 Ramón Bonfil1,2,3*
Ramón Bonfil1,2,3* Píndaro Díaz-Jaimes4
Píndaro Díaz-Jaimes4 Paola Palacios-Barreto3,5Oscar Uriel Mendoza Vargas3Melina Ricaño-Soriano3
Paola Palacios-Barreto3,5Oscar Uriel Mendoza Vargas3Melina Ricaño-Soriano3- 1Department of Systematics and Aquatic Ecology, El Colegio de la Frontera Sur (ECOSUR) Unidad Chetumal, Chetumal, Mexico
- 2Adjunct Direction for Humanistic and Scientific Research, Consejo Nacional de Ciencia y Tecnología, Mexico City, Mexico
- 3Océanos Vivientes AC, Mexico City, Mexico
- 4Unidad Académica de Ecología y Biodiversidad Acuática, Laboratorio de Genética de Organismos Acuáticos. Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 5Fundación colombiana para la investigación y conservación de Tiburones y Rayas, SQUALUS, Cali, Colombia
Sawfishes are considered one of the most threatened marine fish taxa globally, with major contractions in their geographic range documented over the last 50 years across all oceans. Two sawfish species used to be found in Mexico, but a historical lack of research and first-hand baseline information recently raised concerns about the fate of both species there. Recent Local Ecological Knowledge (LEK) and environmental DNA (eDNA) research has shown that: a) Pristis pectinata and P. pristis were formerly abundant and widely distributed in the Mexican Atlantic coast, and the latter also in the Mexican Pacific coast, and b) sawfishes (at least P. pectinata) are still found in some coastal refugia in central Veracruz state and Campeche state on the Atlantic coast. In the present research we developed improved primers that allow for better species ID resolution for Pristis pectinata and P. pristis through successful sequencing of eDNA samples by amplifying a 270 bp fragment of the CO1 gene. Our results based on the analysis of 305 eDNA samples showed an improved positive species identification rate through sequencing, demonstrate that P. pectinata occurs in other coastal refugia along the Yucatán Peninsula in the Mexican Caribbean, and suggest this species might be extirpated from southern Veracruz, while no P. pristis eDNA has been found along the Atlantic coast.
1 Introduction
Assessing the presence/absence of endangered, rare, and invasive species has long posed a major challenge to fisheries and conservation scientists due to the often rarity and elusiveness of such species in aquatic habitats. Gillnetting, electrofishing, trapping, trawling, and angling are techniques usually expensive in time and effort, and which have relatively low power to avoid false negatives –that is, no record of the species when it is present in the environment– (Goldberg et al., 2015; Thomsen and Willerslev, 2015; Le Port et al., 2018). The breakthrough and development of environmental DNA (eDNA) protocols for the detection of rare aquatic species (Ficetola et al., 2008) represents an unprecedented methodological innovation that has solved many, but not all of the obstacles for rare species’ detection (Rees et al., 2014; Cristescu and Hebert, 2018) and one that has seen application to a large number of taxa in recent years (Foote et al., 2012; Piaggio et al., 2014; Boussarie et al., 2018). The eDNA revolution, has also seen extended applications in biodiversity assessment and monitoring (Thomsen et al., 2012a; Thomsen et al., 2012b; Port et al., 2016; Bakker et al., 2017; Valdivia-Carrillo et al., 2021).
Sawfishes (Family Pristidae) are shark-like rays that occurred in all tropical coastal seas but are currently considered the marine taxon most threatened with extinction (Harrison and Dulvy, 2014). Overfishing, habitat destruction and degradation, and the vulnerability of sawfishes to fisheries due to their preference for shallow and often inshore coastal habitats, have all contributed to the severe range reduction and abundance decrease of their populations globally (Dulvy et al., 2016; Leeney and Downing, 2016; Bonfil et al., 2018).
Recently, efforts to assess if sawfishes still exist in Mexico yielded positive results when eDNA evidence highlighted their presence in three coastal areas in the southern Gulf of Mexico (Bonfil et al., 2021). That study, based on the primers developed specifically for the genus Pristis in Australia (Simpfendorfer et al., 2016) found sawfish DNA in 12 out of 158 sampled stations. However, only 2 of the 12 sawfish DNA samples from Bonfil et al. (2021) could be successfully sequenced, confirming that the smalltooth sawfish Pristis pectinata is still hiding in some coastal areas of Mexico (Bonfil et al., 2021). For the remaining 10 samples, no clear identity was found to distinguish between P. pristis and P. pectinata. This may have been due to DNA degradation or to the relatively short strain of DNA (145 bp) amplified and sequenced using the primers developed by Simpfendorfer et al. (2016). Thus, because of the known low number of polymorphic sites in the DNA fragment analyzed, longer fragments containing specific sites for each species may need to be sequenced in order to allow for easier species identification through sequencing (Naylor et al., 2012; Tavares et al., 2013).
In the present study, we a) developed a new set of primers to amplify a longer fragment of DNA (270 bp) containing more polymorphic sites that allow to more easily distinguish between Pristis species, and b) designed an eDNA sampling protocol to determine the presence of two endangered sawfishes in coastal areas of the Gulf of Mexico and Caribbean. We also present results of surveys in Quintana Roo and Veracruz showing through eDNA evidence that smalltooth sawfishes are still present in at least two coastal areas of the Mexican Caribbean, where fishing activity has been comparatively low and where human encroachment is less severe and more recent than in the Gulf of Mexico.
2 Materials and methods
2.1 Primer development
To develop our eDNA assay, we modified the primer pair from Simpfendorfer et al. (2016) targeting the cytochrome oxidase I (COI) gene at positions 172-191 and 453-472 for forward and reverse primers respectively. Primers were designed targeting polymorphic sites for species differentiation. Our primers amplified a 270 bp fragment of the CO1 gene and were Ppec-F1 (5’-GTAACTGCCCATGCCTTTGT-3’), and Ppec-R1 (5’-TAGAGGAGATGCCGGCTAAA-3’). The specificity of these primers was tested using tissue samples from a juvenile P. pectinata collected in 2016 and held since then at the Veracruz Aquarium as well as P. pristis dried rostra tissue samples from the first author’s private collection.
2.2 DNA extraction
Prior to DNA extraction, each water sample in the 50 mL collection tubes was precipitated by centrifugation for 35 min at 6°C and 3820 g (maximum speed of the equipment) according to Ficetola et al. (2008) and Turner et al. (2015). DNA extraction was done using the entire precipitate and Qiagen DNeasy Blood & Tissue kit (Qiagen®) (spin column protocol) following manufacturer’s protocol except for the lysis of precipitated material, which was carried out in the 50 ml tubes used for the precipitation overnight and then transferred to a 1.5 ml tube for the rest of the protocol. Additionally, for the final step, we performed an elution with 50 μl AE buffer (supplied with the kit). The precipitation protocol, excepting for the centrifugation step of the 50 ml tubes, was carried out in a DNA clean lab. During the centrifugation step, tubes were always sealed, and the outsides were decontaminated with 10% bleach. Extraction blanks were included to test for contamination during this process. For samples from the P. pectinata aquarium specimen and from the tissue collected from the dried rostrum of P. pristis, we used a traditional phenol:chloroform:isoamyl alcohol (25:24:1 v/v) protocol (Díaz-Jaimes et al., 2018).
2.3 PCR amplification
PCR reactions were performed in a 25 μl final volume containing 1 μl of template DNA, sterile MilliQ water, Taq polymerase buffer (10x), dNTPs mixture (2 mM each), each primer (10 pm/μl), and DreamTaq DNA polymerase (5 U μl). PCR reactions were run below the following PCR conditions: 5 min initial denaturation at 95°C, followed by 35 cycles of 30 sec at 94°C, annealing for 1.5 min at 58°C, extension for 30 sec at 73°C; final extension for 10 min at 72°C, and an infinite hold at 4°C. The PCR products were verified in a 2% agarose gel through electrophoresis. Blanks containing only deionized water instead of template DNA were used in all amplifications. The resulting sequences were trimmed using Geneious Pro v6.0.5 (Biomatters Ltd.) and compared against the National Center for Biotechnology Information (NCBI) nonredundant nucleotide database. The minimal criterion of similarity values above 98% was adopted for species identification following Hebert et al. (2003).
2.4 Field surveys
Based on our previous Local Ecological Knowledge work (Bonfil et al., 2018) and taking into consideration locations we had previously surveyed (Bonfil et al., 2021), in this study we sampled three large areas in the southern Gulf of Mexico and Caribbean to investigate if sawfishes still exist there (Figure 1). During Feb 25-Mar 16, 2018, we visited the northeast tip of the Yucatan Peninsula (NEYP from now on) in the state of Quintana Roo from Yalahau Lagoon to Puerto Juárez, including all coastal lagoons and the waters off Contoy Island, all known as former sawfish habitat (Bonfil et al., 2018). During this expedition we also surveyed Bahía Espíritu Santo (BES from now on) and surrounding waters, all part of the Sian Ka’an Biosphere Reserve; a live sawfish was reported in BES to one of us (RB) by fishers in 2016. During Sep 18-Oct 8 2018 we surveyed the southern coast of Veracruz State (SVS from now on), from Veracruz to Coatzacoalcos, including del Ostión, Sontecomapan, and Alvarado lagoons, the Coatzacoalcos and Papaloapan river systems, and the coral reefs off Antón Lizardo and Veracruz.
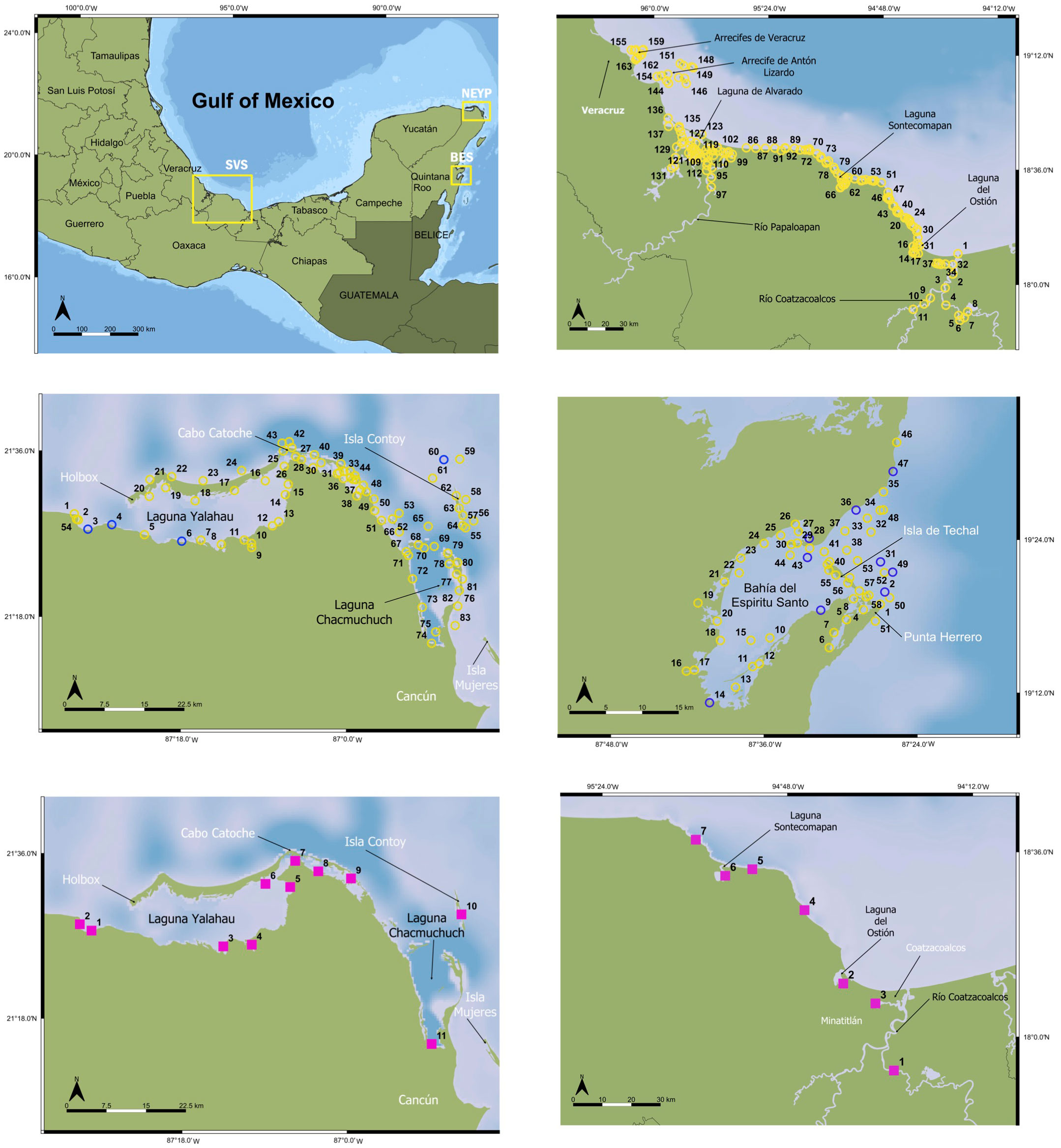
Figure 1 Sampling sites and sample stations for sawfish eDNA (yellow circles) and gillnet sets (fuchsia squares) in the southern Gulf of Mexico and Caribbean. eDNA stations in: Southern Veracruz State coast (SVS; upper right panel), North East Yucatan Peninsula [NEYP; middle left panel), and Bahía Espíritu Santo (BES; middle right panel)]; gillnet sets in: NEYP (lower left panel), and SVS (lower right panel); blue circles denote stations that resulted positive for sawfish DNA.
We centered our study on eDNA detection, supported whenever possible by gillnetting and aerial surveys with drones (known also as unamend aerial vehicles or UAVs). Gillnetting was rather limited due to time and human power constraints, as well as due to fishing prohibitions in some of the Marine Protected Areas surveyed (chiefly, BES).
2.4.1 eDNA sampling
Sampling for eDNA followed Ficetola et al. (2008) as detailed in our previous work (Bonfil et al., 2021). Each water sample was taken in triplicate 75 m apart, and consisted of taking 15 ml of surface water with a 15 ml falcon tube and poured into a 50 ml falcon tube containing 1.5 ml 3M, 5.2 pH sodium acetate buffer solution, which was then filled with 96% ethanol (Ficetola et al., 2008; Thomsen et al., 2012b; Sigsgaard et al., 2015; Figure 2). New 15 ml falcon tubes and sterile examination gloves were used to take each sample and all triplicate samples were kept in pre-labeled ziplock bags inside a sterilized cooler containing a negative control. The negative control consisted of 15 ml of distilled water, 1.5 ml 3M, 5.2 pH sodium acetate buffer solution, and 96% ethanol.

Figure 2 Water samples for eDNA were taken with a 15 ml falcon tube (A), then were poured into a 50 ml falcon tube containing 1.5 ml 3M, 5.2 pH sodium acetate buffer solution (B), which was then filled with 96% ethanol (C). Aerial surveys with two DJI Phantom drones were made in the clear shallow waters of NEYP and BES (D).
2.4.2 Gillnet sampling
Gillnets rented to local fishers were set perpendicular to the shoreline inside coastal lagoons or around keys (Figure 3); soaking times were between 35 and 481 min. Either polyamide or monofilament gillnets were used, and these ranged from 97 to 120 m in length, and from 76 to 89 mm in mesh sizes.

Figure 3 Gillnets were set parallel to the shoreline or closing the mouths of rivers and channels, and left to fish for 35 to 481 min.
2.4.3 Aerial surveys
Where water clarity allowed, a DJI Phantom 3 Advanced or a DJI Phantom 4 UAVs were used to carry out aerial transects recording video during the entire track. Transects followed either the shoreline of the areas surveyed, or the path of inner channels along coastal lagoons. The UAV was flown at a maximum altitude of 20 m during surveys. Whenever elasmobranch fishes were detected from above, the drone was flown at a lower altitude (down to a minimum of 2 m) and individuals photographed and filmed for posterior identification. After this, the transects continued as described above. Transect duration depended heavily on the UAV’s battery life and safety measures, but lasted mainly 15-22 min.
We took eDNA samples at 83 stations in NEYP, at 58 stations in BES, and at 168 stations in SVS (Figure 1) for a total of 305 stations. Gillnetting was limited to only 11 sets in NEYP, and 7 in SVS (Figure 1); gillnetting is not allowed in the Sian Ka’an Biosphere Reserve where Bahía Espíritu Santo is located. We carried out 8 aerial survey transects in NEYP and 10 transects in BES (Figure 4). No aerial surveys were carried out in Veracruz due to the high turbidity of the waters in the region.
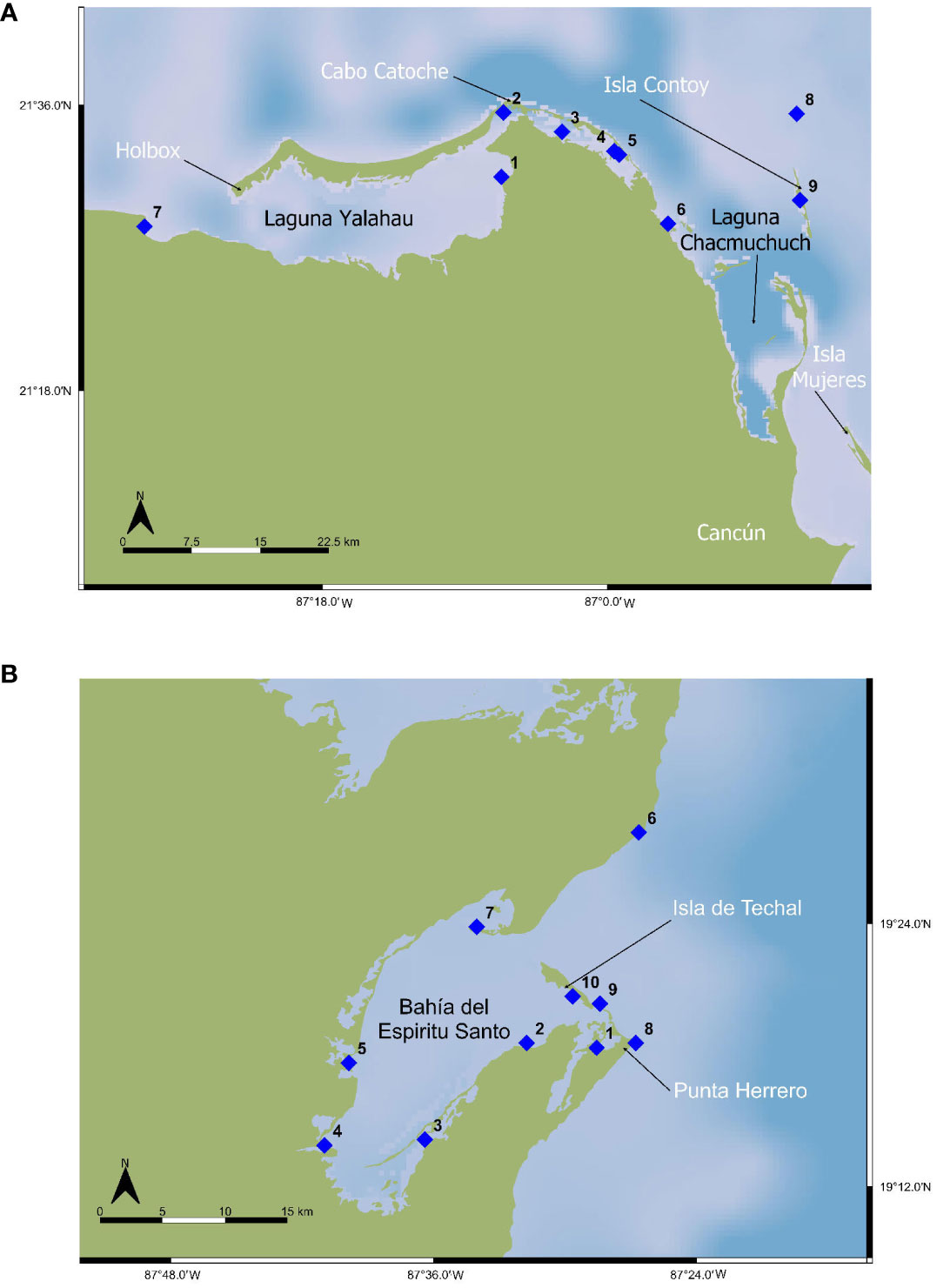
Figure 4 Location of aerial transects with UAVs in the study sites. (A) North East Yucatan Peninsula (NEYP), (B) Bahía Espíritu Santo (BES) inside Sian Ka’an Biosphere Reserve.
3 Results
3.1 Primer and assay tests
Our custom primers amplified a 270 bp fragment of the COI target sequence in Pristis, confirming the primer specificity of the assay for conventional PCR. During the initial trial of our custom primers, we successfully identified DNA from tissue samples of a live specimen of Pristis pectinata, kept at the Veracruz Aquarium since 2016, as well as from dried tissue obtained from P. pristis rostra (Bonfil et al., 2017); both samples amplified as expected. Furthermore, when testing water samples collected from the exhibition tank where the specimen of P. pectinata is kept at the Veracruz Aquarium, we confirmed through sequencing the presence of DNA from P. pectinata in the water samples.
3.2 eDNA surveys
DNA from Pristis pectinata was detected and confirmed in water samples collected from two of the three areas we surveyed in the Southern Gulf of Mexico and the Mexican Caribbean. Out of a total of 305 sampled stations, we obtained 22 positives (i.e. samples that amplified during PCR with our primers), 17 of which were successfully sequenced (see Supplementary Table 1). Of these, 13 were positively identified through BLAST as smalltooth sawfish Pristis pectinata based on a similarity criterion above 98% (Hebert et al., 2003; Table 1). None of the negative controls resulted in a positive reaction and all extraction blanks were negative. We found 4 samples positive for smalltooth sawfish DNA in NEYP, 9 positives in BES, and no positives in the entire coast of SVS (Figure 1). Of the positive samples from NEYP, 3 were from the southern coast of Yalahau Lagoon (2 close to the mouth of the lagoon and 1 from about the middle coast) and 1 was from open waters north of Contoy Island (Figure 1). The positive samples from BES were 1 from deep inside the bay in the southernmost coast, 1 from the southern coast just inside the mouth of the bay, 1 from the main channel between Techal Island and the northern part of the bay, and 1 from a shallow protected small lagoon at the northern entrance of the bay (Figure 1). The remaining 5 positives were all from within the reef lagoon at the mouth of the bay: two in the northern coast, and 3 from the central-southern mouth of the bay just behind the barrier reef. Summarizing, we obtained a total of 13 eDNA samples positively-sequenced as Pristis pectinata, 4 from the NEYP and 9 from BES.
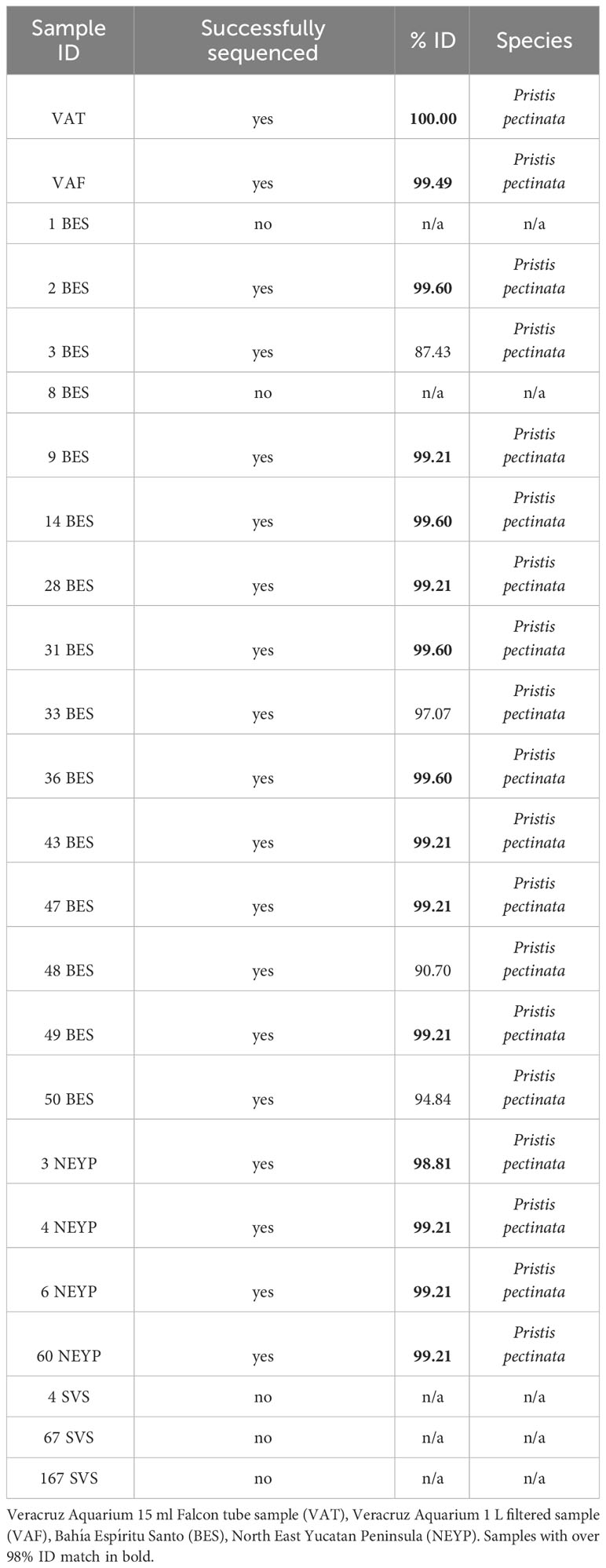
Table 1 Results of BLAST analysis of nucleotide sequences from CO1 gene for positive eDNA samples.
3.3 Gillnetting and aerial surveys
No sawfishes were caught in our gillnet sets or located during our aerial surveys. We caught 3 species of batoids during gillnet surveys in NEYP: one specimen each of Styracura schmardae, Hypanus americanus, and Urobatis jamaicensis (Table 2). In addition, we observed 16 individuals of 3 batoid and 1 shark species during our aerial surveys: Styracura schmardae (n=5), and Hypanus americanus (n=3) in NEYP; Styracura schmardae (n=5), Hypanus americanus (n=1), Aetobatus narinari (n=1), and Carcharhinus leucas (n=1), in BES (Table 3). All batoids caught in the gillnets were measured, sexed, photographed and released alive bearing a plastic spaghetti tag announcing a reward for reporting to us the date and site of recapture.
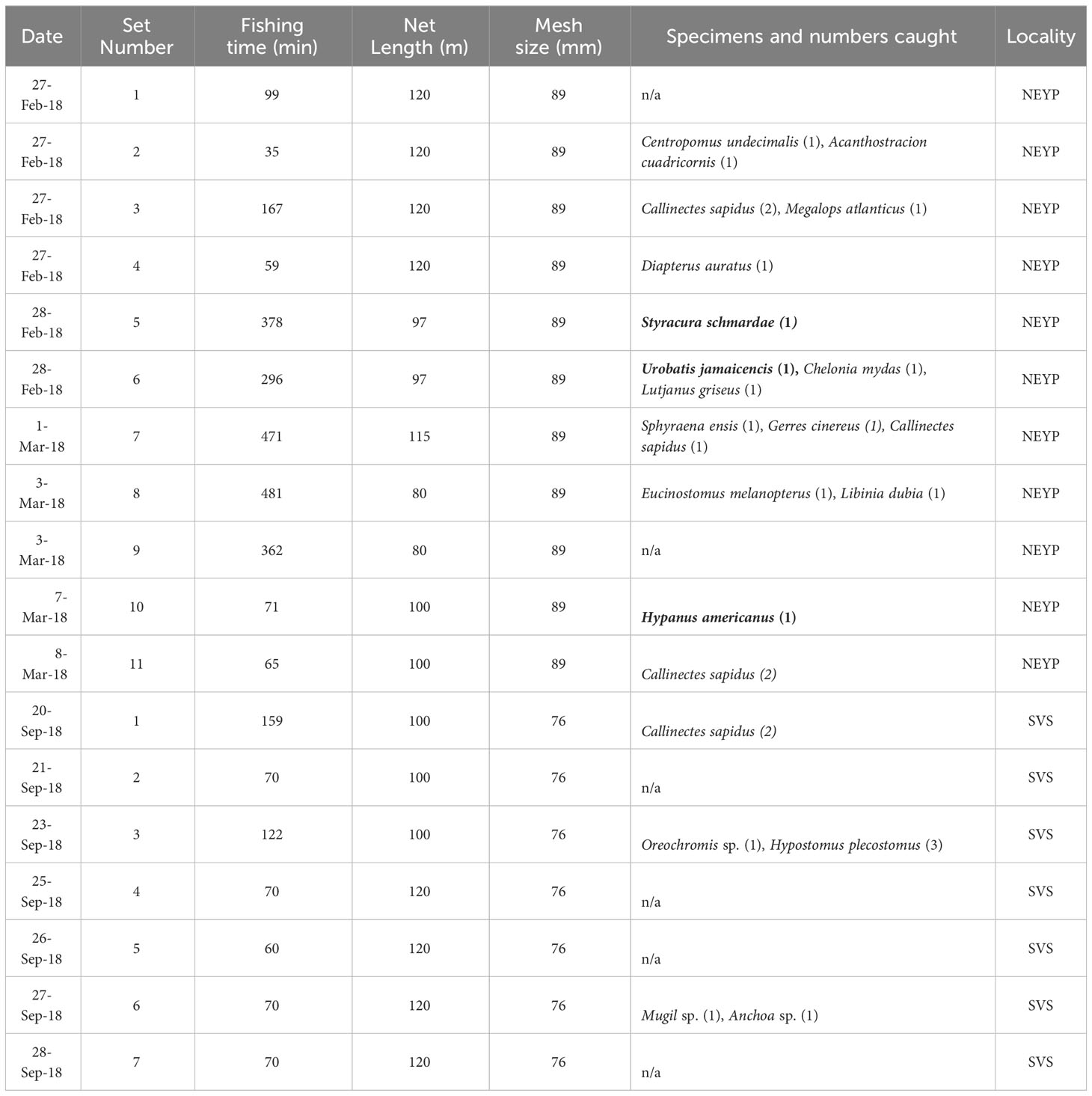
Table 2 Details of gillnets deployed and specimens caught during sawfish surveys (elasmobranchs in bold, n/a = not applicable).
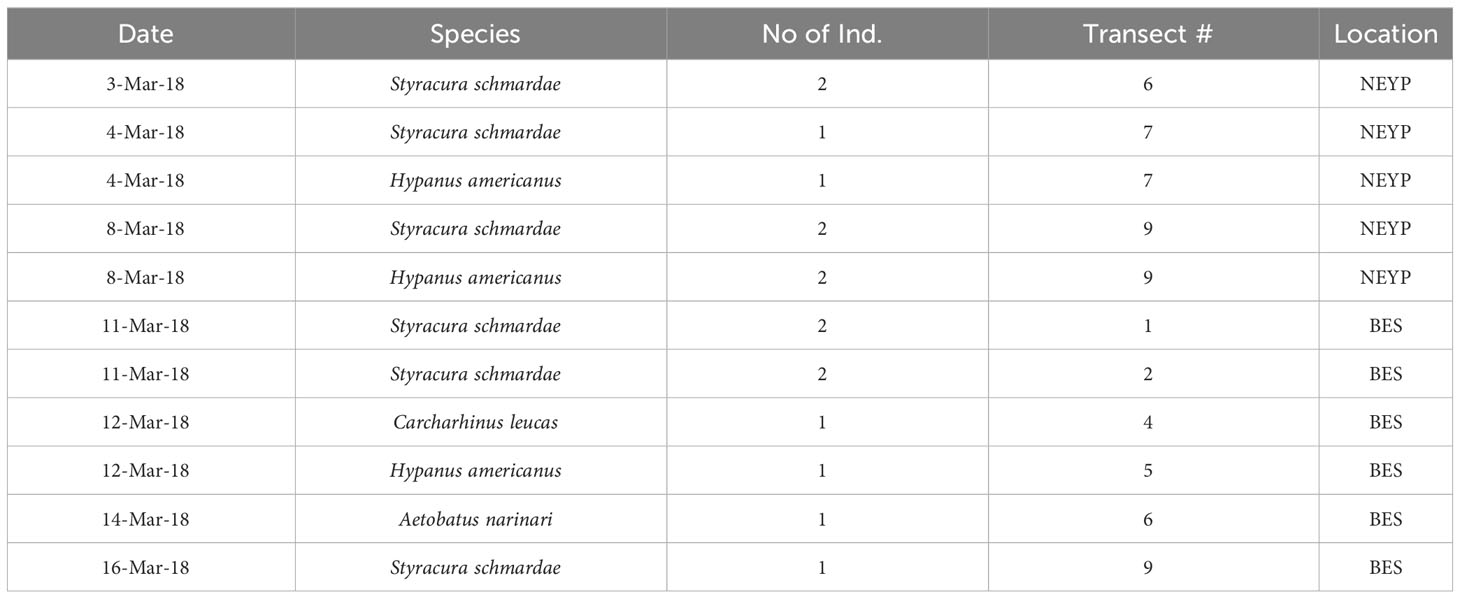
Table 3 Elasmobranchs recorded during drone aerial transects (all 2018, Quintana Roo).
4 Discussion
The primers developed in this study proved to be a useful tool yielding improved results for sawfish DNA detection and evidencing for the first time in over two decades the presence of sawfishes in several locations in the Mexican Caribbean. Notably, these primers gave better species definition during sequencing of positive samples compared to a previous study (Bonfil et al., 2021) where we used the primers for Pristis spp. developed by Simpfendorfer et al. (2016). Specifically, in the present study we failed to obtain sequences for only 23% (5 out of 22 positives samples, i.e., those that amplified during PCR), while our previous study had 83% of samples that failed to be sequenced (10 out of 12 positive samples; Bonfil et al., 2021). The use of short sequences is a major limiting issue for eDNA assays because it can complicate species identification through sequencing, especially for elasmobranchs, which are characterized by a low number of variable sites in the mtDNA genome (Martin et al., 1992). A lower number of variable sites means a lower possibility to find sufficient variation for species discrimination. The primers developed in this study allowed the amplification of a 270 bp fragment containing 32 variable sites to differentiate between P. pristis and P. pectinata, both species formerly distributed in the study area. Four out of the 17 sequenced samples, had identity values below the 98% and we observed no records for identity of P. pristis among our samples. The closest records were <90% for P. zijsron, a species distributed in the Indo-Pacific Region. Considering the average number of differences in COI between both Pristis species known from Mexico (16%), we should expect at least identities below 90% matching to P. pectinata or P. pristis but the Blast did not retrieve any comparison lower than 90% between any of these species. The percentage of identity between the four haplotype sequences we found (reported in Genbank with accession numbers OR362728-OR362731) and both, the tissue sample from the smalltooth sawfish specimen in the Aquarium (100%), and the water samples collected in the surveyed areas (>99%), confirms that the species we detected is Pristis pectinata whereas no evidence for the presence of P. pristis was recovered.
Our results demonstrate that at least the smalltooth sawfish still exists in some refugia in the Mexican Caribbean and suggest that likely, their abundance is very low. We obtained only 4 positives out of 83 samples in NEYP, 9/58 in BES and none in the entire set of 164 samples from SEVS. However, it must be noted that 3 of the positives from NEYP are from relatively close-together sampling stations inside Yalahau Lagoon (stations 3, 4 and 6). Yalahau lagoon used to be a center of abundance and probably a nursery area for sawfishes in the mid 20th century, but no specimens have been reported there in the last few decades (Bonfil et al., 2018; Rubio-Cisneros et al., 2019). Also, the waters off Cabo Catoche and Contoy Island are known as former areas of abundance for sawfishes (Bonfil et al., 2018, Bonfil unpublished data), thus the positive sample in open waters north of Contoy Island is not surprising. In fact, our Local Ecological Knowledge data indicates that the last largetooth sawfish Pristis pristis caught in NEYP back in 1997, was caught off northern Contoy Island (Bonfil et al., 2017).
Our data also indicate that smalltooth sawfishes are either absent or extremely scarce in the waters off SEVS. However, the possibility of having false negatives in our samples from that region always exists since false negatives are a difficult and nearly unavoidable issue that will always remain in eDNA studies (Le Port et al., 2018). This possibility has to be considered in the light of the capture of a live specimen of this species in the northern part of the State of Veracruz in 2016 (Bonfil et al., 2018).
Data availability statement
The datasets presented in this study can be found in online repositories. The haplotype sequences presented in this study can be found in Genbank with accession numbers OR362728, OR362729, OR362730, and OR362731. They can also be seen accessed using the following link https://www.ncbi.nlm.nih.gov/nuccore/?term=OR362728:OR362731[accn].
Ethics statement
The animal study was approved by Océanos Vivientes AC ethics committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. PD: Conceptualization, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. PP: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. OM: Investigation, Writing – review & editing. MS: Investigation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Principal funding was provided by the Save Our Seas Foundation through Keystone Grant SOSF-375. Additional funding was provided by Ripley’s Myrtle Beach, Gatlinburg and Smokies Aquariums, the International Sawfish Day Initiative, as well as the Marine Conservation Action Fund of the New England Aquarium all these, without any specific grant number.
Acknowledgments
Invaluable volunteer fieldwork assistance was provided by Miguel Tello, Eréndira Reza Guzmán, and Rebecca Bateman-John. Rubén Valerio Parra, Francisco Cobá Ríos, and Serviliano Carrillo Moreno, kindly rented us boats and provided expertise during fieldwork in NEYP and BES, while a large number of fishers provided similar support in SVS. Violeta Monserrath Andrade González and Paola Díaz Chávez prepared the maps for publication. The authors wish to thank the financial support provided to RB for this project by: the Save Our Seas Foundation; the Ripley’s Myrtle Beach, Gatlinburg and Smokies Aquariums; the Marine Conservation Action Fund of the New England Aquarium; and an International Sawfish Day Conservation Grant. SEMARNAT and CONAPESCA granted the corresponding research permits and managers of the Sian Ka’an Biosphere Reserve and the Área de Protección de Flora y Fauna Yum Balam facilitated entrance to work inside those natural protected areas.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1290661/full#supplementary-material
References
Bakker J., Wangensteen O. S., Chapman D. D., Boussarie G., Buddo D., Guttridge T. L., et al. (2017). Environmental DNA reveals tropical shark diversity in contrasting levels of anthropogenic impact. Sci. Rep. 7, 1–11. doi: 10.1038/s41598-017-17150-2
Bonfil R., Mendoza-Vargas O. U., Ricaño-Soriano M., Palacios-Barreto P. Y., Bolaño-Martínez N. (2017). Former widespread abundance and recent downfall of sawfishes in Mexico as evidenced by historical photographic and trophy records. Fisheries 42, 256–259. doi: 10.1080/03632415.2017.1276339
Bonfil R., Palacios-Barreto P. Y., Mendoza-Vargas O. U., Ricaño-Soriano M., Díaz-Jaimes P. (2021). Detection of critically endangered marine species with dwindling populations in the wild using eDNA gives hope for sawfishes. Mar. Biol. 168, 1–2. doi: 10.1007/s00227-021-03862-7
Bonfil R., Ricaño-Soriano M., Mendoza-Vargas O. U., Méndez-Loeza I., Pérez-Jiménez J. C., Bolaño-Martínez N., et al. (2018). Tapping into local ecological knowledge to assess the former importance and current status of sawfishes in Mexico. Endang. Species. Res. 36, 213–228. doi: 10.3354/esr00899
Boussarie G., Bakker J., Wangensteen O. S., Mariani S., Bonnin L., Juhel J. B., et al. (2018). Environmental DNA illuminates the dark diversity of sharks. Sci. Advan 4, eaap9661. doi: 10.1126/sciadv.aap9661
Cristescu M. E., Hebert P. D. (2018). Uses and misuses of environmental DNA in biodiversity science and conservation. Annu. Rev. Ecol. Evol. Syst. 49, 209–230. doi: 10.1146/annurev-ecolsys-110617-062306
Díaz-Jaimes P., Bonfil R., Palacios-Barreto P., Bolaño-Martínez N., Bayona-Vásquez N. J. (2018). Mitochondrial genome of the critically endangered smalltooth sawfish Pristis pectinata from Veracruz, Mexico. Conserv. Genet. Res. 10, 663–666. doi: 10.1007/s12686-017-0896-9
Dulvy N. K., Davidson L. N. K., Kyne P. M., Simpfendorfer C. A., Harrison R. L., Carlson J. K., et al. (2016). Ghosts of the coast: global extinction risk and conservation of sawfishes. Aquat. Conserv. 26, 134–153. doi: 10.1002/aqc.2525
Ficetola G. F., Miaud C., Pompanon F., Taberlet P. (2008). Species detection using environmental DNA from water samples. Biol. Lett. 4, 423–425. doi: 10.1098/rsbl.2008.0118
Foote A. D., Thomsen P. F., Sveegaard S., Wahlberg M., Kielgast J., Kyhn L. A., et al. (2012). Investigating the potential use of environmental DNA (eDNA) for genetic monitoring of marine mammals. PloS One 7, e0041781. doi: 10.1371/journal.pone.0041781
Goldberg C. S., Strickler K. M., Pilliod D. S. (2015). Moving environmental DNA methods from concept to practice for monitoring aquatic macroorganisms. Biol. Conserv. 183, 1–3. doi: 10.1016/j.biocon.2014.11.040
Hebert P. D., Cywinska A., Ball S. L., deWaard J. R. (2003). Biological identifications through DNA barcodes. P. Roy. Soc. Lond. B. 272, 313–321. doi: 10.1098/rspb.2002.2218
Harrison L. R., Dulvy N. L. (Eds.) (2014). Sawfish: A global strategy for conservation (Vancouver: IUCN Species Survival Commission’s Shark Specialist Group).
Leeney R. H., Downing N. (2016). Sawfishes in The Gambia and Senegal–shifting baselines over 40 years. Aquat. Conserv. 26, 265–278. doi: 10.1002/aqc.2545
Le Port A., Bakker J., Cooper M. K., Huerlimann R., Mariani S. (2018). “Environmental DNA (eDNA): a valuable tool for ecological inference and management of sharks and their relatives,” in Shark Research: emerging technologies and applications for the field and laboratory. Eds. Carrier J. C., Heithaus M. R., Simpfendorfer C. A. (Boca Raton, FL: CRC Press), 255–284.
Martin A. P., Naylor G. J. P., Palumbi S. R. (1992). Rates of mitochondrial DNA evolution in sharks are slow compared with mammals. Nature 357, 153–155. doi: 10.1038/357153a0
Naylor G. J. P., Caira J. N., Jensen K., Rosana K. A. M., White W. T., Last P. R. (2012). A DNA sequence-based approach to the identification of shark and ray species and its implications for global elasmobranch diversity and parasitology. AMNH Bull. 367, 263. p. doi: 10.1206/754.1
Piaggio A. J., Engeman R. M., Hopken M. W., Humphrey J. S., Keacher K. L., Bruce W. E., et al. (2014). Detecting an elusive invasive species: a diagnostic PCR to detect Burmese python in Florida waters and an assessment of persistence of environmental DNA. Mol. Ecol. Resour. 14, 374–380. doi: 10.1111/1755-0998.12180
Port J. A., O'Donnell J. L., Romero-Maraccini O. C., Leary P. R., Litvin S. Y., Nickols K. J., et al. (2016). Assessing vertebrate biodiversity in a kelp forest ecosystem using environmental DNA. Mol. Ecol. 25, 527–541. doi: 10.1111/mec.13481
Rees H. C., Maddison B. C., Middleditch D. J., Patmore J. R., Gough K. C. (2014). The detection of aquatic animal species using environmental DNA–a review of eDNA as a survey tool in ecology. J. Appl. Ecol. 51, 1450–1459. doi: 10.1111/1365-2664.12306
Rubio-Cisneros N. T., Moreno-Báez M., Glover J., Rissolo D., Sáenz-Arroyo A., Götz C., et al. (2019). Poor fisheries data, many fishers, and increasing tourism development: Interdisciplinary views on past and current small-scale fisheries exploitation on Holbox Island. Mar. Policy 100, 820. doi: 10.1016/j.marpol.2018.10.003
Sigsgaard E. E., Carl H., Møller P. R., Thomsen P. F. (2015). Monitoring the near-extinct European weather loach in Denmark based on environmental DNA from water samples. Biol. Conserv. 183, 46–52. doi: 10.1016/j.biocon.2014.11.023
Simpfendorfer C. A., Kyne P. M., Noble T. H., Goldsbury J., Basiita R. K., Lindsay R., et al. (2016). Environmental DNA detects Critically Endangered largetooth sawfish in the wild. Endang. Species. Res. 30, 109–116. doi: 10.3354/esr00731
Tavares W., da Silva Rodrigues-Filho L. F., Sodré D., Souza R. F., Schneider H., Sampaio I., et al. (2013). Multiple substitutions and reduced genetic variability in sharks. Biochem. Syst. Ecol. 49, 21–29. doi: 10.1016/j.bse.2013.02.004
Thomsen P. F., Kielgast J., Iversen L. L., Møller P. R., Rasmussen M., Willerslev E. (2012a). Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PLoS One 7, e41732. doi: 10.1371/journal.pone.0041732
Thomsen P. F., Kielgast J. O. S., Iversen L. L., Wiuf C., Rasmussen M., Gilbert M. T. P., et al. (2012b). Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 21, 2565–2573. doi: 10.1111/j.1365-294X.2011.05418.x
Thomsen P. F., Willerslev E. (2015). Environmental DNA–An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi: 10.1016/j.biocon.2014.11.019
Turner C. R., Uy K. L., Everhart R. C. (2015). Fish environmental DNA is more concentrated in aquatic sediments than surface water. Biol. Conserv. 183, 93–102. doi: 10.1016/j.biocon.2014.11.017
Valdivia-Carrillo T., Rocha-Olivares A., Reyes-Bonilla H., Domínguez-Contreras J. F., Munguia-Vega A. (2021). Integrating eDNA metabarcoding and simultaneous underwater visual surveys to describe complex fish communities in a marine biodiversity hotspot. Mol. Ecol. Resour. 21, 1558–1574. doi: 10.1111/1755-0998.13375
Keywords: environmental DNA, coastal refugia, smalltooth sawfish, Pristis pectinata, Mexican Caribbean, conservation
Citation: Bonfil R, Díaz-Jaimes P, Palacios-Barreto P, Mendoza Vargas OU and Ricaño-Soriano M (2024) Improved eDNA assay evidences further refugia for critically endangered smalltooth sawfish (Pristis pectinata) in Mexico. Front. Mar. Sci. 11:1290661. doi: 10.3389/fmars.2024.1290661
Received: 07 September 2023; Accepted: 17 January 2024;
Published: 13 February 2024.
Edited by:
James Scott Maki, Marquette University, United StatesReviewed by:
Khaled Mohammed Geba, Menoufia University., EgyptMd Saydur Rahman, The University of Texas Rio Grande Valley, United States
Copyright © 2024 Bonfil, Díaz-Jaimes, Palacios-Barreto, Mendoza Vargas and Ricaño-Soriano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ramón Bonfil, cmFtb24uYm9uZmlsQGVjb3N1ci5teA==