 Gonzalo Araujo1,2*
Gonzalo Araujo1,2* Rebecca Bateman-John3
Rebecca Bateman-John3 Jacinta L. Shackleton1
Jacinta L. Shackleton1 Chris Garraway1Nigel Marsh4Daisy Whicheloe5Jade Rosadoni6Aimee Jan7Kohei Saito8
Chris Garraway1Nigel Marsh4Daisy Whicheloe5Jade Rosadoni6Aimee Jan7Kohei Saito8 Emily Lester9
Emily Lester9- 1Marine Research and Conservation Foundation, Somerset, United Kingdom
- 2Environmental Science Program, Department of Biological and Environmental Sciences, College of Arts and Sciences, Qatar University, Doha, Qatar
- 3Fin Focus Research, Exmouth, WA, Australia
- 4Nigel Marsh Photography, Brisbane, QLD, Australia
- 5Ningaloo Discovery, Exmouth, WA, Australia
- 6Sail Ningaloo, Coral Bay, WA, Australia
- 7OceanAimee Photography, Exmouth, WA, Australia
- 8Heppoko Taylor Inc, Yaese-cho, Okinawa, Japan
- 9Centre for Marine Ecosystem Research, Edith Cowan University, Joondalup, WA, Australia
Introduction
The Spotted Eagle Ray Aetobatus ocellatus (Kuhl 1823) is the largest member of the Aetobatidae family reaching 300 cm disc width (DW) (Last et al., 2016), but little is known about the ecology of this species. Aetobatus ocellatus is distributed throughout the Indo-West Pacific and possess k-selected life history traits, such as slow growth and late maturity (Schluessel et al., 2010), which render this species susceptible to overexploitation. In parts of its range, for example in Borneo, A. ocellatus is commonly caught in coastal fisheries including trawlers and demersal longlines (Last et al., 2010). Such threats have led to inferred population declines of 50-79% over the last three generations (39 years) across much of its range, resulting in A. ocellatus being uplisted to Endangered in the recent IUCN Red List of Threatened Species assessment (Finucci et al., 2024). Given the magnitude of these threats to the species, it is necessary to identify critical habitats essential for management and conservation.
Due to their evasive nature, behavioural observations of A. ocellatus can be logistically challenging to obtain. To date, ten distinct behaviours of A. ocellatus have been described, with the predominant behaviours being swimming, foraging, chafing, cruising and escaping threats (Berthe et al., 2018). Chafing in A. ocellatus has been reported in French Polynesia and inferred as a behaviour to rid itself of parasites (Berthe et al., 2017), likely analogous to cleaning behaviour associated with cleaning stations that has been observed in other elasmobranch species, such as the Oceanic Manta Ray Mobula birostris and the Reef Manta Ray M. alfredi (Jaine et al., 2012; O’Shea et al., 2010; Barr and Abelson, 2019). Escaping is an avoidance behaviour in response to swimmers or close vessel approaches or loud boat engine noise (Berthe and Lecchini, 2016). In response to disturbance or a predator, eagle rays speed-up and swim away from the threat (Berthe and Lecchini, 2016), which is likely to incur an energetic cost. Recovery from these excursions may take the form of resting behaviour, as documented in A. narinari in Belize (Flowers and Kelley, 2020) and the Bahamas (Silliman and Gruber, 1999) where rays were recorded as being completely stationary on the substrate for prolonged periods of time. Although the drivers of this resting behaviour remains unknown, this behaviour does suggest that pelagic eagle rays utilise benthic environments for purposes beyond foraging.
Understanding the behaviour of wide-ranging and evasive elasmobranchs is not trivial and tends to be both resource and time intensive. Citizen science can complement dedicated research efforts by increasing the amount of in-water survey effort and thus data produced (Chin and Pecl, 2018; Bateman et al., 2024). For batoid species, citizen science has been used to understand the connectivity of reef manta rays Mobula alfredi across the Indonesian archipelago (Germanov and Marshall, 2014), to discern the range and occurrence of the rare ornate eagle ray Aetomylaeus vespertilio (Araujo et al., 2020a), and to understand the occurrence and abundance of 11 species in the Canary Islands (Tuya et al., 2021), amongst others. Behavioural studies have also made use of citizen science, for example, in the Philippines to understand the philopatric behaviour of whale sharks Rhincodon typus (McCoy et al., 2018), and in Fiji to measure the effects of provisioning on the occurrence and abundance of sharks (Ward-Paige et al., 2020). Citizen science can therefore play a key role in our understanding of threatened and rare species, including shedding light on their elusive behaviours.
Here, we present eight examples of previously unreported resting behaviour by A. ocellatus, six of which are from Australia, in Ningaloo Reef, Western Australia, and at Lady Elliot Island (LEI) and Mooloolaba in Queensland (Figure 1), with additional observations from both the Maldives and Japan (Table 1). Additionally, we report resting individuals also engaging in cleaning behaviour in association with the blue-streak cleaner wrasse Labroides dimidiatus Valenciennes 1839, the moon wrasse Thalassoma lunare Linnaeus 1758 and the schooling bannerfish Heniochus diphreutes Jordan 1903.

Figure 1. Map and examples of resting and cleaning behaviour of the Spotted Eagle Ray Aetobatus ocellatus. The highlighted areas indicate the known range of the species (IUCN Red List of Threatened Species, Finucci et al., 2024) and points indicate recorded observations of resting behaviour. Example photographs of resting behaviour are from (A) Radshoo Atoll, Maldives, (B) Ningaloo Reef, Australia, (C, D). Lady Elliot Island, Australia. Note the red circles in (C, D) highlight the cleaner fish species identified in association with A. ocellatus: the blue-streak cleaner wrasse Labroides dimidiatus, the moon wrasse Thalassoma lunare and the schooling bannerfish Heniochus diphreutes.
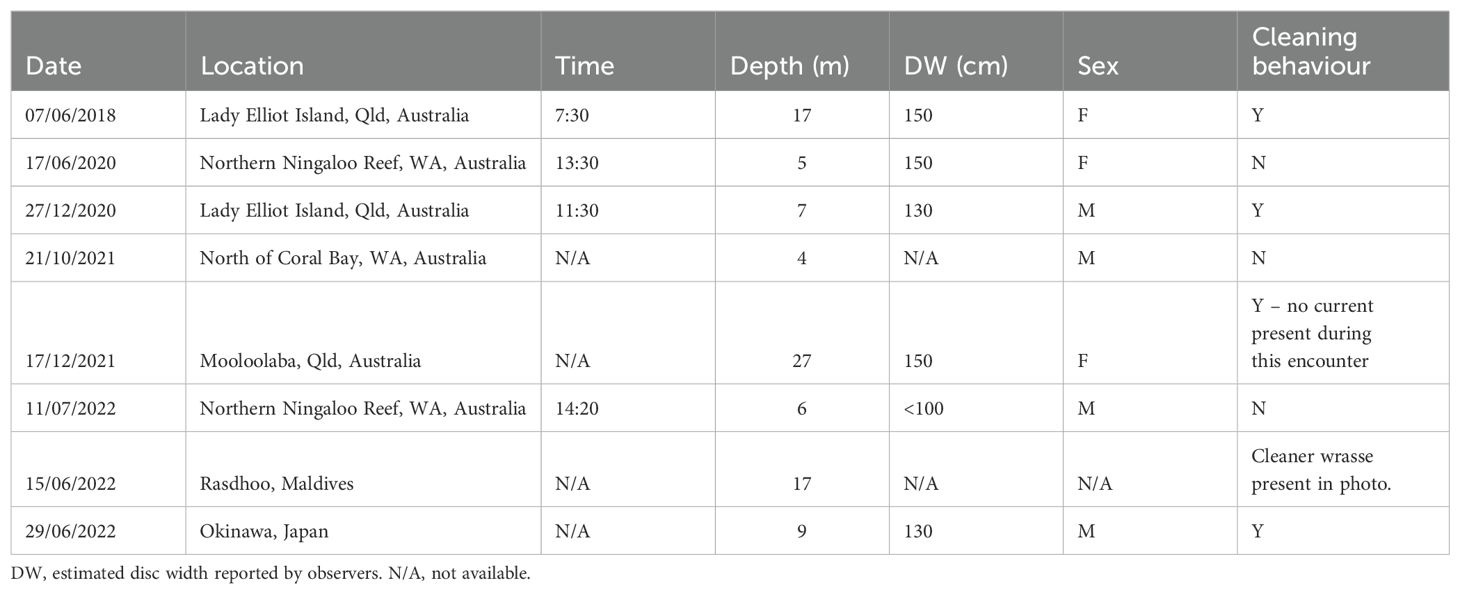
Table 1. Location of resting Spotted Eagle Rays Aetobatus ocellatus and further information collected where available.
Methods and results
Observations of resting and cleaning behaviours by A. ocellatus were sourced and collated from social media posts, outreach and existing citizen science initiatives, including Fin Focus Research that invites photo submissions of elasmobranchs from recreational ocean users and ecotourism industry crew while on tour (see https://www.facebook.com/finfocusresearch). Where possible, observers recorded estimate disc width (in m), behaviour and sex. From the collected footage, cleaner fish species were visually identified using Allen et al. (2003) and validated through FishBase (www.fishbase.org).
Here we describe six of the eight reported examples of resting A. ocellatus for which further information was available (Table 1). On 07 June 2018 at 7:30 am (moon irradiance 40%, low tide c. 9:10 am) a female A. ocellatus was observed resting at ‘Lighthouse Bommies’ in LEI (24.11 S, 152.71 E) albeit no depth information was provided. While resting, this individual was also being cleaned by three species of fish: blue-streak cleaner wrasse Labroides dimidiatus, the moon wrasse Thalassoma lunare Linnaeus 1758 and the schooling bannerfish Heniochus diphreutes Jordan 1903 (Figure 1C). This individual was visually estimated at 150 cm DW and 230 cm total length. The ray departed following collection of photo-ID footage. Similarly, on 17 June 2020, at 1:30 pm (moon irradiance 14%, low tide c. 2:30 pm) at ‘Ranger Bommies’ on the Ningaloo Reef (21.93 S, 113.94 E; Figure 1B), an A. ocellatus female individual was observed resting on the substrate at c. 5 m depth, visually estimated at 150 cm disc width (DW) and total length of 230 cm (Figures 1A, B). There was no predator (i.e., shark or cetacean) noted within the vicinity of the resting eagle ray.
On 27 December 2020 at 11:30 am (moon irradiance 94%, low tide c. 12:50 pm) at ‘Lighthouse Bommies’ on LEI (Figures 1C, D) a male A. ocellatus was observed resting on the substrate at c. 7 m depth, visually estimated at 130 cm DW. Upon close inspection, this individual ray was being cleaned by a blue-streak cleaner wrasse L. dimidiatus Valenciennes 1839. A fourth observation was filmed and reported on 21 October 2021 at c. 9:30 am (moon irradiance 99.9%, low tide c. 4:45 am) from ‘Mooch Patch’ in approximately 4 m depth north of Coral Bay (22.91 S, 113.77 E) on the Ningaloo Reef. The male A. ocellatus was observed resting (with a remora on the substrate behind the ray) for a few minutes, and when the observer dived down adjacent to the ray it slowly rose and swam away. No other information was recorded for this encounter.
A fifth observation was made at the HMAS Brisbane dive site in Mooloolaba, Queensland, on 17 December 2021 at 9:30 am (moon irradiance 97.5%, low tide c. 1:30 am). The ray was resting at 27 m whilst getting cleaned by at least four blue-streak cleaner wrasse L. dimidiatus (Supplementary Figure 1). A sixth observation was made at 2:20 pm on 11 July 2022 (moon irradiance 92.8%, low tide c. 2:39 pm), again at ‘Ranger Bommies’ on the Ningaloo Reef during a snorkel ecotour. A male A. ocellatus was observed resting on the substrate at c. 6 m depth, visually estimated at >100 cm DW, with a large Bottlenose Wedgefish Rhynchobatus australiae also seen resting approximately 5 m away. The photographer making the observation dived down very close to the A. ocellatus for photographs, and no cleaner fish were observed. As the diver ascended and broke the surface, the ray began to move away. A female zebra shark Stegostoma tigrinum was also observed resting at the same site during the snorkel.
On 15 June 2022, a seventh observation was reported in Rasdhoo atoll, Maldives (moon irradiance 99.8%), of a resting A. ocellatus. Although at least two blue-streak cleaner wrasses can be observed in the photo, we cannot assume active cleaning was taking place. An eight observation was reported from Diamond Beach, Okinawa, Japan, on 29 June 2022, of a resting A. oceallatus at 9 m depth. The ray was also getting cleaned by at least two blue-streak cleaner wrasse L. dimidiatus (see Supplementary Video 1).
Discussion
The resting behaviour of benthopelagic rays is not fully understood, nor well documented. For which size was estimated herein were adult-sized (100-150 cm DW; Last et al., 2016; Schluessel et al., 2010). There are several plausible hypotheses for the observed resting behaviour. The first, is that resting behaviour is a result of post-copulatory or post-courtship behaviour. This explanation seems unlikely, however, given the lack of mating scars on the three confirmed female A. ocellatus reported here, and that courtship/copulation usually involves the male grasping or biting the female (Berthe, 2017; McCallister et al., 2020). A second theory, is that eagle rays might rest for short periods of time before foraging in the sand (Smith and Merriner, 1985) or potentially after feeding. Matern et al. (2000) suggest that feeding on deeply burrowed prey items may be energetically demanding, and that Bat Rays Myliobatis californicus feed in shallow, warm waters and subsequently move to rest in cooler water, potentially acquiring metabolic or reproductive benefits. Furthermore, Flowers and Kelley (2020) noted that the resting A. narinari had sand streaks on its dorsum, which they generate during foraging with sediment jetting through their spiracles, further highlighting the likelihood of this as post-foraging behaviour. Finally, resting behaviour may be a form of predator avoidance, as reported in French Polynesia (C. Berthe, pers. comm.). A decrease in activity can lower the probability of an animal being predated upon (Werner and Anholt, 1993; Gerritsen and Strickler, 1977), by reducing the probability of being detected or encountered (Lima, 1998); by remaining stationary, the ray may be able to exercise vigilance against potential predators by scanning its environment for the presence of predators. This may allow the animal to avoid or delay a flight response, therefore reducing the energetic costs associated with fleeing a predator (Møller and Erritzøe, 2014), albeit A. ocellatus have a dark dorsal colouration and tend to forage in sandy substrates. Given the high costs of escape behaviour, an alternative theory is that these observations may represent resting behaviour following an escape from a predator (Heithaus and Vaudo, 2012). Further research is needed, however, to elucidate the energetics of predator-prey interactions involving A. ocellatus, as suggested by Ajemian et al. (2012). Although no predators (e.g. large sharks such as hammerhead sharks) were observed in the cases presented here, their presence in these locations is not uncommon (MacNeil et al., 2020). It should be noted that humans were in-water in the vicinity of the rays during the behaviours reported herein, and short-term behavioural responses of elasmobranchs to humans have been documented (e.g. whale sharks, Araujo et al., 2017; reef sharks, Bradley et al., 2017; manta rays, Murray et al., 2020), as it has for A. ocellatus (e.g. Berthe and Lecchini, 2016).
In the Galápagos, Pacific eagle rays (Aetobatus laticeps) have also been observed resting with no obvious indication of the drivers (D. Pazmiño, pers. comm.). Our observations corroborate that observed for A. narinari in Belize (Flowers and Kelley, 2020) and indicate a possible association with cleaning behaviour. To confirm the drivers of these behaviours, future research needs to move beyond opportunistic observations. One tractable avenue could be to combine the use of accelerometer tags with animal-borne cameras (e.g. Huveneers et al., 2018) to help elucidate the drivers of eagle ray behaviour. For example, a complete decline in overall dynamic body acceleration (ODBA) following a rapid increase could indicate a predator avoidance, or a routine ODBA followed by a complete stop could indicate a resting period following an extended period of foraging. The camera footage could then ground truth the drivers of these behaviours for A. ocellatus, as has been performed on manta rays (Stewart et al., 2019).
Our observations of resting A. ocellatus also show cleaning behaviour on four occasions – likely five if counting the Maldives report – both from LEI, Mooloolaba and Japan. It remains unclear whether the cleaning was opportunistic, or whether the resting in association with cleaning is common behaviour. Cleaner-client associations have been documented for various species elasmobranchs, like the reef manta ray Mobula alfredi Krefft 1868 (Kashiwagi et al., 2011) and the pelagic thresher shark Alopias pelagicus (Nakamura, 1935; Oliver et al., 2011). In the Indo-West Pacific the moon wrasse T. lunare and blue-streaked cleaner wrasse L. dimidiatus are often the main cleaner species (e.g. Araujo et al., 2020b), though other species like the schooling bannerfish H. diphreutes are also common. Their ecological role is important for healthy communities as they remove ectoparasites, dead tissue or mucus (Côté, 2000). The relationship between A. ocellatus and cleaning stations has not been thoroughly assessed, yet our results herein highlight the species’ association with healthy reef communities, and habitat use of reef ecosystems beyond the simple need to forage. Further research is necessary to determine their fidelity to specific cleaning stations, like that observed in reef manta rays for example (Venables et al., 2020), and whether resting behaviour is indeed closely associated with cleaning. The cleaning and resting behaviour reported from Mooloolaba, Australia, noted no current present during the encounter, which raises the question whether resting behaviour during cleaning interactions could be attributable to a lack of current (NM, pers. obs.).
The observations reported here highlight the value of citizen science for documenting elasmobranch behaviour in the wild. Although further research is required to fully understand the drivers of resting behaviour in A. ocellatus, we can continue to engage members of the general public to contribute rare and interesting observations that shed light on the behavioural ecology of elusive elasmobranchs.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for this study involving animals in accordance with the local legislation and institutional requirements. No animal was restrained for this study and observations were non-invasive and opportunistic in nature, made by citizen scientists.
Author contributions
GA: Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing, Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation. RB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. CG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. NM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. DW: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JR: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AJ: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. KS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. EL: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We would like to thank Pipat Cat Kosumlaksamee for sharing photographic evidence of his encounter with a resting A. ocellatus in the Maldives.
Conflict of interest
Author NM is the owner of Nigel Marsh Photography. Author DW was employed by company Ningaloo Discovery. Author JR was employed by company Sail Ningaloo. Author AJ is the owner of OceanAimee Photography. Author KS was employed by the company Heppoko Taylor Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1406803/full#supplementary-material
References
Ajemian M. J., Powers S. P., Murdoch T. J. T. (2012). Estimating the potential impacts of large mesopredators on benthic resources: integrative assessment of spotted eagle ray foraging ecology in Bermuda. PloS One 7, e40227. doi: 10.1371/journal.pone.0040227
Allen G., Steene R., Humann P., DeLoach N. (2003). Reef fish identification: tropical pacific (Jacksonville, FL, USA: New World Publications Inc.).
Araujo G., Legaspi C., Matthews K., Ponzo A., Chin A., Manjaji-Matsumoto B. M. (2020a). Citizen science sheds light on the cryptic ornate eagle ray Aetomylaeus vespertilio. Aquat. Conservation: Mar. Freshw. Ecosyst. 30, 2012–2018. doi: 10.1002/aqc.3457
Araujo G., Miranda J. A., Allen H. L., Labaja J., Snow S., Ponzo A., et al. (2020b). Whale sharks Rhincodon typus get cleaned by the blue-streak cleaner wrasse Labroides dimidiatus and the moon wrasse Thalassoma lunare in the Philippines. J. Fish Biol. 97, 1247–1251. doi: 10.1111/jfb.14464
Araujo G., Vivier F., Labaja J. J., Hartley D., Ponzo A. (2017). Assessing the impacts of tourism on the world's largest fish rhincodon typus at panaon island, southern leyte, philippines. Aquat. Conservation: Mar. Freshw. Ecosyst. 27 (5), 986–994. doi: 10.1002/aqc.2762
Barr Y., Abelson A. (2019). Feeding – cleaning trade-off: manta ray “decision-making” as a conservation tool [Original research]. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00088
Bateman R. L., Morgan D. L., Wueringer B. E., McDavitt M., Lear K. O. (2024). Collaborative methods identify a remote global diversity hotspot of threatened, large-bodied rhino rays. Aquat. Conservation: Mar. Freshw. Ecosyst. 34, e4047. doi: 10.1002/aqc.4047
Berthe C. (2017). First ecological, biological and behavioral insights of the ocellated eagle ray Aetobatus ocellatus in French Polynesia. Biodiversity and Ecology. Available online at: https://ephe.hal.science/hal-01690359 (Accessed April 29, 2021).
Berthe C., Lecchini D. (2016). Influence of boat noises on escape behaviour of white-spotted eagle ray Aetobatus ocellatus at Moorea Island (French Polynesia). Comptes rendus biologies 339, 99–103. doi: 10.1016/j.crvi.2016.01.001
Berthe C., Lecchini D., Mourier J. (2017). Chafing behavior on a patch of sandy bottom by ocellated eagle ray (Aetobatus ocellatus). Mar. Biodiversity 47, 379–380. doi: 10.1007/s12526-016-0463-8
Berthe C., WaqaleVu V. P., Latry L., Besson M., Lerouvreur F., Siu G., et al. (2018). Distribution patterns of ocellated eagle rays, Aetobatus ocellatus, along two sites in Moorea Island, French Polynesia. Cybium 42, 313–320. doi: 10.26028/cybium/2018-424-002
Bradley D., Papastamatiou Y. P., Caselle J. E. (2017). No persistent behavioural effects of SCUBA diving on reef sharks. Mar. Ecol. Prog. Ser. 567, 173–184. doi: 10.3354/meps12053
Chin A., Pecl G. (2018). Citizen science in shark and ray research and conservation: strengths, opportunities, considerations and pitfalls (Boca Raton, FL, USA: CRC Press).
Côté I. M. (2000). Evolution and ecology of cleaning symbioses in the sea. Oceanography Mar. Biol. 38, 311–355.
Finucci B., Rigby C. L., Armstrong A. O., Rezaie-Atagholipour M. (2024). Aetobatus ocellatus. IUCN Red List Threatened Species 2024, e.T42566169A124549514. Available at: https://www.iucnredlist.org/species/42566169/124549514 (Accessed September 3, 2024).
Flowers K., Kelley M. (2020). Resting whitespotted eagle ray aetobatus narinari (Aetobatidae). J. Ichthyol. 60, 122–123. doi: 10.1134/S003294522001004X
Germanov E. S., Marshall A. D. (2014). Running the gauntlet: regional movement patterns of Manta alfredi through a complex of parks and fisheries. PloS One 9, e110071. doi: 10.1371/journal.pone.0110071
Gerritsen J., Strickler J. R. (1977). Encounter probabilities and community structure in zooplankton: a mathematical model. J. Fisheries Res. Board Canada. 34, 73–82. doi: 10.1139/f77-008
Heithaus M., Vaudo J. J. (2012). “Predator prey interactions,” in Biology of sharks and their relatives. Eds. Carrier Jeffrey C., Musick J. A., Heithaus M. R. (Boca Raton: CRC press).
Huveneers C., Watanabe Y. Y., Payne N. L., Semmens J. M. (2018). Interacting with wildlife tourism increases activity of white sharks. Conserv. Physiol. 6, coy019. doi: 10.1093/conphys/coy019
Jaine F. R. A., Couturier L. I. E., Weeks S. J., Townsend K. A., Bennett M. B., Fiora K., et al. (2012). When giants turn up: sighting trends, environmental influences and habitat use of the manta ray manta alfredi at a coral reef. PloS One 7 (10), e46170. doi: 10.1371/journal.pone.0046170
Kashiwagi T., Marshall A. D., Bennett M. B., Ovenden J. R. (2011). Habitat segregation and mosaic sympatry of the two species of manta ray in the indian and pacific oceans: manta alfredi and m. birostris. Mar. Biodiversity Records 4, e53. doi: 10.1017/S1755267211000479
Last P., Naylor G., Séret B., White W., de Carvalho M., Stehmann M. (2016). Rays of the world (Clayton South, VIC: CSIRO publishing).
Last P. R., White W. T., Caira J. N., Fahmi D., Jensen K., Lim A. P. K., et al. (2010). Sharks and rays of borneo (Collingwood: CSIRO Publishing).
Lima S. L. (1998). Stress and decision making under the risk of predation: recent developments from behavioral, reproductive, and ecological perspectives. Adv. Study Behavior. 27, 215–290. doi: 10.1016/S0065-3454(08)60366-6
MacNeil M. A., Chapman D. D., Heupel M., Simpfendorfer C. A., Heithaus M., Meekan M., et al. (2020). Global status and conservation potential of reef sharks. Nature 583, 801–806. doi: 10.1038/s41586-020-2519-y
Matern S. A., Cech J. J. J., Hopkins T. E. (2000). Diel movements of bat rays, myliobatis californica, in tomales bay, california: evidence for behavioral thermoregulation? Environ. Biol. Fishes 58, 173–182. doi: 10.1023/A:1007625212099
McCallister M., Mandelman J., Bonfil R., Danylchuk A., Sales M., Ajemian M. (2020). First observation of mating behavior in three species of pelagic myliobatiform rays in the wild. Environ. Biol. Fish 103, 163–173. doi: 10.1007/s10641-019-00943-x
McCoy E., Burce R., David D., Aca E. Q., Hardy J., Labaja J., et al. (2018). Long-term photo-identification reveals the population dynamics and strong site fidelity of adult whale sharks to the coastal waters of Donsol, Philippines. Front. Mar. Sci. 271. doi: 10.3389/fmars.2018.00271
Møller A. P., Erritzøe J. (2014). Predator–prey interactions, flight initiation distance and brain size. J. Evol. Biol. 27, 34–42. doi: 10.1111/jeb.12272
Murray A., Garrud E., Ender I., Lee-Brooks K., Atkins R., Lynam R., et al. (2020). Protecting the million-dollar mantas; creating an evidence-based code of conduct for manta ray tourism interactions. J. Ecotourism 19, 132–147. doi: 10.1080/14724049.2019.1659802
Nakamura H. (1935). On the two species of the thresher shark from formosan waters Vol. 14 (Memoirs of the Faculty of Science and Agriculture, Taihoku Imperial University), 1–6.
O’Shea O. R., Kingsford M. J., Seymour J. (2010). Tide-related periodicity of manta rays and sharks to cleaning stations on a coral reef. Mar. Freshw. Res. 61 (1), 65–73. doi: 10.1071/MF08301
Oliver S. P., Hussey N. E., Turner J. R., Beckett A. J. (2011). Oceanic sharks clean at coastal seamount. PloS One 6 (3), e14755. doi: 10.1371/journal.pone.0014755
Silliman W. R., Gruber S. H. (1999). Behavioral biology of the spotted eagle ray, aetobatus narinari (Euphrase), in bimini, bahamas; an interim report. Bahamas J. Sci. 7, 13–20.
Schluessel V., Bennett M. B., Collin S. P. (2010). Diet and reproduction in the white-spotted eagle ray Aetobatus narinari from Queensland, Australia and the Penghu Islands, Taiwan. Mar. Freshw. Res. 61, 1278–1289. doi: 10.1071/MF09261
Smith J. W., Merriner J. V. (1985). Food habits and feeding behavior of the cownose ray, Rhinoptera bonasus, in lower Chesapeake Bay. Estuaries 8, 305–310. doi: 10.2307/1351491
Stewart J. D., Smith T. T., Marshall G., Abernathy K., Fonseca-Ponce I. A., Froman N., et al. (2019). Novel applications of animal-borne Crittercams reveal thermocline feeding in two species of manta ray. Mar. Ecol. Prog. Ser. 632, 145–158. doi: 10.3354/meps13148
Tuya F., Aguilar R., Espino F., Bosch N. E., Meyers E. K., Jiménez-Alvarado D., et al. (2021). Differences in the occurrence and abundance of batoids across an oceanic archipelago using complementary data sources: Implications for conservation. Ecol. Evol. 11, 16704–16715. doi: 10.1002/ece3.8290
Venables S. K., van Duinkerken D. I., Rohner C. A., Marshall A. D. (2020). Habitat use and movement patterns of reef manta rays Mobula alfredi in southern Mozambique. Mar. Ecol. Prog. Ser. 634, 99–114. doi: 10.3354/meps13178
Ward-Paige C. A., Brunnschweiler J., Sykes H. (2020). Tourism-driven ocean science for sustainable use: A case study of sharks in Fiji. bioRxiv. doi: 10.1101/2020.02.04.932236
Keywords: ecology, participatory science, novel, Aetobatidae, Batoidea
Citation: Araujo G, Bateman-John R, Shackleton JL, Garraway C, Marsh N, Whicheloe D, Rosadoni J, Jan A, Saito K and Lester E (2024) Citizen science highlights rare behaviours by the spotted eagle ray Aetobatus ocellatus Kuhl 1823. Front. Mar. Sci. 11:1406803. doi: 10.3389/fmars.2024.1406803
Received: 25 March 2024; Accepted: 22 August 2024;
Published: 17 September 2024.
Edited by:
Austin Gallagher, Beneath the Waves, Inc., United StatesReviewed by:
Chi Hin Lam, University of Massachusetts Boston, United StatesMatthew Ajemian, Florida Atlantic University, United States
Copyright © 2024 Araujo, Bateman-John, Shackleton, Garraway, Marsh, Whicheloe, Rosadoni, Jan, Saito and Lester. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gonzalo Araujo, Z29uem9AbWFyZWNvLm9yZy51aw==