buoyant in seawater (Jokiel et al., 1978). Balances were placed over a full 10-gallon aquarium with a submerged suspended platform attached below. Seawater in the aquaria was replaced when the temperature fell below 27°C to minimize stress to coral fragments. Buoyant weight values were corrected for seawater density which was calculated from hourly measurements of seawater temperature, salinity, and coral surface area, and were calculated as the difference from T0. The coral fragments were kept in their respective treatment pH throughout the process. Buoyant weight was standardized to surface area to calculate net calcification (mg CaCO3 cm-2 day-1).
2.6 Coral physiology
The fluorescence of coral fragments was measured using a pulse amplitude modulation (PAM) fluorometer (IMAGING-PAM M-Series, MAXI version, Walz, Germany). Results were processed by ImagingWin GigE software (Version 2.41). Each A. cervicornis fragment was analyzed for fluorescence at three timepoints (T0, T1, T2) throughout the experiment. Data was recorded per manually selected “Area of Interest” (AOI) to reduce the volume of data stored. To ensure the AOIs were representative of each coral fragment, one AOI at the base of the main stem and one AOI near the tip of a younger branch were selected. Parameter values were then averaged between both AOIs per coral. Coral fragments were kept in complete darkness for 15 minutes prior to each measurement to ensure dark adaptation. For the Sargassum bundles, ten AOIs were chosen per measured set and averaged. Each coral fragment and Sargassum bundle were exposed to a photosynthetically available radiation (PAR) increment scheme ranging from 1 – 1250 PAR (µmol m2 s-1). Quantum yield of photosystem II (Fv/Fm: Y(II)), yield of non-photochemical quenching (Y(NPQ)), and maximum electron transport rate (ETRmax) were recorded and averaged between treatments over each timepoint.
2.7 Statistical analyses
A three-way repeated measure analysis of variance (ANOVA) was used to test coral responses between seawater pH and Sargassum presence across the three time points (T0, T1, T2). Data were checked for assumptions of the repeated measures ANOVA (i.e., equal variances, normality, and no outliers). When a significant three-way effect was detected, simple two-way interactions were tested at each time point. If there was a significant two-way interaction, then simple main effects were tested. Two response variables had significant two-way interactions, but no three-way interaction; posthoc analyses for these responses were simple main effect analyses and pairwise comparisons. Differences were considered significant at alpha = 0.05. All data analyses and visualizations were done using RStudio version 2023.09.1 + 494 (R Core Team, 2023). Packages used for analysis include tidyverse (Wickham et al., 2019), ggpubr (Kassambara, 2023a), and rstatix (Kassambara, 2023b).
3 Results
3.1 Coral growth rate
There was a significant three-way interaction within time and between pH and algae presence (F1,166 = 6.566, p = 0.011) (Figure 1, Table 1). After one month (T1), corals growing in ambient pH seawater had similar growth rates (± 1 SE) of 0.912 ± 0.071 mg cm-2 d-1 and 0.978 ± 0.073 mg cm-2 d-1 when algae were absent and present, respectively (i.e., the presence of algae did not impact coral growth after one month). Under OA conditions, the corals growing in the presence of Sargassum had significantly lower growth rates at T1 than corals in the absence of Sargassum (F1,80 = 6.03, p = 0.016) (Figure 1). Growth rates of corals in OA conditions and in the absence of Sargassum was 1.143 ± 0.098 mg cm-2 d-1 while corals growing in the presence of Sargassum had growth rate of 0.855 ± 0.074 mg cm-2 d-1. By the second month (T2), there were no significant effects of seawater pH and Sargassum presence on coral growth rates across all treatments (F1,171 = 0.73, p = 0.394).
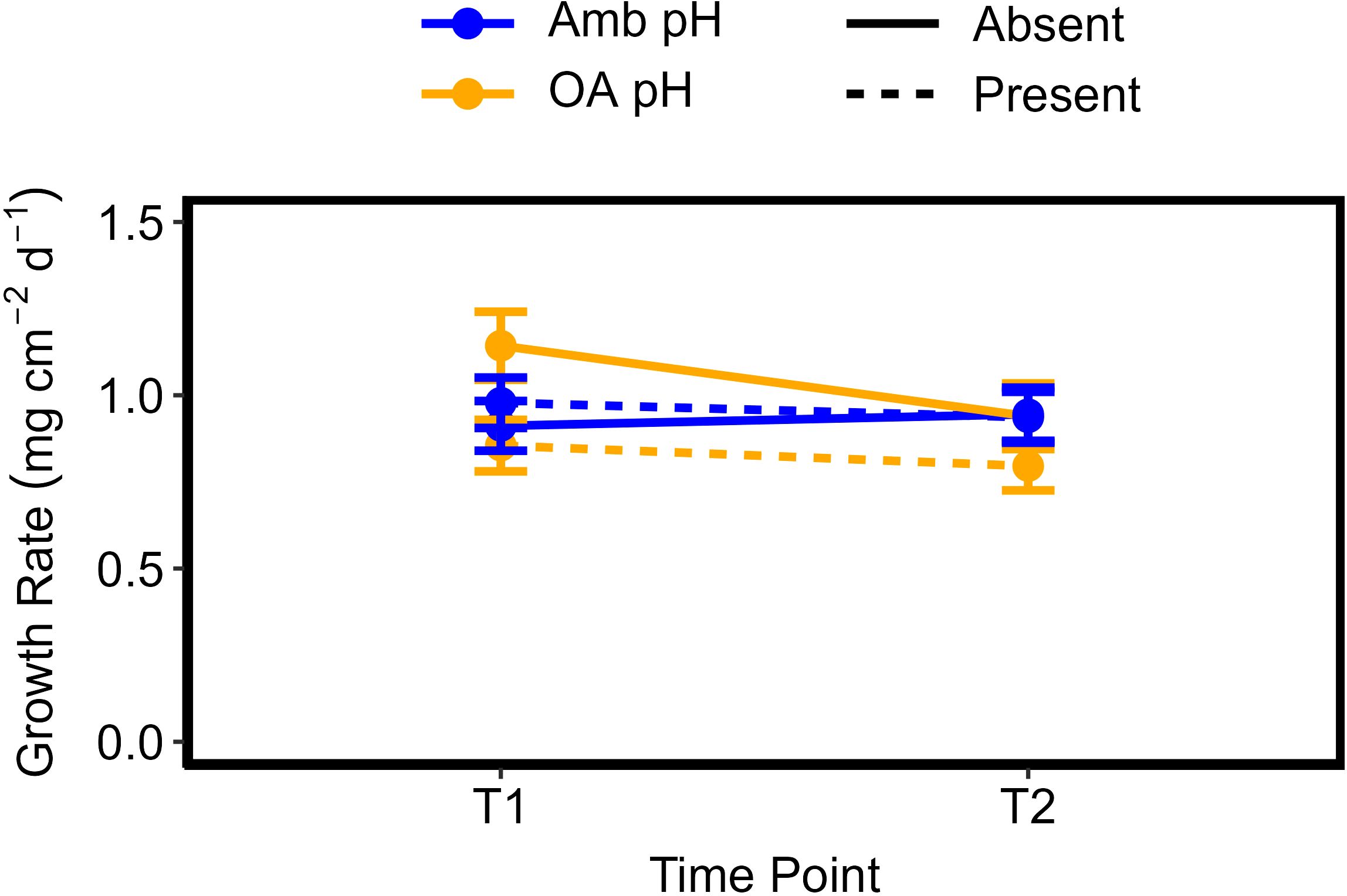
Figure 1. Coral growth rates measured as change in buoyant weight normalized to surface area (mg cm-2 d-1) at measurement time point 1 (T1: 38 days into the experiment) and at time point 2 (T2: 68 days into the experiment). Solid lines indicate Sargassum was absent, dashed lines indicate Sargassum was present. “Amb” = ambient pH, “OA” = Ocean acidification pH. Error bars represent standard error.
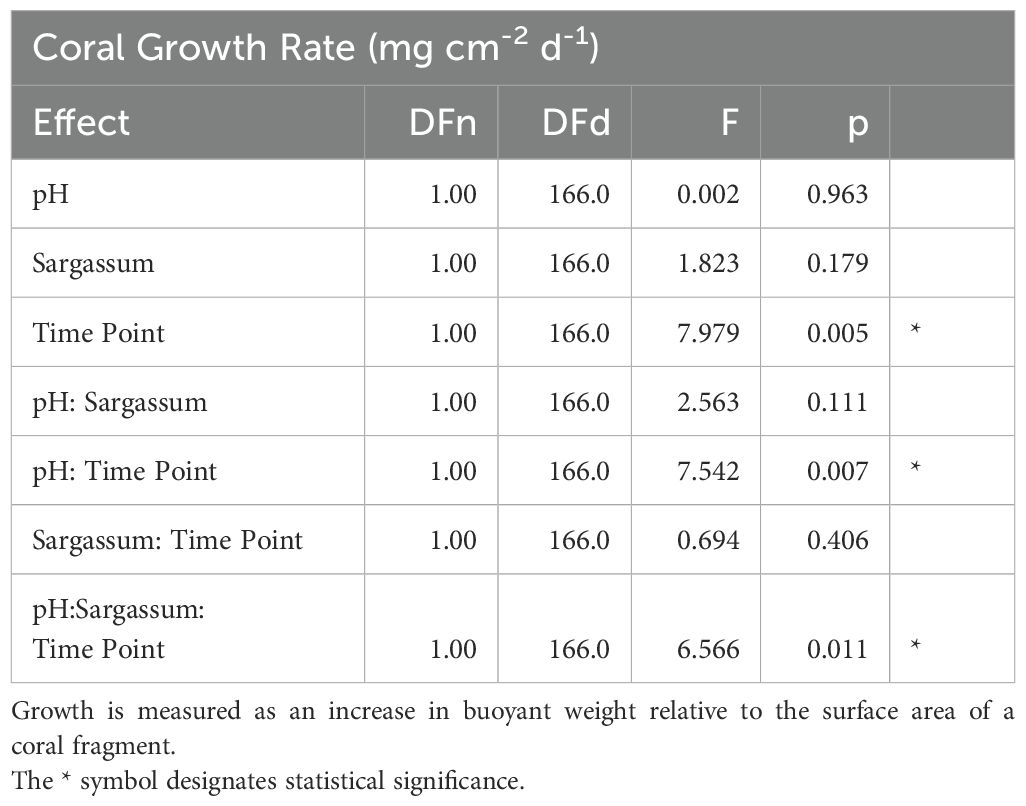
Table 1. Three-way repeated measures analysis of variance (ANOVA) for coral growth measurements.
3.2 Coral physiology
There was a significant three-way interaction within time and between pH and algae presence on quantum yield, Y(II) (F1.92,337.36 = 5.867, p < 0.001). Posthoc analyses highlighted differences between treatments varied throughout the experiment (Table 2). At the beginning of the experiment (T0), there were no significant differences between treatments. After one month (T1), quantum yield decreased in all treatments (Figure 2A, Table 2). Corals in seawater with ambient pH had significantly higher Y(II) than corals in acidified seawater (F1,176 = 45.3, p < 0.001). Within each pH level, Y(II) was significantly higher for corals in the presence of Sargassum algae (F1,176 = 14.9, p < 0.001). However, there were no interactions between pH and presence of algae on coral quantum yield (F1,176 = 1.18, p = 0.279). By the end of the experiment (T2), these differences persisted with a significant interaction between pH and algae presence (F1,176 = 6.17, p = 0.014) emerging. Corals in ambient seawater pH with Sargassum present had the highest quantum yield of 0.556 ± 0.003 while corals in acidified seawater with no algae had the lowest quantum yield of 0.427 ± 0.008.
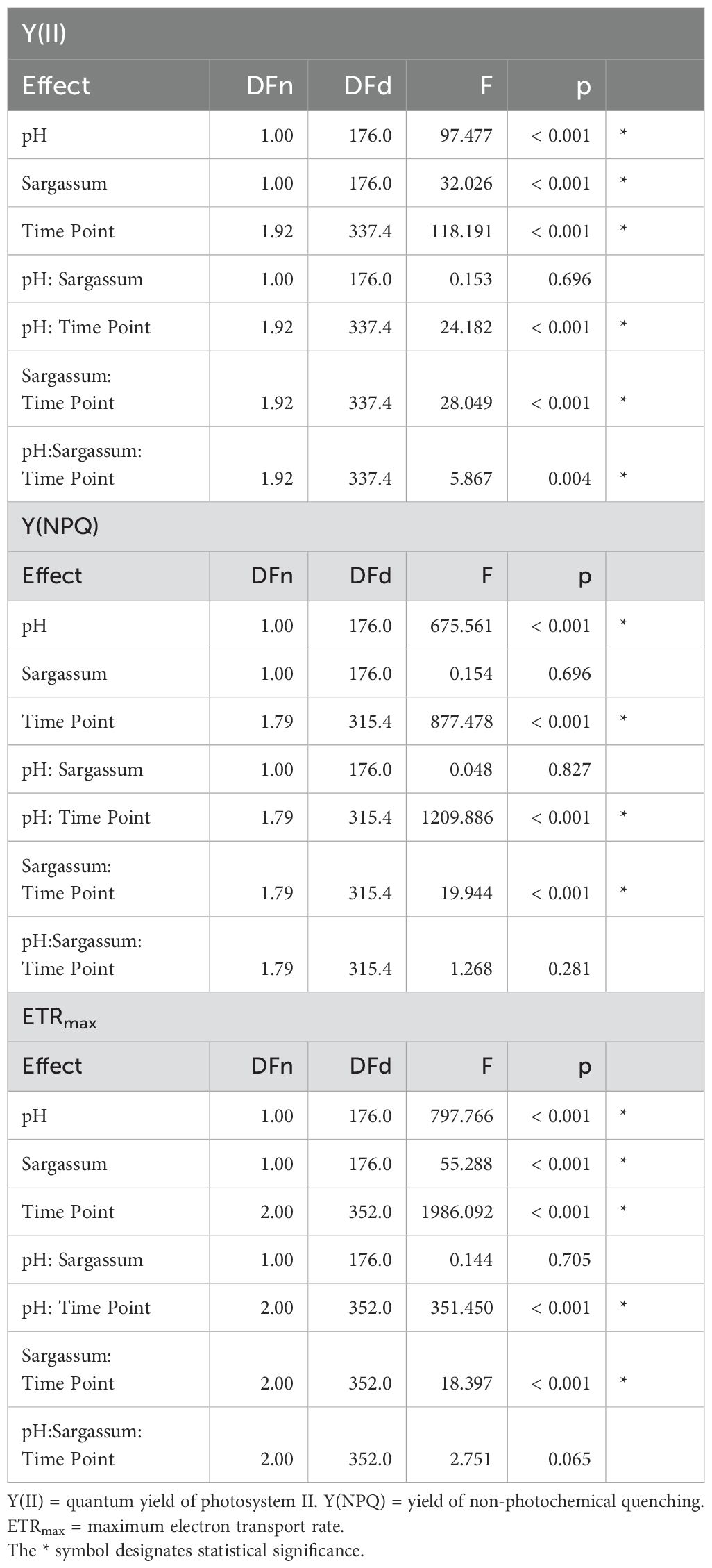
Table 2. Three-way repeated measures analysis of variance (ANOVA) for coral photophysiology results measured using pulse amplitude modulation fluorometry (PAM).
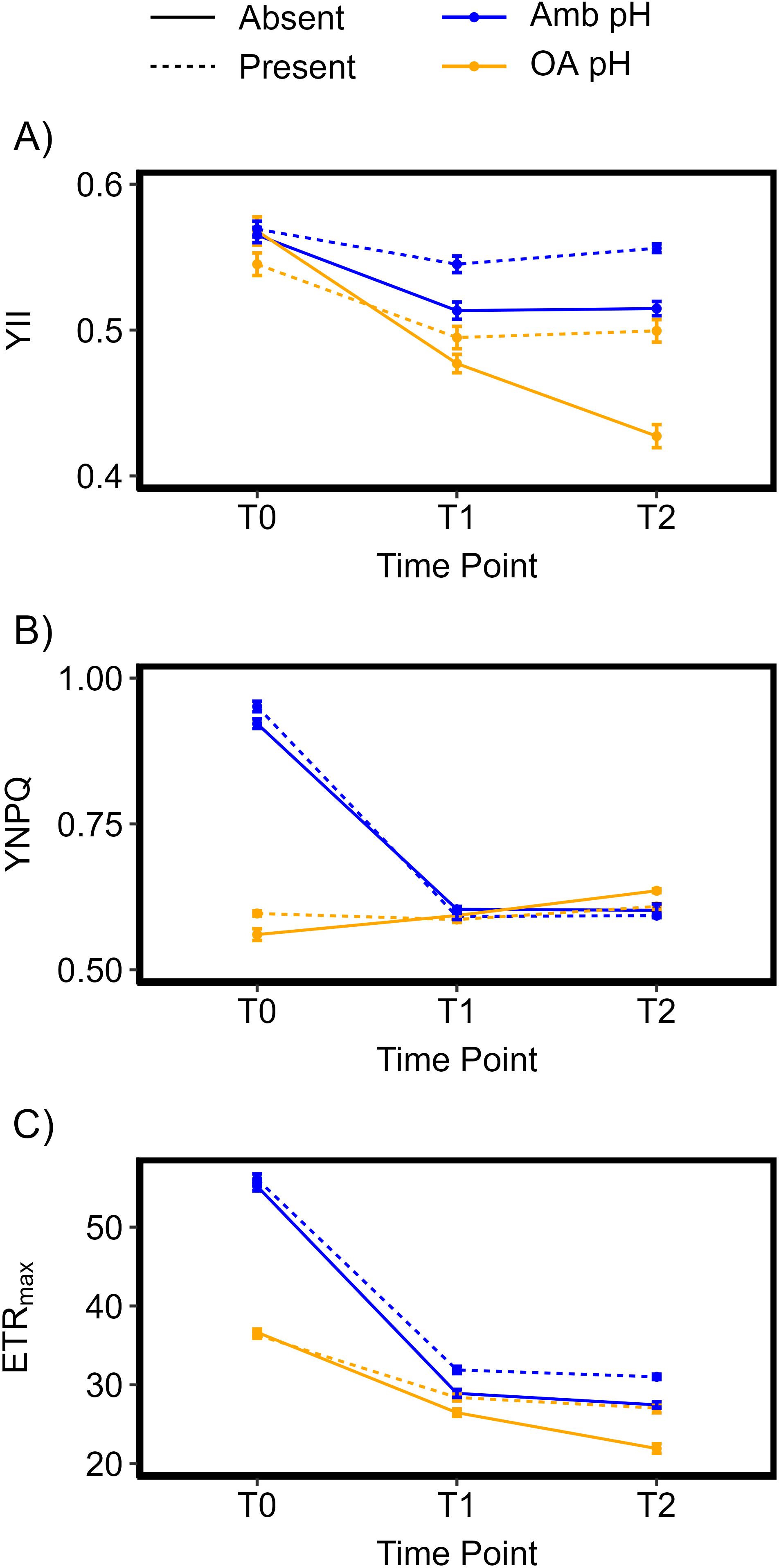
Figure 2. Chlorophyll fluorescence parameters measured through pulse amplitude modulation (PAM). (A) (top) Quantum Yield of photosystem II (Y(II))(± SE), (B) (middle) Yield of Non-Photochemical Quenching (Y(NPQ))(± SE), and (C) (bottom) the maximum Electron Transport Rate of photosynthesis (ETRmax)(± SE). Parameters measured and averaged at measurement time point 0 (T0 = day 8 post acclimation), time point 1 (T1: 38 days) and timepoint 2 (T2: 68 days). “Amb” = ambient pH, “OA” = Ocean acidification pH. Error bars represent standard error.
There was no statistically significant three-way interaction within time and between pH and algae presence on non-photochemical quenching, Y(NPQ) (F1.79, 315.43 = 1.268, p = 0.281) (Figure 2B, Table 2). However, there was a significant two-way interaction between pH and time (F1.72,306.57 = 1092.03, p < 0.001). The biggest difference in Y(NPQ) between pH treatments was at the beginning of the experiment when corals in ambient pH had higher Y(NPQ) compared to corals in acidified seawater. After one month, all corals had similar Y(NPQ) between pH treatments. After two months, corals in acidified seawater had slightly higher Y(NPQ) than corals in ambient seawater.
There was no statistically significant three-way interaction within time and between pH and algae presence on maximum electron transport rate (ETRmax) (F2,352 = 2.751, p = 0.065) (Figure 2C, Table 2). However, there were significant two-way interactions between pH and time (F2,352 = 351.45, p < 0.001) and treatment and time (F2,352 = 18.397, p < 0.001). At the beginning of the experiment (T0), corals in ambient seawater had a higher ETRmax compared to corals in acidified seawater; the ETRmax in ambient pH was 55.6 ± 0.442 while acidified pH had an ETRmax of 36.5 ± 0.296. The presence of Sargassum algae did not impact coral ETRmax. ETRmax decreased across all treatments with a bigger decline for corals in ambient seawater pH. Thus, after one month (T1), rates were similar between pH treatments. However, the effect of Sargassum presence started appearing; corals with Sargassum present had higher ETRmax compared to corals in isolation. These differences persisted to the end of the experiment. The average daily PAR level at solar noon EST across all experimental aquaria from T0-T2 was 373 ± 26.5 µmol m-2 s-1. The PAR range which corresponded to maximum electron transport rate for coral samples was 350-450 µmol m-2 s-1.
3.3 Seawater chemistry in experimental aquaria
Seawater temperature (28.2 ± 0.3°C) and salinity (35.5 ± 1.4 ppt) were similar across treatments (Table 3). Seawater carbonate chemistry for the OA treatment aquaria had higher DIC (2070 ± 47 µmol kg-1 vs. 1974 ± 50 µmol kg-1), lower total pH, (pHT) (7.88 ± 0.08 vs. 8.02 ± 0.05), and lower Ωaragonite (2.8 ± 0.5 vs. 3.6 ± 0.4). The presence of Sargassum algae did not modify seawater carbonate chemistry in DIC or pHT (Table 3) (Supplementary Figure 6).
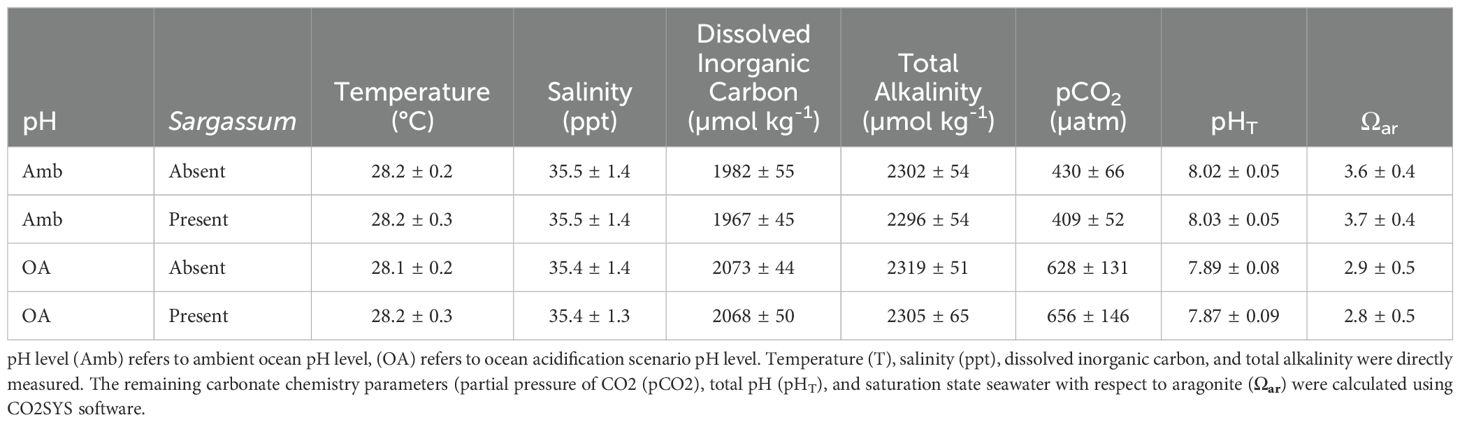
Table 3. Seawater physico-chemical conditions in experimental aquaria.
4 Discussion
Global increases in frequency and size of pelagic Sargassum blooms as well as predicted worsening of ocean acidification (OA) are expected to negatively impact coral reefs in future climate change scenarios. Phase shifts from coral towards algal dominance have already been observed on many coral reefs globally, including in Florida, since benthic macroalgae outcompete corals for space, light, CO2, and other resources through a variety of mechanisms that are often amplified by a changing climate (McCook et al., 2001; Hughes et al., 2007; Enochs et al., 2014; Hillard, 2015; Brown et al., 2018; Enochs et al., 2018; Brown et al., 2019; Biscéré et al., 2019; Muller et al., 2021). Therefore, restoration programs have focused on the addition of historically dominant reef-building species like A. cervicornis to Caribbean reefs. Previous studies have shown negative synergistic impacts of OA and coral-algal competition on coral growth and survivorship, including for acroporids (Anthony et al., 2011; Brown et al., 2019; Vieira, 2020; Rölfer et al., 2021). These studies, however, focused on benthic macroalgae. This is the first study to quantify the combined effects of pelagic Sargassum and OA on an endangered coral species A. cervicornis. Furthermore, we demonstrate that the impacts of Sargassum and OA on coral growth and photo-physiology depend on length of exposure. After 1 month, growth was negatively impacted by the combination of low seawater pH and Sargassum presence. These differences in growth, however, disappeared by the end of the experiment (Figure 1). Coral photo physiology was mostly affected by isolated stressors (low seawater pH or Sargassum presence) but there were no negative synergistic impacts of OA and Sargassum on Y(II), Y(NPQ), or ETRmax, rather, the algal presence improves photosynthetic performance instead (Figure 2, Table 2).
Corals initially showed lower growth rates in the OA + Sargassum treatment after 1 month of exposure, but this difference disappeared after two months, suggesting a degree of acclimation to their environment. This is demonstrated to occur through phenotypic or genotypic plasticity in previous studies (Sekizawa et al., 2017; Muller et al., 2021; Million et al., 2022; Dilworth et al., 2024). Earlier studies show A. cervicornis growth decreases after exposure to OA conditions for 1-2 months (Renegar and Riegl, 2005; Towle et al., 2015; Enochs et al., 2018), but more recent literature has revealed unaffected and even improved growth rates under OA (Bedwell-Ivers et al., 2017; Camp et al., 2017; Enochs et al., 2018; Langdon and Atkinson, 2005; Muller et al., 2021). Negative impacts of OA on A. cervicornis growth can also emerge or be amplified when combined with elevated nutrients (Renegar and Riegl, 2005), making it difficult to isolate the effects of the carbonate environment from other abiotic factors. However, when ideal light and temperature conditions are met, seawater carbonate chemistry does not necessarily limit growth. Enochs et al. (2014) and Comeau et al. (2014a, b) show that the growth rates of Acroporid corals including A. cervicornis are negatively impacted by OA pH levels, but that light plays a crucial modulating role in photosynthetically processing the excess CO2 in acidified seawater (Suggett et al., 2012). Growth rates can increase with CO2 concentration if met with saturating light, as seen in our initial OA controls, but can decrease together with photosynthetic efficiency if other conditions remain unchanged and there is no alternative energy source available. In this study, average peak daylight PAR and seawater temperature across all experimental aquaria throughout the study was 373 ± 26.5 µmol m-2 s-1 and 28.5 ± 0.15°C, respectively. These experimental conditions are within the optimum range for A. cervicornis physiology, resulting in a normal growth rate range of 0.09 to 1.2 mg cm-2 d-1, and expressing commonly cited Y(II) and ETRmax values across multiple studies (Table 4). Given that our PAR exposure was not saturating, it is unlikely that increased growth under OA conditions (T1) as described in Enochs’ and Comeau’s research, can be maintained in the long-term.
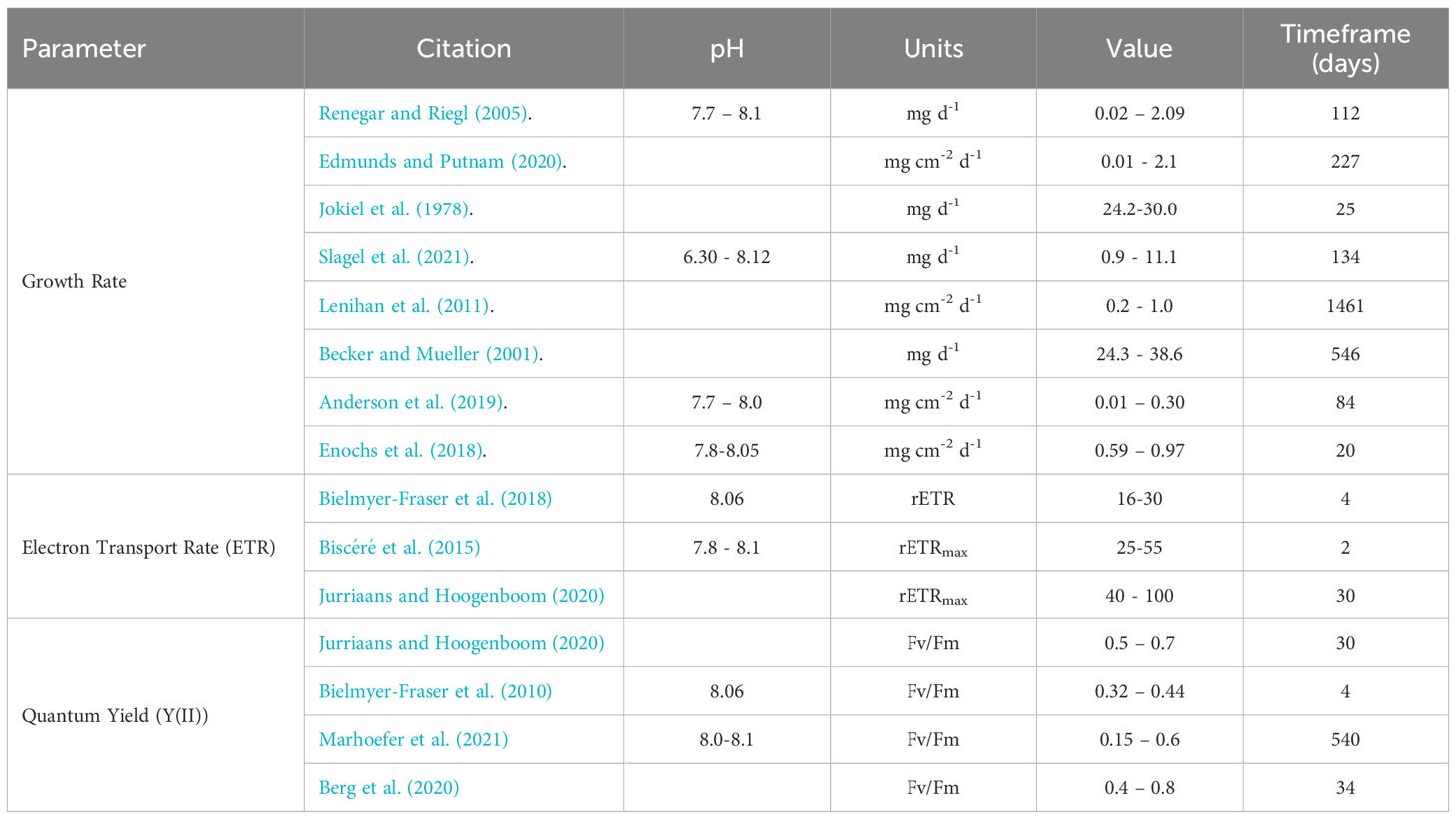
Table 4. Ranges of Acropora cervicornis growth rate and fluorometry (PAM) parameters including data from other acroporids in other literature.
After one month (T1) OA conditions negatively impacted photosynthetic performance as measured through Y(II) (Figure 2A), indicating a reduced percentage of available energy was being used for photosynthesis than in ambient treatments. However, Y(NPQ) and ETRmax were statistically similar. Therefore, energy may have instead been allocated towards maintaining stable calcification rates in the OA + Sargassum treatment. By the end of the experiment, significantly lower Y(II) as well as higher Y(NPQ) in OA controls show corals were still stressed by their environment, indicating that photo-physiology appears to be more sensitive to low seawater pH than growth rates given the differences emerge as early as T0. Research shows mixed results in terms of photosynthetic performance under predicted future OA conditions. Biscéré et al. (2019) suggests that some acroporids show increased gross photosynthesis in OA conditions without highlighting extraordinary changes to growth rate, while Brien et al. (2016) shows that corals including acroporids exposed to OA conditions can experience decreased photosynthetic yield in the short-term without sacrificing growth. Studies like Tanvet et al. (2023) and DeMerlis et al. (2022) further suggest that certain acroporid species express varying degrees of short- and long-term growth and photosynthetic adaptation to changing seawater pH based on whether they live in an environment with fluctuating or stable pH and temperature. In Tanvet et al., acroporids adapt their photosynthetic performance to pH shifts and reduce negative impacts on growth, although this adaptation takes place across longer timescales. Our coral samples came from a reef-based nursery with naturally dynamic diel and seasonal pH levels which may have stabilized their average growth rates in response to pH extremes. In contrast, previous studies show a range of results where A. cervicornis has maintained stable photosynthetic performance but showed improved growth under OA conditions (Bedwell-Ivers et al., 2017) or where carbonate concentrations rather than pH has a stronger effect on photosynthetic performance (Schneider and Erez, 2006). This wide range of results is attributed in part to genotype and multifactorial environmental differences where it is difficult to predict how a particular combination of stressors can amplify or negate each other. However, the pattern of improved photosynthetic performance when exposed to Sargassum in our results has prompted the question of whether the algae was providing a compensatory energy source.
Growth and photosynthesis have been demonstrated to be fueled by the same internal DIC pool in some models (Langdon and Atkinson, 2005) which can be supplemented by heterotrophically sourced carbon as well as photosynthesis, meaning that changes in growth do not have to come at the expense of photosynthesis, and vice versa. Additional feeding by stressed corals could buffer against potential negative growth impacts from acidification (Towle et al., 2015). Therefore, fluctuations in photosynthesis in the short-term do not immediately translate to changes in growth if there are adequate alternative nutrient sources. Sargassum algae can have beneficial or detrimental effects on coral growth and photo-physiology depending on the life stage of the algae. As initially hypothesized, other studies show that healthy benthic Sargassum and other macroalgae can buffer against OA by raising seawater pH through photosynthesis during the day (Jompa and McCook, 2003; Hillard, 2015; Page et al., 2016; Doo et al., 2020), whereas bacterial breakdown of dead or dying macroalgae could have the opposite impact on seawater carbonate chemistry by releasing CO2 and other organic material into the water. Decaying Sargassum also produces hydrogen sulfide which can reduce coral growth (Resiere et al., 2021). We excluded decaying algae from the study by replacing the Sargassum bundles weekly to reduce bacterial breakdown. We cannot guarantee full elimination of bacterial communities; however, measurements of ETRmax, Y(II) and Y(NPQ) of Sargassum thalli at the beginning and end of several 1-week exposure cycles indicated no negative change in Sargassum health (Supplementary Figures 7-9). Throughout the experiment, the Sargassum bundles shed detrital particles into the aquaria which is a typical characteristic of Sargassum blooms during reproduction or over-crowding (Ateweberhan et al., 2008). This shedding likely increased the abundance of particulate organic matter (POM) available in each experimental aquarium leading to unexpected impacts on coral photo-physiology.
Y(II) and ETRmax became higher after one and two months respectively when Sargassum was present relative to controls. Given that the presence of Sargassum did not increase pH in any treatments as we had hypothesized (Supplementary Figure 6), improved photosynthetic performance in corals exposed to algae may have been due to a shift in reliance on heterotrophic behavior rather than pH stress relief (i.e., feeding on Sargassum-derived POM). The energy allocation balance between the zooxanthellae (i.e., primary productivity) and coral polyps (i.e., heterotrophy) can shift to favor heterotrophy in response to environmental stressors such as ocean acidification (Edmunds, 2011; Agostini et al., 2021) and warming (Krueger et al., 2018). Ferrier-Pagès et al. (2003, 2021) describes how heterotrophy provides a complementary pathway for obtaining nutrients that are essential for amino acid synthesis used to maintain zooxanthellae photosynthetic apparatus as well as growth. Given that the improved photosynthetic performance does not translate to improved growth in our samples, we note that heterotrophy does not always guarantee increased growth rates in all corals, but can disproportionately supplement photosynthetic maintenance and efficiency in zooxanthellae (Dubinsky et al., 1990; Titlyanov et al., 2001a, b; Davy and Cook, 2001; Houlbrèque et al., 2003). Some studies show that heterotrophy increases amino acid synthesis used in photosynthetic apparatus repair rather than increasing the production of glycerol and glucose for growth in algae-invertebrate symbiont relationships (Swanson and Hoegh-Guldberg, 1998; Wang and Douglas, 1998). Increased photosynthetic efficiency (ETRmax) despite stable growth in Sargassum exposed corals likely resulted from a higher number of available photosynthetic units, reflected in our Y(II) results (Figure 2A), without growth-directed energy transfer, supporting the notion by Davy and Cook (2001) that nutrients gained from heterotrophy can be retained by the zooxanthellae rather than being transferred to their symbiont hosts for growth. We therefore suspect that the Sargassum treatments show better photosynthetic performance due to the availability of POM that can supplement the maintenance of photosynthetic apparatus across pH treatments but particularly in OA stressed corals which show a steep trend of decreased growth rate when no POM is available.
This study highlights more multi-stressor research is needed across time and ecological scales. Coral reefs in the Florida Keys are exposed to long-term climate change (ocean acidification) combined with faster and more frequent episodic coastal acidification (Strong et al., 2014; Hicks et al., 2022) in response to short-term regional (Sargassum blooms and storms) impacts. Multi-stressor experiments, therefore, need to reflect the varying time scales to capture the full range of effects on coral health. In isolation, our results show that A. cervicornis growth rates were not significantly affected by ocean acidification conditions within the timeframe of this experiment. Likewise, the presence of Sargassum also had no significant effect on growth. However, coral photosynthetic efficiency improved when exposed to Sargassum under current ambient pH levels as well as future ocean acidification scenarios, likely due to providing additional particulate organic matter to a heterotrophic nutrition pathway, supplementing the maintenance and performance of their symbiont zooxanthellae photosynthetic apparatus. Further research is needed to understand coral responses to stressors at a cellular level, and to scale up responses to community and ecosystem scales (Edmunds et al., 2016).
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://figshare.com/projects/Sargassum_Coral_Ocean_Acidification/217402.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
JL: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Validation, Visualization, Writing – original draft. HP: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AQ: Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. JT: Writing – review & editing, Investigation, Methodology. CL: Methodology, Writing – review & editing, Investigation. RN: Conceptualization, Data curation, Formal analysis, Methodology, Writing – review & editing. LB: Investigation, Methodology, Writing – review & editing. EH: Formal analysis, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by crowdfunding, a PADI Foundation grant (Grant #: 40433), a Protect Our Reefs License Plate Grant (POR 2019-M6), and the lead author’s personal funds.
Acknowledgments
Thank you to all staff and interns at the Elizabeth Moore International Center for Coral Reef Research and Restoration who participated in this work.
Conflict of interest
Author JL was employed by company Isometric. Author RN was employed by company Storm Anchor Insights.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1487102/full#supplementary-material
References
Agostini S., Houlbrèque F., Biscéré T., Harvey B. P., Heitzman J. M., Takimoto R. (2021). Greater mitochondrial energy production provides resistance to ocean acidification in “winning” hermatypic corals. Front. Mar. Sci. 7, 600836. doi: 10.3389/fmars.2020.600836
Alvarez-Filip L., Gill J. A., Dulvy N. K., Perry A. L., Watkinson A. R., Côté I. M. (2011). Drivers of region-wide declines in architectural complexity on Caribbean reefs. Coral reefs 30, 1051–1060. doi: 10.1007/s00338-011-0795-6
Anderson K. D., Cantin N. E., Casey J. M., Pratchett M. S. (2019). Independent effects of ocean warming versus acidification on the growth, survivorship and physiology of two Acropora corals. Coral Reefs 38, 1225–1240. doi: 10.1007/s00338-019-01864-y
Anthony K. R., Maynard J. A., Díaz-Pulido G., Mumby P. J., Marshall P. A., Cao L., et al. (2011). Ocean acidification and warming will lower coral reef resilience. Global Change Biol. 17, 1798–1808. doi: 10.1111/j.1365-2486.2010.02364.x
Antonio-Martínez F., Henaut Y., Vega-Zepeda A., Cerón-Flores A. I., Raigoza-Figueras R., Cetz-Navarro N. P. (2020). Leachate effects of pelagic Sargassum spp. on larval swimming behavior of the coral Acropora palmata. Sci. Rep. 10, 3910. doi: 10.1038/s41598-020-60739-z
Aronson R. B., Precht W. F. (2016). Physical and biological drivers of coral-reef dynamics. Coral reefs at crossroads 261–275. doi: 10.1007/978-3-319-22714-1_17
Ateweberhan M., Bruggemann J. H., Breeman A. A. M. (2008). Seasonal module dynamics in Sargassum Subrepandum (fucales, Phaeophyta) 1. J. Phycology 44, 269–283. doi: 10.1111/j.1529-8817.2008.00463.x
Baker P., Minzlaff U., Schoenle A., Schwabe E., Hohlfeld M., Jeuck A. (2018). Potential contribution of surface-dwelling Sargassum algae to deep-sea ecosystems in the southern North Atlantic. Deep Sea Res. Part II: Topical Stud. Oceanography 148, 21–34. doi: 10.1016/j.dsr2.2017.10.016
Becker L. C., Mueller E. (2001). The culture, transplantation and storage of Montastraea faveolata, Acropora cervicornis and Acropora palmata: what we have learned so far. Bull. Mar. Sci. 69, 881–896. doi: 10.5343/bms.2001.881
Bedwell-Ivers H. E., Koch M. S., Peach K. E., Joles L., Dutra E., Manfrino C. (2017). The role of in-hospite zooxanthellae photophysiology and reef chemistry on elevated pCO2 effects in two branching Caribbean corals: Acropora cervicornis and Porites divaricata. ICES J. Mar. Sci. 74, 1103–1112. doi: 10.1093/icesjms/fsw026
Berg J. T., David C. M., Gabriel M. M., Bentlage B. (2020). Fluorescence signatures of persistent photosystem damage in the staghorn coral Acropora cf. pulchra (Anthozoa: Scleractinia) during bleaching and recovery. Mar. Biol. Res. 16, 643–655. doi: 10.1080/17451000.2021.1875245
Bielmyer G. K., Grosell M., Bhagooli R., Baker A. C., Langdon C., Gillette P., et al. (2010). Differential effects of copper on three species of scleractinian corals and their algal symbionts (Symbiodinium spp.). Aquat. Toxicol. 97, 125–133. doi: 10.1016/j.aquatox.2009.12.021
Bielmyer-Fraser G. K., Patel P., Capo T., Grosell M. (2018). Physiological responses of corals to ocean acidification and copper exposure. Mar. Pollut. Bull. 133, 781–790. doi: 10.1016/j.marpolbul.2018.06.048
Biscéré T., Rodolfo-Metalpa R., Lorrain A., Chauvaud L., Thébault J., Clavier J., et al. (2015). Responses of two scleractinian corals to cobalt pollution and ocean acidification. PloS One 10, e0122898. doi: 10.1371/journal.pone.0122898
Biscéré T., Zampighi M., Lorrain A., Jurriaans S., Foggo A., Houlbrèque F., et al. (2019). High p CO2 promotes coral primary production. Biol. Lett. 15, 20180777. doi: 10.1098/rsbl.2018.0777
Bove C. B., Davies S. W., Ries J. B., Umbanhowar J., Thomasson B. C., Farquhar E. B. (2022). Global change differentially modulates Caribbean coral physiology. PLoS One 17, e0273897. doi: 10.1371/journal.pone.0273897
Brandt M. E. (2009). The effect of species and colony size on the bleaching response of reef-building corals in the Florida Keys during the 2005 mass bleaching event. Coral reefs 28, 911–924. doi: 10.1007/s00338-009-0548-y
Brien H. V., Watson S. A., Hoogenboom M. O. (2016). Presence of competitors influences photosynthesis, but not growth, of the hard coral Porites cylindrica at elevated seawater CO2. ICES J. Mar. Sci. 73, 659–669. doi: 10.1093/icesjms/fsv162
Brown K. T., Bender-Champ D., Kenyon T. M., Rémond C., Hoegh-Guldberg O., Dove S. (2019). Temporal effects of ocean warming and acidification on coral–algal competition. Coral Reefs 38, 297–309. doi: 10.1007/s00338-019-01775-y
Brown K. T., Bender-Champ D., Kubicek A., van der Zande R., Achlatis M., Hoegh-Guldberg O. (2018). The dynamics of coral-algal interactions in space and time on the southern Great Barrier Reef. Front. Mar. Sci. 5, 181. doi: 10.3389/fmars.2018.00181
Camp E. F., Nitschke M. R., Rodolfo-Metalpa R., Houlbreque F., Gardner S. G., Smith D. J., et al. (2017). Reef-building corals thrive within hot-acidified and deoxygenated waters. Sci. Rep. 7, 2434. doi: 10.1038/s41598-017-02383-y
Comeau S., Carpenter R. C., Edmunds P. J. (2014a). Effects of irradiance on the response of the coral Acropora pulchra and the calcifying alga Hydrolithon reinboldii to temperature elevation and ocean acidification. J. Exp. Mar. Biol. Ecol. 453, 28–35. doi: 10.1016/j.jembe.2013.12.013
Comeau S., Edmunds P. J., Spindel N. B., Carpenter R. C. (2014b). Fast coral reef calcifiers are more sensitive to ocean acidification in short-term laboratory incubations. Limnology Oceanography 59, 1081–1091. doi: 10.4319/lo.2014.59.3.1081
Cornwall C. E., Comeau S., Kornder N. A., Perry C. T., van Hooidonk R., DeCarlo T. M. (2021). Global declines in coral reef calcium carbonate production under ocean acidification and warming. Proc. Natl. Acad. Sci. 118, e2015265118. doi: 10.1073/pnas.2015265118
Cornwall C. E., Revill A. T., Hall-Spencer J. M., Milazzo M., Raven J. A., Hurd C. L. (2017). Inorganic carbon physiology underpins macroalgal responses to elevated CO2. Sci. Rep. 7, 46297. doi: 10.1038/srep46297
Crabbe J., Rodríguez-Martínez R., Villamizar E., Goergen L., Croquer A., Banaszak A. (2022). Acropora cervicornis. IUCN Red List Threatened Species 2022, e.T133381A165860142. doi: 10.2305/IUCN.UK.2022-2.RLTS.T133381A165860142.en
Cramer K. L., Jackson J. B., Donovan M. K., Greenstein B. J., Korpanty C. A., Cook G. M., et al. (2020). Widespread loss of Caribbean acroporid corals was underway before coral bleaching and disease outbreaks. Sci. Adv. 6, eaax9395. doi: 10.1126/sciadv.aax9395
Davy S. K., Cook C. B. (2001). The influence of ‘host release factor’ on carbon release by zooxanthellae isolated from fed and starved Aiptasia pallida (Verrill). Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 129, 487–494. doi: 10.1016/S1095-6433(01)00428-2
del Monaco C., Hay M. E., Gartrell P., Mumby P. J., Diaz-Pulido G. (2017). Effects of ocean acidification on the potency of macroalgal allelopathy to a common coral. Sci. Rep. 7, 41053. doi: 10.1038/srep41053
DeMerlis A., Kirkland A., Kaufman M. L., Mayfield A. B., Formel N., Kolodziej G. (2022). Pre-exposure to a variable temperature treatment improves the response of Acropora cervicornis to acute thermal stress. Coral Reefs 41, 435–445. doi: 10.1007/s00338-022-02232-z
Devault D. A., Massat F., Lambourdière J., Maridakis C., Dupuy L., Péné-Annette A., et al. (2022). Micropollutant content of Sargassum drifted ashore: arsenic and chlordecone threat assessment and management recommendations for the Caribbean. Environ. Sci. pollut. Res. 29, 66315–66334. doi: 10.1007/s11356-022-20300-3
Diaz-Pulido G., Gouezo M., Tilbrook B., Dove S., Anthony K. R. (2011). High CO2 enhances the competitive strength of seaweeds over corals. Ecol. Lett. 14, 156–162. doi: 10.1111/j.1461-0248.2010.01521.x
Dickson A., Afghan J. D., Anderson G. C. (2003). Reference materials for oceanic CO2 analysis: A method for the certification of total alkalinity. Mar. Chem. 80, 185–197. doi: 10.1016/S0304-4203(02)00133-0
Dickson A. G., Sabine C. L., Christian J. R. (Eds.) (2007). Guide to Best Practices for Ocean CO2 Measurements Vol. 3 (PICES Special Publication), 191 pp.
Dilworth J., Million W. C., Ruggeri M., Hall E. R., Dungan A. M., Muller E. M., et al. (2024). Synergistic response to climate stressors in coral is associated with genotypic variation in baseline expression. Proc. R. Soc. B 291, 20232447. doi: 10.1098/rspb.2023.2447
Doo S. S., Leplastrier A., Graba-Landry A., Harianto J., Coleman R. A., Byrne M. (2020). Amelioration of ocean acidification and warming effects through physiological buffering of a macroalgae. Ecol. Evol. 10, 8465–8475. doi: 10.1002/ece3.v10.15
Dubinsky Z., Stambler N., Ben-Zion M., McCloskey L. R., Muscatine L., Falkowski P. G. (1990). The effect of external nutrient resources on the optical properties and photosynthetic efficiency of Stylophora pistillata. Proc. R. Soc. London. B. Biol. Sci. 239, 231–246. doi: 10.1098/rspb.1990.0015
Edmunds P. J. (2011). Zooplanktivory ameliorates the effects of ocean acidification on the reef coral Porites spp. Limnology oceanography 56, 2402–2410. doi: 10.4319/lo.2011.56.6.2402
Edmunds P. J., Comeau S., Lantz C., Andersson A., Briggs C., Cohen A., et al. (2016). Integrating the effects of ocean acidification across functional scales on tropical coral reefs. Bioscience 66, 350–362. doi: 10.1093/biosci/biw023
Edmunds P. J., Putnam H. M. (2020). Science-based approach to using growth rate to assess coral performance and restoration outcomes. Biol. Lett. 16, 20200227. doi: 10.1098/rsbl.2020.0227
Enochs I. C., Manzello D. P., Carlton R., Schopmeyer S., Van Hooidonk R., Lirman D. (2014). Effects of light and elevated pCO2 on the growth and photochemical efficiency of Acropora cervicornis. Coral Reefs 33, 477–485. doi: 10.1007/s00338-014-1132-7
Enochs I. C., Manzello D. P., Jones P. J., Aguilar C., Cohen K., Valentino L. (2018). The influence of diel carbonate chemistry fluctuations on the calcification rate of Acropora cervicornis under present day and future acidification conditions. J. Exp. Mar. Biol. Ecol. 506, 135–143. doi: 10.1016/j.jembe.2018.06.007
Erez J., Reynaud S., Silverman J., Schneider K., Allemand D. (2011). Coral calcification under ocean acidification and global change. In: Gattuso P. L. F. L. J. M., R. H. (eds.) Coral reefs: an ecosystem in transition. Dordrecht: Springer. p. 151–176.
Farfan G. A., Apprill A., Cohen A., DeCarlo T. M., Post J. E., Waller R. G., et al. (2022). Crystallographic and chemical signatures in coral skeletal aragonite. Coral Reefs 41, 19–34. doi: 10.1007/s00338-021-02198-4
Ferrier-Pagès C., Martinez S., Grover R., Cybulski J., Shemesh E., Tchernov D. (2021). Tracing the trophic plasticity of the coral–dinoflagellate symbiosis using amino acid compound-specific stable isotope analysis. Microorganisms 9, 182. doi: 10.3390/microorganisms9010182
Ferrier-Pagès C., Witting J., Tambutté E., Sebens K. P. (2003). Effect of natural zooplankton feeding on the tissue and skeletal growth of the scleractinian coral Stylophora pistillata. Coral Reefs 22, 229–240. doi: 10.1007/s00338-003-0312-7
Gao K., Beardall J., Häder D. P., Hall-Spencer J. M., Gao G., Hutchins D. A. (2019). Effects of ocean acidification on marine photosynthetic organisms under the concurrent influences of warming, UV radiation, and deoxygenation. Front. Mar. Sci. 6, 322. doi: 10.3389/fmars.2019.00322
Goodwin D. S., Siuda A. N., Schell J. M. (2022). In situ observation of holopelagic Sargassum distribution and aggregation state across the entire North Atlantic from 2011 to 2020. PeerJ 10, e14079. doi: 10.7717/peerj.14079
Gower J. F. R., King S. A. (2011). Distribution of floating Sargassum in the Gulf of Mexico and the Atlantic Ocean mapped using MERIS. Int. J. Remote Sens. 32, 1917–1929. doi: 10.1080/01431161003639660
Hicks T. L., Shamberger K. E. F., Fitzsimmons J. N., et al. (2022). Tropical cyclone-induced coastal acidification in Galveston Bay, Texas. Commun. Earth Environ. 3, 297. doi: 10.1038/s43247-022-00608-1
Hillard H. (2015). The role of Sargassum pacificum in ocean acidification effects on corals (Northridge: Doctoral dissertation, California State University).
Houlbrèque F., Tambutté E., Ferrier-Pagès C. (2003). Effect of zooplankton availability on the rates of photosynthesis, and tissue and skeletal growth in the scleractinian coral Stylophora pistillata. J. Exp. Mar. Biol. Ecol. 296, 145–166. doi: 10.1016/S0022-0981(03)00259-4
Hughes T. P., Anderson K. D., Connolly S. R., Heron S. F., Kerry J. T., Lough J. M. (2018). Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 359, 80–83. doi: 10.1126/science.aan8048
Hughes T. P., Rodrigues M. J., Bellwood D. R., Ceccarelli D., Hoegh-Guldberg O., McCook L., et al. (2007). Phase shifts, herbivory, and the resilience of coral reefs to climate change. Curr. Biol. 17, 360–365. doi: 10.1016/j.cub.2006.12.049
IPCC (2021). Climate change 2021: The physical science basis. contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change. Eds. Masson-Delmotte V., Zhai P., Pirani A., Connors S. L., Péan C., Berger S., et al (Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press). doi: 10.1017/9781009157896
Johnson D., Ko D. S., Franks J. S., Moreno P. A., Sanchez-Rubio G. (2013). The Sargassum invasion of the Eastern Caribbean and dynamics of the Equatorial North Atlantic. Available online at: https://aquadocs.org/handle/1834/36258 (Accessed February 14, 2025).
Jokiel P., Maragos J., Franzisket L. (1978). Coral growth: buoyant weight technique. Coral reefs: research methods. Eds. Stoddart D., Johannes R. E.. Paris: UNESCO, 529–542.
Jompa J., McCook L. J. (2003). Coral-algal competition: macroalgae with different properties have different effects on corals. Mar. Ecol. Prog. Ser. 258, 87–95. doi: 10.3354/meps258087
Jurriaans S., Hoogenboom M. O. (2020). Seasonal acclimation of thermal performance in two species of reef-building corals. Mar. Ecol. Prog. Ser. 635, 55–70. doi: 10.3354/meps13203
Kassambara A. (2023a). ggpubr: “ggplot2” Based Publication Ready Plots (0.6.0). [R package]. Available online at: https://rpkgs.datanovia.com/ggpubr/ (Accessed February 14, 2025).
Kassambara A. (2023b). rstatix: Pipe-Friendly Framework for Basic Statistical Tests (0.7.2). [R package]. Available online at: https://rpkgs.datanovia.com/rstatix/ (Accessed February 14, 2025).
Koch S., Piadyk Y. (2022). “Hardware design and accurate simulation for benchmarking of 3D reconstruction algorithms,” Proceedings of the Neural Information Processing Systems (NeurIPS) Track on Datasets and Benchmarks, 1. NeurIPS 2021, New Orleans, Louisiana, USA (held virtually). Available online at: https://datasets-benchmarks-proceedings.neurips.cc/paper/2021/file/d09bf41544a3365a46c9077ebb5e35c3-Paper-round2.pdf (Accessed June 18, 2023).
Krueger T., Bodin J., Horwitz N., Loussert-Fonta C., Sakr A., Escrig S. (2018). Temperature and feeding induce tissue level changes in autotrophic and heterotrophic nutrient allocation in the coral symbiosis–A NanoSIMS study. Sci. Rep. 8, 12710. doi: 10.1038/s41598-018-31094-1
Kumar A., AbdElgawad H., Castellano I., Lorenti M., Delledonne M., Beemster G. T., et al. (2017). Physiological and biochemical analyses shed light on the response of Sargassum vulgare to ocean acidification at different time scales. Front. Plant Sci. 8, 570. doi: 10.3389/fpls.2017.00570
Langdon C., Atkinson M. J. (2005). Effect of elevated pCO2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. J. Geophysical Research: Oceans 110, 1–16. doi: 10.1029/2004JC002576
Lenihan H. S., Holbrook S. J., Schmitt R. J., Brooks A. J. (2011). Influence of corallivory, competition, and habitat structure on coral community shifts. Ecology 92, 1959–1971. doi: 10.1890/11-0108.1
Lewis E., Wallace D., Allison L. J. (1998). Program developed for CO2 system calculations (No. ORNL/CDIAC-105) (Upton, NY: Brookhaven National Lab.).
Louime C., Fortune J., Gervais G. (2017). Sargassum invasion of coastal environments: a growing concern. Am. J. Environ. Sci. 13, 58–64. doi: 10.3844/ajessp.2017.58.64
Lueker T. J., Dickson A. G., Keeling C. D. (2000). Ocean pCO2 calculated from dissolved inorganic carbon, alkalinity, and equations for K1 and K2: validation based on laboratory measurements of CO2 in gas and seawater at equilibrium. Mar. Chem. 70, 105–119. doi: 10.1016/S0304-4203(00)00022-0
Marhoefer S. R., Zenger K. R., Strugnell J. M., Logan M., van Oppen M. J., Kenkel C. D., et al. (2021). Signatures of adaptation and acclimatization to reef flat and slope habitats in the coral Pocillopora damicornis. Front. Mar. Sci. 8, 704709. doi: 10.3389/fmars.2021.704709
McCook L., Jompa J., Diaz-Pulido G. (2001). Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral reefs 19, 400–417. doi: 10.1007/s003380000129
McManus J. W., Polsenberg J. F. (2004). Coral–algal phase shifts on coral reefs: ecological and environmental aspects. Prog. Oceanography 60, 263–279. doi: 10.1016/j.pocean.2004.02.014
Miller S., Chiappone M. (2013). Elkhorn and Staghorn Coral Surveys - Florida Keys National Marine Sanctuary. Key West, Florida, USA: Florida Keys National Marine Sanctuary. Available online at: https://nmsfloridakeys.blob.core.windows.net/floridakeys-prod/media/archive/scisummaries/elkhornstaghorn2013.pdf (Accessed August 01, 2024).
Miller A. W., Richardson L. L. (2015). Emerging coral diseases: a temperature-driven process? Mar. Ecol. 36, 278–291. doi: 10.1111/maec.2015.36.issue-3
Million W. C., Ruggeri M., O’Donnell S., Bartels E., Conn T., Krediet C. J., et al. (2022). Evidence for adaptive morphological plasticity in the Caribbean coral, Acropora cervicornis. Proc. Natl. Acad. Sci. 119, e2203925119. doi: 10.1073/pnas.2203925119
Muller E. M., Dungan A. M., Million W. C., Eaton K. R., Petrik C., Bartels E., et al. (2021). Heritable variation and lack of tradeoffs suggest adaptive capacity in Acropora cervicornis despite negative synergism under climate change scenarios. Proc. R. Soc. B 288, 20210923. doi: 10.1098/rspb.2021.0923
Muller E. M., van Woesik R. (2009). Shading reduces coral-disease progression. Coral reefs 28, 757–760. doi: 10.1007/s00338-009-0504-x
Muñiz-Castillo A. I., Rivera-Sosa A., Chollett I., Eakin C. M., Andrade-Gómez L., McField M. (2019). Three decades of heat stress exposure in Caribbean coral reefs: a new regional delineation to enhance conservation. Sci. Rep. 9, 11013. doi: 10.1038/s41598-019-47307-0
Orr J. C., Fabry V. J., Aumont O., Bopp L., Doney S. C., Feely R. A. (2005). Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437, 681–686. doi: 10.1038/nature04095
Page H. N., Andersson A. J., Jokiel P. L., Rodgers K. U. S., Lebrato M., Yeakel K. (2016). Differential modification of seawater carbonate chemistry by major coral reef benthic communities. Coral Reefs 35, 1311–1325. doi: 10.1007/s00338-016-1490-4
Page H. N., Hewett C., Tompkins H., Hall E. R. (2021). Ocean acidification and direct interactions affect coral, macroalga, and sponge growth in the florida keys. J. Mar. Sci. Eng. 9, 739. doi: 10.3390/jmse9070739
Perry C. T., Murphy G. N., Kench P. S., Smithers S. G., Edinger E. N., Steneck R. S., et al. (2013). Caribbean-wide decline in carbonate production threatens coral reef growth. Nat. Commun. 4, 1402. doi: 10.1038/ncomms2409
R Core Team (2023). R: A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed October 9, 2024).
Renegar D. A., Riegl B. M. (2005). Effect of nutrient enrichment and elevated CO2 partial pressure on growth rate of Atlantic scleractinian coral Acropora cervicornis. Mar. Ecol. Prog. Ser. 293, 69–76. doi: 10.3354/meps293069
Resiere D., Mehdaoui H., Florentin J., Gueye P., Lebrun T., Blateau A. (2021). Sargassum seaweed health menace in the Caribbean: Clinical characteristics of a population exposed to hydrogen sulfide during the 2018 massive stranding. Clin. Toxicol. 59, 215–223. doi: 10.1080/15563650.2020.1789162
Rodríguez-Martínez R. E., Medina-Valmaseda A. E., Blanchon P., Monroy-Velázquez L. V., Almazán-Becerril A., Delgado-Pech B., et al. (2019). Faunal mortality associated with massive beaching and decomposition of pelagic sargassum. Mar. Pollut. Bull. 146, 201–205.
Rölfer L., Reuter H., Ferse S. C., Kubicek A., Dove S., Hoegh-Guldberg O., et al. (2021). Coral-macroalgal competition under ocean warming and acidification. J. Exp. Mar. Biol. Ecol. 534, 151477. doi: 10.1016/j.jembe.2020.151477
Schneider K., Erez J. (2006). The effect of carbonate chemistry on calcification and photosynthesis in the hermatypic coral Acropora eurystoma. Limnology Oceanography 51, 1284–1293. doi: 10.4319/lo.2006.51.3.1284
Schopmeyer S. A., Lirman D., Bartels E., Gilliam D. S., Goergen E. A., Griffin S. P. (2017). Regional restoration benchmarks for Acropora cervicornis. Coral reefs 36, 1047–1057. doi: 10.1007/s00338-017-1596-3
Sekizawa A., Uechi H., Iguchi A., Nakamura T., Kumagai N. H., Suzuki A. (2017). Intraspecific variations in responses to ocean acidification in two branching coral species. Mar. pollut. Bull. 122, 282–287. doi: 10.1016/j.marpolbul.2017.06.061
Sissini M. N., de Barros Barreto M. B. B., Széchy M. T. M., De Lucena M. B., Oliveira M. C., Gower J., et al. (2017). The floating Sargassum (Phaeophyceae) of the South Atlantic Ocean–likely scenarios. Phycologia 56, 321–328. doi: 10.2216/16-29.1
Slagel S., Lohr K., O’Neil K., Patterson J. (2021). Growth, calcification, and photobiology of the threatened coral Acropora cervicornis in natural versus artificial light. Zoo Biol. 40, 201–207. doi: 10.1002/zoo.21589
Strong A. L., Kroeker K. J., Teneva L. T., Mease L. A., Kelly R. P. (2014). Ocean acidification 2.0: managing our changing coastal ocean chemistry. BioScience 64, 581–592. doi: 10.1093/biosci/biu072
Suggett D. J., Dong L. F., Lawson T., Lawrenz E., Torres L., Smith D. J. (2012). Light availability determines susceptibility of reef building corals to ocean acidification. Coral reefs 32, 327–337. doi: 10.1007/s00338-012-0996-7
Sully S., Burkepile D. E., Donovan M. K., Hodgson G., Van Woesik R. (2019). A global analysis of coral bleaching over the past two decades. Nat. Commun. 10, 1–5. doi: 10.1038/s41467-019-09238-2
Swanson R., Hoegh-Guldberg O. (1998). Amino acid synthesis in the symbiotic sea anemone Aiptasia pulchella. Mar. Biol. 131, 83–93. doi: 10.1007/s002270050299
Tanvet C., Camp E. F., Sutton J., Houlbrèque F., Thouzeau G., Rodolfo-Metalpa R. (2023). Corals adapted to extreme and fluctuating seawater pH increase calcification rates and have unique symbiont communities. Ecol. Evol. 13, e10099. doi: 10.1002/ece3.10099
Titlyanov E. A., Titlyanova T. V., Yamazato K., van Woesik R. (2001a). Photo-acclimation dynamics of the coral Stylophora pistillata to low and extremely low light. J. Exp. Mar. Biol. Ecol. 263, 211–225. doi: 10.1016/S0022-0981(01)00309-4
Titlyanov E. A., Titlyanova T. V., Yamazato K., Van Woesik R. (2001b). Photo-acclimation of the hermatypic coral Stylophora pistillata while subjected to either starvation or food provisioning. J. Exp. Mar. Biol. Ecol. 257, 163–181. doi: 10.1016/S0022-0981(00)00308-7
Toth L. T., Aronson R. B., Cobb K. M., Cheng H., Edwards R. L., Grothe P. R., et al. (2015). Climatic and biotic thresholds of coral-reef shutdown. Nat. Climate Change 5, 369–374. doi: 10.1038/nclimate2541
Towle E. K., Enochs I. C., Langdon C. (2015). Threatened Caribbean coral is able to mitigate the adverse effects of ocean acidification on calcification by increasing feeding rate. PLoS One 10, e0123394. doi: 10.1371/journal.pone.0123394
van Tussenbroek B. I., Arana H. A. H., Rodríguez-Martínez R. E., Espinoza-Avalos J., Canizales-Flores H. M., González-Godoy C. E. (2017). Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. pollut. Bull. 122, 272–281. doi: 10.1016/j.marpolbul.2017.06.057
van Woesik R., McCaffrey K. R. (2017). Repeated thermal stress, shading, and directional selection in the Florida reef tract. Front. Mar. Sci. 4, 182. doi: 10.3389/fmars.2017.00182
van Woesik R., Randall C. J. (2017). Coral disease hotspots in the Caribbean. Ecosphere 8, e01814. doi: 10.1002/ecs2.2017.8.issue-5
Vieira C. (2020). Lobophora–coral interactions and phase shifts: summary of current knowledge and future directions. Aquat. Ecol. 54, 1–20. doi: 10.1007/s10452-019-09723-2
Virgen-Urcelay A., Donner S. D. (2023). Increase in the extent of mass coral bleaching over the past half-century, based on an updated global database. PLoS One 18, e0281719. doi: 10.1371/journal.pone.0281719
Walton C. J., Hayes N. K., Gilliam D. S. (2018). Impacts of a regional, multi-year, multi-species coral disease outbreak in Southeast Florida. Front. Mar. Sci. 5, 323. doi: 10.3389/fmars.2018.00323
Wang J. T., Douglas A. E. (1998). Nitrogen recycling or nitrogen conservation in an alga–invertebrate symbiosis? J. Exp. Biol. 201, 2445–2453. doi: 10.1242/jeb.201.16.2445
Wang M., Hu C., Barnes B. B., Mitchum G., Lapointe B., Montoya J. P. (2019). The great Atlantic sargassum belt. Science 365, 83–87. doi: 10.1126/science.aaw7912
Wells R. J., Rooker J. R. (2004). Spatial and temporal patterns of habitat use by fishes associated with Sargassum mats in the northwestern Gulf of Mexico. Bull. Mar. Sci. 74, 81–99. doi: 10.5343/bms.2004.81
Wickham H., Averick M., Bryan J., Chang W., D’Agostino McGowan L., François R., et al. (2019). Welcome to the {tidyverse}. J. Open Source Software 4, 1686. doi: 10.21105/joss.01686
Xiao J., Wang Z., Liu D., Fu M., Yuan C., Yan T. (2021). Harmful macroalgal blooms (HMBs) in China’s coastal water: Green and golden tides. Harmful Algae 107, 102061. doi: 10.1016/j.hal.2021.102061
Xu Z., Gao G., Xu J., Wu H. (2017). Physiological response of a golden tide alga (Sargassum muticum) to the interaction of ocean acidification and phosphorus enrichment. Biogeosciences 14, 671–681. doi: 10.5194/bg-14-671-2017
Young C. N., Schopmeyer S. A., Lirman D. (2012). A review of reef restoration and coral propagation using the threatened genus Acropora in the Caribbean and Western Atlantic. Bull. Mar. Sci. 88, 1075–1098. doi: 10.5343/bms.2011.1143
Keywords: Sargassum, Acropora cervicornis, ocean acidification, algal bloom, future oceans
Citation: Lankes JD, Page HN, Quasunella A, Torkelson JF, Lemaire C, Nowicki RJ, Blasius L and Hall ER (2025) Quantifying coral-algal interactions in an acidified ocean: Sargassum spp. exposure mitigates low pH effects on Acropora cervicornis health. Front. Mar. Sci. 12:1487102. doi: 10.3389/fmars.2025.1487102
Received: 27 August 2024; Accepted: 03 February 2025;
Published: 28 February 2025.
Edited by:
Guang Gao, Xiamen University, ChinaReviewed by:
Jinlin Liu, Tongji University, ChinaMenglin Bao, Incheon National University, Republic of Korea
Copyright © 2025 Lankes, Page, Quasunella, Torkelson, Lemaire, Nowicki, Blasius and Hall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johann D. Lankes, Si5kYXZpZC5sYW5rZXNAZ21haWwuY29t
†These authors have contributed equally to this work