
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 08 January 2024
Sec. Marine Biology
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1288770
This article is part of the Research TopicThe Use of Stable Isotope Ecology in SharksView all 6 articles
 Diego Páez-Rosas1*
Diego Páez-Rosas1* Jenifer Suarez-Moncada2
Jenifer Suarez-Moncada2 Camila Arnés-Urgellés3Eduardo Espinoza2Yolani Robles4
Camila Arnés-Urgellés3Eduardo Espinoza2Yolani Robles4 Pelayo Salinas-De-León5,6
Pelayo Salinas-De-León5,6The scalloped hammerhead shark (Sphyrna lewini) is one of the most iconic marine species across the Eastern Tropical Pacific (ETP), yet little is still known about its reproductive biology and the origin of their nursery areas, despite a continuous global decline in their populations. Understanding the origin of shark nurseries is a complex task; however, it is known that the obtention of resources in the first stages of life of placental sharks occurs through maternal reminiscence, thus it is possible to infer the origin of the energy that neonates accumulate during their gestation. In this study we assessed the ecological connectivity and nursery ground use by undertaking a stable isotope analysis of carbon (δ13C) and nitrogen (δ15N) in skin samples of neonates S. lewini across much of their distribution in the ETP, including the Galapagos Marine Reserve (GMR). Our isotopic mixing models revealed that the S. lewini neonates found in the GMR, have isotopic information similar to adult females inhabit in the Galapagos archipelago, suggesting that they are direct descendants of the females that inhabit the GMR. While neonates found off the continental coasts of Ecuador and Panama have isotopic signatures like those of adult females inhabiting several locations across the ETP. This work provides insight into the origin of nursery areas used by adult S. lewini that aggregate in large numbers across oceanic islands of the ETP, highlighting the value of isotopic models to generate useful information for the conservation of sharks.
Overfishing has caused populations of many shark species to decline dramatically across most ocean basins (Queiroz et al., 2019; Pacoureau et al., 2021). As a result, one third of all shark and ray’s species currently face extinction risks unless urgent policies are implemented to reverse ongoing declines (Dulvy et al., 2021; Yan et al., 2021). The creation of Marine Protected Areas (MPA), where marine life is protected from destructive human activities, such as overfishing, has proven to be an effective tool for protecting key species and ecosystems (Carlisle et al., 2019; White et al., 2020; Klimley et al., 2022). However, many shark species move in and out of the boundaries of MPA during different stages of their life cycles (Knip et al., 2012; Gallagher et al., 2014; Dwyer et al., 2020). Thus, a better understanding of the population’s connectivity of these species is necessary to inform additional management actions aimed at reverting these alarming declines.
The Eastern Tropical Pacific (ETP) is recognized for its relative high abundance of sharks (Salinas-de-León et al., 2016; López-Angarita et al., 2021), among which the populations of the scalloped hammerhead shark (Sphyrna lewini) stand out (Nalesso et al., 2019; Harned et al., 2022). This species occurs circumglobally in tropical and warm temperate waters, occupying habitats spanning from near-shore to pelagic environments (Klimley and Nelson, 1981; Compagno, 1988). Neonates and juveniles of S. lewini live in shallow coastal areas, where individuals may find higher food abundance or lower predation risk compared to other habitats (Gallagher and Klimley, 2018; Zanella et al., 2019); whereas adult sharks migrate to open waters for feeding and likely mating, and then pregnant females return to coastal waters for pupping (Ketchum et al., 2014; Salinas-de-León et al., 2017; Coiraton et al., 2020). Along the ETP, nursery areas for S. lewini have already been identified across coastal areas of the Pacific coasts of central and south America (Alejo-Plata et al., 2007; Zanella et al., 2009; Loor-Andrade et al., 2015; Quintanilla et al., 2015; Robles et al., 2015; Coiraton et al., 2020; Rodriguez-Arana et al., 2022). However, neonates have also been reported in shallow bays of the oceanic islands of Galapagos in Ecuador and Coco Island in Costa Rica (Zanella et al., 2016; Chiriboga-Paredes et al., 2022). Therefore, these oceanic ecosystems may also play an important role in early life stages, and in the connectivity of their populations.
The scalloped hammerhead shark is listed by the IUCN as Critically Endangered (Rigby et al., 2019), nevertheless, this species is still harvested and traded legally and illegally worldwide (Abercrombie et al., 2005; Rigby et al., 2019). Within the ETP, S. lewini form great adult aggregation on oceanic islands such as Galapagos, Malpelo and Cocos (Bessudo et al., 2011; Ketchum et al., 2014; Nalesso et al., 2019), all of which are protected from industrial fishing (Klimley et al., 2022). However, it is necessary to extend these protection mechanisms to the coastal mainland areas of this region, as they are particularly important for reproductive connectivity (López-Angarita et al., 2021; Harned et al., 2022). Despite this, the persistence of S. lewini populations in the ETP remains largely unidentified and likely lacks fishing protection. Hence, it is imperative to apply alternative techniques to identify adult female habitat use from different populations of S. lewini in the ETP and establish its relationship with the scarce nursery areas already identified for this species.
Understanding the foraging strategies of marine predators allows us to infer their ecological aspects, migratory patterns, and population viability (Bird et al., 2018; Rangel-Morales et al., 2022). Several studies regarding the trophic ecology of S. lewini in the ETP have been performed via anecdotal observations and analysis of stomach contents from fisheries catches (Estupiñán-Montaño et al., 2009; Torres-Rojas et al., 2010). Although diet observations are scarce, stomach contents include an array of cephalopods and pelagic fishes from both coastal and oceanic environments (Klimley, 1987; Castañeda-Suárez and Sandoval-Londoño, 2007; Torres-Rojas et al., 2014; Flores-Martínez et al., 2016). However, this information is limited for the entire region due to the existence of Protected Marine Areas where their capture is not allowed (i.e., Galapagos Marine Reserve). The temporal resolution of this technique is another limitation when relating it to the ecological aspects of this species (Rosende-Pereiro et al., 2020; Estupiñán-Montaño et al., 2021a), since the stomach content of a high-consumption predator only provides recent information (i.e., hours or days) of the food consumed prior to its capture (Vaudo and Heithaus, 2011; Kim et al., 2012a), which prevents defining habitat use over time.
On the contrary, the use of stable isotope analysis of carbon (δ13C) and nitrogen (δ15N) is an alternative tool that can provide retrospective information on the foraging patterns of migratory predators, since the isotopic composition of predator reflects the assimilated resource information over time (Graham et al., 2010; Layman et al., 2012). The isotopic signatures of marine primary producers vary depending on the biogeochemical and oceanographic processes in each region, and these differences are spread through local food webs, resulting in consumers with isotopic signatures resembling the food webs they feed on (Hobson et al., 1995; Newsome et al., 2007). Although isotopic baseline characterization can be challenging for highly migratory species (Graham et al., 2010; García-Rodríguez et al., 2021), isotopic data for S. lewini across the ETP has corroborated offshore foraging and intermediate trophic levels of prey consumption such as squid and small pelagic fish (Loor-Andrade et al., 2015; Estupiñán-Montaño et al., 2021b). However, their dietary preferences have also been related to their geographic location and life stages (Rosende-Pereiro et al., 2020; Estupiñán-Montaño et al., 2021c).
Since the energy used by neonates and early juvenile sharks is highly dependent on their maternal energy reserves available in the yolk sacs (McMeans et al., 2009; Vaudo et al., 2010), their tissues reveal a reminiscence from maternal isotopic signatures (Tamburin et al., 2019; Páez-Rosas et al., 2021). Therefore, using isotopic models, it is possible to establish the origin of the energy reserves used by neonates (Olin et al., 2011; Niella et al., 2021), and thus infer the populations from which their mothers originate. The use of Isotopic Mixing Models has been proposed as a complementary method to isotopic analysis because the percentage contributions of different food items to the diet of predators exhibit a substantial isotopic enrichment between consumers and their prey (Malpica-Cruz et al., 2012; Phillips et al., 2014). However, isotopic mixing models require assumptions regarding isotopic enrichment that exists between trophic levels of a predator’s food web. This information is obtained from the Trophic Enrichment Factors (TEFs) between consumers and their sources to accurately assess the isotopic contribution of each prey item to the consumer’s diet (Caut et al., 2009; Hussey et al., 2010). Unfortunately, there are few experimental studies examining TEFs between neonates and female sharks. Therefore, the application of calculated TEFs for related species is an option when applying these analyses (Salinas-de-León et al., 2019).
To understand the reproductive connectivity of S. lewini across the ETP, we determined the δ13C and δ15N signatures of neonates inhabiting different nursery areas of the region. Then we used an isotopic mixture model to evaluate the energy contribution of pregnant females that inhabit the ETP and thus establish the geographical provenance of their mothers. To our knowledge and understanding, this is the first study that evaluates the origin of nursery areas across the ETP used by the adult S. lewini females that seasonally aggregate at the Galapagos Marine Reserve (GMR) and the ETP using isotopic information. Therefore, this information can improve monitoring and conservation strategies for this critically endangered species in the region.
This research was approved by the Galapagos National Park Directorate (GNPD) under research permits PC-27-17, PC-46-18, PC-53-19 granted to Dr. Pelayo Salinas-de-León from the Charles Darwin Foundation (CDF), and PC-24-17, PC-69-18, PC-86-19 granted to Dr. Diego Páez-Rosas from the University San Francisco de Quito (USFQ). The methods described here were reviewed and approved by the GNPD, CDF, and USFQ committees responsible for assessing animal welfare in research activities.
Sampling was carried out in nursery areas on the mainland coasts of Panama, Ecuador, and the oceanic islands of Galapagos in the ETP (Figure 1). A total of 86 tissue biopsies were collected from S. lewini neonates from Panamá (n=31), Ecuador (n=22) and Galapagos Islands (n=33) between 2016 and 2018 (Table 1). A gill net 100 m long and 3 m wide, with a 7.6 cm mesh size was used for sampling, each net was deployed for three hours at the entrance of nursery areas to catch the largest number of specimens. Biological and morphometric data (total length and sex) were recorded for each shark, to subsequently extract a skin sample of the first dorsal using a sterile 6-mm biopsy punch. Since this species is protected within the GMR, we collected only ~10 mg of tissue in the deep layer of the sample to avoid causing damage (Páez-Rosas et al., 2021). After sampling, neonate scalloped hammerhead sharks were reanimated and returned to the sea.
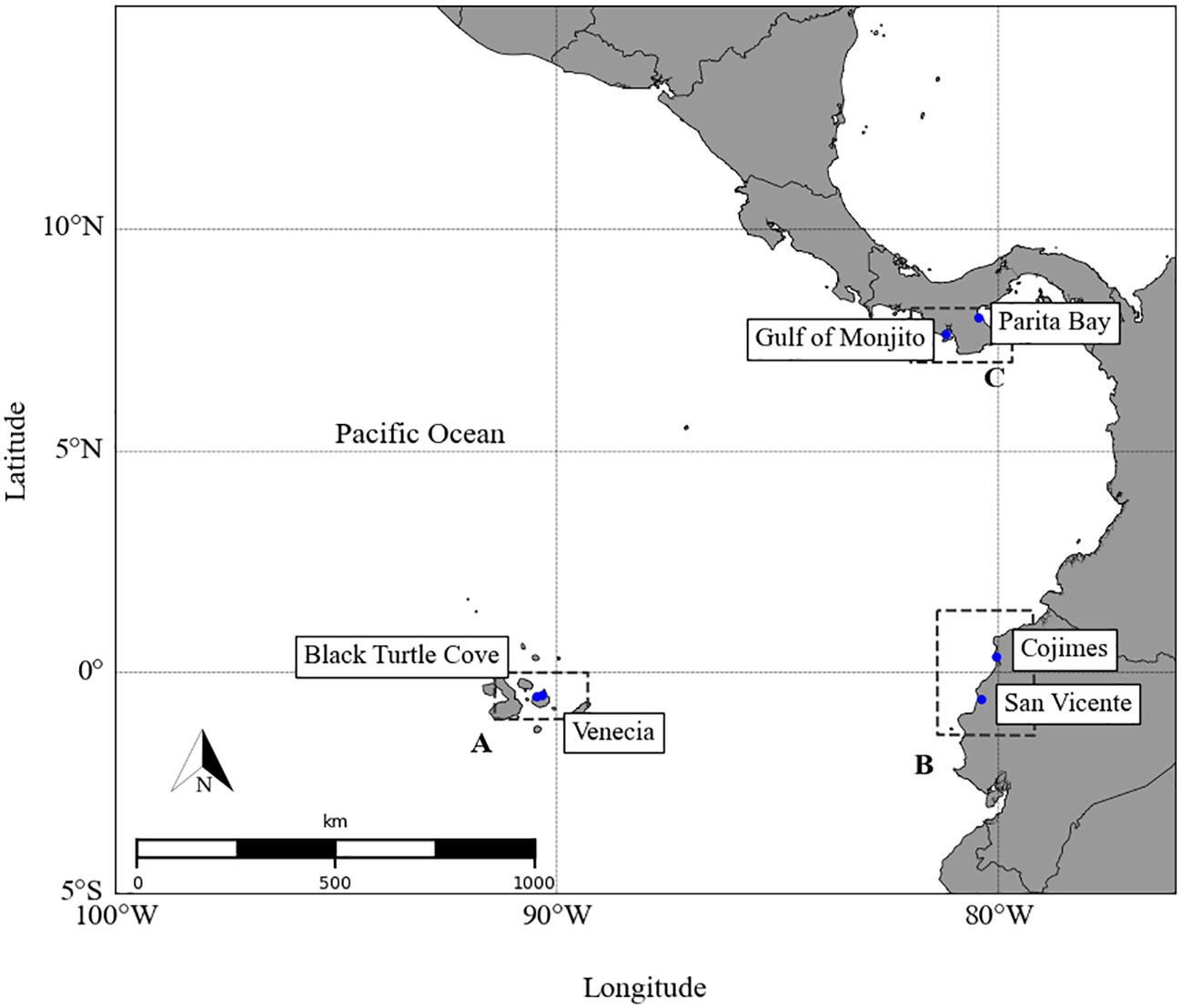
Figure 1 Geographic location of scalloped hammerhead shark nurseries in the Eastern Tropical Pacific. The sampling sites were around coasts of (A) Galapagos, (B) Ecuador and (C) Panamá.
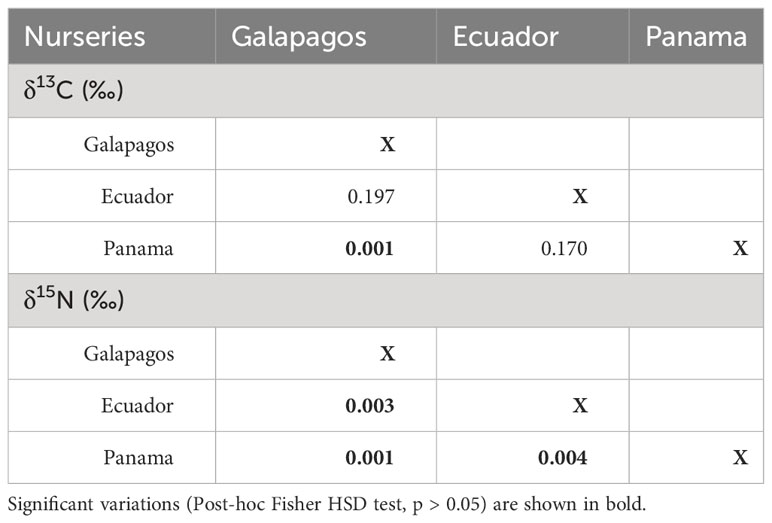
Table 1 P values in comparisons of δ13C and δ15N signatures in neonates scalloped hammerhead shark of different nurseries in the Eastern Tropical Pacific.
All samples were rinsed with distilled water to eliminate residues that could cause contamination and placed in glass vials previously treated with chromic acid (Páez-Rosas et al., 2018). Subsequently, they were dried in an oven at 60 °C for 24 h to remove all moisture, and a lipid extraction protocol was applied via three sequential 24 h soaks in a 2:1 chloroform: methanol solution, then rinsed with deionized water and dried in an oven at 80 °C for 24 h (Páez-Rosas et al., 2020). Each sample was homogenized with an agate mortar and ~1 mg of a fine powder was weighed using an analytical microbalance with a precision of 0.001 mg, and then transferred into a tin capsule for shipment to the Stable Isotope Facility at the University of California Davis (CA, USA).
Isotopic signatures (presented in δ notation) were determined using a PDZ Europa ANCA-GSL elemental analyser interfaced with a PDZ Europa 20-20 isotope ratio mass spectrometer (Sercon Ltd., Cheshire, United Kingdom). Within-run analytical precision was estimated via analysis of two proteinaceous internal standards, which were ±0.2‰ for δ13C and δ15N values. Signatures were expressed in parts per thousand (‰) using the equation: δx = [(Rsample/Rstandard)-1]*1000, where x is the heavy isotope, Rsample is the ratio of heavy to light isotopes in the sample, and Rstandard is the ratio of heavy to light isotopes in the reference standard (DeNiro & Epstein, 1981). The standard reference material for carbon was Pee Dee Belemnite (PDB) and for nitrogen was atmospheric N2. Finally, we measured the weight percentage of carbon and nitrogen concentrations in each sample and used the C/N ratio as a proxy for lipid content (Logan et al., 2008).
To estimate the relative isotopic contributions of female S. lewini inhabiting the ETP to the isotopic signatures of neonate sharks sampled in the region, we used a Bayesian Isotopic Mixing Model (Parnell et al., 2013). The isotopic signatures of females were obtained from published data on this species across the ETP, generating four groups of “potential mothers” according to their sampling distribution: Gulf of Tehuantepec - Mexico (Torres-Rojas et al., 2014), Malpelo island - Colombia (Estupiñán-Montaño et al., 2021a), Manabí coast - Ecuador (Loor-Andrade et al., 2015), and Galapagos Islands - Ecuador (Cerutti-Pereyra et al., 2022). Overall, the mean ± standard deviation for each group was used as the input for the isotope-mixing models. The percentage isotopic contribution of females to neonates was evaluated via SIMMR method (Stable Isotope Mixing Model in R) (Parnell, 2016). This model was run with uninformed priors, four Markov Chain Monte Carlo (MCMC) chains, with 1,000 burn-in and 10,000 iterations, managing to estimate the probability distribution of the contribution of n sources (females) to a mixture and evaluate the uncertainty associated with the isotopic values of the sources and the consumer (Parnell et al., 2010). The variability in the percentage contribution of each source to the neonates was determined by the coefficient of variation, and the results were reported as the percentage contribution of each female group with 97.5% credibility intervals.
Controlled feeding studies have shown that TEFs are both species- and tissue-specific (Hussey et al., 2010). However, TEFs can also be based on values reported in the literature (Phillips et al., 2014). Unfortunately, there are few experimental studies examining TEFs for hammerhead sharks. Therefore, in this work we assumed an average TEF for Δδ15N of 3.01 ± 0.9‰ and 1.3‰± 0.5‰ for Δδ13C, values estimated in controlled feeding experiments by Kim et al. (2012b) and Malpica-Cruz et al. (2012) using muscle samples of leopard sharks (Triakis semifasciata) that inhabit the ETP.
Bayesian Standard Ellipse Areas (SEA) were used to estimate isotopic niche breadth and potential trophic overlap among neonates using the package SIBER (Stable Isotope Bayesian Ellipses in statistical software R); (Jackson et al., 2011). This method creates a convex hull that encompasses all isotopic signatures and generates an ellipse that represents the “core isotopic niche” of consumers (Jackson et al., 2011). Bayesian ellipses provide a measure of the isotopic niche area at the population level, expressed as the SEA in units of area (‰2) and is corrected using posteriori randomly replicated sequences (SEAc Standard Ellipse Area correction); (Jackson et al., 2011). We used Monte Carlo simulations to correct the bivariate ellipses surrounding the data points in the 95% confidence interval for the distributions of δ13C and δ15N values (Jackson et al., 2011). This approach is more robust than other analyses because it is less sensitive to extreme values and small sample sizes, facilitating the estimation of the isotopic niche in sharks of the ETP (Páez-Rosas et al., 2018; Salinas-de-León et al., 2019; Páez-Rosas et al., 2021). In order to quantify possible isotopic overlap between different sampled populations, the magnitude of the isotopic overlap (‰2) among nursery areas was estimated using the estimations of the ellipses via maximum likelihood methods (Jackson et al., 2011).
Finally, we tested normality and homoscedasticity using the Shapiro-Wilk and Levene tests, to examine the statistical differences between the isotopic signatures of the nursery areas. Differences in δ13C and δ15N values were tested using one-way ANOVA along posteriori multiple comparison tests. All statistical analyses were performed in R language (version 3.4.3, The R Foundation for Statistical Computing), using a significance level of P < 0.05.
The total length (TL) of sampled sharks ranged from 55 to 80 cm, with open umbilical scars observed in all individuals. The C/N ratios ranged from 2.73 to 3.44, which is within the theoretical range established for the assimilation of protein from the diet of a predator. The mean δ13C and δ15N values in the skin of S. lewini in Galapagos nurseries were −13.42 ± 0.65‰ and 12.47 ± 0.59‰; in Ecuador −13.93 ± 0.85‰ and 13.99 ± 0.99‰; and in Panama −15.22 ± 0.58‰ and 16.43 ± 0.89‰, respectively (Figure 2). The δ13C and δ15N signatures were significantly different among nurseries (Kruskal–Wallis test, p = 0.001 and p < 0.001, respectively), where δ13C values of Panama differed from those of Galapagos and Ecuador, while δ15N values differed among all nursery areas (multiple comparisons of median ranks, p < 0.005); (Table 1).
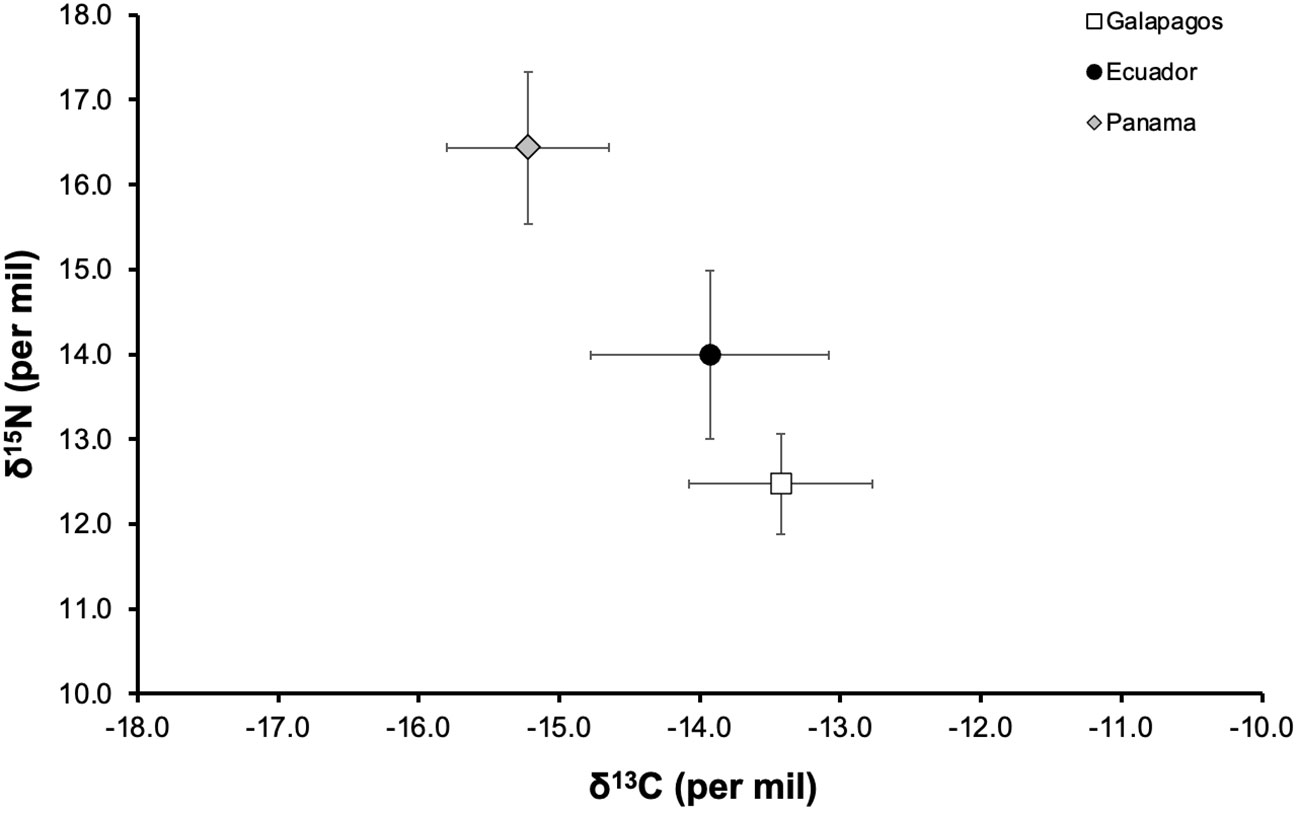
Figure 2 Values of δ13C and δ15N (mean ± SD in ‰) in the scalloped hammerhead shark nurseries in the Eastern Tropical Pacific: Galapagos (white square), Ecuador (black circle) and Panama (grey diamond).
We identified four potential mother groups inhabiting the ETP and executed a SIMMR mixing model to estimate the relative isotopic contribution of each group in different nurseries. The SIMMR revealed the isotopic predominance of S. lewini females inhabiting the GMR in the Galapagos nurseries, representing a mean proportion of 78.5% (Figure 3). The isotopic signatures of females that inhabit the GMR (40.6%), Ecuador (27.1%), and Colombia (26.9%) predominated in the Ecuador nursery (Figure 3). While females inhabiting Colombia (51.8%) and Ecuador (36.4%) presented an isotopic dominance in the Panama nurseries (Figure 3). The SIMMR output values for all groups of potential mothers and their convergence diagnostics to model fit validate are presented in Table 2.
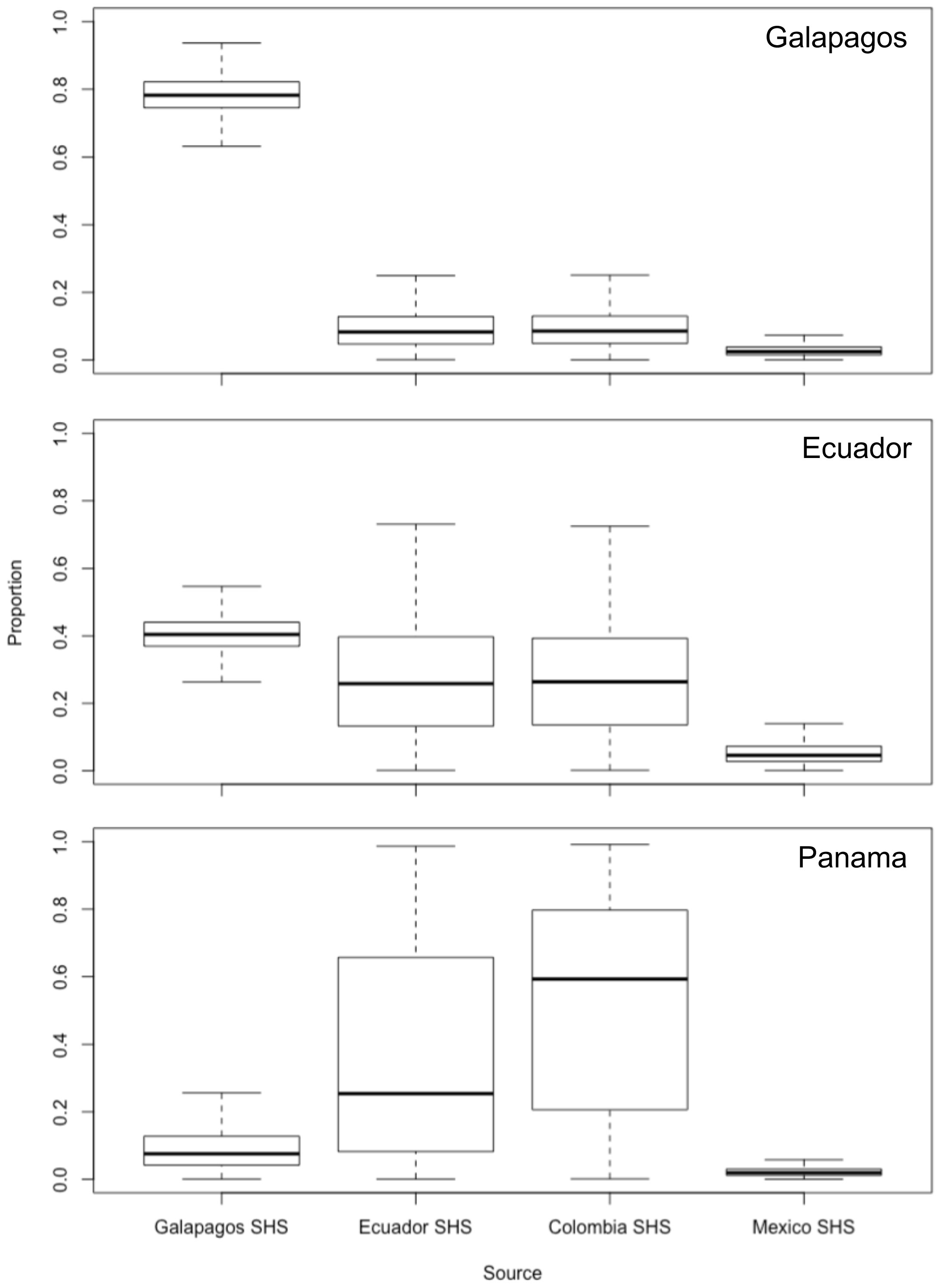
Figure 3 Isotopic contributions (%) to scalloped hammerhead shark nurseries at Eastern Tropical Pacific. Box plot produced by the SIMMR model using individual female S. lewini isotopic values. Boxes show median value and 95%, 75% and 50% credibility intervals.
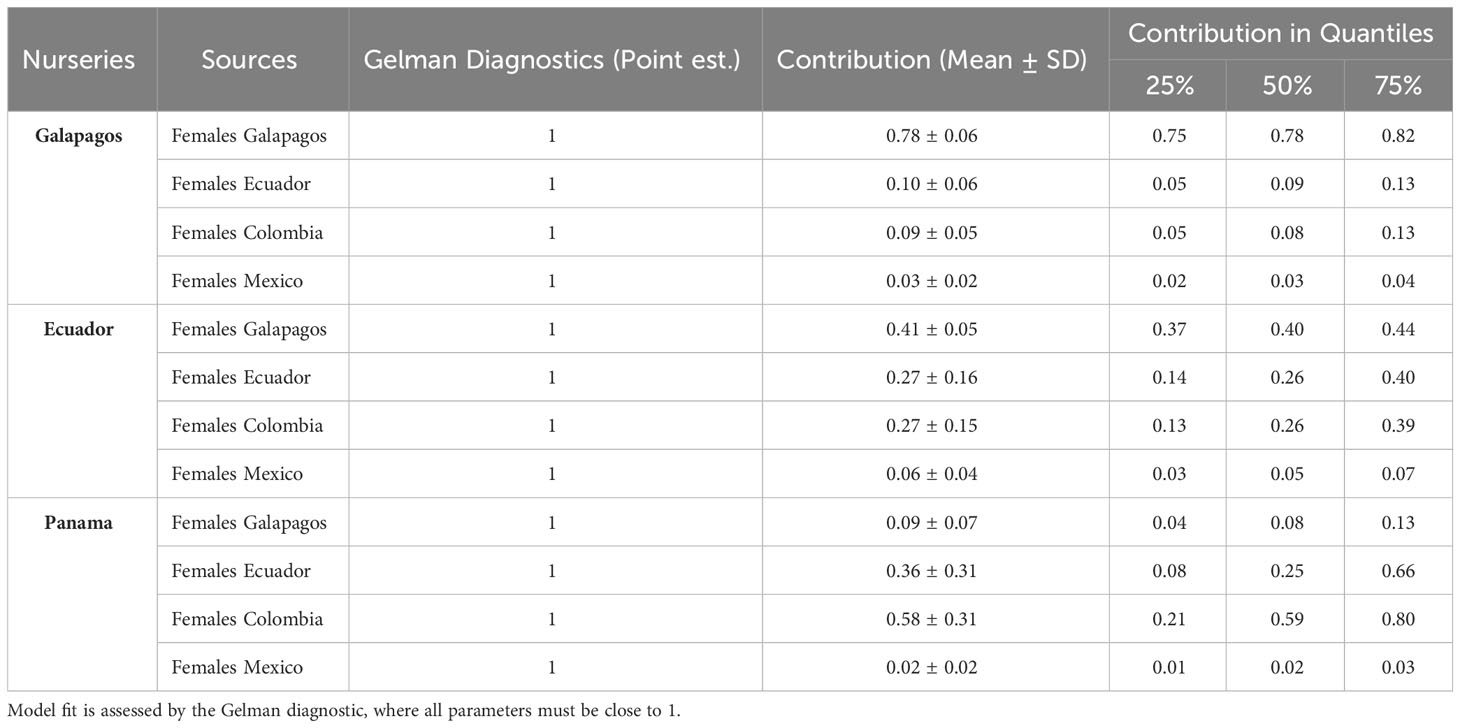
Table 2 Mean proportion, standard deviation (SD) and convergence diagnostics of sources items on the δ13C and δ15N signatures of the neonates scalloped hammerhead shark of different nurseries in the Eastern Tropical Pacific.
The corrected standard ellipse area (SEAc) showed that all S. lewini nurseries reflect different isotopic inputs (Figure 4), thus confirming different resource use patterns for these groups. The Ecuador nursery presented a wider isotopic niche (SEAc = 0.275‰2) compared to Galapagos and Panama nurseries (Figure 4 and Table 3), suggesting a higher diversity of potential mothers (i.e., isotopic contributions from females inhabiting several regions). In contrast, the Galapagos nursery presented the smallest isotopic niche (SEAc = 0.123‰2); (Figure 4), suggesting that the isotopic contributions in this nursery are limited to signatures from females sampled within the GMR.
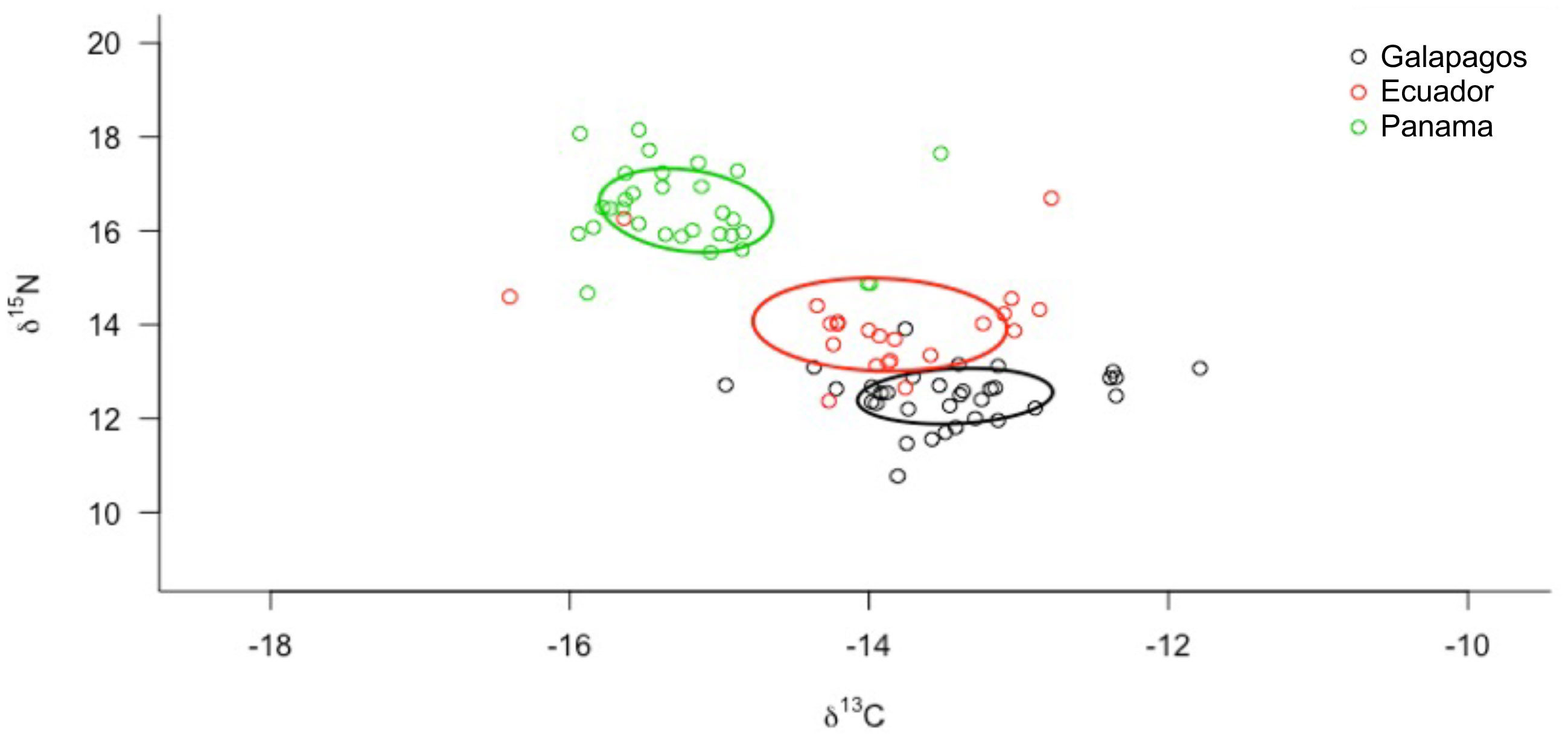
Figure 4 Isotopic niche area (δ13C and δ15N values) of scalloped hammerhead shark nurseries at Eastern Tropical Pacific. Values estimated by a convex hull area and the ellipse corrected for the SIBER analysis.
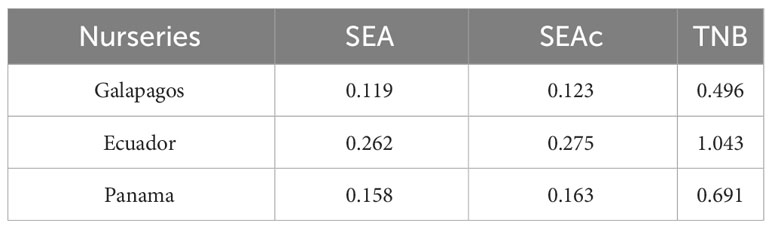
Table 3 Basic Standard Ellipse Area (SEA) and Corrected Standard Ellipse Area (SEAc) measured using Stable Isotope Bayesian Ellipses (SIBER) as an estimate of the isotopic niche breadth (TNB) in different scalloped hammerhead shark nurseries within the Eastern Tropical Pacific.
Isotopic studies investigating neonate shark ecology have shown it is possible to trace biological aspects of females due to the maternal influence on their isotopic signatures (Tamburin et al., 2020; Niella et al., 2021). The most parsimonious explanation for this, is that neonate sharks tissues are influenced by maternal provisioning (McMeans et al., 2009; Hussey et al., 2010), thus the isotopic signatures obtained in neonates reflect the maternal signature, as the development of embryos occurs through the maternal in-uterine nutrient supply (Vaudo et al., 2010; Olin et al., 2011). In elasmobranchs, muscle and skin tissues have slow isotopic turnover due to their low turnover rates (Hussey et al., 2010; Kim et al., 2012b) and they are commonly used to investigate patterns of long-term variation in spatial ecology (Niella et al., 2021). Our results confirm that by applying isotopic enrichment models, it is possible to validate the reproductive philopatry of S. lewini populations across the TEP (Coiraton and Amezcua, 2020).
Neonates of S. lewini are known to occur across the coastlines of the ETP, particularly around the Galapagos Islands, Ecuador, Colombia, Costa Rica, and Mexico (Chiriboga-Paredes et al., 2022; Elizondo-Sancho et al., 2022; Harned et al., 2022; Rodriguez-Arana et al., 2022), with the highest frequency of neonates recorded during the summer months (Klimley, 1987; Corgos and Rosende-Pereiro, 2021). However, the origin of pupping females to each specific nursery remains uncertain, even though it has been proposed that scalloped hammerheads inhabiting the ETP could be philopatric (Hearn et al., 2010; Ketchum et al., 2014; Coiraton and Amezcua, 2020), and that females of this species might use only one calving site per season (Daly-Engel et al., 2012). Our results provide evidence on reproductive philopatric behaviour for GMR nurseries, since neonates within GMR nurseries were found to be more closely related to the isotopic values of adult females inhabiting this region. Despite this, results inferring relatedness from isotopic data from nurseries should be used with caution, since isotopic contributions from GMR females was also observed in the mainland Ecuador nursery, revealing those females sampled around the GMR can also undertake long-range reproductive migrations within the ETP, however, there is still no direct evidence of this (e.g., satellite tracks).
It is known that the female fidelity to specific nurseries in ETP could define reproductive units if females are returning to the same location during each gestation cycle (Marie et al., 2019; Coiraton and Amezcua, 2020; Elizondo-Sancho et al., 2022). In fact, recent genetic evidence on neonates and adults sampled across the TEP has suggested that some female S. lewini could be showing philopatry to Galapagos nurseries (Harned et al., 2022). A regional philopatry in S. lewini has also been inferred in Colombian and Mexican Pacific, where, through molecular analysis, they observed female philopatry in their nurseries (Quintanilla et al., 2015; Rangel-Morales et al., 2022). However, little research exists on specific nursery use in sharks, with patterns so far appearing to be species-specific. Scalloped hammerheads use geomagnetic topotaxis for navigation between seamounts and back to foraging locations offshore with prey availability (Klimley, 1993). Therefore, it is possible for them to return to the aggregation site for their social and reproductive activities (Klimley and Nelson, 1984).
The reduced isotopic niche recorded in the GMR nursery also suggests some residence by adult females. It has been shown that S. lewini females tracked with acoustic transmitters within the GMR remain for long periods of time within the reserve, and the high availability of food given the high productivity of the archipelago has been proposed as the reason behind this residency (Hearn et al., 2010; Ketchum et al., 2014). Previous isotopic analysis have also revealed that these high levels of residency stabilizes their feeding strategies and causes a reduction in their isotopic niche (Arnés-Urgellés et al., 2021; Cerutti-Pereyra et al., 2022). Alternatively, the changes in isotopic niche sizes observed in this study could be a consequence of the fluctuations in the marine productivity and isotopic baseline along ETP (Graham et al., 2010; Páez-Rosas et al., 2020). Therefore, future studies including multi-annual sampling across multipe locations would be required to confirm reproductive philopatry across the region.
Since the migratory capability of S. lewini can reach thousands of kilometres (Klimley et al., 2022), we focused our analysis on specific demographic groups, basing our evaluation on the published isotopic signatures for females (i.e., potential mothers) along the ETP. Our findings in the Ecuadorian and Panamanian nurseries agree with the high genetic connectivity reported for S. lewini across the ETP (Harned et al., 2022). However, these findings contrast with the apparent philopatric behaviour of females in the GMR. Several studies have suggested that S. lewini females may use connected nurseries along the coast for parturition (Duncan et al., 2006; Quintanilla et al., 2015), which would cause greater foraging efforts in coastal populations (Loor-Andrade et al., 2015). Therefore, the females would move away from their usual nurseries. These conditions do not occur in insular habitats, such as the Galapagos Islands, where S. lewini populations have sufficient food (Salinas-de-León et al., 2016), facilitating their permanence on the islands over time (Ketchum et al., 2014; Arnés-Urgellés et al., 2021).
Understanding how sharks use their nursery areas is essential for conservation efforts, since the effectiveness of a marine protected area is closely related to the time that a species spends within its boundaries (MacKeracher et al., 2019; Klimley et al., 2022). The potential philopatric behaviour of S. lewini within the GMR highlights that the reserve could be effectively protecting key pupping grounds for this critically endangered species. Despite this, the overall conservation value of marine protected areas for this species may be limited across the region, since most females use other unprotected nurseries along the ETP, which are currently subject to illegal, unreported, and unregulated fishing (Pérez-Jiménez, 2014; Quintanilla et al., 2015). These conditions highlight the need for a coordinated multinational management cooperation among ETP countries to facilitate the conservation of this species on the brink of extinction.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by Galapagos National Park Directorate. The study was conducted in accordance with the local legislation and institutional requirements.
DP-R: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JS: Data curation, Investigation, Writing – review & editing. CA: Data curation, Investigation, Writing – review & editing. EE: Data curation, Investigation, Writing – review & editing. YR: Data curation, Investigation, Writing – review & editing. PS: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Resources, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Project funding and support was provided by the Helmsley Charitable Trust, the Gordon and Betty Moore Foundation, The Save Our Seas Foundation, USFQ Grants 2018-2019.
We thank the Charles Darwin Foundation, the Galapagos National Park Directorate (GNPD), and Universidad San Francisco de Quito for the institutional support. We also thank the Galapagos Science Centre for providing the facilities for information processing and analysis. Thanks to the E/V Queen Mabel crew, the P/V Sierra Negra, Ecuadorian Navy R/V Sirius for their fieldwork support, as well as the rangers and CDF volunteers that assisted with this study. We are also grateful to the artisanal fishermen from Ecuador and Panama that contributed with samples for analysis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer, AT, declared a shared parent affiliation with the author, CA.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abercrombie D. L., Clarke S. C., Shivji M. S. (2005). Global-scale genetic identification of hammerhead sharks: Application to assessment of the international fin trade and law enforcement. Conserv. Genet. 6, 775–788. doi: 10.1007/s10592-005-9036-2
Alejo-Plata C., Gómez-Márquez J. L., Ramos S., Herrera E. (2007). Presencia de neonatos y juveniles del tiburón martillo Sphyrna lewini (Griffith & Smith 1834) y del tiburón sedoso Carcharhinus falciformis (Müller & Henle 1839) en la costa de Oaxaca, México. Rev. Biol. Mar. Oceanogr. 42, 403–413. doi: 10.4067/S0718-19572007000300020
Arnés-Urgellés C., Salinas-de-León P., Rastoin-Laplane E., Vaca-Pita L., Suárez-Moncada J., Páez-Rosas D. (2021). The effects of climatic variability on the feeding ecology of the scalloped hammerhead shark (Sphyrna lewini) in the tropical eastern Pacific. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.625748
Bessudo S., Soler G. A., Klimley A. P., Ketchum J. T., Hearn A., Arauz R. (2011). Residency of the scalloped hammerhead shark (Sphyrna lewini) at Malpelo Island and evidence of migration to other islands in the eastern tropical Pacific. Envir. Biol. Fish. 91, 165–176. doi: 10.1007/s10641-011-9769-3
Bird C. S., Veríssimo A., Magozzi S., Abrantes K. G., Aguilar A., Al-Reasi H., et al. (2018). A global perspective on the trophic geography of sharks. Nat. Ecol. Evol. 2, 299–305. doi: 10.1038/s41559-017-0432-z
Carlisle A. B., Tickler D., Dale J. J., Ferretti F., Curnick D. J., Chapple T. K., et al. (2019). Estimating space use of mobile fishes in a large marine protected area with methodological considerations in acoustic array design. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00256
Castañeda-Suárez J. D., Sandoval-Londoño L. J. (2007). “Hábitos alimentarios del tiburón martillo, Sphyrna lewini (Griffith & Smith 1834) en el Pacífico Ecuatoriano,” in Tiburones en el Ecuador: casos de estudio. Eds. Martínez-Ortiz J., Galvan-Magana F. (Manta: EPESPO-PMRC Press), 66–76.
Caut S., Angulo E., Courchamp F. (2009). Variation in discrimination factors (d15N and d13C): the effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 46, 443–453. doi: 10.1111/j.1365-2664.2009.01620.x
Cerutti-Pereyra F., Salinas-De-León P., Arnés-Urgellés C., Suarez-Moncada J., Espinoza E., Vaca L., et al. (2022). Using stable isotopes analysis to understand ontogenetic trophic variations of the scalloped hammerhead shark at the Galapagos Marine Reserve. PLoS One 17, e0268736. doi: 10.1371/journal.pone.0268736
Chiriboga-Paredes Y., Palomino Á., Goodman L., Córdova F., Páez V., Yépez M., et al. (2022). Discovery of a putative scalloped hammerhead shark Sphyrna lewini (Carcharhiniformes: Sphyrnidae) nursery site at the Galapagos Islands, Eastern Tropical Pacific. Environ. Biol. Fish. 105, 181–192. doi: 10.1007/s10641-021-01207-3
Coiraton C., Amezcua F. (2020). In utero elemental tags in vertebrae of the scalloped hammerhead shark Sphyrna lewini reveal migration patterns of pregnant females. Sci. Rep. 10, 1799. doi: 10.1038/s41598-020-58735-8
Coiraton C., Amezcua F., Ketchum J. (2020). New insights into the migration patterns of the scalloped hammerhead shark Sphyrna lewini based on vertebral microchemistry. Mar. Biol. 167, 58. doi: 10.1007/s00227-020-3668-0
Compagno L. J. V. (1988). Sharks of the order Carcharhiniformes (Princeton: Princeton University Press).
Corgos A., Rosende-Pereiro A. (2021). Nursery habitats use patterns of the scalloped hammerhead shark, Sphyrna lewini, in coastal areas of the central Mexican Pacific. J. Fish Biol. 100, 117–133. doi: 10.1111/jfb.14925
Daly-Engel T. S., Seraphin K. D., Holland K. N., Coffey J. P., Nance H. A., Toonen R. J., et al. (2012). Global phylogeography with mixed-marker analysis reveals male-mediated dispersal in the endangered scalloped hammerhead shark (Sphyrna lewini). PloS One 7, e29986. doi: 10.1371/journal.pone.0029986
DeNiro M. J., Epstein S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 45, 341–351. doi: 10.1016/0016-7037(81)90244-1
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4778. doi: 10.1016/j.cub.2021.08.062
Duncan K. M., Martin A. P., Bowen B. W., De Couet H. G. (2006). Global phylogeography of the scalloped hammerhead shark (Sphyrna lewini). Mol. Ecol. 15, 2239–2251. doi: 10.1111/j.1365-294X.2006.02933.x
Dwyer R. G., Krueck N. C., Udyawer V., Heupel M. R., Chapman D., Pratt H. L., et al. (2020). Individual and population benefits of marine reserves for reef sharks. Curr. Biol. 30, 480–489. doi: 10.1016/j.cub.2019.12.005
Elizondo-Sancho M., Rodríguez-Arriatti Y., Albertazzi F. J., Bonilla-Salazar A., Arauz-Naranjo D., Arauz R., et al. (2022). Population structure and genetic connectivity of the scalloped hammerhead shark (Sphyrna lewini) across nursery grounds from the Eastern Tropical Pacific: Implications for management and conservation. PLoS One 17, e0264879. doi: 10.1371/journal.pone.0264879
Estupiñán-Montaño C., Cedeño-Figueroa L., Galván-Magaña F. (2009). Hábitos alimentarios del tiburón martillo Sphyrna lewini (Griffith & Smith 1834) (Chondrichthyes) en el Pacífico ecuatoriano. Rev. Biol. Mar. Oceanogr. 44, 379–386. doi: 10.4067/S0718-19572009000200011
Estupiñán-Montaño C., Galván-Magaña F., Elorriaga-Verplancken F., Zetina-Rejón M., Sánchez-González A., Polo-Silva C., et al. (2021a). Ontogenetic feeding ecology of the scalloped hammerhead shark Sphyrna lewini in the Colombian Eastern Tropical Pacific. Mar. Ecol. Prog. Ser. 663, 127–143. doi: 10.3354/meps13639
Estupiñán-Montaño C., Tamburin E., Delgado-Huertas A. (2021b). New insights into the trophic ecology of the scalloped hammerhead shark, Sphyrna lewini, in the eastern tropical Pacific Ocean. Envir. Biol. Fish. 104, 1611–1627. doi: 10.1007/s10641-021-01187-4
Estupiñán-Montaño C., Tamburin E., Delgado-Huertas A. (2021c). Stable isotope evidence for movements of hammerhead sharks Sphyrna lewini, connecting two natural protected areas in the Colombian Pacific. Mar. Biodivers. 51, 74. doi: 10.1007/s12526-021-01215-7
Flores-Martínez I. A., Torres-Rojas Y. E., Galván-Magaña F. (2016). Diet comparison between silky sharks (Carcharhinus falciformis) and scalloped hammerhead sharks (Sphyrna lewini) off the south-west coast of Mexico. J. Mar. Biol. Assoc United Kingdom 97, 337–345. doi: 10.1017/S0025315416000424
Gallagher A. J., Hammerschlag N., Shiffman D. S., Giery S. T. (2014). Evolved for extinction: the cost and conservation implications of specialization in hammerhead sharks. Bioscience 64, 619–624. doi: 10.1093/biosci/biu071
Gallagher A. J., Klimley A. P. (2018). The biology and conservation status of the large hammerhead shark complex: The great, scalloped, and smooth hammerheads. Rev. Fish Biol. Fisheries. 28, 777–794. doi: 10.1007/s11160-018-9530-5
García-Rodríguez E., Herza S. Z., Sosa-Nishizaki O., Lowe C., O´Sullivan J. B. (2021). Stable isotope analysis of juvenile white sharks inside a nursery area reveals foraging in demersal-inshore habitats and trophic overlap with sympatric sharks. Front. Mar. Sci 8, 687738. doi: 10.3389/fmars.2021.687738
Graham B. S., Koch P. L., Newsome S. D., McMahon K. W., Aurioles-Gamboa D. (2010). “Using isoscapes to trace the movements and foraging behavior of top predators in oceanic ecosystems,” in Isoscapes: Understanding movement, pattern, and process on Earth through isotope mapping. Eds. West J. B., Bowen G. J., Dawson T. E., Tu K. P. (Dordrecht: Springer Netherlands). doi: 10.1007/978-90-481-3354-3_14
Harned S. P., Bernard A. M., Salinas-de-León P., Mehlrose M. R., Suarez J., Robles Y., et al. (2022). Genetic population dynamics of the critically endangered scalloped hammerhead shark (Sphyrna lewini) in the Eastern Tropical Pacific. Ecol. Evol. 12, e9642. doi: 10.1002/ece3.9642
Hearn A., Ketchum J., Klimley A. P., Espinoza E., Peñaherrera C. (2010). Hotspots within hotspots? Hammerhead shark movements around Wolf Island, Galapagos Marine Reserve. Mar. Biol. 157, 1899–1915. doi: 10.1007/s00227-010-1460-2
Hobson K. A., Ambrose W. G. Jr., Renaud P. E. (1995). Sources of primary production, benthic-pelagic coupling, and trophic relationships within the Northeast Water Polynya: insights from d13C and d15N analysis. Mar. Ecol. Prog. Ser. 128, 1–10. doi: 10.3354/meps128001
Hussey N. E., Brush J., McCarthy I. D., Fisk A. T. (2010). d15N and d13C diet-tissue discrimination factors for large sharks under semi-controlled conditions. Comp. Biochem. Physiol. A. 155, 445e453. doi: 10.1016/j.cbpa.2009.09.023
Jackson A., Inger R., Parnell A., Bearhop S. (2011). Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses. J. Animal. Ecol. 80, 595–602. doi: 10.1111/j.1365-2656.2011.01806.x
Ketchum J. T., Hearn A., Klimley A. P., Peñaherrera C., Espinoza E., Bessudo S., et al. (2014). Inter-island movements of scalloped hammerhead sharks (Sphyrna lewini) and seasonal connectivity in a marine protected area of the Eastern Tropical Pacific. Mar. Biol. 161, 939–951. doi: 10.1007/s00227-014-2393-y
Kim S. L., Martínez del Rio C., Casper D., Koch P. L. (2012b). Isotopic incorporation rates for shark tissues from a long-term captive feeding study. J. Exp. Biol. 215, 2495–2500. doi: 10.1242/jeb.070656
Kim S. L., Tinker M. T., Estes J. A., Koch P. L. (2012a). Ontogenetic and among-individual variation in foraging strategies of northeast Pacific white sharks based on stable isotope analysis. PLoS One 7, e45068. doi: 10.1371/journal.pone.0045068
Klimley A. P. (1987). The determinants of sexual segregation in the scalloped hammerhead shark, Sphyrna lewini. Environ. Biol. Fish. 18, 27–40. doi: 10.1007/BF00002325
Klimley A. P. (1993). Highly directional swimming by scalloped hammerhead sharks, Sphyrna lewini, and subsurface irradiance, temperature, bathymetry, and geomagnetic field. Mar. Biol. 117, 1–22. doi: 10.1007/BF00346421
Klimley A. P., Arauz R., Bessudo S., Chávez E. J., Chinacalle N., Espinoza E., et al. (2022). Studies of the movement ecology of sharks justify the existence and expansion of marine protected areas in the Eastern Pacific Ocean. Environ. Biol. Fish. 105, 2133–2153. doi: 10.1007/s10641-021-01204-6
Klimley A. P., Nelson D. R. (1981). Schooling of the scalloped hammerhead shark, Sphyrna lewini, in the Gulf of California. Fish. Bull. 79 (2), 356–360.
Klimley A. P., Nelson D. R. (1984). Diel movement patterns of the scalloped hammerhead shark (Sphyrna lewini) in relation to El Bajo Espiritu Santo: a refuging central position social system. Behav. Ecol. Sociobiol. 15, 45–54. doi: 10.1007/BF00310214
Knip D. M., Heupel M. R., Simpfendorfer C. A. (2012). Evaluating marine protected areas for the conservation of tropical coastal sharks. Biol. Conserv. 148, 200–209. doi: 10.1016/j.biocon.2012.01.008
Layman A., Araújo M. S., Boucek R., Hammerschlag-Peyer C. M., Harrison E., Jud Z. R., et al. (2012). Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol. Rev. 87, 545–562. doi: 10.1111/j.1469-185X.2011.00208.x
Logan J., Jardine T., Miller T. J., Bunn S., Cunjak R., Lutcavage M. (2008). Lipid corrections in carbon and nitrogen stable isotope analyses: comparison of chemical extraction and modelling methods. J. Anim. Ecol. 77, 838–846. doi: 10.1111/j.1365-2656.2008.01394.x
Loor-Andrade P., Galván-Magaña F., Elorriaga-Verplancken F. R., Polo-Silva C., Delgado-Huertas A. (2015). Population and individual foraging patterns of two hammerhead sharks using carbon and nitrogen stable isotopes. Rapid Commun. Mass Spectrom. 29, 821–829. doi: 10.1002/rcm.7169
López-Angarita J., Villate-Moreno M., Diáz J. M., Cubillos-M J. C., Tilley A. (2021). Identifying nearshore nursery habitats for sharks and rays in the Eastern Tropical Pacific from fishers’ knowledge and landings. Ocean Coast. Manage. 213, 105825. doi: 10.1016/j.ocecoaman.2021.105825
MacKeracher T., Diedrich A., Simpfendorfer C. A. (2019). Sharks, rays and marine protected areas: A critical evaluation of current perspectives. Fish Fish. 20, 255–267. doi: 10.1111/faf.12337
Malpica-Cruz L., Herzka S. Z., Sosa-Nishizaki O., Lazo J. P. (2012). Tissue-specific isotope trophic discrimination factors and turnover rates in a marine elasmobranch: empirical and modeling results. Can. J. Fish. Aquat. Sci. 69, 551–564. doi: 10.1139/f2011-172
Marie A. D., Herbinger C., Fullsack P. M., Rico C. (2019). First reconstruction of kinship in a scalloped hammerhead shark aggregation reveals the mating patterns and breeding sex ratio. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00676
McMeans B. C., Olin J. A., Benz G. W. (2009). Stable-isotope comparisons between embryos and mothers of a placentatrophic shark species. J. Fish Biol. 75, 2464–2474. doi: 10.1111/j.1095-8649.2009.02402.x
Nalesso E., Hearn A., Sosa-Nishizaki O., Steiner T., Antoniou A., Reid A., et al. (2019). Movements of scalloped hammerhead sharks Sphyrna lewini at Cocos Island, Costa Rica, and between oceanic islands in the Eastern Tropical Pacific. PloS One 14, e0213741. doi: 10.1371/journal.pone.0213741
Newsome S. D., Martínez del Rio C., Bearhop S., Phillips D. L. (2007). A niche for isotopic ecology. Front. Ecol. Environ. 5, 429–436. doi: 10.1890/060150.1
Niella Y., Raoult V., Gaston T., Peddemors V. M., Harcourt R., Smoothey A. F. (2021). Overcoming multi-year impacts of maternal isotope signatures using multi-tracers and fast turnover tissues in juvenile sharks. Chemosphere 269, 129393. doi: 10.1016/j.chemosphere.2020.129393
Olin J. A., Hussey N. E., Fritts M., Heupel M. R., Simpfendorfer C. A., Poulakis G. R., et al. (2011). Maternal meddling in neonatal sharks: implications for interpreting stable isotopes in young animals. Rapid Commun. Mass Spectrom. 25, 1008–1016. doi: 10.1002/rcm.4946
Pacoureau N., Rigby C., Kyne P., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a centry of global decline in oceanic sharks and rays. Nature 589, 567–571. doi: 10.1038/s41586-020-03173-9
Páez-Rosas D., Galván-Magaña F., Baque-Menoscal J., Tripp-Valdez A., Fischer C., Hearn A. (2020). Trophic preferences of three pelagic fish inhabiting the Galapagos marine reserve: ecological inferences using multiple analyses. Environ. Biol. Fish. 103, 647–665. doi: 10.1007/s10641-020-00967-8
Páez-Rosas D., Insuasti-Zarate P., Riofrío-Lazo M., Galván-Magaña F. (2018). Feeding behavior and trophic interaction of three shark species in the Galapagos Marine Reserve. PeerJ 6, e4818. doi: 10.7717/peerj.4818
Páez-Rosas D., Suarez-Moncada J., Elorriaga-Verplancken F. R., Proaño A., Arnés-Urgellés C., Salinas-de-León P., et al. (2021). Trophic variation during the early stages of blacktip sharks (Carcharhinus limbatus) within coastal nurseries of the Galapagos Marine Reserve. J. Sea Res. 102023. doi: 10.1016/j.seares.2021.102023
Parnell A. C. (2016). Simmr: a stable isotope mixing model (R package version 0, 3). Available at: https://CRAN.R-project.org/package=simmr.
Parnell A. C., Inger R., Bearhop S., Jackson A. (2010). Source partitioning using stable isotopes: coping with too much variation. PLoS One 5, e9672. doi: 10.1371/journal.pone.0009672
Parnell A. C., Phillips D. L., Bearhop S., Semmens B. X., Ward E. J., Moore J. W., et al. (2013). Bayesian stable isotope mixing models. Environmetrics 24, 387–399. doi: 10.1002/env.2221
Pérez-Jiménez J. C. (2014). Historical records reveal potential extirpation of four hammerhead sharks (Sphyrna spp.) in Mexican Pacific waters. Rev. Fish. Biol. Fisheries. 24, 671–683. doi: 10.1007/s11160-014-9353-y
Phillips D. L., Inger R., Bearhop S., Jackson A. L., Moore J. W., Parnell A. C., et al. (2014). Best practices for use of stable isotope mixing models in food-web studies. Fish. Aquat. Sci. 92, 823–835. doi: 10.1139/cjz-2014-0127
Queiroz N., Humphries N., Couto A., Vedor M., Costa I., Sequeira A., et al. (2019). Global spatial risk assessment of sharks under the footprint of fisheries. Nature 572, 461–466. doi: 10.1038/s41586-019-1444-4
Quintanilla S., Gómez A., Mariño-Ramírez C., Sorzano C., Bessudo S., Soler G., et al. (2015). Conservation genetics of the scalloped hammerhead shark in the Pacific Coast of Colombia. J. Hered. 106, 448–458. doi: 10.1093/jhered/esv050
Rangel-Morales J. M., Rosales-López L. P., Díaz-Jaimes P., Amezcua-Martínez F., Ketchum J., Hoyos-Padilla M., et al. (2022). Regional philopatry of scalloped hammerhead sharks (Sphyrna lewini) to nursery areas in the Mexican Pacific. Hydrobiologia 849, 3083–3099. doi: 10.1007/s10750-022-04880-2
Rigby C. L., Dulvy N. K., Barreto R., Carlson J., Fernando D., Fordham S., et al. (2019). Sphyrna lewini (The IUCN Red List of Threatened Species). Available at: https://www.iucnredlist.org/es/species/39385/2918526/.
Robles P. Y. A., Montes L. A., Vega Á.J. (2015). Caracterización de la captura de tiburones por la pesca artesanal en los manglares de David, Golfo de Chiriquí, Pacífico de Panamá. Tecnociencia 17, 11–30.
Rodriguez-Arana J. P., Hernández S., González-Armas R., Galván-Magaña F., Tripp-Valdez A., Hoyos-Padilla M., et al. (2022). A priority nursery area for the conservation of the scalloped hammerhead shark Sphyrna lewini in Mexico. J. Fish Biol. 101, 1623–1627. doi: 10.1111/jfb.15227
Rosende-Pereiro A., Flores-Ortega J. R., González-Sansón G., Corgos A. (2020). Stomach content and stable isotopes reveal an ontogenetic dietary shift of young-of-the-year scalloped hammerhead sharks (Sphyrna lewini) inhabiting coastal nursery areas. Envir. Biol. Fish. 103, 49–65. doi: 10.1007/s10641-019-00932-0
Salinas-de-León P., Acuña-Marrero D., Rastoin E., Friedlander A. M., Donovan M. K., Sala E. (2016). Largest global shark biomass found in the northern Galapagos Islands of Darwin and Wolf. PeerJ 4, e1911. doi: 10.7717/peerj.1911
Salinas-de-León P., Fierro-Arcos D., Suarez-Moncada J., Proaño A., Guachisaca-Salinas J., Páez-Rosas D. (2019). A matter of taste: spatial and ontogenetic variations on the trophic ecology of the tiger shark at the Galapagos Marine Reserve. PLoS One 14, 9. doi: 10.1371/journal.pone.0222754
Salinas-de-León P., Hoyos-Padilla E. M., Pochet F. (2017). First observation on the mating behaviour of the endangered scalloped hammerhead shark Sphyrna lewini in the tropical eastern Pacific. Envir. Biol. Fish. 100, 1603–1608. doi: 10.1007/s10641-017-0668-0
Tamburin E., Amador-Capitanachi M. J., Galván-Magaña F., Norris T., Elorriaga- Verplancken F. R. (2020). Isotopic variation in blood components based on their biochemistry and physiology: a comparison between sharks and fur seals. J. Exp. Zool. A Ecol. Integr. Physiol. 333, 104–110. doi: 10.1002/jez.2330
Tamburin E., Kim S. L., Elorriaga-Verplancken F. R., Madigan D. J., Hoyos-Padilla M., Sánchez-González A., et al. (2019). Isotopic niche and resource sharing among young sharks (Carcharodon carcharias and Isurus oxyrinchus) in Baja California, Mexico. Mar. Ecol. Prog. Ser. 613, 107–124. doi: 10.3354/meps12884
Torres-Rojas Y. E., Hernández-Herrera A., Galván-Magaña F., Alatorre-Ramírez V. G. (2010). Stomach content analysis of juvenile, scalloped hammerhead shark Sphyrna lewini captured off the coast of Mazatlán, Mexico. Aquat. Ecol. 44, 301–308. doi: 10.1007/s10452-009-9245-8
Torres-Rojas Y. E., Páez-Osuna F., Camalich J., Galván-Magaña F. (2014). Diet and trophic level of scalloped hammerhead shark (Sphyrna lewini) from the Gulf of California and Gulf of Tehuantepec, Mexico. Iran. J. Fish. Sci. 14, 767–785.
Vaudo J. M., Heithaus M. R. (2011). Dietary niche overlap in a nearshore elasmobranch mesopredator community. Mar. Ecol. Prog. Ser. 425, 247–260. doi: 10.3354/meps08988
Vaudo J. J., Matich P., Heithaus R. (2010). Mother–offspring isotopes fractionation two species if placentatrophic sharks. J. Fish Biol. 77, 1724–1727. doi: 10.1111/j.1095-8649.2010.02813.x
White T. D., Ong T., Ferretti F., Block B. A., McCauley D. J., Micheli F., et al. (2020). Tracking the response of industrial fishing fleets to large marine protected areas in the Pacific Ocean. Conserv. Biol. 34, 1571–1578. doi: 10.1111/cobi.13584
Yan H. F., Kyne P. M., Jabado R. W., Leeney R. H., Davidson L. N. (2021). Overfishing and habitat loss drives range contraction of iconic marine fishes to near extinction. Sci. Adv. 7, eabb6026. doi: 10.1126/sciadv.abb6026
Zanella I., López-Garro A. L., Cure K. (2019). Golfo Dulce: critical habitat and nursery area for juvenile scalloped hammerhead sharks Sphyrna lewini in the Eastern Tropical Pacific Seascape. Envir. Biol. Fish. 102, 1291–1300. doi: 10.1007/s10641-019-00907-1
Zanella I., López-Garro A. L., McComb-Kobza D. M., Golfín-Duarte G., Pérez-Montero M., Morales J. (2016). First record of young-of-the-year scalloped hammerhead shark, Sphyrna lewini (Carcharhiniformes: Sphyrnidae) from Isla del Coco National Park, Costa Rica. Rev. Biol. Trop. 64, 201. doi: 10.15517/rbt.v64i1.23448
Keywords: Eastern Tropical Pacific, isotopic mixing models, maternal reminiscence, shark nurseries, Sphyrna lewini
Citation: Páez-Rosas D, Suarez-Moncada J, Arnés-Urgellés C, Espinoza E, Robles Y and Salinas-De-León P (2024) Assessment of nursery areas for the scalloped hammerhead shark (Sphyrna lewini) across the Eastern Tropical Pacific using a stable isotopes approach. Front. Mar. Sci. 10:1288770. doi: 10.3389/fmars.2023.1288770
Received: 04 September 2023; Accepted: 15 December 2023;
Published: 08 January 2024.
Edited by:
Carlos Polo, Universidad de Bogotá Jorge Tadeo Lozano, ColombiaReviewed by:
James T. Ketchum, Pelagios Kakunjá, MexicoCopyright © 2024 Páez-Rosas, Suarez-Moncada, Arnés-Urgellés, Espinoza, Robles and Salinas-De-León. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego Páez-Rosas, ZHBhZXpAdXNmcS5lZHUuZWM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.