 Ana M. García-Cegarra
Ana M. García-Cegarra Javier Oña
Javier Oña Camilo Arancibia2
Camilo Arancibia2- 1Instituto de Ciencias Naturales Alexander von Humboldt, Facultad de Ciencias del Mar y Recursos Biológicos, Universidad de Antofagasta, Antofagasta, Chile
- 2Laboratorio de Estudio de Megafauna Marina, CETALAB, Antofagasta, Chile
- 3Programa Doctorado en Ciencias Aplicadas, Facultad de Ciencias del Mar y Recursos Biológicos, Universidad de Antofagasta, Antofagasta, Chile
- 4Escuela de Medicina Veterinaria, Facultad de Recursos Naturales y Medicina Veterinaria, Universidad Santo Tomás, Viña del Mar, Chile
- 5ONG Panthalassa, Red de Estudios de Vertebrados Marinos en Chile, Santiago, Chile
Orcas (Orcinus orca) are the top marine predators of the ocean, targeting multiple taxa including teleost fishes, elasmobranchs, seabirds, sea turtles, pinnipeds, odontocetes and other large cetaceans. According to their foraging strategies, genetics, acoustics, and morphology, orcas differentiate into ecotypes. Despite their cosmopolitan distribution, orcas are poorly studied in areas such as the Humboldt Current System of the Southeast Pacific coast. Since 2018 in northern Chile, presumed Type A orcas have been observed foraging on South American sea lions (Otaria byronia),benefiting from the presence of fishing vessels facilitating their hunt. These orcas were observed also pursuing large mixed groups of dusky (Lagenorhynchus obscurus) and long-beaked common dolphins (Delphinus capensis); however, predation on dusky dolphins was not documented. Here, we provide 28 additional sightings of orcas in two upwelling hotspots along northern Chile (15 around the Chañaral, Choros, and Damas Islandsin the Humboldt Penguin National Reserve from 2010–2023 and 13 in the Mejillones Peninsula from 2022–2023) and the first records of orcas effective hunting on dusky dolphins in this area. We present three new observations of orca tooth rake marks on fin whales (Balaenoptera physalus) dorsal fins in northern Chile. Furthermore, we observed the same orcas on two occasions foraging on dusky dolphins and sharing dolphin meat among group members. Overall, these observations suggest that orcas in northern Chile forage on marine mammals, especially on dusky dolphins, and the number of such predation events seems to be increasing near the coast.
1 Introduction
With no natural predators, orcas (Orcinus orca) are top ocean predators that forage on a wide variety of organisms such as fishes, elasmobranchs, sea turtles, pinnipeds and even other cetaceans, such as dolphins and whales (Nottestad et al., 2002; Ford et al., 2005; Álava and Merlen, 2009; Álava et al., 2013; Ford, 2018; Totterdell et al., 2022; Higuera-Rivas et al., 2023; Towner et al., 2024). Orcas are cosmopolitan and found in ocean basins throughout the globe,yet they remain data deficient according to the IUCN Red List of Threatened Species (Reeves et al., 2017). Depending on their morphology, genetics, acoustics, and foraging strategies, orcas differentiate into ecotypes (Pitman and Ensor, 2003; LeDuc et al., 2008; Morin et al., 2015). Currently, five ecotypes are found in the northern hemisphere (i.e., residents, transient, offshore, type 1 eastern north Atlantic, and type 2 eastern north Atlantic) and five in the southern hemisphere, mainly around the Antarctic Peninsula (i.e., type A, type B1, type B2, type C, and type D) (Pitman and Ensor, 2003; Van Waerebeek et al., 2010; Pitman et al., 2011; Durban et al., 2016). Certain ecotypes specialize in marine mammals, such as Type 2 in the north Atlantic foraging on minke whales (Balaenoptera acutorostrata) (Foote et al., 2009), whereas others have a mixed diet of marine mammals and fishes, such as the northeast Atlantic Type 1 orcas which are considered mainly fish-eaters although some seals and small cetaceans compose a portion of their diet. Orcas consume herring (Clupea harengus) and some seals in Norway and the Faeroe Islands, herring and harbor porpoises (Phocena phocoena) in Iceland, baleen whales and porpoises in eastern Canada, and narwhals (Monodon monoceros) and belugas (Delphinapterus leucas) in the Arctic (Remili et al., 2023). Transient orcas in the Gulf of Alaska, Washington, and British Columbia are also known to prey on cetaceans such as harbor porpoises, Dall’s porpoises (Phocoenoides dalli), and gray whales (Eschrichtius robustus) (Ford et al., 2013; Houghton et al., 2015; McInnes et al., 2020).
In the southern hemisphere, scientific efforts focus mainly on Antarctic and subantarctic waters and foraging strategies as described in the Type A ecotype (i.e., specialty on Antarctic minke whales, Balaenoptera bonaerensis, and southern elephant seals, Mirounga leonina) (Visser et al., 2008). Type B1 orcas forage on Weddell seals (Leptonychotesw eddellii), while the Type B2 forage on penguins and fish (Pitman and Durban, 2010). Type C forages on Antarctic toothfish (Dissostichus mawsoni) (Pitman and Ensor, 2003; Tixier et al., 2019) and type D on fish from longlines (Bruyn et al., 2012; Tixier et al., 2016). However, foraging strategies are not clear as they are free ranging animals, moving long distances and taking prey swiftly, all of which makes in situ observations challenging. Stomach content analyses of stranded individuals is often the primary source of information regarding foraging ecology (Santos et al., 2005) along with few direct observations of hunting (Higuera-Rivas et al., 2023; Ayres et al., 2024; Towner et al., 2024), or other indirect approximations such as stable isotope analyses or fatty acids analyses of free-living or stranded individuals (Loizaga et al., 2023; Remili et al., 2023). The latter two methods have contributed to our understanding of the foraging strategies of orcas Types 1 and 2 in the northeast and northwest Atlantic ocean (Remili et al., 2023) as well as Type A in Argentina. For this Type A, Loizaga et al. (2023) identified three potentially distinct groups related with three ecosystems—Patagonia, Subtropical, and Antarctica—suggesting that different isotopic groups exploit different habitats on the Patagonia shelf.
Orcas belonging to each ecotype have been observed sharing their hunting strategies with conspecifics and may transmit this behavior to future generations through social learning (Whitehead, 2020). When learning, calves are accompanied by an adult female, usually the grandmother, who presumably shows the calf how to engage in different hunting strategies, such as cooperative hunting behavior when knocking seals from ice floes or beach stranding for pinnipeds (e.g., Hoelzel, 1991; Pitman and Ensor, 2003; Pitman and Durban, 2012; Black et al., 2023). Beach strand foraging is well reported for Type A orcas in Peninsula Valdés, Argentina, where matriarchs strand themselves on sandy shores accompanied by a calf to hunt South American sea lions and elephant seals (López and López, 1985; Hoelzel, 1991; Iñiguez, 2001). In other studies, food sharing between two or more individuals has been described to occur in North Pacific resident orcas feeding on salmon, Type A orcas in Argentina feeding on both sea lions and dusky dolphins (Lagenorhynchus obscurus) (Hoelzel, 1991; Coscarella et al., 2015),orcas in New Zealand feeding on stingrays (Visser, 1999), orcas in the Gulf of California sharing bulk sharks (Carcharhinus leucas) (Ayres et al., 2024) and different species of rays (Higuera-Rivas et al., 2023). In these situations, the prey has been described as broken apart prior sharing with juveniles or potentially disabled group members that are unable to hunt (Ford and Ellis, 2006; Coscarella et al., 2015).
Little is known regarding the cooperative foraging and prey sharing strategies of orcas in the southeastern (SE) Pacific Ocean. Orcas have been observed, primarily opportunistically, along the Chilean coastline from Arica city in the north to Cabo de Hornos in the south (Capella et al., 1999; Häussermann et al., 2013; Capella et al., 2014; García-Cegarra, 2022; Haro et al., 2023). During the last decade, orcas sightings have become more frequent in the Magellanes Strait; Capella et al. (2014) observed 55 different individuals foraging on South American sea lions, South American fur seals (Arctocephalu saustralis), kelp gulls (Larus dominicanus), Magellanic diving petrels (Pelecanoides magellani), and unidentified fishes with low site fidelity. García-Cegarra (2022) provided evidence of presumed Type A orcas predating on large groups of dusky and common dolphins in the Humboldt Current System of northern Chile. However, the authors did not observe successful hunts on dolphins; that is, they documented the pursuit of dolphins by five orcas comprised of two males, one female, one juvenile, and one calf, named the Menacho group (García-Cegarra, 2022). The authors observed this pod multiple times along the Humboldt Current System of northern Chile foraging on South American sea lions, likely using the purse-seine vessels present to corner the sea lions and possibly displaying this strategy to their calf (García-Cegarra, 2022). To date, there are no sightings of orcas hunting dolphins or published descriptions of their forage behavior in the SE Pacific Ocean. Here, we describe new orca sightings along northern Chile, provide the first evidence of successful hunts on dusky dolphins by known individuals, and describe three new sightings of orca tooth rake marks on fin whale dorsal fins.
2 Methods
2.1 Study area
The Mejillones Peninsula is in the Antofagasta Region of northern Chile (23°5’57.65’’S: 70°27’043.31’’W) (Figure 1A). This region is influenced by the Humboldt Current System; upwelling cells are constant throughout the year, but more intense during austral autumn and spring (Marin et al., 2003). Upwelling favors the presence of the Humboldt Current marine trophic chain primary and secondary productivity, leading to the presence of keystone species such as krill (Euphausia mucronata) and anchovy (Engraulis ringens) that are the main prey of many cetaceans such as dusky dolphins, long-beak common dolphins, fin whales, and humpback whales (Megaptera novaeangliae) as well as pinnipeds like South American sea lions and fur seals (García-Cegarra et al., 2021; Andrade et al., 2023). Around 800 km south of the Mejillones Peninsula is the second study site group: the Chañaral, Choros, and Damas Islands in the Humboldt Penguin National Reserve, hereafter HPNR (29°2’6.69’’S; 71°34’43.96’’W) (Figure 1B). Here, the area is influenced also by the Humboldt Current System and upwelling cells with high marine diversity (Thiel et al., 2007).
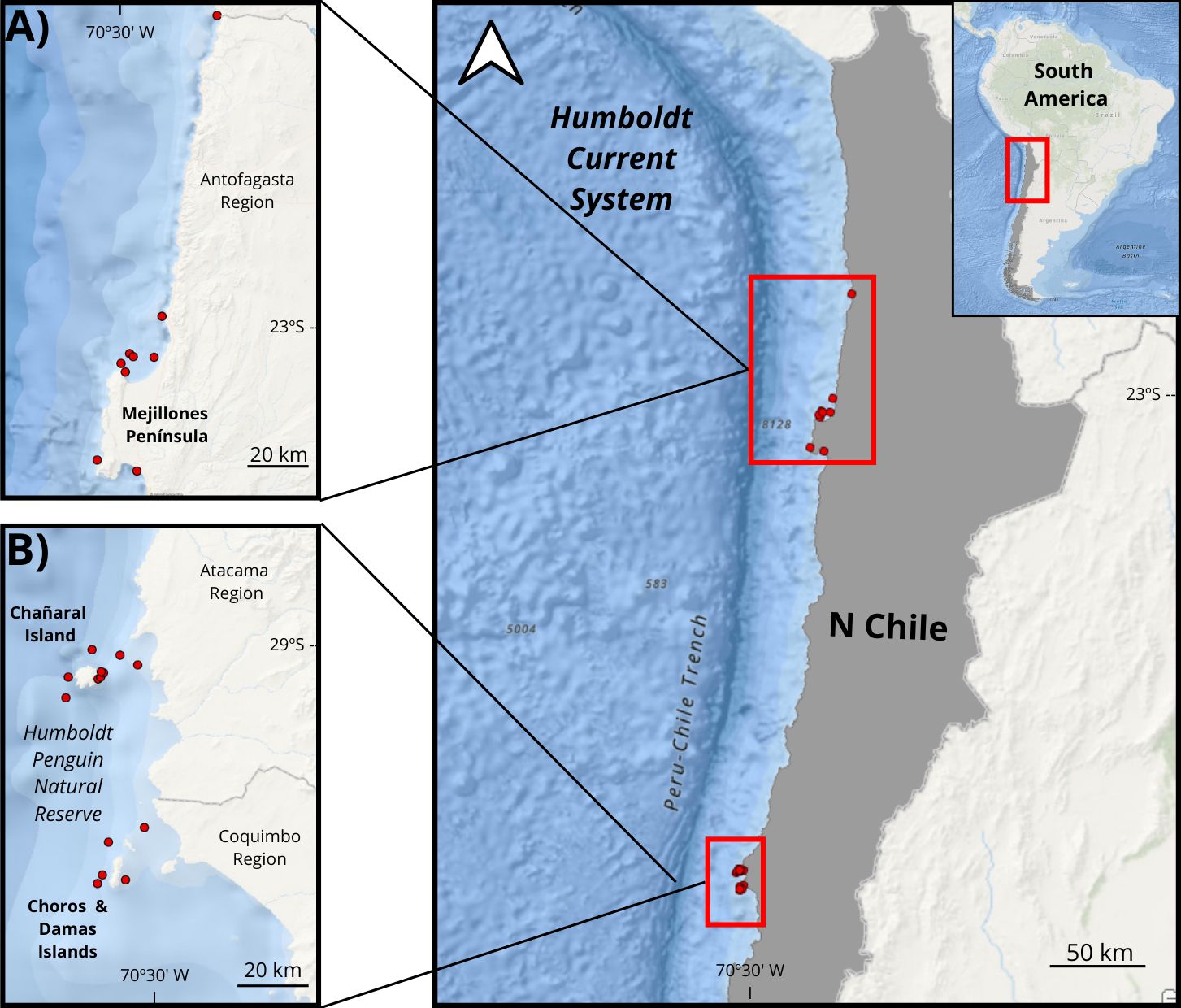
Figure 1. Study area map of northern Chile indicating the two locations of orcas sightings: (A) Mejillones Peninsula in Antofagasta region, (B) Chañaral Island in Atacama Region and Choros&Damas Islands in Coquimbo region, both belonging to the Humboldt Penguin National Reserve.
2.2 Data collection
We collected orca sightings via citizen science and systematic boat surveys.
2.2.1 Citizen science
At both study sites, local fishers or recreational whale watching tours reported sightings via social media. At the Chañaral, Choros, and Damas Islands, whale-watching boats were 11m long, each equipped with a 100HP engine. Tours were from 8:00 to 17:00 h with each trip lasting 2–3 h throughout January and February from 2010 to 2021. Two observers onboard recorded the orcas’ geographical position with GPS and logged group composition and behavior on a datasheet. Once off the boat, researchers received the sighting information for formal documentation. Observers collected pictures of orca dorsal fins and the white eye patch as well as mobile phone videosof orca behavior when observed and possible.
2.2.2 Systematic boat surveys
Along the Mejillones Bay, we conducted systematic line transect boat surveys to observe cetaceans following a line transect starting from Hornitos Port to Punta Angamos covering a total distance of 8 nautic miles. The boat was 5 m long with a 40 HP motor. Surveys took place from 8:30 to 14:30 h with two observers equipped with binoculars Nikon 10x50 who scanned the sea surface until cetacean fins or blows were observed. When orcas were observed, a researcher launched and retrieved an unmanned aerial vehicle (drone) DJI Phantom 4 Pro (DJI Innovation, Shenzhen, China) equipped with a gimbaled camera (1” CMOS, 20M effective pixels) at the stern or bow of the research vessel. The drone was fitted with a flight controller and equipped with an Android system tablet with a 5.5-inch screen. The four smart batteries (LiPo4S, 15.2 V, 5870 mAh) allowed a maximum flight time of 25 min each for a total of 100 min. We calibrated the drone’s gyro sensors on land prior to data collection. When hovering above orcas, the drone recorded videos to register the animals with the camera facing directly down (90°). We also obtained pictures of dorsal fins for photo identification using distinctive features such as scars and nicks on dorsal fins and white eye patches using a Reflex camera Nikon 6D with a 18–200 mm lens (Bigg et al., 1987). We logged the orcas’ geographic positions with a GPS and noted group composition and behavior on a datasheet.
We obtained fin whale photoidentification catalogues for both the Mejillones Peninsula and the Chañaral, Choros, and Damas Islands from 2010–2023. During boat surveys, we collected fin whale dorsal fin photos for photoidentification with a Canon Rebel T5i, Canon 9D, or Nikon D5200 with a 70–300 mm lens. We analyzed and catalogued dorsal fin images for both sites following the criteria of Guevara et al. (2024). We also defined orca tooth rake marks as a set of three or more parallel linear scars approximately equidistant from each other on a whale’s body (Mehta et al., 2007; García-Cegarra, 2022; Minton et al., 2022).
2.3 Data analysis
We analyzed the drone images using Behavioral Observation Research Interactive Software (BORIS, v. 8.22.6; Fiard and Gamba, 2016). We analyzed videos to identify orca group composition and behavior. We defined group composition as adult female(s) and male(s), calf with mother, and juveniles according to size relative to the boat and the height and shape of their dorsal fins. We defined behavior categories as normal swimming, cohesion of the group (individuals at a distance of up 20 m from each other), cohesion between potential mother and calf (individuals at a distance of up 10 m from each other), and foraging behavior (individuals with the entire prey or pieces of prey in their mouth). We constructed an orca distribution map using ArcGIS pro v.2 (ESRI, 2022) by mapping individual positions using the layer of coastal northern Chile. We also performed photo identification of individuals in Image J (v.1.8.0). We identified orcas by dorsal fin morphology and the presence of nicks and scars in the dorsal fins trailing edge according to Black et al. (1997), which allowed us to confirm group composition.
3 Results
3.1 Sightings in the Mejillones Peninsula
In the Mejillones Peninsula, we documented 13 new orcas sightings from 2022–2023 (10 from citizen science, 3 from boat surveys) (Table 1). These sightings are updated from García-Cegarra (2022). Group size ranged from 1–6 individuals (average group size 3.4 individuals). We observed single adult male orcas once and two adult males on five occasions. The remaining groups of 5–6 individuals consisted of adult males, adult females, juveniles, and calves. Calves were present during six sightings, and in one sighting, the calf was a newborn as determined by the yellow eye patch and slow swimming near the presumed adult mother. Between July 24, 2022 and August 2, 2023, a new orca pod was observed in Mejillones Bay. We observed lone adult males on two occasions, while the rest of the sightings presented what we believe was the entire pod. This group was not the Menacho group previously described in the area by García-Cegarra (2022) as individual photoidentification did not match the adults males of the Menacho group (García-Cegarra, 2022). However, during May 14–27, 2023 Menacho pod was observed in Mejillones Bay (Figure 2). The group consisted of one adult male (named Aguilar), one adult female (presumably the matriarch Dakota), one adult female (presumably the calf’s mother), one presumed male juvenile, and one calf. These IDs corresponded with the Menacho pod identification; however, the Menacho adult male (Menacho) was not present during these sightings.
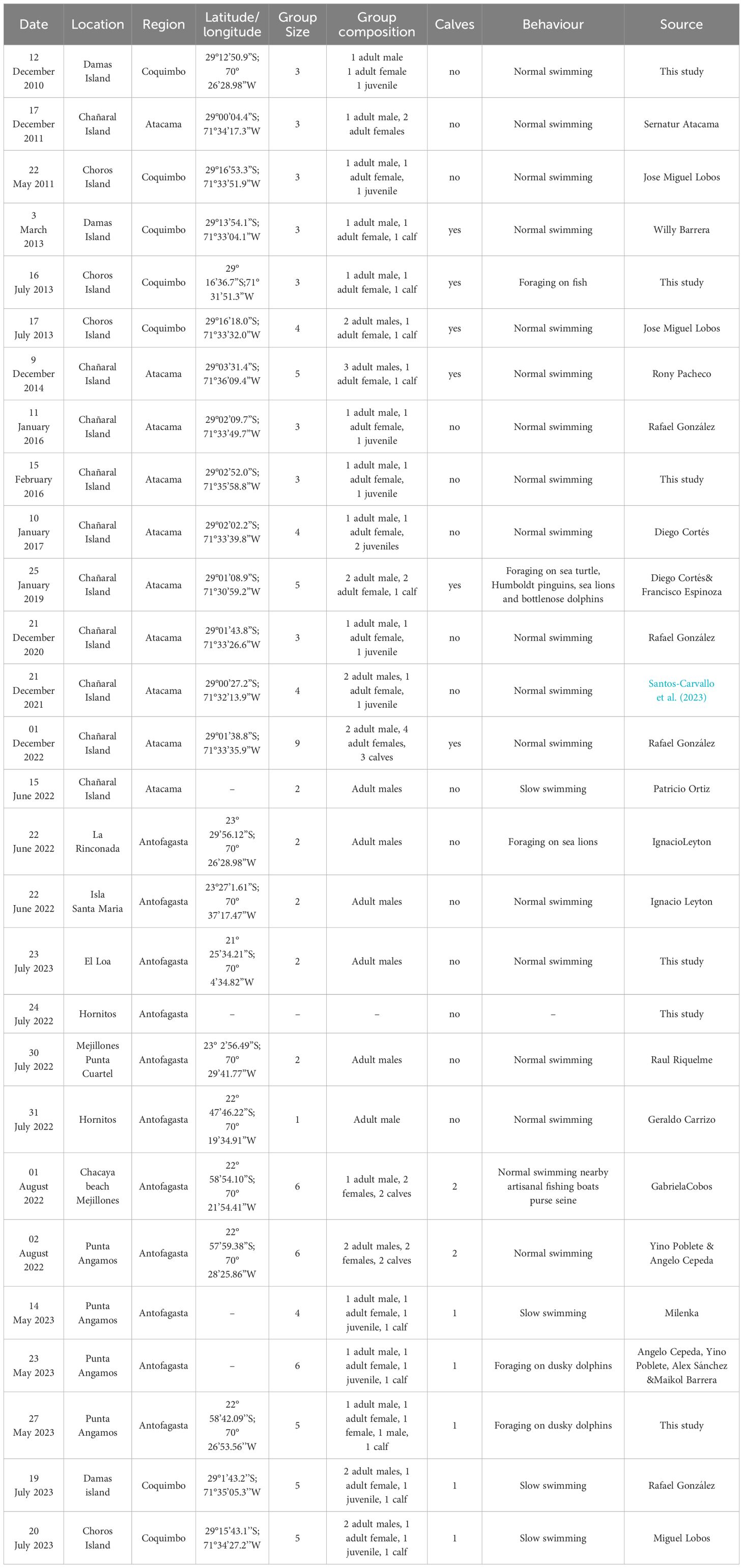
Table 1. Orcas sightingsalong northern Chile.

Figure 2. The “Menacho” pod of orcas hunting a dusky dolphin the 27 May 2023 in Mejillones Bay. (A) The adult male named as “Aguilar”; (B) the adult female presumably the matriarch and grandmother named as “Dakota”; (C) the female of the group presumably the mother of E; (D) a juvenile male; (E) the calf of the pod, presumably son of C; (F) the juvenile male D showing a portion of the hunted dusky dolphins to human observers. White arrows indicate nicks in dorsal fins of individuals. Photo credits: Camilo Arancibia (C, D, F) and Luis Aguilar (A, B, E).
3.2 Sightings in Humboldt Penguin Natural Reserve
Around HPNR, we collected15 orca sightings from 2011–2023 (12 from citizen science, 3 from boat surveys) (Table 1). We did not observe a pattern of regular presence, so we considered sightings occasional in the study area. Group size ranged from 1–5 individuals (average group size 3.9 individuals), and calves were present in three sightings. During three sightings, we observed the orcas foraging. On July 16, 2013, we saw the group hunting a group of unidentified fish. For HPNR, we have photographs of seven sightings from January 11, 2016, wherein we identified the adult male Aguilar but not any of the others. On January 10, 2017, we identified the adult female Dakota and Aguilar male, and on January 25, 2019, we identified Dakota, Aguilar, and the other adult male Menacho. Additionally, we identified four different adult males in 2011, 2013, 2020, and 2022. On January 25, 2019, the group fed on multiple species: first a leatherback turtle (Dermochelys coriacea) and then a bottlenose dolphin (Tursiops truncatus) 1 h later. We also observed the group chasing Humboldt penguins (Spheniscus humboldti) and South American fur seals. This sighting lasted approximately 3 h and 30 min.
3.3 First hunting record on dusky dolphins in the Mejillones Peninsula
We observed the Menacho group hunting dusky dolphins in pairs. On May 23, 2023, a group of observers recorded the group’s behavior and described an adult female (maybe Dakota) eating a dusky dolphin and sharing the carcass with the calf and the rest of the group. One observer captured an image of the matriarch female throwing the dusky dolphin into the air (Figures 3A, B). The group ate the dusky dolphin, and one individual (either the matriarch or the other adult female) held the dolphin in its mouth and allowed the rest of the individuals to tear off smaller portions of dolphin meat to eat. The next sighting, May 17, 2023, was during this study the documentation of the Menacho pod eating a dusky dolphin (Figures 3C-F); 15 min before the encounter, we observed a mixed group of dusky and long-beak common dolphins in the bay, swimming slowly and erratically. When the orcas were observed, they were already eating a dusky dolphin. We launched the drone and recorded the eating behavior. We observed how the adult female (presumably the calf’s mother) held the dusky dolphin with her mouth, allowing to the calf to bite off smaller pieces of meat (Figures 3C, D). After the calf ate, Aguilar also ate parts of the dolphin, starting with the head (Figure 3E). Finally, the young mother and calf seemed to eat the last piece of dolphin together (Figure 3F). After 15 min, the orcas had consumed the dusky dolphin carcass and started swimming slowly to the south of the Mejillones Peninsula. This was the last sighting of this orcas pod in the area for 2023.

Figure 3. Menacho pod observed foraging on dusky dolphins in Mejillones Bay. (A, B) corresponds to the sighting of the 23rd of May 2023, when the female matriarch Dakota(Figure 2B) was observed pursing a dusky dolphin and throwing to the air and finally hunting it. From C to F corresponds to the same Menacho pod observed the 27th of May 2023 foraging on another dusky dolphin in Mejillones bay. (C, D) indicates how the matriarch Dakota shares the dolphin with the juvenile male; (E) show the adult male Aguilar eating the dolphin; (F) show how finally the young female presumably the mother of the calf eat the last piece of dolphin meat. Photocredits: Barrera (A, B), Luis Aguilar (C-F).
3.4 Orcas tooth rake marks in fin whales dorsal fins from Mejillones and HPNR
From a total of 122 different fin whales individuals photo-identified and cataloged in the Mejillones Peninsula by Guevara et al. (2024), five fin whales showed orcas tooth rake marks in their dorsal fins. García-Cegarra (2022) previously described four, and here, we provide evidence of a new fin whale sighting in the Mejillones Bay with orca tooth rake marks in its dorsal fin. From a total of 88 different fin whales individuals photo-identified in Chañaral, Choros, and Damas Islands, two showed evidence of potential orca tooth rakes in their dorsal fins (Supplementary Figure S1). These observations correspond to a 2.27% of the catalogued fin whales for HPNR.
4 Discussion
Here, we describe more recent (2010–2023) sightings of orcas along northern Chile and show that orcas in the Humboldt Current System include dusky dolphins in their diet. Although these orcas have been observed previously foraging on South American sea lions and pursuing mixed groups of dusky and long-beaked common dolphins likely in an attempt to hunt them (García-Cegarra, 2022), we describe the first successful hunt by orcas on dusky dolphins.
4.1 Updated orcas sightings along northern Chile
Citizen science is growing especially in remote places as the Atacama Desert coastline where they have proved to provide valuable sightings of cetaceans species along the coast (García-Cegarra et al., 2021). Here we prove again that citizen science may help researchers to describe natural behaviors such as orcas hunting strategies and be useful for update orca sightings along northern Chile. In this study most of orca sightings along northern Chile were performed through citizen scientist (22 of 28 sightings) and thanks to the use of social network and mobile phone cameras sightings were registered by researchers to confirm species identification and group composition.
Sightings along the Humboldt Current System in northern Chile were confirmed as small groups of 1–9 orcas, which is similar to that observed for Type A orcas foraging on elephant seals and South American sea lions in Argentina (southwestern Atlantic) and in Chilean Patagonia (Häussermann et al., 2013; Capella et al., 2014). This group size is also similar to what was found in the Galapagos Islands (Alarcón et al., 2012). Smaller groups seem to be marine mammal specialists in the North Pacific and North Atlantic (Baird and Dill, 1996; Ford, 2018). Although we could not collect skin biopsy samples for genetic analyses to identify the Humboldt Current System ecotype, García-Cegarra (2022) highlighted that the white eye patch in the orcas observed in the Menacho pod is smaller than Type A and bigger than Type B orcas, an observation that we corroborated here. Further studies should investigate this detail and identify whether orcas in the Humboldt Current System might be the Type A ecotype of the southern Hemisphere.
4.2 Dolphin hunters and prey sharing among group members
Hunting strategies had not been described previously for Humboldt Current orcas; only García-Cegarra (2022) had described them as foraging on South American sea lions and pursuing mixed groups of dusky and long-beaked common dolphinsin HPNR. In this study, with drone footage, we confirmed successful hunting on dusky dolphins as well asprey sharing behaviors between group members. Wright et al. (2016) has described prey sharing for resident orcas foraging on salmon in the NE Pacific. There, field observations indicate that the prey is broken apart prior to sharing, and all pod members consume small pieces of salmon. Wright et al. (2016) suggested that prey sharing is kin-directed, with adult members trying to benefit their offspring; however, for marine mammal-eating orcas, food sharing might be the result of hunting large prey that must be cooperatively acquired (Hemilich-Boran, 1988; Hoelzel, 1993; Ford and Ellis, 2006). We were able to record just one sighting of the matriarch hunting a dusky dolphin. García-Cegarra (2022) observed the same Menacho pod pursuing a mixed group of dusky and long-beak common dolphins, although only the adult males Aguilar and Menacho were involved, excluding the adult female Dakota and her calf. In our study, we observed the Menacho pod sharing the dolphin carcass, but not in coordinated hunting. Therefore, we cannot confirm if dolphin hunting is a cooperative technique in the Humboldt Current System, although an image provided by colleagues during the sighting of Menacho pod hunting dusky dolphins on May 23,2023, showed Dakota chasing the dusky (Figure 3A). Cooperative hunting has been described for Type B orcas foraging on seals in the Antarctic Peninsula (Pitman and Durban, 2012). They are pack-ice specialists that create a wave-wash attack that often results in the capture of Weddell seals. Type B orcas share their prey in every hunting event and all group members feed, which is also how Type A orcas of Argentina are described to feed after stranding themselves to catch South American sea lions and elephant seals (López and López, 1985; Hoelzel, 1991; Iñiguez, 2001). Type A orcas share food with group members even though only one individual performs the stranding event (Hoelzel, 1991). These sharing events are considered to be kin directed when members of the same matriline will have the highest inclusive fitness benefits (Wright et al., 2016). Although we could not collect skin tissue samples for genetic analysis to assess kinship in the Menacho pod, we know this pod is led by the recognized matriarch Dakota, as she has been observed nine times along the Humboldt Current System in northern Chile, from Chañaral Island to El Loa from 2018–2023 (García-Cegarra, 2022). Food sharing from this matriarch first to the juvenile male, later the lone adult male, and finally the adult female who also shared with her presumed offspring, confirms a kin-related food sharing that could benefit all group members’ fitness.
There are no abundance estimates of dusky dolphins along northern Chile. Using citizen science observations, García-Cegarra et al. (2021) reported 6,670 dusky dolphin individuals in groups averaging 167 individuals (min = 1 max = 1000) on 58 occasions in a 5-y study in the Mejillones Bay. During the same period, the authors report 3,360 long-beak common dolphin individuals in groups averaging 134 individuals (min = 10, max = 500) on 45 occasions (García-Cegarra et al., 2021). From this one study, it would seem that dusky dolphins are more abundant in the area than long-beaked common dolphins and might be more available for orcas to hunt in larger groups.
In Chilean Patagonia, orcas have been observed foraging on sea lions and fur seals as well as attacking sei whales (Balaenoptera borealis) and fin whales (Häussermann et al., 2013; Capella et al., 2014; Pitman et al., 2023). Our observations provide new evidence of fin whale dorsal fins with orca tooth rake marks, perhaps indicating that orcas at least attack fin whales in northern Chile. None of the orcas described in this study have been observed in Patagonia, as we have no match among catalogues. This may suggest that orcas in northern Chile do not migrate to the Chilean Patagonia or the Antarctic. Further studies should compare orcas present in northern Chile with those in catalogues of the Antarctic Peninsula orcas. Whether orcas in this area eat sea lions more than dusky dolphins is unknown; however, Type A orcas from Argentina and New Zealand have been observed also foraging on dusky dolphins (Constantine, 1998; Coscarella et al., 2015). For orcas off Argentina, Coscarella et al. (2015) suggest that they also share dusky dolphins among group members that are kin-related via the matriarch. Further studies should use genetic analysis to examine kinship among the Menacho pod individuals; similarly, investigation of fatty acid composition in orcas blubber or stable isotopes in their skin may facilitate understanding of the contribution of dolphins, sea lions, and fin whales to their diet.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Certificado Comité de Ética, Universidad de Antofagasta (N&z.ousco;323/2021). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AG: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. JO: Data curation, Writing – review & editing. CA: Data curation, Writing – review & editing. LA: Data curation, Methodology, Writing – review & editing. FT: Conceptualization, Data curation, Formal analysis, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Boat surveys were funded by CETALAB.
Acknowledgments
The authors thank all the citizen scientists and whale-watching companies for providingreports of orca sightings along northern Chile. We especially thank M. Barrera for his contribution of the pictures of Dakota, the female orcas hunting a dusky dolphin in the Mejillones Peninsula as well as D. Cortes, F. Espinoza, and R. Gonzalez for contributing the sightings from Chañaral Island. We warmly thank A. Arriagada and C. Guevara for their effort to develop the fin whale photo-identification catalog for the Mejillones Peninsula. A. García-Cegarra was funded by ANID FONDECYT Postdoctoral 3210483.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1450624/full#supplementary-material
References
Alarcón D., Denkinger J., Gudiño C. (2012). Orcinus orca Ecuador ID catalogue 2012. Galápagos Science Center (Quito, Ecuador: Universidad San Francisco de Quito).
Álava J. J., Merlen G. (2009). Video-documentation of a orcas (Orcinus orca) predatory attack on a giant manta (Manta birostris) in the Galapagos Islands. Lat. Am. J. Aquat. Mam. 7, 81–84. doi: 10.5597/lajam00139
Álava J. J., Smith K. J., O´Hern J., Alarcón D., Merlen G., Denknger J. (2013). Observations of orcas (Orcinus orca) attacks on Bryde’s whales (Balaenopteraedeni) in the Galapagos Islands. Aquat. Mamm. 39, 196–201. doi: 10.1578/AM.39.2.2013.196
Andrade D., García-Cegarra A. M., Docmac F., Ñacari L. A., Harrod C. (2023). Multiple stable isotopes (C, N & S) provide evidence for fin whale (Balaenopteraphysalus) trophic ecology and movements in the Humboldt Current System of northern Chile. Mar. Environm. Res. 192, 106178. doi: 10.1016/j.marenvres.2023.106178
Ayres K. A., Gallagher A. J., Higuera-Rivas J. E. (2024). Orca (Orcinus orca) and shark predator-prey interactions within Cabo Pulmo National Park in the Gulf of California, Mexico. Front. Mar. Sci. 11. doi: 10.3389/fmars.2024.1407379
Baird R. W., Dill L. M. (1996). Ecological and social determinants of group size in transient orcas. Behav. Ecol. 7, 408–416. doi: 10.1093/beheco/7.4.408
Bigg M. A., Ellis G. M., Ford J. K. B., Balcomb K. C. (1987). Killer whales: A study of their identification, genealogy and natural history in British Columbia and Washington State (Nanaimo, British Columbia: Phantom Press).
Black N. A., Schulman-Janiger A., Ternullo R. L., Guerrero-Ruiz M. (1997). Killer whales of California and Western Mexico: a catalog of photo-identified individuals (NOAA-TM-NMFS-SWFSC-247).
Black N. A., Talty C. M., Schulman-Janiger A., Srinivasan M. (2023). “Mammal hunting orcass off Monterey, California: a 30 year synthesis,” in Social strategies of carnivorous mammalian predators. Fascinating Life Sciences. Eds. Srinivasa M., Würsig B. (Springer, Cham). doi: 10.1007/978-3-031-29803-5_8
Bruyn P. J. N., Tosh C. A., Terauds A. (2012). Killer whale ecotypes: is there a global model? Biolog. Rev. 88 (1), 62-80. doi: 10.1111/j.1469-185X.2012.00239.x
Capella J., Abramson J. Z., Vilina Y. A., Gibbons J. (2014). Observations of killer whales (Orcinus orca) in the fjords of Chilean Patagonia. Pol. Biol. 37, 1533–1539. doi: 10.1007/s00300-014-1535-5
Capella J., Gibbons J., Vilina Y. (1999). The orcas, Orcinus orca (Delphinidae) in Chilean waters between Arica and Cabo de Hornos. Anales. del. Inst. la. Patagonia. 27, 63–72.
Constantine R. (1998). Orcas (Orcinus orca) predation on dusky dolphins (Lagenorhynchusobscurus) in Kaikoura, New Zealand. Mar. Mamm. Sci. 14, 324–330. doi: 10.1111/j.1748-7692.1998.tb00721.x
Coscarella M. A., Bellazzi G., Leonì-Gaffet M., Berzano M., Degrati M. (2015). Technique used by killer whales (Orcinus orca) when hunting for dolphins in Patagonia, Argentina. Aquat. Mamm. 41, 192–197. doi: 10.1578/AM.41.2.2015.192
Durban J. W., Fearnbach H., Burrows D. G., Ylitalo G. M., Pitman R. L. (2016). Morphological and ecological evidence for two sympatric forms of Type B killer whale around the Antarctic Peninsula. Polar Biol. doi: 10.1007/s00300-016-1942-x
Fiard O., Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Meth. Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Foote A. D., Newton J., Piertney S. B., Willerslev E., Gilbert M. T. P. (2009). Ecological, morphological and genetic divergence of sympatric North Atlantic orcas populations. Molec. Ecol. 18, 5207–5217. doi: 10.1111/1365-2656.13920
Ford J. K. B. (2018). “Orcas: Orcinus orca,” in The encyclopedia of marine mammals, 3rd ed. Eds. Würsig B., Thewissen J. G. M., Kovacs K. M. (Academic Press/Elsevier), 531–537.
Ford J. K. B., Ellis G. M. (2006). Selective foraging by fish-eating killer whales Orcinus orca in British Columbia. Mar. Ecol. Prog. Ser. 316, 185–199. doi: 10.3354/meps316185
Ford J. K. B., Ellis G. M., Matkin D. R., Balcomb K. C., Briggs D., Morton A. B. (2005). Killer whales attacks on minke whales: Prey capture and antipredator tactics. Mar. Mamm. Sci. 21, 603–618. doi: 10.1111/j.1748-7692.2005.tb01254.x
Ford J. K. B., Stredulinsky E. H., Towers J. R., Ellis G. M. (2013). Information in support of the identification of critical habitat for transient Killer whales (Orcinus orca) off the west coast of Canada. DFO. Can. Sci. Advis. Sec. Res. Doc., Iv +46p. 2012/155. doi: 10.1111/j.1748-7692.2012.00572.x
García-Cegarra A. M. (2022). Evidence of Type A-like killer whales (Orcinus orca) predating on marine mammals along the Atacama Desert Coast, Chile. Aquat. Mamm. 48, 436–448. doi: 10.1578/AM.48.5.2022.436
García-Cegarra A. M., Toro F., González-Borasca V. (2021). Citizen science as a tool to assess cetacean diversity in Atacama Desert coast. Oce. Coast. Manage. 213, 105858. doi: 10.1016/j.ocecoaman.2021.105858
Guevara C., Arriagada A., Fishbach M., Van Bressem M. F., Van Waerebeek K., García-Cegarra A. M. (2024). Site fidelity and visual health assessment of two distinct fin whale populations from the Northeast and Southeast Pacific Ocean (Valdivia, Chile: University of Antofagasta).
Haro D., Blank O., Garrido G., Cáceres B., Cáceres M. (2023). Unrevealing the enigmatic type D killer whales (Orcinus orca): mass stranding in the Magellan Strait, Chile. Pol. Biol. 46, 801–807. doi: 10.1007/s00300-023-03156-2
Häussermann V., Acevedo J., Försterra G., Bailey M., Aguayo-Lobo A. (2013). Killer whales in Chilean Patagonia: Additional sightings, behaviouralobservations and individual identifications. Rev. Biología. Marina. y Oceanografía. 48, 73–85. doi: 10.4067/S0718-19572013000100007
Hemilich-Boran J. R. (1988). Behavioral ecology of killer whales (Orcinus orca) in the Pacific Northwest. Canad. J. Zool. 66, 565–578. doi: 10.1139/z88-084
Higuera-Rivas J. E., Hoyos-Padilla E. M., Elorriaga-Verplancken F. R., Rosales_Nanduca H., Rosenthal R., Urbán R. J. (2023). Short note: orcas (Orcinus orca) use different strategies to prey on rays in the Gulf of California. Aquat. Mammals. 49, 7–18. doi: 10.1578/AM.49.4.2023.7
Hoelzel A. R. (1991). Killer whales predation on marine mammals at Punta Norte, Argentina; food sharing, provisioning and foraging strategy. Behav. Ecol. Sociobiol. 29, 197–204. doi: 10.1007/BF00166401
Hoelzel A. R. (1993). Foraging behavior and social group dynamics in Puget Sound killer whales. Anim. Behav. 45, 581–591. doi: 10.1006/anbe.1993.1068
Houghton J., Holt M. M., Giles D. A., Hanson M. B., Emmons C. K., Hogan J. T., et al. (2015). The relationship between vessel traffic and noise levels received by killer whales (Orcinus orca). PLoSOne 10, e0140119. doi: 10.1371/journal.pone.0140119
Iñiguez M. (2001). Seasonal distribution of killer whales (Orcinus orca) in Northern Patagonia, Argentina. Aquat. Mamm. 27, 154–161.
LeDuc R. G., Robertson K. M., Pitman R. L. (2008). Mitochondrial sequence divergence among Antarctic killer whale ecotypes is consistent with multiple species. Biol. Lett. 4, 426–429. doi: 10.1098/rsbl.2008.0168
Loizaga R., García N., Vales D. G., Duarte C. A. (2023). Killer whales at northern Patagonia, Argentina: Evidence of different foraging groups from stable isotopes. Mar. Mamm. Sci. 39 (4), 1121–1135. doi: 10.1111/mms.13048
López J. C., López D. (1985). Killer whales (Orcinus orca) of Patagonia and their behavior of intentional stranding while hunting nearshore. J. Mammal. 66, 181–183. doi: 10.2307/1380981
Marin V., Delgado L., Escribano R. (2003). Upwelling shadows at Mejillones Bay (northern Chilean coast): a remote sensing in situ analysis. Investigaciones. Marinas,Valparaiso. 31, 47–55.
McInnes J. D., Buckmaster J. N., Cullen K. D., Mathieson C. R., Tawse J. P. (2020). International stranding by mammal-hunting killer whales (Orcinus orca) in the Salish Sea. Aquat. Mamm. 46, 556–560. doi: 10.1578/AM.46.6.2020.556
Mehta A. V., Allen J. M., Constantine R., Garrigue C., Jann B., Jenner C., et al. (2007). Baleen whales are not important as prey for killer whales Orcinus orca in high-latitude regions. Mar. Ecol. Progr. Ser. 348, 297–307. doi: 10.3354/meps07015
Minton G., Van Bressem M. F., Willson A., Collins T., Al Harthi S., Willson M. S., et al. (2022). Visual health assessment and evaluation of anthropogenic threats to Arabian Sea humpback whales in Oman. J. Cetac. Res. Manag. 23, 59–79. doi: 10.47536/jcrm.v23i1.336
Morin P. A., Parsons K. M., Archer F. I., Avila Arcos M. C., Barrett-Lennard L. G., Dalla Rosa L., et al. (2015). Geographic and temporal dynamics of a global radiation and diversification in the killer whale. Mol. Ecol. 24, 3964–3979. doi: 10.1111/mec.1328
Nottestad L., Fernö A., Axelsen B. E. (2002). Digging in the deep: Orcas’ advanced hunting tactic. Pol. Biol. 25, 939–941. doi: 10.1007/s00300-002-0437-0
Pitman R. L., Durban J. W. (2010). Killer whale predation on penguins in Antarctica. Pol. Biol. 33, 1589–1594. doi: 10.1007/s00300-010-0853-5
Pitman R. L., Durban J. W., Greenfelder M., Guinet C., Jorgensen M., Olson P. A., et al. (2011). Observations of a distinctive morphotype of killer whale (Orcinus orca) type D from subantarctic waters. Polar Biology 34, 303–306.
Pitman R. L., Durban J. W. (2012). Cooperative hunting behavior, prey selectivity and prey handling by pack ice killer whales (Orcinus orca), type B, in Antarctic Peninsula waters. Mar. Mamm. Sci. 28, 16–36. doi: 10.1111/j.1748-7692.2010.00453.x
Pitman R. L., Ensor P. (2003). Three forms of killer whales (Orcinus orca) in Antarctic waters. J. Cet. Res. Manage. 5, 131–140. doi: 10.47536/jcrm.v5i2.813
Pitman R. L., Schulman-Janiger A., Guerrero-Ruiz M. E., Ortega-Gonzalez A. M., Nanduca H. R., Fishbach M., et al. (2023). Records of fatal killer whales (Orcinus orca) attacks on fin whales (Balaenopteraphysalus) with an emphasis on Baja California, Mexico. Aquat. Mamm. 49, 195–207. doi: 10.1578/AM.49.2.2023.195
Reeves R., Pitman R. L., Ford J. K. B. (2017). Orcinus orca. In: The IUCN Red List of Threatened Species 2017 (Accessed 10 June 2024). doi: 10.2305/IUCN.UK.2017-3.RLTS.T15421A50368125.en
Remili A., Dietz R., Sonne C., Samarra F. I. P., Rikardsen A. H., Kettemer L., et al. (2023). Quantitative fatty acid signature analysis reveals a high level of dietary specialization in killer whales across the North Atlantic. J. Anim. Ecol. 92 (6), 1216–1229. doi: 10.1111/1365-2656.13920
Santos M. C. O., Netto D. F. (2005). Killer whale (Orcinus orca) predation onfranciscana dolphin (Pontoporia blainvillei) in Brazilian waters. Latin Am. J. Aquat.Mamm. 4 (1), 69–72. doi: 10.5597/lajam00072
Santos-Carvallo M., Toro F., Perez-Alvarez M. J., González S. M. (2023). Curly tails: rare occurrence of bent flukes in free-ranging cetaceans. Aquatic Mammals 49 (3), 274-281. doi: 10.1578/AM.49.3.2023
Thiel M., Macaya E. C., Acuña E., Arntz W. E., Bastias H., Brokordt K., et al. (2007). The Humboldt Current System of Northern and Central Chile – oceanographic processes, ecological interactions and socio-economic feedback. Oceanography. Mar. Biol.: Annu. Rev. 45, 195–344. doi: 10.1201/9781420050943.ch6
Tixier P., Gasco N., Duhamel G., Guinet C. (2016). Depredation of Patagonian toothfish (Dissostichuseleginoides) by two sympatrically occurring killer whale (Orcinus orca) ecotypes: insights on the behavior of the rarely observed type D killer whales. Ma.rMamm. Sci. 32, 983–1003. doi: 10.1111/mms.12307
Tixier P., Gimenez J., Reisinger R. R., Mendez-Fernandez P., Arnould J. P. Y., Cherel Y., et al. (2019). Importance of toothfish in the diet of generalist subantarctic killer whales: implications for fisheries interactions. Mar. Ecol. Progr. Ser. 613, 197–210. doi: 10.3354/meps12894
Totterdell J., Wellard R., Reeves I. M., Elsdon B., Markovic P., Yoshida M., et al. (2022). The first three records of killer whales (Orcinus orca) killing and eating blue whales (Balaenopteramusculus). Mar. Mamm. Sci. 1, 1–16. doi: 10.1111/mms.12906
Towner A., Miscarelli P., Hurwitz D., Smale M. J., Booth A. J., Stopforth C., et al. (2024). Further insights into killer whales Orcinus orca preying on white sharks Carcharodoncarcharias in South Africa. Afr. J. Mar. Sci. 46, 1–5. doi: 10.2989/1814232X.2024.2311272
Van Waerebeek K., Leaper R., Baker A. N., Papastavrou V., Thiele D., Findlay K., et al. (2010). Odontocetes of the southern ocean sanctuary. J. Cet. Res. Manage. 11, 315–346. doi: 10.47536/jcrm.v11i3.611
Visser I. (1999). Benthic foraging on stingrays by killer whales (Orcinus orca) in New Zealand waters. Mar. Mamm. Sci. 15, 220–227. doi: 10.1111/j.1748-7692.1999.tb00793.x
Visser I. N., Smith T. G., Bullock I. D., Green G. D., Carlsson O. G. L., Imberti S. (2008). Antarctic peninsula killer whales (Orcinus orca) hunt seals and a penguin on floating ice. Marine Mammal Science. 24 (1), 225–234 doi: 10.1111/j.1748-7692.2007.00163.x
Whitehead H. (2020). Cultural specialization and genetic diversity: Killer whales and beyond. J. Theor. Biol. 490, 1–11. doi: 10.1016/j.jtbi.2020.110164
Keywords: Orcinus orca, Lagenorhynchus obscurus, Southeastern Pacific, foraging, sharing
Citation: García-Cegarra AM, Oña J, Arancibia C, Aguilar L and Toro F (2024) New records of odontocete and mysticete predation by orcas in the Humboldt current system, South Pacific Ocean. Front. Mar. Sci. 11:1450624. doi: 10.3389/fmars.2024.1450624
Received: 17 June 2024; Accepted: 19 July 2024;
Published: 26 September 2024.
Edited by:
Austin Gallagher, Beneath the Waves, Inc., United StatesReviewed by:
Shannon E Aldridge, Beneath the Waves, Inc., United StatesJesus Erick Higuera Rivas, Protección y Conservación Pelágica AC, Mexico; Conexiones Terramar AC, Mexico
Copyright © 2024 García-Cegarra, Oña, Arancibia, Aguilar and Toro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frederick Toro, ZnJlZGVyaWNrLnRvcm8uY0BnbWFpbC5jb20=