 Kevin P. Barry1†
Kevin P. Barry1† Keith D. Mullin1*
Keith D. Mullin1* Katherine Maze-Foley2,3
Katherine Maze-Foley2,3 Lynsey A. Wilcox Talbot4
Lynsey A. Wilcox Talbot4 Patricia E. Rosel4
Patricia E. Rosel4 Melissa S. Soldevilla2
Melissa S. Soldevilla2 Laura Aichinger Dias2,3
Laura Aichinger Dias2,3 M. Rafael Ramírez-León5
M. Rafael Ramírez-León5 Jenny A. Litz2
Jenny A. Litz2- 1Marine Mammal and Turtle Division, Southeast Fisheries Science Center, NMFS/NOAA, Pascagoula, MS, United States
- 2Marine Mammal and Turtle Division, Southeast Fisheries Science Center, NMFS/NOAA, Miami, FL, United States
- 3Cooperative Institute for Marine and Atmospheric Studies, University of Miami, Miami, FL, United States
- 4Marine Mammal and Turtle Division, Southeast Fisheries Science Center, NMFS/NOAA, Lafayette, LA, United States
- 5Departamento de Oceanografía Biológica, Centro de Investigación Científicas y de Educación, Superior de Ensenada, Baja California, Mexico
Killer whales occur in the Gulf of Mexico (GoMex) and the North Atlantic, including off the southeastern United States (SEUS). Data from cetacean surveys during 1990 – 2021 and other sources were combined to assess killer whale biology, including spatial and temporal distribution, social structure, genetics, morphology, acoustics, and predatory behavior. GoMex records occurred predominantly in oceanic waters (>200 m) during spring and summer. SEUS records occurred primarily in winter and spring off the North Carolina region along the shelf-edge and deeper waters, and off the east coast of Florida. Photo-identification analysis of GoMex killer whales resulted in 49 individuals sighted up to seven times with sighting histories up to 26 years, and social analysis provided evidence of long-term relationships up to 16 years. The GoMex genetic samples revealed two mtDNA haplotypes, one of which does not match any outside the GoMex. Most GoMex whales had wide non-faint saddle patches and many had cookiecutter shark scars while no scars were noted on SEUS whales. Three groups recorded in the GoMex made few calls, but a group harassing sperm whales produced many. Cetaceans and tuna are known prey in the GoMex and SEUS, respectively. Directed studies of killer whales in the GoMex areas would be difficult to implement as this species is very rare. It is therefore important to pursue ongoing efforts to collect behavioral, acoustic and any biological samples that will contribute to improve our understanding of the biology and ecology of killer whales in tropical and subtropical regions.
1 Introduction
Killer whales (Orcinus orca) are distributed globally with higher densities in temperate and polar waters than tropical regions (Forney and Wade, 2006). Long-term ecological research on killer whales conducted in high latitude habitats of the North Pacific, North Atlantic, and Antarctic describe distinctive external morphology (morphotype), social structure, taxonomy and speciation, population structure, dietary preferences (e.g., marine mammals, fish, sharks, penguins), and distribution and movement patterns (e.g., Bigg et al., 1990; Ford et al., 1998; Baird, 2000; Pitman and Ensor, 2003; Foote et al., 2009; Moura et al., 2014; Morin et al., 2015; Jourdain et al., 2019) among multiple ecotypes or morphotypes (De Bruyn et al., 2013; Jefferson et al., 2015; Ford, 2019). The most extensively studied killer whales are the three well-established ecotypes from the temperate eastern North Pacific (resident, Bigg’s/transient, offshore) (Jones, 2006; Krahn et al., 2007; Dahlheim et al., 2008; Ford, 2009; Ford et al., 2011). Five ecotypes or morphotypes have been described in the Antarctic-Subantarctic (Type A, Type B-large, Type B-small, Type-C, Type-D) (Pitman and Ensor, 2003; Pitman et al., 2007; Andrews et al., 2008; Pitman, 2011; Pitman et al., 2011). Two proposed ecotypes were described in the high latitude North Atlantic (Type 1, Type 2) (Foote et al., 2009) but require further study (Foote, 2022).
Compared to these high latitude populations, information on killer whales in low latitude warm waters (“tropical”) is sparse because they are more dispersed and, in many cases, less accessible in more offshore waters. Our current view of killer whale ecology is largely based on high latitude populations and our understanding of killer whale populations in tropical habitats is just beginning to emerge. Limited studies have been conducted in the eastern tropical Atlantic (Weir et al., 2010), Caribbean Sea (Bolaños-Jiménez et al., 2014; Bolaños-Jiménez et al., 2023), Bahamas (Dunn and Claridge, 2013), Gulf of Mexico (O'Sullivan and Mullin, 1997), eastern tropical Pacific (Olson and Gerrodette, 2008; Vargas-Bravo et al., 2020), Hawaii (Baird et al., 2006), Galápagos Islands (Denkinger et al., 2020), Indian Ocean (Terrapon et al., 2021), and New Guinea (Visser and Bonoccorso, 2003). Because they are comparatively understudied relative to populations at high latitudes, tropical ecotypes/morphotypes are less well defined (Baird et al., 2006; Olson and Gerrodette, 2008; Weir et al., 2010; Bolaños-Jiménez et al., 2014; Vargas-Bravo et al., 2020). Killer whales across the tropics in general appear to have a broad diet and have been observed feeding on a range of prey (e.g., marine mammals, sharks, fish, cephalopods) (e.g., Visser and Bonoccorso, 2003; Baird et al., 2006; Denkinger et al., 2020; Terrapon et al., 2021) and this includes the Caribbean region (Bolaños-Jiménez et al., 2014; Kiszka et al., 2021). In some tropical areas they have only been observed feeding on specific prey (e.g., marine mammals) (Dunn and Claridge, 2013) but no populations have been defined as dietary specialists thus far based on a limited of number observations.
Because killer whales are distributed throughout the world and they have shown a diversity of adaptations (e.g., behavior, prey, social structure and morphology) and, potentially, speciation (Morin et al., 2010, 2024) in high latitude regions, it is important to study them in tropical habitats to understand the complete extent and nature of their potential adaptations. Our goal here is to develop a more comprehensive picture of killer whale ecology in subtropical waters of the Gulf of Mexico and the North Atlantic Ocean off the southeastern United States (U.S.) (Figure 1) and include information on spatial and temporal distribution, strandings, individual sighting histories and movements, potential social structure, genetics, morphology, acoustics and prey. We combine up-to-date killer whale records from published and unpublished sources including U.S. National Marine Fisheries Service (NMFS), Southeast Fisheries Science Center (SEFSC) marine mammal line-transect surveys, other sightings made by the SEFSC and Northeast Fisheries Science Center (NEFSC), public sightings reported directly to the SEFSC and internet reports that could be verified, and stranding reports.
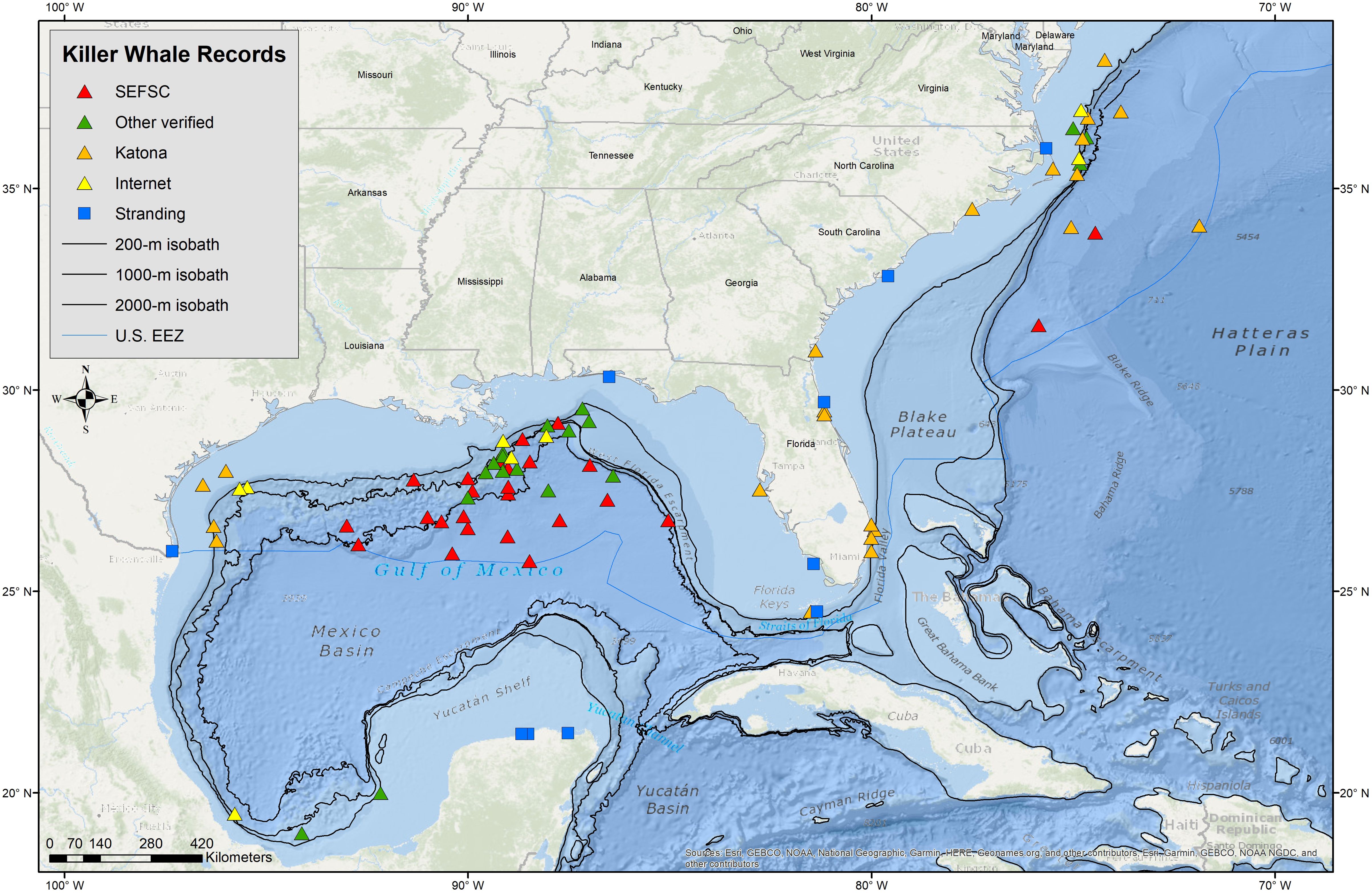
Figure 1. Locations of killer whale sighting (triangles) and stranding (squares) records for the Gulf of Mexico and the SEUS Atlantic Ocean from NOAA SEFSC marine mammal line-transect surveys, verified public reports and other sources, internet reports, strandings, and Katona et al. (1988). SEUS Atlantic study area is confined to U.S. waters from the Gulf of Mexico boundary through the Straits of Florida to (83°W to ~28°N) to Delaware (~38.5°N).
2 Methods
2.1 Spatial and temporal distribution
From 1990 to 2021, the SEFSC has conducted broad-scale, visual, aerial- and ship-based line-transect surveys (Burnham et al., 1980) in continental shelf (20 – 200 m depth) and oceanic waters (>200 m depth) in the U.S. Gulf of Mexico (GoMex) and the North Atlantic Ocean off the southeastern U.S. (SEUS) south of Delaware (Figure 2; Table 1) (e.g., Fulling et al., 2003; Mullin and Fulling, 2004; Mullin et al., 2004; Garrison et al., 2010), primarily to collect data to define cetacean species distribution and estimate abundance (Hayes et al., 2021) to satisfy requirements under the U.S. Marine Mammal Protection Act (MMPA).
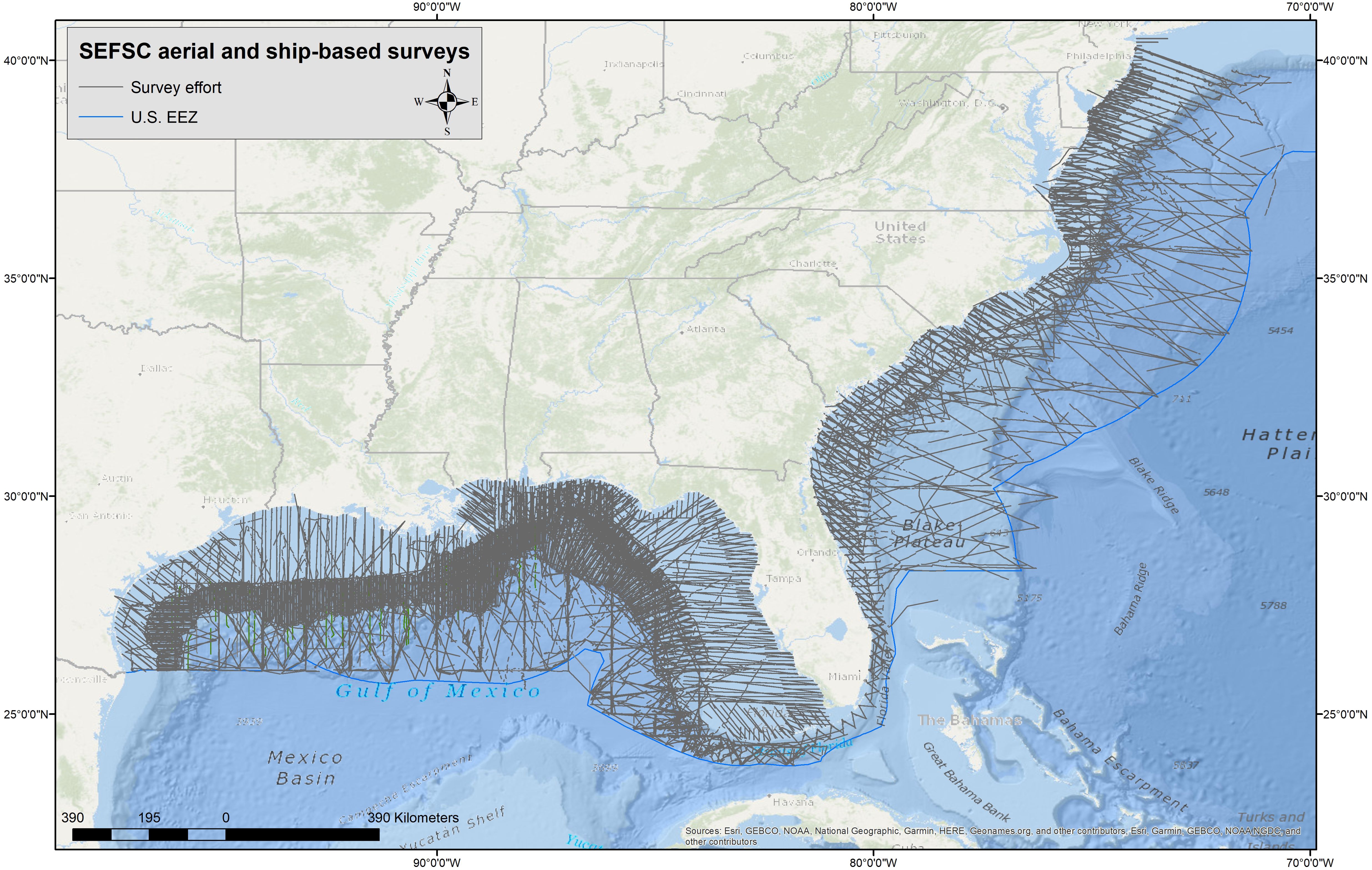
Figure 2. Survey effort during SEFSC aerial- and ship-based line transect surveys from 1990 to 2021 in the Gulf of Mexico and Atlantic Ocean.
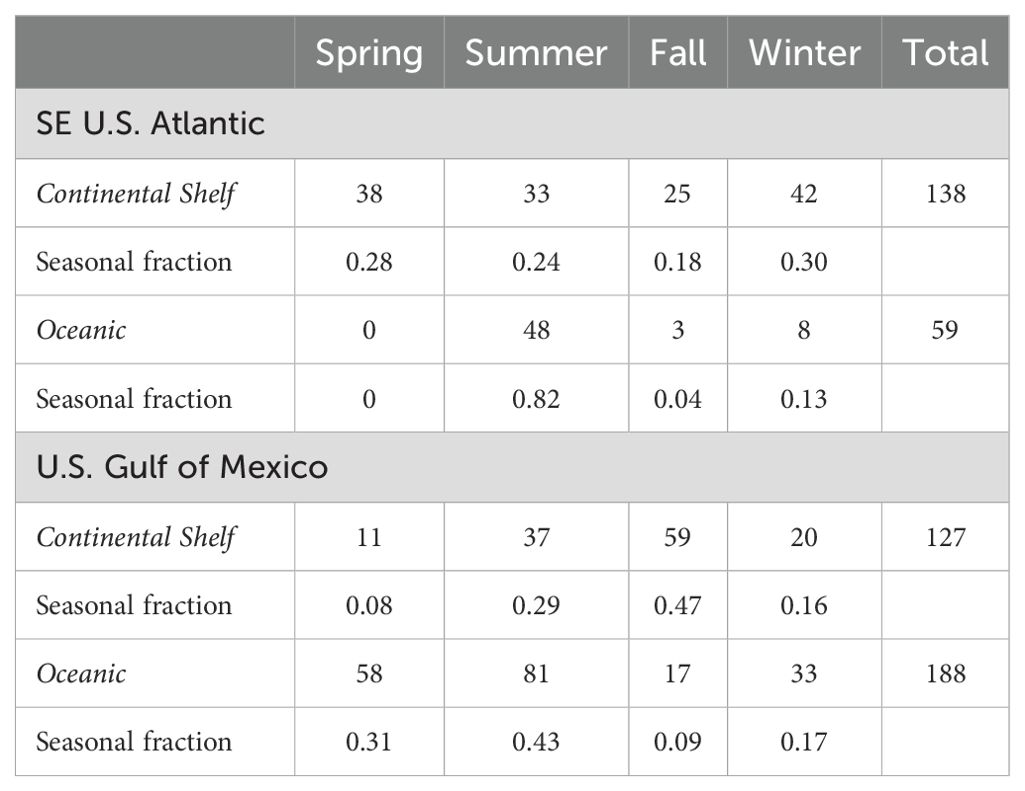
Table 1. Approximate search effort (1000 km) during SEFSC marine mammal research surveys by season in continental shelf (<200 m) and oceanic waters (>200 m) in the Southeast (SE) U.S. Atlantic and U.S. Gulf of Mexico from 1990 to 2021.
When possible, data recorded for killer whale sightings from ship or aerial surveys included group size, sea surface temperature, water depth, the number of calves and adult males, and behavior. Differing survey objectives among years affected the geographic distribution of survey effort. During ship surveys, when possible, photographs were taken to identify individual killer whales primarily based on dorsal fin characteristics (e.g., Bigg et al., 1990). When sighted from the ship in favorable weather, a small boat was often deployed to approach the killer whales to facilitate photography and attempt remote biopsy sampling to obtain a skin and blubber sample for genetic and other analyses. Starting in 2000, an acoustic array was typically deployed during surveys from the ship (see below).
To complement SEFSC line-transect survey sightings, opportunistic killer whale sighting reports were compiled from 1) a 2015 NEFSC line-transect survey in the SEUS study area; 2) NOAA ships conducting fisheries research from 1978 to 2022; 3) those shared by the public directly with the SEFSC from 1989 to mid-2024, with varying degrees of additional information, including photographs, video, date and location; and 4) killer whale sighting information from published sources (i.e., Hairr, 2012; Whitt et al., 2015; Ramírez-León et al., 2020). These sighting records, both opportunistic and from dedicated SEFSC line-transect surveys, include those previously published by O'Sullivan and Mullin (1997).
To supplement spatial and temporal distribution information, opportunistic internet reports of killer whale sightings (e.g., news media, sports fishing blogs) were found by searching with Google® from 2004 to April 2024 using keyword combinations such as appropriate U.S. and Mexico state names with killer whale. Reports were included if they contained sufficiently detailed information to reasonably validate the information (e.g., video, photographs, location, date, and group size).
Finally, sighting and stranding data were compiled to provide a complete overview of killer whales and their spatial and temporal distribution in GoMex and SEUS waters from the following sources: 1) published records from Katona et al. (1988) (Supplementary Table S1) and Schmidly (1981); 2) all database records from OBIS-SEAMAP (https://seamap.env.duke.edu, accessed 27 February 2024); 3) and stranding records from the NOAA National Marine Mammal Health and Stranding Response Database for the Southeast [U.S.] Region (accessed 27 February 2024). Records from each of the sources described were compared so duplicate records could be removed. Data used for analyses came from all or a subset of the data sources described, therefore, except for the SEFSC line-transect sightings, the findings cannot be viewed in the context of survey effort.
2.2 Photo-identification analyses
To study the social structure, external morphology, and ranging and temporal patterns of GoMex killer whales, individual whales were identified using photographic data from SEFSC surveys and from non-NOAA data provided to the SEFSC. Photographs of killer whale dorsal fins and other identifying characteristics were digitized when necessary, cropped, rotated, and sorted per individual. The best photograph(s) were then entered into FinBase (Adams et al., 2006) and quality rated as excellent, good, fair, or poor. Individuals were primarily identified using the notches, scarring, and shape of their dorsal fins, as well as distinguishing characteristics on their bodies [i.e., eye patches (Visser and Mäkeläinen, 2000), saddle patch characteristics, cookiecutter shark (Isistius spp.) scars, and other scars] and assigned a distinctiveness category: very distinctive, distinctive, slightly distinctive, or not distinctive (Mcsweeney et al., 2007; Young et al., 2011). Non-distinctive whales and photos of poor quality were excluded from the analyses. While not ideal, both left- and right-side photos were not available for each identified whale. Three people experienced with photo identification (photo-ID) examined the photos for individual whale identification and matches, and those without agreement from all three people were also excluded.
2.3 Gulf of Mexico social structure
To assess the social structure of GoMex killer whales, groups were defined as all whales sighted in the same location at the same time and showing coordinated behavior, and included all whales within visual range of the vessel (Baird and Dill, 1996). Only one group was ever within visual range at any given time due to the scarcity of killer whale sightings. Groups could contain multiple subgroups. In general, whales within a sighting were described as “tight” groups with less than a body length between individuals, but the maximum separation noted for individuals was 200 m, and the maximum separation noted for subgroups was 50 – 500 m. Affiliate refers to an individual whale sighted in the same group as a specific whale. The total number of affiliates across all sightings of each identified whale were calculated.
We examined the associations of individual killer whales using the program SOCPROG2.9 (Whitehead, 2009). Data were restricted to 49 whales that were slightly to very distinctive and had photos of fair to excellent quality to examine lagged association rate. Analyses of association indices (AIs), displays of the matrix of AIs, and tests for preferred/avoided companions were performed on a subset of 13 whales that were sighted three or more times.
Because not every individual was identified in each group, the half-weight index was chosen to calculate AIs (Cairns and Schwager, 1987) as: 2ab/(a + b), where a = total number of times individual a was seen, b = total number of times individual b was seen, and ab = total number of times a and b were seen together. A dendogram was created using hierarchical cluster analysis (using average linkage) to display the matrix of AIs.
To test for preferred or avoided companions (whales who preferentially associate or avoid one another), it is recommended to use animals sighted a minimum of five times (or in 5 sampling periods) (Whitehead, 2008); however, despite having data spanning 26 years, data were insufficient to use animals sighted five or more times, and hence a lower cutoff of three sightings was used in order to make a preliminary examination of preferred/avoided companions. Permutation tests, as in Bejder et al. (1998) with modifications as in Whitehead et al. (2005), were used to test the null hypothesis that there are no preferred or avoided companions given the number of groups each individual whale was sighted in during each sampling period. The permute groups within samples test and the permute associations within samples test were both performed. The permute groups within samples test tests for preferred companions both within and between sampling periods (short-term and long-term). The sampling period for the permutation tests was set to 5 years. Long-term preferred companionships are shown by a significantly high CV of the real (observed) AIs. Short-term preferred companionships are shown by a significantly low mean of the real AIs. If some individuals avoid each other, the proportion of non-zero AIs should be lower in the real data than in the random data. The permute association within samples test tests for long-term (between sampling period) preferred/avoided companionships, and the sampling period was also set to 5 years. Long-term preferred/avoided companionships are shown by a significantly high SD and CV of the real AIs.
The temporal analyses module in SOCPROG2.9 was used to estimate standardized lagged and null association rates. The lagged association rate is the estimate of the probability that if two individuals are associating now, they will still be associated various time lags later (Whitehead, 1995, 2009). The null association rate is the expected value of the lagged association rate if there is no preferred association given the sighting histories of the individual whales and the number of associations of each individual whale in each sampling period. It will generally be less than or equal to the lagged association rate. When the lagged association rate equals the null association rate, this indicates no preferred associations over these time lags (Whitehead, 2009). “Standardized” rates were used because not all true associates of an individual were recorded during a sampling period in which it was seen (Whitehead, 1995, 2009). Mathematical models were fit to the standardized lagged association rates, and the quasi Akaike information criterion (QAIC) was used to select the best model (Whitehead, 2007).
2.4 Genetic and sex data analyses
Twelve biopsy skin samples were collected from killer whales in the GoMex between 2001 and 2010 (Table 2). DNA was extracted from skin using a standard proteinase K digestion followed by phenol-chloroform extraction (Rosel and Block, 1996) or using a Qiagen DNeasy Blood and Tissue kit following the manufacturer’s protocol. DNA quality was assessed by agarose gel electrophoresis and quantity was measured by fluorometry (Hoefer DyNA Quant 200, GE Healthcare).
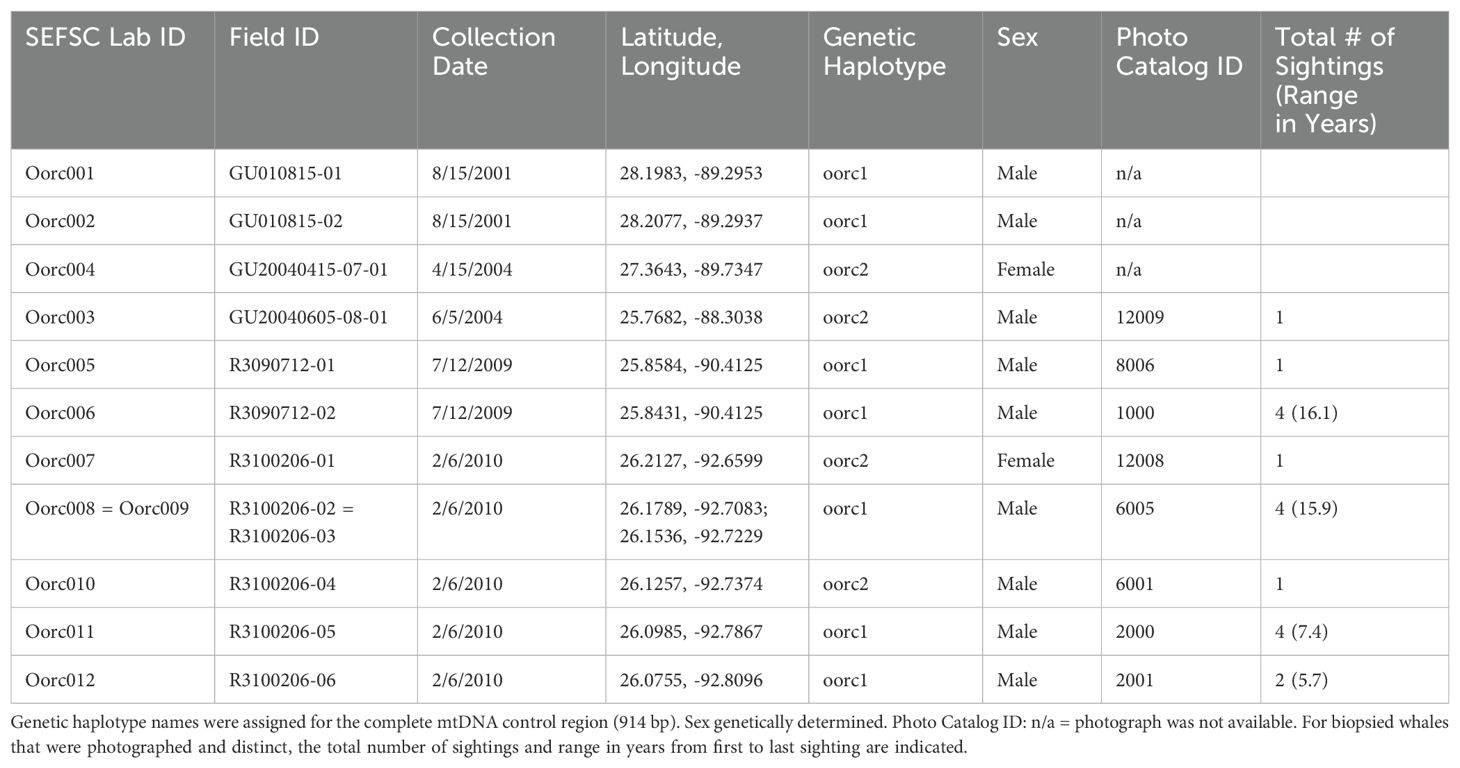
Table 2. Biopsy samples collected from killer whales in the Gulf of Mexico.
The complete mitochondrial DNA (mtDNA) control region was amplified and sequenced in two overlapping fragments. The 5’ portion was amplified using the primer pair L15824 and H16265 (Rosel et al., 1999) with an annealing temperature (Ta) of 55°C. The 3’ end was amplified using the primer pair L16061 (Tolley and Rosel, 2006) and H00651 (Kocher et al., 1989), Ta = 52°C. Each PCR reaction included 25 ng of DNA in 25 µl reactions with 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl2, 150 µM dNTPs, 0.3 µM of each primer, and 1 U Taq DNA Polymerase (Invitrogen). Bovine serum albumin (BSA; Sigma-Aldrich) was added to the reaction at a final concentration of 0.24 mg ml-1 to enhance amplification when needed. The PCR cycling profile was 95°C for 30 s followed by 36 cycles of 95°C for 30 s, Ta as listed above for 30 s, and 72°C for 30 s with a final extension at 72°C for 10 min. PCR products were purified via extraction from low melting point agarose followed by agarose digestion, then sequenced in both directions using the BigDye Terminator v1.1 cycle sequencing kit (Applied Biosystems) and run on an ABI 3500 Genetic Analyzer. Forward and reverse sequences were independently edited and final consensus sequences assembled for each sample using Geneious Prime 2019.2 (https://www.geneious.com). Unique haplotypes were identified in Geneious Prime and then compared to published sequences for killer whales in the GenBank database (www.ncbi.nlm.nih.gov/genbank) using NCBI’s BLAST (Johnson et al., 2008) on 17 May 2023.
The sex of each sample was genetically determined by PCR amplification of ZFX and SRY gene fragments using primer pairs and PCR conditions as described in Rosel (2003) except 25 ng of DNA and 0.75 – 1.5 U Taq DNA Polymerase was used in each reaction. When needed, BSA was added to the PCR reaction at a final concentration of 0.3 mg ml-1. Fragment sizes produced by PCR were visualized via electrophoresis on a 2.5% agarose gel in 1x SB (sodium borate) buffer to determine sex.
When sex could not be determined genetically, some individual killer whales were identified to sex using dorsal fin characteristics. Sexually mature males were defined as individuals with an upright, pronounced dorsal fin in relation to body size (Bigg, 1982). Sexually mature females are individuals with falcate and smaller dorsal fins in relation to body size (Bigg, 1982), however sexually mature females are difficult to distinguish from juvenile males whose dorsal fins have not yet reached full size. To differentiate sexually mature females from juvenile males, whales were only positively identified as females using dorsal fin characteristics if they had a sighting history spanning 15 years indicating they likely had reached sexual maturity (Olesiuk et al., 2005). Animals with a sighting history spanning 10 or 11 years were considered probable females.
2.5 External morphology
We assessed a subset of characteristics following Bolaños-Jiménez et al. (2014) for GoMex killer whales that are used to describe their external morphology: (1) eye patch size – the ratio of the eye patch length and the distance from the blowhole to anterior base of the dorsal fin (Visser and Mäkeläinen, 2000), (2) eye patch orientation relative to the long axis of the body – parallel, up or down (Evans et al., 1982), (3) saddle patch width – wide or narrow, (4) saddle patch intensity – faint, intermediate or conspicuous (Olson and Gerrodette, 2008; Jefferson et al., 2015; Vargas-Bravo et al., 2020; Olson et al., 2023), (5) saddle patch shape – open or closed, and (6) dorsal cape – present or absent (Evans et al., 1982) (Figure 3; Supplementary Figure S1). For GoMex individuals, most of these characteristics were visible in the photographs and they were assessed with an important caveat that all photographs that were generally perpendicular to the dorsal fin were included. Ideally, all assessments would be made from photographs taken from a right angle to the dorsal fin. Photographs were not taken strictly to assess morphology and were collected under a range of field conditions that were many times opportunistic, and where the whales were often evasive. Due to the limited killer whale sighting opportunities, these are most likely the best photographs that will be available for the foreseeable future.

Figure 3. Examples of killer whales from (A-C) the Gulf of Mexico and from (D) the Atlantic adjacent to the southeastern United States. Each of the killer whales shown have no cape, and medium-sized eye patches that are parallel in orientation. Photo B has a saddle patch that is wide, conspicuous intensity, and closed. Photo C has a saddle patch that is wide, faint intensity, and closed. (Additional examples of external morphology are shown in Supplementary Figure S1).
We also assessed individuals for the presence of fresh wounds or scars from bites of cookiecutter sharks (e.g., Dwyer and Visser, 2011; Grace et al., 2018) and presence of pseudostalked barnacles (Xenobalanus globicipitus) (Kane et al., 2008; Whitehead et al., 2015).
2.6 Acoustic data collection and analysis
Acoustic recordings were frequently collected during SEFSC ship-based marine mammal surveys starting in 2000, and recordings associated with five killer whale sightings were available. During these surveys, a multi-element hydrophone array was towed behind the ship to record and localize acoustic signals from vocalizing cetaceans concurrent with visual surveys. The recordings from the summer 2001 cruise were collected with two pre-amplified hydrophones (Benthos AQ4, Benthos, Falmouth, USA), which were recorded to hard-drives at a 48 kHz sample rate with a 400 Hz high pass filter (Edirol USB Audio Interface UA-3 sound card, Roland Corp. USA, Los Angeles, U.S.A; detailed in SEFSC (2001)). The recordings from the surveys over the 2016 to 2018 period were collected with two hydrophones (Reson TC4013, Teledyne Marine, Slangerup, Denmark) and were recorded to hard-drives at a 500 kHz sample rate (custom 12 channel SailDAQ soundcard) and were decimated to 192 kHz with a 1 kHz high pass filter (detailed in GU1605 Appendix B, NMFS, 2016).
All acoustic recordings were analyzed using PAMGuard software (version 1.15.17). Spectrograms with approximately 100 Hz frequency resolution and 1 ms time resolution were reviewed for calls from killer whales and other toothed whales. Echolocation clicks were automatically detected and bearings were calculated based on time-difference-of-arrival methods using the Click Detector module (e.g., Keating and Barlow, 2013). Bearing time plots, which show bearing tracks of individual echolocating whales and dolphins as consecutive clicks received at consistent bearing angles, were reviewed concurrent with spectrogram review. This provided additional context for mixed species acoustic encounters, to distinguish acoustic detections of killer whales and other species. Echolocation signals, pulsed calls including codas, discrete or burst-pulsed calls, and whistles were identified to species by one experienced acoustic analyst and verified by other analysts.
2.7 Predatory/aggressive behavior
Predation or aggressive behavior directed toward potential prey by killer whales observed in the GoMex and SEUS during previously unreported sightings or from the published literature were summarized to better understand the feeding ecology of killer whales. Aggressive behavior was defined as killer whales moving repeatedly or consistently toward potential prey that elicited avoidance (e.g., fleeing, rapid travel away) or distress (e.g., apparent defensive formations, defecation) reactions from the prey.
3 Results
3.1 Spatial and temporal distribution
A total of 49 killer whale groups were sighted in the GoMex from 1978 to mid-2024 (Figure 1; Supplementary Table S2), including 23 recorded during SEFSC marine mammal line-transect surveys, 15 opportunistic sightings [7 from other NOAA surveys, 6 shared by public, and 2 from published sources subsequent to O'Sullivan and Mullin (1997)], six found on the internet with verified photographs or video, and five published by Katona et al. (1988) (Supplementary Table S1). While sightings from dedicated line-transect surveys occurred in a broad area in the oceanic waters of the central GoMex, sightings from all sources show a broader distribution that includes the western oceanic GoMex and two sightings in continental shelf waters off Texas, one near Tampa Bay, Florida, and three in the Bay of Campeche (Figure 1). At least one killer whale sighting (i.e., non-stranding) has been reported in the GoMex in each season of the year based on all sources although there is only one record for winter (Table 3). While most of the sightings from all sources (75%) as well as those from SEFSC marine mammal line-transect surveys (83%) were from spring and summer, the vast majority (74%) of SEFSC line-transect survey effort in oceanic waters also occurred in these seasons (Table 1).
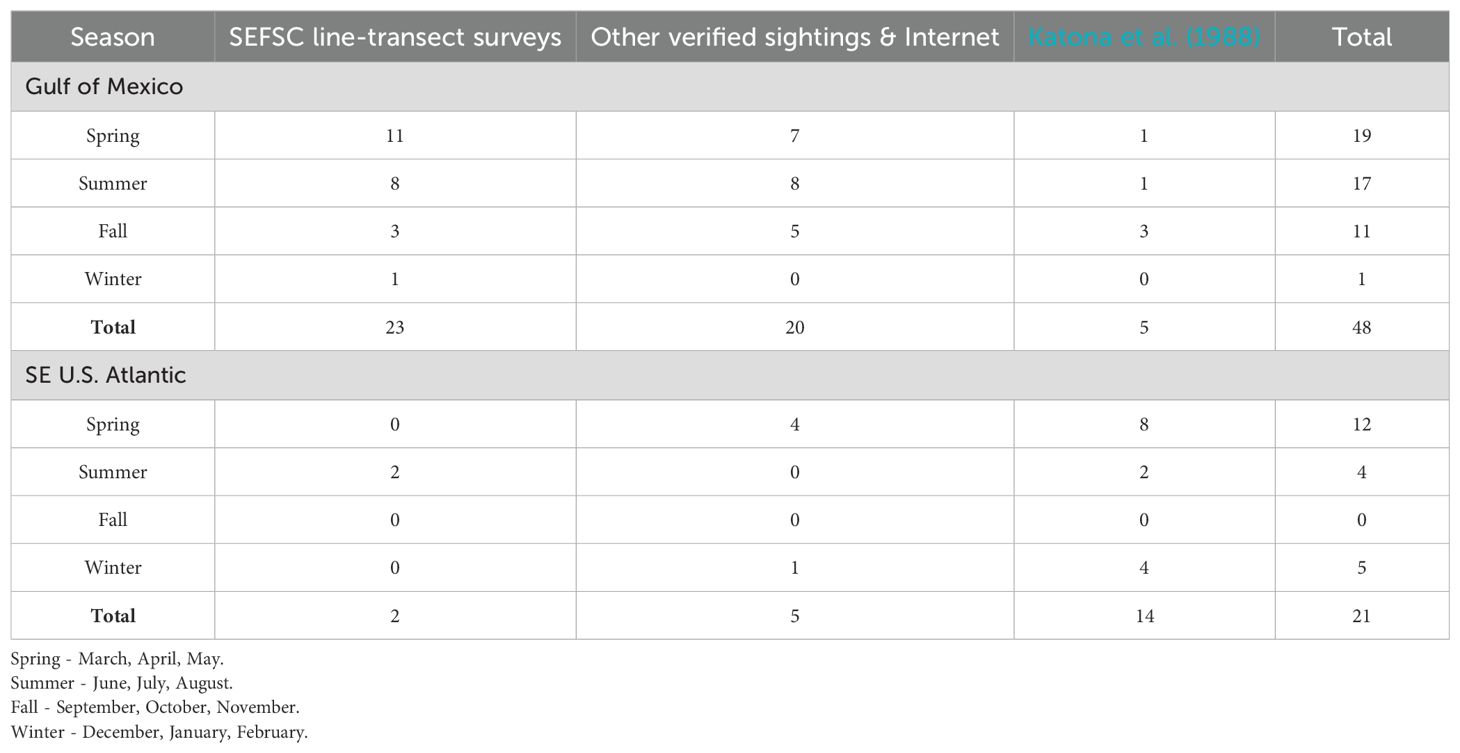
Table 3. Killer whale sighting records by season in the Gulf of Mexico and Southeast (SE) U.S. Atlantic (see Supplementary Tables S1, S2; One Gulf of Mexico and three Atlantic sightings could not be assigned to a season).
More detailed data from SEFSC line-transect marine mammal ship and aerial surveys in the GoMex since 1990 revealed median group size as 6.0 whales and the mean group size, water depth, and sea surface temperature of groups as, 7.1 whales (n = 23, SD = 4.69, range = 1 – 14), 2011 m (n = 23, SD = 826, range = 621 – 3300 m) and 27.3°C (n = 21, SD = 2.35, range = 22.7° – 31.6°C), respectively. Eight of these groups had more than 10 whales and nine had four or fewer whales (Figure 4).
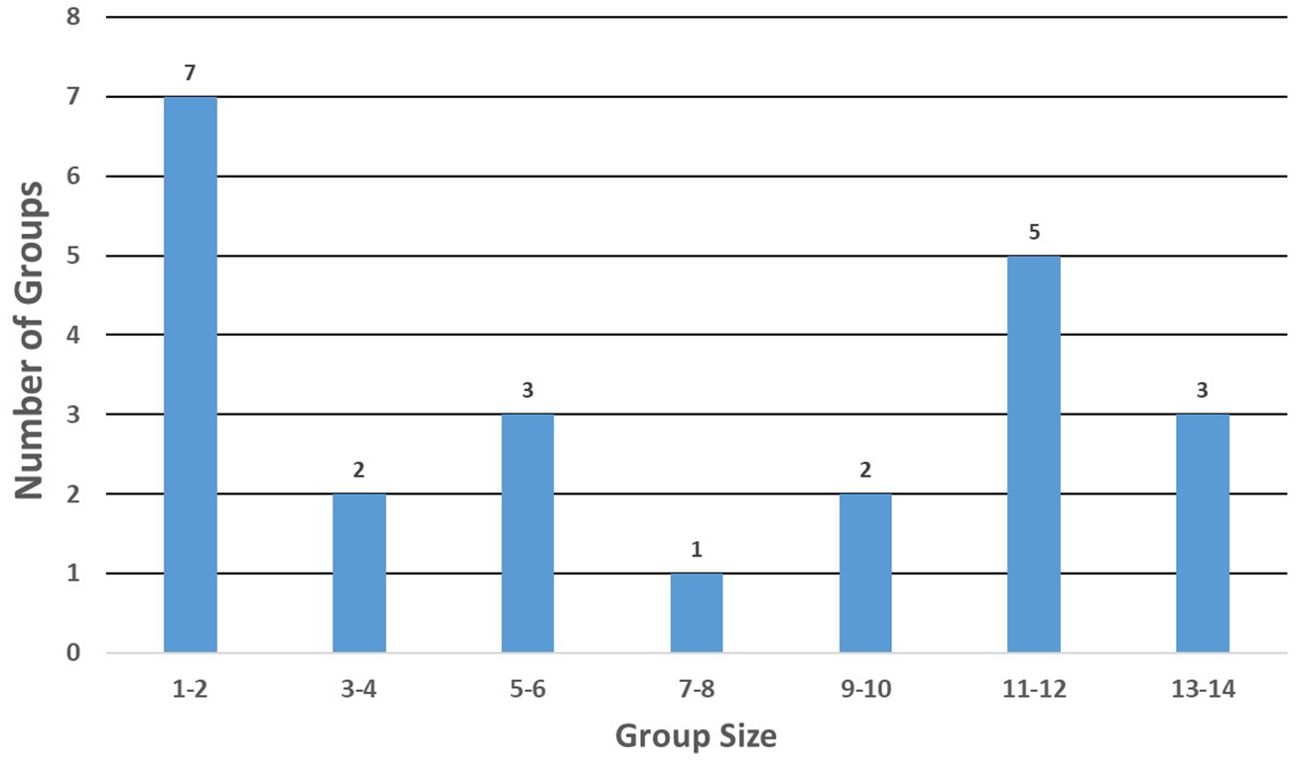
Figure 4. Distribution of group sizes for 23 killer whale sightings made during SEFSC ship and aerial-based line-transect surveys in the Gulf of Mexico from 1990 to 2018.
In the SEUS, a total of 24 killer whale sightings, including three during SEFSC and NEFSC marine mammal line-transect surveys, two from Hairr (2012), two found on the internet with verified photographs or video, and 17 published by Katona et al. (1988) (Supplementary Tables S1, S2), primarily occurred in a broad area off the North Carolina region as well as adjacent to the east coast of Florida (Figure 1). Most killer whale sightings (80%) were from winter and spring, although the two SEFSC sightings were in summer (Table 3) when the majority of survey effort in oceanic waters (82%) in this area occurred (Table 1). Group sizes were 4, 5, and 6 whales from line-transect surveys with two sightings in oceanic waters (>2000 m) and one on the continental shelf (<200 m).
From 1912 through 2023, ten killer whale strandings, all single whales except one, have been reported in the GoMex and SEUS (Figure 1); three in the GoMex and three in the SEUS by Katona et al. (1988) (Supplementary Table S1) and subsequently, three in the GoMex, all on the Yucatán Peninsula, one of which was of five whales (M. R. Ramírez-León, pers. comm.); and one in the SEUS in northern Florida (Supplementary Table S2).
3.2 Photo-ID catalog
Fifty-nine individual whales (22 males, 5 females, 5 probable females and 27 females/juvenile males), including 10 non-distinct whales, from the GoMex are in the photo-ID catalog and were given a unique catalog number (Supplementary Table S3). In the SEUS, photographs were taken during two of the three sightings but only one distinct adult male was clearly identifiable in each group and these males were different. The two males did not match any whale in the GoMex catalog.
3.3 Gulf of Mexico social structure
In the GoMex, 49 distinct individual killer whales were sighted 1 to 7 times (Figure 5) and individuals ranged widely over the north-central GoMex (Supplementary Figure S2). The most frequently sighted killer whale also had the longest sighting history - a male (ID 5000) with seven sightings spanning 26 years (1992 – 2018). The next most frequently sighted whales were two females that were each sighted five times, one (ID 7002) with sightings spanning 18 years (1994 – 2012) and the other (ID 7007) with sightings spanning 17 years (1993 – 2010). A distinctive male (ID 1000) with approximately the top third of his dorsal fin missing was sighted on four occasions spanning 16 years (1993 – 2009), but with a 15-year gap between the most recent two sightings.
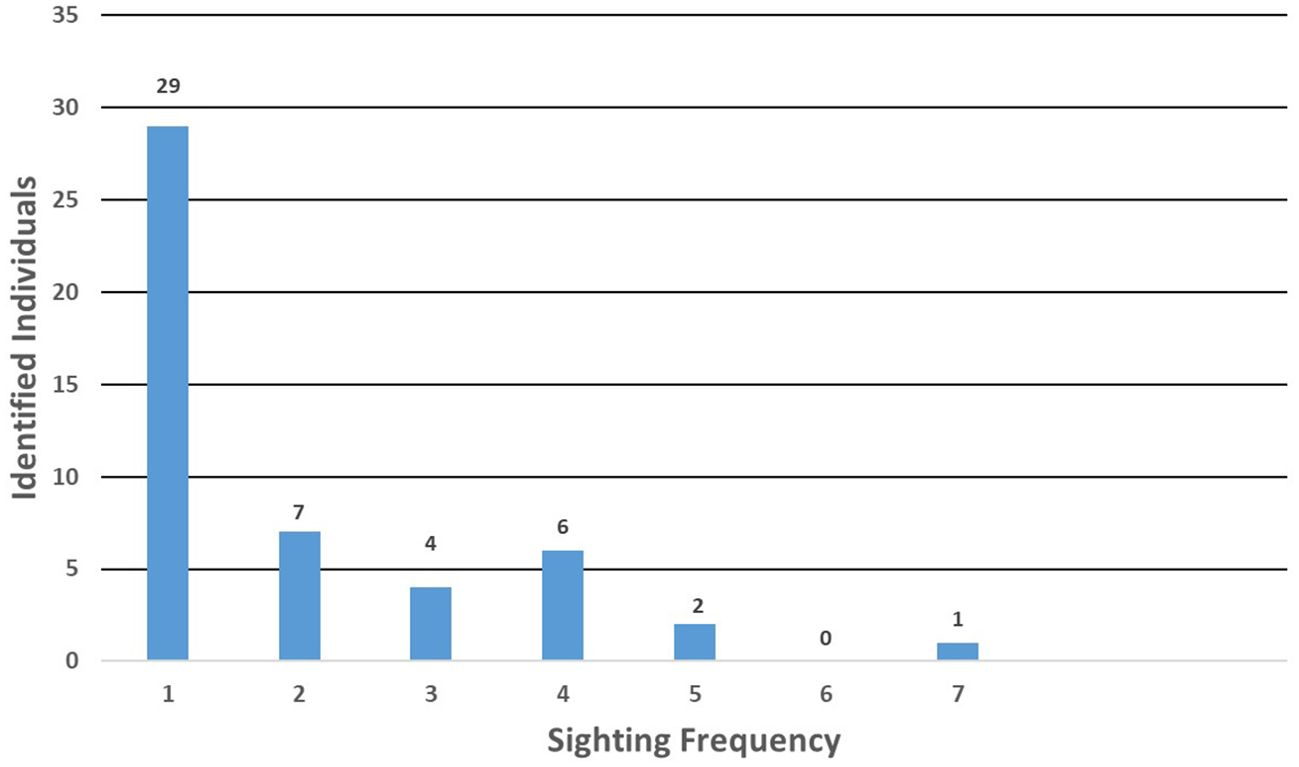
Figure 5. Sighting frequency of 49 individually identified killer whales sighted in the Gulf of Mexico during 21 sightings made during SEFSC vessel surveys from 1992 to 2018 in which photographs were taken for photo-identification.
Total number of affiliates across sightings ranged from 1 to 28 (mean = 9.3). There was evidence of long-term relationships between affiliates. A male (ID 6005) and a female (ID 8005) with severe propeller scarring along her entire peduncle (Supplementary Figure S3) were sighted together four times spanning 16 years (2001 – 2017), and neither whale was sighted absent of the other (Supplementary Table S3). Four whales [1 female (ID 7002), 3 probable females (IDs 7003, 8003, 8004)] were sighted together on 8 June 1994 and again on 15 April 2004, nearly 10 years later. Two whales [1 female (ID 7007), 1 probable female (ID 8004)] sighted together on 21 May 1996 were sighted together on 15 April 2004, nearly eight years later. Three whales [2 males (IDs 2000, 2001) and 1 unknown (ID 6003)] sighted together on 28 May 2004 were resighted together on 6 February 2010, more than five years later. Five whales [1 male (ID 6006), 2 females (IDs 7002, 8005), and 2 unknown (IDs 6003, 7005)] were sighted together on 6 February 2010 and again on 20 September 2012, more than two and a half years later. Two whales [both males (IDs 5000, 6000)] were sighted together on 5 June 1992 and again on 13 June 1993, just over a year apart.
Though based on a small number of sightings, there are multiple lines of evidence for long-term preferred companions: 1) the permute groups within samples test indicated the CV of the real AIs was significantly higher than the randomly generated AIs (p = 0.0360); 2) the permute associations within samples test showed both the SD and CV of the AIs were higher in the real data (p = 0.0040 for both tests) (Table 4) versus the randomly generated data; and 3) the lagged association rate was higher than the null association rate for over 4000 days (~11 years) and the preferred companions model was the best fit (lowest QAIC) (see Table 5). The proportion of non-zero AIs was significantly lower in the real data (p = 0.0230) than the random data, indicating some individuals may avoid each other.
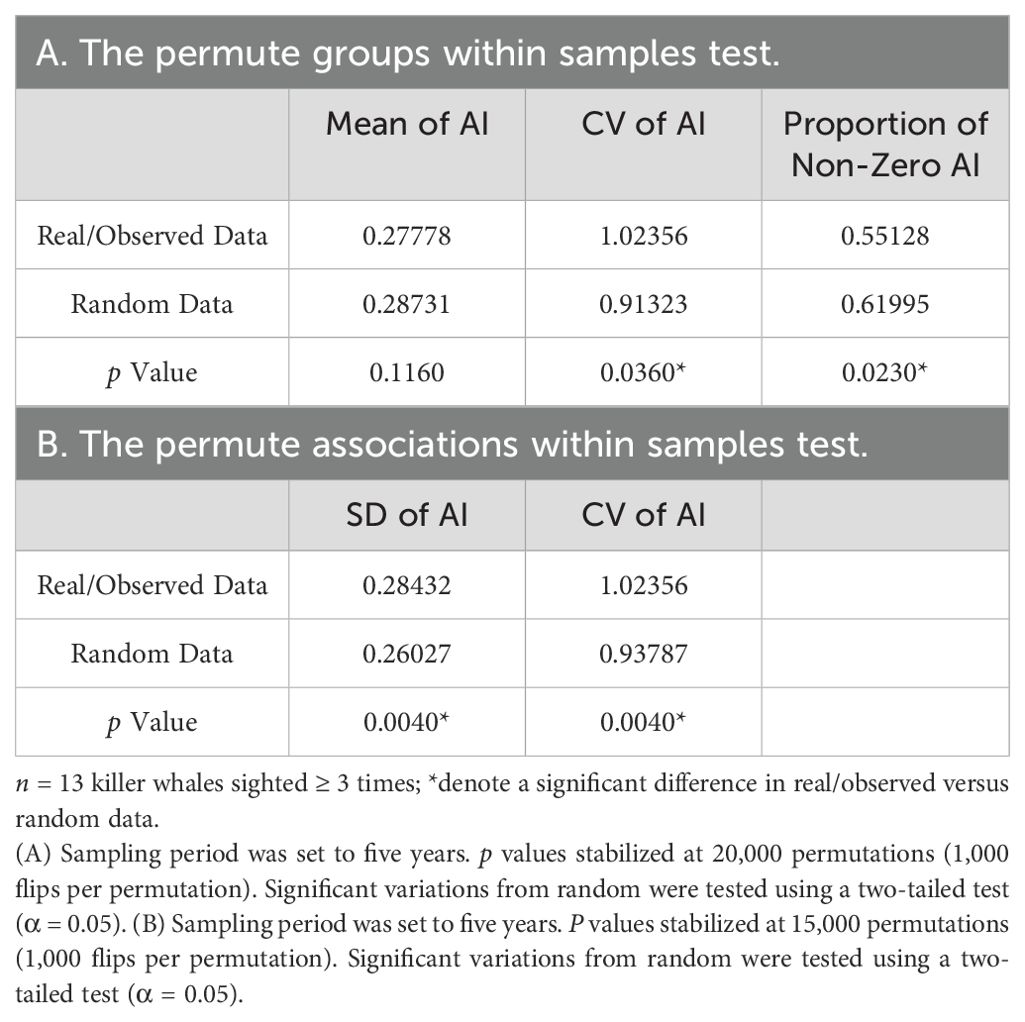
Table 4. Results of permutation tests for preferred/avoided companions.
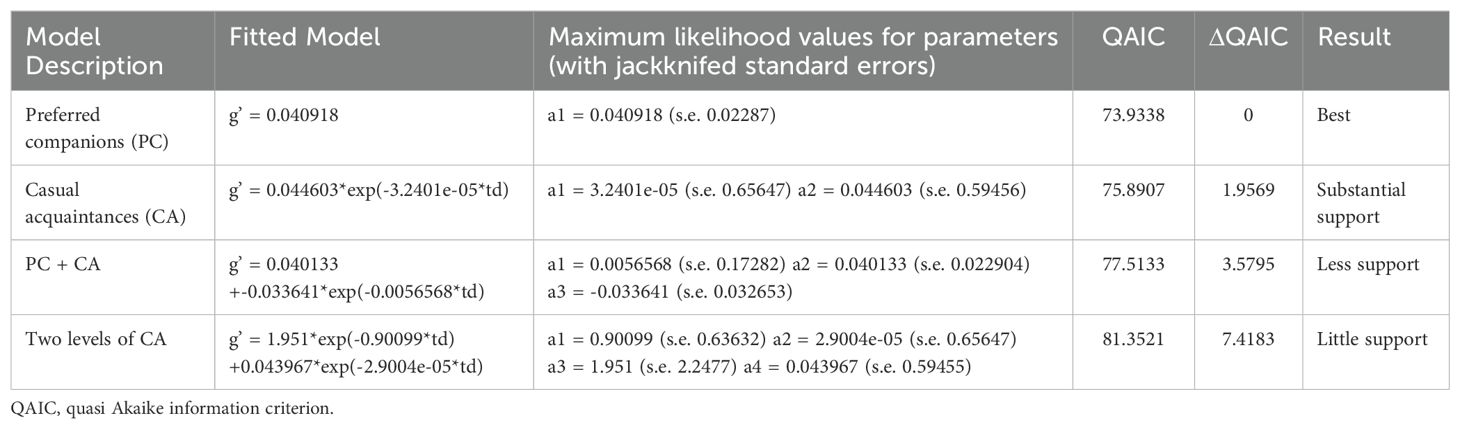
Table 5. Exponential models (td = time lag in days) fit to data on standardized lagged association rates (g’).
The dendogram created using hierarchical cluster analysis (using average linkage) of the 13 whales sighted 3 or more times indicated three major groupings of whales associating at a rate of ~0.2, and within-group associations of 0.5 to 1 (Figure 6).
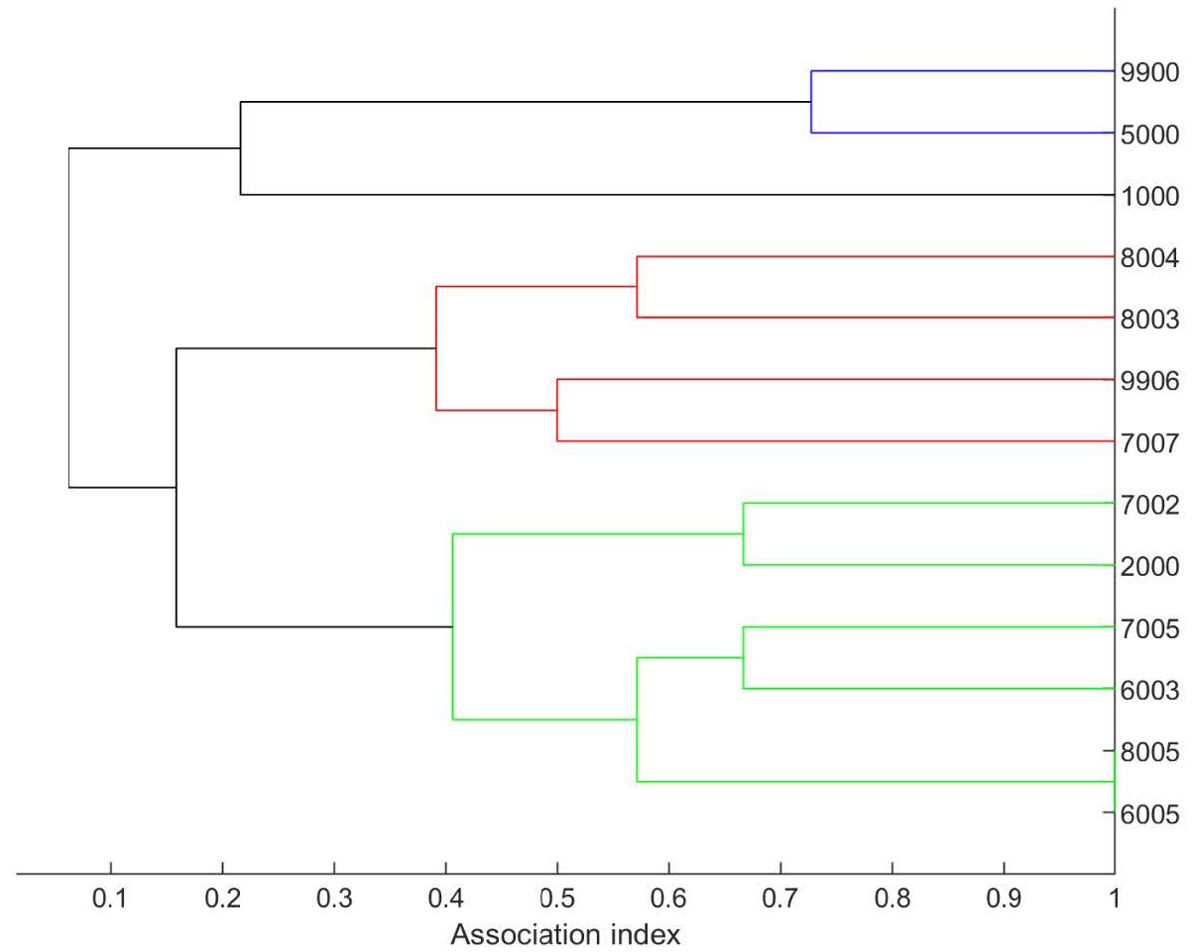
Figure 6. Dendogram generated using average linkage cluster analysis for 13 individual killer whales identified in three or more sightings in the Gulf of Mexico during SEFSC vessel surveys from 1992 to 2018. The numbers along the right axis represent identification numbers for individuals. However, in the case of 9900 and 9906, these identification numbers were shortened from the actual identification for acceptance into SOCPROG2.9. Animal 9900 in Figure 5 is Catalog ID 12000, and 9906 is Catalog ID 12006 as noted in Supplementary Table S2.
3.4 Genetics
The complete mtDNA control region [914 base pairs (bp)] was sequenced for 12 biopsy samples from GoMex killer whales. Two haplotypes were identified among the samples with seven transitional differences between them. Using photo-ID data, one individual (ID 6005) was identified as being sampled more than once and the two genetic samples had matching control region haplotypes and sex. Removal of the duplicate sample resulted in a total of 11 individual animals in the genetic data set (Table 2). Of these 11, eight exhibited distinctive dorsal fins. No dorsal fin photos were available for the remaining three animals. However, based on field notes and the genetic data, we could determine that those three animals were unique from one another – one was a female biopsied in 2001 and two were males biopsied during the same sighting in 2004. Field observations for the latter indicate the two biopsies were collected from an animal with the dorsal fin of an adult male and an animal with a smaller, “female-type” dorsal fin. While these three represent different animals, without dorsal fin photos, we cannot rule out they were re-sampled in later years. Therefore, the 12 biopsy samples represent at least eight confirmed individuals (seven males and one female) and could represent up to 11 individuals (nine males and two females) (Table 2).
Control region haplotype oorc1 (GenBank accession number DQ845456) was found in seven individuals (five with photo-ID data) and in three of the four sampling years (2001, 2009, and 2010). This haplotype is identical to the control region sequence of two other entries in GenBank [accession numbers GQ303372 (Foote et al., 2009) and GU187216 (Morin et al., 2010)], both from the same sample collected in the GoMex in August 2001 and which is also one of the individuals used in our study (Field ID GU010815-02). Oorc1 does not match any known killer whale haplotypes from samples collected outside of the GoMex. The second haplotype, oorc2 (GenBank accession number PP910826), was found in four individuals (three with photo-ID data) during years 2004 and 2010. This haplotype matches the control region sequence of two samples collected in the South Atlantic Ocean: one from the northeastern coast of Brazil and the other from waters near the Cape of Good Hope, Africa (KR180322; Morin et al., 2015), as well as a sample collected in the northwest Pacific near Taiji, Wakayama, Japan (KR180299; Morin et al., 2015).
Genetic samples were collected from multiple individuals during three sightings. On 15 Aug 2001 and 12 July 2009, two genetic samples were collected each and those individuals had matching oorc1 control region haplotypes. Five individuals were sampled during the 06 Feb 2010 sighting and of those individuals, three had the oorc1 haplotype and two the oorc2 haplotype, resulting in one group with mixed mtDNA control region haplotypes.
3.5 External morphology
All 59 whales in the GoMex photo-ID catalog were assessed for morphological characteristics, however, not every whale could be assessed for all characteristics. The ratio of the eye patch length relative to the length from the blowhole to the anterior base of the dorsal fin averaged 31% (median = 31%, range 22 – 38%, n = 36). Eye patch orientation (n = 36) was parallel (80%), down (14%) and up (6%) although in none of the non-parallel cases was the departure dramatic. Saddle patches (n = 49) were wide (76%) or narrow (24%) and were closed (100%). Saddle patch intensity (n = 38) was faint (39%), intermediate (50%), and conspicuous (11%) although lighting could be critical to the interpretation. None of the individuals had a cape. The shape of the dorsal fin tip was not assessed, partially, because they have a range of shapes from very round to more pointed (Supplementary Figure S1). Only seven whales from the SEUS could be assessed. None had a cape and all had eye patches that were parallel and ranged from 28 – 35% in relative length. The saddle patches on only two whales could be assessed and both were faint with one wide and one narrow.
Forty-seven percent of GoMex whales (27/58) had at least one cookiecutter shark bite wound or scar. Some individuals had numerous scars (Supplementary Figure S1). In contrast, no whale from the SEUS had any obvious cookiecutter shark bite wound or scar. Only four whales from the GoMex, each in a different sighting, and one from the SEUS, could be found with a pseudostalked barnacle, and in each case, there appeared to be 1 – 3 barnacles on the tip of the dorsal fin (Supplementary Figure S1).
3.6 Acoustics
Acoustic recordings were available for four visual GoMex sightings and one visual SEUS sighting with killer whales during SEFSC surveys. Of the three GoMex line-transect survey sightings during 2017 – 2018, two had no killer whale calls present and one had only one likely killer whale whistle of 3 s duration detected (Figure 7).
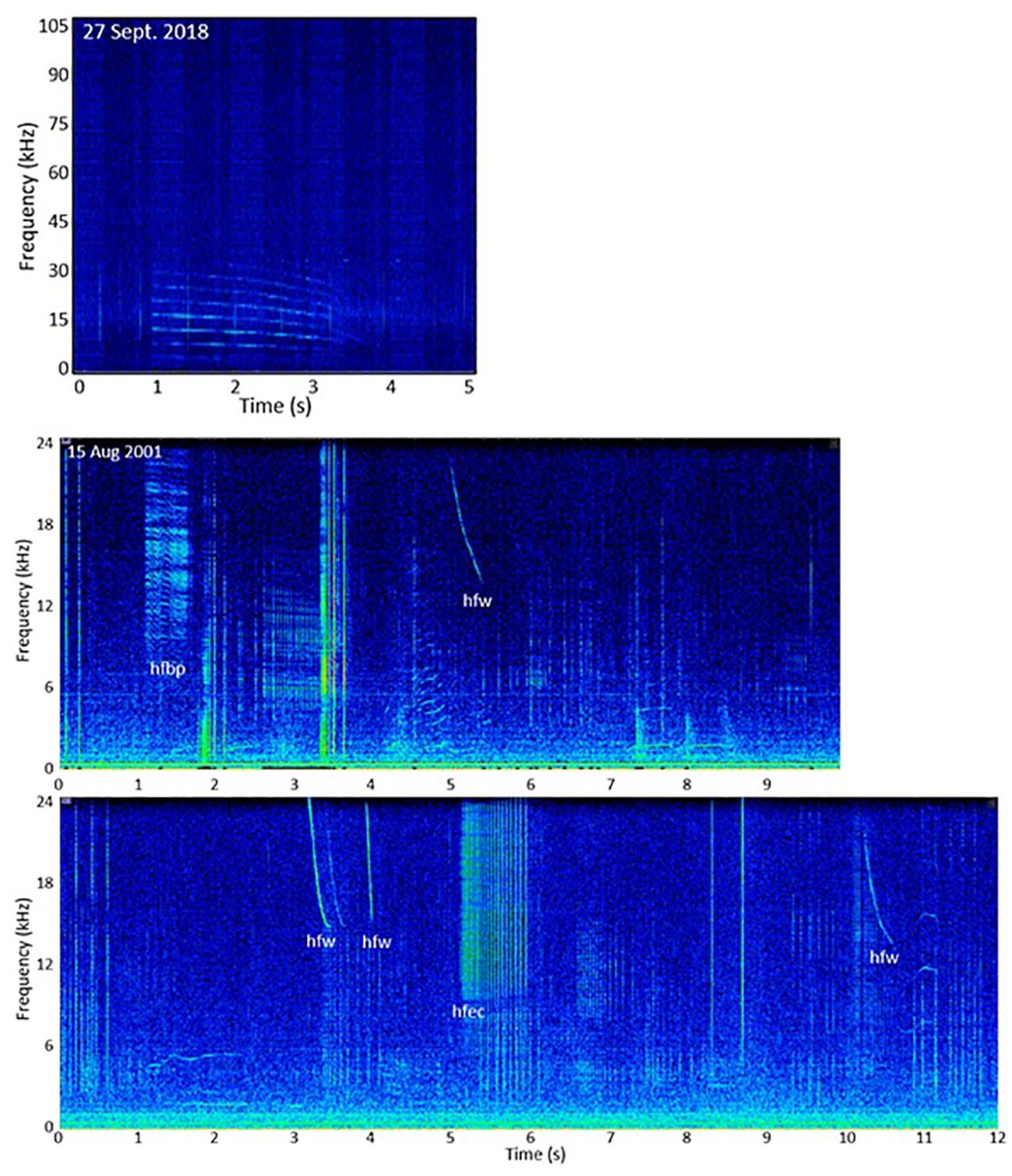
Figure 7. Spectrograms of (upper panel) probable killer whale whistle from 27 September 2018 in the Gulf of Mexico, and (lower two panels) mixed killer whale and sperm whale encounter in the Gulf of Mexico on 15 August 2001 with sperm whale clicks and codas, killer whale high frequency whistles (hfw), burst-pulses (hfbp), and echolocation click packets (hfec), and calls of undetermined origin.
The fourth GoMex encounter with recordings was the 15 August 2001 sighting in which a group of seven killer whales that included one adult male, five female/juvenile males, and one calf was observed harassing sperm whales (see Predatory/Aggressive Behavior Section). In this encounter, sperm whales were originally acoustically detected producing regular clicks (primarily used for foraging/feeding), then switched to producing codas, creaks, rapid clicks, squeals and chirrups along with regular clicks around the same time visual observers noticed the killer whales approaching the sperm whale group, and continued these vocalizations throughout the interaction. Many calls from this encounter could be attributed to killer whales (Figure 7) including 29 high-frequency downswept whistles (24 – 16 kHz, 0.1 – 0.5 s duration), 25 rapid click trains of 20 – 40 clicks (10 – 24 kHz) over 1 – 2 s, and seven high frequency burst-pulses (10 – 24 kHz). The rapid click train bearing tracks followed closely with sperm whale click bearing tracks, suggesting the male killer whale documented to be interacting within the sperm whale group may have produced them. Many lower frequency burst-pulsed calls or whistles (750 – 900 Hz fundamental, 1500 – 1800 Hz harmonic) were also detected throughout the encounter (Figure 7), but it could not be determined whether those were produced by the killer whales or sperm whales. Note, during this encounter, our system bandwidth upper limit was 24 kHz, so it is possible GoMex high frequency whistle ranges for killer whales extend above 24 kHz.
The one SEUS killer whale encounter with recordings was also a mixed species recording, with pilot whales (Globicephala sp.) present and producing sounds. Echolocation clicks with peaks in the 4 – 12 kHz range, attributed to pilot whales, were common throughout this detection. Additionally, there were stereotyped burst-pulsed or discrete calls of 0.5 – 0.75 s duration, rapid click packets (5 – 60 kHz with around 15 clicks in 0.2 s), and graded buzz-burst-pulsed calls of 0.1 to 1 s duration, but these could not be differentiated to whether the killer whales or pilot whales produced them.
3.7 Predatory/aggressive behavior
On 20 September 2012, a group of approximately 15 killer whales was observed from a vessel conducting sea turtle research in the GoMex just south of the Mississippi River delta (613 m deep) attacking several dwarf sperm whales (Kogia sima) (C. McKenzie, pers. comm.). The killer whales circled the dwarf sperm whales, with the larger whales, including at least one adult male, “hitting and wounding” them although no wounds were visible in the photographs. The smaller killer whales would then approach and attack the dwarf sperm whales (Figure 8).

Figure 8. (A-D) Killer whales attacking a Kogia in the Gulf of Mexico on 20 September 2012. (A) The animal is clearly a Kogia based on the size, tapered head, and small underslung jaw. (D) It was identified as a dwarf sperm whale (K. sima) based on the large, erect and pointed dorsal fin (white outline) (Jefferson et al., 2015). Photographs by Caz McKenzie. (Photos are heavily cropped from the original photos).
On 10 June 2024 a group of 15 – 25 killer whales that included two adult males and calves was observed in multiple subgroups from a recreational fishing boat south of the Florida Panhandle in the GoMex (786 m deep). Short cell phone videos show a subgroup of 4 – 5 with females/juveniles and calves pursuing 2 – 3 dolphins that were most likely from the genus Stenella. Due to darkness it is unknown how this event ended (T. Ream, pers. comm.).
On 15 August 2001 while the SEFSC was conducting sperm whale research in the GoMex south of the Mississippi River delta (1070 m deep) a group of seven killer whales was observed harassing sperm whales for 1 hr. This interaction was observed through 25x binoculars from a large ship towing an acoustic array. A small boat was already in the water to collect biopsies from sperm whales located over 3.5 km from the killer whale-sperm whale interaction site and subsequently collected a biopsy from two killer whales and photographs. The sperm whales made notable vocalizations (see Acoustic sections) and displayed some notable behaviors that included more distant sperm whales swimming rapidly to the interaction site, an apparent “defensive” formation, and defecation (Appendix S1 inside Supplementary Data Sheet S1). There was no indication that the killer whales killed or wounded any of the sperm whales.
4 Discussion
4.1 Spatial and temporal distribution
Killer whales inhabit oceanic waters throughout the northern GoMex but there are few marine mammal surveys of the southern oceanic GoMex. Killer whales have been sighted in the southern GoMex and the overall GoMex distribution remains similar to that reported by O'Sullivan and Mullin (1997). While there is a gap in sightings in the western GoMex during SEFSC surveys, there are a number of other sightings off Texas (Figure 1). Also, despite a very large amount of survey effort in continental shelf waters during both SEFSC marine mammal (Table 1; Figure 2) and fisheries research surveys, no killer whales have been reported. While three sightings were reported in GoMex continental shelf waters (Katona et al., 1988), more reports would be expected if killer whales frequented shelf waters, given both the large amount of survey effort and the volume of commercial and recreational activity in GoMex shelf waters.
It is curious that killer whales do not inhabit continental shelf waters in both the GoMex and SEUS more frequently. Shelf waters are productive and in both areas there are tens of thousands of bottlenose dolphins (Tursiops spp.) and Atlantic spotted dolphins (Stenella frontalis), and other potential prey (e.g., fish, sharks and turtles) (Hayes et al., 2021).
Similar to the oceanic GoMex, killer whales are found throughout the eastern tropical Pacific, which is vast, and is one of few other tropical areas that has been systematically surveyed for cetaceans. Over 8000 killer whales are estimated to inhabit the eastern tropical Pacific (19 million km2) (Wade and Gerrodette, 1993) whereas estimates range from 28 (CV = 1.02) to 277 (CV = 0.42) for the oceanic northern GoMex (380 thousand km2) (Waring et al., 2013). Killer whales are also found throughout the Caribbean Sea associated with the Windward and Leeward islands and near the coasts of Central and South America with few records from the open Caribbean. This distribution may be an artifact of lack of survey effort far from land and the distribution of human activities (Bolaños-Jiménez et al., 2023). Otherwise, few broadscale marine mammal assessments of other tropical regions have been made. It is unknown if killer whales are present in the GoMex year-round or if their abundance varies throughout the year. While there are sightings from all seasons there is only one from winter (Table 3). Killer whales are highly mobile and could shift distribution to the southern GoMex (where the SEFSC does not survey) and/or leave the GoMex completely during winter, but this is difficult to ascertain since there is less effort in winter with SEFSC marine mammal oceanic surveys occurring primarily from late spring through early fall and likely less recreational activity in winter. If killer whale photo-ID catalogs in adjacent waters of the Bahamas and Caribbean can be identified, comparisons may help determine whether GoMex individuals move throughout the western tropical North Atlantic. Similarly, satellite tags (e.g., Durban and Pitman, 2012) could potentially help address movement questions.
In the SEUS between 1990 and 2022, only two groups of killer whales were sighted during SEFSC surveys from the Delaware border (~38.5°N) south to the north end of the Bahamas (~27°N) despite considerable survey effort in both continental shelf and oceanic waters (Figure 2, Table 1), and an additional sighting was made during a NEFSC survey in this area. Similar to the GoMex, killer whales in the SEUS appear to prefer deeper waters but in the SEUS sightings are more concentrated along the shelf-edge region. More survey effort has been made in GoMex oceanic waters (~188,000 km) compared to the SEUS (~59,000 km) (Table 1) which may partially account for the difference in the number of killer whale sightings between the two areas during SEFSC surveys. While two survey sightings were off the North Carolina region, killer whales have also been documented off the east coast of Florida (Katona et al., 1988; Dunn and Claridge, 2013; Bolaños-Jiménez et al., 2023). Sightings off the North Carolina region have been previously reported and SEFSC survey results are similar to those of Katona et al. (1988) that show few records from off Georgia and South Carolina.
Few killer whale strandings have been documented in the GoMex and SEUS. In both areas they occur in low abundance, and in the GoMex they are usually distributed well offshore. In general in the GoMex, the stranding/recovery rates of all species that occur offshore tend to be low (Williams et al., 2011). Regardless, killer whales tend to not strand even in areas where they occur closer to shore (Dahlheim et al., 1982). It is interesting that the GoMex strandings on the Yucatán were just west of four sightings in the Caribbean Sea in the Yucatán Channel reported by Bolaños-Jiménez et al. (2023).
4.2 Genetics
Two mtDNA control region haplotypes were identified from killer whales sampled in the northern GoMex. The most common control region haplotype (oorc1) appears to be unique to this region when compared to global data. Interestingly, animals with the oorc1 haplotype were sighted up to four times, with a range of 5.7 to 16.1 years between sightings whereas animals with the less common haplotype (oorc2) were each only sighted once during the course of this research (Table 2; Supplementary Table S3), raising support for some degree of a geographically restricted matriline in the GoMex for some killer whales. Further sampling in adjacent waters of the Caribbean would help clarify the range of this unique haplotype.
Earlier studies revealed that control region data exhibit poor resolution for determining phylogeographic relationships among killer whales, due to the low level of variability in this relatively short DNA fragment; however whole mitogenome analysis can improve inference of evolutionary divergence among ecotypes (Morin et al., 2010; Duchêne et al., 2011; Foote et al., 2013). The mitogenome from a GoMex killer whale with the oorc1 control region haplotype was sequenced in previous research and phylogenetic analysis placed it in a clade with samples collected near Antarctica and the Crozet Archipelago, the western South and eastern North Atlantic, and the western South Pacific oceans (mitogenome MtGen_60 in phylogenetic clade 6; Morin et al., 2015). The whole mitogenome has yet to be sequenced for an individual from the GoMex with the oorc2 control region haplotype. However, this oorc2 haplotype sequence is identical to multiple samples for which the whole mitogenome has been sequenced (mitogenomes MtGen_120, MtGen_124 and WNPjap in phylogenetic clade 12; Morin et al., 2015). These mitogenomes grouped together in a phylogenetic clade separate from the GoMex animals with the oorc1 haplotype (Morin et al., 2015). Samples within mitogenome clade 12 were collected from the North and South Atlantic, western North Pacific, and Indian oceans, as well as waters near southwest Australia, western Thailand, and Antarctica (Morin et al., 2015). To understand whether the GoMex killer whales are a genetically distinct population, further sampling is needed to fill in geographic gaps, particularly in the Caribbean and Atlantic Ocean, with genetic analysis using mitogenome or nuclear data to help resolve the evolutionary relationship among individuals in the GoMex and nearby waters.
4.3 Gulf of Mexico social structure
Although the dataset was small, some aspects of social structure were apparent. Individual killer whales in the GoMex do preferentially associate with other killer whales, and they associate with these animals over the long-term, for at least one to two decades. However, an individual’s network of affiliates is large, meaning individuals were sighted with many other individuals, and animals were found in a range of group sizes. Denkinger et al. (2020) suggested that in the Galápagos and eastern tropical Pacific region, another generally oligotrophic area, killer whales live in a fission-fusion society (Connor et al., 2000) where small groups may fuse to form larger groups when it is beneficial to forage on larger prey, such as baleen whales. Our results are consistent with fission-fusion societies for GoMex killer whales but given the limitations of our data it is too early to make that conclusion. If GoMex killer whales are generalists and consume a variety of prey, as has been found for other subtropical to tropical areas (see below), then we would expect group sizes to vary and social structure to be somewhat flexible. The mtDNA control region data identified multiple matrilines within one group (during the 06 Feb 2010 sighting) with one haplotype more globally dispersed and the other representing a separate maternal lineage possibly restricted to the GoMex. The evidence for a geographically restricted mtDNA haplotype might be suggestive of a historically more geographically isolated group of whales, with the total range of this population still undefined. More work is needed to better understand the social structure of GoMex killer whales and how it relates to predatory behavior, prey, and evolutionary history of this population.
4.4 External morphology
GoMex and SEUS killer whales generally look like the ‘classic’ black and white killer whales found worldwide (Pitman and Ensor, 2003) (Figure 3). GoMex killer whales can generally be described as having no dorsal cape and oval medium-sized eye patches (see Pitman and Ensor, 2003) that are parallel to the axis of the body. Saddle patches were all closed but varied in intensity (39% faint and 61% non-faint) and width (76% wide and 24% narrow). There was no indication that these characteristics were confined to a group of whales or to whales with a specific mtDNA haplotype. For example, similar to a group in the Pacific (Olson et al., 2023), whales in the GoMex with both narrow and wide saddle patches were seen together in the same groups (Supplementary Figure S1). Eastern North Pacific ecotypes have external morphological characteristics that are different from each other but are uniformly consistent across ecotypes that allows them to be differentiated by experienced observers in the field (Morin et al., 2024). This does not appear to be the case for GoMex and none of the morphological characteristics evaluated allowed us to group GoMex whales together or to diagnose a single whale as a GoMex whale.
This is generally similar to other tropical populations, e.g., those described for the Caribbean (Bolaños-Jiménez et al., 2014), tropical West Africa (Weir et al., 2010), Hawaii (Baird et al., 2006), and the wider eastern tropical Pacific (Olson and Gerrodette, 2008), where no unique morphotypes have been clearly defined. However, an “Eastern Tropical Pacific” ecotype has been proposed for a nearshore population based on studies from the Central Mexican Pacific (Vargas-Bravo et al., 2020). It should be noted that sample sizes from these tropical studies are usually small.
Morphological characteristics have been described for other tropical areas but only quantified for the Caribbean (Bolaños-Jiménez et al., 2014) and the Mexican Central Pacific (Vargas-Bravo et al., 2020). Aspects of morphology of GoMex killer whales appear to be similar to those in both areas (i.e., no cape, closed saddle patches, and a preponderance of parallel eye patches). However, saddle patches in the GoMex were primarily non-faint saddle patches whereas about 85% and 100% were faint in the Caribbean and Mexican Central Pacific, respectively. Also, saddle patches were primarily wide in the GoMex but all were narrow in the Caribbean. Similarly, saddle patches for Hawaii (Baird et al., 2006) and the wider eastern tropical Pacific (Olson and Gerrodette, 2008) are also generally described as faint and narrow, although there is variation. A better understanding of the relationship between Caribbean and GoMex killer whales will require genetic studies along with tagging and photo-ID studies. Given the mobility of killer whales and the adjacent proximity of the GoMex and Caribbean, including the Bahamas, it seems likely that individuals would move between these areas at least to some degree.
Cookiecutter shark scars or wounds were commonly observed, being clearly visible on nearly half of the GoMex killer whales. This is certainly a minimum because seeing scars in photographs depends on lighting, the part of the whale that is visible, and photo quality. Scars are most visible on the saddle patch and tend to be obscured on the darkest part of the whale. Cookiecutter shark wounds commonly occur on deepwater cetaceans in the GoMex and elsewhere (Pérez-Zayas et al., 2002; Dwyer and Visser, 2011; Wenzel and López Suárez, 2012; Best and Photopoulou, 2016; Grace et al., 2018). In contrast, the few whales sighted and photographed in the SEUS did not appear to have cookiecutter shark bites or scars. If they really do not have bites/scars, it would indicate that these whales do not spend a lot of time in deep tropical to warm temperate waters inhabited by cookiecutter sharks (see Wenzel and López Suárez, 2012), which could suggest they do not mix with GoMex killer whales.
In the eastern tropical Pacific, 38% of killer whale groups had Xenobalanus infestations (Kane et al., 2008) compared to 19% in the GoMex. Both areas are largely oligotrophic but the eastern tropical Pacific, which is much larger, has regions of higher productivity. In both areas the intensity of the infestation was similar with typically three or fewer barnacles per whale.
4.5 Acoustics
In the GoMex, three of the four killer whale visual encounters with recordings had no or only one obvious killer whale call detected throughout the encounter, though other marine mammals were acoustically detected during each. Mammal-eating killer whales vocalize infrequently and may use passive listening when hunting, instead of echolocation, to avoid alerting their prey, with vocalization rates increasing after a successful hunt (Barrett-Lennard et al., 1996; Deecke et al., 2005). The limited vocalizations from these three groups are compatible with hypotheses that they were hunting marine mammals, and is consistent with findings of limited vocalizations from killer whales during sperm whale harassment and Kogia spp. predation events in the GoMex and Bahamas, respectively (Dunn and Claridge, 2013; Whitt et al., 2015). However, this does not rule out alternative hypotheses, as fish eating whales can also be silent for extended periods.
Conversely, the killer whales harassing sperm whales in the GoMex in August 2001 produced many calls, including high frequency whistles (HFW) similar to whistles described for multiple ecotypes from all oceans over the last decade (Filatova et al., 2012; Simonis et al., 2012; Trickey et al., 2014; Andriolo et al., 2015; Samarra et al., 2015). The downswept HFWs we recorded in the 24 to 16 kHz range were distinctly different from the higher frequency and variable frequency modulation HFWs produced by herring eating killer whales from Iceland and Norway (Samarra et al., 2010, 2015). On the other hand, they share strong similarities to those recorded from mammal-eating killer whales from Antarctica (Trickey et al., 2014) and the western South Atlantic (Andriolo et al., 2015), as well as those from offshore and unknown ecotypes in the North Pacific (Samarra et al., 2010; Filatova et al., 2012). Interestingly, killer whales produced the HFWs recorded in the western South Atlantic during an attack on sperm whales (Andriolo et al., 2015), but in contrast to the GoMex encounter in 2001, they were only produced when traveling and not during close approaches to the sperm whales. The function of HFWs remains unknown. They may be used for short-range communication (Samarra et al., 2010), in situations where high frequencies would not be detected by prey or competitive species (Simonis et al., 2012), or they could be an important component of the attack stage during sperm whale or other marine mammal interactions.
4.6 Predatory/aggressive behavior
The dwarf sperm whale predation event described here is the first reported for killer whales in the GoMex, though Kogia spp. have previously been identified as a killer whale prey species (Mcalpine, 2009; Dunn and Claridge, 2013; Olson et al., 2023). In fact, Madsen et al. (2005) hypothesized that K. breviceps evolved to use narrow-band high frequency (NBHF) clicks as a way of evading acoustic detection by killer whales. Pitman et al. (2003) described killer whales in the GoMex killing and consuming at least one pantropical spotted dolphin (Stenella attenuata). In addition to Kogia spp. and pantropical spotted dolphins, other cetacean species that occur in the GoMex and SEUS [e.g., Atlantic spotted dolphins, Fraser’s dolphins (Lagenodelphis hosei)] have been documented as killer whale prey elsewhere (e.g., Jefferson et al., 1991; Dunn and Claridge, 2013).
Whitt et al. (2015) describe a 2011 event where killer whales harassed sperm whales in the GoMex that did not appear to result in any sperm whale injury or mortality. Many of the behaviors observed for both species during the killer whale-sperm whale interaction on 15 August 2001 in the GoMex were similar to those observed by Whitt et al. (2015), and also in the Bahamas (Dunn and Claridge, 2013) and near the Galápagos Islands (Arnbom et al., 1987) where there was no evidence of sperm whales being killed or seriously wounded. However, killer whales killing or wounding sperm whales was observed in the Pacific Ocean (Pitman et al., 2001) and Atlantic Ocean off Africa (Weir et al., 2010).
In some areas, killer whales feed on Atlantic bluefin tuna (Thunnus thynnuss) (e.g., Guinet et al., 2007) and they have been observed feeding on tuna off North Carolina (Hairr, 2012). Killer whales have not been reported preying on tuna in the GoMex but it is one of the primary western North Atlantic bluefin tuna spawning areas (Knapp et al., 2014) and there is a commercial pelagic longline fishery for yellowfin tuna (T. albacares) (Teo and Block, 2010). Off Brazil, killer whales depredate tuna (Thunnus spp.) and swordfish (Xiphias gladius) caught in the longline fishery (Dalla Rosa and Secchi, 2007). There has been one recorded killer whale take in the commercial pelagic longline fishery in the GoMex where an adult male was entangled and released alive (Garrison et al., 2009; S. Cushner, pers. comm.). The species composition from the haul was primarily mahi mahi (Coryphaena hippurus) and yellowfin tuna, but no damage to the catch indicative of depredation was noted in the data (S. Cushner, pers. comm.). There have been no known observations of GoMex killer whales preying on non-marine mammals but, in addition to tuna, other known prey species such as leatherback sea turtles (Dermochelys coriacea) (e.g., Pitman and Dutton, 2004; Elwen and Leeney, 2011), elasmobranchs (e.g., Fertl et al., 1996; Pyle et al., 1999; Reyes and García-Borboroglu, 2004; Visser, 2005; Alava and Merlen, 2009; Terrapon et al., 2021), and sunfish (Mola mola) (e.g., Weir et al., 2010; Ryan and Holmes, 2012), also occur in both the GoMex and SEUS.
While studies have shown that high latitude subpopulations of killer whales appear to have rigidly-defined prey preferences (e.g., fish or marine mammals) (Jefferson et al., 2015), recent studies indicate, at least in the North Atlantic, there are not rigid prey preferences but more general tendencies in populations with individual variation (Lefort et al., 2020; Remili et al., 2023). While not observed in the GoMex, consumption of non-marine mammal prey such as fish cannot be ruled out because of the likelihood that capture and/or consumption would occur below the surface and be less observable or unobservable, as opposed to the inherently surface-oriented capture/consumption of air-breathing marine mammals (e.g., Hanson et al., 2010).
In the high latitude North Atlantic and North Pacific, killer whale group size may be indicative of their primary prey where, while variable, those that prey primarily on pinnipeds, dolphins or small whales tend to occur in small groups (e.g., ≤5) (Baird and Dill, 1996; Ford et al., 1998; Saulitis et al., 2000; Higdon et al., 2012) whereas those that feed primarily on fish occur in larger groups (e.g., >10) (Ford et al., 1998; Baird, 2000; Tavares et al., 2017; Olsen et al., 2020). In areas where killer whales may not be dietary specialists, the relationship between prey and group size may be harder to define. Baird et al. (2006) suggest that killer whales in low-productivity tropical and oceanic areas should have a wider-range of prey than those inhabiting productive waters where specific prey is more abundant, and evidence from several tropical areas indicates that killer whales feed on both marine mammals and other species (Baird et al., 2006; Sorisio Sonnino et al., 2006; Alava and Merlen, 2009; Weir et al., 2010; Alava et al., 2013; Bolaños-Jiménez et al., 2014; Terrapon et al., 2021). Groups sizes reported from many tropical areas tend to be relatively small with means ranging from 3.4 to 5.6 whales (Wade and Gerrodette, 1993; García-Godos, 2004; Baird et al., 2006; Weir et al., 2010; Dunn and Claridge, 2013; Testino et al., 2019; Vargas-Bravo et al., 2020; Castro-Azofeifa, 2021; Bolaños-Jiménez et al., 2023). In the GoMex, where killer whales have only been observed feeding on marine mammals, the mean group size was larger, 7.1, and the groups observed preying on S. attenuatta and K. sima were estimated to have 12 and 15 whales, respectively, and those harassing sperm whales had 5 and 7 whales. However, given the small sample size it is too early to draw conclusions about the prey in the GoMex. The situation in the Bahamas is similar where killer whales have only been observed preying on marine mammals (Dunn and Claridge, 2013) but other species, such as large pelagic fish, cannot be ruled out.
5 Conclusions
Historical data including 30 years of discontinuous survey data on killer whales from the GoMex and SEUS provide insights into their biology and ecology:
1. Killer whales in the GoMex almost exclusively inhabit oceanic waters (>200 m deep) and there are few records from the extensive continental shelf. Those in the SEUS are more concentrated in the shelf-edge region but also inhabit deeper waters. For both areas, there is an abundance of potential prey, both marine mammal and other species, throughout continental shelf waters so it is curious that killer whales do not inhabit shelf waters more frequently.
2. Killer whales are uncommon but routinely sighted in the GoMex in all seasons but winter and are part of the oceanic cetacean community, and this includes specific individuals sighted over decades. Like other killer whales in the world, there is evidence of long-term associations of individual whales.
3. Killer whales, including individuals, range widely in the oceanic GoMex, whereas most records for the SEUS are off the east coast of Florida and the North Carolina region.
4. There is a genetic haplotype, thus far, found only in the GoMex which leaves open the possibility of a GoMex population or at least a group of individuals that are descendants of a maternal lineage not found elsewhere.
5. GoMex and SEUS killer whales are, except for Antarctic/subantarctic morphotypes Types B, C and D, broadly similar in external appearance to other killer whales found throughout the world. Compared to other tropical populations killer whales in the GoMex appear to have a higher prevalence of wide non-faint saddle patches.
6. Based on limited information, killer whales may be only situationally vocal. Killer whales harassing sperm whales were very vocal, otherwise they made few vocalizations.
7. No conclusions can be made with respect to prey specialization in either area, only that GoMex whales have been observed feeding on marine mammals and those in the SEUS have been observed feeding on tuna.
Clearly there is much more to learn about all aspects of killer whale biology in the GoMex and SEUS and the tropics in general. Directed killer whale studies in these areas would be extremely difficult and inefficient because killer whales occur far from shore and are infrequently encountered. Therefore, it is important that cetacean researchers working in these areas are prepared with sampling protocols to collect a full suite of photo-ID, behavioral, morphological, acoustic, and genetic data when killer whales are encountered, and to take advantage of additional information that can be gleaned from sightings shared by other researchers and the public. Additionally, it is critical that researchers throughout the GoMex, western North Atlantic, and Caribbean work cooperatively and share information and insights so that a regional understanding of killer whale biology can be achieved.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, PP910826 and https://www.fisheries.noaa.gov/inport/item/30211.
Ethics statement
The animal study was approved by NMFS Atlantic Institutional Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements. All NMFS marine mammal research was conducted under MMPA Research Permits issued to the SEFSC (Nos. 738, 779-1339, 779-1633, 14450, 21938) and the NEFSC (No. 17355).
Author contributions
KB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Validation. KM: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing, Funding acquisition, Project administration, Supervision. KM-F: Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. LWT: Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing. PR: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. MS: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. LAD: Data curation, Investigation, Methodology, Visualization, Writing – review & editing. MR-L: Data curation, Investigation, Methodology, Visualization, Writing – review & editing. JL: Conceptualization, Funding acquisition, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. NMFS sighting data were collected during ship and aerial surveys funded by NMFS and the Bureau of Ocean Energy Management, U.S. Department of Interior. This research was carried out (in part) under the auspices of the Cooperative Institute for Marine and Atmospheric Studies (CIMAS), a Cooperative Institute of the University of Miami and the National Oceanic and Atmospheric Administration, cooperative agreement # NA20OAR4320472.
Acknowledgments
We thank the officers and crews of NOAA ships Oregon II, Gordon Gunter and Pisces for their assistance on research surveys. We thank the many scientists who have participated on these surveys and helped collect and review data including Anthony Martinez, Wayne Hoggard, Carrie Sinclair, and Annie Gorgone. We thank Jason Parkhurst, Andrew Millet, Anthony Wyler, Ian Searles, Travis Ream, and Justin Weeks for sharing information on the killer whale sightings they made. Many thanks to Caz McKenzie for providing photographs and a description of killer whales predating on dwarf sperm whales that he witnessed. We thank Debra Palka for sharing photographs and information on the killer whale sighting made by the NEFSC that is included in this paper. We are grateful to Raena Cota for assisting with genetic data collection. We thank Sascha Cushner for providing information on the killer whale interaction with the pelagic longline fishery, and Paula Olson for sharing insights into killer whale biology. We thank Mirdula Srinivasan and Lance Garrison for reviewing drafts of the manuscript. Finally, we are thankful for the joyous life of Kevin Barry.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
The views expressed here are those of the authors and do not necessarily reflect the views of the U.S. National Marine Fisheries Service, NOAA.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1460314/full#supplementary-material
References
Adams J. D., Speakman T., Zolman E., Schwacke L. H. (2006). Automating image matching, cataloging, and analysis for photo-identification research. Aquat. Mammals 32, 374–384. doi: 10.1578/AM.32.3.2006.374
Alava J., Merlen G. (2009). Video-documentation of a killer whale (Orcinus orca) predatory attack on a giant manta (Manta birostris) in the Galápagos Islands. Latin Am. J. Aquat. Mammals 7, 81–84. doi: 10.5597/lajam00139
Alava J. J., Smith K. J., O'hern J., Alarcón D., Merlen G., Denkinger J. (2013). Observations of killer whale (Orcinus orca) attacks on Bryde's whales (Balaenoptera edeni) in the Galápagos Islands. Aquat. Mammals 39, 196. doi: 10.1578/AM.39.2.2013.196
Andrews R. D., Pitman R. L., Ballance L. T. (2008). Satellite tracking reveals distinct movement patterns for Type B and Type C killer whales in the southern Ross Sea, Antarctica. Polar Biol. 31, 1461–1468. doi: 10.1007/s00300-008-0487-z
Andriolo A., Reis S. S., Amorim T. O., Sucunza F., De Castro F. R., Maia Y. G., et al. (2015). Killer whale (Orcinus orca) whistles from the western South Atlantic Ocean include high frequency signals. J. Acoustical Soc. America 138, 1696–1701. doi: 10.1121/1.4928308
Arnbom T., Papastavrou V., Weilgart L. S., Whitehead H. (1987). Sperm whales react to an attack by killer whales. J. Mammalogy 68, 450–453. doi: 10.2307/1381497
Baird R. W. (2000). “The killer whale: Foraging specializations and group hunting,” in Cetacean societies: field studies of dolphins and whales. Eds. Mann J., Connor R. C., Tyack P. L., Whitehead H. (Chicago, IL: The University of Chicago Press), 127–153.
Baird R. W., Dill L. M. (1996). Ecological and social determinants of group size in transient killer whales. Behav. Ecol 7, 408–416. doi: 10.1093/beheco/7.4.408
Baird R. W., Mcsweeney D. J., Bane C., Barlow J., Salden D. R., Antoine L. R. K., et al. (2006). Killer whales in Hawaiian waters: information on population identity and feeding habits. Pacific Sci 60, 523–530. doi: 10.1353/psc.2006.0024
Barrett-Lennard L. G., Ford J. K., Heise K. A. (1996). The mixed blessing of echolocation: differences in sonar use by fish-eating and mammal-eating killer whales. Anim. Behav 51, 553–565. doi: 10.1006/anbe.1996.0059
Bejder L., Fletcher D., Bräger S. (1998). A method for testing association patterns of social animals. Anim. Behav 56, 719–725. doi: 10.1006/anbe.1998.0802
Best P. B., Photopoulou T. (2016). Identifying the “demon whale-biter”: patterns of scarring on large whales attributed to a cookie-cutter shark. Isistius PloS One sp, 11. doi: 10.1371/journal.pone.0152643
Bigg M. (1982). An assessment of killer whale (Orcinus orca) stocks off Vancouver Island, British Columbia. Rep. Int. Whaling Commission 32, 655–666.
Bigg M., Olesiuk P., Ellis G. M., Ford J., Balcomb K. C. (1990). Social organization and genealogy of resident killer whales (Orcinus orca) in the coastal waters of British Columbia and Washington State. Rep. Int. Whaling Commission 12, 383–405.
Bolaños-Jiménez J., Kiszka J. J., Bouveret L., Ferrer G. R., Ramos E. A., Henriquez A., et al. (2023). The killer whale in the Caribbean Sea: An updated review of Its ecology, exploitation, and interactions with fisheries. Aquat. Mammals 49, 184–194. doi: 10.1578/AM.49.2.2023.184
Bolaños-Jiménez J., Mignucci-Giannoni A. A., Blumenthal J., Bogomolni A., Casas J. J., Henríquez A., et al. (2014). Distribution, feeding habits and morphology of killer whales Orcinus orca in the Caribbean Sea. Mammal Rev 44, 177–189. doi: 10.1111/mam.12021
Burnham K. P., Anderson D. R., Laake J. L. (1980). Estimation of density from line transect sampling of biological populations. Wildlife Monogr 72, 3–202.
Cairns S. J., Schwager S. J. (1987). A comparison of association indices. Anim. Behav 35, 1454–1469. doi: 10.1016/S0003-3472(87)80018-0
Castro-Azofeifa C. (2021). Avistamientos de Orcinus orca (Linnaeu 1758)(Cetartiodactyla: Odontoceti: Delphinidae) en el Pacífico costarricense (1990-2020). J. Mar. Coast. Sci 13, 29–47. doi: 10.15359/revmar.13-2.3
Connor R. C., Wells R. S., Mann J., Read A. J. (2000). “The bottlenose dolphin: Social relationships in a fission-fusion society,” in Cetacean societies: field studies of dolphins and whales. Eds. Mann J., Connor R. C., Tyack P. L., Whitehead H. (Chicago, IL: The University of Chicago Press), 91–125.
Dahlheim M., Leatherwood S., Perrin W. F. (1982). Distribution of killer whales in the warm temperate and tropical eastern Pacific. Rep. Int. Whaling Commission 32, 647–653.
Dahlheim M. E., Schulman-Janiger A., Black N., Ternullo R., Ellifrit D., Balcomb III K. C. (2008). Eastern temperate North Pacific offshore killer whales (Orcinus orca): Occurrence, movements, and insights into feeding ecology. Mar. Mammal Sci 24, 719–729. doi: 10.1111/j.1748-7692.2008.00206.x
Dalla Rosa L., Secchi E. R. (2007). Killer whale (Orcinus orca) interactions with the tuna and swordfish longline fishery off southern and south-eastern Brazil: a comparison with shark interactions. J. Mar. Biol. Assoc. United Kingdom 87, 135–140. doi: 10.1017/S0025315407054306
De Bruyn P. N., Tosh C. A., Terauds A. (2013). Killer whale ecotypes: is there a global model? Biol. Rev 88, 62–80. doi: 10.1111/j.1469-185X.2012.00239.x
Deecke V. B., Ford J. K., Slater P. J. (2005). The vocal behaviour of mammal-eating killer whales: communicating with costly calls. Anim. Behav 69, 395–405. doi: 10.1016/j.anbehav.2004.04.014
Denkinger J., Alarcon D., Espinosa B., Fowler L., Manning C., Oña J., et al. (2020). Social structure of killer whales (Orcinus orca) in a variable low-latitude environment, the Galápagos Archipelago. Mar. Mammal Sci 36, 774–785. doi: 10.1111/mms.12672
Duchêne S., Archer F. I., Vilstrup J., Caballero S., Morin P. A. (2011). Mitogenome phylogenetics: the impact of using single regions and partitioning schemes on topology, substitution rate and divergence time estimation. PloS One 6, e27138. doi: 10.1371/journal.pone.0027138
Dunn C., Claridge D. (2013). Killer whale (Orcinus orca) occurrence and predation in the Bahamas. J. Mar. Biol. Assoc. United Kingdom 94, 1305–1309. doi: 10.1017/S0025315413000908
Durban J., Pitman R. (2012). Antarctic killer whales make rapid, round-trip movements to subtropical waters: evidence for physiological maintenance migrations? Biol. Lett 8, 274–277. doi: 10.1098/rsbl.2011.0875
Dwyer S. L., Visser I. N. (2011). Cookie cutter shark (Isistius sp.) bites on cetaceans, with particular reference to killer whales (Orca)(Orcinus orca). Aquat. Mammals 37, 111–138. doi: 10.1578/AM.37.2.2011.111
Elwen S. H., Leeney R. H. (2011). Interactions between leatherback turtles and killer whales in Namibian waters, including possible predation. Afr. J. Wildlife Res 41, 205–209. doi: 10.3957/056.041.0207
Evans W., Yablokov A., Bowles A. (1982). Geographic variation in the color pattern of killer whales. Rep. Int. Whaling Commission 32, 687–694.
Fertl D., Acevedo-Gutierrez A., Darby F. L. (1996). A report of killer whales (Orcinus orca) feeding on a carcharhinid shark in Costa Rica. Mar. Mammal Sci 12, 606–611. doi: 10.1111/j.1748-7692.1996.tb00075.x
Filatova O. A., Deecke V. B., Ford J. K., Matkin C. O., Barrett-Lennard L. G., Guzeev M. A., et al. (2012). Call diversity in the North Pacific killer whale populations: implications for dialect evolution and population history. Anim. Behav 83, 595–603. doi: 10.1016/j.anbehav.2011.12.013
Foote A. D. (2022). Are “Type 2“ killer whales long in the tooth? A critical reflection on the discrete categorization of Northeast Atlantic killer whales. Mar. Mammal Sci 39, 345–350. doi: 10.1111/mms.12964
Foote A. D., Newton J., Ávila-Arcos M. C., Kampmann M.-L., Samaniego J. A., Post K., et al. (2013). Tracking niche variation over millennial timescales in sympatric killer whale lineages. Proc. R. Soc. B: Biol. Sci 280, 20131481. doi: 10.1098/rspb.2013.1481
Foote A. D., Newton J., Piertney S. B., Willerslev E., Gilbert M. T. P. (2009). Ecological, morphological and genetic divergence of sympatric North Atlantic killer whale populations. Mol. Ecol 18, 5207–5217. doi: 10.1111/j.1365-294X.2009.04407.x
Ford J. K. (2009). “Killer whale - Orcinus orca,” in Encyclopedia of marine mammals, 2nd. Eds. Perrin W. F., Würsig B., Thewissen J. G. M. (Switzerland: Academic Press, Amsterdam), 650–657.
Ford J. K. (2019). “Killer whales: behavior, social organization, and ecology of the oceans’ apex predators,” in Ethology and behavioral ecology of odontocetes. Ed. Würsig B. (Switzerland: Springer), 239–259.
Ford J. K., Ellis G. M., Barrett-Lennard L. G., Morton A. B., Palm R. S., Balcomb III K. C. (1998). Dietary specialization in two sympatric populations of killer whales (Orcinus orca) in coastal British Columbia and adjacent waters. Can. J. Zoology 76, 1456–1471. doi: 10.1139/z98-089
Ford J. K., Ellis G. M., Matkin C. O., Wetklo M. H., Barrett-Lennard L. G., Withler R. E. (2011). Shark predation and tooth wear in a population of northeastern Pacific killer whales. Aquat. Biol 11, 213–224. doi: 10.3354/ab00307
Forney K. A., Wade P. R. (2006). “Worldwide distribution and abundance of killer whales,” in Whales, whaling and ocean ecosystems. Eds. Estes J. A., DeMaster D. P., Doak D. F., Williams T. M., Brownell R. L. Jr (Berkeley, CA: University of California Press), 145–162.
Fulling G. L., Mullin K. D., Hubard C. W. (2003). Abundance and distribution of cetaceans in outer continental shelf waters of the US Gulf of Mexico. Fishery Bull 101, 923–932.
García-Godos I. (2004). Killer whale (Orcinus orca) occurrence off Peru 1995-2003. Latin Am. J. Aquat. Mammals 48, 177–180. doi: 10.5597/lajam00064
Garrison L. P., Martinez A., Maze-Foley K. (2010). Habitat and abundance of cetaceans in Atlantic Ocean continental slope waters off the eastern USA. J. Cetacean Res. Manage 11, 267–277. doi: 10.47536/jcrm.v11i3
Garrison L. P., Stokes L., Fairfield C. (2009). Estimated bycatch of marine mammals and sea turtles in the U.S. Atlantic pelagic longline fleet during 2008. NOAA Technical Memorandum NOAA NMFS-SEFSC-591. NOAA Tech. Mem.
Grace M. A., Dias L. A., Maze-Foley K., Sinclair C., Mullin K. D., Garrison L., et al. (2018). Cookiecutter shark bite wounds on cetaceans of the Gulf of Mexico. Aquat. Mammals. 44, 491–499. doi: 10.1578/AM.44.5.2018.491
Guinet C., Domenici P., De Stephanis R., Barrett-Lennard L., Ford J., Verborgh P. (2007). Killer whale predation on bluefin tuna: exploring the hypothesis of the endurance-exhaustion technique. Mar. Ecol. Prog. Ser 347, 111–119. doi: 10.3354/meps07035
Hairr J. (2012). Killer whales (Orcinus orca) off the north carolina coast 1709–2011. J. North Carolina Acad. Sci 128, 39–43. doi: 10.7572/2167-5880-128.2.39
Hanson M. B., Baird R. W., Ford J. K., Hempelmann-Halos J., Van Doornik D. M., Candy J. R., et al. (2010). Species and stock identification of prey consumed by endangered southern resident killer whales in their summer range. Endangered Species Res 11, 69–82. doi: 10.3354/esr00263
Hayes S. A., Josephson E., Katherine Maze-Foley K., Rosel P. E. (2021). US atlantic and gulf of Mexico marine mammal stock assessments - 2020. NOAA Tech. Memorandum NMFS-NE-271. doi: 10.25923/nbyx-p656
Higdon J. W., Hauser D. D., Ferguson S. H. (2012). Killer whales (Orcinus orca) in the Canadian Arctic: distribution, prey items, group sizes, and seasonality. Mar. Mammal Sci 28, E93–E109. doi: 10.1111/j.1748-7692.2011.00489.x
Jefferson T. A., Stacey P. J., Baird R. W. (1991). A review of killer whale interactions with other marine mammals: predation to co-existence. Mammal Rev 21, 151–180. doi: 10.1111/j.1365-2907.1991.tb00291.x
Jefferson T. A., Webber M. A., Pitman R. L. (2015). Marine mammals of the world: a comprehensive guide to their identification, 2nd edition (Academic Press).
Johnson M., Zaretskaya I., Raytselis Y., Merezhuk Y., McGinnis S., Madden T. L. (2008). NCBI BLAST: a better web interface. Nucleic Acids Research 36(suppl_2), W5–W9.
Jones I. M. (2006). A northeast Pacific offshore killer whale (Orcinus orca) feeding on a Pacific halibut (Hippoglossus stenolepis). Mar. Mammal Sci 22, 198–200. doi: 10.1111/j.1748-7692.2006.00013.x
Jourdain E., Ugarte F., Víkingsson G. A., Samarra F. I., Ferguson S. H., Lawson J., et al. (2019). North Atlantic killer whale Orcinus orca populations: a review of current knowledge and threats to conservation. Mammal Rev 49, 384–400. doi: 10.1111/mam.12168
Kane E. A., Olson P. A., Gerrodette T., Fiedler P. C. (2008). Prevalence of the commensal barnacle Xenobalanus globicipitis on cetacean species in the eastern tropical Pacific Ocean, and a review of global occurrence. Fishery Bull 106, 395–404.
Katona S. K., Beard J. A., Girton P. E., Wenzel F. (1988). Killer whales (Orcinus orca) from the Bay of Fundy to the Equator, including the Gulf of Mexico. Rit Fiskideildar 11, 205–224.
Keating J. L., Barlow J. (2013). Summary of PAMGuard beaked whale click detectors and classifiers used during the 2012 Southern California Behavioral Response Study. NOAA-TM-NMFS-SWFSC-517.
Kiszka J. J., Caputo M., Méndez-Fernandez P., Fielding R. (2021). Feeding ecology of elusive Caribbean killer whales inferred from Bayesian stable isotope mixing models and whalers’ ecological knowledge. Front. Mar. Sci 8, 648421. doi: 10.3389/fmars.2021.648421
Knapp J. M., Aranda G., Medina A., Lutcavage M. (2014). Comparative assessment of the reproductive status of female Atlantic bluefin tuna from the Gulf of Mexico and the Mediterranean Sea. PloS One 9, e98233. doi: 10.1371/journal.pone.0098233
Kocher T. D., Thomas W. K., Meyer A., Edwards S. V., Pääbo S., Villablanca F. X., et al. (1989). Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci 86, 6196–6200. doi: 10.1073/pnas.86.16.6196
Krahn M. M., Herman D. P., Matkin C. O., Durban J. W., Barrett-Lennard L., Burrows D. G., et al. (2007). Use of chemical tracers in assessing the diet and foraging regions of eastern North Pacific killer whales. Mar. Environ. Res 63, 91–114. doi: 10.1016/j.marenvres.2006.07.002
Lefort K. J., Matthews C. J., Higdon J. W., Petersen S. D., Westdal K. H., Garroway C. J., et al. (2020). A review of Canadian Arctic killer whale (Orcinus orca) ecology. Can. J. Zoology 98, 245–253. doi: 10.1139/cjz-2019-0207
Madsen P. T., Carder D., Bedholm K., Ridgway S. (2005). Porpoise clicks from a sperm whale nose—Convergent evolution of 130 kHz pulses in toothed whale sonars? Bioacoustics 15, 195–206. doi: 10.1080/09524622.2005.9753547
Mcalpine D. F. (2009). “Pygmy and Dwarf Sperm Whales - Kogia breviceps and K. sima,” in eds. Encyclopedia of marine mammals, 2nd edn. Ed. Perrin Würsig Thewissen W. F. B.J. G. M. (Academic Press, Amsterdam), 936–938.
Mcsweeney D. J., Baird R. W., Mahaffy S. D. (2007). Site fidelity, associations, and movements of Cuvier's (Ziphius cavirostris) and Blainville's (Mesoplodon densirostris) beaked whales off the island of Hawai ‘i. Mar. Mammal Sci 23, 666–687. doi: 10.1111/j.1748-7692.2007.00135.x
Morin P. A., Archer F. I., Foote A. D., Vilstrup J., Allen E. E., Wade P., et al. (2010). Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species. Genome Res 20, 908–916. doi: 10.1101/gr.102954.109
Morin P. A., Mccarthy M. L., Fung C. W., Durban J. W., Parsons K. M., Perrin W. F., et al. (2024). Revised taxonomy of eastern North Pacific killer whales (Orcinus orca): Bigg’s and resident ecotypes deserve species status. R. Soc. Open Sci 11, 231368. doi: 10.1098/rsos.231368
Morin P. A., Parsons K. M., Archer F. I., Ávila-Arcos M. C., Barrett-Lennard L. G., Dalla Rosa L., et al. (2015). Geographic and temporal dynamics of a global radiation and diversification in the killer whale. Mol. Ecol 24, 3964–3979. doi: 10.1111/mec.13284
Moura A., Janse Van Rensburg C., Pilot M., Tehrani A., Best P., Thornton M., et al. (2014). Killer whale nuclear genome and mtDNA reveal widespread population bottleneck during the last glacial maximum. Mol. Biol. Evol 32, 1121–1131. doi: 10.1093/molbev/msu058
Mullin K. D., Fulling G. L. (2004). Abundance of cetaceans in the oceanic northern Gulf of Mexico 1996–2001. Mar. Mammal Sci 20, 787–807. doi: 10.1111/j.1748-7692.2004.tb01193.x
Mullin K. D., Hoggard W., Hansen L. J. (2004). Abundance and seasonal occurrence of cetaceans in outer continental shelf and slope waters of the north-central and northwestern Gulf of Mexico. Gulf Mexico Sci 22, 6. doi: 10.18785/goms.2201.06
NMFS (2016). 2016 annual report of a comprehensive assessment of marine mammal, marine turtle, and seabird abundance and spatial distribution in US waters of the western North Atlantic Ocean – AMAPPS II. doi: 10.25923/gbap-g480
O'Sullivan S., Mullin K. D. (1997). Killer whales (Orcinus orca) in the northern Gulf of Mexico. Mar. Mammal Sci 13, 141–147. doi: 10.1111/j.1748-7692.1997.tb00618.x
Olesiuk P. F., Ellis G. M., Ford J. K. (2005). Life history and population dynamics of northern resident killer whales (Orcinus orca) in British Columbia (ON, Canada: Canadian Science Advisory Secretariat Ottawa).
Olsen D. W., Matkin C. O., Mueter F. J., Atkinson S. (2020). Social behavior increases in multipod aggregations of southern Alaska resident killer whales (Orcinus orca). Mar. Mammal Sci 36, 1150–1159. doi: 10.1111/mms.12715
Olson P. A., Gerrodette T. (2008). Killer whales of the eastern tropical Pacific: a catalog of photo-identified individuals. NOAA Tech. Memorandum NMFS-SWFSC-428.
Olson P. A., Hancock-Hanser B. L., McCullough J. L., Dahlheim M., Schulman-Janiger A., Black N., et al. (2023). Killer whale predation on a pygmy sperm whale in an oceanic habitat. Mar. Mammal Sci 39, 281–288. doi: 10.1111/mms.12956
Pérez-Zayas J. J., Mignucci-Giannoni A. A., Toyos-González G. M., Rosario-Delestre R. J., Williams E. (2002). Incidental predation by a largetooth cookiecutter shark on a Cuvier's beaked whale in Puerto Rico. Aquat. Mammals 28, 308–311.
Pitman R. (2011). Antarctic killer whales: top of the food chain at the bottom of the world. J. Am. Cetacean Soc 40, 39–45.
Pitman R. L., Ballance L. T., Mesnick S. I., Chivers S. J. (2001). Killer whale predation on sperm whales: observations and implications. Mar. Mammal Sci 17, 494–507. doi: 10.1111/j.1748-7692.2001.tb01000.x
Pitman R. L., Durban J. W., Greenfelder M., Guinet C., Jorgensen M., Olson P. A., et al. (2011). Observations of a distinctive morphotype of killer whale (Orcinus orca), type D, from subantarctic waters. Polar Biol 34, 303–306. doi: 10.1007/s00300-010-0871-3
Pitman R. L., Dutton P. H. (2004). Killer whale predation on a leatherback turtle in the northeast Pacific. Pacific Sci 58, 497–498. doi: 10.1353/psc.2004.0034
Pitman R. L., Ensor P. (2003). Three forms of killer whales (Orcinus orca) in Antarctic waters. J. Cetacean Res. Manage 5, 131–140. doi: 10.47536/jcrm.v5i2
Pitman R. L., Osullivan S., Mase B. (2003). Killer whales (Orcinus orca) attack a school of pantropical spotted dolphins (Stenella attenuata) in the Gulf of Mexico. Aquat. Mammals 29, 321–324. doi: 10.1578/01675420360736488
Pitman R. L., Perryman W. L., Leroi D., Eilers E. (2007). A dwarf form of killer whale in Antarctica. J. Mammalogy 88, 43–48. doi: 10.1644/06-MAMM-A-118R1.1
Pyle P., Schramm M. J., Keiper C., Anderson S. D. (1999). Predation on a white shark (Carcharodon carcharias) by a killer whale (Orcinus orca) and a possible case of competitive displacement. Mar. Mammal Sci 15, 563–568. doi: 10.1111/j.1748-7692.1999.tb00822.x
Ramírez-León M. R., García-Aguilar M. C., Aguayo-Lobo A., Fuentes-Allen I., Sosa-Nishizaki O. (2020). What do we know about cetaceans in the Mexican waters of the Gulf of Mexico? A review. Aquat. Mammals 46, 623–632. doi: 10.1578/AM.46.6.2020.623
Remili A., Dietz R., Sonne C., Samarra F. I., Rikardsen A. H., Kettemer L. E., et al. (2023). Quantitative fatty acid signature analysis reveals a high level of dietary specialization in killer whales across the North Atlantic. J. Anim. Ecol 92, 1–14. doi: 10.1111/1365-2656.13920
Reyes L. M., García-Borboroglu P. (2004). Killer whale (Orcinus orca) predation on sharks in Patagonia, Argentina: a first report. Aquat. Mammals 30, 376–379. doi: 10.1578/AM.30.3.2004.376
Rosel P. E. (2003). PCR-based sex determination in Odontocete cetaceans. Conserv. Genet 4, 647–649. doi: 10.1023/A:1025666212967
Rosel P., Block B. (1996). Mitochondrial control region variability and global population structure in the swordfish, Xiphias gladius. Mar. Biol 125, 11–22. doi: 10.1007/BF00350756
Rosel P., France S., Wang J., Kocher T. (1999). Genetic structure of harbour porpoise Phocoena phocoena populations in the northwest Atlantic based on mitochondrial and nuclear markers. Mol. Ecol 8, S41–S54. doi: 10.1046/j.1365-294X.1999.00758.x
Ryan C., Holmes J. (2012). Killer whale Orcinus orca predation on sunfish Mola mola. Mar. Biodiversity Records 5, e10. doi: 10.1017/S1755267211001187
Samarra F. I. P., Deecke V. B., Simonis A. E., Miller P. J. O. (2015). Geographic variation in the time-frequency characteristics of high-frequency whistles produced by killer whales (Orcinus orca). Mar. Mammal Sci 31, 688–706. doi: 10.1111/mms.12195
Samarra F. I., Deecke V. B., Vinding K., Rasmussen M. H., Swift R. J., Miller P. J. (2010). Killer whales (Orcinus orca) produce ultrasonic whistles. J. Acoustical Soc. America 128, EL205–EL210. doi: 10.1121/1.3462235
Saulitis E., Matkin C., Barrett-Lennard L., Heise K., Ellis G. (2000). Foraging strategies of sympatric killer whale (Orcinus orca) populations in Prince William Sound, Alaska. Mar. mammal Sci 16, 94–109. doi: 10.1111/j.1748-7692.2000.tb00906.x
Schmidly D. (1981). Marine mammals of the southeastern United States coast and the Gulf of Mexico (Washington, DC: US Fish and Wildl. Serv., Office Biol. Serv.). FWS/OBS-80/41: 1-163.
SEFSC (2001). Cruise results – NOAA Ship Gordon Gunter Cruise GU-01-04(013) 17 July – 22 August 2001: a study of sperm whales in the north-central Gulf of Mexico. Southeast Fisheries Science Center Reference Document MMTD-2024-10. doi: 10.25923/8n3f-nn77.
Simonis A. E., Baumann-Pickering S., Oleson E., Melcon M. L., Gassmann M., Wiggins S. M., et al. (2012). High-frequency modulated signals of killer whales (Orcinus orca) in the North Pacific. J. Acoustical Soc. America 131, EL295–EL301. doi: 10.1121/1.3690963
Sorisio Sonnino L., De Maddalena A., Visser I. N. (2006). Interaction between killer whales (Orcinus orca) and hammerhead sharks (Sphyrna sp.) in Galápagos waters. Latin Am. J. Aquat. Mammals 5, 69–71. doi: 10.5597/lajam00095
Tavares S. B., Samarra F. I., Miller P. J. (2017). A multilevel society of herring-eating killer whales indicates adaptation to prey characteristics. Behav. Ecol 28, 500–514. doi: 10.1093/beheco/arw179
Teo S. L., Block B. A. (2010). Comparative influence of ocean conditions on yellowfin and Atlantic bluefin tuna catch from longlines in the Gulf of Mexico. PloS One 5, e10756. doi: 10.1371/journal.pone.0010756
Terrapon M., Kiszka J. J., Wagner J. (2021). Observations of killer whale (Orcinus orca) feeding behavior in the tropical waters of the northern Mozambique Channel Island of Mayotte, Southwest Indian Ocean. Aquat. Mammals 47, 196–205. doi: 10.1578/AM.47.2.2021.196
Testino J. P., Petit A., Alcorta B., Pacheco A. S., Silva S., Alfaro-Shigueto J., et al. (2019). Killer whale (Orcinus orca) occurrence and interactions with marine mammals off Peru. Pacific Sci 73, 261–273. doi: 10.2984/73.2.7
Tolley K. A., Rosel P. E. (2006). Population structure and historical demography of eastern North Atlantic harbour porpoises inferred through mtDNA sequences. Mar. Ecol. Prog. Ser 327, 297–308. doi: 10.3354/meps327297
Trickey J. S., Reyes M. V. R., Baumann-Pickering S., Melcón M. L., Hildebrand J. A., Iñíguez M. A. (2014). Acoustic encounters of killer and beaked whales during the 2014 SORP cruise. IWC SC/65b/SM12re.
Vargas-Bravo M. H., Elorriaga-Verplancken F. R., Olivos-Ortiz A., Morales-Guerrero B., Liñán-Cabello M. A., Ortega-Ortiz C. D. (2020). Ecological aspects of killer whales from the Mexican Central Pacific coast: Revealing a new ecotype in the Eastern Tropical Pacific. Mar. Mammal Sci 2020, 1–16. doi: 10.1111/mms.12748
Visser I. N. (2005). First observations of feeding on thresher (Alopias vulpinus) and hammerhead (Sphyrna zygaena) sharks by killer whales (Orcinus orca) specialising on elasmobranch prey. Aquat. Mammals 31, 83–88. doi: 10.1578/AM.31.1.2005.83
Visser I. N., Bonoccorso F. J. (2003). New observations and a review of killer whale (Orcinus orca) sightings in Papua New Guinea waters. Aquat. Mammals 29, 150–172. doi: 10.1578/016754203101024004
Visser I. N., Mäkeläinen P. (2000). Variation in eye-patch shape of killer whales (Orcinus orca) in New Zealand waters. Mar. Mammal Sci 16, 459–469. doi: 10.1111/j.1748-7692.2000.tb00938.x
Wade P. R., Gerrodette T. (1993). Estimates of cetacean abundance and distribution in the eastern tropical Pacific. Rep. Int. Whaling Commission 43, 477–493.
Waring G. T., Josephson E., Maze-Foley K., Rosel P. E. (2013). U.S. Atlantic and Gulf of Mexico marine mammal stock assessments – 2012. NOAA Tech. Memorandum NMFS-NE-223.
Weir C. R., Collins T., Carvalho I., Rosenbaum H. C. (2010). Killer whales (Orcinus orca) in Angolan and Gulf of Guinea waters, tropical West Africa. J. Mar. Biol. Assoc. United Kingdom 90, 1601–1611. doi: 10.1017/S002531541000072X
Wenzel F. W., López Suárez P. (2012). What is known about cookiecutter shark (Isistius spp.) interactions with cetaceans in Cape Verde seas. Zoologia Caboverdiana 3, 57–66.
Whitehead H. (1995). Investigating structure and temporal scale in social organizations using identified individuals. Behav. Ecol 6, 199–208. doi: 10.1093/beheco/6.2.199
Whitehead H. (2007). Selection of models of lagged identification rates and lagged association rates using AIC and QAIC. Commun. Statistics—Simulation Computation® 36, 1233–1246. doi: 10.1080/03610910701569531
Whitehead H. (2008). Analyzing animal societies: quantitative methods for vertebrate social analysis (Chicago, IL: University of Chicago Press).
Whitehead H. (2009). SOCPROG programs: analysing animal social structures. Behav. Ecol. Sociobiology 63, 765–778. doi: 10.1007/s00265-008-0697-y
Whitehead H., Bejder L., Ottensmeyer C. A. (2005). Testing association patterns: issues arising and extensions. Anim. Behav 69, e1. doi: 10.1016/j.anbehav.2004.11.004
Whitehead T. O., Rollinson D. P., Reisinger R. R. (2015). Pseudostalked barnacles Xenobalanus globicipitis attached to killer whales Orcinus orca in South African waters. Mar. Biodiversity 45, 873–876. doi: 10.1007/s12526-014-0296-2
Whitt A. D., Baran M. A., Bryson M., Rendell L. E. (2015). First report of killer whales harassing sperm whales in the Gulf of Mexico. Aquat. Mammals 41, 252–255. doi: 10.1578/AM.41.3.2015.252
Williams R., Gero S., Bejder L., Calambokidis J., Kraus S. D., Lusseau D., et al. (2011). Underestimating the damage: interpreting cetacean carcass recoveries in the context of the Deepwater Horizon/BP incident. Conserv. Lett 4, 228–233. doi: 10.1111/conl.2011.4.issue-3
Keywords: acoustics, genetics, Gulf of Mexico, killer whale, morphology, North Atlantic Ocean, social structure
Citation: Barry KP, Mullin KD, Maze-Foley K, Wilcox Talbot LA, Rosel PE, Soldevilla MS, Dias LA, Ramírez-León MR and Litz JA (2024) Killer whales in the Gulf of Mexico and North Atlantic off the Southeastern United States. Front. Mar. Sci. 11:1460314. doi: 10.3389/fmars.2024.1460314
Received: 05 July 2024; Accepted: 03 October 2024;
Published: 12 November 2024.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Ann Pabst, University of North Carolina Wilmington, United StatesOlga Filatova, University of Southern Denmark, Denmark
Copyright © 2024 Barry, Mullin, Maze-Foley, Wilcox Talbot, Rosel, Soldevilla, Dias, Ramírez-León and Litz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Keith D. Mullin, S2VpdGguRC5NdWxsaW5Abm9hYS5nb3Y=
†Deceased