 Benny K. K. Chan
Benny K. K. Chan Hiromi Kayama Watanabe
Hiromi Kayama Watanabe Chong Chen
Chong Chen- 1Biodiversity Research Center, Academia Sinica, Taipei, Taiwan
- 2X-STAR, Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Kanagawa, Japan
by Mao N, Shao W, Cai Y, Kong X, Ji N and Shen X (2024). Front. Mar. Sci. 10:1269411. doi: 10.3389/fmars.2023.1269411
Mao et al. (2024) recently published an article comparing the mitochondrial genome and transcriptome of the deep-sea hydrothermal vent barnacle Vulcanolepas fijiensis and the relatively shallower barnacle Scalpellum stearnsi (contradictory to the title, no new species was described). This paper concluded that the mitogenomes and key genes found in the transcriptomic analysis were subjected to positive selection related to high-temperature and high-pressure conditions, allowing V. fijiensis to adapt to the deep-sea environment. However, the neolepadid hydrothermal vent barnacle used in their study was misidentified and actually represents a different species, Neolepas marisindica. We present our evidence and reasoning for this below. Although the main conclusions presented by Mao et al. (2024) still stand because N. marisindica is also a deep-sea vent species, their misidentification has connotations on the context, interpretation, and of course future use of their data.
The deep-sea hydrothermal vent barnacle specimen referred to as V. fijiensis in Mao et al. (2024) was collected at Longqi hydrothermal vent field on the Southwest Indian Ridge at a depth of 2759 m in the Indian Ocean. Vulcanolepas fijiensis was originally described from the North Fiji Basin in the Southwest Pacific (Chan et al., 2019) and had never been reported in the Indian Ocean. Watanabe et al. (2018) were the first to explore the diversity of neolepadid hydrothermal vent barnacles in the Indian Ocean and found only one species, Neolepas marisindica, from several vent fields including the Longqi vent field. Though N. marisindica exhibits two morphotypes, these have been shown to be the same species using molecular data (Watanabe et al., 2018).
Using the mitochondrial genome of V. fijiensis supplied by Mao et al. (2024) in GenBank (MZ772032), we show in a phylogenetic tree that the mitochondrial COI (cytochrome c oxidase subunit I) barcode sequence of V. fijiensis in Mao et al. (2023) is nested into a well-supported clade with other available sequences of N. marisindica and not with V. fijiensis sequences from the original description by Chan et al. (2019) that includes barcodes of the holotype (Figure 1). This sequence from Mao et al. (2024) was identical to published sequences of N. marisindica from both Kairei (type locality; LC350008, LC350015) and Solitaire (LC350013) vent fields on the Central Indian Ridge (CIR). Searching on GenBank revealed another mitogenome listed as V. fijiensis (MW291947) linked to the Mao et al. (2024) paper, but this too was nested within N. marisindica (Figure 1). Furthermore, a mitogenome of the real V. fijiensis from the North Fiji Basin had already been published (Lee et al., 2019; MN061491) using one of the original specimens sequenced in the description paper, and a BLAST comparison with the mitogenome (MZ772032) presented by Mao et al. (2024) revealed a percentage identity of only 88.84% between these two mitogenomes. Altogether, these evidences show that the species identification in Mao et al. (2024) was incorrect, and their hydrothermal vent barnacle was actually N. marisindica.
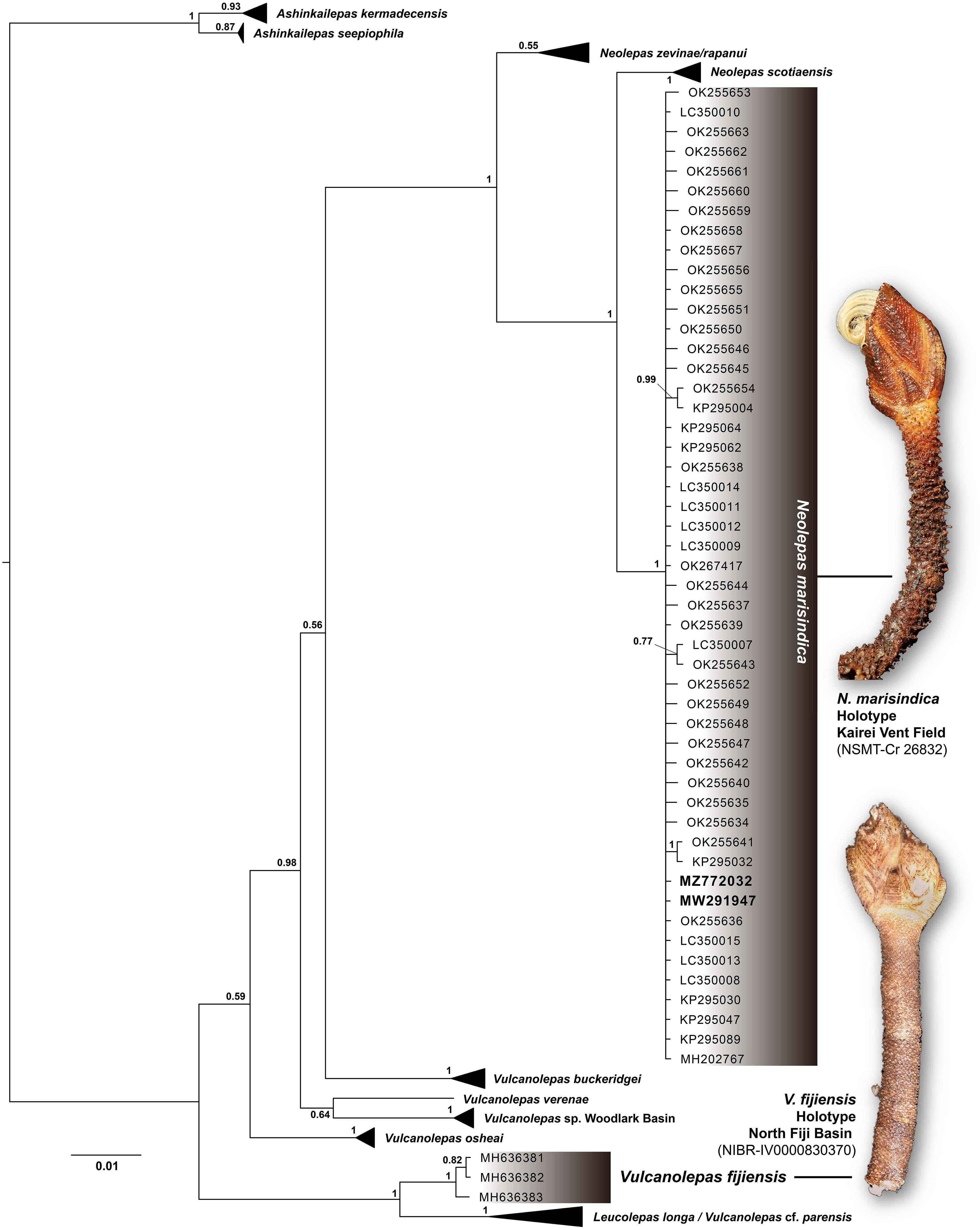
Figure 1 Phylogenetic tree of neolepadid stalked barnacles reconstructed by Bayesian inference with the GTR+I+G model, following published methods in Watanabe et al. (2023). For simplicity, taxa other than Vulcanolepas fijiensis and Neolepas marisinidica have been collapsed. Accession numbers in bold refer to sequences referred to as V. fijiensis in Mao et al. (2024). Node values indicate Bayesian posterior probability. The 528 bp alignment used contained all 202 COI barcoding sequences openly available on GenBank, accession numbers as follows: JX036420-JX036464, KF739820-KF739838, KP295001, KP295004-KP295005, KP295007-KP295009, KP295013-KP295014, KP295018-KP295019, KP295022, KP295026-KP295028, KP295030-KP295037, KP295039-KP295042, KP295045-KP295053, KP295055-KP295058, KP295060-KP295064, KP295067-KP295069, KP295073, KP295076, KP295078, KP295080, KP295082-KP295085, KP295089-KP295091, KP295094, KP295097-KP295098, KY502196-KY502197, LC350007-LC350015, MH202767, MH636381-MH636383, MN722546-MN722548, MN722550-MN722551, MT662001, MW291947, MW602536-MW602540, MW602548-MW602549, MW602552-MW602566, MZ772032, OK255634-OK255663, OK267417. Holotype of V. fijiensis (NIBR-IV0000830370; Korea National Institute for Biological Resources, Incheon, Korea) as well as N. marisindica (NSMT-Cr 26832; National Museum of Nature and Science, Tsukuba, Japan) shown in inset, photographs were taken by Benny K. K. Chan.
We also note that another layer of confusion came from an inset specimen photo used in Figure 1A of Mao et al. (2024), which shows a photo of the real V. fijiensis directly taken from figure 3F of Chan et al. (2019) (original authors were not consulted about this use prior to publication). This photograph is of the holotype of V. fijiensis housed in the collection of the Korea National Institute for Biological Resources (Incheon, Korea; NIBR-IV0000830370) where it still resides. As such, the photo of V. fijiensis presented in Mao et al. (2024) is misleadingly not linked to the actual specimen used in their study.
Hydrothermal vent field stalked barnacles in the family Neolepadidae often exhibit morphological variations in the capitular shells and peduncular scales (Watanabe et al., 2021), which may have contributed to the misidentification in Mao et al. (2024). A more reliable identification for deep-sea hydrothermal vent barnacles should be based on an integrative approach, a combination of morphological and barcoding sequence matching with reference sequences available in GenBank. With the misidentification in Mao et al. (2024) corrected, Neolepas marisindica remains the only neolepadid species known from the Indian Ocean so far (Watanabe et al., 2018; Kim et al., 2020; 2021, Zhou et al., 2018; 2022), known from nine vent fields across four ridges (SWIR, CIR, Southeast Indian Ridge, and Carlsberg Ridge). At present, the hydrothermal vent stalked barnacle species identified in the Pacific include V. fijiensis, V. osheai, V. buckeridgei, V. verenae, Leucolepas longa, Neolepas rapanuii, and N. zevinae (see Watanabe et al., 2021); an undescribed species is also known from Woodlark Basin (Boulart et al., 2022). Such a contrasting diversity pattern between the Pacific and Indian Ocean vent fields supports the Dispersal Corridor Hypothesis, where the ancestors of the Indian Ocean neolepadid barnacles originated from the Pacific Ocean and entered the Indian Ocean via the Southern Ocean (see Watanabe et al., 2018).
Correct species identification is of utmost importance to ensure the correct interpretation and usage of molecular and distribution data. The data in Mao et al. (2024) presents the first assembled and published mitogenome sequence for N. marisinidica, a valuable piece of new data that will be useful in the future for understanding the evolutionary history of this species and vent barnacles as a whole. Although previously another transcriptome for this species was published using specimens from Onnuri vent field on the CIR (Ryu et al., 2019), this means the Longqi transcriptome data from Mao et al. (2024) provides an opportunity to make an interesting comparison with Onnuri to reveal potential differences between conspecific barnacles in two different vents across two ridge systems. We hope this case serves as an example for biologists working on difficult-to-identify deep-sea species to embrace integrative taxonomy and to ensure the species identification and taxonomy is correct prior to publication.
Author contributions
BC: Funding acquisition, Writing – original draft, Validation, Investigation, Conceptualization. HW: Writing – original draft, Visualization, Software, Resources, Methodology, Investigation, Formal analysis, Data curation, Conceptualization. CC: Writing – original draft, Visualization, Validation, Methodology, Investigation, Data curation, Conceptualization.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. BKKC acknowledges intramural research funding from the Biodiversity Research Center, Academia Sinica, Taiwan.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Boulart C., Rouxel O., Scalabrin C., Le Meur P., Pelleter E., Poitrimol. C., et al. (2022). Active hydrothermal vents in the Woodlark Basin may act as dispersing centres for hydrothermal fauna. Commun. Earth Env. 3, 64. doi: 10.1038/s43247–022-00387–9
Chan B. K. K., Ju S., Kim S. J. (2019). A new species of hydrothermal vent stalked barnacle Vulcanolepas (Scalpelliforms: Eolepadidae) from the North Fiji Basin, Southwestern Pacific Ocean. Zootaxa 4563, 135–148. doi: 10.11646/ZOOTAXA.4563.1.7
Kim J., Son S.-K., Kim D., Pak S.-J., Yu O. H., Walker S. L., et al. (2020). Discovery of active hydrothermal vent fields along the Central Indian Ridge, 8–12°S. Geochemistry Geophysics Geosystems 21, e2020GC009058. doi: 10.1029/2020GC009058
Lee W.-K., Kang H. M., Chan B. K. K., Ju S.-J., Kim S.-J. (2019). Complete mitochondrial genome of the hydrothermal vent stalked barnacle Vulcanolepas Fijiensis (Cirripedia, Scalpelliforms, Eolepadidae). Mitochondrial DNA Part B 4, 2725–2726. doi: 10.1080/23802359.2019.1644564
Mao N., Shao W., Cai Y., Kong X., Ji N., Shen X. (2024). Comparative omics analysis of a new deep-sea barnacle species (Cirripedia, Scalpellomorpha) and shallow-water barnacle species provides insights into deep-sea adaptation. Front. Mar. Sci. 10, 1269411. doi: 10.3389/fmars.2023.1269411
Ryu T., Woo S., Lee N. (2019). The first reference transcriptome assembly of the stalked barnacle, Neolepas marisindica, from the Onnuri Vent Field on the Central Indian Ridge. Mar. Genomics 48, 100679. doi: 10.1016/j.margen.2019.04.004
Watanabe H. K., Chen C., Chan B. K. K. (2021). A new deep-sea hot vent stalked barnacle from the Mariana Trough with notes on the feeding ecology of Vulcanolepas. Mar. Biodiversity 51, 9. doi: 10.1007/s12526-020-01144-x
Watanabe H. K., Chen C., Marie D. P., Takai K., Fujikura K., Chan B. K. K. (2018). Phylogeography of hydrothermal vent stalked barnacles: a new species fills a gap in the Indian Ocean ‘dispersal corridor’ hypothesis. R. Soc Open Sci. 5, 172408. doi: 10.1098/rsos.172408
Watanabe H. K., Uyeno D., Yamamori L., Jimi N., Chen C. (2023). From commensalism to parasitism within a genus-level clade of barnacles. Biol. Lett. 19, 20220550. doi: 10.1098/rsbl.2022.0550
Zhou Y., Chen C., Zhang D., Wang Y., Watanabe H. K., Sun J., et al. (2022). Delineating biogeographic regions in Indian Ocean deep-sea vents and implications for conservation. Diversity Distributions 28, 2858–2870. doi: 10.1111/ddi.13535
Zhou Y., Zhang D., Zhang R., Liu Z., Tao C., Lu B., et al. (2018). Characterization of vent fauna at three hydrothermal vent fields on the Southwest Indian Ridge: Implications for biogeography and interannual dynamics on ultraslow-spreading ridges. Deep Sea Res. Part I: Oceanographic Res. Papers 137, 1–12. doi: 10.1016/j.dsr.2018.05.001
Keywords: chemosynthetic ecosystem, integrative taxonomy, Neolepadidae, phylogeny, species identification, systematics
Citation: Chan BKK, Watanabe HK and Chen C (2024) Commentary: Comparative omics analysis of a new deep-sea barnacle species (Cirripedia, Scalpellomorpha) and shallow-water barnacle species provides insights into deep-sea adaptation. Front. Mar. Sci. 11:1374419. doi: 10.3389/fmars.2024.1374419
Received: 22 January 2024; Accepted: 26 April 2024;
Published: 28 May 2024.
Edited by:
Andrew R. Thurber, Oregon State University, United StatesCopyright © 2024 Chan, Watanabe and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chong Chen, Y2NoZW5AamFtc3RlYy5nby5qcA==
‡ORCID: Benny K. K. Chan, orcid.org/0000-0001-9479-024X
Hiromi Kayama Watanabe, orcid.org/0000-0001-5031-9018
Chong Chen, orcid.org/0000-0002-5035-4021