 Lis Bittencourt1,2*
Lis Bittencourt1,2* Rafael Carvalho1Elitieri B. Santos-Neto1
Rafael Carvalho1Elitieri B. Santos-Neto1 Tatiana L. Bisi1,2
Tatiana L. Bisi1,2 Jose Lailson-Brito Jr.1,2Alexandre F. Azevedo1,2
Jose Lailson-Brito Jr.1,2Alexandre F. Azevedo1,2- 1Laboratorio de Mamiferos Aquaticos e Bioindicadores “Profa. Izabel M. G. do N. Gurgel” (MAQUA/UERJ), Faculdade de Oceanografia, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
- 2Programa de Pos-graduacao em Oceanografia, Faculdade de Oceanografia, Universidade do Estado do Rio de Janeiro (UERJ), Rio de Janeiro, Brazil
At present, there are still populations of different balaenopterids that have never been acoustically recorded and observed simultaneously. In an opportunistic sighting of dwarf minke whales during winter in southeastern Brazil, we have registered six individuals and recorded over 200 acoustic signals. Signals were quantified and had their acoustic parameters extracted; the calling rate was estimated as the number of sounds per minute, and the repetition rate was estimated as the number of calls of the same type emitted per minute. Four call types were described: Ba1, Ba2, Ba3 and Ba4. The most common and distinctive call was the Ba1, composed of three components: one had a peak frequency of 615.0 ± 189.8 Hz, and the other had peak frequencies of 1632.0 ± 191.5 and 5038.2 ± 195.1 Hz. Ba1 repetition varied from 2.4 to 11.1 repetitions/min. Ba2 was the second most common call with a peak frequency of 485.2 ± 421.9 Hz. The total calling rate varied from 4.4 to 11.1 calls/min. The signals are in the same frequency range reported for other areas but contain different structures.
1 Introduction
The dwarf minke whale, Balaenoptera acutorostrata, is a smaller form of the common minke whale that inhabits the waters of the Southern Hemisphere alongside the Antarctic minke whale, Balaenoptera bonaerensis (Perrin et al., 2018). Dwarf minke whale distribution covers the entire hemisphere, with most records coming from stranding data on the coasts of Brazil, South Africa, Chile, and Australia (Risch et al., 2019). In the Southwestern Atlantic Ocean, the majority of strandings occur during the winter, even though there are records in all seasons, and mostly between 33.45 ˚S and ˚22.93 S (Milmann et al., 2020). They have been sighted on 500m deep waters mostly during spring in the continental slope area (Di Tullio et al., 2016) and engaging in feeding behavior in an upwelling environment during summer (Hassel et al., 2003). However, live observation is still rare and underreported.
Minke whale sounds include pulse trains, down sweeps, and variable pulsed signals (Mellinger et al., 2000; Rankin and Barlow, 2005; Dominello and Sirović, 2016; Filun et al., 2020). Singing whales have been recorded in the Southern Hemisphere through passive acoustic monitoring in different areas. Signs attributed to minke whales were registered throughout the year in the South Pacific, where it peaked during July (Buchan et al., 2020). In the South Atlantic Ocean, signals attributed to minke whales are identified as produced by B. bonaerensis (Rossi-Santos et al., 2022; Filun and van Opzeeland, 2023). Complex sounds produced by dwarf minke whales have been recorded and characterized in Australian waters (Gedamke et al., 2001; Risch et al., 2019). Still, there is a large gap in our knowledge of the acoustic repertoire of this species throughout its distribution.
This study aimed to describe the acoustic signals opportunistically collected during an encounter with dwarf minke whales in a Southwestern Atlantic Ocean coastal area.
2 Method
The present study was an opportunistic observation of a single encounter with a group of dwarf minke whales during regular field surveys for cetacean sightings in Ilha Grande Bay, southeastern Brazil (Supplementary Figure 1 from the Supplementary material). Field surveys have occurred consistently since the year 2016 to register and monitor cetacean species in the area. There are no records in the literature of this species sighting in the area, and after the encounter, they have not been observed again since (Lailson-Brito et al., 2020).
The sighting occurred during a field survey on July 19th, 2022, with an outboard-powered boat of approximately 30 ft, and it lasted from 07:00 to 14:00. The first minke whale sighting occurred at approximately 09:00, and the monitoring continued until around 12:00 when the individuals started to disperse and spread over a large area. The local depth was about 30 m, approximately 9 km from the coast (23.09200 S, 44.47700 W). One group of minke whales was sighted; they were identified as the dwarf form of Balaenoptera acutorostrata due to their dorsal dark gray and ventral white coloration patterns (Figure 1), but especially the white patch on each flipper visible even from above the surface (Jefferson et al., 2011).

Figure 1 Underwater image of a dwarf minke whale highlighting the white patch on the flipper, taken during the sighting of a group of six individuals at Ilha Grande Bay, southeastern Brazil on July 19th, 2022.
The total number of whales in the field was six individuals. Visually comparing the animals size to the length of the survey boat, their total lengths were estimated to be around 7 m long, suggesting they were all adult individuals. During the observation time, the animals swam, interacting with each other. On one occasion, the six whales moved cohesively together nearby, but most of the time, the group was dispersed, interacting with three individuals. The Supplementary Material has a short video (S1) comprising aerial and underwater footage of the minke whales, taken through an unmanned aerial vehicle and a GoPro during the sighting.
During visual observation, underwater acoustic recordings were conducted. The recording equipment consisted of a digital recorder model SM4BatFS coupled to a hydrophone model HTI-99-UHF, the sampling rate was set at 192 kHz, and the recorder was scheduled to make continuous recordings of 2-minute files at a 0 dB gain. The hydrophone was kept at approximately four meters below the surface. During this period, the boats engine and the echo sounder remained turned off so they would not interfere with the recording. The total recording time was 40 minutes, broken into three sections: from 10:22 to 10:36 (00:17:00 duration), from 10:42 to 11:00 (00:17:44 duration), and from 11:25 to 11: 11:31 (00:06:25 duration). The intervals were due to relocating the boat to better observe the animals and obtain closer recordings. Within the recording time, there were moments in which the animals were so close that they swam beneath the boat and moments when they were further apart, but we estimate that during recordings, there were always at least three whales within a 500 m radius of the hydrophone. Due to our limitation of a single hydrophone, it was not possible to determine which whale was responsible for each call and the exact distance between individuals and the hydrophone.
Recordings were down-sampled to a 16 kHz sampling rate to obtain higher resolution at lower frequency bands. Sound analyses were performed with Raven Pro 1.6 (Cornell Lab of Ornithology, 2023), where spectrograms of 512 Hann window and 75% overlap were manually inspected to search for acoustic signals by an experienced observer. The acoustic environment of Ilha Grande Bay has been under regular monitoring since 2016 (Bittencourt et al., 2020; Lailson-Brito et al., 2020) as part of acoustic studies conducted along the Rio de Janeiro state coast since 2011. In the recording, there were sounds identified as produced by fishes, due to their characteristic pulsed and rhythmic nature, and due to having been previously encountered at other sites in the bay where soniferous fish species are typically encountered (Bittencourt et al., 2020). Constant noise from the shipping lane and ship mooring areas adjacent to the recording sites was also present, which is common for this portion of the bay (Bittencourt et al., 2020). All known signals, identified as being produced by fish or machinery from man-made activities, were ignored in the analyses while we searched for signals that could be attributed to the whales. Upon encountering new signals that had not been previously recorded in Ilha Grande Bay, we have separated these signals for further inspection and compared to minke whale calls available in the literature to compare frequencies and signal structure (Gedamke et al., 2001; Rankin and Barlow, 2005; Risch et al., 2019; Rossi-Santos et al., 2022; Filun and van Opzeeland, 2023). Although there was no exact match, there were signals that had significant similarities to calls recorded from dwarf minke whales in Australia, as we explain in the discussion of this manuscript. These were counted and classified into types named: Ba (for Balaenoptera acutorostrata) + a number (indicating the order in which they first appeared in the analysis).
The total calling rate was estimated by counting all sounds and dividing by the total time of each of the three recording sessions, providing us with an estimated number of calls per minute (calls/min). During visual inspection of the spectrograms, it was observed that signals were usually repeated within a short interval, so to estimate the repetition rate, we counted calls of the same type. We divided the total time of each recording session (repetitions/min), and then we measured the time interval between each repetition. Intervals were defined as the period between consecutive calls starts. These estimations were performed with all calls with a precise start time, even if they were not of high enough quality for acoustic parameter extraction. This procedure was applied separately in each recording session since these intervals could not be estimated during the break between sessions.
These signals had varying apparent source levels, their energy increasing and decreasing, as would have been expected from animals swimming around our hydrophone. Calculating precise source levels with a single hydrophone and multiple animals would not be possible, so we measured the signal-to-noise ratio through the SNR NIST Quick tool from Raven 1.6, and discarded sounds with less than 15 dB of SNR. Then, if the signals had a start and end that could be clearly identified, and if other calls did not overlap them, their acoustic parameters were extracted: duration (ms), minimum frequency (Hz), maximum frequency (Hz), delta frequency (Hz), center frequency (Hz) and peak frequency (Hz). Call types were regularly visually compared to signals available in the literature to see if there were possible matches to sounds previously reported by dwarf minke whales.
3 Results
A total of 222 acoustic signals in the recordings were attributed to the whales, occurring mostly in four call types named Ba1, Ba2, Ba3, and Ba4. The Ba1 call had the most distinct characteristics because, in the majority of occurrences, it had three different parts occupying different frequency bands: a low-frequency component (LFC) composed of pulsed moans where three to four bands were visible; and a high-frequency component (HFC) that had two to four pulses and two to three tones. Ba2 was the second most common type, composed of short burst pulses. Ba3 call type occurred three times and Ba4 occurred only once, each. Figure 2 shows the observed types, and in the Supplementary Material Supplementary Figures 2-4 show the different components of the Ba1 call in detail.
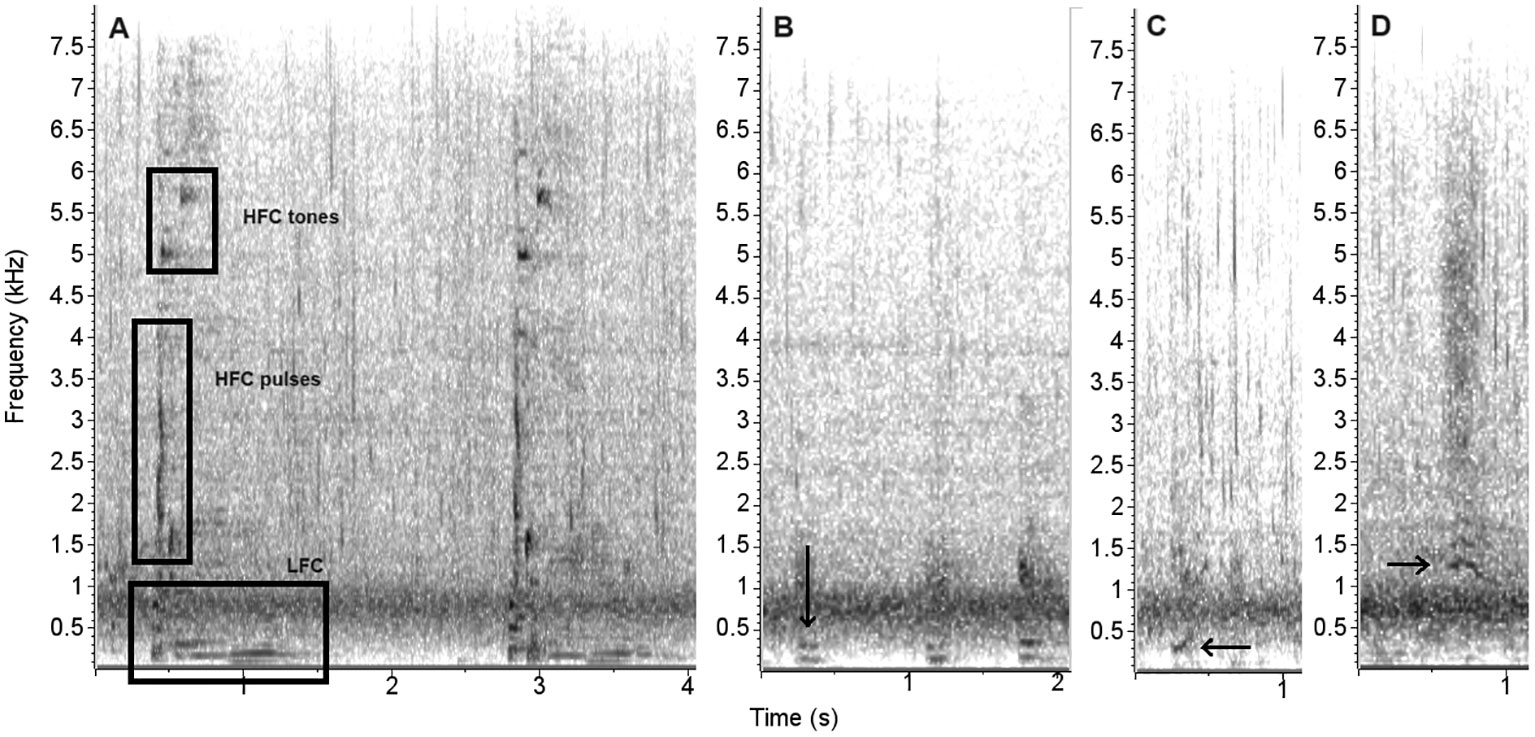
Figure 2 Dwarf minke whale call types and repetition sequences recorded in Ilha Grande Bay, southeastern Brazil: (A) two Ba1 signals, (B) a sequence of three Ba2 signals, (C) Ba3, and (D) Ba4. Squares indicated the different components of Ba1, and arrows point to the other signals locations. Spectrograms with 10s window scale and a 512-Hann window with 75% overlap.
During the three recording sessions, the calling rate varied from 4.4 to 6.7 calls/min, then to 11.1 calls/min respectively. As can be observed in Table 1, the repetition rate of Ba1 followed this pattern, increasing from the first session to the last while intervals decreased. The overlapping of Ba1 calls also occurred in the second and third sessions. In contrast, Ba2 repetitions decreased from the beginning to the end of the recordings. It is possible that some of these repetitions could not be counted due to noise masking and overlapping of signals.
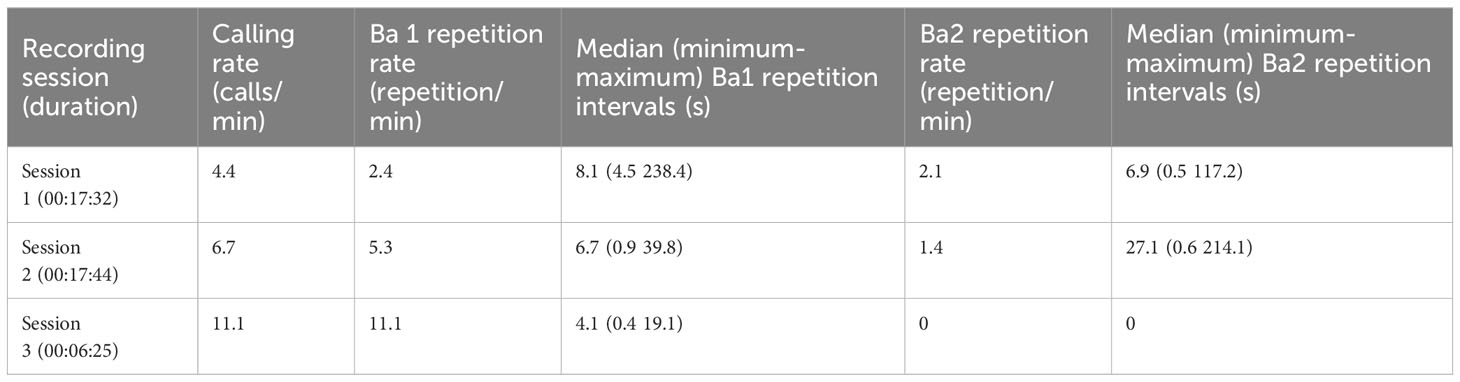
Table 1 Dwarf minke whale total calling rate and repetition rates for two call types (Ba1 and Ba2) recorded on the Brazilian coast.
59 calls fit our analysis criteria (Table 2). We analyzed LFC and HFC parts separately since Ba1 calls had multiple components with varied structures. There was only one occurrence in which Ba1 contained only the LFC part and none in which only the HFCs appeared alone. Still, many weak signals could not be analyzed, and it is possible that whales emitted more varied combinations.
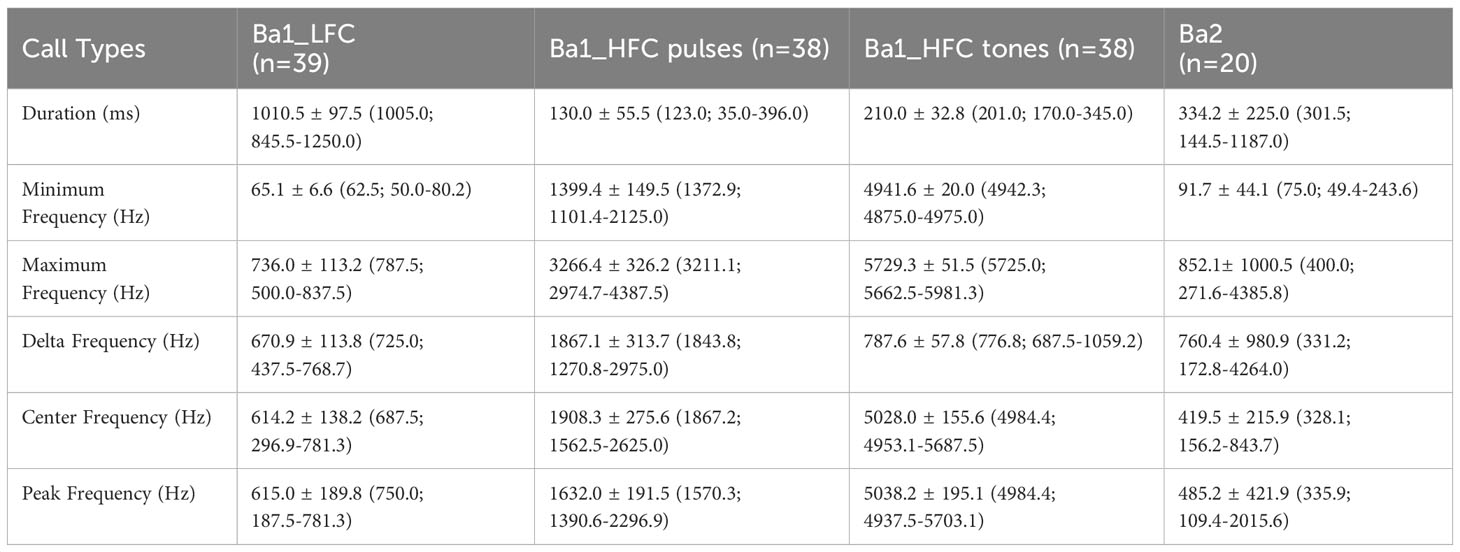
Table 2 Mean ± standard deviation (median; minimummaximum) values of dwarf minke whale calls acoustic parameters recorded on the Brazilian coast.
4 Discussion
Although the data reported here is from a brief encounter, the calls we have described contribute to our constantly expanding knowledge of the minke whale acoustic repertoire, emphasizing the dwarf form from the southern hemisphere. The direct visual observation concomitant to the recordings is not possible across all studies, and this opportunity allowed us to identify new calls that might have been otherwise discarded as artificial sounds. Minke whale sounds are known for their strange aural characteristics (Nieukirk et al., 2016), often like mechanical sounds. This is exemplified by the boing sound (Rankin and Barlow, 2005) produced by the regular form of B. acutorostrata in the North Pacific, and by the “Star Wars” sound (SW) produced by the dwarf form in the Great Barrier Reef area (Gedamke et al., 2001).
The complex structure of the Ba1 calls is rare and similar sounds in the literature have been reported only twice: the SW sound, described by Gedamke et al. (2001), and the Western Pacific Biotwang (WPB), described by Nieukirk et al. (2016). The WPB was recorded through autonomous recording and had no visual confirmation of the source, but it is possibly a minke whale sound (Nieukirk et al., 2016). Although these three sounds (SW, WPB, and Ba1) are not exactly matched, they occupy similar frequency bands and are a complex combination of pulses and tones. The SW and the Ba1 are especially alike at the higher frequency portion, in which the ‘A structure from SW and the HFC pulses have similar spectral characteristics. Various factors can influence intraspecific variation of acoustic signals. It can occur from population to population, so we may observe a biogeographic acoustic variation similar to the call differences of blue and pygmy blue whales (McDonald et al., 2006), and the apparent biogeographic variation observed in Antarctic minke whales in the Weddel Sea (Filun and van Opzeeland, 2023).
The function of many whale calls is largely associated with reproduction and feeding behavior (McDonald et al., 2006). Winter is the most common season of baleen whale reproduction (Pomeroy et al., 2018), and sounds attributed to minke whales typically peak during this season in the South Pacific (Buchan et al., 2020) and in the Antarctic (Filun et al., 2020). In this opportunistic study, we could not classify the behavioral state during which the calls were emitted. It is possible that this was a reproduction display, but many of the balaenopterids behavioral displays are still unknown (Pomeroy et al., 2018). The animals could be feeding, but no fish were visible in the water or the boats echosounder during the moments it was on; no birds were diving or following the whales.
Although we have a limited sample size and time frame, our findings add more range to the characteristics of known sound emissions of Balaenoptera acutorostrata on the South Atlantic Ocean, expanding the knowledge of this species. Those acoustic signals had similar bandwidth, but some variation in structure from those signals reported for dwarf minke whales in the Pacific Ocean. Despite regular monitoring, this is the first record of dwarf minke whales direct observation in Ilha Grande Bay. This is one of the less noise-impacted environments in the Rio de Janeiro state coastal area (Bittencourt et al., 2020). Still, the whales remained precisely where there is a higher density of anchored and trafficking ships, with shipping noise visible throughout and impacting all the recordings. They overlapped with some frequency bands of both Ba1 and Ba2 calls. It is beyond the scope of this study to evaluate if shipping noise interfered with their behavior and communication. This occurrence highlights the need to monitor and maintain the acoustic preservation of the bay.
Data availability statement
The acoustic data that support the findings of this study are available upon request from the corresponding author. The data are not publicly available due to university regulation restrictions. Requests to access the datasets should be directed to LB,bGlzLmJpdHRAZ21haWwuY29t.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
LB: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. RC: Data curation, Investigation, Methodology, Validation, Writing – review & editing. ES-N: Data curation, Investigation, Methodology, Validation, Writing – review & editing. TB: Data curation, Funding acquisition, Project administration, Resources, Writing – review & editing. JL-B: Data curation, Funding acquisition, Project administration, Resources, Validation, Writing – review & editing. AA: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Fundacao Carlos Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq) have supported researches developed by MAQUA/UERJ. LB has a scholarship from FAPERJ (PDR-10 E-26/202.266/2019), and AA has research grants from CNPq (PQ-1B 307458/2022-9), FAPERJ (CNE E-26/200.397/2023) and UERJ (Prociencia), TB has three research grants (FAPERJ-JCNE E-26/201.318/2022, CNPq-PQ-2 312142/2020-0 and UERJ-Prociencia), and JL-B has research grants from CNPq (PQ-1D 315.276/2021-5), and UERJ (Prociencia).
Acknowledgments
The authors thank the MAQUA/UERJ team for their support during field campaigns. Tamoios Ecological Station (ESEC Tamoios ICMBio) has collaborated in the conduction of this study. The Brazilian National Institute of Science and Technology- INCT Biodiversidade da Amazônia Azul (CNPq proc. 405999/2022-4) also contributed to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1338538/full#supplementary-material
References
Bittencourt L., Barbosa M., Bisi T. L., Lailson-Brito J. Jr., Azevedo A. F. (2020). Anthropogenic noise influences on marine soundscape variability across coastal areas. Mar. pollut. Bull. 160, 111648. doi: 10.1016/j.marpolbul.2020.111648
Buchan S. J., Balcazar-Cabrera N., Stafford K. M. (2020). Seasonal acoustic presence of blue, fin, and minke whales off the Juan Fernandez Archipelago, Chil 2016). Mar. Biodivers. 50. doi: 10.1007/s12526-020-01087-3
Cornell Lab of Ornithology, K. L. Y. C. for C. B (2023). Raven pro: Interactive Sound Analysis Software (Version 1.6.4). Available online at: https://ravensoundsoftware.com/.
Di Tullio J. C., Gandra T. B. R., Zerbini A. N., Secchi E. R. (2016). Diversity and distribution patterns of cetaceans in the subtropical southwestern atlantic outer continental shelf and slope. PloS One 11, e0155841. doi: 10.1371/journal.pone.0155841
Dominello T., Sirović A. (2016). Seasonality of Antarctic minke whale (Balaenoptera bonaerensis) calls off the western Antarctic Peninsula. Mar. Mammal Sci. 32 (3), 826–838. doi: 10.1111/mms.12302
Filun D., Thomisch K., Boebel O., Brey T., Sirović A., Spiesecke S., et al. (2020). Frozen verses: Antarctic minke whales (Balaenoptera bonaerensis) call predominantly during austral winter: Antarctic minke whales acoustic presence. R. Soc Open Sci. 7. doi: 10.1098/rsos.192112
Filun D., van Opzeeland I. (2023). Spatial and temporal variability of the acoustic repertoire of Antarctic minke whales (Balaenoptera bonaerensis) in the Weddell Sea. Sci. Rep. 13, 1–12. doi: 10.1038/s41598-023-38793-4
Gedamke J., Costa D. P., Dunstan A. (2001). Localization and visual verification of a complex minke whale vocalization. J. Acoust. Soc Am. 109, 3038. doi: 10.1121/1.1371763
Hassel L. B., Venturotti A., Magalhaes F. A., Cuenca S., Siciliano S., Marques F. F. C. (2003). Summer sightings of dwarf minke whales (Balaenoptera acutorostrata) off the eastern coast of Rio de Janeiro State, Brazil. Lat. Am. J. Aquat. Mamm. 2, 47–50. doi: 10.5597/lajam00030
Jefferson T., Webber M., Pitman R. (2011). Marine mammals of the World: A Comprehensive Guide to Their Identification: A Comprehensive Guide to Their Identification (London: Academic Press).
Lailson-Brito J., Azevedo A. F., Santos-Neto E. B., Bisi T. L. (2020). Botos cinza: e outros cetaceos das baias da Ilha Grande e de Sepetiba (São Paulo, SP - Brazil: DBA Editora).
McDonald M. A., Mesnick S. L., Hildebrand J. A. (2006). Biogeographic characterisation of blue whale song worldwide: using song to identify populations. J. Cetacean Res. Manage. 8, 55–65. doi: 10.47536/jcrm.v8i1.702
Mellinger D. K., Carson C. D., Clark C. W. (2000). Characteristics of minke whale (Balaenoptera acutorostrata) pulse trains recorded near Puerto Rico. Mar. Mammal Sci. 16, 739–756. doi: 10.1111/j.1748-7692.2000.tb00969.x
Milmann L., Siciliano S., Morais I., Tribulato A. S., MaChado R., Zerbini A. N., et al. (2020). A review of Balaenoptera strandings along the east coast of South America. Reg. Stud. Mar. Sci. 37. doi: 10.1016/j.rsma.2020.101343
Nieukirk S. L., Fregosi S., Mellinger D. K., Klinck H. (2016). A complex baleen whale call recorded in the Mariana Trench Marine National Monument. J. Acoust. Soc Am. 140, EL274–EL279. doi: 10.1121/1.4962377
Perrin W. F., Mallette S. D., Brownell R. L. (2018). “Minke whales,” in Encyclopedia of Marine Mammals, (United States of America: Academic Press) 608–613. doi: 10.1016/b978-0-12-804327-1.00175-8
Pomeroy P. P., Twiss S. D., Robinson K. J. (2018). “Reproductive Behavior,” in Encyclopedia of Marine Mammals, (United States of America: Academic Press) 797–807. doi: 10.1016/b978-0-12-804327-1.00213-2
Rankin S., Barlow J. (2005). Source of the North Pacific “boing” sound attributed to minke whales. J. Acoust. Soc Am. 1183346–3351. doi: 10.1121/1.2046747
Risch D., Norris T., Curnock M., Friedlaender A. (2019). Common and antarctic minke whales: conservation status and future research directions. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00247
Rossi-Santos M. R., Filun D., Soares-Filho W., Paro A. D., Wedekin L. L. (2022). “Playing the beat”: Occurrence of Bio-duck calls in Santos Basin (Brazil) reveals a complex acoustic behaviour for the Antarctic minke whale (Balaenoptera bonaerensis). PloS One 17, 1–13. doi: 10.1371/journal.pone.0255868
Keywords: bioacoustics, Ilha Grande Bay, balaenopterids, soundscape, animal communication
Citation: Bittencourt L, Carvalho R, Santos-Neto EB, Bisi TL, Lailson-Brito Jr. J and Azevedo AF (2024) Dwarf minke whales (Balaenoptera acutorostrata) acoustic signals from the South Atlantic Ocean. Front. Mar. Sci. 11:1338538. doi: 10.3389/fmars.2024.1338538
Received: 14 November 2023; Accepted: 01 February 2024;
Published: 20 February 2024.
Edited by:
Todd Atwood, U.S. Geological Survey, United StatesReviewed by:
Ilse Catharina Van Opzeeland, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), GermanyDiego Filun, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), Germany
Copyright © 2024 Bittencourt, Carvalho, Santos-Neto, Bisi, Lailson-Brito and Azevedo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lis Bittencourt, bGlzLmJpdHRAZ21haWwuY29t