 Laura Ciriminna
Laura Ciriminna Geraldina Signa
Geraldina Signa Giovanna Cilluffo
Giovanna Cilluffo Arnold Rakaj
Arnold Rakaj Salvatrice Vizzini
Salvatrice Vizzini
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci., 21 March 2024
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1381836
This article is part of the Research TopicSea Cucumbers: The Sustainability of Emergent and Historical ResourcesView all 12 articles
Sea cucumbers are considered a traditional food delicacy in Asian countries. In last decades high market demand, uncontrolled exploitation and inadequate management have led to stock overexploitation, driving commercial interest in new target species and new areas. In recent years, Mediterranean and North-East Atlantic sea cucumbers have become fishing targets, and, due to overfishing, a significant decline of wild populations have been observed, with detrimental effects on benthic communities and ecosystems. Indeed, in addition to being an important economic resource, sea cucumbers play a key ecological role in benthic dynamics where they are involved as ecosystem engineers in the processing of organic matter in the detrital food web pathway. In this context, aquaculture can play a crucial role in supporting the conservation of natural stocks by reducing harvesting pressure on wild populations and enabling restoration programmes for depleted stocks. Commercial sea cucumber farming is already an established reality in the Indo-Pacific region, where it has become one of the most profitable sectors of aquaculture production. In Europe, on the other hand, sea cucumber aquaculture is still an emerging sector, whose development is hampered by poor and fragmented information on the farming requirements of native species. In this context, this systematic review was carried out with the aim of providing a comprehensive summary of the state of the art of farming practices of the European sea cucumbers considered as a target of commercial fisheries. A total of 34 original articles were included in this review, after title, abstract and full-text screening. The results highlighted a sharp increase in the number of studies over the last six years. Portugal, Turkey and Italy are the countries where most research has been carried out. Among the selected species, Holothuria tubulosa was the most studied, while only one article was found on Parastichopus regalis. The main aims of this review were i) to gather all available literature on the farming practices of North-East Atlantic and Mediterranean sea cucumber species, ii) to outline the most common farming practices, iii) to identify gaps and future directions in this emerging aquaculture sector.
Holothurians, or sea cucumbers, have been a popular and traditional food in Asian countries since ancient times and, today, represent an economically important market due to their high market value (Conand, 2018; Purcell et al., 2023). Sea cucumbers are processed and preserved as a dried product, commonly called “Trepang” or “Bêche-de-Mer” in the global market. The edible part of the animals, the body wall, is an appreciated food, characterised by a high protein content and a low concentration of fats, a large proportion of which are ω3-fatty acids, molecules of great value for their beneficial effects on human health (Aydin et al., 2011; Roggatz et al., 2018). Sea cucumbers are also used in traditional Chinese medicine for their bioactive compounds with anti-tumoral, antioxidant and anti-inflammatory properties, and are also applied in the cosmetic industry (Santos et al., 2016). In the Indo-Pacific region, the high harvesting pressure on sea cucumbers has led to the overexploitation of many natural stocks, even towards lower-value species (Conand, 2018). Similarly, due to the decline in tropical species abundance, Mediterranean and Atlantic sea cucumbers have recently been subject to strong fishing pressure, with some European countries starting to export their wild catches to the main Asian markets (Taiwan, Hong-Kong or Singapore; Conand, 2018). Several species inhabit European seas, but only eight are currently considered of commercial interest (Purcell et al., 2023; Rakaj and Fianchini, 2024). Holothuria poli (Delle Chiaje, 1823), H. tubulosa (Gmelin, 1791), H. sanctori (Delle Chiaje, 1823), H. forskali (Delle Chiaje, 1823), H. arguinensis (Koehler & Vaney, 1906) and H. mammata (Grube, 1840) belonging to the family Holothuriidae, Parastichopus regalis (Cuvier, 1817) and P. tremulus (Gunnerus, 1767) belonging to the family Stichopodidae, are the species that have recently experienced a significant increase in catches to meet the demands of international markets. P. regalis, is the only species that is consumed in Europe and precisely in coastal localities of Catalonia and Valencia regions, where the muscular bands are extracted and commercialized as a culinary delicacy known as “espardeña”. In the European Atlantic H. arguinensis, H. forskali, and H. mammata are collected, while in the Mediterranean Sea the most harvested species are H. poli, H. tubulosa and H. mammata, with Turkey being the main exporter (González-Wangüemert et al., 2018; Dereli and Aydın, 2021; Aydin et al., 2023; Rakaj and Fianchini, 2024).
The increase in harvesting pressure has led to a rapid decline in natural stocks of marketable species, resulting in reduction of abundance, size and genetic diversity, with detrimental effects on benthic communities and ecosystems (González-Wangüemert et al., 2014, 2015; Maggi and González-Wangüemert, 2015; Ventura et al., 2023). Indeed, in addition to being an important economic resource, sea cucumbers play a key ecological role as ecosystem engineers, and their decline may affect the entire ecosystem. In fact, sea cucumbers, being deposit-feeders, ingest large amount of sediment contributing to nutrient cycling and energy fluxes in benthic ecosystems, regulating dissolved oxygen and nutrient concentration and promoting nitrification and denitrification processes (Purcell et al., 2013; Boncagni et al., 2019).
To counteract stock depletion and protect natural populations from the spread of illegal, unreported and unregulated (IUU) fishing, some countries have introduced restrictions to regulate harvesting and catches. In Galicia, the harvest of H. forskali has been regulated since 2016 (González-Wangüemert et al., 2018). In Italy, the increasing number of massive and uncontrolled catches, especially in the southern regions, led the Italian Ministry of Agriculture, Food and Forestry (MIPAAF) to impose a ban on “fishing, holding on board, transhipping, or disembarking of sea cucumbers”. The ban has been extended annually pending the establishment of a scientific framework to support the adoption of structural measures in the management of sea cucumber stocks (Ministerial Decree n.156 of 02/27/2018).
In Asia, particularly in China and Japan, farming practices of the most popular species have been developed since the 1950s to make up for the lack of wild sea cucumbers and, in particular, aquaculture activities are focused on Apostichopus japonicus (Selenka 1867), the most prized temperate species (Brown and Eddy, 2015). In China, the cultivation of this species allows the production of juveniles (6 billion per year) to be reared at sea, in cages and in ponds, producing more than a quarter of the global wild catch in dry weight (Purcell et al., 2012).
As in the Indo-Pacific, aquaculture could be a potential solution for North-Eastern Atlantic and Mediterranean sea cucumbers. The farming system could sustain market demand by counteracting harvesting pressure on wild stocks, and the development of full-cycle production may support restocking programmes in depleted areas. Finally, the diffusion of sea cucumber aquaculture practices may contribute to the diversification of aquaculture products and the development of Multitrophic Integrated Aquaculture models (IMTA) which are essential to cope with principles of sustainable development. In this context, a systematic review of the available literature on the status of sea cucumber aquaculture in Europe was carried out. Specifically, among the species found in the North East Atlantic and the Mediterranean Sea, those currently subject to fishing activities were selected (Purcell et al., 2023) and all literature found was reviewed with the aim of providing a comprehensive summary of the most common farming practices and identifying gaps and future directions in this emerging area of aquaculture.
Scientific literature was searched in Web of Science (https://www.webofscience.com) and Scopus (https://www.scopus.com) from 2000 to 21 August 2023, using the following search string:
“Holothuria arguinensis” OR “Holothuria forskali*” OR “Holothuria mammata” OR “Holothuria poli*” OR “Holothuria sanctori” OR “Holothuria tubulosa” OR “Parastichopus regalis” OR “*Stichopus tremulus” AND “*cultur*” OR “IMTA” OR “farm*” OR “rear*” OR “diet*” OR “feed*” OR “food*” OR “bioremediat*” OR “bioturbat*” OR “nutriti*” OR “growth” OR “biomass” OR “*product*”.
Only original articles (i.e. experimental field studies, observational studies) published in English and in full text, providing information on rearing practices were considered eligible. In particular, studies on culture conditions (feed, temperature, stocking density, salinity, sediment size) and reproduction (spawning induction, larval development, juvenile culture) were included in the present review.
Exclusion criteria were: (i) studies on the ecology and biology of selected species; (ii) studies on the contribution of holothurians to pollution control without information on their fitness or growth; (iii) studies on the quality of species of commercial interest; (iv) studies on bioactive compounds of pharmaceutical or biotechnological applications.
Titles and abstracts of articles identified by the search strategy were reviewed independently by two authors, and full-text articles potentially meeting the inclusion criteria were retrieved. From the full texts, the same two authors independently established whether the studies met the inclusion criteria. Studies that did not fulfil all the inclusion criteria were excluded, and the reasons for exclusion were reported. Any disagreements were resolved by consensus. If the two review authors disagreed, a third review author was involved to resolve the disagreement. The review authors were not blinded to authors, journals, results, etc.
The search yielded a total of 417 results, 253 in Web of Science and 164 in Scopus. 79 items were removed (68 in Web of Science and 11 in Scopus), resulting in 338 articles. Duplicates (125) were removed and the remaining 213 articles were screened for inclusion or exclusion (Figure 1). After title and abstract screening, 161 articles were excluded and 52 articles were selected for full-text screening. After final screening, 34 articles were retained for review (Supplementary Table S1).
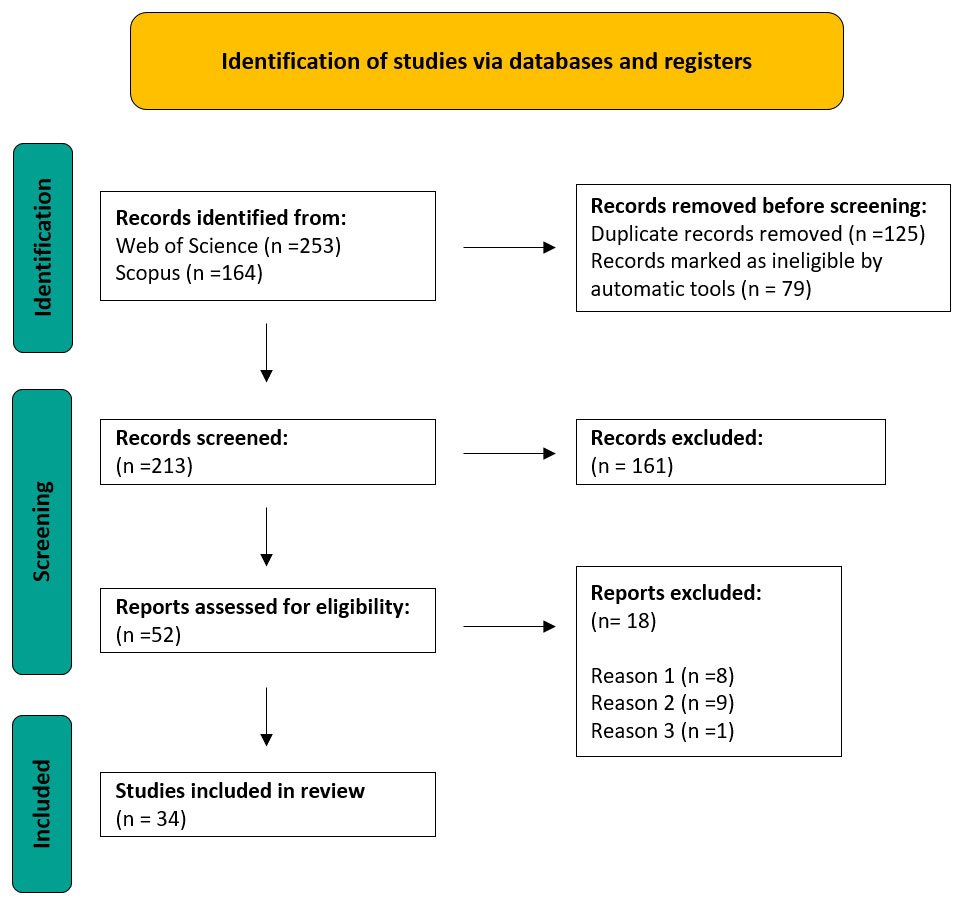
Figure 1 Search strategy following the PRISMA flow diagram. Reason 1: Articles on ecology and biology of selected species; Reason 2: Articles on the contribution of holothurians to pollution abatement without information on their well-being or ability to grow in those conditions; Reason 3: Articles on molecules of pharmaceutical or biotechnological interest.
The distribution over the years showed that production was low from 2000 to 2015 with only 5 articles published, 1 in 2005 and 2013 and 3 in 2015 (Figure 2). In contrast, from 2017 to 2023 (August), 29 studies were conducted on North-Eastern Atlantic and Mediterranean sea cucumber aquaculture, with a peak in 2018 with 6 articles published.
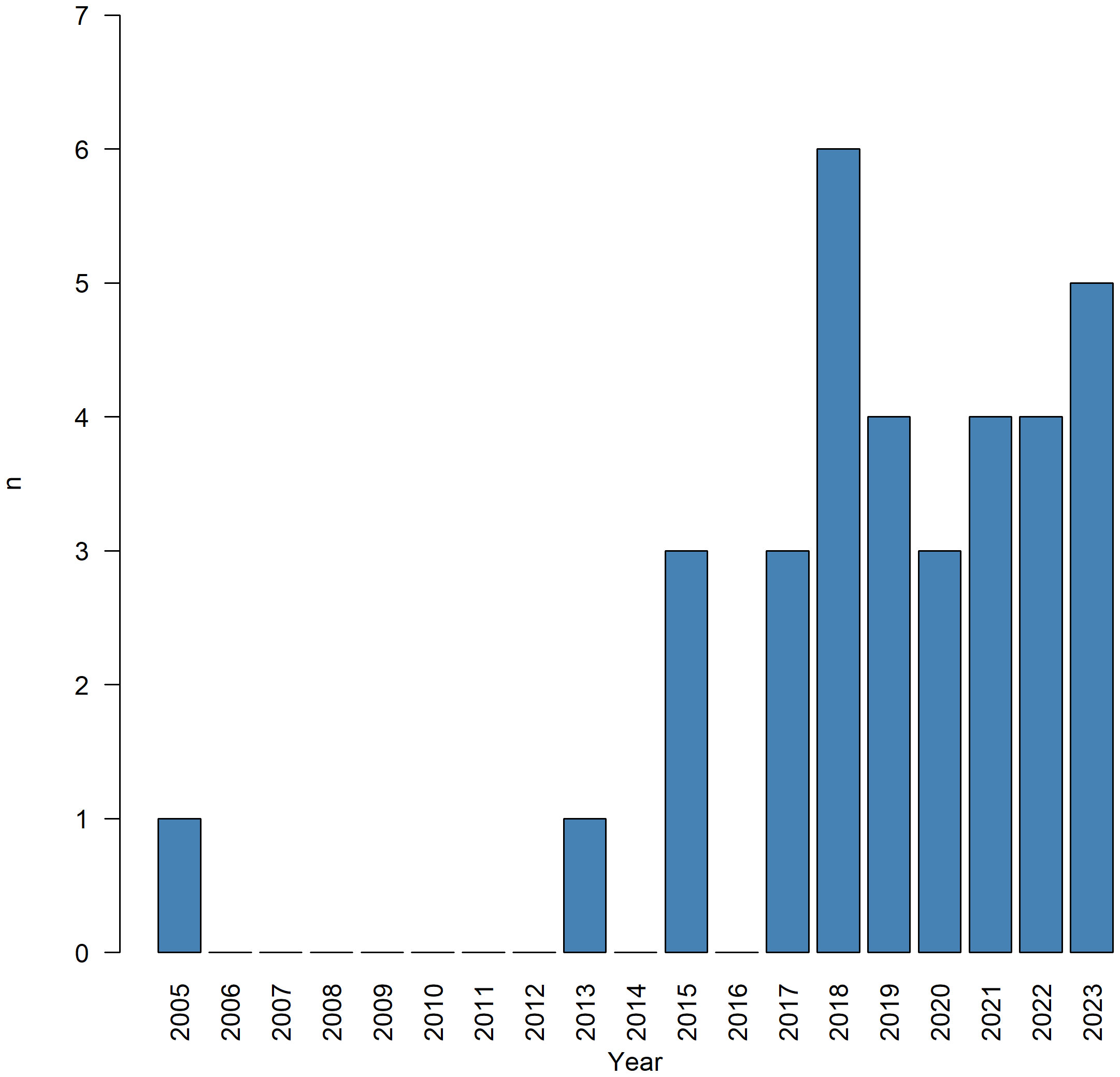
Figure 2 Distribution of publications across the years.
Among the 34 articles included in the review, the highest number of studies was found in Portugal (11), followed by Turkey (6), Italy (5), Spain, France, Sweden, Norway and Malta (2), Egypt, Greece and United Kingdom (1) (Figure 3). Among the selected species Holothuria tubulosa was the most investigated with 11 papers, followed by H. arguinensis and H. poli with 8 articles each, while only 1 paper dealt with on Parastichopus regalis (Figure 4). Most of the included papers were related to culture conditions to improve fitness and promote fattening of adult or juvenile sea cucumbers.
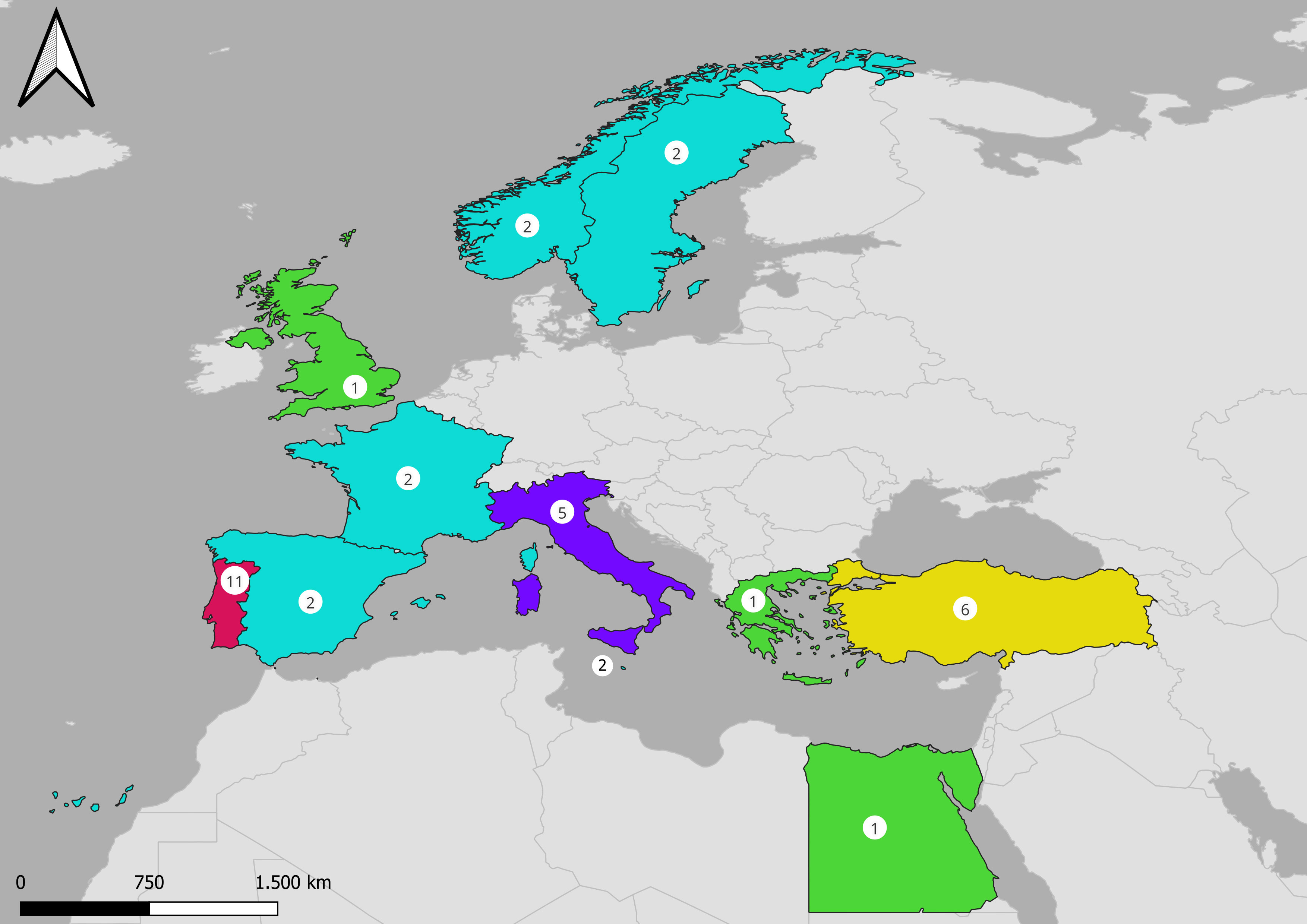
Figure 3 Geographical distribution of the studies included in the review.
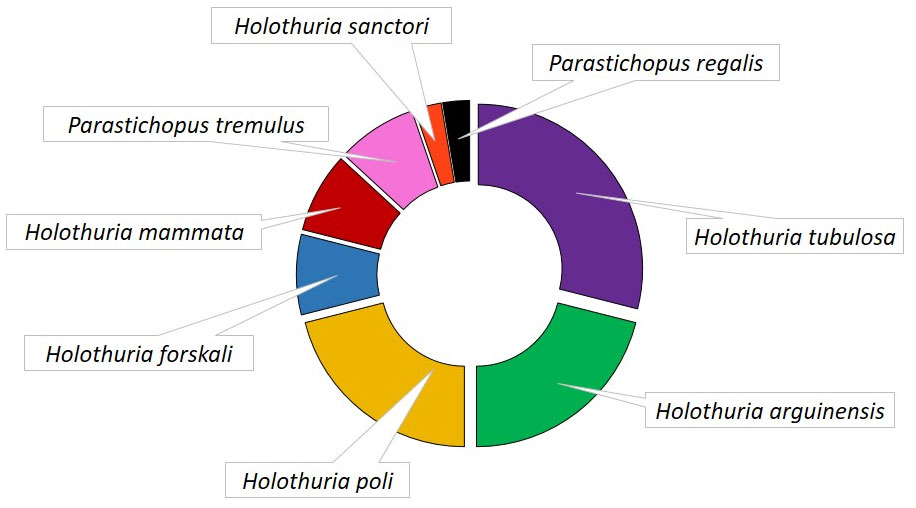
Figure 4 Frequency of the species used in the selected studies.
Feed was the most studied issue, with 10 experiments conducted under integrated multitrophic aquaculture (IMTA) conditions, in mariculture (5) or in-land systems (5). Fish were the most commonly associated species in IMTA studies, with only two experiments combining sea cucumbers with other invertebrates. Temperature and stocking density were the most commonly investigated variables. Only 9 studies were conducted on artificial reproduction and larval rearing, focusing on spawning induction methods, larval feeding regimes and larval development (Figure 5).
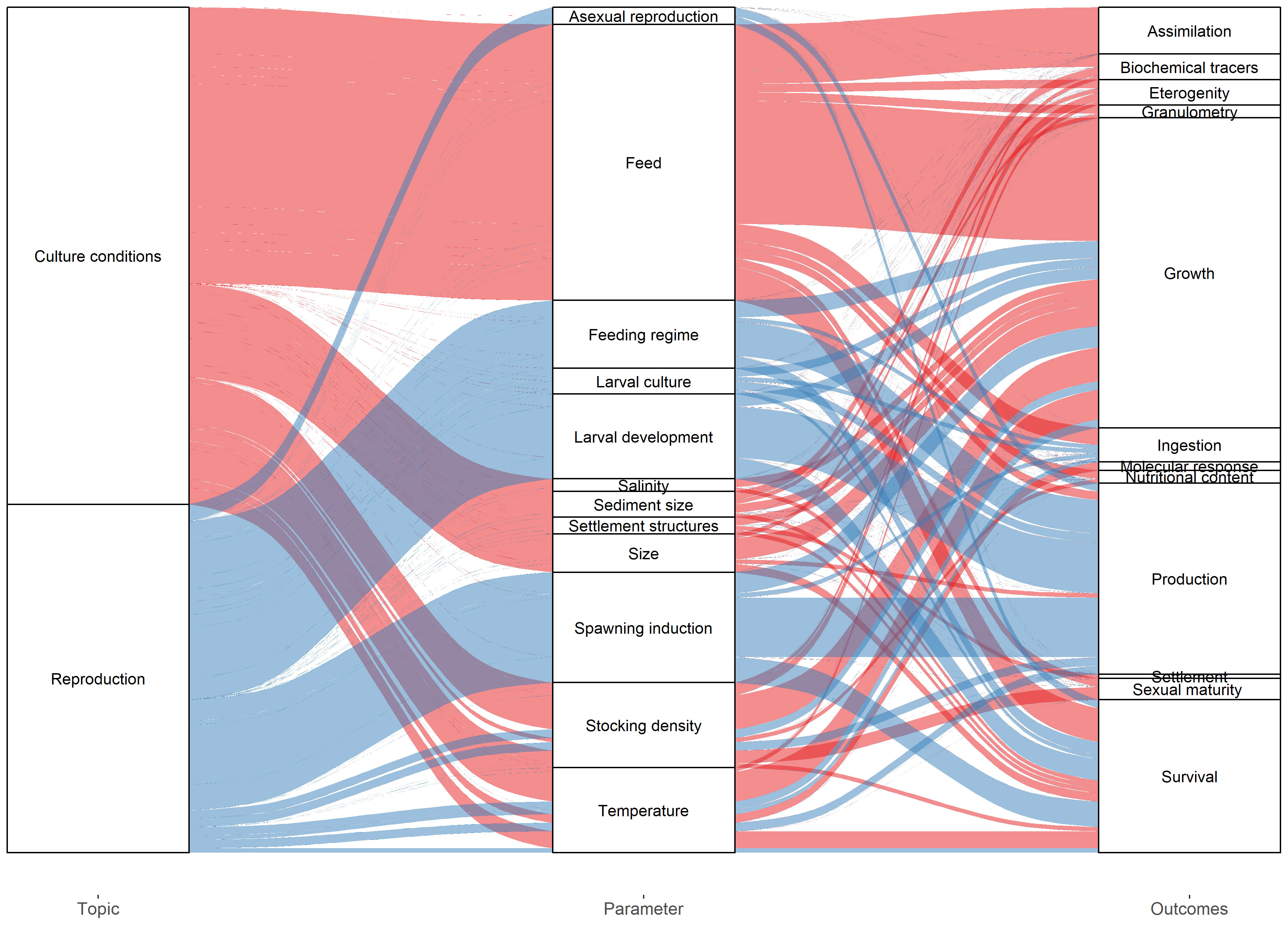
Figure 5 Alluvial diagram showing the distribution of Topic, Parameter and Outcomes investigated in the included studies.
Among the outcomes used to evaluate the response of sea cucumbers to culture conditions, the most investigated were survival (survival, survival rate or mortality rate) and growth (specific growth rate, growth rate, weight gain, relative weight gain) (Figure 6).

Figure 6 Word cloud analysis using outcomes used in the selected studies. The size of the words is proportional to the observed frequency. Same colours indicate same frequency.
Given the recent increase in sea cucumber catches and trade in North-Eastern Atlantic and Mediterranean countries, the development of aquaculture practices represents a promising solution to protect natural populations without losing the economic business generated by sea cucumber sales. The findings of this systematic review showed a sharp increase in scientific production related to sea cucumber farming in recent years (from 2017), probably in response to the natural stock depletion observed for commercial holothurians. The countries most involved in scientific research were Portugal and Turkey, which have recently reported the first signs of stock depletion in their seas (González-Wangüemert et al., 2014, 2015; Maggi and González-Wangüemert, 2015). Similarly, the most investigated species were Holothuria tubulosa, H. poli and H. arguinensis, Atlantic species also reported in the western Mediterranean Sea (Rodrigues, 2012; González-Wangüemert et al., 2014; Mezali and Thandar, 2014). Surprisingly, only one study was found for Parastichopus regalis, the most valuable European species (Roggatz et al., 2018). This is probably because this species occurs at great depths (50 - 1200 m), making it more difficult to study than shallower water species (Ramón et al., 2022). The results showed that a much high percentage of papers focused on the study of environmental variables and feeding, as optimal culture conditions are fundamental to ensure animal fitness and promote growth of sea cucumbers at all life stage. Most studies were conducted on adult specimens, whit less attention paid to larval and juvenile stages and, in particular, studies on integrated multitrophic aquaculture (IMTA) have only been carried out on sub-adult and adult specimens. Knowledge of the biology and ecology of European sea cucumber species is generally poor and almost absent for the early stages of the life cycle. Indeed, to date, there are no full-cycle farming systems able to produce juveniles and the few facilities working on the reproduction process under controlled conditions are still at the experimental stage. It is therefore not surprising that most experiments have been conducted on large specimens, which are easy to find in the wild and can be handled under controlled conditions. The large number of outcomes used to assess the response to experimental conditions also underlines the lack of defined protocols and consolidated information. Although several authors analysed the survival and growth ability of sea cucumbers, the data are not always comparable. For example, to express sea cucumber growth, some authors use total wet weight, others eviscerated weight or dry weight, while in some cases length measurements are also included. However, the ability of holothurians to change in size and weight significantly depending on the amount of water contained in their bodies makes growth determinations critical, which is probably the reason for such different measurement protocols. The lack of knowledge on the biology and ecology of marketable North-Eastern Atlantic and Mediterranean sea cucumbers is one of the main bottlenecks for the development of their culture. In particular, little information is available on their natural food sources and even less is known about effective diets that could promote the growth of sea cucumbers in culture condition at each stage of their life cycle. As the most investigated topics were reproductive techniques for hatchery production and culture conditions to promote sea cucumber growth, these two categories were used to organise the information of the reviewed articles.
The development of an effective protocol for full-cycle production is fundamental to counteract fishing pressure on natural sea cucumber stocks, as it would reduce the dependence of culture practices and market demand on wild specimens. Two studies have been conducted on the use of asexual reproduction with encouraging results the (Toscano and Cirino, 2018; Moussa et al., 2019), while other experiments have been carried out aiming at the production of larvae and juveniles in hatcheries. Protocols for artificial reproduction are available for all North-Eastern Atlantic and Mediterranean sea cucumbers of commercial interest, with the exception of P. regalis. Induction of gamete emission is a key challenge to optimise spawning and fertilisation rates in invertebrate hatcheries, so it is important to know the appropriate spawning stimulation method for each species. For H. arguinensis and H. mammata, spawning was successfully induced by thermal stimulation (gradual increase in water temperature) (Domínguez-Godino et al., 2015; Domínguez-Godino and González-Wangüemert, 2018a). Differently, for H. poli (Rakaj et al., 2019), H. forskali (Laguerre et al., 2020) and P. tremulus (Schagerström et al., 2022), spawning was obtained with thermal shock (rapid increase in water temperature). A combination of induction methods was used to induce gamete release in H. tubulosa (thermal stimulation and thermal shock, Rakaj et al., 2018) and H. sanctori (mechanical and algal stimulation, Magdy et al., 2021). Like other echinoderms, sea cucumbers showed sex order, with male specimens spawning first and presumably stimulating female gamete release, with the exception of H. forskali, which showed synchronous spawning (Laguerre et al., 2020).
The larval cycle was similar in all species, with three planktonic and feeding stages (early, mid- and late auricularia) followed by two metamorphic non-feeding stages (doliolaria and pentactula respectively). However, some species-specific differences in the duration of each larval stages were reported. In H. poli and H. arguinensis the juvenile stage was reached after 15 and 18 days respectively, whereas H. tubulosa, H. forskali, H. sanctori and P. tremulus needed more than 20 days to reach the late auricularia stage.
Larval development seems to be influenced by culture conditions. Temperature and rearing time are inversely proportional, with temperate species characterised by a faster life cycle than species typical of cold waters (Schagerström et al., 2022). Stocking density affected larval survival, with low survival rates at high density (>3 larvae/ml; Domínguez-Godino et al., 2015; Rakaj et al., 2018; Domínguez-Godino and González-Wangüemert, 2018a; Magdy et al., 2021). In all selected studies, microalgae were used as feed for larval culture stages. Tetraselmis chuii, Chaetoceros calcitrans, Isochrysis galbana, T. suecica, Rhodomonas baltica, Skeletonema marinoi, Phaeodactylum tricornutum, Dunaliella tertiolecta and C. mulleri, were considered suitable to sustain larval development. However, the mixture of microalgae promoted higher larval survival rates than treatments with single algae (Domínguez-Godino and González-Wangüemert, 2019b). In terms of nutritional content, diets rich in lipids resulted more suitable for larval development since the lipids assimilated from the food during the first larval stages are stored in specific cells, called hyaline spheres, and become important energy resources for the subsequent non-feeding larval phases (Ramofafia et al., 2003; Sousa et al., 2023). Larval development was also influenced by the feeding regime with overfeeding conditions promoting morphological malformations, while underfeeding regime resulted in high mortality rates (Rakaj et al., 2018; Schagerström et al., 2022). Also in this case, optimal conditions were species-specific with low algal concentration suitable for H. tubulosa and P. tremulus, while H. arguinensis, H. mammata, H. poli and H. forskali preferred high algal concentration (Domínguez-Godino et al., 2015; Rakaj et al., 2018, 2019; Laguerre et al., 2020; Venâncio et al., 2021; Schagerström et al., 2022).
The differences observed in larval development could be the result of specie-specific adaptive strategies, since the length of the planktonic phase influences the mortality rate and hence their range of colonisation (Domínguez-Godino and González-Wangüemert, 2018a). In addition, the duration of larval development may be related to the different reproductive effort. Indeed, it was observed that H. poli, which is characterised by faster larval development, produces large eggs (around 240 µm of diameter), suggesting the ability of this species to store high amounts of nutrient and take advantage of high feeding regimes (Rakaj et al., 2019). Similarly, H. tubulosa produced smaller eggs, preferring lower feeding conditions and showing slower larval development (Rakaj et al., 2018).
Finally, as already observed in other marine invertebrates characterized by planktonic larval stages, high mortality occurs during settlement. Settlement is induced by chemical cues provided, for example, by benthic diatoms (Schagerström et al., 2022). In addition, survival rates during this phase appear to be influenced more by the available surface area of the settlement structure than by its shape (Sousa et al., 2021).
Temperature is one of the most important physical variables affecting metabolism, growth and physiological performance of sea cucumbers (Yang et al., 2005; Dong et al., 2006). Sea cucumbers can respond to extreme temperatures by entering in a hypometabolic state called aestivation (high temperature) or hibernation (low temperature), in which specimens become inactive, stop feeding and start intestine digestion and metabolic rate depression (Zhao et al., 2022). Temperate species, such as H. tubulosa and H. arguinensis, can grow at temperature range of 20-25°C while lower temperatures (12-18°C) induce weight loss and low enzymatic and feeding activity (Günay et al., 2015, 2020; Domínguez-Godino and González-Wangüemert, 2019a; Madruga et al., 2023). High temperatures (23-28°C) are above the tolerance range of P. regalis, a species that lives in high depth where water temperatures are generally much lower. However, the ability of this species to survive at 18°C opens up the possibility of testing its cultivation also in shallow waters, throughout winter months (Galimany et al., 2018). Temperature also affects the ability of sea cucumbers to tolerate changes in salinity. H. tubulosa can grow between 30 and 45 psu in summer (25°C), reducing its tolerance to high salinity at winter temperatures (15°C).Similarly, H. poli can only tolerate salinity increases with warm water. However, both species are not able to survive under low salinity conditions (< 25 psu) (Tolon, 2017).
The stocking density is another important variable to take into account in sea cucumber culture practices in order to avoid intraspecific competition for space and food. Low stocking density is adequate for H. tubulosa (250 g/m2) and H. arguinensis (520 g/m2), while H. mammata prefers medium density (720 g/m2, Tolon et al., 2017b; Domínguez-Godino and González-Wangüemert, 2018b). In general, growth performance decreases with increasing stocking density but, while medium densities (600 – 700 g/m2) can be maintained for short periods, high densities must be avoided as negative growth was observed for all species studied (Tolon et al., 2017b; Domínguez-Godino and González-Wangüemert, 2018b). However, the critical biomass values vary between species, so species-specific tests are needed to define the appropriate rearing protocol.
Finally, as since sea cucumbers have the ability to select organic particles of different size (Ramón et al., 2019), it is important to understand size class preference for by North-Eastern Atlantic and Mediterranean sea cucumbers in order to facilitate sediment ingestion, and consequently food intake. Large sediment particles (7 mm) are not easily handled by juvenile specimens, while too small particles (0.05 – 0.2 mm), although preferred by sea cucumbers, may not be able to reach the bottom due to the aeration and water flow typical of tank systems (Hudson et al., 2005; Tolon et al., 2015). The best sediment size range seems to be between 1 and 3 mm, but as for other environmental variables, particle size class must be selected based on the cultured species and culture system.
In summary, food quality and availability are among the main factors influencing the fitness and growth of sea cucumbers under controlled conditions. Holothurians include both suspension feeders, able to collect phytoplankton and organic particles from the water column, or, like all European species of commercial interest, deposit feeders, able to ingest large amounts of sediment from the upper layers of the seabed or tank bottom. They are able to extract organic matter from sediment, which is composed of bacteria, microalgae and detritus, thus contributing to the recycling of sediment organic matter and improving sediment oxygenation. In line with these considerations, firsts experiments on the maintenance of sea cucumbers in tank conditions were carried out by feeding adults of H. arguinensis with natural sediment (Domínguez-Godino and González-Wangüemert, 2019a). The positive feeding rates (FRs) and absorption efficiency (AE), despite some variability associated with temperature changes, confirmed the suitability of this method and highlighted the ability of H. arguinensis to actively select organic-rich sediments and increase AE to compensate for the reduction in FR. Based on these encouraging results, further studies were conducted to stimulate sea cucumber growth using natural sediments enriched with macroalgae or seagrasses. Enriched substrates promoted higher growth rates compared with natural treatment, in both adult and juvenile specimens, but with similar survival rates (Rodrigues et al., 2023). In addition, while growth rates were high regardless of the ratio of sand to macrophyte, FR and AE increased with sediment enrichment (Sunde and Christophersen, 2023). Sediments enriched with seagrasses (Cymodocea nodosa and Zostera noltii) showed different performance, probably due to the composition of the seagrass litter (Domínguez-Godino et al., 2020). As deposit-feeding sea cucumbers have low cellulase activity, the high cellulose content typical of some seagrasses might have affected their growth. Sediment enrichment seems to favour the feeding activity providing easily access to food and reducing searching time and energy expenditure. In addition, a lower heterogeneity in growth performance was observed in juveniles when fed with enriched substrates, suggesting that enriched sediments might contrast the typical non-homogeneous growth patterns of sea cucumbers, an issue that could seriously affect their culture, as cohorts tend to reach commercial sizes at different times (Rodrigues et al., 2023).
As sea cucumbers are considered suitable extractive species to be included in integrated multitrophic aquaculture (IMTA) systems, the effect of sediments enriched with waste deriving from the farming system was also tested. Most studies associated sea cucumbers with fish (Sparus aurata, Dicentrarchus labrax, Seriola dumerili, Salmo salar), while only two studies used other invertebrates (the sea urchin Paracentrotus lividus and the mussel Mytilus galloprovincialis) as primary species. The choice to combine sea cucumbers mainly with fish is probably due to the fact that fish farming is the most common aquaculture activity in Europe. In addition, fish farming may impact the surrounding environment due to organic enrichment caused by the accumulation of faeces and uneaten feed (Janssen et al., 2017), conditions that could be favourable for sea cucumber growth. Although fish farming is the most widespread in Europe, mussel farming produces more than 1/3 of aquaculture products, making it one of the most important farming practices also in terms of extension (Avdelas et al., 2021). The high extension makes mussel farms particularly suitable for the development of co-culture with sea cucumbers, as the growth of holothurians is strongly affected by high stocking densities under culture conditions (Grosso et al., 2023). Finally, the development of a co-culture between sea cucumbers and P. lividus, the most prized sea urchin in the Mediterranean Sea, could be a valuable solution to reduce the impact of fishing pressures on both organisms, since P. lividus, like sea cucumbers, is now overexploited in several Mediterranean areas (Grosso et al., 2021; Elmasry et al., 2023). According with the literature, the ability of sea cucumbers to survive and grow under IMTA conditions is species-specific. H. tubulosa showed high survival rates and positive growth with fish, mussels and sea urchins (Tolon et al., 2017a; Grosso et al., 2021, 2023), but also showed signs of disease in highly eutrophic conditions, such as those that may occur near fish cages (Sadoul et al., 2022). Similarly, P. tremulus growth rates were enhanced by the addition of salmon sludge to natural sediments, suggesting the suitability of this species to reduce the impact of salmon farming, which is widespread in northern European countries (Sunde and Christophersen, 2023). Fish waste was also found to be palatable to H. forskali but further studies on growth performance are needed (MacDonald et al., 2013). In contrast, survival and growth of H. poli were generally affected by IMTA conditions, regardless of the primary species associated (Chatzivasileiou et al., 2022; Cutajar et al., 2022a, 2022b; Grosso et al., 2023), and similarly, H. arguinensis showed a reduction in weight, low FRs and negative AE when fed sediment enriched with fish (seabream) waste (Domínguez-Godino and González-Wangüemert, 2019a). The differences observed between species could be caused by several factors, including a different ecological compatibility due to a specific feeding strategy in the wild. For example, H. tubulosa actively selects organic matter from the upper sediment layer, while H. poli, which lives inside the sediment, consumes deeper sediments, which may be characterized by unfavourable physicochemical conditions when affected by waste enrichment, especially if the enrichment is due to fish waste accumulation (Grosso et al., 2023). The responses of sea cucumber to IMTA conditions is also influenced by the feed administered to the associated primary species (Grosso et al., 2021). Finally, stocking density in IMTA systems may also play a fundamental role in driving growth patterns of sea cucumbers. Exceeding the optimal stocking density may improve organic matter bioremediation, but compromise the growth yield of sea cucumbers (Slater and Carton, 2007; Domínguez-Godino and González-Wangüemert, 2018b). However, the survival of sea cucumbers in co-culture with fish or other invertebrates confirmed their compatibility with many of the rearing conditions expected in IMTA systems, and suggest a high potential of these species for the development of sustainable aquaculture project. Finally, as highlighted by Grosso et al. (2021), the use of sea cucumbers in an IMTA would also bring significant economic benefits due to their high market value, especially when combined with other prized species (P. lividus). However, in order to encourage the development of these farming practices, a more comprehensive economic evaluation of the inclusion of sea cucumbers in large-scale IMTA systems would be appropriate.
The number of papers on North-Eastern Atlantic and Mediterranean holothurian aquaculture has increased considerably in recent years, probably due to the growing interest in these organisms for commercial purposes. Most of the studies concern Holothuria tubulosa, H. poli and H. arguinensis and H. mammata, which are the most exported species, while little knowledge is available for the other marketable European sea cucumbers. Experiments revealed species-specific responses to the main environmental parameters (temperature and salinity) and stocking density. Hatchery production protocols are successfully described for all European species except Parastichopus regalis. The settlement stage is considered to be the main bottleneck, while there is still little and fragmentary information on the optimal maintenance conditions for larvae, both in terms of stocking density and food source and feeding regime. The most commonly used method to efficiently feed sea cucumbers, juveniles or adults, is the enrichment of natural sediments by the addition of macroalgae. Several algal species have been tested, but no studies have been conducted to compare the nutritional composition of the algae with the growth performance of sea cucumbers. However, enzymatic activity analysis suggests that protein-rich diets may be most suitable for these organisms. Finally, it has been confirmed that Mediterranean holothurians can be successfully included in multi-trophic aquaculture systems, achieving good survival and growth performance both by in combination with fish and other marine invertebrates. Again, interspecific differences were observed, hence further research is needed. In particular, more information on stocking densities and tolerable organic enrichment is essential to develop and consolidate this farming practice. Finally, there is important to stress the lack of information on the culture of P. regalis, a species that may soon be overexploited, due to its high value.
LC: Investigation, Writing – original draft, Writing – review & editing, Data curation, Conceptualization. GS: Writing – review & editing, Validation, Investigation, Data curation. GC: Writing – review & editing, Visualization, Validation, Formal Analysis. AR: Writing – review & editing, Supervision, Data curation. SV: Conceptualization, Funding acquisition, Supervision, Validation, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by the PON Project (CUP: E86C18001440008), the INSAIL Project (CUP: B66C18000480005) both funded by the Italian Ministry of Education, University, and Research and by the National Inter-University Consortium for Marine Sciences (CoNISMa) through the Terna Elba- Continent project (Monitoring of the abiotic and biotic components of the marine area between the island of Elba and the continental coast).
Project PON Blue Growth (CUP: J89J22001370007) and MIPAAF PO FEAMP 2014-2020 1.39 (CUP: J89J22001370007) of the Italian Ministry of Environment and Energy Security (MASE).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1381836/full#supplementary-material
Avdelas L., Avdic-Mravlje E., Borges Marques A. C., Cano S., Capelle J. J., Carvalho N., et al. (2021). The decline of mussel aquaculture in the European Union: causes, economic impacts and opportunities. Rev. Aquac. 13, 91–118. doi: 10.1111/raq.12465
Aydin M., Aksay K., Karadurmuş U. (2023). Sea cucumber trade and sustainability in Türkiye: progress, problems, and opportunities. Aquat. Sci. Eng. 38, 180–188. doi: 10.26650/ASE20231276000
Aydin M., Sevgili H., Tufan B., Emre Y., Köse S. (2011). Proximate composition and fatty acid profile of three different fresh and dried commercial sea cucumbers from Turkey. Int. J. Food Sci. Technol. 46, 500–508. doi: 10.1111/j.1365-2621.2010.02512.x
Boncagni P., Rakaj A., Fianchini A., Vizzini S. (2019). Preferential assimilation of seagrass detritus by two coexisting Mediterranean sea cucumbers: Holothuria polii and Holothuria tubulosa. Estuar. Coast. SHELF Sci. 231, 106464. doi: 10.1016/j.ecss.2019.106464
Brown N. P., Eddy S. D. (2015). Echinoderm Aquaculture (Hoboken: Wiley Blackwell). doi: 10.1002/9781119005810
Chatzivasileiou D., Dimitriou P. D., Theodorou J., Kalantzi I., Magiopoulos I., Papageorgiou N., et al. (2022). An IMTA in Greece: co-culture of fish, bivalves, and holothurians. J. Mar. Sci. Eng. 10, 776–791. doi: 10.3390/jmse10060776
Conand C. (2018). Tropical sea cucumber fisheries: changes during the last decade. Mar. pollut. Bull. 133, 590–594. doi: 10.1016/j.marpolbul.2018.05.014
Cutajar K., Falconer L., Massa-Gallucci A., Cox R. E., Schenke L., Bardócz T., et al. (2022a). Culturing the sea cucumber Holothuria poli in open-water integrated multi-trophic aquaculture at a coastal Mediterranean fish farm. Aquaculture 550, 737881. doi: 10.1016/j.aquaculture.2021.737881
Cutajar K., Falconer L., Massa-Gallucci A., Cox R. E., Schenke L., Bardócz T., et al. (2022b). Stable isotope and fatty acid analysis reveal the ability of sea cucumbers to use fish farm waste in integrated multi-trophic aquaculture. J. Environ. Manage. 318, 115511. doi: 10.1016/j.jenvman.2022.115511
Dereli H., Aydın M. (2021). Sea cucumber fishery in Turkey: management regulations and their efficiency. Reg. Stud. Mar. Sci. 41, 101551. doi: 10.1016/j.rsma.2020.101551
Domínguez-Godino J. A., González-Wangüemert M. (2018a). Breeding and larval development of Holothuria mammata, a new target species for aquaculture. Aquac. Res. 49, 1430–1440. doi: 10.1111/are.13597
Domínguez-Godino J. A., González-Wangüemert M. (2018b). Does space matter? Optimizing stocking density of Holothuria arguinensis and Holothuria mammata. Aquac. Res. 49, 3107–3115. doi: 10.1111/are.2018.49.issue-9
Domínguez-Godino J. A., González-Wangüemert M. (2019a). Assessment of Holothuria arguinensis feeding rate, growth and absorption efficiency under aquaculture conditions. New Zeal. J. Mar. Freshw. Res. 53, 60–76. doi: 10.1080/00288330.2018.1480499
Domínguez-Godino J. A., González-Wangüemert M. (2019b). Improving the fitness of Holothuria arguinensis larvae through different microalgae diets. Aquac. Res. 50, 3130–3137. doi: 10.1111/are.14267
Domínguez-Godino J. A., Santos T. F., Pereira H., Custódio L., González-Wangüemert M. (2020). Seagrass debris as potential food source to enhance Holothuria arguinensis’ growth in aquaculture. Aquac. Res. 51, 1487–1499. doi: 10.1111/are.14495
Domínguez-Godino J. A., Slater M. J., Hannon C., González-Wangüermert M. (2015). A new species for sea cucumber ranching and aquaculture: breeding and rearing of Holothuria arguinensis. Aquaculture 438, 122–128. doi: 10.1016/j.aquaculture.2015.01.004
Dong Y., Dong S., Tian X., Wang F., Zhang M. (2006). Effects of diel temperature fluctuations on growth, oxygen consumption and proximate body composition in the sea cucumber Apostichopus japonicus Selenka. Aquaculture 255, 514–521. doi: 10.1016/j.aquaculture.2005.12.013
Elmasry E., Abdelrazek F. A., El-Sayed A. F. M. (2023). Growth pattern and population status of the sea urchin Paracentrotus lividus (Lamarck 1816) on the mediterranean coast of Egypt. Egypt. J. Aquat. Res. 49, 409–416. doi: 10.1016/j.ejar.2023.07.001
Galimany E., Baeta M., Ramon M. (2018). Immune response of the sea cucumber Parastichopus regalis to different temperatures: implications for aquaculture purposes. AQUACULTURE 497, 357–363. doi: 10.1016/j.aquaculture.2018.08.005
González-Wangüemert M., Aydin M., Conand C. (2014). Assessment of sea cucumber populations from the Aegean Sea (Turkey): first insights to sustainable management of new fisheries. Ocean Coast. Manage. 92, 87–94. doi: 10.1016/j.ocecoaman.2014.02.014
González-Wangüemert M., Domínguez-Godino J. A., Canovas F., González-Wangüemert M., Domínguez-Godino J. A., Cánovas F. (2018). The fast development of sea cucumber fisheries in the Mediterranean and NE Atlantic waters: from a new marine resource to its over-exploitation. Ocean Coast. Manage. 151, 165–177. doi: 10.1016/j.ocecoaman.2017.10.002
González-Wangüemert M., Valente S., Aydin M. (2015). Effects of fishery protection on biometry and genetic structure of two target sea cucumber species from the Mediterranean Sea. Hydrobiologia 743, 65–74. doi: 10.1007/s10750-014-2006-2
Grosso L., Rakaj A., Fianchini A., Morroni L., Cataudella S., Scardi M. (2021). Integrated Multi-Trophic Aquaculture (IMTA) system combining the sea urchin Paracentrotus lividus, as primary species, and the sea cucumber Holothuria tubulosa as extractive species. Aquaculture 534, 736268. doi: 10.1016/j.aquaculture.2020.736268
Grosso L., Rampacci M., Pensa D., Fianchini A., Batir E., Aydin I., et al. (2023). Evaluating sea cucumbers as extractive species for benthic bioremediation in mussel farms. Sci. Rep. 13, 1457. doi: 10.1038/s41598-023-28733-7
Günay D., Emiroğlu D., Suzer C. (2020). Seasonal variations of digestive enzymes in sea cucumbers (Holothuria tubulosa, G. 1788) under culture conditions. J. Exp. Zool. Part A. Ecol. Integr. Physiol. 333, 144–150. doi: 10.1002/jez.2336
Günay D., Emiroğlu D., Tolon T., Özden O., Saygi H. (2015). Growth and survival rate of juvenile sea cucumbers (Holothuria tubulosa, Gmelin 1788) at various temperatures. Turkish J. Fish. Aquat. Sci. 15, 533–541. doi: 10.4194/1303-2712-v15_2_41
Hudson I. R., Wigham B. D., Solan M., Rosenberg R. (2005). Feeding behaviour of deep-sea dwelling holothurians: inferences from a laboratory investigation of shallow fjordic species. J. Mar. Syst. 57, 201–218. doi: 10.1016/j.jmarsys.2005.02.004
Janssen K., Chavanne H., Berentsen P., Komen H. (2017). Impact of selective breeding on European aquaculture. Aquaculture 472, 8–16. doi: 10.1016/j.aquaculture.2016.03.012
Laguerre H., Raymond G., Plan P., Améziane N., Bailly X., Le Chevalier P. (2020). First description of embryonic and larval development, juvenile growth of the black sea-cucumber Holothuria forskali (Echinodermata: Holothuroidea), a new species for aquaculture in the north-eastern Atlantic. Aquaculture 521, 734961. doi: 10.1016/j.aquaculture.2020.734961
MacDonald C. L. E., Stead S. M., Slater M. J. (2013). Consumption and remediation of European Seabass (Dicentrarchus labrax) waste by the sea cucumber Holothuria forskali. Aquac. Int. 21, 1279–1290. doi: 10.1007/s10499-013-9629-6
Madruga A. S., Félix P. M., Sousa J., Azevedo e Silva F., Brito A. C., Mendes S., et al. (2023). Effect of rearing temperature in the growth of hatchery reared juveniles of the sea cucumber Holothuria arguinensis (Koehler & Vaney 1906). Aquaculture 562, 738809. doi: 10.1016/j.aquaculture.2022.738809
Magdy M., Otero-Ferrer F., de Vicose G. C. (2021). Preliminary spawning and larval rearing of the sea cucumber Holothuria sanctori (Delle Chiaje 1823): a potential aquaculture species. Aquac. Rep. 21, 100846. doi: 10.1016/j.aqrep.2021.100846
Maggi C., González-Wangüemert M. (2015). Genetic differentiation among Parastichopus regalis populations in the Western Mediterranean Sea: potential effects from its fishery and current connectivity. Mediterr. Mar. Sci. 16, 489–501. doi: 10.12681/mms.1020
Mezali K., Thandar A. S. (2014). First record of Holothuria (Roweothuria) arguinensis (Echinodermata: Holothuroidea: Aspidochirotida: Holothuriidae) from the Algerian coastal waters. Mar. Biodivers. Rec. 7, 1–4. doi: 10.1017/S1755267214000438
Moussa R. M., Nasef L. E. S., Abdel-Latif H. (2019). Innovative approach of asexual reproduction induction in Holothuria polii (Delle Chiaje 1824). Turkish J. Fish. Aquat. Sci. 19, 585–592. doi: 10.4194/1303-2712-v19_7_05
Purcell S. W., Conand C., Hamel J. F., Toral-Granada M. V., Lovatelli A., Uthicke S. (2013). Sea cucumber fisheries: global analysis of stocks, management measures and drivers of overfishing. Fish Fish. 14, 34–59. doi: 10.1111/j.1467-2979.2011.00443.x
Purcell S. W., Hair C. A., Mills D. J. (2012). Sea cucumber culture, farming and sea ranching in the tropics: progress, problems and opportunities. Aquaculture 368–369, 68–81. doi: 10.1016/j.aquaculture.2012.08.053
Purcell S. W., Lovatelli A., González-Wangüermert M., Solis-Marin F. A., Samyn Y., Conand C. (2023). Commercially important sea cucumbers of the world. 2nd ed. (Rome: FAO).
Rakaj A., Fianchini A. (2024). “Mediterranean sea cucumbers - Biology, ecology, and exploitation,” in The World of Sea Cucumbers: Challenges, Advances, and Innovations. Eds. Mercier A., Hamel J. F., Suhrbier A., Pearce C. (London: Academic Press), 753–773.
Rakaj A., Fianchini A., Boncagni P., Lovatelli A., Scardi M., Cataudella S. (2018). Spawning and rearing of Holothuria tubulosa: a new candidate for aquaculture in the Mediterranean region. Aquac. Res. 49, 557–568. doi: 10.1111/are.2018.49.issue-1
Rakaj A., Fianchini A., Boncagni P., Scardi M., Cataudella S. (2019). Artificial reproduction of Holothuria polii: a new candidate for aquaculture. Aquaculture 498, 444–453. doi: 10.1016/j.aquaculture.2018.08.060
Ramofafia C., Byrne M., Battaglene S. C. (2003). Development of three commercial sea cucumbers, Holothuria scabra, H. fuscogilva and Actinopyga mauritiana: larval structure and growth. Mar. Freshw. Res. 54, 657–667. doi: 10.1071/MF02145
Ramón M., Amor M. J., Galimany E. (2022). Reproductive biology of the holothurian Parastichopus regalis in the Mediterranean Sea and its implications for fisheries management. Fish. Res. 247, 106191. doi: 10.1016/j.fishres.2021.106191
Ramón M., Simarro G., Galimany E., Lleonart J. (2019). Evaluation of sediment particle size selection during feeding by the holothurian Parastichopus regalis (Cuvier 1817). Reg. Stud. Mar. Sci. 31, 100763. doi: 10.1016/j.rsma.2019.100763
Rodrigues N. V. (2012). New geographic distribution records for Northeastern Atlantic species from Peniche and Berlengas Archipelago. Life Mar. Sci. 29, 1–4.
Rodrigues T., Azevedo e Silva F., Sousa J., Félix P. M., Pombo A. (2023). Effect of enriched substrate on the growth of the sea cucumber Holothuria arguinensis Koehler and Vaney 1906 juveniles. Diversity 15, 458–468. doi: 10.3390/d15030458
Roggatz C. C., González-Wangüemert M., Pereira H., Vizetto-Duarte C., Rodrigues M. J., Barreira L., et al. (2018). A first glance into the nutritional properties of the sea cucumber Parastichopus regalis from the Mediterranean Sea (SE Spain). Nat. Prod. Res. 32, 116–120. doi: 10.1080/14786419.2017.1331224
Sadoul B., Caprioli J.-P., Barrier-Loiseau C., Cimiterra N., Laugier T., Lagarde F., et al. (2022). Is Holothuria tubulosa the golden goose of ecological aquaculture in the Mediterranean Sea? Aquaculture 554, 738149. doi: 10.1016/j.aquaculture.2022.738149
Santos R., Dias S., Pinteus S., Silva J., Alves C., Tecelão C., et al. (2016). Sea cucumber Holothuria forskali, a new resource for aquaculture? Reproductive biology and nutraceutical approach. Aquac. Res. 47, 2307–2323. doi: 10.1111/are.2016.47.issue-7
Schagerström E., Christophersen G., Sunde J., Bakke S., Matusse N. R., Dupont S., et al. (2022). Controlled spawning and rearing of the sea cucumber, Parastichopus tremulus. J. World Aquac. Soc 53, 224–240. doi: 10.1111/jwas.12816
Slater M. J., Carton A. G. (2007). Survivorship and growth of the sea cucumber Australostichopus (Stichopus) mollis (Hutton 1872) in polyculture trials with green-lipped mussel farms. Aquaculture 272, 389–398. doi: 10.1016/j.aquaculture.2007.07.230
Sousa J., Félix P. M., Brito A. C., Venâncio E., Azevedo e Silva F., Simões T., et al. (2021). The effects of stocking density on physiological traits in Holothuria forskali broodstock. Aquac. Res. 52, 1548–1560. doi: 10.1111/are.15007
Sousa J., Félix P. M., Brito A. C., Venâncio E., Azevedo e Silva F., Simões T., et al. (2023). Optimizing growth and rearing techniques for larvae and juveniles of the sea cucumber Holothuria arguinensis. Diversity 15, 722–742. doi: 10.3390/d15060722
Sunde J., Christophersen G. (2023). Appetite in captivity - feeding studies of the red sea cucumber Parastichopus tremulus. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1052968
Tolon T. (2017). Effect of salinity on growth and survival of the juvenile sea cucumbers Holothuria tubulosa (Gmelin 1788) and holothuria poli (Delle chiaje 1923). FRESENIUS Environ. Bull. 26, 163–171. doi: 10.11598/btb.2019.26.3.1041
Tolon T., Emiroğlu D., Günay D., Hancı B. (2017b). Effect of stocking density on growth performance of juvenile sea cucumber Holothuria tubulosa (Gmelin 1788). Aquac. Res. 48, 4124–4131. doi: 10.1111/are.2017.48.issue-8
Tolon M. T., Emiroglu D., Gunay D., Ozgul A. (2017a). Sea cucumber (Holothuria tubulosa Gmelin 1790) culture under marine fish net cages for potential use in integrated multi-trophic aquaculture (IMTA). Indian J. Geo-Marine Sci. 46, 749–756.
Tolon T., Emiroğlu D., Günay D., Saygi H. (2015). Effect of sediment grain size on growth performance of juvenile sea cucumber (Holothuria tubulosa). Turkish J. Fish. Aquat. Sci. 15, 555–559.
Toscano A., Cirino P. (2018). First evidence of artificial fission in two Mediterranean species of Holothurians: Holothuria tubulosa and Holothuria polii. TURKISH J. Fish. Aquat. Sci. 18, 1141–1145. doi: 10.4194/1303-2712-v18_10_01
Venâncio E., Félix P. M., Brito A. C., Sousa J., Azevedo e Silva F., Simões T., et al. (2021). Do broodstock diets influence viability and larval development of Holothuria mammata? Aquaculture 536, 1–11. doi: 10.1016/j.aquaculture.2021.736431
Ventura D., Grosso L., Pensa D., Casoli E., Mancini G., Valente T., et al. (2023). Coastal benthic habitat mapping and monitoring by integrating aerial and water surface low-cost drones. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1096594
Yang H., Yuan X., Zhou Y., Mao Y., Zhang T., Liu Y. (2005). Effects of body size and water temperature on food consumption and growth in the sea cucumber Apostichopus japonicus (Selenka) with special reference to aestivation. Aquac. Res. 36, 1085–1092. doi: 10.1111/j.1365-2109.2005.01325.x
Keywords: holothurian, echinoderm, Trepang, IMTA, co-culture
Citation: Ciriminna L, Signa G, Cilluffo G, Rakaj A and Vizzini S (2024) Aquaculture of emerging species in North-Eastern Atlantic and Mediterranean Sea: a systematic review on sea cucumber farming and potential development. Front. Mar. Sci. 11:1381836. doi: 10.3389/fmars.2024.1381836
Received: 04 February 2024; Accepted: 04 March 2024;
Published: 21 March 2024.
Edited by:
Miquel Planas, Spanish National Research Council (CSIC), SpainReviewed by:
Adriana Giangrande, University of Salento, ItalyCopyright © 2024 Ciriminna, Signa, Cilluffo, Rakaj and Vizzini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Arnold Rakaj, YXJub2xkLnJha2FqQHVuaXJvbWEyLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.