
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 03 October 2023
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1234772
This article is part of the Research TopicBlue Foods Security and SustainabilityView all 15 articles
 Xiaofan Hong1,2,3
Xiaofan Hong1,2,3 Kui Zhang1,2
Kui Zhang1,2 Jiajun Li1,2
Jiajun Li1,2 Youwei Xu1,2
Youwei Xu1,2 Mingshuai Sun1,2
Mingshuai Sun1,2 Jingyuan Jiang4
Jingyuan Jiang4 Shannan Xu1,2Yancong Cai1,2
Shannan Xu1,2Yancong Cai1,2 Yongsong Qiu1,2
Yongsong Qiu1,2 Zuozhi Chen1,2*
Zuozhi Chen1,2*A detailed understanding of the impact of climate variability on fish life histories provides a fundamental basis for the ecosystem approach to fisheries management. In this study, we evaluated the relationship between fish life-history trait parameters and climate periods, using survey data from 2006−2020 for the Beibu Gulf of the northwestern South China Sea, a fishing ground with high commercial and ecological value. We show that climate events can significantly alter the structure of the life history for major commercial fishes, with more diverse life-history strategies during warm events and more concentrated mortality during cold events. Additionally, we detected special relationships between climate events and the life-history trait composite index (LTCI) in Psenopsis anomala and Trichiurus lepturus. It is possible that other indicators would be more appropriate than variability in Oceanic Niño Index (ONI), such as variation in fishing effort or the environmental resistance of fish. Anthropogenic disturbance and biological features may also help explain the magnitude of population variability. We found that changes in life-history traits of only a few commercial fishes (Decapterus maruadsi, Pennahia macrocephalus, and Upeneus sulphureus) were related to the shift in climate periods; these fishes were generally characterized by faster growth and higher mortality rates in the normal and El Niño periods, with the opposite trends in La Niña periods. However, the emergence of climate events has led to a clear complementarity of life-history strategies among some fish in the same ecological niche (same genus and/or feeding group), which may be explained by a balance between habitat conditions and fishing pressures. This study of fish life-history strategies under climate anomalies provides key insights into important attributes for managers to consider when implementing relevant measures to promote fisheries sustainability in the subtropical bay.
Climate anomalies, such as the El Niño–Southern Oscillation (ENSO), are one of the main drivers of marine ecosystem changes (Timmermann et al., 2018). Although the major signal of ENSO occurs in the equatorial Pacific with an intensity that can vary considerably from one event to another, it is unique among climate phenomena in its great influence on the patterns of weather variability worldwide (Lehodey et al., 2006; Mcphaden et al., 2006). The physical-biogeochemical changes in the marine ecosystem caused by ENSO affect the distribution and biomass of plankton, fish and invertebrates that depend on them (Gomez et al., 2019). The literature devoted to effects of climate variability on these components of the marine ecosystem is rapidly growing (Holbrook et al., 2020), as is also the case for the literature on freshwater and terrestrial ecosystems. However, there is growing evidence for interactions between climate change and ENSO dynamics and effects, complicating analyses of underlying mechanisms (Lehodey et al., 2020). A better understanding of these mechanisms will be useful for the exploration of relationship between climate anomalies and fishery resources, as well as formulate polices for fisheries management.
From the tropical Pacific open sea to the coasts of Asia and the Americas, ENSO affects the mortality, recruitment, and habitat distribution of commercially valuable fishes (Stenseth et al., 2002). ENSO events provide an excellent opportunity to observe and evaluate how changes in habitats influence the life-history traits, abundance, or distribution of fishes. Identifying changes in these population characteristics will increase understanding of the habitat requirements of important marine resources and will help local government departments and/or organizations better manage fisheries. Life-history traits and population dynamics of fish depend closely on environmental conditions (Chaparro-Pedraza and De Roos, 2019). Lots of studies have suggested that changes in meteorological variables and hydrographic, such as salinity, cloudiness, storminess, and temperature, will affect fish life histories (e.g., recruitment, mortality rates, migration patterns, growth, and spatial distributions) (Hunter et al., 2002; Hossain et al., 2012; Pécuchet et al., 2015; Chaparro-Pedraza and De Roos, 2019). These variables can therefore impact fish population dynamics substantially; for example, populations may exhibit oscillations in abundance and/or alternative pattern in distribution due to changes in the populations of prey (Blanchot et al., 1992) or predators (Sprogis et al., 2018). If these populations are commercially exploited, the oscillating trends in abundance and distribution will, in turn, guide the appropriate and efficient management of local fisheries (e.g., updating fisheries law, registering fishing vessels, and establishing closed fishing areas and seasons). Particular examples of populations that have undergone such fluctuations are the small pelagic fish populations in the northwestern offshore of Baja California in 1997–1998, these populations were concentrated in specific areas not influenced directly by ENSO events, and researchers suggested that it is necessary to regulate the fishing during climate anomalies (Rojas-Mendez and Mendoza, 2008).
Life history refers to the pattern of survival and reproduction events during the life of fish. Life-history traits include fecundity, age-at-maturity, maximum size, and longevity (Oli and Coulson, 2016; Brown and Choe, 2019). Life history theory holds that these traits have been shaped by natural selection to optimize trade-offs related to growth, reproduction, and survival (Petrik, 2019). Moreover, life-history traits are the underlying determinants for population responses to environmental forcing (King and Mcfarlane, 2003). There is no doubt that the diversity of life-history trait combinations is due to inherent trade-offs, as more energy allocated to reproduction reduces energy available for growth or maintenance and vice versa (Waples and Audzijonyte, 2016). Since the finite amount of energy cannot fully meet the demands of reproduction, growth, and maintenance, only certain combinations of life-history traits produce an organism with high fitness (Charnov, 1993). The particular life-history traits of any given fish can be shaped by the environment. For example, the impact of global warming will not only directly influence the duration of embryonic development, egg survival, size at hatching, developmental rate, pelagic larval duration, and survival but will also alter larval behavior and impair sensory capabilities through its companion effect (ocean acidification) (Pankhurst and Munday, 2011). The age-at-maturity and energy allocated to reproduction depend on growth conditions of the juvenile (Ward et al., 2017), and this will in turn impact population recruitment and connectivity patterns in marine fishes (Pankhurst and Munday, 2011). Life-history traits are the underlying determinants of population responses to environmental forcing. Accordingly, it is important to consider life-history strategies in fisheries management.
The Beibu Gulf is located in the northwestern South China Sea (17–22°N, 105–110°E) and covers a total area of 1.3 × 105 km2 with an average depth of 50 m (Gao et al., 2017). As one of the four most prominent fishing grounds in China (Su et al., 2022), the vessel monitoring system shows that Beibu Gulf is one of the main areas where fishing vessels operate in the South China Sea (Qian et al., 2022), the rich fishery resources of this area are particularly important as a key contributor to ecosystem services. The Beibu Gulf serves as a spawning and feeding ground for many major commercial fish because of its unique geographic location and climatic conditions (Chen et al., 2009; Gao et al., 2017). In total, > 1000 fish species have been reported in the region to date, more than half of the fish species recorded in the northern South China Sea continental shelf (Liang, 2022), most of which are demersal (Khanh et al., 2013). However, the ecosystem structure has been altered considerably during the last decades of the 20th century, with overfishing leading to changes in species compositions and a decline in the biomass of most commercial species (Chen et al., 2011; Xu et al., 2011). The typical dominant species, including crimson snapper (Lutjanus malabaricus) and Pomadasys hasta, have been replaced by fishes of lower trophic levels, such as Acropoma japonicum and Argyrosomus macrocephalus (Qiu et al., 2010; Wang et al., 2013). Fortunately, the government of China has taken a series of strict management measures for the domestic fishery industry over the last 20 years, such as the implementation of a summer fishing moratorium (Shen and Heino, 2014) and the adjustment of fishery fuel subsidies for fishing vessels (Su M. et al., 2021). The mean trophic level of fish resources from bottom-trawl survey data in Beibu Gulf has slightly increased since 2000 (Su L. et al., 2021). However, increasing fishing pressure in the Beibu Gulf by Vietnam fishery vessels appears to have made these measurements less effective, annual total catch in this region is decreasing (Huang and Huang, 2013; Tian et al., 2022). In addition, ENSO events can influence marine ecosystems via global atmospheric and oceanic processes. These effects could be particularly important in a subtropical gulf, such as the Beibu Gulf, where complex circulation and strong horizontal gradients in salinity, temperature, and oxygen concentrations control the reproduction and distribution of most species. Although most studies of fishery resources in the Beibu Gulf have focused on effects of anthropogenic activities and seasonality (Wang Y. et al., 2020; Su L. et al., 2021; Zhang et al., 2022), few published studies have considered the relationship between population status and climate variability. Therefore, this study aims to fill the gap in knowledge regarding the impact of climatic variability on fish populations in the subtropical semi-enclosed Beibu Gulf.
Here, we used the electronic length frequency analysis (ELEFAN) to estimate von Bertalanffy growth function (VBGF) parameters (Pauly, 1980) for major commercial fishes in the Beibu Gulf. Trends in life-history strategies (what kind of ways that organisms allocate limited resources to growth, health maintenance, and various life processes)(Wang et al., 2021) across climate periods were evaluated. Natural mortality and growth parameters were also examined for synchronicity across species to determine the potential for climate events to generate a synchronous trend in life-history strategies. Therefore, the relationships between the Oceanic Niño Index (ONI) and life-history traits of fishes were examined to analyze the potential impact of climate events.
Fish sampling data were obtained from 34 otter trawl surveys at 52 stations (Figure 1) from 2006 to 2020. Apart from four survey cruises per year in January, April, July, and October in 2006 and 2007, surveys since 2008 were performed twice a year in January and July. Most survey cruises did not cover all 52 stations for a variety of reasons, but the samples were generally coherent in methodology. The surveys were undertaken by a commercial fishing vessel with a main engine of 600 HP. In every survey, each station was trawled once for 1 hour with a towing speed of 2.5~3.5 knots. The headrope length of the otter trawl net was 37.7 m and the cod-end mesh was 40 mm. Every trawl sample was sorted onboard and identified to the species level. Samples of each fish were counted, weighed, and recorded, and length and weight frequency data were also collected for major commercial fish. When fewer than 50 individuals were caught in a trap, all individuals were measured; otherwise, 50 individuals were randomly sampled for measurement. For each marine fish, total length (nearest cm) and body weight (nearest g) were measured. Based on the availability of data, we selected 13 species of major commercial fish (include 9 fish families, Table 1) with the highest frequencies of occurrence for analyses; according to data from the China Fishery Statistical Yearbook (2006-2020), these species account for about 45% of the total catch recorded in the Beibu Gulf.

Figure 1 Map showing stations for otter trawl surveys in the Beibu Gulf during 2006–2020. (A) the location of the Beibu Gulf in the South China Sea; (B) the sampling station (dots) in the Beibu Gulf.
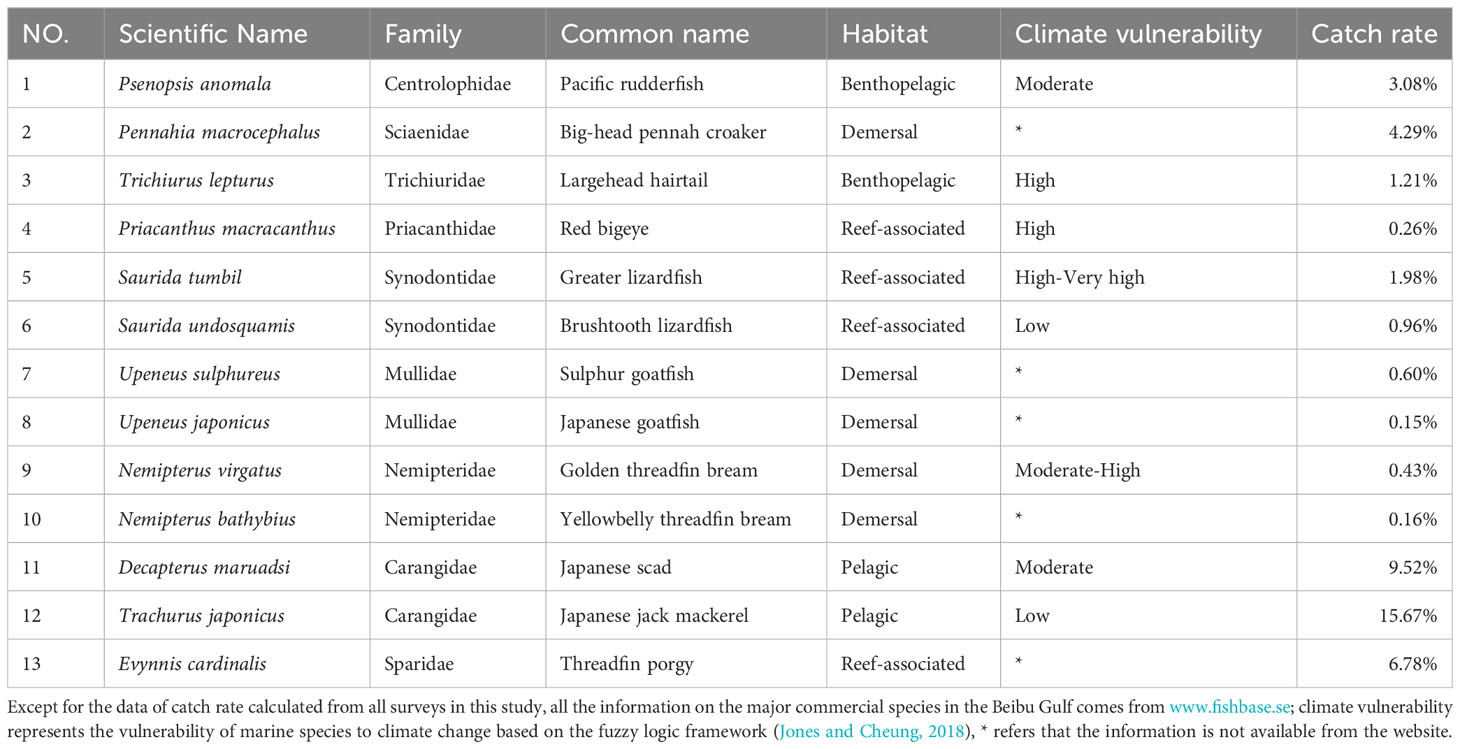
Table 1 The 13 major commercial fishes in the Beibu Gulf.
To characterize ENSO events, data for the ONI in each month from 2006 to 2020 were obtained from products of the NOAA (National Oceanic and Atmospheric Administration) Climate Prediction Center (CPC; https://origin.cpc.ncep.noaa.gov/). The ONI is defined as 3-month running mean sea surface temperature (SST) departures in the Niño 3.4 region (5°N to 5°S, 120 to 170°W) and is a principal measure for monitoring, assessing, and predicting ENSO. Climate events are defined as five consecutive overlapping 3-month periods at or above a +0.5°C anomaly for warm (El Niño) events and at or below the −0.5°C anomaly for cold (La Niña) events. The threshold is further broken down into weak (with a 0.5–0.9°C SST anomaly), moderate (1.0–1.4°C), Strong (1.5–1.9°C), and very strong (≥2.0°C) events. The CPC considers that these anomalies must also be forecasted to persist for three consecutive months. In addition, the 1/4-resolution SST data were obtained from the OI SST V2 High Resolution Dataset provided by NOAA’s Physical Science Laboratory, Boulder, Colorado, USA (https://psl.noaa.gov, accessed on 1 October 2022). Monthly mean SST data were collected at all survey stations to calculate the monthly mean SST in the Beibu Gulf from 2006 to 2020.
Length frequency data of major commercial fishes from 2006 to 2020 was pooled and converted to monthly catches with the assumption that the samples were representative of the total catch of the month (Zhang et al., 2023). We used ELEFAN in the R package “TropFishR (1.6.3)” (Mildenberger et al., 2017) to estimate the natural mortality, asymptotic length, and growth coefficient of major commercial fishes, and the optimization routine for the fitting of von Bertalanffy growth function (VBGF) parameters is the “ELEFAN_GA” function uses the Genetic Algorithm package “GA” (Scrucca, 2013; Scrucca, 2017). Growth parameters are described by the VBGF (Von Bertalanffy, 1938):
where Lt represents mean length (cm) at age t, L∞ represents asymptotic length (cm), k represents growth coefficient, and t0 is the theoretical age when Lt = 0.
To improve the precision of the growth parameters (L∞ & k) estimation, the length frequency data of major commercial fishes was binned according to the maximum body length observed for the fish species (Wang K. et al., 2020), number indicating over how many length classes the moving average should be performed:
where Lmax represents maximum body length (mm) of each fish being studied.
Natural mortality (M) was estimated by applied an updated version of the (Pauly, 1980) growth-based method (Then et al., 2015):
For optimum search and improvement in the accuracy of the output of ELEFAN_GA, we appropriately tuned key input settings (including Fix L∞ (initial seed value of L∞) and upper and lower limits of k) based on relevant literature (Taylor and Mildenberger, 2017; Mzingirwa et al., 2020; Wang K. et al., 2020) and used default TropFishR settings for all other parameters.
Based on the fundamental trade-offs between natural mortality (M) and parameters defining the maximum size and rate of growth in marine fish proposed by Gislason et al. (2010) and the empirical equation demonstrated by Charnov et al. (2013) (Equation (4), a life-history trait composite index (LTCI) was constructed to show the combinations of life-history traits, that is, life-history strategies. Given that for any combination of life-history traits, the points formed by the natural mortality (M) and growth state parameter ((L/L∞)-1.5•k, a higher value indicates how fast the fish grows) of marine fishes on the coordinate axis are basically along the y = x line (Charnov et al., 2013), the LTCI was calculated as the distance between the point projected on the y = x line of each point and the zero point (origin). The value of LTCI was calculated as shown in Equation (5):
where L∞ (cm) and k (year-1) are the VBGF parameters and L (cm) is defined as the mean body length of the fish population. The value of LTCI will be positive if the projected point is on the right-hand side of the zero point, negative if it is on the left-hand side of the zero point, and 0 if it falls on the zero point (Figure 2).
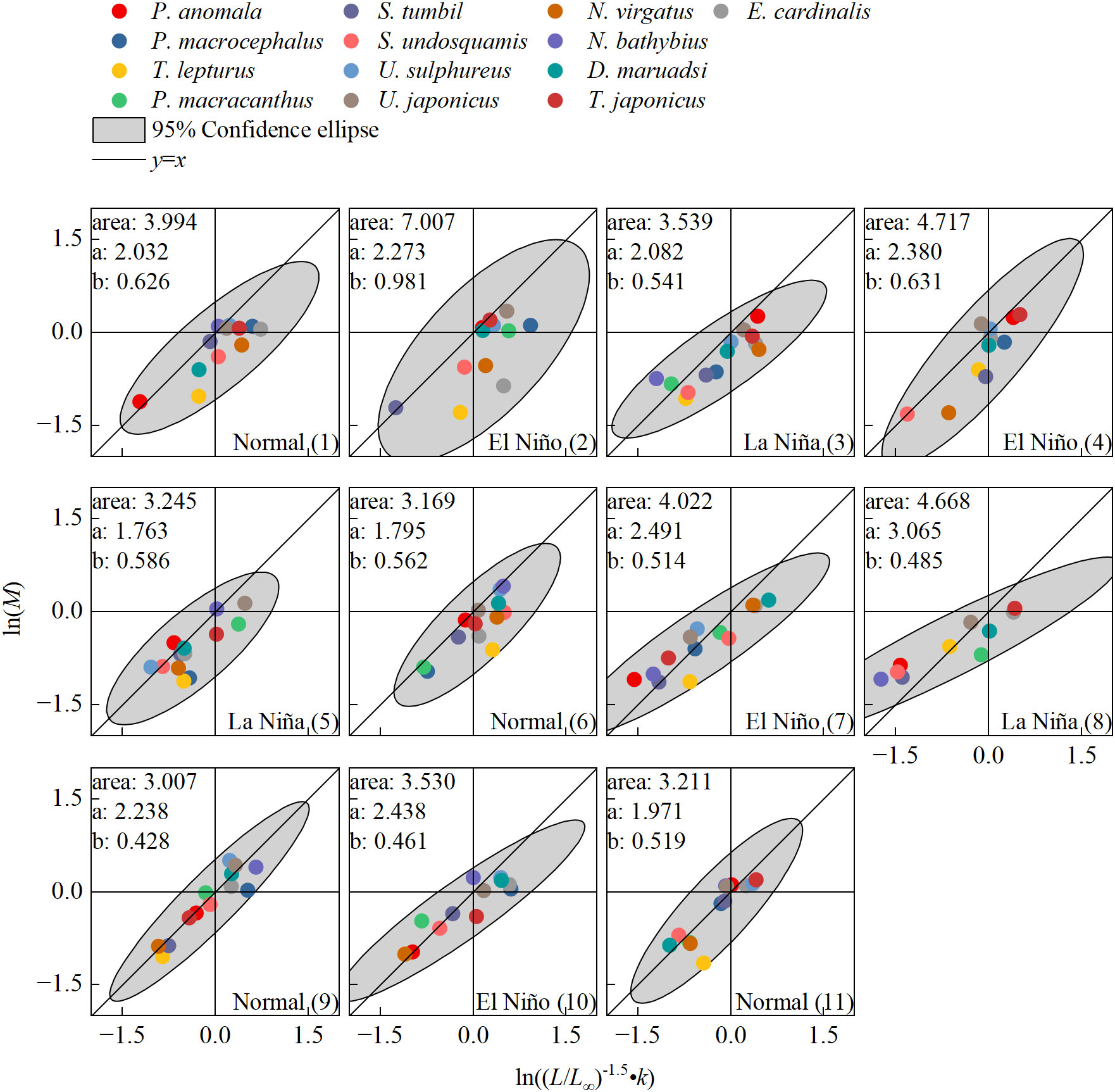
Figure 2 Log-log relationship between natural mortality (M) and growth parameters ((L/L∞)-1.5•k) for fish community in the Beibu Gulf. The numbers represent the climate periods in the same order as Table 2, and the area (gray region) represents a 95% confidence ellipse, a and b refer to the major and minor axes of the 95% confidence ellipse, respectively.
Cross-correlation function (CCF) analysis was used to explore the hysteretic relationship between climate event intensity and SST in time series, for removing spurious correlations from the affected time series (Baumgartner et al., 2018), this study performed prewhitened CCF analysis using the “prewhiten” function from the R package “TSA”. To investigate responses of fish populations to climate events, linear models were used to identify the relationships between the ONI and parameters of fish life-history traits. Spearman correlation coefficients were used to test the significance of these relationships (p < 0.05 was considered significant). Multivariate analysis of variance (MANOVA) and one-way analysis of variance (ANOVA) were employed to compare fish life-history trait parameters among climate periods. SPSS 27.0 (SPSS Inc.) was used to perform these statistical analyses.
Furthermore, the effects of climate events on the life history of the fish community were analyzed by using a hierarchical linear model and moderating effect estimation, and the R package “broom. mixed” and R package “bruceR” were used respectively. All R packages were operated in the R Environment (R Core Team, 2022).
In 2006−2020, 11 climate periods were identified according to the afore-mentioned criteria for the classification of climate events, of which four were normal climate periods, four were El Niño periods, and four were La Niña periods (Table 2).
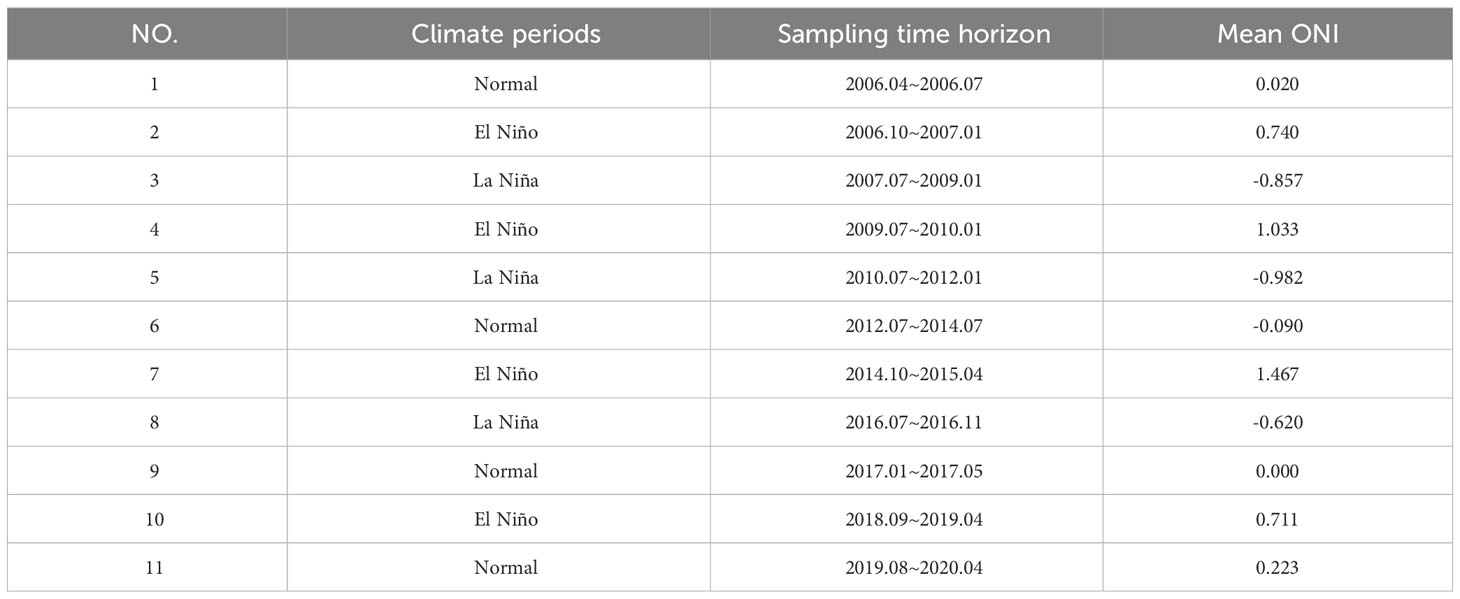
Table 2 Summary statistics of fish community in each climate period from 2006 to 2020.
The hierarchical linear model results indicated that the relationship between the growth status parameter (ln((L/L∞)-1.5•k)) and natural mortality (ln(M)) of fish community differ significantly by climate event type; the normal climate period regression slope (pn = 0.772) was closest to 1.0, and it also was higher than the regression slopes of El Niño period (pe=0.669) and La Niña period (pl=0.593) (Figure 3A) The mean difference estimation also showed that climate events had an impact on LTCI in fish communities, and the mean diffidence of LTIC La Niña events also was the largest. In addition, regression analysis of the life history trait parameters of fish communities for each climate period was also shown the mean area of the 95% confidence ellipse (3.345 ± 0.382) was lowest in the normal period (Figure 2), followed by the La Niña period (3.817 ± 0.613), and was highest in the El Niño period (4.819 ± 1.332). The mean major axis of the ellipse (a) was also lowest in the normal period (2.009 ± 0.158) and larger in the La Niña (2.303 ± 0.554) and El Niño periods (2.396 ± 0.081), while the mean minor axes of the ellipse (b) in the normal (0.534 ± 0.072) and La Niña periods (0.537 ± 0.041) were similar, with the highest value in the El Niño period (0.647 ± 0.203). Overall, the results of MANOVA showed that the growth state parameter, natural mortality, and LTCI of fish in different climatic event periods showed significant differences (p < 0.05).
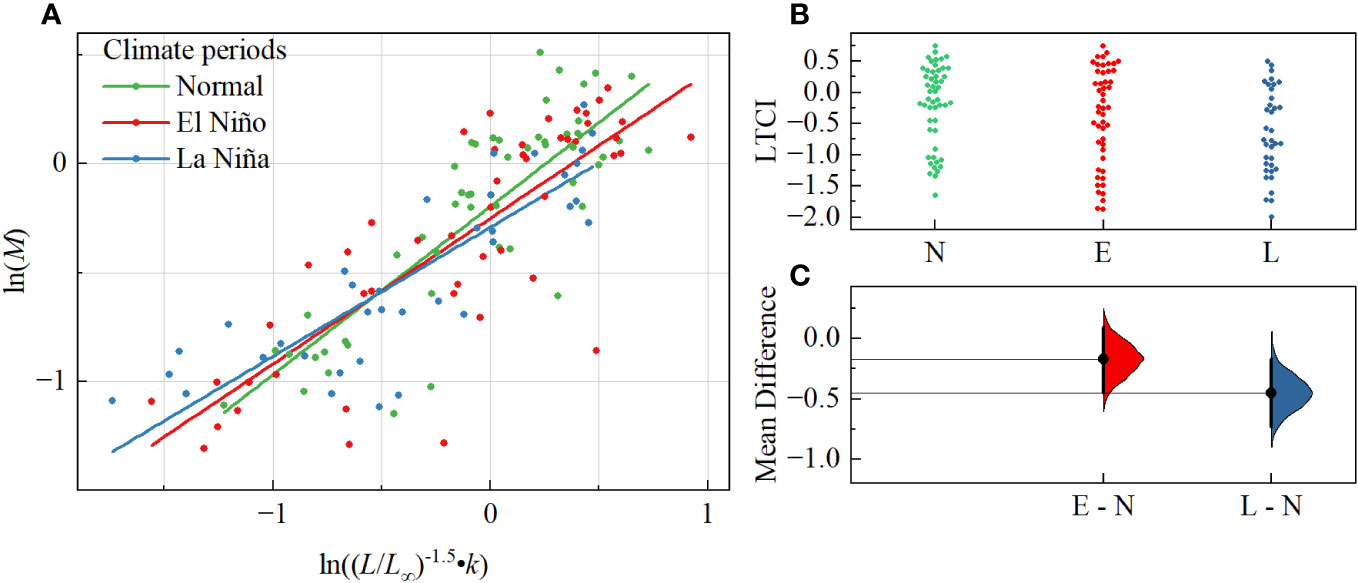
Figure 3 (A) Linear regressions of natural mortality and growth status parameter at the three climate periods since 2006-2020 within the Beibu Gulf; (B, C) Mean differences in LTCI of fish communities during climate event periods versus normal climate periods, N, E and L represent normal, El Niño and La Niña climate, respectively.
The monthly mean SST in the Beibu Gulf (Figure 4A) presented significant and positive associations with ONI during 2006–2020 (r = 0.521). Considering the time with higher correlation values, there was a high correlation between the monthly mean SST in the Beibu Gulf and ONI without any lag (Figure 4B). In addition, although the effects of monthly mean SST on LTCI of fish communities were no significantly different in El Niño climate periods compared with normal and La Niña climate periods based on the results of moderate effect estimation (p = 0.07), the LTCI of fish communities strongly increased with monthly mean SST, while the LTCI of fish communities as a whole was lower than normal climate periods during La Niña climate periods (Figure 5).
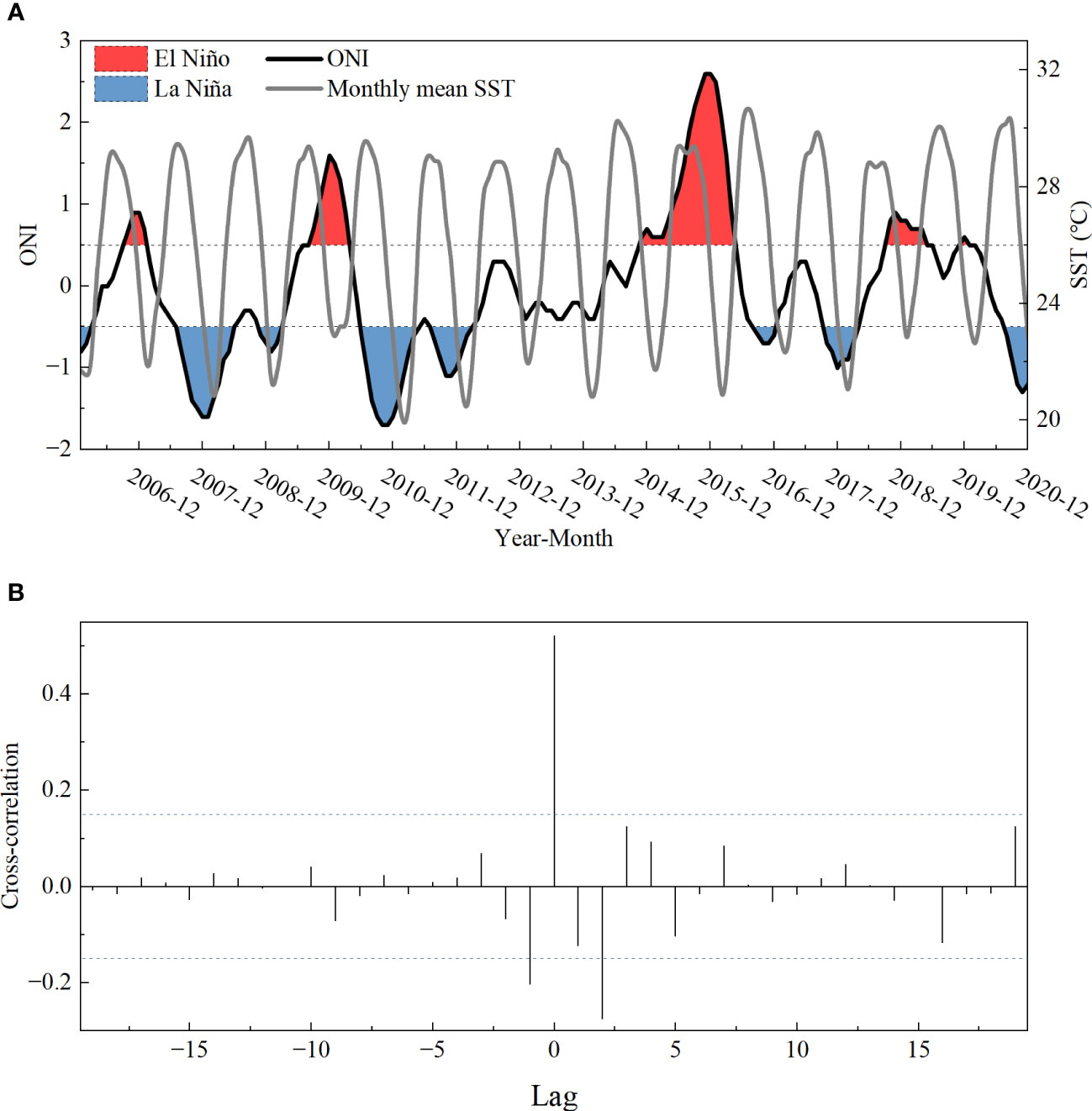
Figure 4 (A) Time-series of monthly mean SST and (ONI). (B) Cross- correlation functions (CCFs) between ONI and monthly mean SST; dashed lines indicate the significance threshold (α = 0.15).
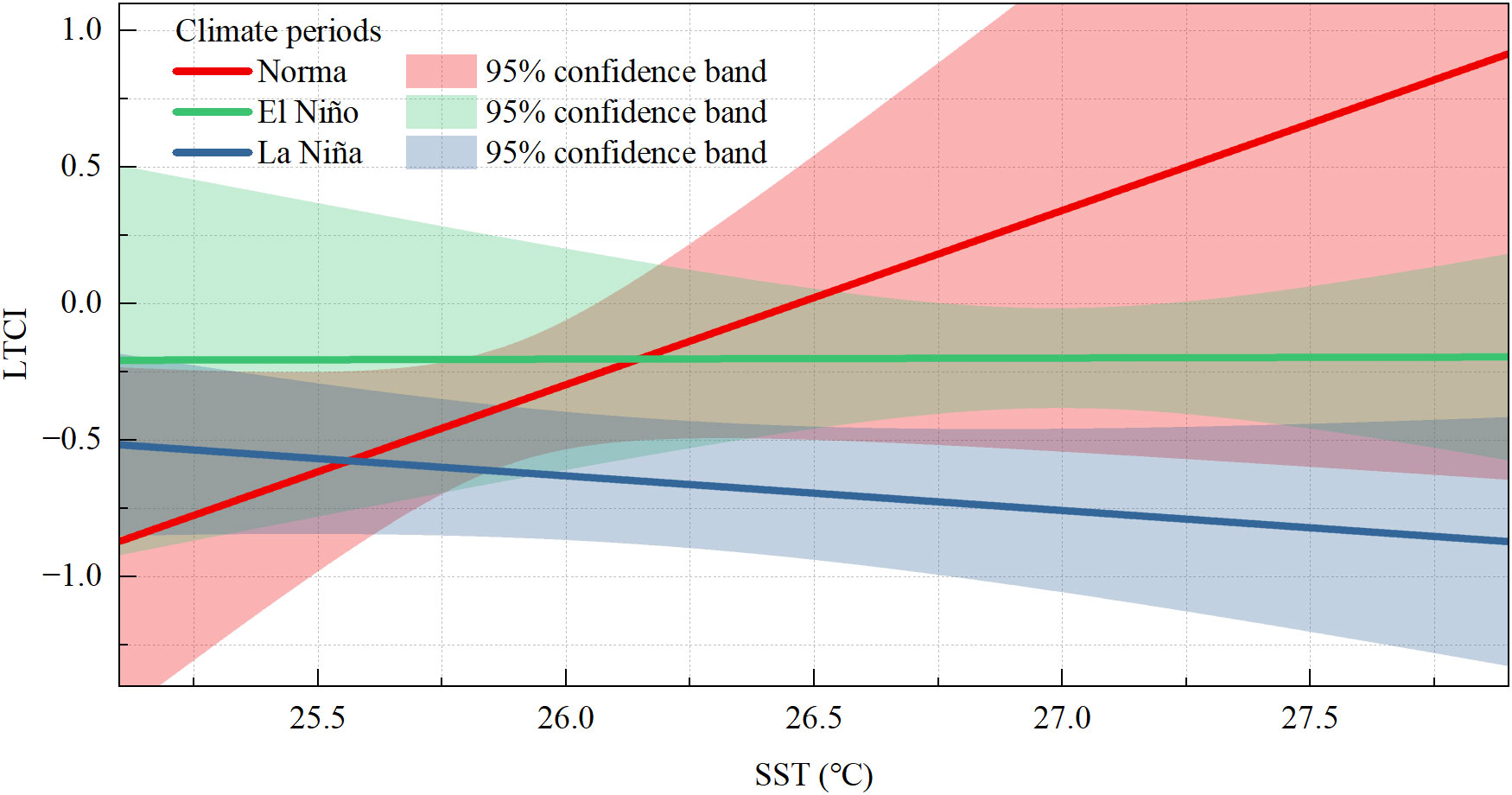
Figure 5 Linear regressions of LTCI and monthly mean SST at the three climate periods since 2006-2020 within the Beibu Gulf.
Except for T. lepturus and E. cardinalis, the natural mortality values and growth parameters of other fish showed a significant positive correlation (Figure 6). Some fish exhibited relatively stable life histories between climate periods; for example, the life-history trait parameters of T. lepturus clustered in the lower-left area of the coordinate plane. In addition, one-way ANOVA showed that these fish life-history trait parameters did not differ significantly among climate periods (p > 0.05), whereas life-history trait composite index (LTCI) of P. macrocephalus and D. maruadsi differed significantly during the transition of climate periods (F = 38.986 and F = 14.921, respectively, all p < 0.05). Except for N. bathybius and P. anomala, all fishes did not display any considerable oscillation in LTCI with the transformation of climate periods (Figure 7).
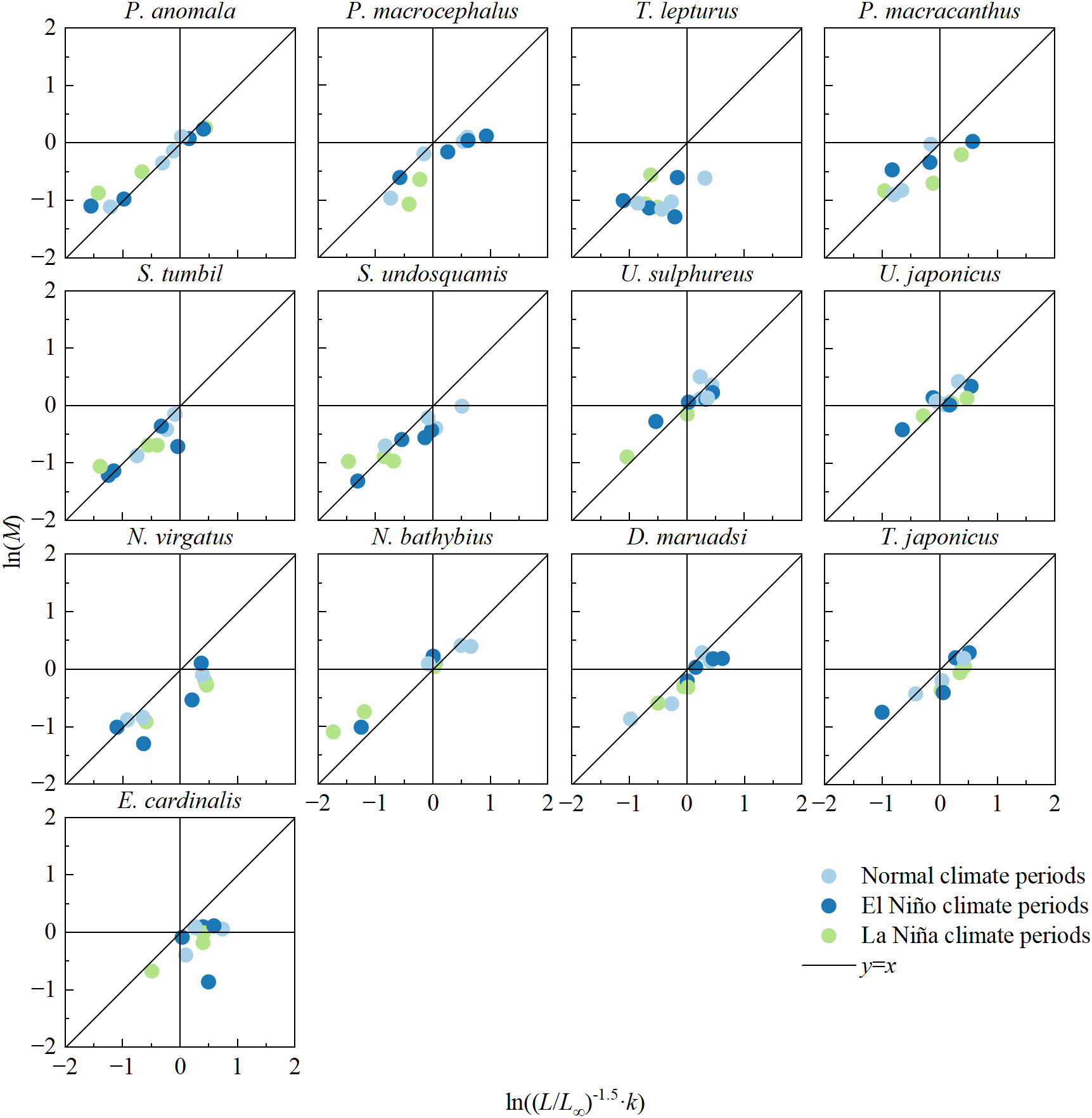
Figure 6 Log-log relationship between natural mortality (M) and growth parameters for 13 major commercial fishes in the Beibu Gulf.
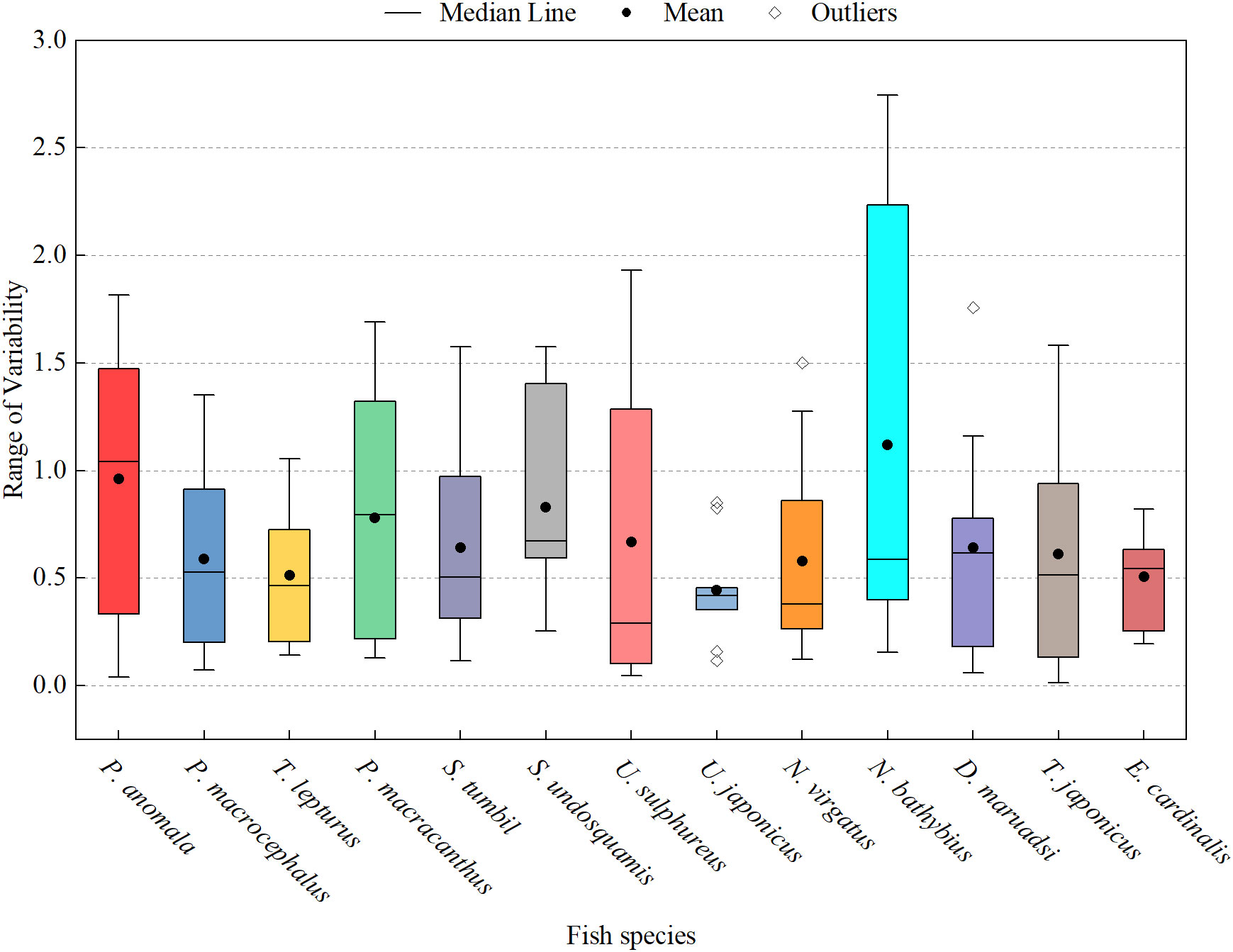
Figure 7 Boxplot of LTCI for 13 major commercial fishes. Each box represents fluctuation of LTCI of a fish between the climate periods; median value of LTCI is indicated by a filled point.
A Spearman correlation analysis indicated that the LTCI values for D. maruadsi, P. macrocephalus, and U. sulphureus were significantly correlated with ONI in 2006–2020 (r = 0.697, 0.762, and 0.905, respectively, all p < 0.05). The most consistent changes in the LTCI of the three fish species occurred in 2006–2012, when transitions between warm and cold events was most frequent (Figure 8). Furthermore, in the latter six climate periods, there was a negative correlation between the LTCI of T. japonicus and D. maruadsi. While the relationships were not statistically significant, the LTCI of fishes belonging to the same genus showed the opposite trends in some climate periods, including U. sulphureus and U. japonicus, S. tumbil and S. undosquamis, and N. virgatus and N. bathybius.
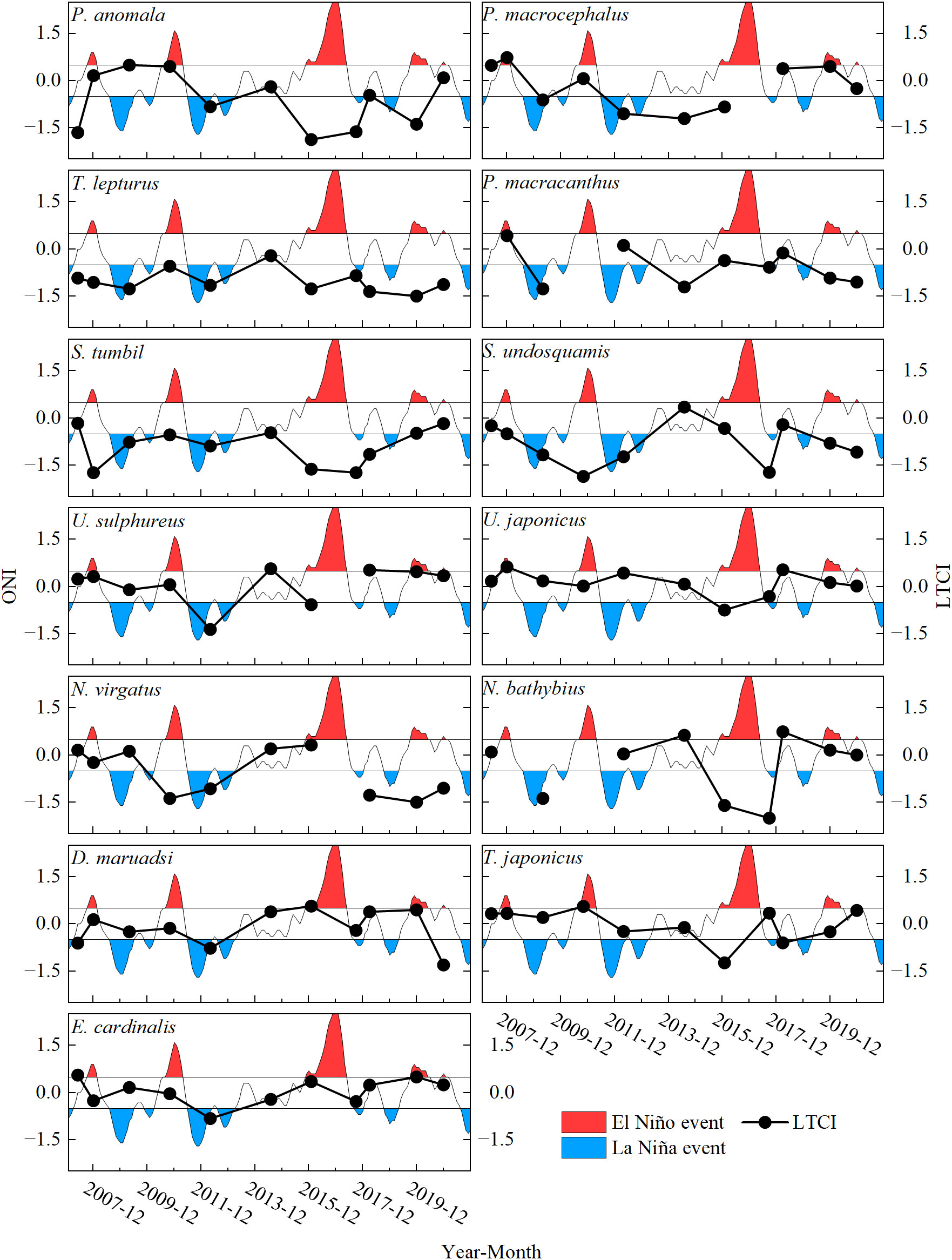
Figure 8 Relationship between the LTCI of major commercial fishes and climate events in the Beibu Gulf.
According to Table 1, the vast majority of the 13 major commercial fishes in the Beibu Gulf that we selected were low vulnerable to fishing, but nearly half were recorded as having moderate to high vulnerability to climate. There is no doubt that it is interesting to analyze the correlation between the parameters of the life-history traits of these fishes and climatic events. By examining patterns of responses across 13 major commercial fishes in the Beibu Gulf, we found evidence that fluctuations in life-history traits and the abundance of some small fishes are best explained by changes in environmental variables caused by ENSO events. Frequent transformations of climate events were correlated with periodic fluctuation in LTCI, while the sensitivity of fishes to various environmental variables explained variation in abundance. However, our results also expand upon theoretical and regional results, highlighting the particularly high sensitivity of some fishes in the subtropical gulf to climate events.
Despite a large and growing body of evidence for the role of anthropogenic activities in the population dynamics of marine species (Pinsky and Palumbi, 2014; Sherman et al., 2017; García-March et al., 2020; Strøm et al., 2021), including dramatic fluctuations of pelagic and demersal fishes in the South China Sea over space and time scales (Masrikat, 2012; Yang et al., 2021; Zhang et al., 2022), our results suggest that the combination of life-history strategies for the Beibu Gulf fish community were most strongly driven by climate events. Climate events significantly altered the slope of the regression of life-history trait parameters for fish communities, with slopes closest to 1.0 in the normal period (Figure 3). However, our results also revealed that both climate events would lead to a decrease in the mean correlation coefficient, especially La Niña periods, which can be explained by the fact that the combinations of life-history traits for most fish species fall in the lower-left quadrant (slower growth and lower M). Although the mean correlation coefficient of the El Niño period was between those for the normal and La Niña periods, the larger mean area of the confidence ellipse for the combinations of life-history traits in the El Niño period differed from those of other periods, and the former period tends to have a higher diversity of fish life-history strategies. While other studies have found that this climate variability may influence the biomass of certain fish species substantially via environmental variables (Yatsu et al., 2008; Yen and Lu, 2016), our results highlight the regular dynamics of life-history strategy selection in fish communities.
An important and relatively underappreciated mechanism underlying this dynamic may be related to ocean-atmosphere coupling. Temperature in all regions has pervasive effects on biological processes. Even though the mean SST in Beibu Gulf is on the rise under the background of global warming, this study found a positive correlation between the monthly mean SST in Beibu Gulf and ONI without any lag. This response to water temperature anomaly in the equatorial Pacific is bound to affect fish growth and development (Gillanders et al., 2012). In general, species characterized by fast growth and short generation times (high M values) would respond quickly to environmental changes (King and Mcfarlane, 2006), and the increase in seawater temperature caused by warm events may accelerate their growth and maturation. From the results of this study analysis, however, the relationship between LTCI of fish community and SST in the Beibu Gulf has a certain correlation with the type of climate events. On the one hand, since Beibu Gulf was a subtropical bay and its fishes were mainly warm-water species (Liang, 2022), temperate fishes tend to have smaller heating tolerances than tropical fishes (Payne et al., 2021). Therefore, an increase in environmental temperature would reduce the physiological function of fish and then affect the fish’s life history (Dembski et al., 2006; Holt and Jorgensen, 2015), especially during the El Niño climate periods when the SST is significantly warmer, and this study observed a significant increase in LTCI in fish with increasing SST. On the other hand, fishes tend to choose more “conservative” life-history strategy (slower growth and lower M) selection because of the lack of suitable water temperature in the environment (Soyano and Mushirobira, 2018). This was also evident in the event of cold events, where significantly lower seawater temperatures undoubtedly result in a lower overall LTCI of fish communities compared to normal climate periods. Life-history traits of signal species evolve based on environmental conditions to maximize sustainable reproduction; however, changes in the life-history strategy of fast-growing species, as the most important component of the ecosystem (Waples and Audzijonyte, 2016), will inevitably affect their ecological functions. In addition, during El Niño periods, the South China Sea generally exhibits weakened circulatory gyres and warming of the SST, consistent with the weakening of East Asian winter monsoon (Qiu et al., 2015). Previous studies have suggested that warmer temperatures increase metabolic rates and therefore increase the energy demand of fishes, while simultaneously decreasing the quality and quantity of available prey (Overland et al., 2010). As a result, combinations of life-history traits of fish within communities have become more diverse, even if some combinations have deviated significantly from evolutionarily stable strategies. This mechanism may explain our finding that life-history strategies for the fish community in Beibu Gulf were sensitive to climate events. While marine species fluctuate strongly, even in the presence of fishing (Adams, 1980; Hu et al., 2015), the strong interaction we found between climate events and life-history traits suggested that the composition and structure of the fish community in the subtropical semi-enclosed bay were also primarily caused by climate teleconnection.
The life-history traits of most fish species were optimized across all periods, i.e., variation in mortality coincided with variation in growth parameters; however, the variation in life-history traits of T. lepturus and E. cardinalis differed significantly from those of other species. Ribbonfish (T. lepturus) is a benthonic-pelagic species found worldwide in tropical and subtropical regions, mainly between 45°S and 60°N (Taghavi Motlagh et al., 2021). While T. lepturus is relatively short-lived and fast-growing (Clain et al., 2023) compared to larger fish in the marine ecosystem, they had the lowest mean mortality rate among the fish analyzed in this study and the distribution of M values was very similar at all periods, suggesting that their life-history traits could mainly be described as “longevity-slow.” Although we were unable to find significant differences between growth parameters among climatic periods, relatively sharp variation in the growth of ribbonfish was clearly influenced by climatic events. Past research has suggested that T. lepturus possesses flexible reproduction strategies; in environments with marked temperature cycles or weak fishing pressure, populations are more likely to reach maturity after longer durations (Al-Nahdi et al., 2009). However, in warmer and less productive environments, lower batch fecundity may be compensated for, in part, by a prolonged spawning period due to a smaller size (Martins and Haimovici, 2000). The increase in growth parameters in the case of relatively fixed mortality will promote a shift in the combination of life-history traits to the fourth quadrant. Therefore, T. lepturus would expend a great deal of energy to mature and begin reproduction early, retaining a small size and not capitalizing on the benefits of increased fecundity that come with a large size and long-life cycle (Waples and Audzijonyte, 2016). However, the reproduction strategy of a multiple batch spawner with a prolonged spawning period seems to allow for such a life-history strategy to exist.
We also observed that the life-history strategy of E. cardinalis was relatively constant in most climate periods. E. cardinalis is a small, warm-water, near-demersal fish with a fast rate of sexual maturity and a high reproductive rate (Zhang et al., 2020). The species exhibits significant periodic fluctuations in stock density (Sun and Lin, 2004) and can withstand a high fishing intensity (Chen, 2005). While E. cardinalis is one of the most economically important species in the northern South China Sea, the substantial increase in catch due to overfishing is a major threat (Zhang et al., 2020; Xu et al., 2021), and the species was even listed as endangered in a recent International Union for Conservation of Nature (IUCN) red list (Xu et al., 2022). Having a relatively unique niche in the ecosystem may be the main reason for the relatively stable life-history strategy selections of these fishes (Olden et al., 2006), with less interspecific competitive pressures making them more resistant to disturbances (Lewis, 2022). We also suggested that the relatively fixed combination of life-history traits may be a response of the population to overfishing; this stable life-history strategy would reduce the negative impacts of heavy exploitation on survival, and the effects of climate events may not be dominant.
Of note, P. anomala and N. bathybius showed substantial differences in LTCI among climate periods. Given that fishing may lead to populations with an elevated sensitivity to climate variability, interaction effects between fishing and climate cannot be ignored. Several recent studies have shown that the fishery resources in the Beibu Gulf have been overfished (Su L. et al., 2021; Zhang et al., 2021; Su et al., 2022), and loopholes in local management measures and the increased fishing pressure from Vietnam have resulted in the annual decreases of fishery resources in the region (Huang and Huang, 2013; Tian et al., 2022). This not only results in the replacement of dominant species with less valuable, small-sized, low trophic level species (Zhang et al., 2020) but also increases the magnitude of population fluctuations driven by environmental variability (Hsieh et al., 2006; Planque et al., 2010; Hidalgo et al., 2011; Rouyer et al., 2011; Shelton and Mangel, 2011). Therefore, synergistic interactions between climate anomalies and fishing are realistic (Waples and Audzijonyte, 2016). Fish whose life-history strategies are significantly affected by climate events are more frequent in overexploited areas.
Among the commercially valuable fish species included in this study, only the LTCI values for D. maruadsi, P. macrocephalus, and U. sulphureus were significantly correlated with the fluctuations in ONI, which represents climate period transformations. Given the delayed response trait of the fish population to environmental changes, it is reasonable to estimate fish life-history traits separately by the types of climate periods over a longer time scale in this study. The life-history traits of these fishes are characterized by higher M and faster growth during the normal or El Niño periods. The LTCI for D. maruadsi generally decreased after the shift from El Niño to La Niña or normal conditions. Climate events will affect a range of abiotic factors that are tightly linked to the production and distribution of fish populations. Shallow areas will exhibit larger increases in sea temperatures than those in deeper waters. As warm-water fishes, D. maruadsi, P. macrocephalus, and U. sulphureus will grow faster with increased reproduction due to seawater warming, which expands aerobic scope initially (Holt and Jorgensen, 2015), and the increase in metabolic requirements intensify foraging and reduce survival. However, under higher or lower mortality, evolution toward a faster or slower growth, respectively, optimizes their combination of life-history traits to ensure the stability of population dynamics. High natural mortality is essential if fish growth is fast, as the stability of the ecosystem only limits population size within the carrying capacity. Furthermore, La Niña events also strongly affect the surface and subsurface patterns of the South China Sea through the anomalous cyclonic circulation in the low-level atmosphere (Kuo and Tseng, 2020). Like the west coast of the Atlantic (Mcphaden et al., 2006), tropical cyclones tend to be reduced in number and intensity during El Niño periods but stronger and more numerous during La Niña periods (Huang and Xu, 2010; Hsu et al., 2013; Chen and Duan, 2018). The frequent occurrence of weather phenomena reduces anthropogenic disturbances and the lower SST during La Niña periods (Qiu et al., 2015) results in a shift to increased growth over rapid reproduction.
We also found that changes in life-history strategies can differ among species within the same genus in response to climate events. In particular, for U. sulphureus and U. japonicus, S. undosquamis and S. tumbil, and N. virgatus and N. bathybius, the LTCI values of two species in the same genus were generally similar in normal periods, with divergence in opposite directions in response to climate events. These species of the same genus not only share very similar biological parameters but also have similar population characteristics, such as diet, habitat preference, and spawning period, suggesting that their ecological niche is basically the same (Lewis, 2022). However, the oceanographic features that change following ENSO events include surface seawater temperatures, the vertical and thermal structure of the ocean (especially in coastal regions) (Wu et al., 2014), and the coastal and upwelling currents (Chu et al., 2017; Dang et al., 2021). These effects will undoubtedly have a significant negative impact on the growth and fecundity of fish and consequently intensify competition for survival between species with similar ecological niches. Thus, species of the same genus, through the complementarity of life-history strategies, avoid intense competition for ecological niches during climate anomalies because the carrying capacity of the ecosystem can be increased by reducing the similarity in life stages.
Finally, T. japonicus and D. maruadsi are generally considered to belong to the same feeding group at the ecosystem level and are both dominant species in offshore areas of China (Li et al., 2019; Su et al., 2022); accordingly, they have very similar ecological niches in long-term cohabitation. While the correlation was not statistically significant, we observed similar trends in LTCI for the two species over the first six climate periods in this study. Combined with the observation that there was a negative correlation between the LTCI value of T. japonicus and D. maruadsi in the latter six climate periods, we can conclude that the regularly alternating climate periods may explain the consistency of direction of change in life history strategies. The continued decline of fishery resources and increasing fishing pressure in the Beibu Gulf may also be responsible for the complementarity of life-history strategies between D. maruadsi and T. japonicus in the latter six climate periods. Overall, given the different relationships between life-history strategies and study periods, we emphasize the crucial importance of analyses of the effect of climate events on fish life history; overfishing weakens the resilience of fish populations to climate events, while interactions within communities also affect the response of fish life-history strategies to climate events.
There are some limitations in this study. The ELEFAN assumes that the body size composition of the survey data is representative of the exploited population major commercial fishes, and the study assumes that these survey data can be adequately analyzed in the corresponding climate event periods. However, the time span of each period of climate event is not uniform, and the composition of seasons varies from period to period. The energy storage of fish to adapt to seasonal environments is also reflected in changes in their life history (Giacomini and Shuter, 2013), which may affect our judgment of the influence of climate events on fish life-history trait parameters. For instance, although the effects of different types of climate events on the life history trait parameters of fish populations can be observed at the community level, these changes were relatively difficult to notice at the individual species level. Furthermore, the ONI has become the de-facto standard that NOAA uses for classifying warm and cool events in the eastern tropical Pacific. Although we have demonstrated a correlation between temperature changes in the Beibu Gulf and anomalies in the distant eastern equatorial Pacific, considering that ENSO’s teleconnections to other regions have impacts beyond just affecting water temperature [e.g., rainfall, seawater salinity (Zhu et al., 2014; Ramos et al., 2019)], so we did not directly use the local sea surface temperature anomalies as a direct factor in this study. In conclusion, our research results have demonstrated a certain correlation between the changes in life-history traits of major commercial fishes in the Beibu Gulf and ENSO events, and identifying specific influencing environmental factors will be the focus of our further research. Therefore, in future studies on the relationship between climate change and fisheries in the Beibu Gulf, more factors will be taken into account when assessing the effects of climate events on the life-history trait parameters of fish, such as the temporal continuity of survey data.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The requirement of ethical approval was waived by The Committee on Laboratory Animal Welfare and Ethics of South China Sea Fisheries Research Institute for the studies involving animals because the study area is in open water where is not a protected area and no endangered or protected marine species were involved. The studies were conducted in accordance with the local legislation and institutional requirements.
XH conceived the study and wrote the first manuscript. KZ, JL, YX, MS, and JJ performed the data analyses and prepared the graphs. SX, YC, YQ, and ZC provided the original length data and revised the manuscript. All the authors contributed to the article and approved the submitted version.
This work was supported by Key research and development project of Guangdong Province (2020B1111030001), the Central Public-interest Scientific Institution Basal Research Fund, Chinese Academy of Fishery Sciences (2023TD05), the Central Public-interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS (2021SD01).
We thank all those involved in the marine surveys that collected the data used in this study. We appreciate the valuable comments made by reviewers, which significantly improved our manuscript. We thank Dr. Gang Zhang from MogoEdit (http://en.mogoedit.com/), for editing a draft of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adams P. B. (1980). Life history patterns in marine fishes and their consequences for fisheries management. Fishery Bull. 78, 1–12.
Al-Nahdi A., Al-Marzouqi A., Al-Rasadi E., Groeneveld J. (2009). The size composition, reproductive biology, age and growth of largehead cutlassfish Trichiurus lepturus Linnaeus from the Arabian Sea coast of Oman. Indian J. Fisheries 56, 73–79.
Baumgartner M. T., De Oliveira A. G., Agostinho A. A., Gomes L. C. (2018). Fish functional diversity responses following flood pulses in the upper Paraná River floodplain. Ecol. Freshw. Fish 27, 910–919. (Amsterdam: Academic Press) doi: 10.1111/eff.12402
Blanchot J., Rodier M., Le Bouteiller A. (1992). Effect of El Niño Southern Oscillation events on the distribution and abundance of phytoplankton in the Western Pacific Tropical Ocean along 165°E. J. Plankton Res. 14, 137–156. doi: 10.1093/plankt/14.1.137
Brown J. L., Choe J. C. (2019). “Behavioral ecology and sociobiology,” in Encyclopedia of animal behavior, (Amsterdam: Academic Press)103–108.
Chaparro-Pedraza P. C., De Roos A. M. (2019). Environmental change effects on life-history traits and population dynamics of anadromous fishes. J. Anim. Ecol. 88, 1178–1190. doi: 10.1111/1365-2656.13010
Charnov E. L. (1993). Life history invariants: some explorations of symmetry in evolutionary ecology. (Oxford: Oxford University Press).
Charnov E. L., Gislason H., Pope J. G. (2013). Evolutionary assembly rules for fish life histories. Fish Fisheries 14, 213–224. doi: 10.1111/j.1467-2979.2012.00467.x
Chen Z. (2005). Ecological distribution of Parargyrops edita Tanaka in the Beibu Gulf. Mar. Fish. Res. 26, 16–21.
Chen Y., Duan Z. (2018). Impact of ENSO on typhoon wind hazard in the coast of southeast China. Natural Hazards 92, 1717–1731. doi: 10.1007/s11069-018-3283-2
Chen Z., Qiu Y., Xu S. (2011). Changes in trophic flows and ecosystem properties of the Beibu Gulf ecosystem before and after the collapse of fish stocks. Ocean Coast. Manage. 54, 601–611. doi: 10.1016/j.ocecoaman.2011.06.003
Chen Z., Xu S., Qiu Y., Lin Z., Jia X. (2009). Modeling the effects of fishery management and marine protected areas on the Beibu Gulf using spatial ecosystem simulation. Fisheries Res. 100, 222–229. doi: 10.1016/j.fishres.2009.08.001
Chu X., Dong C., Qi Y. (2017). The influence of ENSO on an oceanic eddy pair in the South China Sea. J. Geophysical Research: Oceans 122, 1643–1652. doi: 10.1002/2016JC012642
Clain C., Stewart J., Fowler A., Diamond S. (2023). Reproductive biology of largehead hairtail (Trichiurus lepturus) in south-eastern Australia. Aquaculture Fisheries 8, 148–158. doi: 10.1016/j.aaf.2021.09.008
Dang X., Bai Y., Gong F., Chen X., Zhu Q., Huang H., et al. (2021). Different responses of phytoplankton to the ENSO in two upwelling systems of the South China Sea. Estuaries Coasts 45, 485–500. doi: 10.1007/s12237-021-00987-2
Dembski S., Masson G., Monnier D., Wagner P., Pihan J. C. (2006). Consequences of elevated temperatures on life-history traits of an introduced fish, pumpkinseed Lepomis gibbosus. J. Fish Biol. 69, 331–346. doi: 10.1111/j.1095-8649.2006.01087.x
Gao J., Wu G., Ya H. (2017). Review of the circulation in the beibu gulf, South China Sea. Continental Shelf Res. 138, 106–119. doi: 10.1016/j.csr.2017.02.009
García-March J. R., Tena J., Henandis S., Vázquez-Luis M., López D., Téllez C., et al. (2020). Can we save a marine species affected by a highly infective, highly lethal, waterborne disease from extinction? Biol. Conserv. 243, 108498. doi: 10.1016/j.biocon.2020.108498
Giacomini H. C., Shuter B. J. (2013). Adaptive responses of energy storage and fish life histories to climatic gradients. J. Theor. Biol. 339, 100–111. doi: 10.1016/j.jtbi.2013.08.020
Gillanders B. M., Black B. A., Meekan M. G., Morrison M. A. (2012). Climatic effects on the growth of a temperate reef fish from the Southern Hemisphere: a biochronological approach. Mar. Biol. 159, 1327–1333. doi: 10.1007/s00227-012-1913-x
Gislason H., Daan N., Rice J. C., Pope J. G. (2010). Size, growth, temperature and the natural mortality of marine fish. Fish Fisheries 11, 149–158. doi: 10.1111/j.1467-2979.2009.00350.x
Gomez F. A., Lee S. K., Hernandez F. J. Jr., Chiaverano L. M., Muller-Karger F. E., Liu Y., et al. (2019). ENSO-induced co-variability of salinity, plankton biomass and coastal currents in the Northern Gulf of Mexico. Sci. Rep. 9, 178. doi: 10.1038/s41598-018-36655-y
Hidalgo M., Rouyer T., Molinero J. C., Massutí E., Moranta J., Guijarro B., et al. (2011). Synergistic effects of fishing-induced demographic changes and climate variation on fish population dynamics. Mar. Ecol. Prog. Ser. 426, 1–12. doi: 10.3354/meps09077
Holbrook N. J., Claar D. C., Hobday A. J., Mcinnes K. L., Oliver E. C. J., Gupta A. S., et al. (2020). “ENSO-driven ocean extremes and their ecosystem impacts,” in El Niño Southern oscillation in a changing climate. (American Geophysical Union, Hoboken, NJ, Washington, D.C.: John Wiley & Sons, Inc.). 409–428.
Holt R. E., Jorgensen C. (2015). Climate change in fish: effects of respiratory constraints on optimal life history and behaviour. Biol. Lett. 11, 20141032. doi: 10.1098/rsbl.2014.1032
Hossain M. S., Gopal Das N., Sarker S., Rahaman M. Z. (2012). Fish diversity and habitat relationship with environmental variables at Meghna river estuary, Bangladesh. Egyptian J. Aquat. Res. 38, 213–226. doi: 10.1016/j.ejar.2012.12.006
Hsieh C.-H., Reiss C. S., Hunter J. R., Beddington J. R., May R. M., Sugihara G. (2006). Fishing elevates variability in the abundance of exploited species. Nature 443, 859–862. doi: 10.1038/nature05232
Hsu P.-C., Ho C.-R., Liang S.-J., Kuo N.-J. (2013). Impacts of two types of El Niño and La Niña events on typhoon activity. Adv. Meteorology 2013, 1–8. doi: 10.1155/2013/632470
Hu W., Ye G., Lu Z., Du J., Chen M., Chou L. M., et al. (2015). Study on fish life history traits and variation in the Taiwan Strait and its adjacent waters. Acta Oceanologica Sin. 34, 45–54. doi: 10.1007/s13131-015-0625-8
Huang Y., Huang M. (2013). Implementation of the Sino-Vietnamese fishery agreement: mainly Chinese perspective. Beijing Law Rev. 04, 103–119. doi: 10.4236/blr.2013.43014
Huang F., Xu S. (2010). Super typhoon activity over the western North Pacific and its relationship with ENSO. J. Ocean Univ. China 9, 123–128. doi: 10.1007/s11802-010-0123-8
Hunter E., Aldridge J. N., Metcalfe J. D., Arnold G. P. (2002). Geolocation of free-ranging fish on the European continental shelf as determined from environmental variables. Mar. Biol. 142, 601–609. doi: 10.1007/s00227-002-0984-5
Jones M. C., Cheung W. W. L. (2018). Using fuzzy logic to determine the vulnerability of marine species to climate change. Glob Chang Biol. 24, e719–e731. doi: 10.1111/gcb.13869
Khanh N., Phu T., Luong N., Opoku-Acheampong A. (2013). Appropriate fishing depths for squid longline fishery in the Gulf of Tonkin, Vietnam. Fish People 11, 29–32.
King J. R., Mcfarlane G. A. (2003). Marine fish life history strategies: applications to fishery management. Fisheries Manage. Ecol. 10, 249–264. doi: 10.1046/j.1365-2400.2003.00359.x
King J. R., Mcfarlane G. A. (2006). A framework for incorporating climate regime shifts into the management of marine resources. Fisheries Manage. Ecol. 13, 93–102. doi: 10.1111/j.1365-2400.2006.00480.x
Kuo Y. C., Tseng Y. H. (2020). Impact of enso on the south china sea during enso decaying winter–spring modeled by a regional coupled model (a new mesoscale perspective). Ocean Modelling 152, 101655.
Lehodey P., Alheit J., Barange M., Baumgartner T., Beaugrand G., Drinkwater K., et al. (2006). Climate variability, fish, and fisheries. J. Climate 19, 5009–5030. doi: 10.1175/JCLI3898.1
Lehodey P., Bertrand A., Hobday A. J., Kiyofuji H., Mcclatchie S., Menkès C. E., et al. (2020). “ENSO impact on marine fisheries and ecosystems,” in El Niño Southern oscillation in a changing climate. (American Geophysical Union, Hoboken, NJ, Washington, D.C.: John Wiley & Sons, Inc.). 429–451.
Lewis W. M. (2022). “The ecological niche in aquatic ecosystems,” in Encyclopedia of inland waters (Second edition). Eds. Mehner T., Tockner K. (Oxford: Elsevier), 202–206.
Li Y., Wang C., Zou X., Feng Z., Yao Y., Wang T., et al. (2019). Occurrence of polycyclic aromatic hydrocarbons (PAHs) in coral reef fish from the South China Sea. Mar. pollut. Bull. 139, 339–345. doi: 10.1016/j.marpolbul.2019.01.001
Liang Z. (2022). Fish diversity and fauna characteristics in the northern South China Sea continental shelf (Guandong Ocean University).
Martins A. S., Haimovici M. (2000). Reprodution of the cutlassfish Trichiurus lepturus in the southern Brazil subtropical convergence ecosystem. 64.
Masrikat J. A. (2012). Standing stock of demersal fish assessment in southern part of South China Sea. J. Coast. Dev. 15, 276–281.
Mcphaden M. J., Zebiak S. E., Glantz M. H. (2006). ENSO as an integrating concept in earth science. Science 314, 1740–1745. doi: 10.1126/science.1132588
Mildenberger T. K., Taylor M. H., Wolff M., Price S. (2017). TropFishR: an R package for fisheries analysis with length-frequency data. Methods Ecol. Evol. 8, 1520–1527. doi: 10.1111/2041-210X.12791
Mzingirwa F. A., Nyamora J. M., Omukoto J. O., Tuda P. (2020). Stock assessment of the Tigertooth croaker, Otolithes ruber (Bloch & Schneider 1801) from the commercial prawn trawl fishery by-catch in coastal Kenya. Western Indian Ocean J. Mar. Sci. 19, 149–165. doi: 10.4314/wiojms.v19i2.11
Olden J. D., Poff N. L., Bestgen K. R. (2006). LIFE-HISTORY STRATEGIES PREDICT FISH INVASIONS AND EXTIRPATIONS IN THE COLORADO RIVER BASIN. Ecol. Monogr. 76, 25–40. doi: 10.1890/05-0330
Oli M. K., Coulson T. (2016). “Life history, what is?,” in Encyclopedia of evolutionary biology. Ed. Kliman R. M. (Oxford: Academic Press), 394–399.
Overland J. E., Alheit J., Bakun A., Hurrell J. W., Mackas D. L., Miller A. J. (2010). Climate controls on marine ecosystems and fish populations. J. Mar. Syst. 79, 305–315. doi: 10.1016/j.jmarsys.2008.12.009
Pankhurst N. W., Munday P. L. (2011). Effects of climate change on fish reproduction and early life history stages. Mar. Freshw. Res. 62, 1015–1026. doi: 10.1071/MF10269
Pauly D. (1980). On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 39, 175–192. doi: 10.1093/icesjms/39.2.175
Payne N. L., Morley S. A., Halsey L. G., Smith J. A., Stuart-Smith R., Waldock C., et al. (2021). Fish heating tolerance scales similarly across individual physiology and populations. Commun. Biol. 4, 264. doi: 10.1038/s42003-021-01773-3
Pécuchet L., Nielsen J. R., Christensen A. (2015). Impacts of the local environment on recruitment: a comparative study of North Sea and Baltic Sea fish stocks. ICES J. Mar. Sci. 72, 1323–1335. doi: 10.1093/icesjms/fsu220
Petrik C. M. (2019). “Life history of marine fishes and their implications for the future oceans,” in Predicting future oceans. (Amsterdam: Elsevier). 165–172.
Pinsky M. L., Palumbi S. R. (2014). Meta-analysis reveals lower genetic diversity in overfished populations. Mol. Ecol. 23, 29–39. doi: 10.1111/mec.12509
Planque B., Fromentin J.-M., Cury P., Drinkwater K. F., Jennings S., Perry R. I., et al. (2010). How does fishing alter marine populations and ecosystems sensitivity to climate? J. Mar. Syst. 79, 403–417. doi: 10.1016/j.jmarsys.2008.12.018
Qian J., Li J., Zhang K., Qiu Y., Cai Y., Wu Q., et al. (2022). Spatial–temporal distribution of large-size light falling-net fisheries in the South China Sea. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1075855
Qiu Y., Lin Z., Wang Y. (2010). Responses of fish production to fishing and climate variability in the northern South China Sea. Prog. Oceanography 85, 197–212. doi: 10.1016/j.pocean.2010.02.011
Qiu F., Pan A., Zhang S., Cha J., Sun H. (2015). Sea surface temperature anomalies in the South China Sea during mature phase of ENSO. Chin. J. Oceanology Limnology 34, 577–584. doi: 10.1007/s00343-016-4290-3
Ramos R. D., Goodkin N. F., Siringan F. P., Hughen K. A. (2019). Coral records of temperature and salinity in the tropical Western Pacific reveal influence of the pacific decadal oscillation since the late nineteenth century. Paleoceanography Paleoclimatology 34, 1344–1358. doi: 10.1029/2019PA003684
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria). Available at: https://www.R-project.org/.
Rojas-Mendez J. A., Mendoza C. J. R. (2008). Effects of ENSO 1997?1998 on the distribution of small pelagic fish off the west coast of Baja California. Int. J. Environ. Health 2, 45–63. doi: 10.1504/IJENVH.2008.018672
Rouyer T., Ottersen G., Durant J. M., Hidalgo M., Hjermann D.Ø., Persson J., et al. (2011). Shifting dynamic forces in fish stock fluctuations triggered by age truncation? Global Change Biol. 17, 3046–3057. doi: 10.1111/j.1365-2486.2011.02443.x
Scrucca L. (2013). GA: A package for genetic algorithms in R. J. Stat. Software 53, 1–37. doi: 10.18637/jss.v053.i04
Scrucca L. (2017). On Some Extensions to GA Package: Hybrid Optimisation, Parallelisation and Islands Evolution On some extensions to GA package: hybrid optimisation, parallelisation and islands evolution. R J. 9, 187. doi: 10.32614/RJ-2017-008
Shelton A. O., Mangel M. (2011). Fluctuations of fish populations and the magnifying effects of fishing. Proc. Natl. Acad. Sci. 108, 7075–7080. doi: 10.1073/pnas.1100334108
Shen G., Heino M. (2014). An overview of marine fisheries management in China. Mar. Policy 44, 265–272. doi: 10.1016/j.marpol.2013.09.012
Sherman K. D., King R. A., Dahlgren C. P., Simpson S. D., Stevens J. R., Tyler C. R. (2017). Historical Processes and Contemporary Anthropogenic Activities Influence Genetic Population Dynamics of Nassau Grouper (Epinephelus striatus) within The Bahamas. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00393
Soyano K., Mushirobira Y. (2018). The mechanism of low-temperature tolerance in fish. Adv. Exp. Med. Biol. 1081, 149–164. doi: 10.1007/978-981-13-1244-1_9
Sprogis K. R., Christiansen F., Wandres M., Bejder L. (2018). El Nino Southern Oscillation influences the abundance and movements of a marine top predator in coastal waters. Glob Chang Biol. 24, 1085–1096. doi: 10.1111/gcb.13892
Stenseth N. C., Mysterud A., Ottersen G., Hurrell J. W., Chan K.-S., Lima M. (2002). Ecological effects of climate fluctuations. Science 297, 1292–1296. doi: 10.1126/science.1071281
Strøm H., Descamps S., Ekker M., Fauchald P., Moe B. (2021). Tracking the movements of North Atlantic seabirds: steps towards a better understanding of population dynamics and marine ecosystem conservation. Mar. Ecol. Prog. Ser. 676, 97–116. doi: 10.3354/meps13801
Su L., Chen Z., Zhang K., Xu Y., Xu S., Wang K. (2021). Decadal-scale variation in mean trophic level in Beibu gulf based on bottom-trawl survey data. Mar. Coast. Fisheries 13, 174–182. doi: 10.1002/mcf2.10144
Su M., Wang L., Xiang J., Ma Y. (2021). Adjustment trend of China’s marine fishery policy since 2011. Mar. Policy 124. doi: 10.1016/j.marpol.2020.104322
Su L., Xu Y., Qiu Y., Sun M., Zhang K., Chen Z. (2022). Long-term change of a fish-based index of biotic integrity for a semi-enclosed bay in the Beibu gulf. Fishes 7(3), 124. doi: 10.3390/fishes7030124
Sun D., Lin Z. (2004). Variations of major commercial fish stocks and strategies for fishery management in Beibu Gulf. J. Trop. Oceanography 23, 62–68.
Taghavi Motlagh S., Ghodrati Shojaei M., Vahabnezhad A. (2021). Life history traits of ribbonfish Trichiurus lepturus (Linnaeus 1758) in the Persian Gulf and Oman Sea. Iranian J. Fisheries Sci. 20, 298–312.
Taylor M. H., Mildenberger T. K. (2017). Extending electronic length frequency analysis in R. Fisheries Manage. Ecol. 24, 330–338. doi: 10.1111/fme.12232
Then A. Y., Hoenig J. M., Gedamke T., Ault J. S. (2015). Comparison of two length-based estimators of total mortality: a simulation approach. Trans. Am. Fisheries Soc. 144, 1206–1219. doi: 10.1080/00028487.2015.1077158
Tian H., Liu Y., Tian Y., Jing Y., Liu S., Liu X., et al. (2022). Advances in the use of nighttime light data to monitor and assess coastal fisheries under the impacts of human activities and climate and environmental changes: A case study in the Beibu Gulf. Mar. Policy 144, 105227. doi: 10.1016/j.marpol.2022.105227
Timmermann A., An S.-I., Kug J.-S., Jin F.-F., Cai W., Capotondi A., et al. (2018). El Niño–southern oscillation complexity. Nature 559, 535–545. doi: 10.1038/s41586-018-0252-6
Von Bertalanffy L. (1938). A QUANTITATIVE THEORY OF ORGANIC GROWTH (INQUIRIES ON GROWTH LAWS. II). Hum. Biol. 10, 181–213.
Wang I. M., Michalak N. M., Ackerman J. M. (2021). “Life history strategies,” in Encyclopedia of evolutionary psychological science. Eds. Shackelford T. K., Weekes-Shackelford V. A. (Cham: Springer International Publishing), 4560–4569.
Wang X., Qiu Y., Du F., Lin Z., Sun D., Huang S. (2013). Spatio-temporal variability of fish diversity and dominant species in the Beibu Gulf. J. Fishery Sci. China 18, 427–436. doi: 10.3724/SP.J.1118.2011.00427
Wang Y., Yao L., Chen P., Yu J., Wu Q. E. (2020). Environmental influence on the spatiotemporal variability of fishing grounds in the Beibu Gulf, South China Sea. J. Mar. Sci. Eng. 8(12), 957. doi: 10.3390/jmse8120957
Wang K., Zhang C., Xu B., Xue Y., Ren Y. (2020). Selecting optimal bin size to account for growth variability in Electronic LEngth Frequency ANalysis (ELEFAN). Fisheries Res. 225, 105474. doi: 10.1016/j.fishres.2019.105474
Waples R. S., Audzijonyte A. (2016). Fishery-induced evolution provides insights into adaptive responses of marine species to climate change. Front. Ecol. Environ. 14, 217–224. doi: 10.1002/fee.1264
Ward H. G. M., Post J. R., Lester N. P., Askey P. J., Godin T. (2017). Empirical evidence of plasticity in life-history characteristics across climatic and fish density gradients. Can. J. Fisheries Aquat. Sci. 74, 464–474. doi: 10.1139/cjfas-2016-0023
Wu R., Chen W., Wang G., Hu K. (2014). Relative contribution of ENSO and East Asian winter monsoon to the South China Sea SST anomalies during ENSO decaying years. J. Geophysical Research: Atmospheres 119, 5046–5064. doi: 10.1002/2013JD021095
Xu S., Chen Z., Li C., Huang X., Li S. (2011). Assessing the carrying capacity of tilapia in an intertidal mangrove-based polyculture system of Pearl River Delta, China. Ecol. Model. 222, 846–856. doi: 10.1016/j.ecolmodel.2010.11.014
Xu Y., Zhang K., Sun M., Su L., Chen Z. (2022). Tempo-spatial distribution of an endangered fish species, threadfin porgy evynnis cardinalis (Lacepède 1802), in the northern South China Sea. J. Mar. Sci. Eng. 10(9), 1191.
Xu S., Zhang Z., Yu K., Huang X., Chen H., Qin Z., et al. (2021). Spatial variations in the trophic status of Favia Palauensis corals in the South China Sea: Insights into their different adaptabilities under contrasting environmental conditions. Sci. China Earth Sci. 64, 839–852. doi: 10.1007/s11430-020-9774-0
Yang B., Herrmann B., Yan L., Li J., Wang T. (2021). Size selectivity and catch efficiency of diamond-mesh codends in demersal trawl fishery for conger pike (Muraenesox cinereus) of the South China Sea. Ocean Coast. Manage. 211, 105777. doi: 10.1016/j.ocecoaman.2021.105777
Yatsu A., Aydin K. Y., King J. R., Mcfarlane G. A., Chiba S., Tadokoro K., et al. (2008). Elucidating dynamic responses of North Pacific fish populations to climatic forcing: Influence of life-history strategy. Prog. Oceanography 77, 252–268. doi: 10.1016/j.pocean.2008.03.009
Yen K.-W., Lu H.-J. (2016). Spatial–temporal variations in primary productivity and population dynamics of skipjack tuna Katsuwonus pelamis in the western and central Pacific Ocean. Fisheries Sci. 82, 563–571. doi: 10.1007/s12562-016-0992-x
Zhang K., Cai Y., Liao B., Jiang Y., Sun M., Su L., et al. (2020). Population dynamics of threadfin porgy Evynnis cardinalis, an endangered species on the IUCN red list in the Beibu Gulf, South China Sea. J. Fish Biol. 97, 479–489. doi: 10.1111/jfb.14398
Zhang K., Li J., Hou G., Huang Z., Shi D., Chen Z., et al. (2021). Length-based assessment of fish stocks in a data-poor, jointly exploited (China and Vietnam) fishing ground, Northern South China Sea. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.718052
Zhang K., Li M., Li J., Sun M., Xu Y., Cai Y., et al. (2022). Climate-induced small pelagic fish blooms in an overexploited marine ecosystem of the South China Sea. Ecol. Indic. 145, 109598. doi: 10.1016/j.ecolind.2022.109598
Zhang K., Zhang J., Zhang P., Su L., Hong X., Qiu Y., et al. (2023). This is what we know: Assessing the stock status of the data-poor skipjack tuna (Katsuwonus pelamis) fishery in the South China Sea. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1095411
Keywords: Beibu Gulf, major commercial fishes, ELEFAN, life-history trait parameters, El Niño-Southern Oscillation
Citation: Hong X, Zhang K, Li J, Xu Y, Sun M, Jiang J, Xu S, Cai Y, Qiu Y and Chen Z (2023) Impacts of climate events on life history parameters of major commercial fishes in the Beibu Gulf, South China Sea in the last 15 years. Front. Mar. Sci. 10:1234772. doi: 10.3389/fmars.2023.1234772
Received: 05 June 2023; Accepted: 14 September 2023;
Published: 03 October 2023.
Edited by:
Jingzhen Wang, Beibu Gulf University, ChinaReviewed by:
Xiujuan Shan, Chinese Academy of Fishery Sciences (CAFS), ChinaCopyright © 2023 Hong, Zhang, Li, Xu, Sun, Jiang, Xu, Cai, Qiu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zuozhi Chen, Y2hlbnp1b3poaUBzY3NmcmkuYWMuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.