 Yuka Inagaki1†
Yuka Inagaki1† Yoshiyuki Ishitani2Akihiro Tame3
Yoshiyuki Ishitani2Akihiro Tame3 Katsuyuki Uematsu4
Katsuyuki Uematsu4 Naotaka Tomioka5Takayuki Ushikubo5
Naotaka Tomioka5Takayuki Ushikubo5 Yurika Ujiié1,6*
Yurika Ujiié1,6*- 1Department of Biology, Kochi University, Kochi, Japan
- 2Institute for Extra-Cutting-Edge Science and Technology Avant-garde Research (X-star), Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Japan
- 3School of Medical Sciences, University of Fukui, Fukui, Japan
- 4Department of Marine and Earth Sciences, Marine Works Japan Ltd, Yokosuka, Japan
- 5Kochi Institute for Core Sample Research, X-Star, JAMSTEC, Nankoku, Japan
- 6Marine Core Research Institute, Kochi University, Nankoku, Japan
The increase discharge of titanium dioxide (TiO2) nanoparticles, derived from engineered material waste, exerts a detrimental impact on both the marine ecosystem and public health. The cytotoxicity of TiO2 nanoparticles on marine organisms should be imperatively understood to tackle the urgent concern for the well-being of marine life. Various concentrations of TiO2 nanoparticles have proven to reach fatal levels in aquatic organisms, requiring a deeper exploration of cytotoxicity. Notably, certain benthic foraminifers, such as Ammonia veneta, have been identified as capable of incorporating TiO2 nanoparticles into vesicles. However, these organisms exhibit a detoxification mechanism through exocytosis, as indicated by previous transcriptomic inferences. This presents the advantage of assessing the tolerance of foraminifers to TiO2 nanoparticles as pollutants and investigating the long-term effects of cytotoxicity. In this study, we scrutinized the distribution of TiO2 nanoparticles within cells and the growth rates of individuals in seawater media containing 1, 5, 10, and 50 ppm TiO2 nanoparticles, comparing the results with a control group over a 5-week period, utilizing A. veneta stain. Transmission electron microscopy observations consistently revealed high concentrations of TiO2 nanoparticles in vesicles, and their expulsion from cells was evident even with exposure to 5 ppm TiO2 nanoparticles. Under the control and 1 ppm TiO2 conditions, foraminifers increased their cell volume by adding a calcification chamber to their tests every 1 or 2 days. However, the 5-week culturing experiments demonstrated that foraminifers gradually ceased growing under 5 ppm TiO2 nanoparticle exposure and exhibited no growth at > 10 ppm concentrations, despite an ample food supply. Consequently, these findings with A. veneta suggest that the foraminiferal detoxification system could be disrupted by concentrations exceeding 5 ppm of TiO2 nanoparticles. The toxic effect of TiO2 nanoparticles on meiofauna, such as benthic foraminifers, have been poorly understood, though these organisms play an important role in the marine ecosystem. Environmental accumulation of TiO2 nanoparticles on the coast has already exceeded twenty times more than foraminiferal detoxification level. Future studies focusing on toxic mechanism of TiO2 nanoparticles are crucial to prevent the breakdown of the marine ecosystem through accelerating discharge of TiO2 nanoparticles into the ocean.
1 Introduction
Titanium dioxide nanoparticles (TiO2 NPs) stand as the second most widely employed engineered nanomaterial globally, owing to their advantageous physicochemical properties, notably their photocatalytic capability for the oxidative decomposition of organic matter (Zheng and Nowack, 2021). In recent years, these nanoparticles have found application in sunscreen filters, replacing oxybenzone and octinoxate, to safeguard marine ecosystems (Ammendolia et al., 2022). The escalating demand for TiO2 NPs has led to the discharge of manufacturing waste into the environment–specifically, soil and groundwater–eventually culminating in the ocean as the ultimate repository. NPs commonly form aggregates in water and sink down to the bottom (Boxall et al., 2007). Indeed, the accumulation of TiO2 NPs on European coasts was projected to reach up to 123 ppm in 2020, and the current trend indicates an acceleration (Sun et al., 2016; Zheng and Nowack, 2021). This accumulation in the marine environment raises concerns due to its toxicity to aquatic organisms (Moore, 2006; Farré et al., 2009; Delay and Frimmel, 2012; Matranga and Corsi, 2012). Particularly, aggregated nanoparticles are accumulated in the sediments and interacted with sediment-dwelling organisms due to their low mobility (Farré et al., 2009). Experimental studies have previously demonstrated the toxicity of TiO2 NPs to aquatic organisms, particularly microalgae and macroorganisms (Menard et al., 2011; Minetto et al., 2014; Luo et al., 2020). These studies have assessed the toxic effects through cell stress assays, revealing a Median Effect Concentration (EC50) of 35.3 ppm for algae and a Median Lethal Concentration (LC50) of 5.5–20000 ppm for Daphnia magna. Given the observed variability in lethal effects among organisms in exposure experiments with viability examinations, it becomes imperative to extend studies to benthos, encompassing not only macro or microfauna but also benthic foraminifers, to comprehensively understand the impact of TiO2 NPs on marine ecosystems.
Benthic foraminifers, ubiquitous unicellular meiofauna on the seafloor, particularly in coastal areas (Culver and Buzas, 1999), serve as biomonitoring targets to assess the effects of NPs on marine environments (Ciacci et al., 2019). Foraminifers construct a test using calcium carbonate, sediment particles, or an organic membrane (Goldstein, 1999), with the test growing larger by adding chambers. Recent studies utilizing single-cell transcriptome analysis and cellular observations from a time-course experiment with the benthic foraminifers Ammonia species exposed to 1 ppm TiO2 NPs for 24 hours have elucidated the cytotoxic mechanism of TiO2 NPs (Ciacci et al., 2019; Ishitani et al., 2023). Transcriptome analysis with Ammonia veneta successfully unveiled the metabolic pathway associated with TiO2 NPs toxicity and its time-series changes (Ishitani et al., 2023). Foraminifers demonstrated phagocytosis of TiO2 NPs into their cells, manifesting stress indicators such as the production of reactive oxygen species (ROS) and peroxidized lipids. Intriguingly, they exhibited detoxification, wherein aggregated TiO2 NPs were encapsulated in ceramide and expelled as waste within 24 hours. This unique detoxification system enables foraminifers to survive in polluted environments, though the long-term cytotoxicity remains unknown. Furthermore, the limitation of TiO2 NPs concentration that regulates foraminiferal growth or lethality has been never examined. In the other experiments with unicellular diatom, their EC50 and LC50 are ~10 and ~70 ppm of TiO2 NPs, which are ~20–30 nm in diameter (Hartmann et al., 2010; Clément et al., 2013). Understanding such limitation becomes crucial in light of the rapid accumulation of TiO2 NPs in marine environments in recent times.
In this study, we investigated the threshold limits of TiO2 NPs concentrations for the survival of the benthic foraminifer A. veneta. The foraminifer was cultured for five weeks with exposures to 1, 5, 10, and 50 ppm TiO2 NPs, and the potential for long-term survival was assessed based on individual growth rates, evaluated through the increased number of chambers, and the distribution of intracellular TiO2 NPs. Our investigations suggest that foraminiferal growth is gradually inhibited in rising of TiO2 NPs concentration most likely due to the breakdown of detoxification.
2 Materials and methods
2.1 TiO2 NPs – source, characterization, and preparation
AEROXIDE®TiO2, a mixture of rutile and anatase crystals with a nominal diameter of 25 nm (Evonik, Germany), was utilized in the present study. The stock suspension (1000 mg/L) of TiO2 NPs was prepared using AEROXIDE®TiO2 powder in Milli-Q water and sonicated using an NR-50M ultrasonic homogenizer (Microtec, Japan) at 50 W for 60 min with 2 s on/off cycles placed in ice to prevent aggregation. Stock suspensions were shielded from light and stored at -20°C. For each experiment, the thawed stock suspension was similarly sonicated for approximately 60 min to disperse the TiO2 particles, which tend to clump due to their cohesive nature, bringing them to a particle size of 25 nm. The resulting dispersion was diluted with artificial sea water (ASW), which was filtered through a 0.2µm mesh membrane filter (ADVANTEC®, Japan) of 35ppt Coral Pro Salt (Red Sea, USA) in Milli-Q water, according to the experimental conditions.
2.2 Culture experiments of foraminiferal strain Ammonia veneta
The study employed specimens derived from the culture strain A. veneta (Ishitani et al., 2023), which has been maintained in ASW at 23°C under a 12:12 hour light/dark cycle with weekly feedings of fresh Dunaliella salina (NIES-2257). This species forms a calcium carbonate test, which is sequentially increased by the maturity phase prior to reproduction. Adult (ca. 10 chambers) specimens of A. veneta showing reticulopodia extension and activity were randomly selected from subcultures, and three individuals were transferred to each petri dish (Aznor Petri Dish φ55 × 17 mm Azwan, Japan) containing 10 mL of either TiO2 NPs seawater medium or ASW (control). For each medium condition, three replicated petri dishes were prepared and incubated at 23°C under a 12:12 h light/dark cycle for five weeks. All cultures per experiment were run in parallel.
Four different experimental conditions were set for TiO2 NPs concentrations, as well as the frequency of medium changes, feeding frequency, and amount of food (Table 1). In both culture experiments 1 and 2, TiO2 NPs seawater media were set up at 1, 10, and 50 ppm, in addition to the control. The frequency of medium changes differed between culture experiments 1 and 2: once a week in Experiment 1 and twice a week in Experiment 2. At each time, 80 µL of food was added to each petri dish. In culture experiment 3, TiO2 NPs seawater media were set up at 1, 5, and 10 ppm, in addition to the control, with weekly medium changes and with feeding 40 µL of food and increased feeding to a total of 80 µL per week through addition of 20 µL food every few days. Culture experiment 4 involved TiO2 NPs seawater media at 5 and 10 ppm, in addition to the control, with weekly medium changes with 80 µL of food and increased feeding to a total of 160 µL per week through addition of 40 µL food every few days. In experiments 2 and 4, the number of feedings and the amount of food were increased to keep double the amount of fresh D. salina in the petri dishes per week (Table 1).
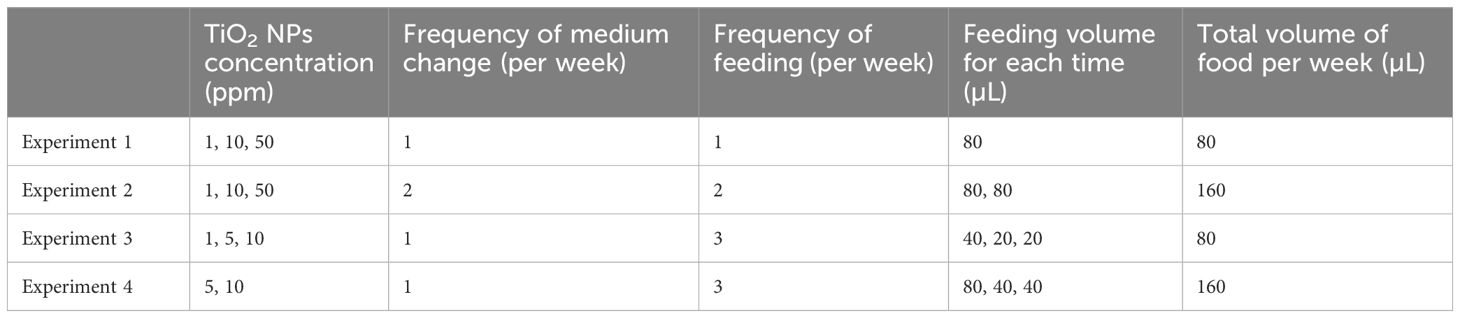
Table 1 Conditions of foraminiferal culture experiments.
At the beginning of culture experiments, we photographed each individual using an inverted research microscope (IX73 Olympus, Japan) with a microscope digital camera (DS-Ri2 Nikon Japan) and assigned an individual identification number. We photographed each of all studied individual weekly to check for signs of life or death, regarding the observations: extension and movement of reticulopodia and the movement of the individual to escape from light emission of the inverted microscope for a few minutes. Specimens, which lacked these three observation points, were categorized as dead. Individual growth was evaluated by the number of chamber increases from the previous week.
We firstly tested normality and variances for total numbers of increased chambers for five weeks at each culture experimental condition with Kolmogorov-Smirnov and F test in R v.4.0.2 (R Core Team, 2021), respectively. According to the normality and variances, we conducted the Student-t and Man-Whitney-U tests in R v.4.0.2. The p values of multiple comparisons were adjusted with the Bonferroni correction.
2.3 Culture experiment of Dunaliella salina
We independently conducted a culture experiment of D. salina, a food source for foraminifers, to examine cell divisions in TiO2 NPs seawater medium. We prepared seven replicates for each of the 10 mL culture media with 1, 5, and 10 ppm TiO2 NPs and ASW (control) in petri dishes and added 80 µL of D. salina to each of the 28 dishes. During the 6-day culture experiments under the same conditions as foraminiferal experiments, we examined the cell density of D. salina in each of the four different media daily. All 10 mL medium was transferred into a 15 mL centrifuge tube, and the petri dish was rinsed with 3 mL ASW to recover all remaining cells. After centrifugation at 3,300 rpm for 10 min at 23°C using a micro high-speed centrifuge (CF 16RX II Hitachi, Japan), the pellet of D. salina and TiO2 NPs was suspended with 30 µL of ASW and transferred to a 1.5 ml tube. We immediately added 0.5% xanthan gum solution to a final concentration of 0.01% to inhibit the motility of D. salina. Three replicates were made, each of which was mounted with 10 µL of cell suspension to a cell counter (AS ONE, Japan). Following the manufacturer’s protocol of the cell counter, we counted the number of cells derived from the control and 1 ppm samples using a stereomicroscope (SMZ18 Nikon, Japan) equipped with an inclined-angle lens tube. As TiO2 NPs aggregations in the 5 and 10 ppm overlapped on D. salina and avoided observation, we captured the autofluorescence of D. salina with a digital camera (DS-Ri2 Nikon, Japan) using fluorescence observation (U-HGLGPS Olympus Japan, Excitation filter: BP530-550, Barrier filter: BA575IF) equipped with an inverted research microscope (IX73 Olympus, Japan) and counted the cell numbers.
2.4 Scanning electron microscope observation of foraminiferal tests
We used six specimens: two for control and four for the 10 ppm TiO2 NPs treatment. Two out of four 10 ppm TiO2 NPs treated specimens formed a new chamber during the culture experiment, while the other two showed no growth. Each test was cleaned with a brush in water and dried in air. These tests were placed on carbon tape and glued to a stage where carbon deposition was performed with the carbon coater (JEE-420 JEOL, Japan). The surface structure and element analysis of foraminiferal tests were conducted with a field emission SEM equipped with an energy dispersive X-ray spectrometer (EF-SEM-EDS) (JSM-6500F JEOL, Japan).
2.5 Transmission electron microscope observation of cells
Ammonia veneta specimens, cultured in the 5 ppm TiO2 NPs medium, were fixed with 2.5% glutaraldehyde in filtered artificial sea water (FASW) for longer than 24 hours at 4°C. The fixed specimens were embedded in 1% aqueous agarose and then decalcified with 0.2% EGTA in 0.81 mol/L aqueous sucrose solution (pH 7.0) for several days. The specimens were rinsed with FASW, then postfixed with 2% osmium tetroxide in FASW for 2 hours at 4°C, dehydrated in a graded ethanol series, and embedded in epoxy resin (Quetol 651). Ultrathin sections (100 nm for observation and 200 nm for analysis) were cut using a diamond knife on an Ultracut S ultramicrotome (Leica Reichert, Germany), stained with 2% aqueous uranyl acetate and lead staining solution (0.3% lead nitrate and 0.3% lead acetate), and examined using TEM equipped with an energy dispersive X-ray spectrometer (TEM-EDS) (FEI Tecnai G2 20, USA) operated at 200 kV. Elemental mapping was performed in scanning TEM (STEM) mode.
3 Results
3.1 Cell observation and viability of Ammonia veneta
Living specimens of A. veneta are filled with cytoplasm inside the test and extend fine reticulopodia radially from the aperture and pores of the test to capture food particles in their surroundings (Figure 1). In 1 ppm TiO2 NPs seawater medium, TiO2 NPs aggregate up to ~2 µm within 24 hours and accumulate with other wastes on the bottom (Ishitani et al., 2023). TiO2 NPs is aggregating faster in high TiO2 NPs concentration: over 1 µm in hydrodynamic diameter within 1 hour in 5 and 10 ppm TiO2 NPs seawater and over 15 µm in 50 ppm TiO2 NPs seawater (Morelli et al., 2018; Reyes et al., 2021; Palmeira-Pinto et al., 2023). These aggregates visibly settled at the bottom of the petri dish as brownish substances during the culture experiment with TiO2 NPs seawater media (Figure 1). However, these depositions were not observed in the surrounding areas of living A. veneta, particularly around the reticulopodia extensions (Figure 1). At the onset of the culture experiment, all studied specimens extended fine reticulopodia. In the control and 1 ppm TiO2 NPs seawater medium, specimens showed reticulopodia activity and also added new chambers and grew cytoplasm, which typically filled the interior of the test. Under TiO2 NPs seawater media concentrations greater than 10 ppm, reticulopodia were retracted, and cytoplasm continuously shrank (Figures 2A–D). After reduction of cytoplasm, any reticulopodia activity and movement of specimens were not observed. In these conditions, aggregations containing TiO2 NPs were observed around foraminiferal specimens.
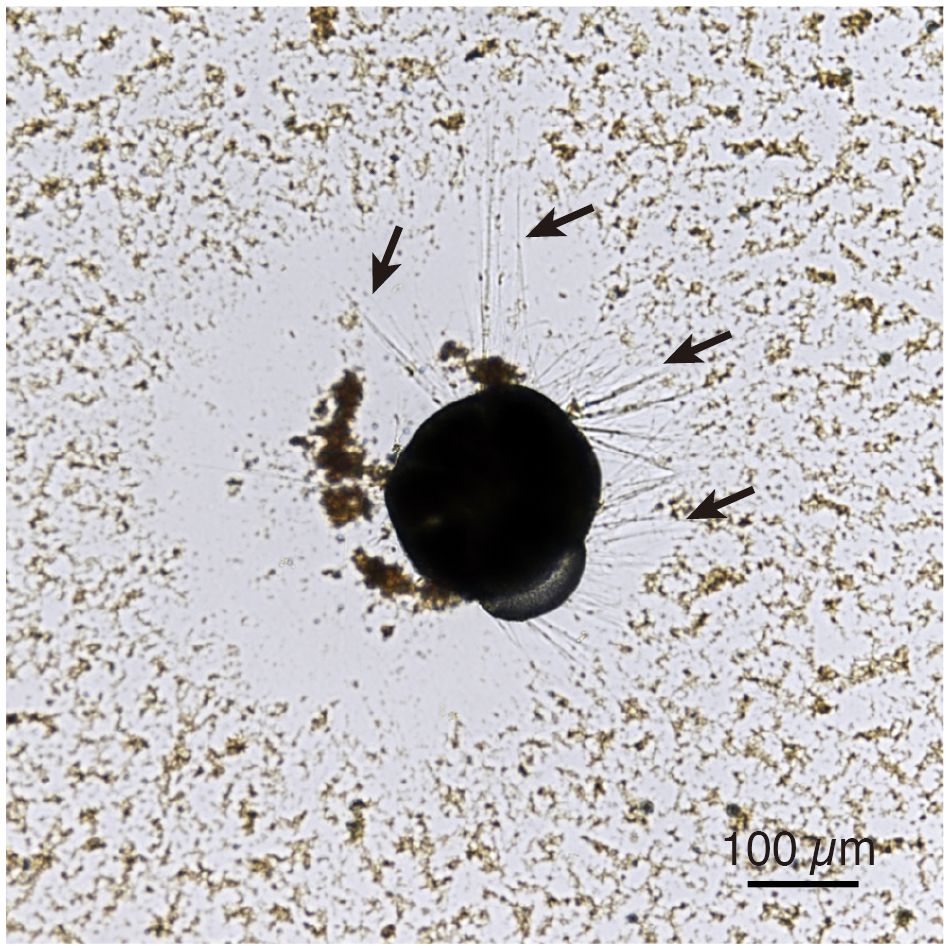
Figure 1 Living Ammonia veneta specimen in 10 ppm TiO2 medium. The dark part is cytoplasm inside the test. Fine fibrous reticulopodia, as shown by black arrows, are extended from the cytoplasm in all directions. The brown substance is an aggregation of TiO2 NPs.
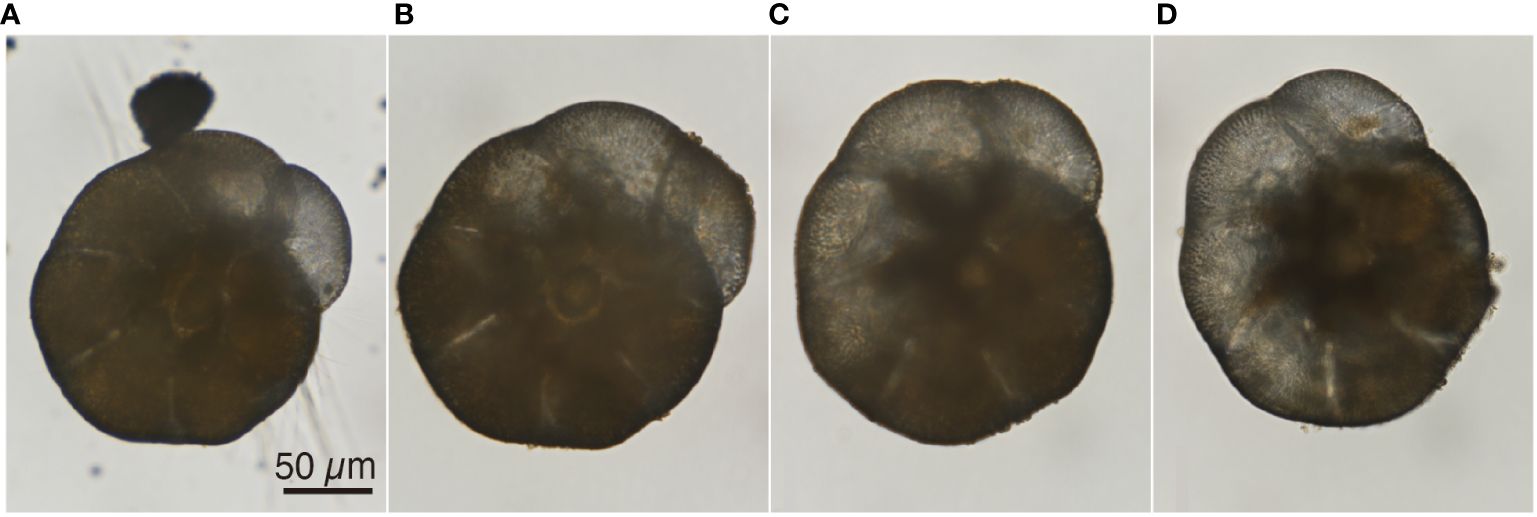
Figure 2 Light microscopy images of an Ammonia veneta specimen exposed to 10 ppm TiO2 medium for four weeks. (A) Reticulopodia were extended from the cytoplasm at the onset of experiment. (B) There were no reticulopodia, and cytoplasm was reduced after a week. (C) Cytoplasm was retracted from the previous week. (D) Cytoplasm completely shrank in the fourth week.
Holes appeared on the surface of the tests in some specimens exposed to TiO2 NPs seawater media concentrations higher than 10 ppm (Figure 3). After a 5-week experiment, additional holes were observed on the umbilical side of the test.

Figure 3 Light microscopy image of the umbilical side of an Ammonia veneta specimen exposed to 50 ppm TiO2 medium for five weeks. Three big holes were opened on the surface of the test, as shown by the black arrows.
3.2 Foraminifera culture experiments
In Experiment 1, control specimens increased by an average of ~2 chambers every week, reaching a total of ~9 chambers over five weeks (Figure 4A; Supplementary Table S1). One out of nine control specimens reproduced in the third week. By the end of the 5-week experiment, three specimens had reproduced clones, forming an average of ~20 chambers during their entire lifespan. In the 1 ppm TiO2 treatment, eight out of nine specimens increased by ~1 chamber per week, although five specimens died after 3 and 5 weeks, respectively. In the 10 ppm TiO2 treatment, six out of nine specimens increased by ~1 chamber per week, but all specimens ceased growth by the third week and died by the fourth week. In the 50 ppm TiO2 treatment, only one specimen increased by 1 chamber in the first week, but all specimens died by the fourth week. The total number of increased chambers over the 5-week experiment showed a significant difference between the control specimens and the 10 and 50 ppm TiO2 treated specimens (Figure 4B).
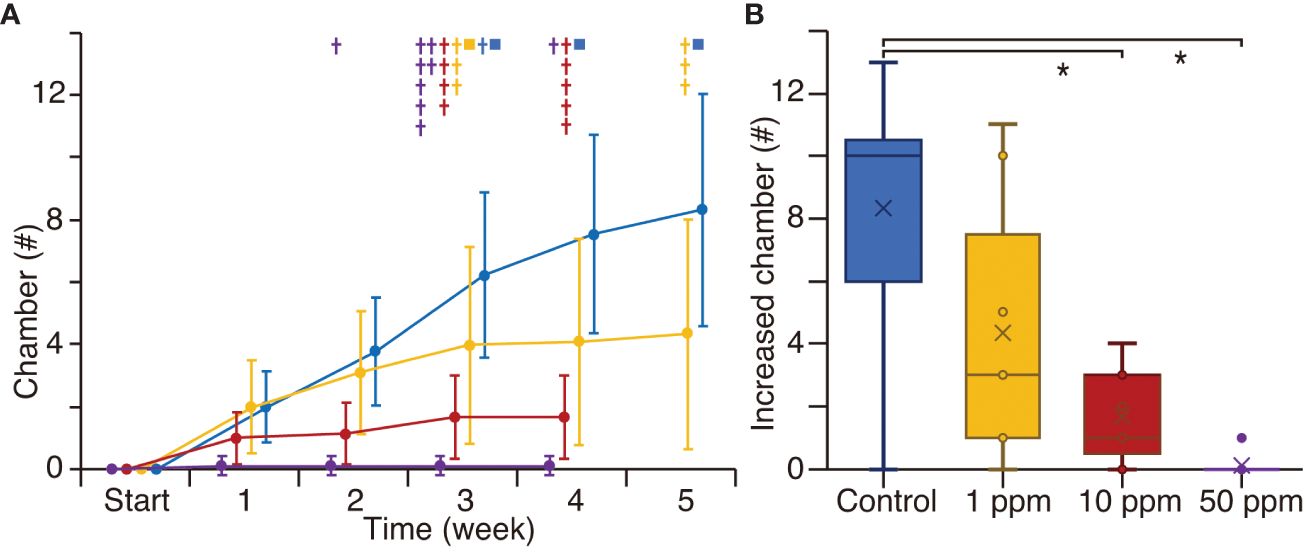
Figure 4 The number of increased chambers in culture experiment 1. Blue, yellow, red, and purple colors show the control, 1 ppm, 10 ppm, and 50 ppm TiO2 treatments, respectively. (A) The average of additional numbers of chambers every week during the 5-week experiment. Crosses indicate dead specimen and squares show clonal reproduction. (B) Box plot of the number of increased chambers for five weeks. Asterisks show significant differences (p < 0.0125 with the Bonferroni correction).
In Experiments 2, all specimens, both in the control and the 1 ppm TiO2 treatment, increased by 6–8 chambers over 5 weeks (Figure 5A; Supplementary Table S1). During the experiment, four and five specimens reproduced clones in the control and the 1 ppm TiO2 treatment, respectively. However, in both the 10 and 50 ppm TiO2 treatments, all specimens showed no growth and died by the fourth week. The total number of increased chambers over the 5-week experiment was the same between the control and the 1 ppm TiO2 treated specimens, but there were significant differences between the controls and the 10 and 50 ppm TiO2 treatments (Figure 5B).
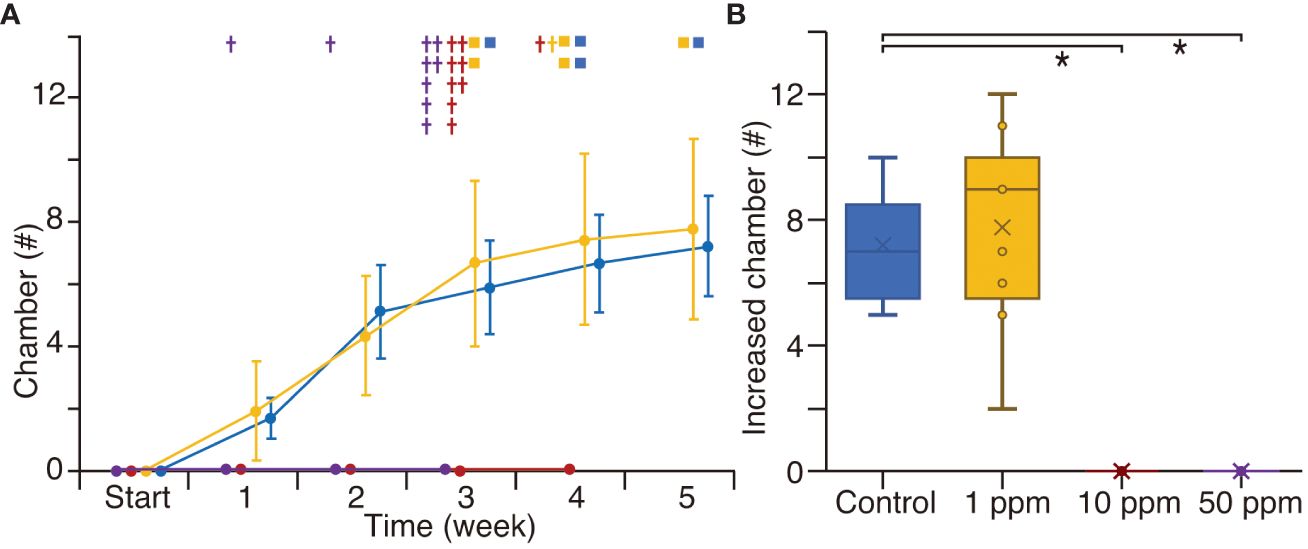
Figure 5 The number of increased chambers in the culture experiment 2. The manner of plots is the same as the in Figure 4.
In Experiment 3, all specimens, both in the control and the 1 ppm TiO2 treatment, increased by ~8 chambers over 5 weeks (Figure 6A; Supplementary Table S1). During this experiment, one specimen reproduced clones in each of the control and the 1 ppm TiO2 treatments, but some specimens died. In the 5 ppm TiO2 treatment, three out of nine specimens increased by ~1 chamber per week, but all had stopped growing and died by the third week. In the 10 ppm TiO2 treatment, all individuals showed no growth and died by the third week. The total number of increased chambers over the 5-week experiment was the same between the control and the 1 ppm TiO2 treated specimens, but there were significant differences between the controls and the 5 and 10 ppm TiO2 treatments (Figure 6B).
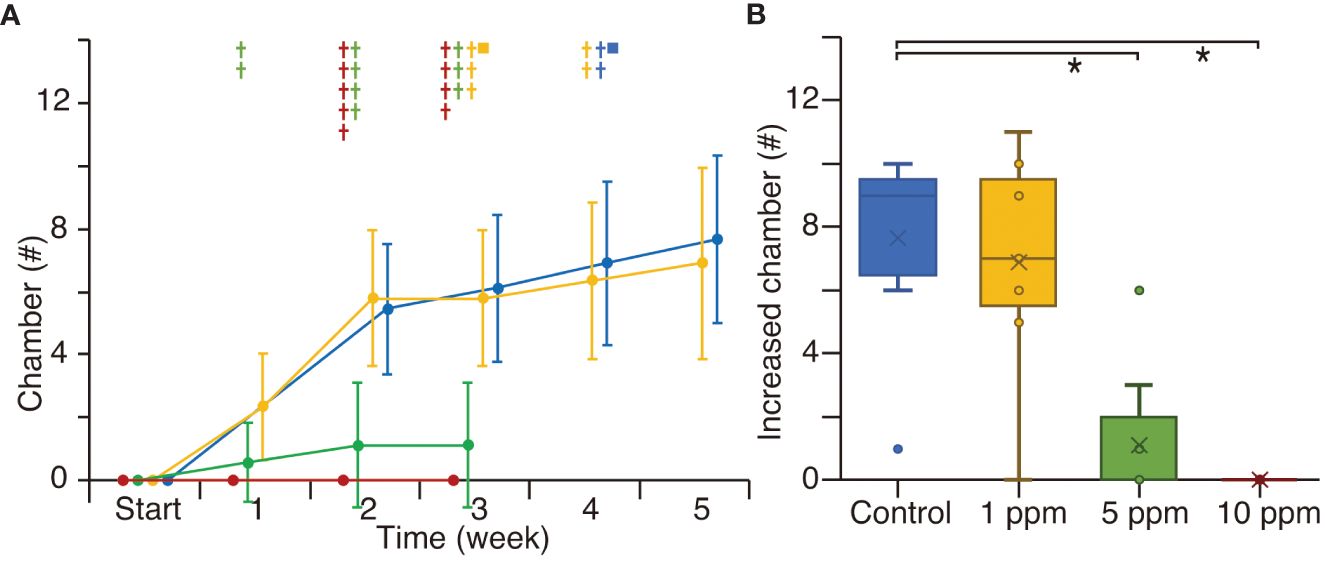
Figure 6 The number of increased chambers in the culture experiment 3. Green color indicates the 5 ppm TiO2 treatment. The other manners of plots are same as Figure 4. The control data for the first week was absent.
In Experiment 4, eight out of nine control specimens increased by ~8 chambers over 5 weeks (Figure 7A; Supplementary Table S1). At the end of the experiment, three specimens reproduced clones. In the 5 ppm TiO2 treatment, four out of nine specimens increased by ~1 chamber per week, but they stopped growing by the third week and all died by the fifth week. In the 10 ppm TiO2 treatment, only two specimens increased by one chamber in the first week, but all individuals died by the third week. The total number of increased chambers over the 5-week experiment showed significant differences between the controls and the 5 and 10 ppm TiO2 treatments, as well as those of Experiment 3 (Figure 7B).
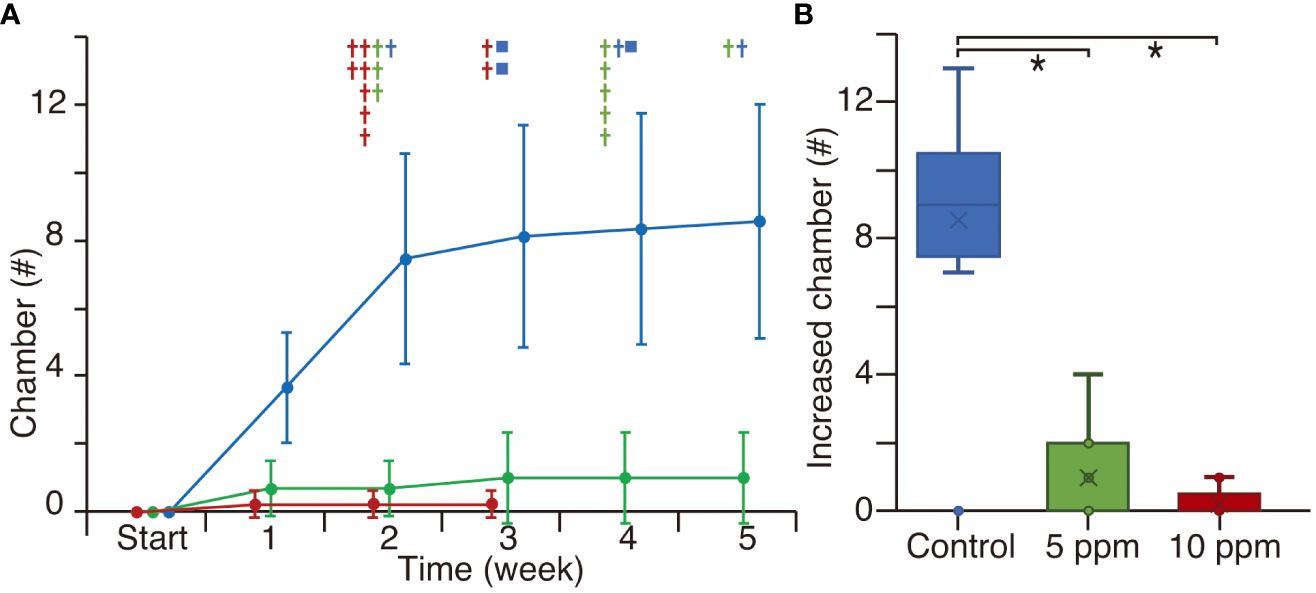
Figure 7 The number of increased chambers in culture experiment 4. Blue, green, and red show the control, 5 ppm, and 10 ppm TiO2 treatments, respectively. The other manners of plots are the same as in Figure 4. Significant differences are p < 0.0167 with the Bonferroni correction.
3.3 Surface structure of Ammonia veneta test
Morphological differences between control and 10 ppm TiO2 treated specimens were observed on the surface of the tests, but pores were clearly opened on both specimens (Figures 8A–D; Supplementary Figure S1). The test surface was smooth on control specimens but slightly rough on 10 ppm TiO2 treated specimens (Figures 8B, D). Moreover, some adhesive materials were observed on the test surface of TiO2 treated specimens, even when cleaned with brushes. These adhesive materials were often observed from the 10 ppm TiO2 treated specimens, which showed no growth during the culture experiment (Supplementary Figure S1). The SEM-EDS mapping showed a trace amount of titanium from adhesive materials and on the surface of the test but not covering the aperture and pores of the test (Figures 8E, F).

Figure 8 SEM images and EDS elemental map. (A) SEM image of the control specimen. (B) The surface test structure of the control specimen. (C) SEM image of a specimen cultured in 10 ppm TiO2 seawater medium. (D) The surface test structure of the 10 ppm TiO2 treated specimen. (E) SEM image around a last chamber, which was formed during a culture experiment in 10 ppm TiO2 medium. (F) EDS elemental map showed the distribution of titanium on the surface of the test (E).
3.4 Increase of Dunaliella salina cell density
Time-series changes in D. salina cell density were recorded for each culture condition, including control and 1, 5, and 10 ppm TiO2 seawater media, over a 6-day period. In the control, D. salina cell density continuously increased for four days, reaching a stationary phase thereafter (Figure 9). Over the 6-day experiment, the cell density was approximately 10 times higher than the initial density. In the 1 ppm TiO2 seawater medium, cell density started a slow increase by the second day, significantly rising afterward and reaching a level comparable to the control by the fifth day. However, in the 5 and 10 ppm TiO2 seawater media, cell densities slightly increased but remained at 2000 cells/mL throughout the experiment. By the end of the experiment, cell densities in the 5 and 10 ppm TiO2 seawater media were less than half of those in the control and the 1 ppm seawater medium.
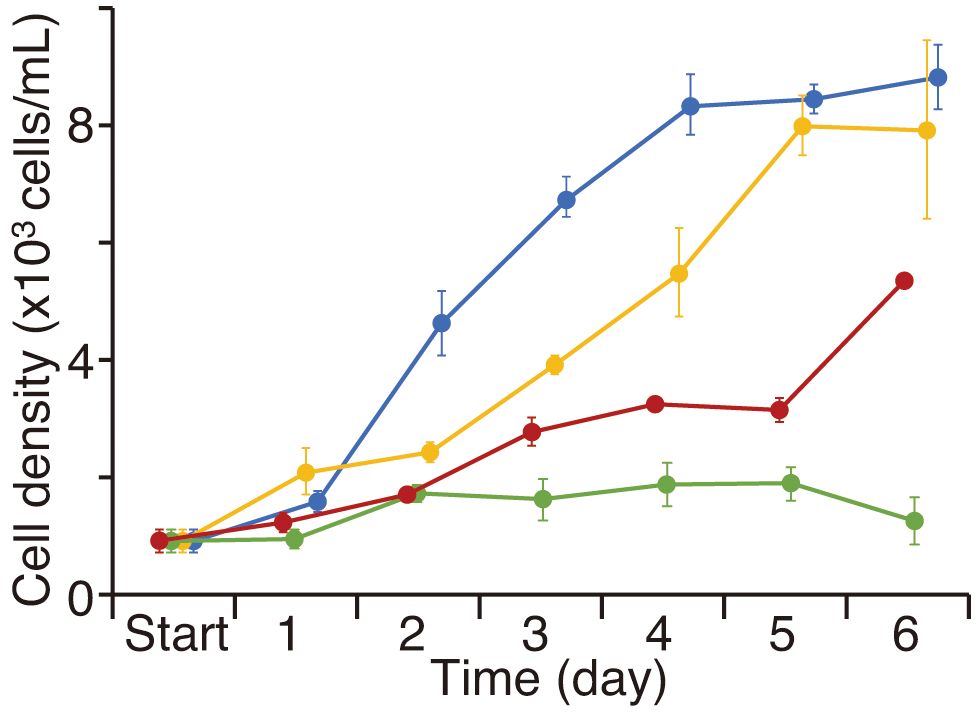
Figure 9 The average of Dunaliella salina cell density every day for the 6-day experiment. Blue, yellow, green, and red colors show the control, 1 ppm, 5 ppm, and 10 ppm TiO2 treatments, respectively.
3.5 TiO2 NPs in Ammonia veneta cell
Angular-shaped grains with distinct contours were observed in vesicles distributed in the latest three chambers by TEM observations (Figures 10A–C; Supplementary Figure S2). These grains, several tens of nanometers in size, exhibited a darker contrast than the surrounding organics and resin, indicating a heavier and more electronically dense element. STEM-EDS analysis detected titanium X-ray peaks from these angular grains (Figures 10D–F), but no titanium peaks were detected from the background (Figures 10G, H). Additionally, STEM-EDS mapping identified abundant titanium-bearing grains in the vesicles, each tens of nanometers in size (Figures 11A, B).
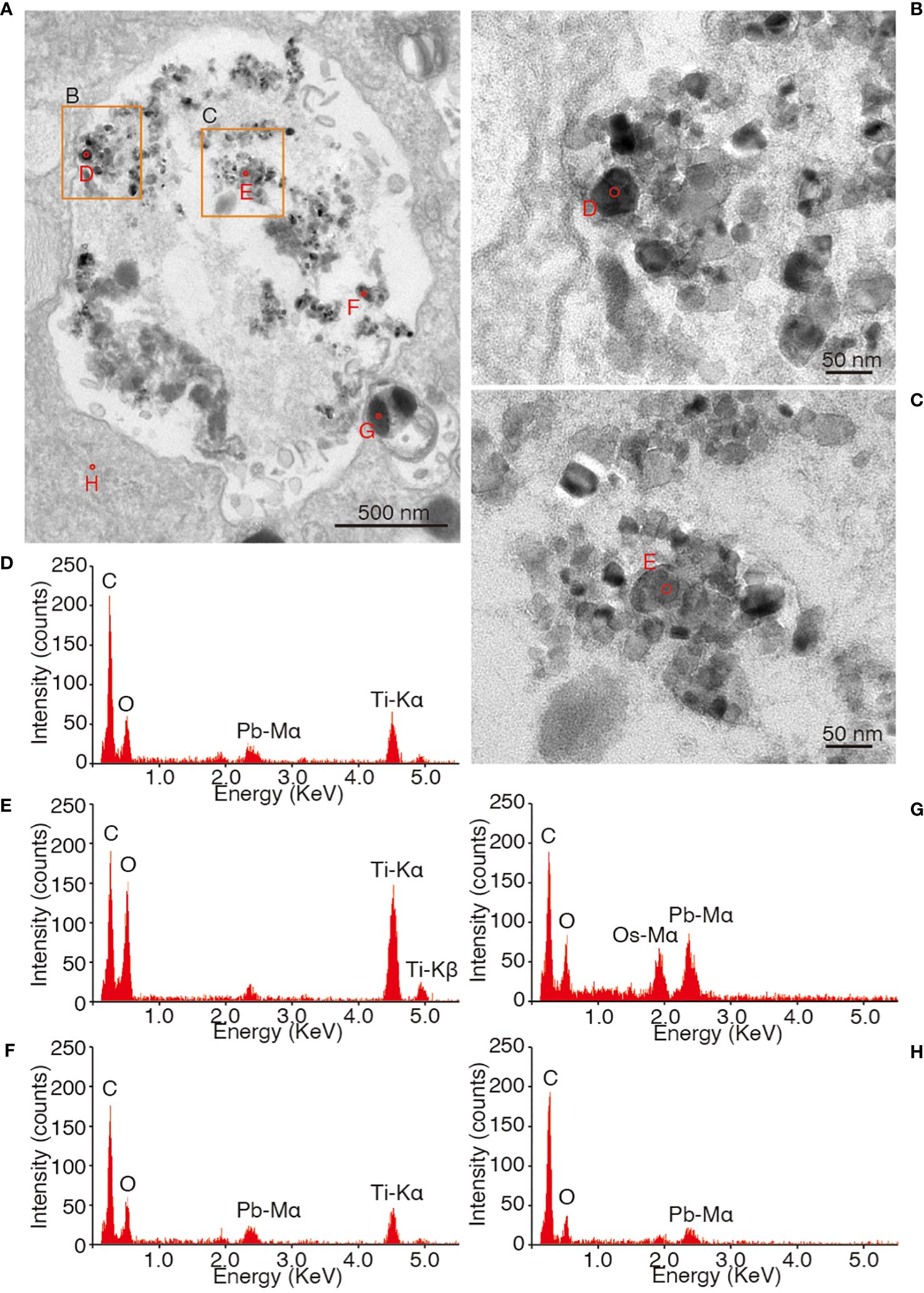
Figure 10 Sectioning images and EDS qualitative spectra of the vesicle, which was placed in the second last chamber of Ammonia veneta cultured in a 5 ppm TiO2 seawater medium. The images are contrast-inverted transmission electron micrographs for better visualization of microstructure. (A) Heavier and electronic dense grains in dark color were distributed in the vesicle. Orange squares correspond to (B, C). Red circles show the spots for EDS analysis. (B, C) Heavier and electronic dense grains were ~50 nm in size with an angular shape. (D–H) Qualitative spectra of EDS analysis at each of the five spots shown in (A). (D–F) are associated with small angular grains, (G) shows an electronic dense body, and (H) places the cytosol as a background.
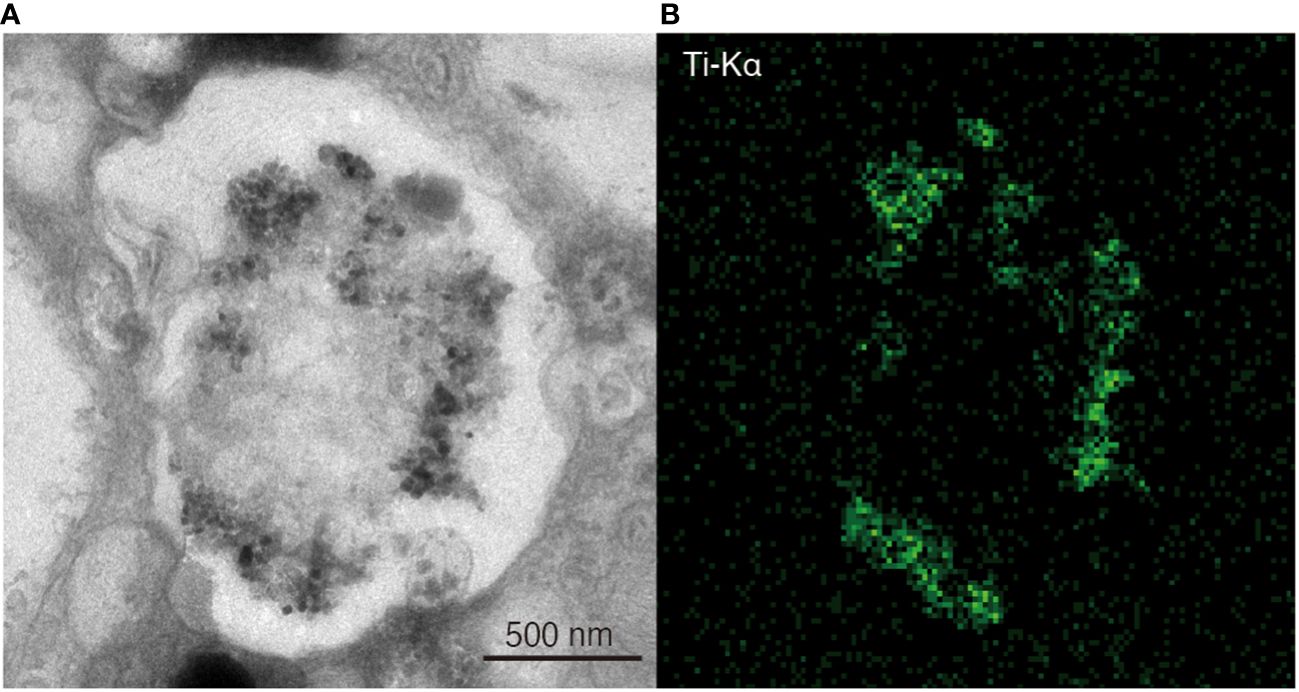
Figure 11 Sectioning images and EDS elemental map of the vesicle, which placed in the second last chamber of Ammonia veneta cultured in 5 ppm TiO2 seawater medium. (A) Contrast-inverted TEM image of vesicle. (B) Distribution of titanium with green color by the EDS elemental map in the same area of the TEM image.
Aggregates of angular grains covered with light-contrast materials were found along the cell, relevant to the surface of the decalcified test (Figure 12A). STEM-EDS analysis detected titanium X-ray peaks from these grains, as well as those in vesicles (Figure 12B), while no titanium peaks were detected from the background (Figure 12C).
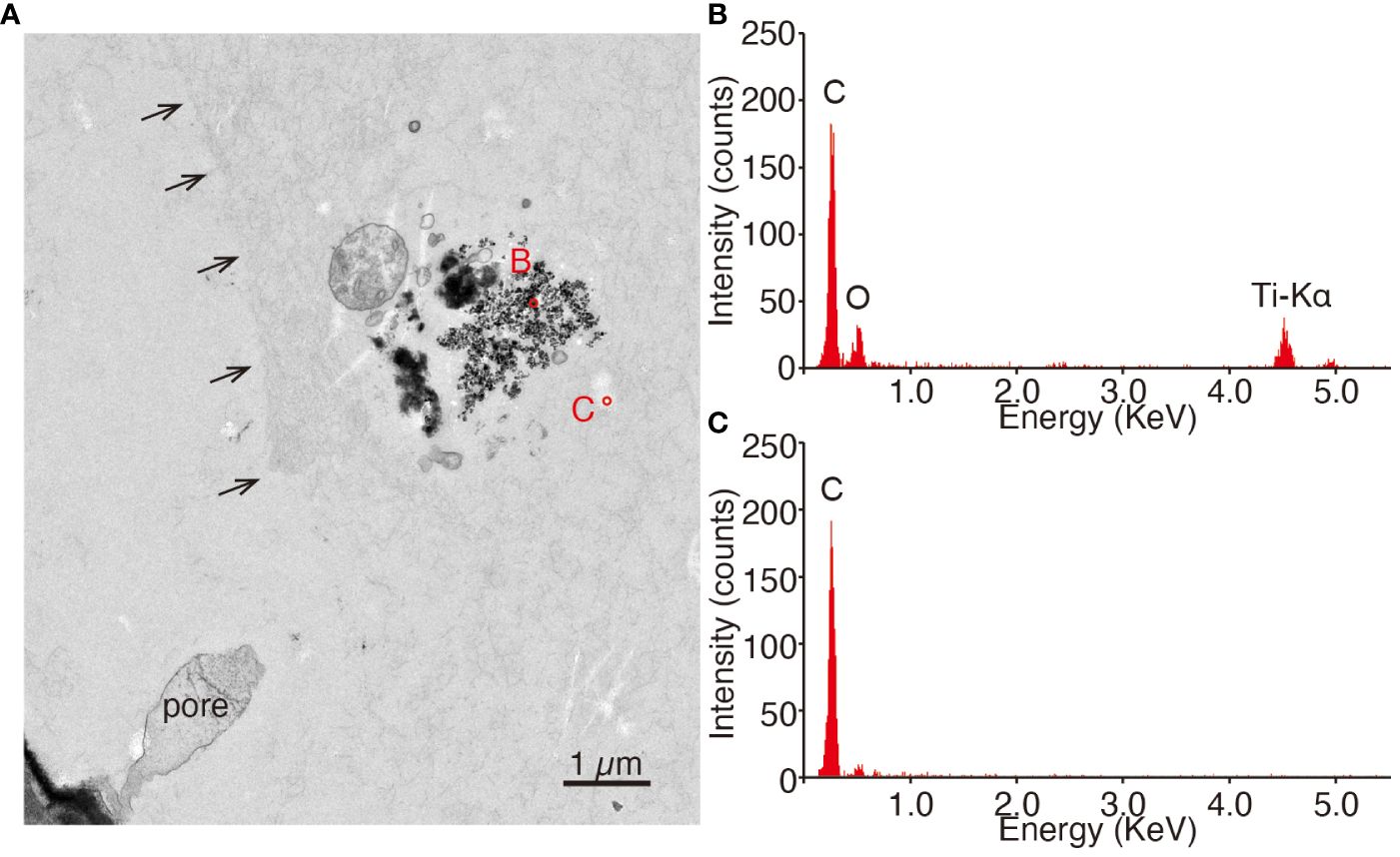
Figure 12 Sectioning images and EDS qualitative spectra of expelled materials placed outside of the decalcified test. (A) Contrast-inverted TEM images. The structure of the pore, as shown by the black arrows, remained. Electronically dense grains were distributed near the pore. Circles show the spot of EDS analysis. (B, C) Qualitative spectra of EDS analysis at the spot shown in (A). (B) is associated with a small angular grain, and (C) is a background.
4 Discussion
We successfully demonstrated the viability of the clonally reproduced strain of the foraminifer based on the activity of the reticulopodia and chamber formation under microscopic observation (Figures 1–3). Ammonia veneta mostly survived for the first three weeks under TiO2 NPs-containing media, even at high TiO2 NPs concentrations exceeding 10 ppm (Figures 4–7). TiO2 NPs did not physically impede the extension of reticulopodia by covering the pores and aperture on the test, as shown by the SEM image (Figures 8C, D). Few TiO2 NPs were observed in the adhesive materials and on the test surface of 10 ppm-treated specimens (Figures 8E, F). The adhesive materials were often found on the shell surface of the 10 ppm-treated specimens, which stopped reticulopodia activity in the early stage of culture experiments and had no growth of chamber (Supplementary Figure S1). Aggregated TiO2 NPs might be attached to the shell surface, when reticulopodia did not extend covering over the shell. Although TiO2 NPs did not physically prevent the extension of reticulopodia, specimens in high concentrations of TiO2 medium over 5 ppm exhibited no extended reticulopodia and shrunk cytoplasm without chamber formation, leading to their demise (Figures 2, 4–7). The control and 1 ppm TiO2 treated specimens showed a gradual increase in chambers up to ~20 in total until the reproduction stage. The methods for counting new chambers in addition to reticulopodia activity have been validated to estimate foraminiferal viability and health in response to anthropogenic pollution (Ben-Eliahu et al., 2020; Losada Ros et al., 2020). Our observation based on the test and cytoplasm including reticulopodia is consistent with the trend of chamber increase over time and is useful for estimating the viability and health of foraminifers.
All four foraminiferal culture experiments showed that the control specimens increased by ~8 chambers and grew into maturity from the young form with ~10 chambers during five weeks (Figures 4–7). The specimens exposed to 1 ppm TiO2 medium showed a slightly low growth rate in Experiment 1, though there was no significant difference from the controls (Figure 4B). When we modified feeding frequency and amount of food in Experiments 2 and 3, the growth rate in 1 ppm TiO2 treatment was the same as the controls, and some specimens reproduced clones (Figures 5B, 6B). The foraminiferal life cycle has been maintained under a 1 ppm TiO2 NPs-contained condition, in congruence with the long-term culturing experiment of the previous study (Ishitani et al., 2023). However, the specimens exposed to high TiO2 NPs concentrations greater than 10 ppm died without growth for four weeks, despite increases of feeding frequency and food volume (Figures 4–7). This indicates that 10 ppm TiO2 NPs concentration is fatal for foraminifers. Furthermore, we found hole-opening on the test surface of specimens exposed to 10 and 50 ppm TiO2 media during the experiments (Figure 3). It is though that foraminifers melt their own tests under conditions of strong stress (Buzas-Stephens and Buzas, 2005), suggesting that high TiO2 NPs concentrations lead foraminifers to strong stress conditions and cell death.
In Experiment 3, most specimens stopped growing in 5 ppm TiO2 medium during the 5-week experiments (Figure 6A). This is possibly caused by a lack of food, D. salina. The culture experiment with D. salina in TiO2 media showed that the cell densities of D. salina in the 5 and 10 ppm TiO2 media were lower than half of those in the control and the 1 ppm TiO2 medium for six days (Figure 9). This result has indicated that the feeding volume in Experiment 3 could not be enough for foraminiferal specimens. However, A. veneta specimens stopped growing in 5 ppm TiO2 medium, even though we increased food volume in Experiment 4 (Figure 7A). Therefore, feeding is not a direct cause to prevent growth of foraminifers. The growth of some microalgae is suppressed in media with metallic NPs as well as TiO2 NPs because of physicochemical effects such as inhibition of photosynthesis by packed TiO2 NPs aggregation and oxidative stress, which damage the cell membrane (Chen et al., 2012; Li et al., 2015; Wang et al., 2016; Ghazaei and Shariati, 2020). The studied specimens have extended reticulopodia at any concentration of TiO2 treatments and seem to take surroundings including TiO2 NPs via protoplasmic streaming of reticulopodia without organismal selection (Figure 1). TEM observation of A. veneta cells derived from 5 ppm TiO2 treatment congruently demonstrates uptake of TiO2 NPs into vesicles (Figures 10A–C, 12B). These specimens could take TiO2 NPs via phagocytosis because of fast aggregation of TiO2 NPs in high concentrations more than 5 ppm (Morelli et al., 2018; Reyes et al., 2021; Palmeira-Pinto et al., 2023). This is congruent with the previous transcriptomic study, which estimated uptake of TiO2 NPs via endocytosis or phagocytosis based on the metabolic pathway in 1 ppm TiO2 treatment (Ishitani et al., 2023). This previous study showed that captured TiO2 NPs induce the Fenton reaction in vesicles, resulting in ROS production, though such TiO2 NPs are encapsulated in ceramide and these capsules are expelled from the cell as mucus after 24 hours (Ishitani et al., 2023). The light-contrast materials, including titanium, around the test could be leftovers of waste excreted by foraminifers (Figure 12B). These observations suggest that the foraminiferal metabolic system is acting even in high concentrations of TiO2 NPs at least 5 ppm condition. This could be caused by two possibilities: a) ROS production is accelerated due to regulation of genes related to stress or releasing toxic substances, and b) overexpression of genes related to releasing toxic substances inhibits cell growth leading to cell death. Future studies will be required to examine the functional changes of detoxification under high TiO2 NPs concentration environments.
The lethal level of our studied organism, A. veneta, is lower than that of microalgae such as green algae and diatoms and those of microorganisms, whose EC50 and LC50 were applicable at concentrations higher than 10 ppm TiO2 NPs (Menard et al., 2011; Minetto et al., 2014). Our results suggest that the previous examination for EC50 and LC50 could be underestimated than the actual effect of TiO2 NPs concentration on marine organisms. The previous studies have used different types of TiO2 NPs: rutile and anatase types with different size ranging from ~10 to ~100 nm. Such material specificity provides different surface charge and other physical characteristics leading to various toxicities reported as cell death by rutile type, while cell necrosis and cell membrane damage by anatase type (Kose et al., 2020). It is still unclear what processes in the metabolic system are affected by the physical characteristics of TiO2 NPs leading to toxicity. Our study, conducted with same type and size of TiO2 NPs as those of the transcriptome study (Ishitani et al., 2023), presented that foraminiferal detoxification is possibly disturbed by overdose (> 5 ppm) of TiO2 NPs. Toxic mechanisms are likely changed associating with concentration of nanoparticles shown by this study, as well as size of nanoparticles. Understanding of both cellular and metabolic mechanisms is important to unveil toxic effects of nanoparticles on various organisms in sets of nanoparticle types. The concentration around 5 ppm of TiO2 NPs, a limit of foraminiferal detoxification, is much lower than ~120 ppm potentially accumulated on European coasts (Sun et al., 2016; Zheng and Nowack, 2021), though nanoparticle pollution has been invisibly but certainly spread out over marine environment. Considering sedimentation of nanoparticles after aggregation, benthic fauna is dominantly exposed to pollutants. Long-term experiments are crucial to examine an extended cytotoxicity of marine organisms including meiofauna as happen in nature. These studies will help to estimate toxic tolerance of marine organisms to nanoparticles for sustaining marine ecosystem under a harmful impact of nanoparticles pollution.
5 Conclusions
This study succeeded to conduct long-term culture experiments based on the exposure of a marine foraminiferal strain, A. veneta, to 1, 5, 10, and 50 ppm TiO2 NPs. This species had been known to have a detoxification mechanism for TiO2 NPs by the previous transcriptome analysis. Our TEM observations showed that these foraminifers took TiO2 NPs into vesicles and ejected some of them even at 5 ppm TiO2 NPs exposure. However, foraminifers stopped growing in the early stage of 5-week culture experiments under 5 ppm TiO2 NPs exposure despite the availability of food. These findings indicate that the foraminiferal detoxification system could be disrupted by over 5 ppm TiO2 NPs. The demand for TiO2 NPs has grown and their concentration is estimated at up to 120 ppm around the European coast nowadays. Such acceleration of discharging TiO2 NPs to the ocean could moderate the foraminiferal detoxification ability leading a breakdown of marine ecosystem, because meio-benthos such as foraminifers play an important role for marine food chain. Future studies will be required to examine toxic mechanisms of various marine organisms, for understanding an effect of nanoparticles to the entire marine ecosystem including meiofauna.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
YIn: Writing – original draft, Methodology, Investigation, Data curation, Conceptualization. YIs: Writing – original draft, Resources, Investigation. AT: Writing – review & editing, Investigation. KU: Writing – review & editing, Investigation. NT: Writing – review & editing, Investigation. TU: Writing – review & editing, Investigation. YU: Writing – original draft, Project administration, Funding acquisition, Conceptualization.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work is supported by ESPEC Foundation for Global Environment Research and Technology and Japan Society for the Promotion of Science (JSPS) grant (20H02016).
Acknowledgments
We thank M. Kakimoto, K. Kukita, I. Ikuma, and K. Miura from Kochi University for their help with the culture experiments.
Conflict of interest
Author KU was employed by Marine Works Japan Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1381247/full#supplementary-material
Supplementary Figure 1 | SEM images and EDS analyses of control and 10 ppm TiO2 treated specimens.
Supplementary Figure 2 | TEM images of vesicles in a 5 ppm TiO2 treated specimen.
Supplementary Table 1 | The number of increased chambers in foraminiferal culture experiments 1–4.
References
Ammendolia C., Hofkirchner C., Plener J., Bussières A., Schneider M. J., Young J. J., et al. (2022). Non-operative treatment for lumbar spinal stenosis with neurogenic claudication: an updated systematic review. BMJ Open 12, e057724. doi: 10.1136/bmjopen-2021-057724
Ben-Eliahu N., Herut B., Rahav E., Abramovich S. (2020). Shell growth of large benthic foraminifera under heavy metals pollution: Implications for geochemical monitoring of coastal environments. Int. J. Environ. Res. Public Health 17, 3741. doi: 10.3390/ijerph17103741
Boxall A. B., Tiede K., Chaudhry Q. (2007). Engineered nanomaterials in soils and water: how do they behave and could they pose a risk to human health? Nanomed. (Lond). 2, 919–927. doi: 10.2217/17435889.2.6.919
Buzas-Stephens P., Buzas M. A. (2005). Population dynamics and dissolution of foraminifera in Nueces Bay, Texas. J. Foraminiferal Res. 35, 248–258. doi: 10.2113/35.3.248
Chen L., Zhou L., Liu Y., Deng S., Wu H., Wang G. (2012). Toxicological effects of nanometer titanium dioxide (nano-TiO2) on Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 84, 155–162. doi: 10.1016/j.ecoenv.2012.07.019
Ciacci C., Grimmelpont M. V., Corsi I., Bergami E., Curzi D., Burini D., et al. (2019). Nanoparticle-biological interactions in a marine benthic foraminifer. Sci. Rep. 9, 19441. doi: 10.1038/s41598-019-56037-2
Clément L., Hurel C., Marmier N. (2013). Toxicity of TiO2 nanoparticles to cladocerans, algae, rotifers and plants - effects of size and crystalline structure. Chemosphere 90, 1083–1090. doi: 10.1016/j.chemosphere.2012.09.013
Culver S. J., Buzas M. A. (1999). “Biogeography of neritic benthic foraminifera,” in Modern Foraminifera, eds. Barun K., Sen G. (Dordrecht: Springer Netherlands). doi: 10.1007/0-306-48104-9_6
Delay M., Frimmel F. H. (2012). Nanoparticles in aquatic systems. Anal. Bioanal. Chem. 402, 583–592. doi: 10.1007/s00216-011-5443-z
Farré M., Gajda-Schrantz K., Kantiani L., Barceló D. (2009). Ecotoxicity and analysis of nanomaterials in the aquatic environment. Anal. Bioanal. Chem. 393, 81–95. doi: 10.1007/s00216-008-2458-1
Ghazaei F., Shariati M. (2020). Effects of titanium nanoparticles on the photosynthesis, respiration, and physiological parameters in Dunaliella salina and Dunaliella tertiolecta. Protoplasma 257, 75–88. doi: 10.1007/s00709-019-01420-z
Goldstein S. T. (1999). “Foraminifera: A biological overview,” in Modern Foraminifera, eds. Barun K., Sen G. (Dordrecht: Springer Netherlands). doi: 10.1007/0-306-48104-9_3
Hartmann N. B., Von der Kammer F., Hofmann T., Baalousha M., Ottofuelling S., Baun A. (2010). Algal testing of titanium dioxide nanoparticles - testing considerations, inhibitory effects and modification of cadmium bioavailability. Toxicology 269, 190–197. doi: 10.1016/j.tox.2009.08.008
Ishitani Y., Ciacci C., Ujiié Y., Tame A., Tiboni M., Tanifuji G., et al. (2023). Fascinating strategies of marine benthic organisms to cope with emerging pollutant: Titanium dioxide nanoparticles. Environ. pollut. 330, 121538. doi: 10.1016/j.envpol.2023.121538
Kose O., Tomatis M., Leclerc L., Belblidia N. B., Hochepied J. F., Turci F., et al. (2020). Impact of the physicochemical features of TiO2 nanoparticles on their in vitro toxicity. Chem. Res. Toxicol. 33, 2324–2337. doi: 10.1021/acs.chemrestox.0c00106
Li F., Liang Z., Zheng X., Zhao W., Wu M., Wang Z. (2015). Toxicity of nano-TiO2 on algae and the site of reactive oxygen species production. Aquat. Toxicol. 158, 1–13. doi: 10.1016/j.aquatox.2014.10.014
Losada Ros M. T., Al-Enezi E., Cesarini E., Canonico B., Bucci C., Alves Martins M. V., et al. (2020). Assessing the cadmium effects on the benthic foraminifer Ammonia cf. parkinsoniana: An acute toxicity test. Water 12, 1018. doi: 10.3390/w12041018
Luo Z., Li Z., Xie Z., Sokolova I. M., Song L., Peijnenburg W. J. G. M., et al. (2020). Rethinking nano-TiO2 safety: overview of toxic effects in humans and aquatic animals. Small 16, e2002019. doi: 10.1002/smll.202002019
Matranga V., Corsi I. (2012). Toxic effects of engineered nanoparticles in the marine environment: model organisms and molecular approaches. Mar. Environ. Res. 76, 32–40. doi: 10.1016/j.marenvres.2012.01.006
Menard A., Drobne D., Jemec A. (2011). Ecotoxicity of nanosized TiO2. Review of in vivo data. Environ. pollut. 159, 677–684. doi: 10.1016/j.envpol.2010.11.027
Minetto D., Libralato G., Ghirardini. A. V. (2014). Ecotoxicity of engineered TiO2 nanoparticles to saltwater organisms: an overview. Environ. Int. 66, 18–27. doi: 10.1016/j.envint.2014.01.012
Moore M. N. (2006). Do nanoparticles present ecotoxicological risks for the health of the aquatic environment? Environ. Int. 32, 967–976. doi: 10.1016/j.envint.2006.06.014
Morelli E., Gabellieri E., Bonomini A., Tognotti D., Grassi G., Corsi I. (2018). TiO2 nanoparticles in seawater: Aggregation and interactions with the green alga Dunaliella tertiolecta. Ecotoxicol. Environ. Saf. 148, 184–193. doi: 10.1016/j.ecoenv.2017.10.024
Palmeira-Pinto L., Emerenciano A. K., Bergami E., Joviano W. R., Rosa A. R., Neves C. L., et al. (2023). Alterations induced by titanium dioxide nanoparticles (nano-TiO2) in fertilization and embryonic and larval development of the tropical sea urchin Lytechinus variegatus. Mar. Environ. Res. 188, 106016. doi: 10.1016/j.marenvres.2023.106016
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Reyes J. P., Lagdameo J. D., Celorico J. R., Almeda R. A., Peralta M. M., Basilia B. A. (2021). Aquatic toxicity studies of titanium dioxide and silver nanoparticles using Artemia franciscana Nauplii and Daphnia magna. Mater. Sci. Eng. A. 11, 107–113. doi: 10.17265/2161-6213/2021.10-12.001
Sun T. Y., Bornhöft N. A., Hungerbühler K., Nowack B. (2016). Dynamic probabilistic modeling of environmental emissions of engineered nanomaterials. Environ. Sci. Technol. 50, 4701–4711. doi: 10.1021/acs.est.5b05828
Wang Y., Zhu X., Lao Y., Lv X., Tao Y., Huang B., et al. (2016). TiO2 nanoparticles in the marine environment: Physical effects responsible for the toxicity on algae Phaeodactylum tricornutum. Sci. Total Environ. 565, 818–826. doi: 10.1016/j.scitotenv.2016.03.164
Keywords: cytotoxicity, detoxification, foraminifera, TiO2 NPs, fatal level
Citation: Inagaki Y, Ishitani Y, Tame A, Uematsu K, Tomioka N, Ushikubo T and Ujiié Y (2024) Foraminiferal detoxification breakdown induced by fatal levels of TiO2 nanoparticles. Front. Mar. Sci. 11:1381247. doi: 10.3389/fmars.2024.1381247
Received: 03 February 2024; Accepted: 17 June 2024;
Published: 01 July 2024.
Edited by:
Xuchun Qiu, Jiangsu University, ChinaReviewed by:
Praxedes Natalia Muñoz, Catholic University of the North, ChileFabrizio Frontalini, University of Urbino Carlo Bo, Italy
Copyright © 2024 Inagaki, Ishitani, Tame, Uematsu, Tomioka, Ushikubo and Ujiié. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yurika Ujiié, yujiie@kochi-u.ac.jp
†Present address: Yuka Inagaki, Graduate School of Integrated Arts and Sciences, Kochi University, Kochi, Japan