 Aiman Imentai
Aiman Imentai Volodymyr Bondarenko2
Volodymyr Bondarenko2 Tomáš Pěnka
Tomáš Pěnka Tomáš Policar
Tomáš Policar- 1Faculty of Fisheries and Protection of Waters, South Bohemian Research Center of Aquaculture and Biodiversity of Hydrocenoses, University of South Bohemia in České Budějovice, Vodňany, Czechia
- 2Faculty of Fisheries and Environmental Sciences, Kherson State Agricultural University, Kherson, Ukraine
In this study, three separate experiments were conducted to optimize the intensive larviculture of pike (Esox lucius L.) under different weaning times, light regimes, and stocking densities. In the first experiment, larvae were fed sequential combinations of live feed (LF), co-feeding with dry starter (DS), and only DS feed for 18 days as follows: 9 days LF followed by 3 days co-feeding and 5 days DS (group A); 6 days LF followed by 3 days co-feeding and 9 days DS (group B); 3 days LF followed by 3 days co-feeding and 12 days DS (group C); and DS alone for 18 days (group D). Fish fed LF had significantly higher growth rates than those fed a dry diet (p< 0.05). Extending the feeding period on LF resulted in a significantly higher growth rate (24.6% d−1), variability (13.5%), and heterogeneity (299.5% d−1) than fish fed a DS. In the second experiment, the effect of the light regime was assessed at eight different levels (L0:D24; L24:D0; L16:D8; L4:D4:L4:D4:L4:D4; L8:D4:L8:D4; L12:D12; L8:D16; and L4:D8:L4:D8). The growth and survival of the larvae increased with increasing light period. Groups exposed to complete light (L24:D0) and 16 h of light (L16:D8) exhibited significantly higher weight gain and specific growth rate (SGR, %) than the other groups. The highest survival was observed in fish reared with complete light (68.5 ± 4.5%) and an L8:D4:L8:D4 light regime (61.4 ± 5.2%). All larvae maintained under L0:D24 died during the experiment. The third experiment tested the effect of the fish density within four experimental groups: 10 ind L−1, 20 ind L−1, 40 ind L−1, and 80 ind L−1. Fish at densities of 20 and 40 ind L−1 had significantly higher weight gain and SGR (%) than those in the other groups. The highest survival was in fish reared with 20 ind L−1 density (72.5 ± 8.5%). Fish at densities of 10 and 80 ind L−1 showed significantly higher cannibalism rates than those in the other groups. These results indicated that weaning time, photoperiod, and stocking density play significant roles in early pike larval performance.
1 Introduction
The northern pike, Esox lucius L., a fish species widely distributed in most water bodies in the Northern Hemisphere (Lucas, 1996), can influence the abundance and distribution of many species (Hubenova et al., 2007; Craig, 2008). It is promising for inland aquaculture diversification (Samarin et al., 2016) because of its high-quality meat and popularity in commercial fishing and angling (Salam and Davies, 1994; Bondarenko et al., 2015b). In inland aquaculture, the northern pike is cultured mainly in ponds owing to its biomelioration effect within the polyculture fish stock (Prejs et al., 1994). However, the effective and stable production of marketable or stocked fish of this species from year to year is difficult to manage because of the pike’s rapacity, gluttony, and tendency for cannibalism (Bondarenko et al., 2015a). Moreover, uncontrollable pond aquaculture, overfishing, water pollution, degraded natural grounds, and limited natural resources are the main factors limiting the stable production of northern pikes in Europe (Lehtonen et al., 2009; Larsson et al., 2015).
Therefore, in the last two decades, researchers, mainly from Central Europe, have attempted to innovate and optimize northern pike-controlled broodstock reproduction, egg incubation (Szabó, 2001, 2003, 2008; Kucska et al., 2006a; Hulak et al., 2008; Hadi Alavi et al., 2009; Bondarenko et al., 2015b, 2015a; Samarin et al., 2016; Kristan et al., 2020), and intensive larval and juvenile culture using Recirculating Aquaculture System (RAS) technology (Kucska et al., 2006b, 2007; Szczepkowski, 2009). The aim of this study was to increase the stable, high-quality production of pike juveniles by optimizing light intensity, water temperature, fish density, feeding regime, food quality, tank shape, water surface shading, and size grading (Kucska et al., 2006b, 2007; Jankowska et al., 2008; Szczepkowski, 2009; Kozłowski and Piotrowska, 2022), mainly by eliminating cannibalistic behavior and supporting high growth and development (Giles et al., 1986; Kucharczyk et al., 1998; Szczepkowski, 2009). However, larval culture of the northern pike during the first period of exogenous feeding has not yet been optimized, and information is limited.
Therefore, this study mainly focused on the culture period of larvae and early juveniles, with the goal of evaluating the effects of weaning time, light regime, and stocking density on northern pike larvae and early juvenile growth, conditions, survival, and cannibalism rate in RAS until an acceptable juvenile size was reached for stocking into a follow-up pond or RAS culture.
2 Materials and methods
2.1 Experimental larvae for three experiments
Larvae used in all three experiments in this study were obtained from pond-cultured northern pike broodstock in Fishery Nove Hrady Ltd. [15 females and 45 males with body weights (BWs) between 0.8 and 1.5 kg], environmentally stimulated by increasing water temperature and sunshine under ambient outdoor conditions. Females and males were separated and maintained in two earthen ponds with a total area of 500 m2 and littoral vegetation covering approximately 100 m2 of pond area. No hormones were injected. Females were checked for ovulation at weekly intervals, when water temperature fluctuated during the day from 6°C to 12°C. Females were captured by hand and with fishing nets in the littoral area of the pond and were tested for egg ovulation. When ovulation was detected, females were moved near the hatchery for egg stripping and fertilization, according to the method of Bondarenko et al. (2015b). For egg fertilization of each female, testicular sperm collected from three killed males was used, according to the method of Kristan et al. (2020). Fresh fertilized eggs were desticked and incubated under controlled condition with an optimal water temperature of 6–10°C (Bondarenko et al., 2015a). Eggs from each female were incubated separately in 10-L Zug jars. For each experiment, fertilization and incubation of eggs and larvae from five females were performed continuously at one 18-day interval and two 14-day intervals.
Larvae obtained at 11 days post-hatching (DPH) were tested by osmotic shock based on Policar et al. (2010) prior to the transport and stock into each experiment, and only high-quality larvae at 12 DPH presenting survival rates >85% after 90 min of shock (Bondarenko et al., 2015a) were transported in plastic bags (30 L) filled with 2/3 oxygen and 1/3 water from the Fishery Nove Hrady Ltd. hatchery to the Experimental Fish Facility of the Faculty of Fisheries and Protection of Waters, University of South Bohemia (FFPW USB), Vodňany, Czech Republic. After transportation, the larvae (12 DPH) were counted and stocked in experimental tanks for acclimatization over a day. The same procedure was repeated for each experiment. Significantly smaller larvae were obtained and stocked at the same age [BW = 8.9–9.0 mg and total length (TL) = 10.8 mm] at the beginning of the first and third experiments, representing the beginning and the end of northern pike reproductive season compared to larvae stocked for the second experiment performed in the middle of the reproductive season (BW = 11.5–12.5 and TL = 10.8 mm).
2.2 Culture facilities, water quality, and use of dry feed during three experiments
At the beginning of each experiment, the larvae were stocked and cultured in 16 cylindrical plastic tanks with an identical volume of 180 L of the same RAS in the FFPW USB. This system comprised culture tanks and water outlets connected with a mechanical parabolic screen filter Ultra Sieve 200 with a maximum water flow of 15,000 L. After the mechanical filtration, water in the RAS continued to the biological filter designed for this study, which comprised two parts: (1) a submerged filter with a total volume of 550 L filled with 50% water and 50% polyurethane foam filter mats (PU PPI30), and (2) a bed-moving filter with a total volume of 550 L filled with 50% water and 50% filtration elements (Random Media BT 10, Ratz Ltd., Leinfelden-Echterdingen, Germany). An EL-S 250 W air blower (Secoh Shanghai Mec. Ltd., Shanghai, China) was used as an air source in both biofilters through air stones and plastic grates installed on the bottom of filters. After biological filtration, the treated water was sterilized with UV light (UV lamp EVO 110, Evolution Aqua Ltd., Wigan, UK) and pumped into a 1,000-L retention tank; from this tank, the water was gravitated back to the fish tanks.
Water temperature (°C) and oxygen saturation (%) were determined during the three experiments using a portable YSI ProODO oximeter (YSI Inc., Yellow Springs, OH, USA) in each tank twice daily (7:30 a.m. and 6:30 p.m.). The pH values were determined using a WTW 3310 pH (WTW GmbH, Weilheim, Germany) meter daily at 8:00 a.m. by submerging it in the biological filter of the RAS system. Total ammonia and nitrite concentrations were measured using a colorimetric reference kit according to Pěnka et al. (2021) once daily. The average water quality parameters for the three experiments are summarized in Table 1.
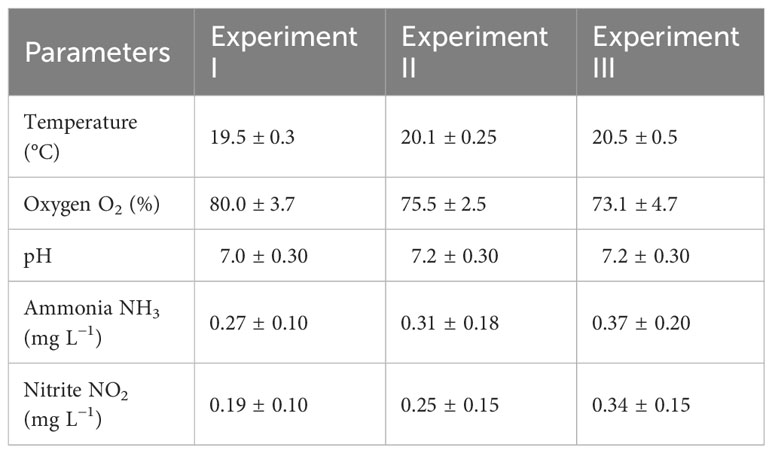
Table 1 Water quality parameters in fish rearing tanks during three separate experiments of this study during larva and juvenile intensive culture of northern pike (Esox lucius L.).
An Otohime dry starter (Marubeni Nisshin Feed, Tokyo, Japan), which is frequently and successfully used in larval pikeperch (Sander lucioperca), was used as a high-quality starter (Ljubobratovic et al., 2017; Kucera et al., submitted). Depending on the larval stage, different Otohime sizes were used: B1 (250–360 µm) for days 1–5, B2 (360–650 µm) for days 6–10, and C1 (580–840 µm) for days 11–18; the nutrient content was as follows: 55.8%–55.1% protein, 14.3%–14.9% fat, 2.8%–2.9% fiber, 15.0%–16.4% ash, 2.5%–2.9% calcium, and 2.2%–2.4% phosphorus, vitamin A 10,000 IU kg−1, vitamin D3 2,000 IU kg−1, vitamin E 1,250 mg kg−1, and Cu 7.0 mg kg−1, (Marubeni Nisshin feed Co. Ltd., Japan), which was applied by hand at 15-min intervals during the light regime of the dry feeding period of all three experiments.
2.3 Experimental groups, design, and aim of each experiment
2.3.1 Effects of weaning time on efficiency of pike larvae and early juvenile intensive culture (Experiment I)
The first experiment tested the effects of weaning time on the growth, condition, survival, and cannibalism rates of northern pike larvae and early juveniles. The experiment comprised four experimental groups, with four replicates tested for 18 days from the first day of exogenous feeding of pike larvae. Larvae were fed sequential combinations of live feed (LF), co-feeding with dry starter (DS), and only DS feed for 18 days as follows: 9 days LF followed by 3 days co-feeding and 5 days DS (group A); 6 days LF followed by 3 days co-feeding and 9 days DS (group B); 3 days LF followed by 3 days co-feeding and 12 days DS (group C); and DS alone for 18 days (group D). Pond-cultured, harvested, and ozone-treated zooplankton instars and adult Daphnia sp. (with body size 200–3,500 µm) were used as natural feed. At the beginning of live feed, daily rate was 35% of fish biomass updated every 3 days by checking the fish growth rate. The co-feeding with Daphnia sp. (90%–60%–30%) and dry starter (10%–40%–70%) at a proportion of 20% of fish biomass was applied for 3 days. The pike juveniles were fed a dry starter (daily rate ad libitum). A schematic of Experiment I is shown in Figure 1. In total, 57,600 larvae were stocked (counted in a volumetric manner) in all 16 tanks at an initial density of 20 larvae/L. The initial BW (8.9 ± 1.21 mg) and TL (10.8 ± 0.70 mm) were measured using a stereo microscope SMZ75T (Nikon, Tokyo, Japan) with Quick PHOTO MICRO 3 and weighted using the analytical scale Mettler – model AE 200 in 300 larvae on stocking day (13 DPH) and 30 larvae from each group (10 per replicate) 3, 6, 9, 12, 15, and 18 days after stocking. All larvae were randomly sampled and anesthetized with MS-222 (tricaine methane sulfonate, Sigma-Aldrich, MO, USA; 100 mg L−1) prior to handling. In addition, 90 larvae per group (30 per replicate) were individually measured and weighed under anesthesia as described above, and the total deformity rate, including lordosis, scoliosis, and zigzag-shaped deformity, was evaluated in all fish measured by one expert according to Policar et al. (2016) at the end of the trial. The tanks were cleaned twice daily (7:00 a.m. and 2:30 p.m.) by siphoning all dead fish, feces, and uneaten food. Mortality was recorded daily by removing dead fish during the cleaning procedure and calculating the cannibalism rate.
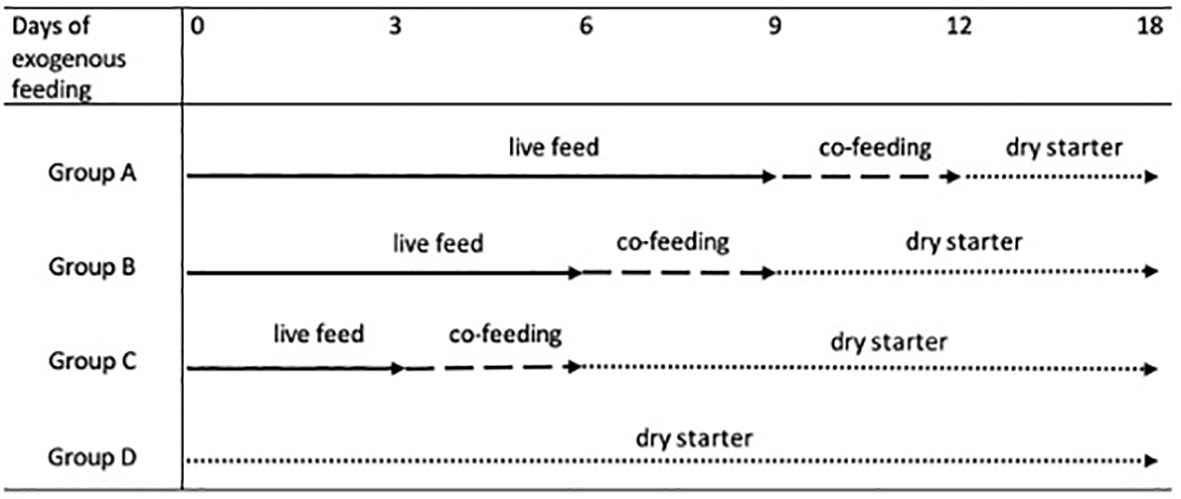
Figure 1 The schedule of the first experiment with the different weaning time of northern pike (Esox lucius) larval and early juvenile intensive culture: feeding with live zooplankton instars and adult of Daphnia sp., co-feeding (mix of zooplankton and dry starter - Otohime), and feeding with dry starter Otohime.
Growth, condition, survival, cannibalism, and deformity rates were calculated using the following formulas:
where BW (g) is the average BW; TL (cm) is the average TL; σ is the standard deviation of BW; μ is the arithmetic mean of BW; lnBWI and lnBWF are natural logarithms for the initial and final BWs at the beginning and end of the experiment, respectively; d (days) is the duration of the experiment; NF (pcs) is the final number of produced fish; NI (pcs) is the initial number of stocked fish; RM is the recorded mortality (number of dead fish); and TDR (pcs) is the number of all deformed produced fish.
2.3.2 Effects of light regime on efficiency of pike larvae and early juvenile intensive culture (Experiment II)
The second experiment tested the effects of different light regimes on the growth, condition, survival, cannibalism, and deformity rates of northern pike larvae and early juveniles using 57,600 larvae at 13 DPH (mean BW = 11.5 ± 1.55 mg, TL= 12.5 ± 0.81 mm). Fish were randomly divided into eight experimental groups with two repetitions under the same initial stocking density (20 larvae/L) as in the first experiment. Eight experimental groups were prepared using the light regimes listed in Table 2. During this experiment, only the dry starter Otohime was applied by hand at 15-min intervals during the light period with daily rate ad libitum.
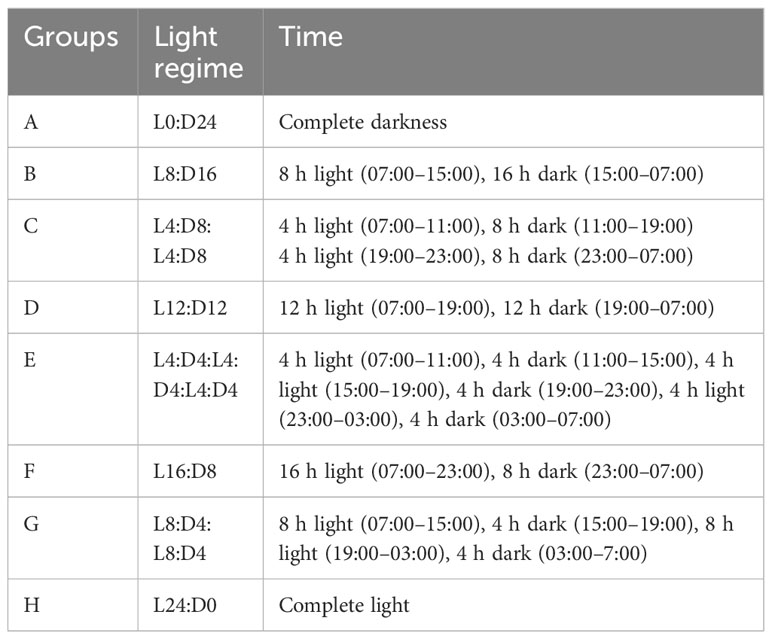
Table 2 Experimental groups presenting different light regimes during the second experiment of northern pike (Esox lucius L.) larval and early juvenile culture under RAS.
The same daily procedures related to tank cleaning and water quality monitoring were performed during this experiment, according to the routines of Experiment I. The same initial and final measurements and weights of TL and BW of the same number of larvae at 13 DPH and at the end of the experiment were also applied to calculate and compare the growth, condition, survival, cannibalism, and deformity rates among the tested groups.
2.3.3 Effects of the initial stocking density on efficiency of pike larvae and early juvenile intensive culture (Experiment III)
At the beginning of the third experiment, the same number of larvae (57,600 ind with initial BW = 9.00 ± 0.8 mg, TL = 10.8 ± 0.4 mm) were randomly stocked to the four tested groups with four repetitions (16 tanks in total) using the same RAS. The four experimental groups were assigned to four initial stocking densities: 10 ind L−1, 20 ind L−1, 40 ind L−1, and 80 ind L−1. The duration of this trial was 13 days until the juvenile age at 27 DPH. All procedures in Experiment III were performed identically to those in Experiment II with the same measurements and parameters at the end of the trial.
2.4 Statistical analysis
Data analysis for all three experiments was performed using one-way analysis of variance (ANOVA) in Statistica version 13 for Windows (StatSoft CR s.r.o., Prague, Czech Republic). The homogeneity of variance and normality of distribution were evaluated using Levene’s test. Post hoc comparisons of means were conducted using the Tukey HSD method when ANOVA detected significant differences among groups, with statistical significance set at p< 0.05. When the data did not meet the parametric assumptions, the Kruskal–Wallis test was used for global analyses and Dunn’s test was used for pairwise comparisons. All data are presented as the mean ± standard deviation.
3 Results
3.1 Effects of weaning time on efficiency of pike larvae and early juvenile intensive culture (Experiment I)
The effects of the first exogenous feeding regime on the growth performance of pike larvae and early juveniles are shown in Table 3. Fish fed live feed had significantly higher growth rates than those fed a dry diet (p< 0.05). Extending the feeding period to live feeding resulted in a significantly higher growth rate (24.6% d−1), variability (13.5%), and heterogeneity (299.5% d−1), whereas the opposite was observed for fish fed the dry starter. A similar trend was observed for FC values, where the group offered only the dry starter exhibited the lowest FC value. However, the highest survival rate (49%) and lowest cannibalism rate (21%) were recorded in fish fed solely dry starter (p< 0.05). BW was significantly affected by the weaning time over time (Figure 2). A detailed comparison of the size and condition of the northern pike larvae and juveniles for 18 days is presented in Table 4.
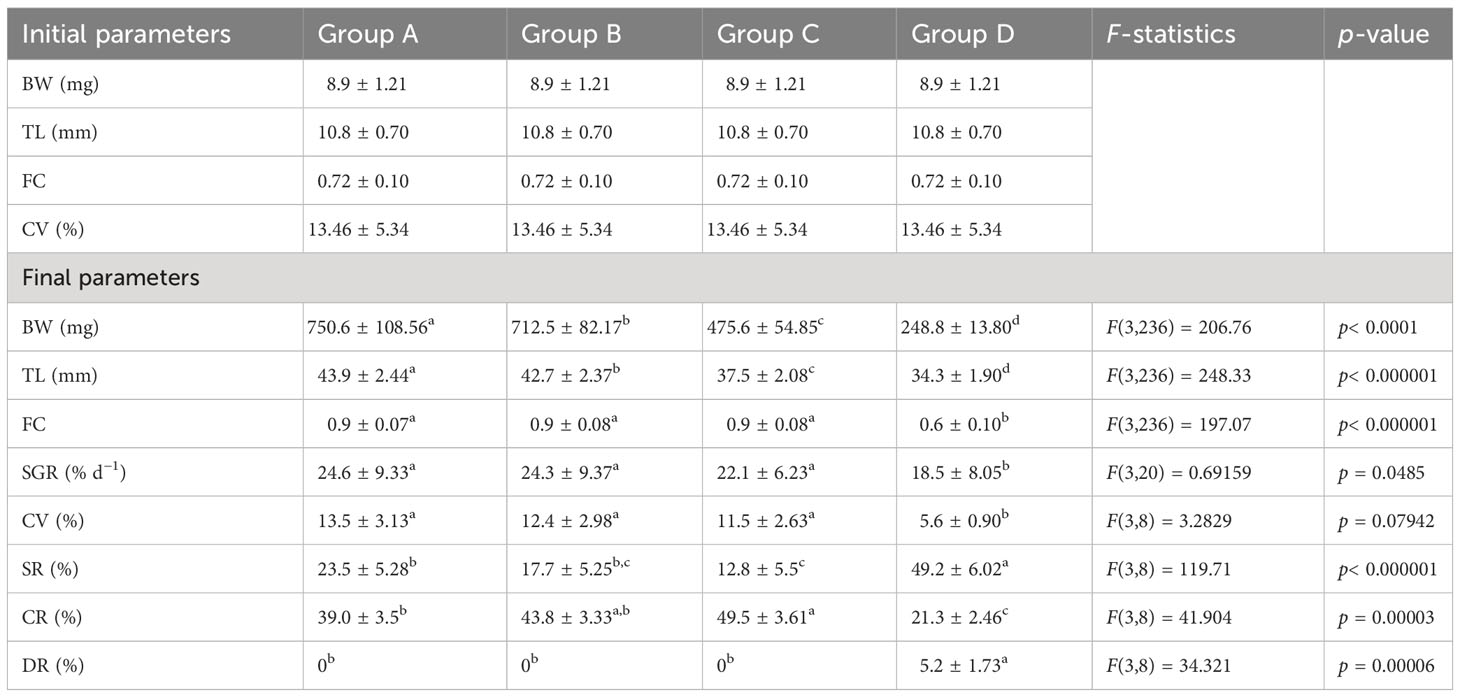
Table 3 Comparison of growth, condition, survival, cannibalism, and deformity rates between different weaning times of northern pike (Esox lucius L.) larvae and juveniles for the 18-day experiment (Experiment I).
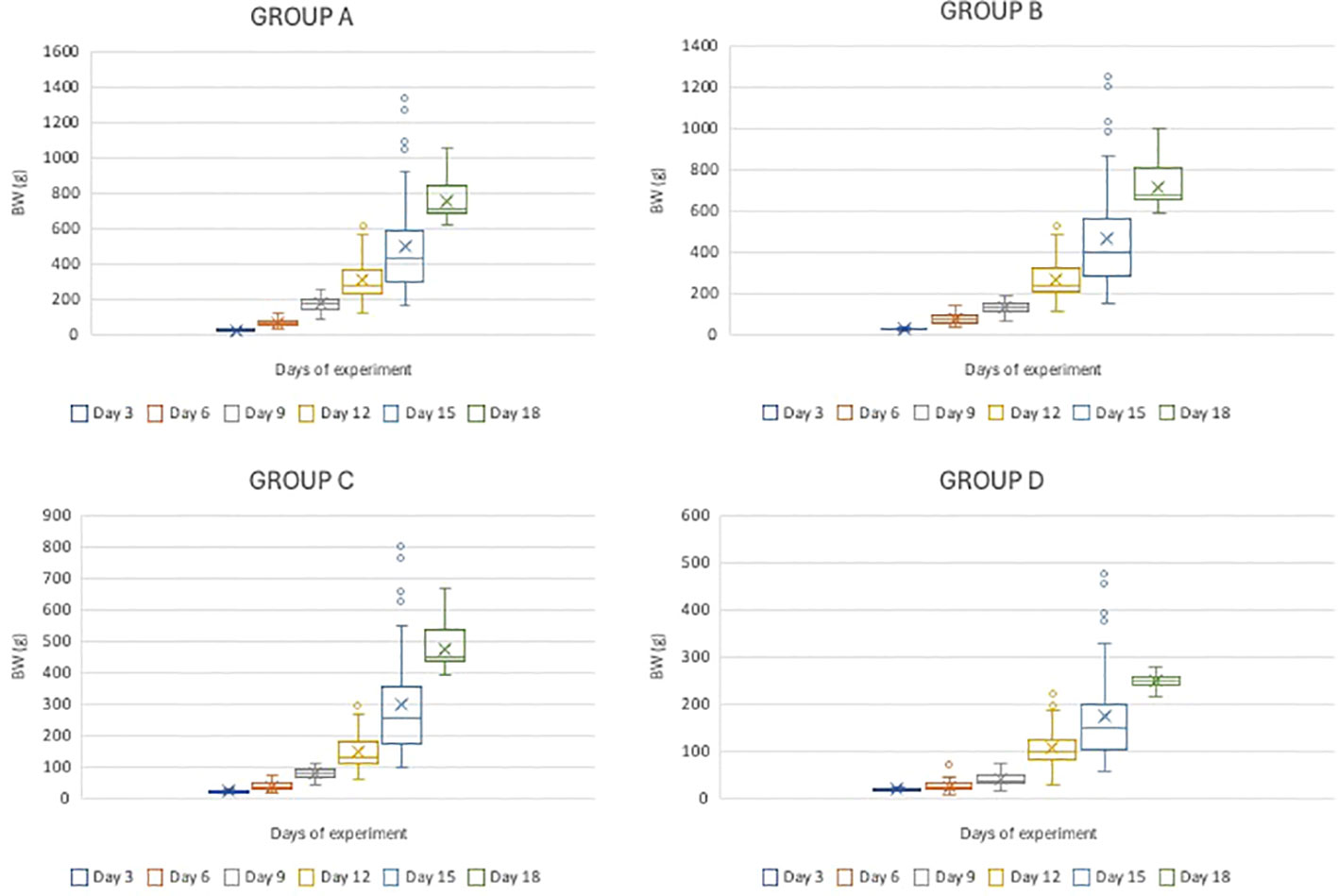
Figure 2 The comparison of body weight (g) of northern pike (Esox lucius L.) larvae and juveniles for the 18-day period (Experiment I).
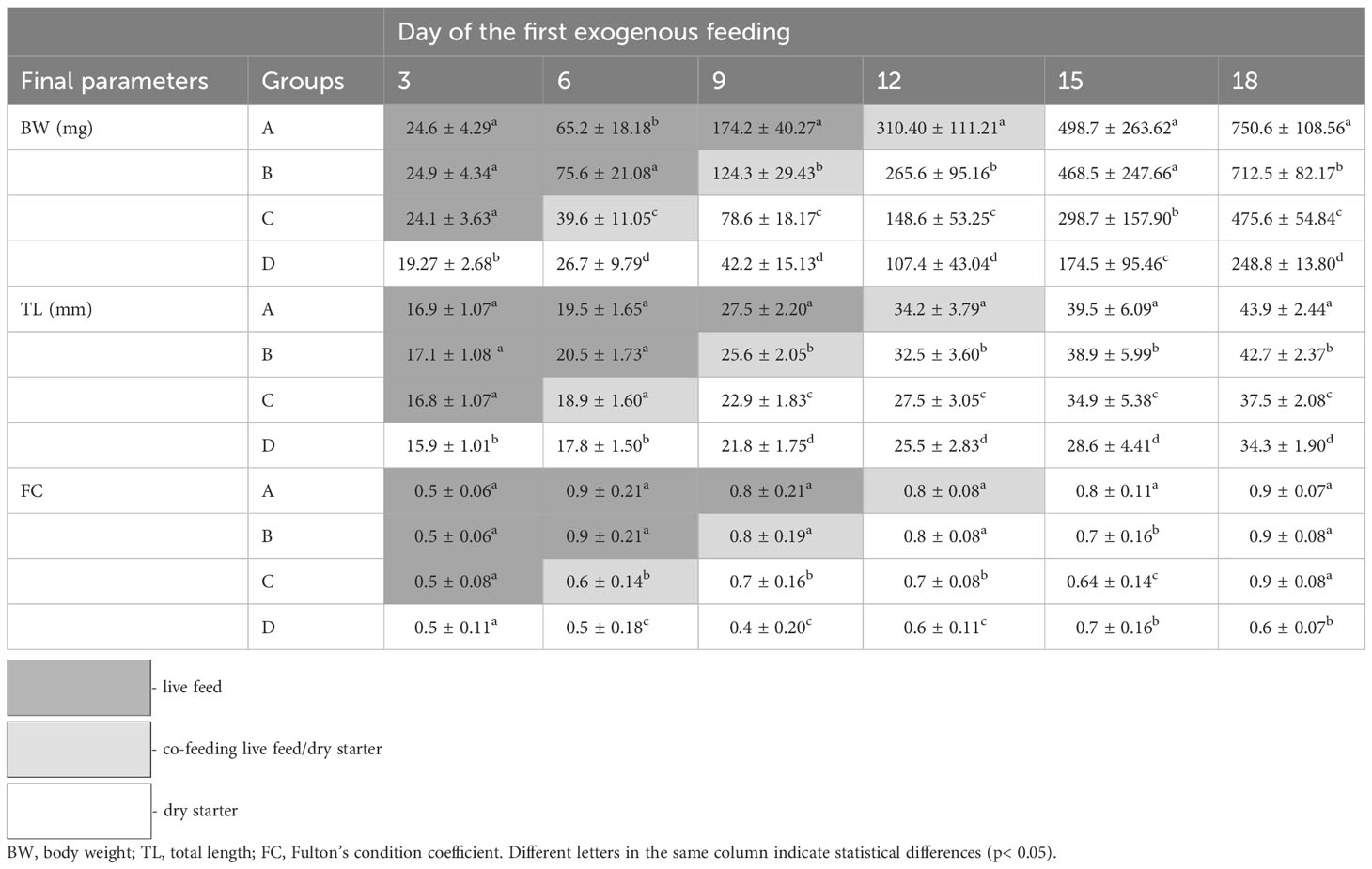
Table 4 Comparison of detailed size and condition of northern pike (Esox lucius L.) larvae and juveniles for the 18-day period (Experiment I).
3.2 Effects of light regime on efficiency of pike larvae and early juvenile intensive culture (Experiment II)
Groups exposed to complete light (L24:D0) and 16 h of light (L16:D8) exhibited significantly higher weight gain and SGR (%) than the other groups (Table 5). The highest survival rate was observed in fish reared with complete light (68.5%) and under L8:D4:L8:D4 light regimes (61.4%), whereas the highest mortality was in fish reared under L4:D8:L4:D8 light regimes. Cannibalism was significantly affected by photoperiod. The cannibalism rate was significantly higher in fish reared under 4 h light and 4 h dark (L4:D8:L4:D8), and the lowest rate was observed in fish reared with complete light and 16 h light (L16:D8; L8:D4:L8:D4).
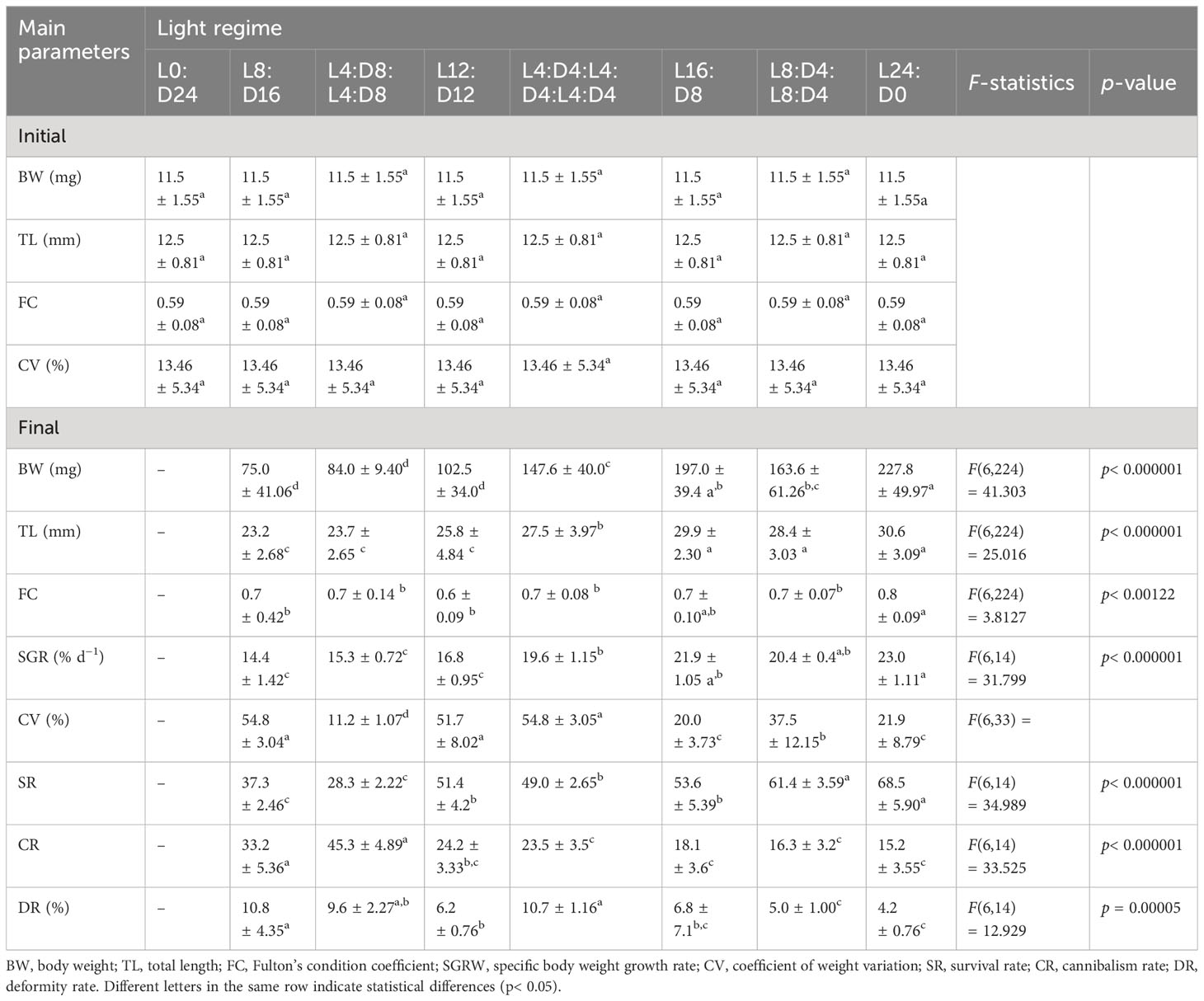
Table 5 Comparison of growth, condition, survival, cannibalism, and deformity rates between different light regimes of northern pike (Esox lucius L.) larvae and juveniles during the 13-day experiment (Experiment II).
3.3 Effects of the initial stocking density on efficiency of pike larvae and early juvenile intensive culture (Experiment III)
The results showed significant effects of stocking density on the growth, survival, and cannibalism rates of pikeperch juveniles (p< 0.05; Table 6). Fish with densities of 20 and 40 ind L−1 had significantly higher BWs (205.4–218.6 mg), TL (30.5–31.6 mm), SGR (24.1–24.5% d−1), and SR (58.5%–61.1%) than those in the other groups. The survival rates at the end of the experiment were 72% and 53% for larvae at densities of 20 ind L−1 and 10 ind L−1, respectively. These values were significantly different (p< 0.05). Fish at densities of 10 ind L−1 and 80 ind L−1 showed significantly higher cannibalism rates than those in the other groups. The highest FC value (0.7) and the lowest cannibalism rate (8.5%) were found in groups H40 and H20, respectively. Other parameters such as CV and DR were unaffected by the initial stocking larval density.
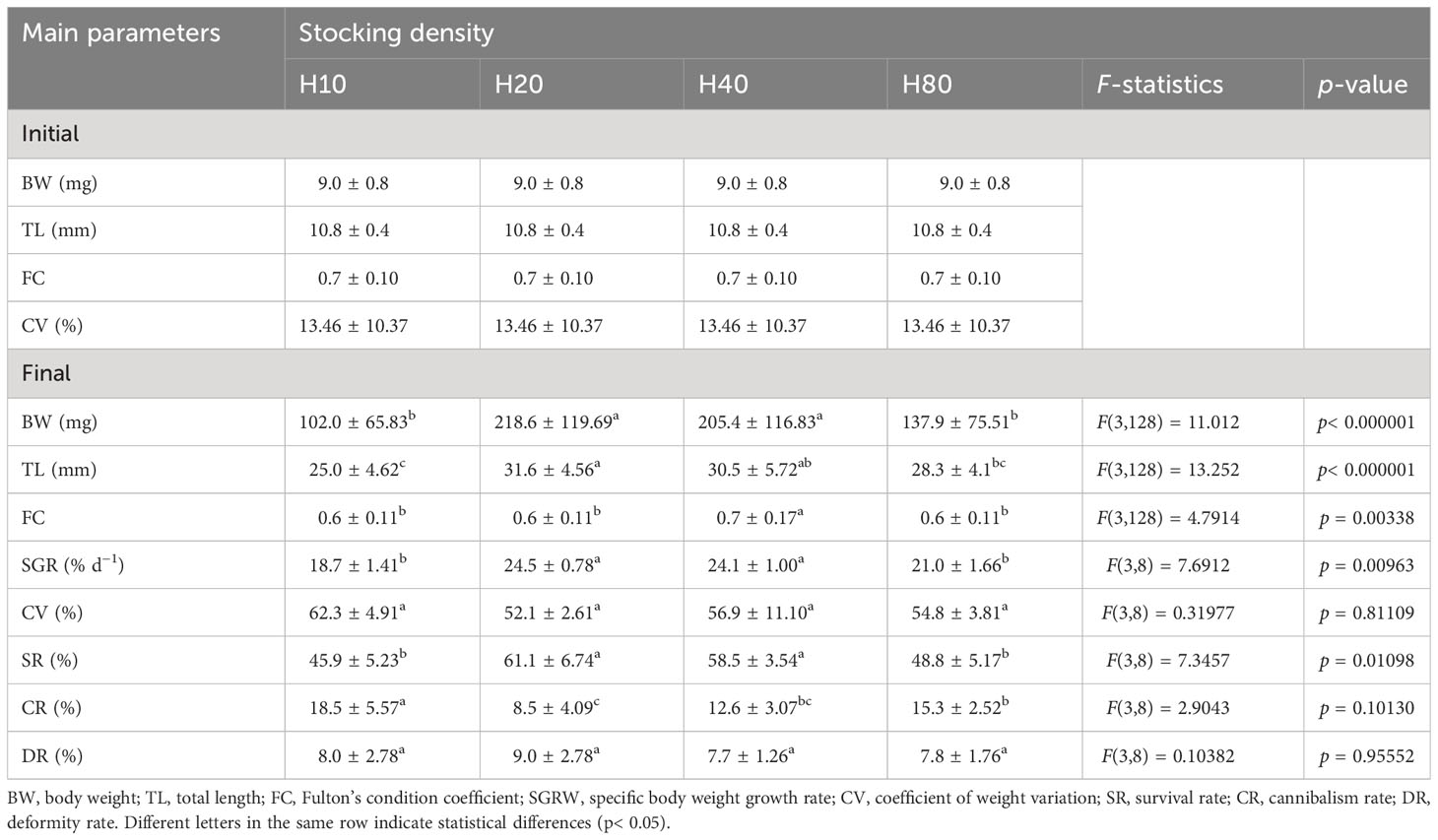
Table 6 Comparison of final growth, condition, survival, cannibalism, and deformity rates between different stocking densities of northern pike (Esox lucius L.) larvae and juveniles during the 13-day experiment (Experiment III).
4 Discussion
European freshwater inland aquaculture requires continuous technological developments for species and product diversification. Currently, most produced freshwater fish species in Europe are common carp (Cyprinus carpio) and rainbow trout (Oncorhynchus mykiss), which account for >70% of fish freshwater production in Europe (Policar and Adámek, 2013). Continuous and stable controlled reproduction and intensive farming of highly demanding carnivorous species, such as the Eurasian perch (Perca fluviatilis), pikeperch (S. lucioperca), northern pike (E. lucius L.), burbot (Lota lota), and European catfish (Silurus glanis), can substantially increase freshwater fish production and consumption in Europe, resulting in less independence on exports from Asia (Wocher et al., 2012; Bondarenko et al., 2015b; Nebeský et al., 2016; Khendek et al., 2017; Linhart et al., 2020).
In this study, differences in the body size of northern pike larvae were identified during different periods of its reproductive season, with the largest larvae being produced in the middle of the reproductive season. The same trend was found in the common barbel (Policar et al., 2010), which reached the largest fish size in the middle of the reproductive season compared to the beginning and end of the season. Moreover, the quality of hatched common barbel larvae (checked for osmotic shock resistance) was highest in the middle of the reproductive season. However, this statement was not confirmed in this study because only high-quality northern pike larvae were used for all three experiments, and no effect on this parameter was found during the reproductive season of the northern pike. The different spawning periods during the reproductive season affected sperm quality in male common barbel and northern pike (Alavi et al., 2008; Bondarenko et al., 2018). Bondarenko et al. (2018) reported the highest percentage of northern pike sperm at the end of the reproductive season and the highest sperm velocity at 30 and 45 s post-activation in the middle and at the end of the reproductive season, respectively. Alavi et al. (2008) reported the highest percentage of motile sperm and highest velocity in the middle or beginning of the reproductive season in male common barbel.
4.1 Effects of the weaning time on efficiency of pike larvae and early juvenile intensive culture (Experiment I)
Weaning time is crucial for larval rearing. Fish farms are interested in introducing inert diets as early as possible into larval cultures to achieve full replacement of live diets. In the present study, weaning time significantly affected the growth of pike larvae. Upon completion of the experiment, fish fed live feed showed significantly higher growth rates compared to fish fed solely a starter diet, indicating that an artificial diet is currently unsuitable for first feeding larvae. This result is in agreement with previous studies on Vimba vimba (Hamáčková et al., 2009), Clarias gariepinus (Appelbaum and Mcgeer, 1998), C. carpio (Szlamińska and Przybył, 1986), Pleteobagrus fulvidraco (Wang et al., 2005), Clarias batrachus (Giri et al., 2002), and O. mykiss (Akbary et al., 2010). Moreover, there were significantly higher numbers of larvae in fish fed live feed than in those fed artificial feed, 3 days after initiating feeding. Although there was significantly lower growth in larvae fed only the starter diet, the survival rates in this group were the highest. The difference in survival rate observed between fish fed the starter diet alone and those fed live feed was associated with the cannibalism rate. Surprisingly, northern pike displayed the lowest cannibalism rate in fish fed only a starter diet (21%), whereas the highest cannibalism rate (49.5%) was seen in group C under early weaning (day 3), with a survival rate of 13%. This is possibly because larvae fed a starter diet grew slower without developing large differences in size, allowing for higher survival rates. This study indicates that northern pike larvae require at least 9 days of live feed feeding before switching to co-feeding and then to artificial diets.
4.2 Effects of light regime on efficiency of pike larvae and early juvenile intensive culture (Experiment II)
Photoperiod is an important factor that influences the growth and survival of fish larvae (Fielder et al., 2002; Villamizar et al., 2011). Similar results have been reported for fish larvae, in which the growth rates improved with increasing light periods. The growth of Sparus aurata (Tandler and Helps, 1985) and Siganus guttatus (Duray and Kohno, 1988) larvae was significantly higher under the 24-h light regime. In contrast, increased photoperiod decelerated growth and reduced survival in Dicentrarchus labrax and Archosargus rhomboidalis, possibly due to higher energy expenditure (Barahona-Fernandes, 1979; Dowd and Houde, 1980).
In the present study, the growth of pike larvae in the complete light regime was significantly different from that of larvae reared in complete darkness. The highest survival rate was observed in fish reared under complete light (68.5%) and the L8:D4:L8:D4 light regime (61.4%). Under 24 h of the dark regime, pike larvae mortality reached 100%. The same trend has been observed in previous studies on fish larvae, as demonstrated by decreased growth performance and complete mortality (Fielder et al., 2002; Villamizar et al., 2009; Shi et al., 2010). A possible explanation for mortality under complete darkness is that larvae did not establish first feeding and required light to start feeding. The better growth and survival of pike larvae during longer light periods in the present study is possibly because, during the long light phases, sufficient time was available to establish a robust rhythmicity, and larvae learned quicker how to catch prey (Barlow et al., 1995; Fielder et al., 2002). Cannibalism was significantly affected by photoperiod. Longer light periods also had a positive impact on the cannibalism rate. The current study revealed a low cannibalism rate in larvae reared under complete light and 16-h light groups (L16:D8; L8:D4:L8:D4). The low cannibalism rate observed in the present study was related to the high survival rate of pike larvae.
4.3 Effects of the initial stocking density on efficiency of pike larvae and early juvenile intensive culture (Experiment III)
The optimal density varies among species and depends on the developmental stage of the fish (Baldwin, 2010). Our results demonstrated that stocking density significantly affected pike larvae (p< 0.05; Table 6). Larvae reared at low and high stocking densities had significantly lower BWs, SGR, and survival rates than those reared at medium densities. The negative correlation between growth and high stocking density in pikes is consistent with previous studies on fish larvae (Sharma and Chakrabarti, 2003; Bolasina et al., 2006) and can be explained by several factors. An increase in stocking density results in a lower amount of food being consumed by the fish (UEMATSU and Ogawa, 1975); deteriorated water quality (Kebus et al., 1992) and higher cortisol levels (Barnett and Pankhurst, 1998; Montero et al., 1999; Bolasina et al., 2006) lead to decreased growth rates. In addition, a high stocking density results in a greater probability of encounter between a cannibal and potential prey (Baras et al., 2003), allowing a few large larvae to dominate the area and become cannibals. The results of cannibalism in the present study clearly indicate a density-dependent cannibalism rate in pike larvae. Fish at densities of 10 and 80 ind L−1 showed significantly higher cannibalism rates than those in the other groups. This study revealed that a decrease in stocking density resulted in a high cannibalism rate. Similarly, lower growth rates at lower densities were observed in the juvenile African catfish, C. gariepinus (Hecht and Uys, 1997), and according to the authors’ observations, fish at lower densities most likely maintain the territory rather than looking for food.
5 Conclusion
The results of the current study indicated successful rearing of pike larvae from the standpoint of growth performance, survival rate, and cannibalism rate with weaning times as follows: 9 days live feed followed by 3 days co-feeding and 5 days dry starter, at a 24-h complete light regime and a stocking density of 20 ind L−1. These results have significant applications in pike aquaculture using indoor recirculating systems, as they improve our understanding of the roles of weaning time, photoperiod, and stocking density in fish growth and survival. Future work is needed to establish optimal weaning periods, photoperiod, and stocking density using advanced molecular techniques and histology.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the permission of the Departmental Expert Committee for the Authorization of Experimental Projects of the Ministry of Education, Youth and Sports of the Czech Republic (permit no. MSMT-8155/2022-4). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AI: Formal Analysis, Writing – original draft, Writing – review & editing. VB: Data curation, Writing – review & editing. TPě: Formal analysis, Writing – review & editing. TPo: Data curation, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The study was supported by the Ministry of Agriculture of the Czech Republic (project NAZV QK22020144) and by the Ministry of Education, Youth and Sports of the Czech Republic (project CENAKVA LM2018099).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akbary P., Hosseini S. A., Imanpoor M., Sudagar M., Makhdomi N. M. (2010). Comparison between live food and artificial diet on survival rate, growth and body chemical composition of Oncorhynchus mykiss larvae. Iran J. Fish Sci. 9, 19–32. doi: 10.22092/IJFS.2018.114080
Alavi S. M. H., Psenicka M., Rodina M., Policar T., Linhart O. (2008). Changes of sperm morphology, volume, density and motility and seminal plasma composition in Barbus bar bus (Teleostei: Cyprinidae) during the reproductive season. Aquat Living Resour 21, 75–80. doi: 10.1051/alr:2008011
Appelbaum S., Mcgeer J. C. (1998). Effect of diet and light regime on growth and survival of African catfish (Clarias gariepinus) larvae and early juveniles. Aquac Nutr. 4, 157–164. doi: 10.1046/j.1365-2095.1998.00064.x
Baldwin L. (2010). The effects of stocking density on fish welfare. Plymouth Student Scientist 4, 372–383.
Barahona-Fernandes M. H. (1979). Some effects of light intensity and photoperiod on the sea bass larvae (Dicentrarchus labrax (L.)) reared at the Centre Oceanologique de Bretagne. Aquaculture 17, 311–321. doi: 10.1016/0044-8486(79)90086-3
Baras E., Kestemont P., Mélard C. (2003). ). Effect of stocking density on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions. Aquaculture 219, 241–255. doi: 10.1016/S0044-8486(02)00349-6
Barlow C. G., Pearce M. G., Rodgers L. J., Clayton P. (1995). Effects of photoperiod on growth, survival and feeding periodicity of larval and juvenile barramundi Lates calcarifer (Bloch). Aquaculture 138, 159–168. doi: 10.1016/0044-8486(95)01073-4
Barnett C. W., Pankhurst N. W. (1998). The effects of common laboratory and husbandry practices on the stress response of greenback flounder Rhombosolea tapirina (Gunther 1862). Aquaculture 162, 313–329. doi: 10.1016/S0044-8486(98)00202-6
Bolasina S., Tagawa M., Yamashita Y., Tanaka M. (2006). Effect of stocking density on growth, digestive enzyme activity and cortisol level in larvae and juveniles of Japanese flounder, Paralichthys olivaceus. Aquaculture 259, 432–443. doi: 10.1016/j.aquaculture.2006.05.021
Bondarenko V., Blecha M., Policar T. (2018). Changes of sperm morphology, volume, density, and motility parameters in northern pike during the spawning period. Fish Physiol. Biochem. 44, 1591–1597. doi: 10.1007/s10695-018-0546-5
Bondarenko V., Drozd B., Policar T. (2015a). Effect of water temperature on egg incubation time and quality of newly hatched larvae of northern pike (Esox lucius L). J. Appl. Ichthyology 31, 45–50. doi: 10.1111/jai.12851
Bondarenko V., Podhorec P., Svinger V. W., Policar T. (2015b). Evaluation of treatments for induction of ovulation in northern pike (Esox lucius L.). Turk J. Fish Aquat Sci. 15, 575–581. doi: 10.4194/1303-2712-v15_2_46
Craig J. F. (2008). A short review of pike ecology. Hydrobiologia 601, 5–16. doi: 10.1007/s10750-007-9262-3
Dowd C., Houde E. (1980). Combined effects of prey concentration and photoperiod on sirvival and growth of larval sea bream archosargus rhomboidalis (Sparidae). Mar. Ecol. Prog. Ser. 3, 181–185. doi: 10.3354/meps003181
Duray M., Kohno H. (1988). Effects of continuous lighting on growth and survival of first-feeding larval rabbitfish, Siganus guttatus. Aquaculture 72, 73–79. doi: 10.1016/0044-8486(88)90147-0
Fielder D. S., Bardsley W. J., Allan G. L., Fielder D. S., Pankhurst P. M. (2002). Effect of photoperiod on growth and survival of snapper Pagrus auratus larvae. Aquaculture 211, 135–150. doi: 10.1016/S0044-8486(02)00006-6
Giles N., Wright R. M., Nord M. E. (1986). Cannibalism in pike fry, Esox lucius L.: some experiments with fry densities. J. Fish Biol. 29, 107–113. doi: 10.1111/j.1095-8649.1986.tb04930.x
Giri S. S., Sahoo S. K., Sahu B. B., Sahu A. K., Mohanty S. N., Mukhopadhyay P. K., et al. (2002). Larval survival and growth in Wallago attu (Bloch and Schneider): Effects of light, photoperiod and feeding regimes. Aquaculture 213, 151–161. doi: 10.1016/S0044-8486(02)00012-1
Hadi Alavi S. M., Rodina M., Viveiros A. T. M., Cosson J., Gela D., Boryshpolets S., et al. (2009). Effects of osmolality on sperm morphology, motility and flagellar wave parameters in Northern pike (Esox lucius L.). Theriogenology 72, 32–43. doi: 10.1016/j.theriogenology.2009.01.015
Hamáčková J., Prokeš M., Kozák P., Peňáz M., Stanny L. A., Policar T., et al. (2009). Growth and development of vimba bream (Vimba vimba) larvae in relation to feeding duration with live and/or dry starter feed. Aquaculture 287, 158–162. doi: 10.1016/j.aquaculture.2008.10.059
Hecht T., Uys W. (1997). Effect of density on the feeding and aggressive behaviour in juvenile African catfish, Clarias gariepinus. S Afr J. Sci. 93.
Hubenova T., Zaikov A., Vasileva P. (2007). Investigation on fecundity, follicles and free embryo size of pond-reared pike (Esox lucius) of different age and size. in Aquaculture International. 15 (3), 235–240. doi: 10.1007/s10499-007-9077-2
Hulak M., Rodina M., Linhart O. (2008). Characteristics of stripped and testicular Northern pike (Esox lucius) sperm: Spermatozoa motility and velocity. Aquat Living Resour 21, 207–212. doi: 10.1051/alr:2008022
Jankowska B., Zakś Z., Zmijewski T., Szczepkowski M. (2008). Fatty acid composition of wild and cultured northern pike (Esox lucius). J. Appl. Ichthyology 24, 196–201. doi: 10.1111/j.1439-0426.2007.01011.x
Kebus M. J., Collins M. T., Brownfield M. S., Amundson C. H., Kayes T. B., Malison J. A. (1992). Effects of rearing density on the stress response and growth of rainbow trout. J. Aquat Anim. Health 4, 1–6. doi: 10.1577/1548-8667(1992)004<0001:EORDOT>2.3.CO;2
Khendek A., Alix M., Viot S., Ledoré Y., Rousseau C., Mandiki R., et al. (2017). How does a domestication process modulate oogenesis and reproduction performance in Eurasian perch? Aquaculture 473, 206–214. doi: 10.1016/j.aquaculture.2017.02.003
Kozłowski M., Piotrowska I. (2022). Effect of size grading on growth, survival, and cannibalism in larval and juvenile pike, Esox lucius (L.), reared in recirculating systems. Aquaculture Int. 30, 2231–2244. doi: 10.1007/s10499-022-00899-3
Kristan J., Samarin A. M., Malinovskyi O., Policar T. (2020). Gamete management for artificial reproduction of northern pike Esox lucius (Linnaeus 1758). Aquaculture 528, 735575. doi: 10.1016/j.aquaculture.2020.735575
Kucharczyk D., Mamcarz A., Kujawa R., Skrzypczak A. (1998). Development of cannibalism in larval northern pike, esox lucius (Esocidae). Ital. J. Zoology 65, 261–263. doi: 10.1080/11250009809386829
Kucska B., Müller T., Bercsényi M. (2007). The effect of feeding frequency on the growth and survival of pike (Esox lucius L.) using floating pellets. J. Appl. Ichthyology 23, 193–194. doi: 10.1111/j.1439-0426.2006.00809.x
Kucska B., Pál L., Müller T., Bódis M., Bartos Á., Wágner L., et al. (2006a). Changing of fat content and fatty acid profile of reared pike (Esox lucius) fed two different diets. Aquac Res. 37, 96–101. doi: 10.1111/j.1365-2109.2005.01402.x
Kucska B., Pál L., Müller T., Bódis M., Bartos Á., Wágner L., et al. (2006b). Changing of fat content and fatty acid profile of reared pike (Esox lucius) fed two different diets. Aquac Res. 37, 96–101. doi: 10.1111/j.1365-2109.2005.01402.x
Larsson P., Tibblin P., Koch-Schmidt P., Engstedt O., Nilsson J., Nordahl O., et al. (2015). Ecology, evolution, and management strategies of northern pike populations in the Baltic Sea. Ambio 44, 451–461. doi: 10.1007/s13280-015-0664-6
Lehtonen H., Leskinen E., SeléN. R., Reinikainen M. (2009). Potential reasons for the changes in the abundance of pike, Esox lucius, in the western Gulf of Finland 1939-2007. Fish Manag Ecol. 16, 484–491. doi: 10.1111/j.1365-2400.2009.00701.x
Linhart O., Cheng Y., Rodina M., Gela D., Tučková V., Shelton W. L., et al. (2020). AQUA_2020_1080: Sperm management of European catfish (Silurus glanis L.) for effective reproduction and genetic conservation. Aquaculture 529, 735620. doi: 10.1016/j.aquaculture.2020.735620
Ljubobratovic U., Kosanovic D., Vukotic G., Molnar Z., Stanisavljevic N., Ristovic T., et al. (2017). Supplementation of lactobacilli improves growth, regulates microbiota composition and suppresses skeletal anomalies in juvenile pike-perch (Sander lucioperca) reared in recirculating aquaculture system (RAS): A pilot study. Res. Vet. Sci. 115, 451–462. doi: 10.1016/j.rvsc.2017.07.018
Lucas M. C. (1996). Pike: biology and exploitation. Fish Res. 28, 240–242. doi: 10.1016/0165-7836(96)82494-5
Montero D., Izquierdo M. S., Tort L., Robaina L., Vergara J. M. (1999). High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 20, 53–60. doi: 10.1023/A:1007719928905
Nebeský V., Policar T., Blecha M., Křišťan J., Svačina P. (2016). Trends in import and export of fishery products in the Czech Republic during 2010–2015. Aquaculture Int. 24, 1657–1668. doi: 10.1007/s10499-016-0066-1
Pěnka T., Malinovskyi O., Křištan J., Imentai A., Policar T. (2021). Effect of density and mixed culture of largemouth bass (Micropterus salmoides) and pikeperch (Sander lucioperca) on growth, survival and feed conversion rate in intensive culture. Czech J. Anim. Sci. 66 (10), 428–440. doi: 10.17221/59/2021-CJAS
Policar T., Blecha M., Křišťan J., Mráz J., Velíšek J., Stará A., et al. (2016). Comparison of production efficiency and quality of differently cultured pikeperch (Sander lucioperca L.) juveniles as a valuable product for ongrowing culture. Aquaculture International. 24, 1607–1626.
Policar T., Podhorec P., Stejskal V., Hamackova J., Alavi S. M. H. (2010). Fertilization and hatching rates and larval performance in captive common barbel (Barbus barbus L.) throughout the spawning season. J. Appl. Ichthyology 26, 812–815. doi: 10.1111/jai.2010.26.issue-5
Prejs A., Martyniak A., Boroń S., Hliwa P., Koperski P. (1994). Food web manipulation in a small, eutrophic Lake Wirbel, Poland: effect of stocking with juvenile pike on planktivorous fish. Hydrobiologia, 275–276. doi: 10.1007/BF00026700
Salam A., Davies P. M. C. (1994). Body composition of northern pike (Esox lucius L.) in relation to body size and condition factor. Fish Res. 19, 193–204. doi: 10.1016/0165-7836(94)90038-8
Samarin A. M., Blecha M., Uzhytchak M., Bytyutskyy D., Zarski D., Flajshans M., et al. (2016). Post-ovulatory and post-stripping oocyte ageing in northern pike, Esox lucius (Linnaeu), and its effect on egg viability rates and the occurrence of larval malformations and ploidy anomalies. Aquaculture 450, 431–438. doi: 10.1016/j.aquaculture.2015.08.017
Sharma J., Chakrabarti R. (2003). Role of stocking density on growth and survival of Catla, Catla catla, and rohu, Labeo rohita, larvae and water quality in a recirculating system. J. Appl. Aquaculture 14, 171–178. doi: 10.1300/J028v14n01_14
Shi Y., Zhang G., Zhu Y., Liu J. (2010). Effects of photoperiod, temperature, and salinity on growth and survival of obscure puffer Takifugu obscurus larvae. Aquaculture 309, 103–108. doi: 10.1016/j.aquaculture.2010.09.004
Szabó T. (2001). Hormonally induced ovulation of northern pike via sustained-release vehicles. N Am. J. Aquac 63, 137–148. doi: 10.1577/1548-8454(2001)063<0137:hioonp>2.0.co;2
Szabó T. (2003). Ovulation induction in northern pike Esox lucius L. using different GnRH analogues, Ovaprim, Dagin and carp pituitary. Aquac Res. 34, 479–486. doi: 10.1046/j.1365-2109.2003.00835.x
Szabó T. (2008). Use of Carbopol resin for carp pituitary administration improves the fertilization percentage of northern pike (Esox lucius Linnaeus) eggs in commercial hatcheries. Hydrobiologia 601, 91–97. doi: 10.1007/s10750-007-9269-9
Szczepkowski M. (2009). Impact of selected abiotic and biotic factors on the results of rearing juvenile stages of northern pike Esox lucius L. @ in recirculating systems. Arch. Polish Fisheries 17, 107–147. doi: 10.2478/v10086-009-0010-3
Szlamińska M., Przybył A. (1986). Feeding of carp (Cyprinus carpio L.) larvae with an artificial dry food, living zooplankton and mixed food. Aquaculture 54, 77–78. doi: 10.1016/0044-8486(86)90258-9
Tandler A., Helps S. (1985). The effects of photoperiod and water exchange rate on growth and survival of gilthead sea bream (Sparus aurata, Linnaeus; Sparidae) from hatching to metamorphosis in mass rearing systems. Aquaculture 48, 71–82. doi: 10.1016/0044-8486(85)90053-5
UEMATSU T., OGAWA S. (1975). Social facilitation of feeding behavior in fresh-water fish. (3) the himedaka, oryzias latipes. Annu. Anim. Psychol. 25, 57–64. doi: 10.2502/janip1944.25.57
Villamizar N., Blanco-Vives B., Migaud H., Davie A., Carboni S., Sánchez-Vázquez F. J. (2011). Effects of light during early larval development of some aquacultured teleosts: A review. Aquaculture 315, 86–94. doi: 10.1016/j.aquaculture.2010.10.036
Villamizar N., García-Alcazar A., Sánchez-Vázquez F. J. (2009). Effect of light spectrum and photoperiod on the growth, development and survival of European sea bass (Dicentrarchus labrax) larvae. Aquaculture 292, 80–86. doi: 10.1016/j.aquaculture.2009.03.045
Wang C., Xie S., Zheng K., Zhu X., Lei W., Yang Y., et al. (2005). Effects of live food and formulated diets on survival, growth and protein content of first-feeding larvae of Plelteobagrus fulvidraco. J. Appl. Ichthyology 21, 210–214. doi: 10.1111/j.1439-0426.2005.00620.x
Keywords: recirculating systems, growth performance, survival, fish density, photoperiod
Citation: Imentai A, Bondarenko V, Pěnka T and Policar T (2024) Effects of weaning time, light regime, and stocking density on growth, condition, survival, and cannibalism rates in northern pike (Esox lucius L.) larvae and early juveniles under intensive culture. Front. Mar. Sci. 11:1352699. doi: 10.3389/fmars.2024.1352699
Received: 08 December 2023; Accepted: 04 March 2024;
Published: 21 March 2024.
Edited by:
Seyyed Morteza Hoseini, Iranian Fisheries Science Research Institute (IFSRI), IranReviewed by:
Alessandra Roncarati, University of Camerino, ItalyMelika Ghelichpour, University of Tehran, Iran
Copyright © 2024 Imentai, Bondarenko, Pěnka and Policar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aiman Imentai, YWltZW50YWlAZnJvdi5qY3UuY3o=