 Peili Mao
Peili Mao Qingzhi Lin†
Qingzhi Lin† Ruiqiang Ni
Ruiqiang Ni Xin Han
Xin Han
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 22 August 2023
Sec. Marine Ecosystem Ecology
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1231928
This article is part of the Research Topic Degradation, Ecological Restoration and Adaptive Management of Estuarine Wetlands under Intensifying Global Changes, volume II View all 11 articles
Elucidating the effect of soil moisture on the adaptation of dominant plants in coastal wetlands is important for predicting the evolution of vegetation in the region. In this paper, Tamarix chinensis, a dominant species in the Yellow River Delta, was used as the object to study the changes of its growth and physiological parameters with increasing soil salinity under different moisture conditions (normal watering, persistent drought and persistent waterlogging). Different salt stress (2‰, 5‰, 8‰, 12‰, 16‰, and 20‰) using pot experiments was also used to reveal the mechanism of soil moisture on its salt tolerance. The results showed that the relative growth rate between 5‰-8‰ soil salinity was the largest, and growth was significantly inhibited above 20‰. Among different moisture conditions, the difference in relative growth rate under normal watering and persistent drought were nonsignificant, while both were significantly lower than those under persistent waterlogging. With increasing soil salinity, relative water content and total chlorophyll content significantly decreased, and cell membrane permeability (malondialdehyde), sodium ion, osmoregulatory substances (proline, soluble protein), and protective enzyme activity (SOD) significantly increased, while changes in non-structural carbohydrates (NSC) were not significant. Compared with normal watering and persistent waterlogging, persistent drought had the lowest leaf relative water content, total chlorophyll content, and sodium ions, and the highest cell membrane permeability, osmoregulatory substances and protective enzyme activity. With increasing treatment time, the relative leaf water content and total chlorophyll content significantly decreased, and cell membrane permeability, osmoregulatory substances and protective enzyme activity increased more significantly than normal watering and persistent waterlogging. NSC increased under normal watering and persistent waterlogging, while significantly decreased under persistent drought. Correlation analysis showed that the relationships between sodium ions, total chlorophyll content and malondialdehyde were various under different moisture conditions. Under persistent drough, malondialdehyde was significantly positively correlated with relative conductivity, superoxide dismutase, proline, soluble protein and soluble sugar. Total chlorophyll content was the key indicator reflecting the salt and waterlogging tolerance of T. chinensis under normal watering and persistent waterlogging, while cell membrane damage was under persistent drought. In summary, T. chinensis has strong salt and waterlogging tolerance, but persistent drought with salt stress can have serious impacts on its growth and survival.
Along with the frequent occurrence of extreme climate events from global warming, the increase in temperature leads to more evaporation of water and exacerbates drought in wetlands (Middleton and Kleinebecker, 2012). Water availability and variability will have a significant impact on the evolution of global vegetation (Smith and Boers, 2023). Climate warming has increased the probability of coastal flooding by melting iceberg and permafrost, and thus rising sea levels (Taherkhani et al., 2020), which severely impact on coastal wetland ecosystems (Schuerch et al., 2018). Jiang et al. (2013) found that climate features play a dominant role in seasonal variation in vegetation cover in the coastal wetland. Lu et al. (2016) reported that the increasing in annual average temperature, annual precipitation, and humidity index of the Yellow River Delta from 1986 to 2005 led to a significant decrease in river runoff and reduced the water supply to the marshes. The inflow of freshwater has a significant impact on the water-salt status of coastal wetlands, which in turn affects the plant community and vegetation dynamics (Cui et al., 2009). Therefore, it is important to reveal the adaptation of plants to water and salt changes to predict the dynamic of vegetation in the coastal wetland (Sun et al., 2022).
Climate change is predicted to lead soil moisture and salinity particularly in the coastal regions through sea level rise and salt water intrusion (Rahmstorf, 2007; Dasgupta et al., 2009). Soil moisture has an important influence on the adaptation of plants in coastal saline lands (Souid et al., 2018). During the rainy season in this region, soil water content increases and soil salinity decreases, but in the dry season the changes are reversed (Souid et al., 2018; Guo et al., 2021). Studies on the halophyte Limonium delicatulum have found that the salt ion content and protective enzyme activity in its aboveground parts were significantly higher in the dry season than in the rainy season (Souid et al., 2018), and osmoregulatory substances had similar changes (Maaloul et al., 2021). Severe drought can cause extensive vegetation mortality in coastal wetlands (Watson et al., 2016; Liu et al., 2020a). In a restoration of vegetation in an estuarine wetland of the Liao River in China, it was found that Suaeda salsa mortality significantly increased during the dry season, which was closely related to the synergistic effects of drought and high salinity (Liu et al., 2020b). Seedling height and seedling biomass of S. salsa increased under light salt and drought co-stress treatments, but significantly decreased under heavy salt and drought stress, much more severely than single salt stress (Jia et al., 2018). Waterlogging is one of the key factors determining the distribution of vegetation in coastal wetlands (Hou et al., 2020). Flooding significantly reduced the biomass of the halophyte Triglochin maritima and Argentina pacifica, and the aboveground biomass of Carex lyngbyei and A. pacifica was significantly affected by a combination of salt and flooding (Buffington et al., 2020). In the Yellow River Delta, the growth of Phragmites australis (Guan et al., 2017) and S. salsa (Guan et al., 2011) has also been inhibited by flooding. Smith and Lee (2015) found that prolonged flooding for a few months can eradicate Spartina alterniflora. Thus, salinity and waterlogging are key factors in determining distribution of vegetation in coastal wetlands (Hou et al., 2020).
The growth and development of plants were inhibited under salt stress. With increasing salt concentration, Na+ accumulation caused a decrease in chlorophyll content and decomposition (Borzouei et al., 2020; Irshad et al., 2021; Mushtaq et al., 2021), leading to a decrease in photosynthetic capacity (Muchate et al., 2016). Salt stress leads to the accumulation of reactive oxygen in plants, and the excess of reactive oxygen peroxidizes cell membrane lipids, disrupting the intra-membrane environment and affecting all physiological metabolism (Nxele et al., 2017; Kiarash et al., 2020). Excessive salinity in the soil makes the soil water potential usually lower than the plant cells, and plants have difficulty in water uptake and water retention reduction (Mao et al., 2016; Nxele et al., 2017). Salt stress increases Na+ content in plants, inhibiting the uptake and transport of nutrients such as Ca2+ and K+ and disrupting ionic balance (Muchate et al., 2016). Osmoregulation and antioxidant regulation are important mechanisms by which plants adapt to salt stress (Rangani et al., 2018). Under salt stress, osmoregulatory substances significantly increase, reducing the osmotic potential and increasing the ability to absorb water (Benjamin et al., 2019; Borzouei et al., 2020). These nitrogenous compounds significantly enhance the antioxidant effect of plants as well as acting as osmoregulatory substances (Khoshbakht et al., 2018; Arbelet-Bonnin et al., 2020). To avoid cell membrane damage resulting from the massive accumulation of reactive oxygen under salt stress, plants synthesize and accumulate antioxidant enzymes and antioxidants to protect cell membranes (Muchate et al., 2016; Irshad et al., 2021; Mushtaq et al., 2021). However, at high salt concentrations, osmoregulatory substances such as soluble sugars, soluble proteins, and proline content are significantly reduced, and osmoregulatory capacity is significantly reduced (Ghaderi et al., 2018; Mushtaq et al., 2021), and protective enzyme activity is also reduced (Ghaderi et al., 2018).
Non-structural carbohydrates (NSC) are substrates for plant growth and metabolism, consist mainly of soluble sugars and starch (Hoch et al., 2003). NSC are products of photosynthesis, a temporary storage material between photosynthesis and utilization. When photosynthesis is insufficient, plants consume NSC to provide energy (O’Brien et al., 2014). NSC also has important roles in osmoregulation and defense against diseases and insects under stressful conditions (Sala et al., 2012; Dietze et al., 2014). Soluble sugars are an important osmoregulatory substance and provide direct available substrates and energy for physiological activities such as plant growth and respiration (Hartmann and Trumbore, 2016), while starch is stably stored in organs as a storage carbon source (Macneill et al., 2017). It was found that plants actively increase the concentration of nonstructural carbohydrates in their bodies under adversity (Gao et al., 2021). When plants are subjected to stress condition, starch and sugar can be converted to each other in response to the external environment (Kozlowski, 1992; da Silva et al., 2019). Under stress, the consumption of photosynthesis, respiration, and osmoregulation in plants determines the amount of NSC (Guo et al., 2020). A comparative study revealed that plant NSC changes under drought stress and salt stress are not the same (Cui et al., 2019). And how water conditions affect the accumulation of NSC under salt stress is still not clear.
The Yellow River Delta is the most extensive and youngest coastal wetland in the primary stage of succession in the temperate zone of China (Cong et al., 2019). Previous study (Li et al., 2019) has shown that soil salinity as a key environmental factor direactly and hydrological factors by altering soil salinity indirectly affect the spatial differentiation pattern of plant communities. T. chinensis, one of the dominant species in this area, is mainly distributed in supratidal wetlands on higher terrain or far from the sea and inland saline areas generally below 4 m in elevation, which are characterized by seasonal waterlogging and heavy salinization. Seasonal waterlogging occurs in summer, when a shallow water is connected to surface waterlogging, creating a highly saline and waterlogged environment. In spring, late autumn, and winter, precipitation is often scarce, and T. chinensis faces the combined effects of salinity and drought. Moisture and salinity are key factors affecting the growth of T. chinensis forests in this region, leading to low quality and inefficiency of the stands (Xia et al., 2013). Current studies on T. chinensis mainly focused on soil nutrient characteristics at the community level (Yang et al., 2021), distribution patterns at the population level (Jiao et al., 2021), and physiological and biochemical characteristics under salt and drought stress conditions (Liu et al., 2014). Also, the effects of soil moisture (Gao et al., 2017) and groundwater level (Xia et al., 2017) on the photosynthetic physiology of T. chinensis had been systematically studied. However, studies on the effect of soil moisture on the salt acclimation of T. chinensis are still insufficient. Precipitation in the Yellow River Delta gradually increased in recent decades (Lu et al., 2016). Thus, the study on physiological and ecological responses of T. chinensis seedlings under different soil salinity and soil moisture conditions (normal watering, persistent drought and persistent waterlogging) using pot experiments will contribute to systematically reveal the mechanisms of moisture conditions on its salt acclimatization. The aim is to provide theoretical and technical support for vegetation restoration and ecological construction in coastal wetland ecosystems.
The study was conducted in Dongying City (36°55´~38°10´ N, 118°07´~119°10´ E) in the Yellow River Delta of China. It has a warm temperate monsoon continental climate, where the annual average temperature is 12.3°C, the annual average frost-free period is 210 d, and the annual average precipitation is 559 mm, 70% of which is distributed in summer. Due to the influence of the Yellow River flooding sediment, the soil is mainly coastal tidal salt soil, the soil salt content is high, depending on the salt content of the growth of various types of salt vegetation. The main vegetation types in this area are scrub and saline meadows, and the common ones are S. salsa, Miscanthus sacchariflorus, P. australis, T. chinensis.
The experiment was conducted in the greenhouse of Shengda Eco-Forestry, Shengda Branch of Shengli Oilfield (37°20’53” N, 118°37’10” E), which is located in Hekou District, Dongying City, Shandong Province (Figure 1). The soil in the forest farm has a salinity of about 2‰, a soil pH of 8.21, a total nitrogen content of 0.54 g/kg, a hydrolytic nitrogen content of 50.79 mg/kg, an effective phosphorus content of 4.42 mg/kg, a fast-acting potassium content of 140.55 mg/kg, and organic matter content of 10.58 g/kg. The soil from this forest farm was used for potted test of T. chinensis seedlings. The T. chinensis seedlings used for the pot test were annual cuttings with an average height of 20 cm and an average ground diameter of 15 mm. The cuttings were transplanted into 15.5 cm × 22.8 cm × 19 cm (bottom diameter × diameter × height) pots on April 30, 2021, and one plant individual in each pot.
Figure 1 Geographical location of the study area and experiment site (Shengda Eco-Forestry, Shengda Branch of Shengli Oilfield).
After transplanting, the T. chinensis annual cuttings were placed in a refinement shed for growth, during which regular watering and weeding were carried out. Seedlings of uniform growth were selected for the experiment on June 19th, and the soil salinity (soil salinity/soil dry weight) was controlled by configuring different concentration gradients of NaCl solution in multiple waterings, with salt concentrations of 5‰, 8‰, 12‰, 16‰, and 20‰, and the soil salinity of 2‰ in the woodland was used as the control. The bottom of the pots were equipped with trays. To prevent salt loss, the leaking water from the trays was poured back into the pots and the trays were cleaned, and the cleaning water was also poured into the pots. The whole experiment was divided into three groups based on water treatment, each group including two factors of salinity and treatment time, and arranged in a randomized group design with 15 replicates for each treatment. The soil salinity in the pots reached the design level on 0d. The three moisture treatments on the same day were normal watering, persistent drought and persistent waterlogging.
Normal watering test: the normal watering amount was 80% of the soil field water holding capacity, using the weighing method and regular water supplementation. Mature leaves of T. chinensis with the same growing parts and normal growth were collected at 1d, 11d, 21d and 31d, respectively, for index determination. The normal watering lasted for 31 days.
Persistent waterlogging test: the double set pot method was used so that the bottom of the pots did not leak and watered heavily until the level was parallel with the sides of the pots, after which water was replenished daily. Similar to the normal watering experiment, mature leaves with the same growth site and normal growth were collected on days 1, 11, 21, and 31 for indicator measurement. The persistent waterlogging test lasted for 31 days.
Persistent drought test: watering was done by small and repeated watering until the water seeped out from the bottom of the pot, and then no watering was needed. Leaves were collected at 1d, 11d, and 21d of stress and soil was taken at 10 cm from the bottom of the pot using the perforation method to determine the water and salinity content of the soil. Due to the death of all the aboveground parts of T. chinensis on the 31st day, no sampling was conducted. The average soil moisture content was 13.21%, 9.49%, and 3.77% on days 1, 11, and 21, respectively. The soil moisture content gradually decreases with the duration of stress, and the soil salt content gradually increased (Table 1). The persistent drought test lasted for 21 days.

Table 1 Variation of soil salt content under persistent drought (‰).
At the beginning and end of the test, the height and ground diameter of T. chinensis seedlings were measured, the seedling height was measured using a meter ruler, and the ground diameter was measured using a vernier caliper. At the end of the experiments, all seedlings were dug out and the whole root system was obtained, and the root system was slowly rinsed with running water. The above-ground parts and roots were placed in an oven at 75°C for 48 h and then weighed for dry weight using a one-thousandth electronic balance.
Soil moisture content (SMC, %) was measured by the drying method; soil salt content (SSC, ‰) was determined using the conductivity method, and the standard curve of soil salt concentration was measured using the drying residue method and the conductivity meter.
Total chlorophyll content (Chl, mg/g) was determined by acetone extraction; leaf cell membrane permeability was determined by relative conductivity (RC, %) method; Malondialdehyde (MDA, μmol/g) content was determined by the thiobarbituric acid method; superoxide dismutase (SOD, U/g) activity was determined by the nitrogen blue tetrazolium photoreduction method, and 50% of the nitrogen blue tetrazolium photoreduction was used as one enzyme activity unit (U); Proline (Pro, μg/g) content was determined by acidic ninhydrin colorimetric method; soluble sugar (SS, mg/g) and starch (ST, mg/g) content were determined by anthrone colorimetric method; soluble protein (SP, mg/g) content was determined by coomassie blue staining (Wu, 2018). The contents of Na+ (%) were determined by the water bath extraction method using a mixture of perchloric acid and concentrated nitric acid (Ou et al., 2019).
Relative height growth rate (RGRH, cm/(cm.d)):
Relative ground diameter growth rate (RGRD, mm/(mm.d)):
Where: H1 is the seedling height at the beginning of the trial, H2 is the seedling height at the end of the trial, D1 is the seedling ground diameter at the beginning of the trial, D2 is the seedling ground diameter at the end of the trial, T1 represents the start time of the trial, and T2 represents the end time of the trial.
FIGO, International Federation of Gynecology and Obstetrics; NOS, Newcastle-Ottawa Scale; PD-1, programmed death-1.
Excel 2010 was used to organize and calculate the data and draw graphs, and SPSS 17.0 was used for all experimental data analysis. A two-way ANOVA was performed on the relative growth rates of T. chinensis seedlings for water treatment and salt gradient, and a three-factor ANOVA was performed on the physiological indicators (Chl, RC, MDA, SOD, Pro, SP,RWC, ST, SS, NSC and Na+) for water treatment, salt gradient, and stress time. If the differences were significant, multiple comparisons were then performed using the least significant difference (LSD) method with a significance difference test level of P< 0.05. Correlation analyses and principal component analysis were conducted separately for the physiological indicators (Chl, RC, MDA, SOD, Pro, SP,RWC, ST, SS, NSC and Na+) under different moisture treatments.
Soil salt treatment had a significant effect on RGRH (F=2.62, P<0.05) and RGRD (F=4.63, P<0.01), moisture conditions had a highly significant effect on RGRH (F=6.48, P<0.01) and RGRD (F=19.81, P<0.01), and the interactions between the two factors had insignificant effects on RGRH (F=0.16, P>0.05) and RGRD (F=0.66, P>0.05). With increasing soil treatment, both RGRH and RGRD increased and then decreased, with RGRH being maximum at 5‰ (Figure 2A), while RGRD was maximum at 5‰ under normal watering and maximum at 8‰ under persistent drought and persistent waterlogging (Figure 2B). Among moisture conditions, both RGRH and RGRD were not significantly different between normal watering and persistent drought (P>0.05), and both were highly significantly lower than persistent waterlogging (P<0.01).
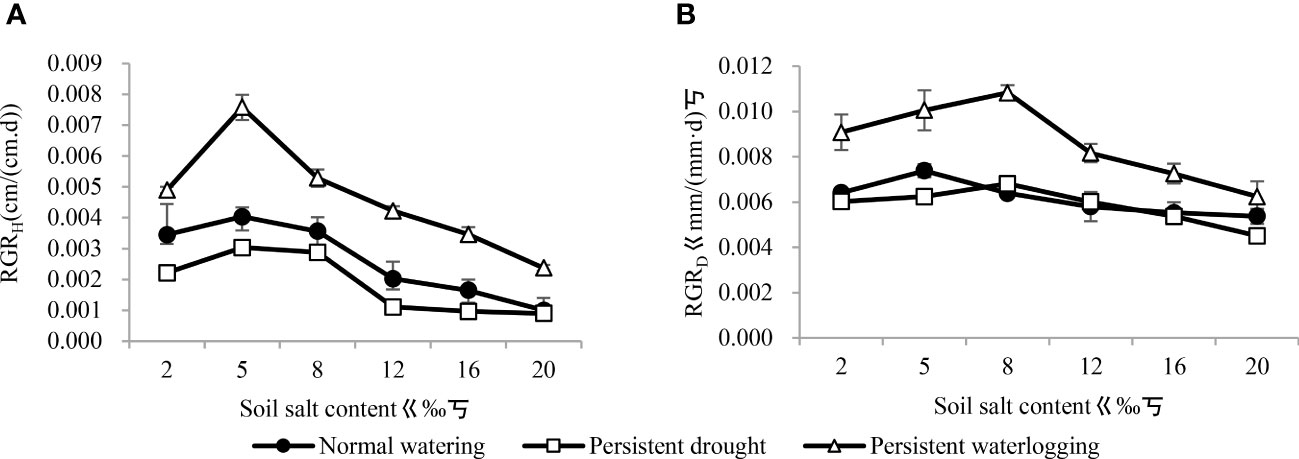
Figure 2 Changes in relative growth rates of Tamarix chinensis seedling height (A) and ground diameter (B) under different water conditions with increasing soil salinity.
Soil salt treatment, moisture conditions and treatment time and the interactions among these three factors all had highly significant effects on leaf relative water content (Table 2). The leaf relative water content decreased significantly with increasing soil salt concentration (Figure 3). Under moisture conditions, the relative water content showed as persistent drought< normal watering< persistent waterlogging (P<0.01). The pattern of variation of leaf relative water content with the duration of treatment was different among moisture conditions. As the treatment time increased, the leaf relative water content decreased slowly under normal watering (Figure 3A), rapidly under persistent drought (Figure 3B), and not significantly under persistent waterlogging (Figure 3C). Under normal watering and persistent drought, the higher the soil treatment and the longer the treatment time, the lower the leaf relative water content.
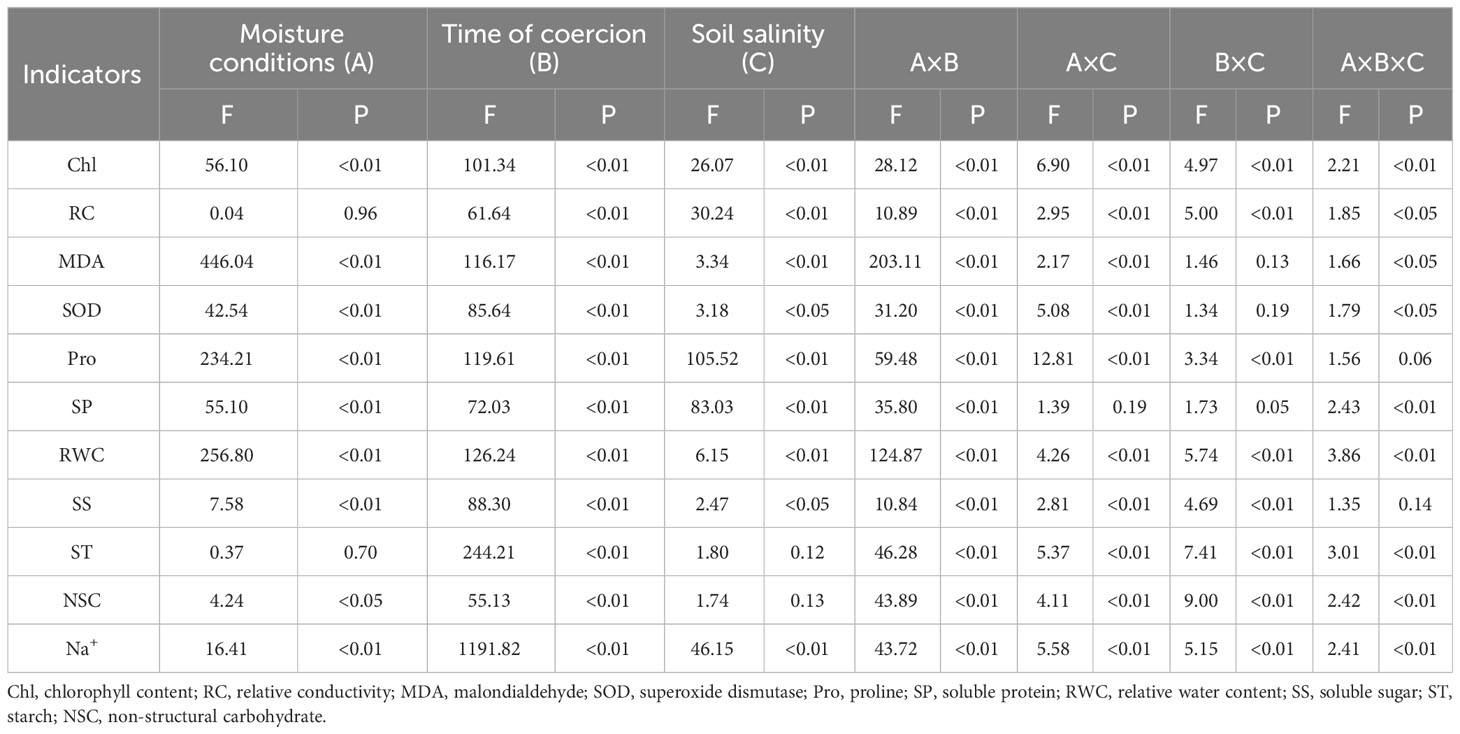
Table 2 Three-way ANOVA of the effects of water conditions, stress time and soil salinity on physiological indicators of Tamarix chinensis seedlings.
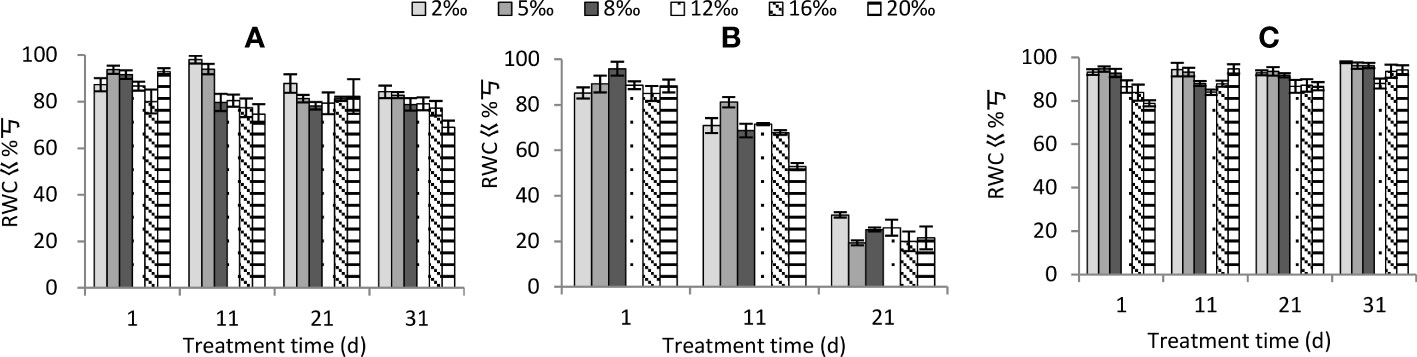
Figure 3 Changes in relative water content of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time. (A) normal watering; (B) persistent drought; (C) persistent waterlogging.
Total chlorophyll content was highly significantly affected by soil salt treatment, water conditions and time of treatment, and the effect of interactions among the three factors were also significant (Table 2). Total chlorophyll content significantly decreased with increasing soil salt content (Figure 4). Among different water conditions, the ranking of total chlorophyll content was persistent drought< normal watering< persistent waterlogging (P<0.01). With increasing treatment time, total chlorophyll content gradually increased under normal watering (Figure 4A) and persistent waterlogging (Figure 4C), while it increased and then decreased under persistent drought (Figure 4B). Overall, the poorer the water conditions, the higher the soil treatment and the lower the total chlorophyll content.
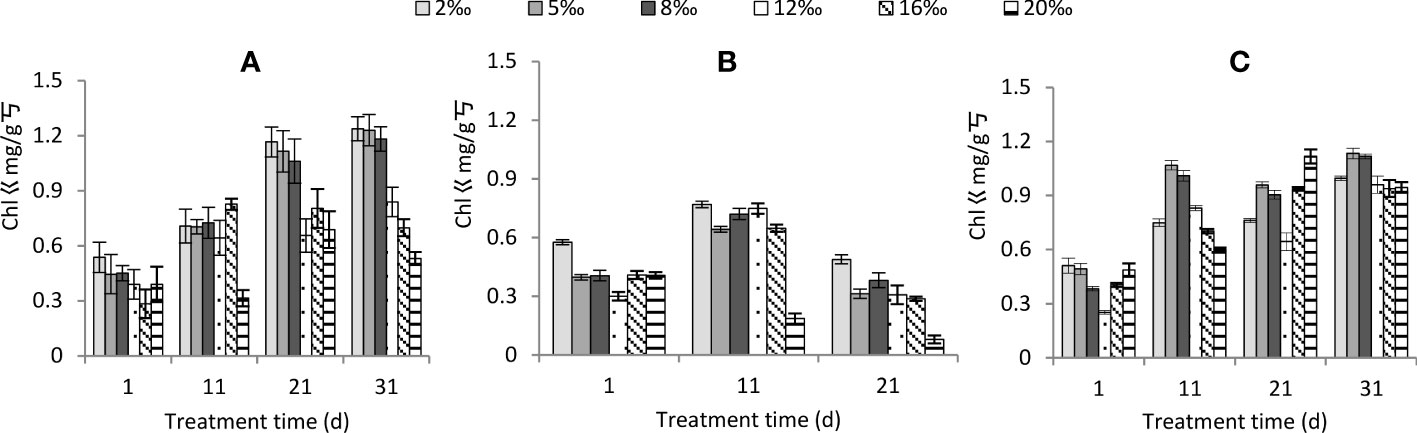
Figure 4 Changes in total chlorophyll content of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time. (A) normal watering; (B) persistent drought; (C) persistent waterlogging.
Soil salt treatment and treatment time had significant effects on relative conductivity, moisture conditions had insignificant effect, and the interactions of all three factors were significant (Table 2). As the soil salt content increased, the relative conductivity decreased and then increased, with 12‰ being the lowest and 20‰ being the highest (Figure 5). The relative conductivity gradually increased with increasing treatment time (Figure 5). Overall, the relative conductivity increased less under normal watering and persistent impregnation, and the relative conductivity increased the most under each soil salt content and the most under persistent drought.
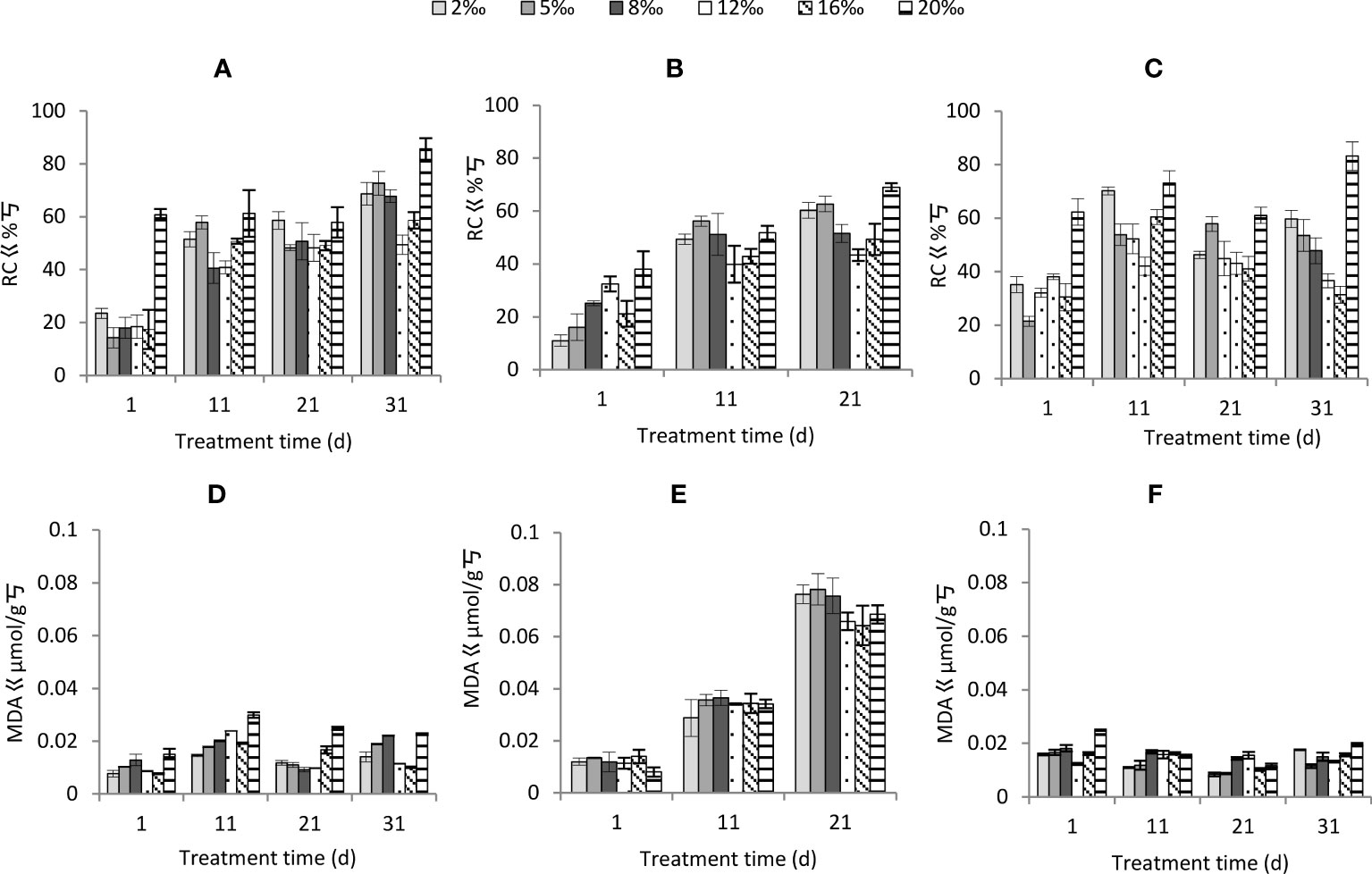
Figure 5 Changes in cell membrane permeability of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time. (A) Relative conductivity under normal watering; (B) Relative conductivity under persistent drought; (C) Relative conductivity under persistent waterlogging; (D) Relative conductivity under normal watering; (E) Relative conductivity under persistent drought; (F) Relative conductivity under persistent impregnated water.
Malondialdehyde was significantly affected by soil salt treatment, soil conditions and time of treatment, and the interactions of the three factors (except time and salt) also had a significant effect (Table 2). The malondialdehyde content fluctuated and increased with increasing soil salt content, with the highest at 20‰ (Figure 5). The difference in malondialdehyde content between moisture conditions was not significant between normal watering and persistent water logging and was significantly lower than that of persistent drought. Malondialdehyde content fluctuated with increasing treatment time under normal watering (Figure 5) and persistent waterlogging for each soil salt content (Figure 5C), but significantly increased under persistent drought (Figure 5B).
Na+ content was significantly affected by soil treatment, water conditions and treatment time, and the interactions among the three factors were also significant (Table 2). The Na+ content increased significantly with increasing soil treatment (Figure 6). Under moisture conditions, the Na+ content were ranked as persistent drought< normal watering< persistent water logging. The Na+ content also increased significantly with the increasing duration of treatment. Overall, the better the moisture conditions, the higher the soil treatment and the longer the duration of treatment, the higher the Na+ content (Figure 6).
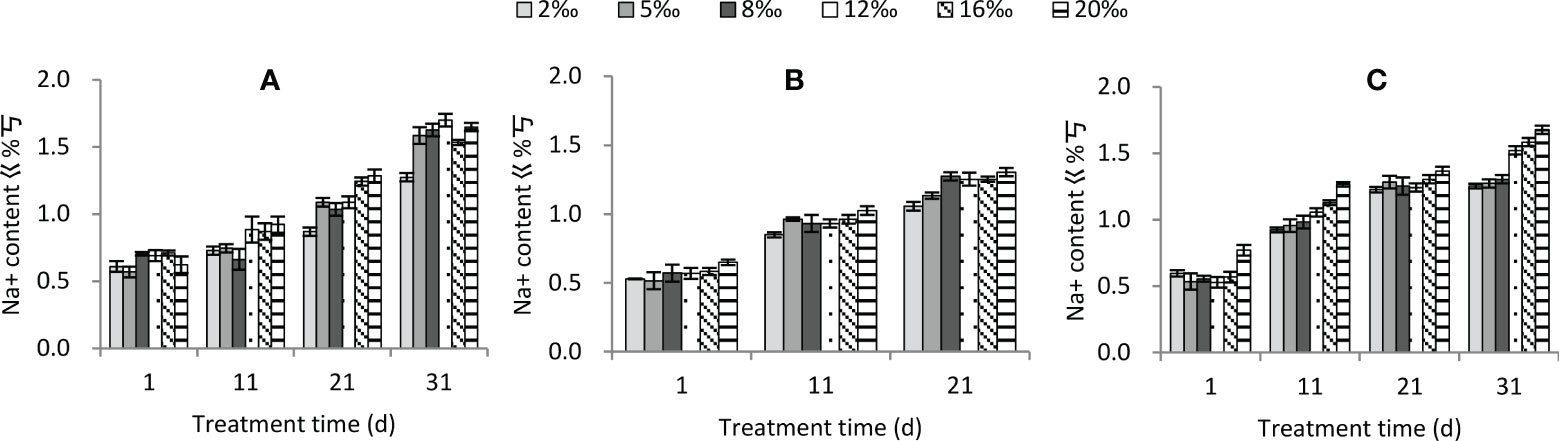
Figure 6 Changes in Na+ content of Tamarix chinensis seedlings with increasing treatment time under different soil moisture conditions and soil salt content. (A) normal watering; (B) persistent drought; (C) persistent waterlogging.
Soil treatment gradient, moisture conditions and time of treatment all had significant effects on proline and soluble protein content (Table 2). Both proline and soluble protein content significantly increased with increasing soil treatment (Figure 7). Among water conditions, proline was ranked as persistent waterlogging< normal watering< persistent drought, and soluble protein was ranked as normally watering< persistent waterlogging< persistent drought. Both proline and soluble protein content significantly increased with increasing treatment time. In terms of interaction of three factors, except for soil treatment gradient × moisture condition × stress time, the other interaction of three factors had significant effects on proline. And for soluble protein, except for soil treatment gradient × moisture condition and soil treatment gradient × treatment time, the other interaction of three factors had significant effects (Table 2). Overall, the higher the soil treatment and the longer the treatment time, the higher the proline and soluble protein content under persistent drought, while the normal watering and persistent waterlogging did not vary much in the stress time (Figure 7).
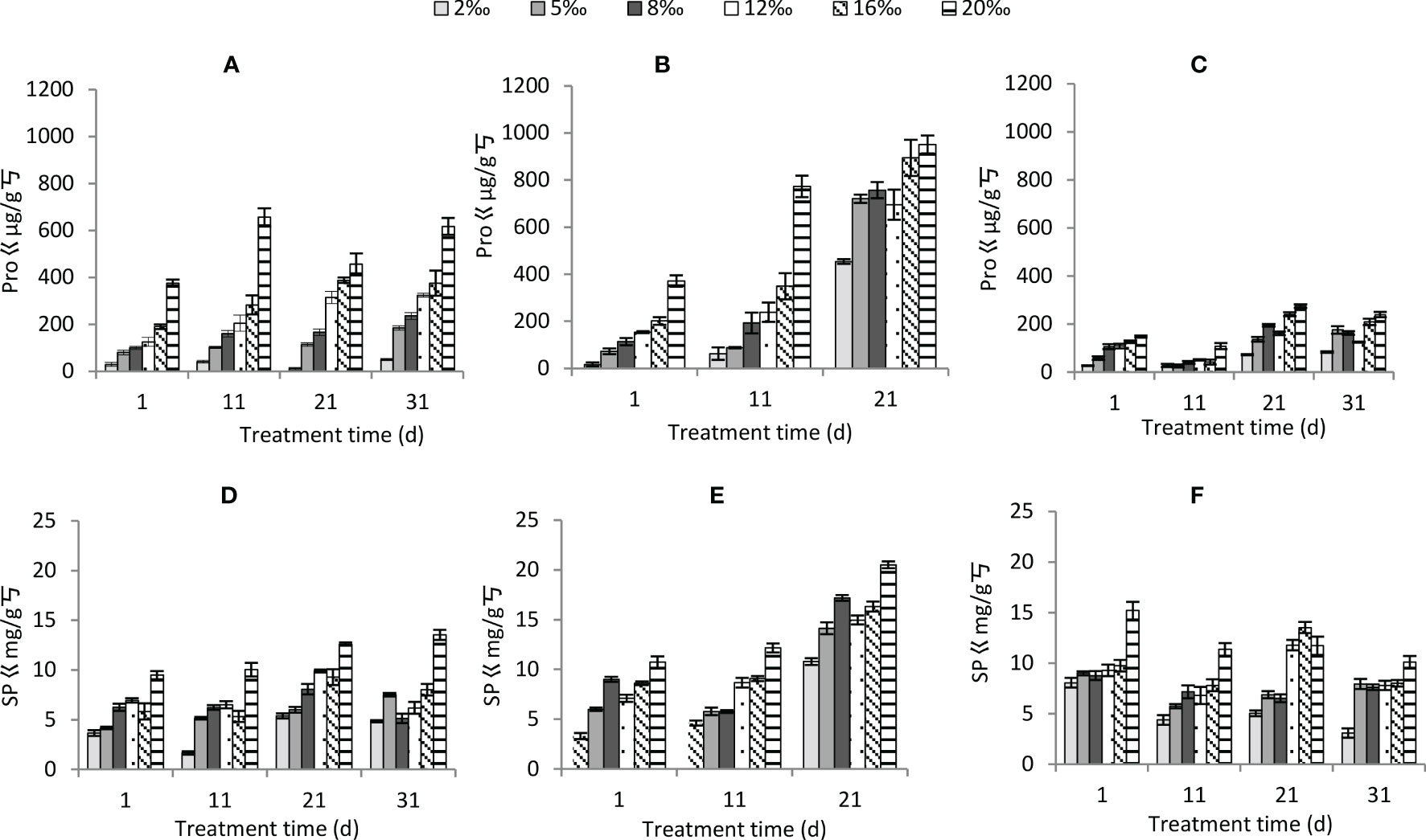
Figure 7 Changes in osmoregulatory substances of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time (A) proline under normal watering; (B) proline under persistent drought; (C) proline under persistent waterlogging (D) soluble protein under normal watering; (E) soluble protein under persistent drought; (F) soluble protein under persistent waterlogging.
SOD activity was significantly affected by salt treatment, moisture conditions and treatment time (Table 2). The interactions of the three factors, except soil salt gradient × treatment time, also had a significant effect on it (Table 2). Among the soil salt treatments, SOD activity was greatest at 20‰ (P<0.01), while the differences between the other treatments were not significant (P>0.05). The differences between normal watering and persistent waterlogging were not significant (P>0.05), and both were significantly lower (P<0.01) than persistent drought. The SOD activity increased significantly with increasing treatment time (Figure 8). Under persistent drought, the increase in SOD activity with increasing treatment time was significantly higher than that of normal watering and persistent waterlogging.
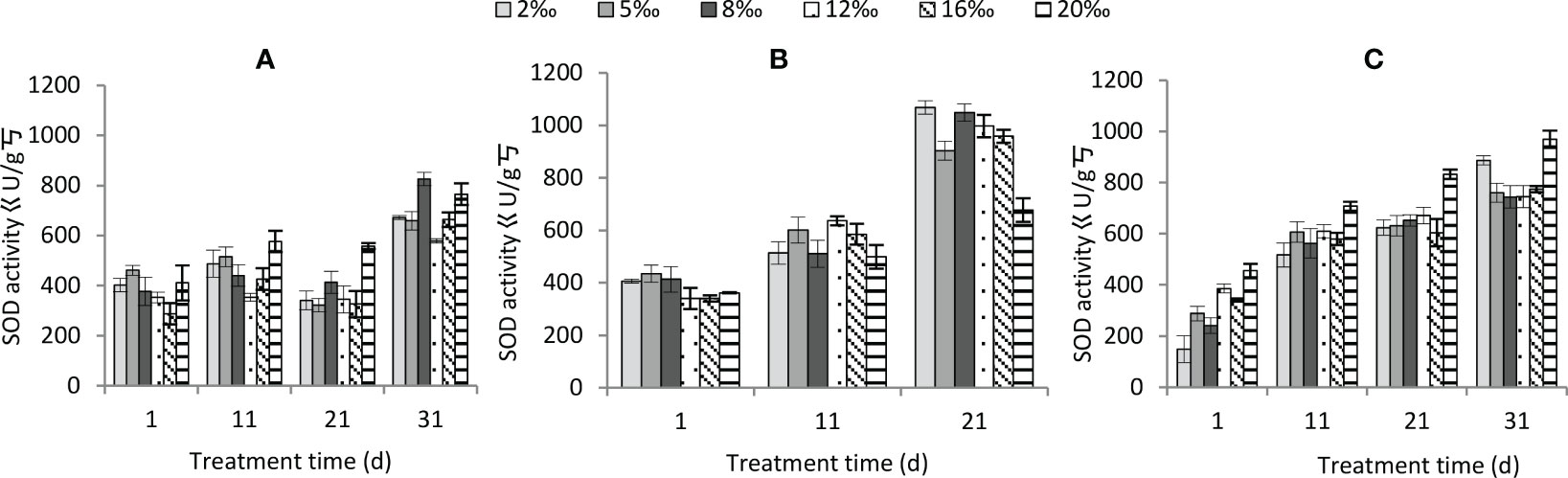
Figure 8 Changes in SOD enzyme activity of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time. (A) normal watering; (B) persistent drought; (C) persistent waterlogging.
Soil salt gradient, moisture conditions and treatment time had significant effects on soluble sugars. Soil salt gradient and moisture conditions did not have significant effects on starch, while treatment time had significant effects on starch (Table 2). The soil salt gradient had no significant effect on NSC, and moisture conditions and treatment time had significant effects on it. Among moisture conditions, soluble sugars were ranked as normal watering > persistent waterlogging > persistent drought; NSC content was significantly greater in normal watering than in persistent drought and persistent waterlogging (P<0.05), and the difference between persistent drought and persistent waterlogging was not significant (P>0.05).
The interaction of the three factors significantly affected soluble sugars, starch and NSC (except for soil treatment gradient × moisture condition and soil treatment gradient × treatment time which did not significantly affect soluble sugars) (Table 2). Soluble sugars, starch and NSC increased gradually with stress time under both normal watering (Figures 9A, D, G) and persistent waterlogging (Figures 9C, F, I), whereas they all decreased gradually under persistent drought (Figures 9B, E, H).
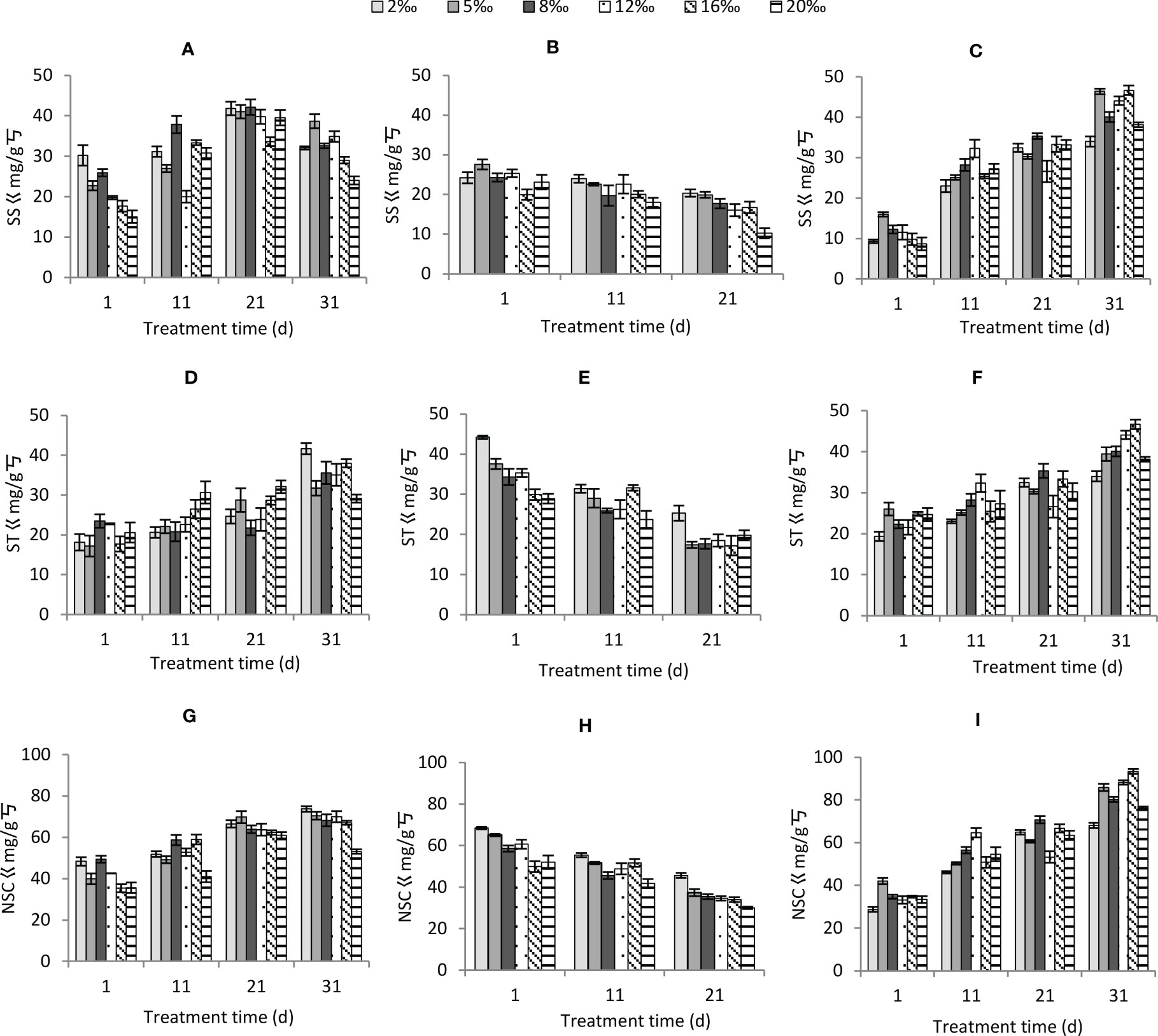
Figure 9 Changes in non-structural carbohydrates of Tamarix chinensis seedlings under different soil moisture conditions and soil salt content with increasing treatment time (A) soluble sugars under normal watering; (B) soluble sugars under persistent drought; (C) soluble sugars under persistent waterlogging (D) starch under normal watering; (E) starch under persistent drought; (F) starch under persistent waterlogging (G) Non-structural carbohydrates under normal watering; (H) Non-structural carbohydrates under persistent drought; (I) Non-structural carbohydrates under persistent waterlogging.
Under normal watering (Table 3), Na+ was significantly and positively correlated with total chlorophyll content, relative conductivity, SOD activity, proline and soluble sugar content (P< 0.01), while had highly significantly negative correlations (P< 0.01) with relative leaf water content and soluble starch. MDA was highly significantly and positively correlated with SOD (P< 0.01), and highly and negatively correlated with ST (P< 0.01) and NSC (P< 0.01). ST was highly significantly and negatively correlated with RC (P< 0.01), Pro (P< 0.01) and SS (P< 0.01), and SS was positively correlated with RC (P< 0.01).
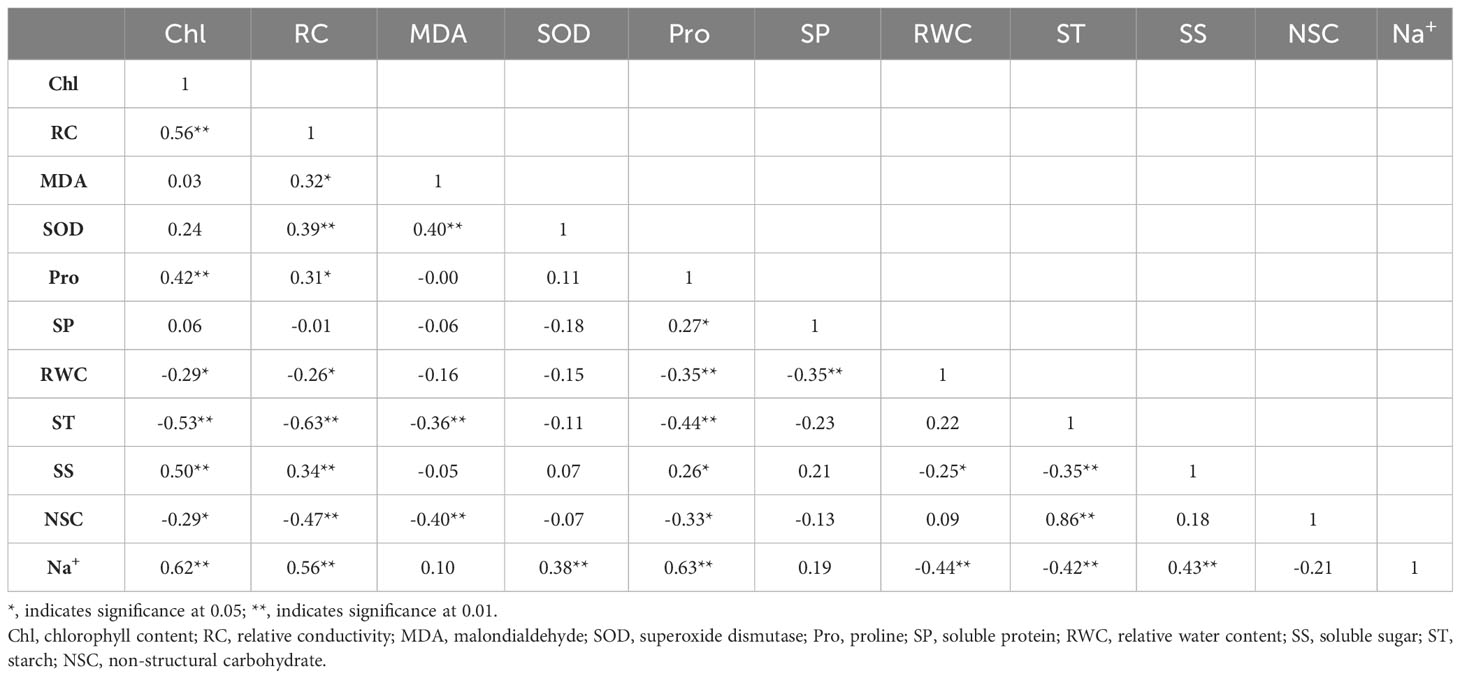
Table 3 Correlation analysis of physiological indices of Tamarix chinensis seedlings response to salinity under normal watering.
Four principal components were obtained under normal watering conditions, with contributions of 24.64%, 21.16%, 15.01%, and 14.57% in the order, with a cumulative contribution of 75.38% (Table 4). The first principal component consisted of chlorophyll, soluble sugars, sodium ions, relative conductivity, and proline, reflecting the changes in chlorophyll. The second principal component consisted mainly of NSC and starch, reflecting the changes in nonstructural carbon.
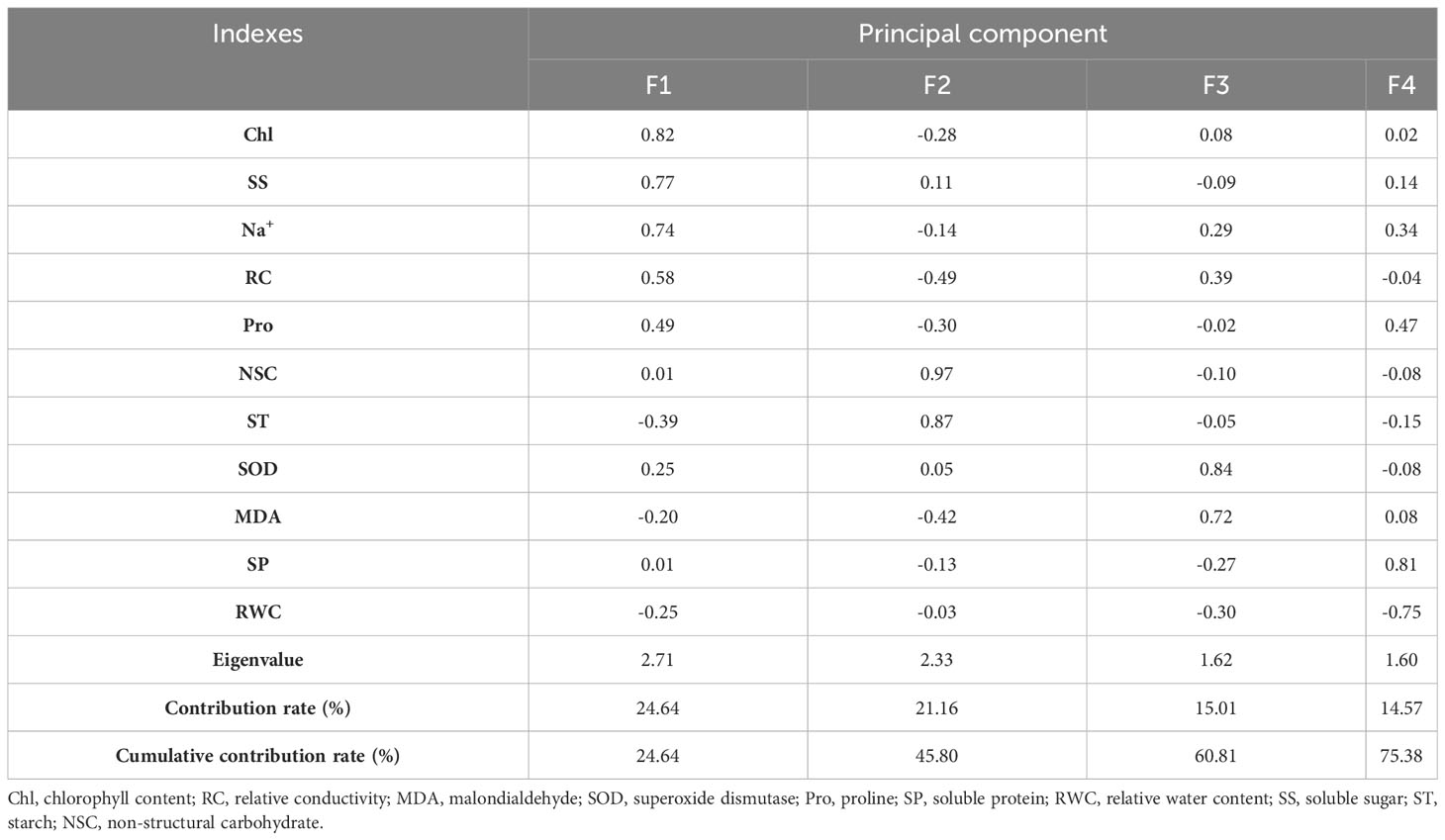
Table 4 Principal component analysis of physiological indexes of Tamarix chinensis seedlings under normal watering.
As shown in Table 5, under persistent drought, Na+ was significantly and positively correlated with malondialdehyde (P< 0.01), SOD activity (P< 0.01), proline (P< 0.01), soluble protein (P< 0.01), relative conductivity (P< 0.01), soluble sugar (P< 0.01), and negatively correlated with leaf relative water content (P< 0.01) and soluble starch (P< 0.01). MDA was significantly and positively correlated with RC (P< 0.01), SOD (P< 0.01), Pro (P< 0.01), SP (P< 0.01), SS (P< 0.01), and negatively correlated with RWC (P< 0.01), ST (P< 0.01) and NSC (P< 0.01). ST was significantly and negatively correlated with RC (P< 0.01), SOD (P< 0.01), Pro (P< 0.01), SP (P< 0.01) and SS (P< 0.01); SS was positively correlated with SOD (P< 0.05) and negatively correlated with RWC (P< 0.01).
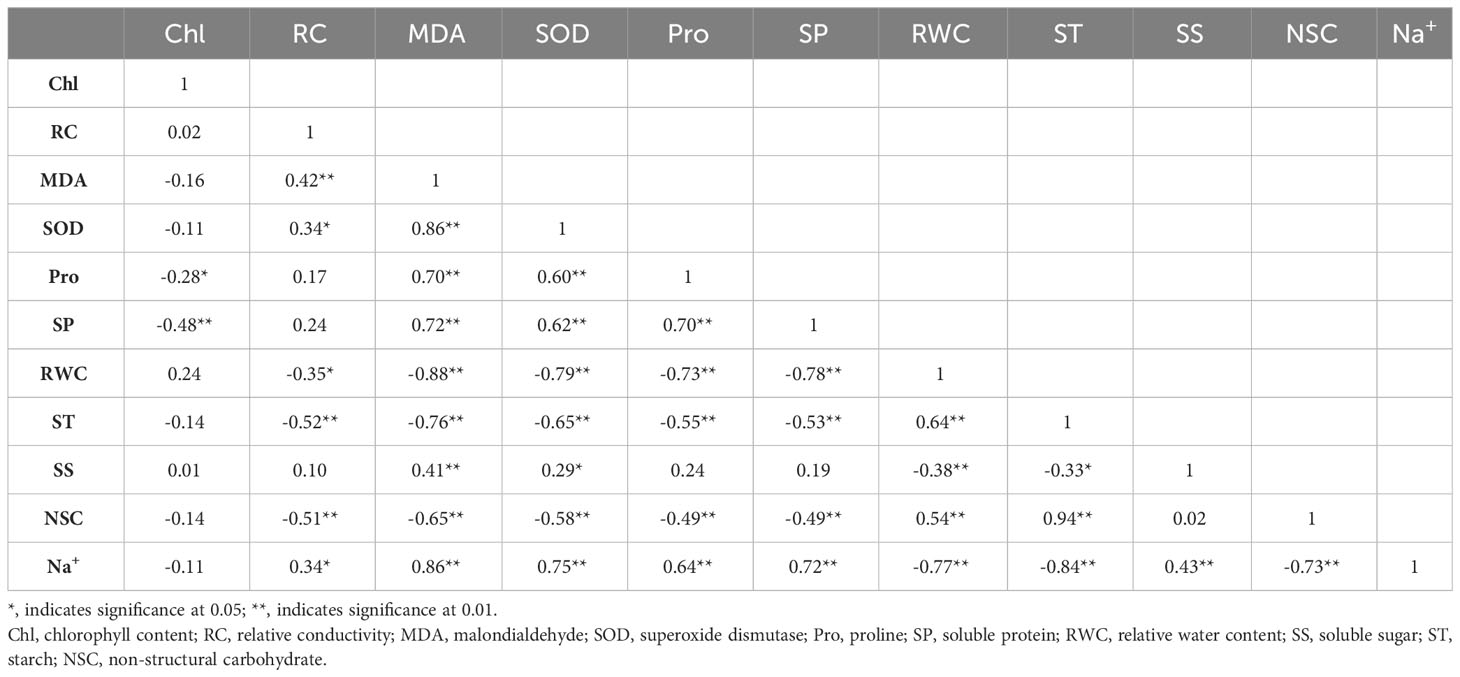
Table 5 Correlation analysis of physiological indexes of Tamarix chinensis seedlings in response to salt under persistent drought. .
Three principal components were obtained under persistent drought stress, with contributions of 35.58%, 28.75%, and 16.49% in the order, and the cumulative contribution was 80.82% (Table 6). The first principal component consisted of NSC, starch, relative conductivity, malondialdehyde, and SOD, reflecting the integrity of the cell membrane structure. The second principal component consisted of soluble protein, chlorophyll, proline, and relative water content, reflecting the changes in chlorophyll.
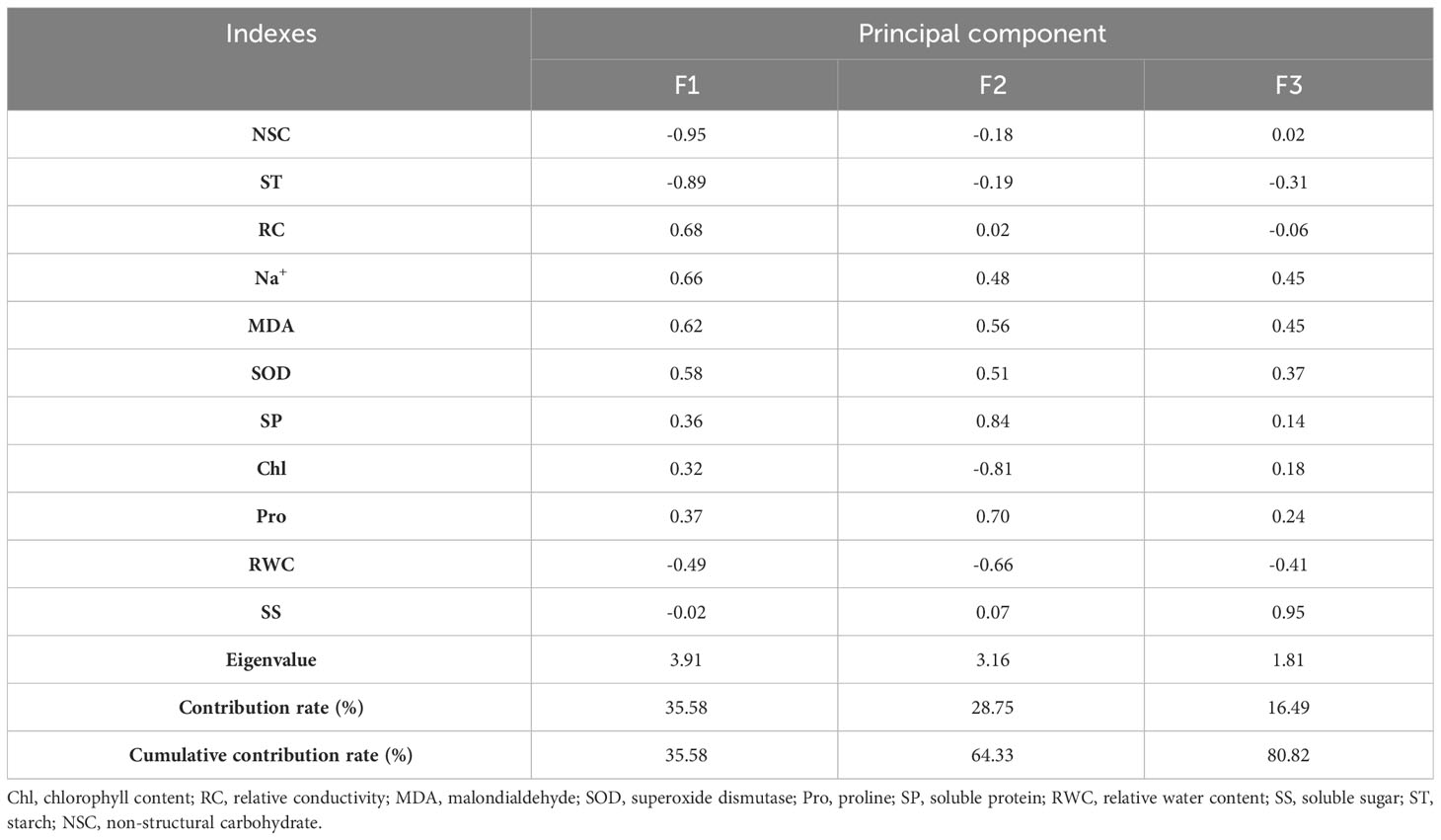
Table 6 Principal component analysis of physiological indicators of Tamarix chinensis seedlings under persistent drought conditions.
Under persistent waterlogging (Table 7), sodium ions were significantly and positively correlated with total chlorophyll (P< 0.01), relative conductivity (P< 0.01), SOD activity (P< 0.01), proline (P< 0.01), soluble sugars (P< 0.01) and NSC content (P< 0.01), and negatively correlated with MDA (P< 0.05). MDA was negatively correlated with Pro (P< 0.05) and SS (P< 0.05), and positively correlated with SP (P< 0.05) and ST (P< 0.01). SS was positively correlated with Pro (P< 0.01) and RWC (P< 0.05), and negatively correlated with SP (P< 0.05).
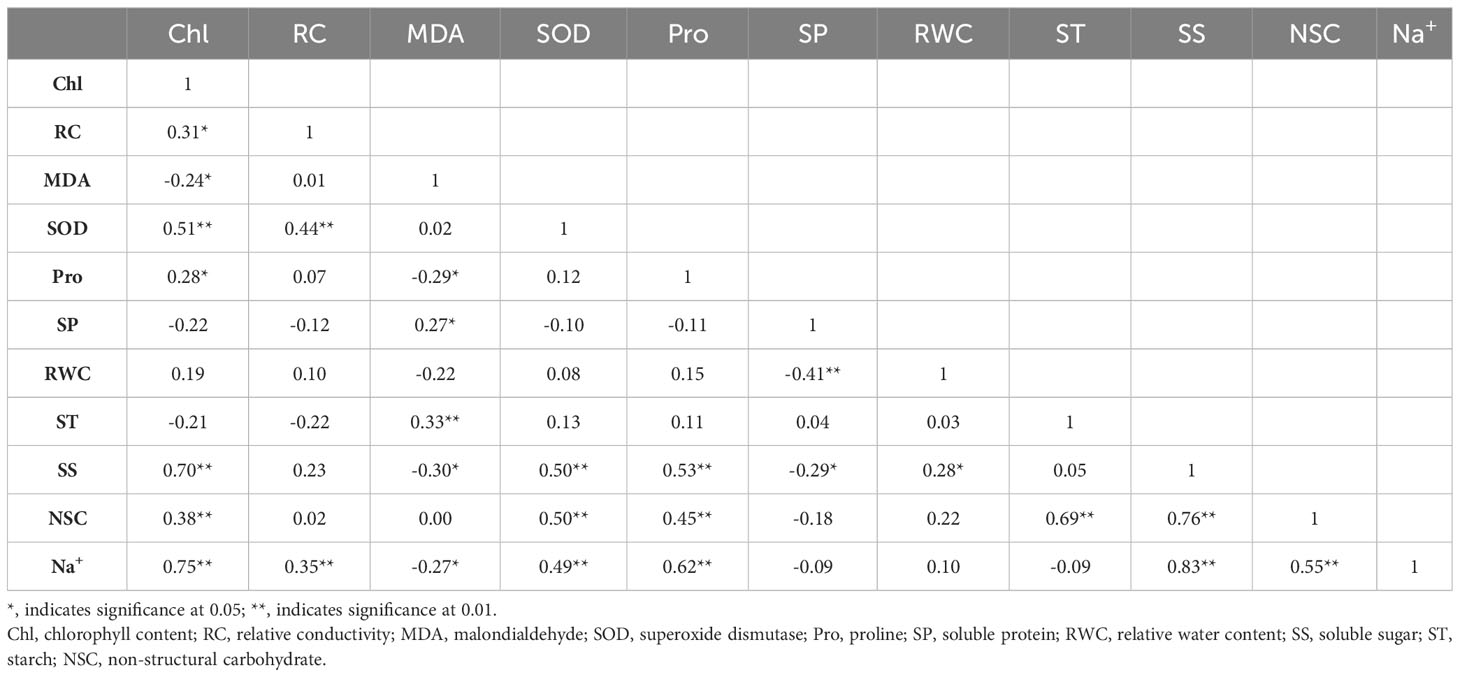
Table 7 Correlation analysis of physiological indexes of Tamarix chinensis seedlings in response to salt under persistent waterlogging.
Four principal components were obtained under persistent waterlogging, with contributions of 27.96%, 18.19%, 16.98%, and 14.58% in the order, with a cumulative contribution of 77.70% (Table 8). The first principal component consisted of sodium ions, soluble sugars, proline, chlorophyll, and malondialdehyde, which reflected the changes in chlorophyll. The second principal component consisted of relative conductivity and SOD, reflecting the integrity of the cell membrane structure.
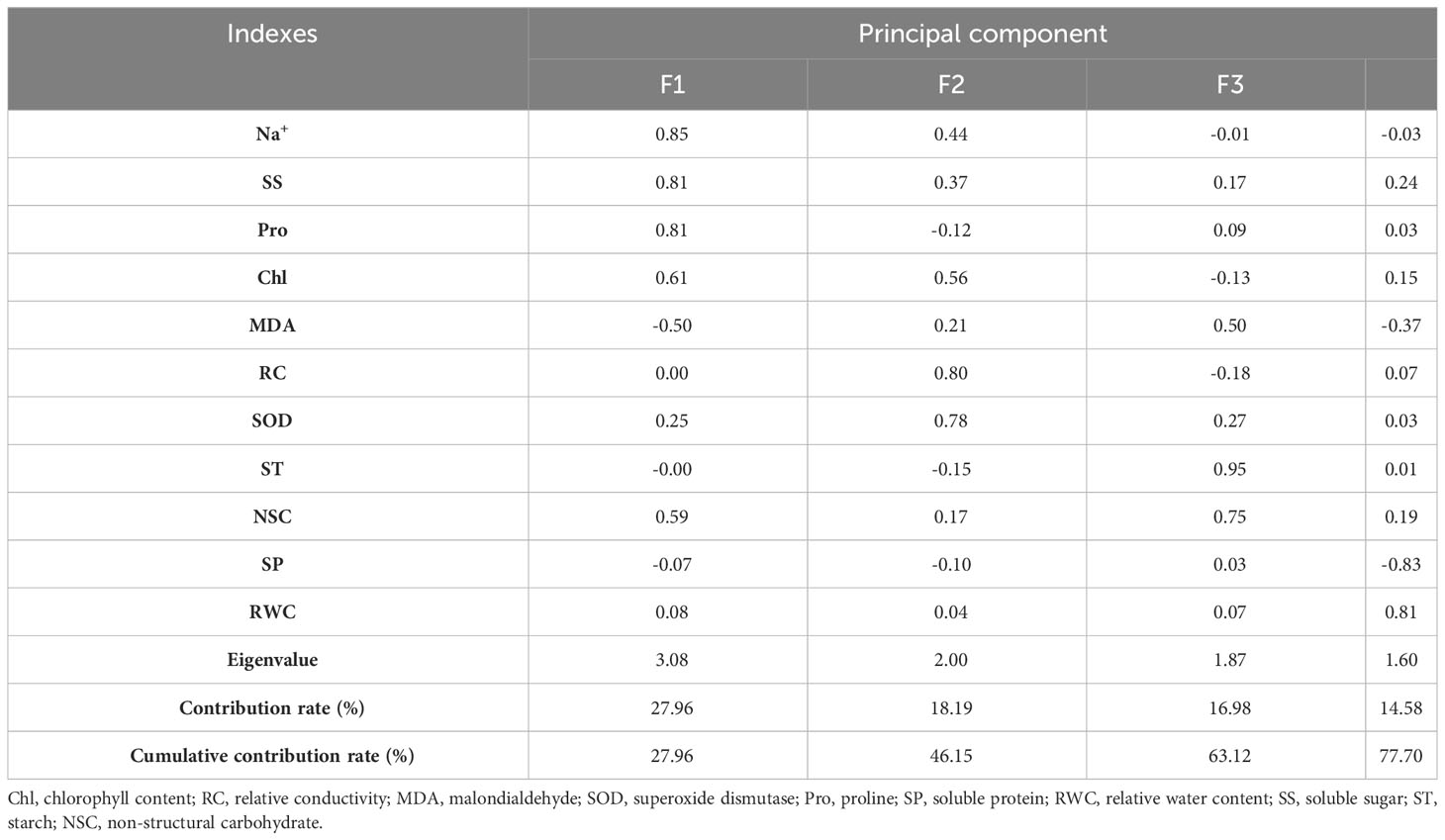
Table 8 Principal component analysis of physiological indicators of Tamarix chinensis seedlings under persistent waterlogging conditions.
In coastal wetland ecosystems, plants are combinedly affected by soil moisture and salinity. Soil water change regulating the soil salt content obviously influence the plant adaptability. In the present study, we found that the physiological adaptability of T. chinensis siginificantly changed along the soil moisture. The combined roles of persistent drought and soil salinity further intensified the inhibition of T. chinensis growth.
The relative seedling height and ground diameter growth rates of T. chinensis seedlings first increased and then decreased with increasing salt concentration. Studies on S. salsa also found that low salt promoted their growth while high salt significantly inhibited it (Jia et al., 2018). The relative seedling height, ground diameter growth rate and chlorophyll content of seedlings were the highest under persistent waterlogging, indicating that T. chinensis is highly tolerant to flooding. Chlorophyll is an extremely important pigment affecting photosynthesis, and the level of content reflects the photosynthetic capacity of the plant (Muchate et al., 2016). The changes in chlorophyll were the first principal component under normal watering and persistent waterlogging and the second principal component under persistent drought, suggesting that the maintenance of chlorophyll is an important physiological basis for its adaptation to coastal saline sites, in agreement with the results of the study on Cakile maritima (Farhat et al., 2021). Under persistent drought stress, the higher the salt concentration and longer the treatment time, the lower the chlorophyll content, with the lowest chlorophyll content of only 0.08 mg/g at the 21st d salt concentration of 20‰, and almost all of the T. chinensis leaves lost their green color. In dry seasons, soil salinity increases in coastal saline sites (Souid et al., 2018). We similarly found that soil treatment gradually increased with the increase of persistent stress time in our study, which was an important reason for the aboveground mortality of T. chinensis seedlings in the experiment. With global climate change, T. chinensis natural forests may experience prolonged drought, which will threaten the maintenance of their populations in coastal wetlands.
The salt treatment caused osmotic stress to T. chinensis seedlings, and the leaf relative water content under high salt treatment was lower than that of low salt treatment, which was related to the difficulty of water uptake by the soil due to low osmotic potential caused by high salt concentration in the soil. A decrease in leaf relative water content with increasing salt concentration was also found in Robinia pseudoacacia (Mao et al., 2016) and Lonicera japonica (He et al., 2021). The decrease in leaf relative water content indicates that an increase in water deficit occurs in them (Mao et al, 2016). Increased proline (Nxele et al., 2017; Per et al., 2017) and soluble protein content (Arbelet-Bonnin et al., 2020) are important ways to increase the osmoregulatory capacity of plants under salt stress. Proline and soluble protein content in T. chinensis in this study gradually increased with increasing salt concentration, which suggested that both were involved in osmoregulation. In this paper, we found that the relative water content of T. chinensis leaves under three moisture conditions was lowest under persistent drought and highest under persistent waterlogging. A regular leaf water content could ensure normal physiological metabolism (Mao et al., 2016), which is an important reason for the highest relative growth rate under persistent waterlogging. Under the three different water conditions, the content of proline and soluble protein increased with increasing treatment time, but they were the highest under persistent drought stress. It wad suggested that although proline increased significantly under drought and salt stress, the osmoregulatory capacity was limited (Nxele et al., 2017). Arbelet-Bonnin et al. (2020) concluded that the increase in proline was related to the tolerant ability of halophyte C. maritima to salt stress. Benjamin et al. (2019), on the other hand, concluded that there were differences in osmoregulatory substances in different saline plants. Correlation analysis showed that the relative leaf water content was significantly negatively correlated with proline and soluble protein. This suggested that osmoregulatory substances, although significantly increased under persistent drought, were not sufficient to counteract water deficit due to stress, which indicated that there might be other substances involved in the process.
The relative conductivity and malondialdehyde content of salt concentration 20‰ in this study was higher than other salt treatments, indicating that heavy salt stress severely damaged cell membranes (Muchate et al., 2016). Malondialdehyde content was highest under persistent drought and increased rapidly with increasing treatment time, while it did not change much under normal watering and persistent waterlogging. The MDA content of halophyte L. delicatulum was also highest in the dry season (Souid et al., 2018). Studies on S. salsa have shown that cell membrane persecution is more severe under combined salt-drought effects (Jia et al., 2018). The full use of sodium ions to adapt to salt environments is a common performance of saline plants (Muchate et al., 2016; He et al., 2021). In this paper, it was found that T. chinensis absorbed large amounts of sodium ions under all moisture conditions. However, correlation analysis showed that malondialdehyde and sodium ions were significantly and positively correlated with malondialdehyde under persistent drought, but not under normal watering and persistent waterlogging, suggesting that increased sodium ions under drought stress was an important factor contributing to cell membrane disruption in T. chinensis. Cell membrane disruption under persistent drought was the first principal component and was the main mode leading to severe stress in T. chinensis. Studies on Sorghum bicolor have also identified cell membrane disruption as an important cause of its cell death under drought and salt stress (Nxele et al., 2017). In response to oxidative stress damage to cell membranes, T. chinensis SOD activity increased with increasing soil salt content. Under persistent drought, SOD activity was significantly higher than that under normal watering and persistent waterlogging, indicating that SOD is an important way to eliminate superoxide ions under drought stress (Souid et al., 2018). It has been shown that proline (Per et al., 2017) and other osmoregulatory substances are also important ways to eliminate oxygen radicals (Khoshbakht et al., 2018; Arbelet-Bonnin et al., 2020). Correlation analysis showed that under persistent drought MDA was significantly and positively correlated with Pro, SP, and SS, in addition to SOD. Thus, protective enzymes and osmoregulatory substances together protected cell membranes under persistent drought. As a whole, MDA was an important indicator in response to the stress suffered by T. chinensis.
Soluble sugars, starch and NSC increased first and then decreased with increasing salt concentration under normal water and persistent waterlogging, and gradually decreased under persistent drought. Arbelet-Bonnin et al. (2020) suggested that sugar content decreased under high salt stress due to the inhibition of photosynthesis. It has been shown that NSC increases under adversity suggesting plants with strong resilience (Gao et al., 2021). The second principal component of the highest soluble sugars and NSC under normal watering was correlated with NSC changes, suggesting that NSC accumulation was an important way of its adaptation to salt stress. The persistent increase in NSC of T. chinensis under persistent waterlogging also indicated its strong adaptation to waterlogging. In this paper, we found that the total chlorophyll content decreased under persistent drought. Therefore, the decrease in photosynthesis is an important reason for the decrease in NSC under persistent drought. The significantly negative correlation between SS and ST under normal watering and persistent drought indicated that there was a conversion between them. This further supported the founding that the interconversion between soluble sugars and starches was an important way to cope with the external environment (McDowell, 2011). ST was significantly negatively correlated with Pro and SS under normal watering and with SOD, Pro and SS under persistent drought. These suggested that the breakdown of starch to soluble sugars under stress, which provided a carbon shelf for nitrogenous compounds, was an important way to adapt to adversity (Hartmann and Trumbore, 2016). The reduction in soluble sugar content of T. chinensis under persistent drought indicated that depletion was much greater than photosynthetic production, which maybe one of important reasons for the decrese in NSC. After drought stress was eliminated, NSC content was significantly reduced in trees that did not die and was used for tissue repair (Tomasella et al., 2019). Therefore, the high depletion of NSC under persistent drought was detrimental to the survival of T. chinensis in coastal saline sites.
This study found that the seedling growth of T. chinensis is significantly inhibited with a soil salinity above 20‰. Soil moisture conditions significantly influenced the physiological processes of tolerance of T. chinensis in different soil salinity. Under normal watering and persistent waterlogging, the physiology of T. chinensis was less affected under different soil salinity, especially the highest relative growth rate under persistent waterlogging. These results illustrated that T. chinensis in persistent waterlogging conditions could tolerate different salt stress. Under persistent drought stress, with the decrease of soil water content and the significant increase of soil salinity, T. chinensis suffered from both drought and salt stress, the relative water content in the body was significantly reduced, chlorophyll and cell membrane suffered serious damage, and although osmoregulation and protective enzyme activities were significantly increased, they were not sufficient to offset the damage caused by water deficit, which eventually led to aboveground death. Under the regular water condition of normal and persistent waterlogging, sodium ion was an important osmoregulatory substance, while under persistent drought it is a key factor leading to cell membrane disruption. NSC significantly increased under normal and persistent waterlogging while significantly decreased under persistent drought. In this process, starch played an important role in carbon storage function and was an important carbon source for various physiological maintenance. Malondialdehyde correlated well with various indicators under persistent drought and was a good indicator to reflect the tolerance under drought stress. With the intensification of global climate change, the change of temperature and precipitation will intensify the hydrological changes of coastal wetlands. It is of great significance to strengthen the research on the dynamic monitoring of precipitation and water-salt and their impacts on dominant plants in the Yellow River Delta in the future, which is meaningful for predicting the dynamic evolution of vegetation and improving the management level.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
PM and BC designed and reviewed the manuscript. QL, YP and KW collected the data. QL drafted the manuscript. YP analyzed the date. RN and XH reviewed and improved the manuscript. All authors contributed to the article and approved the submitted version.
This research was funded by Major Scientific Technological Innovation Projects of Shandong Province (2017CXGC0316), China Natural Science Foundation (31770668), Central Finance Forestry Reform and Development Fund (2020TG08) and Germplasm resources nursery project of saline alkali tolerant tree species in the Yellow River Delta (2019-370505-05-03-035206).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Arbelet-Bonnin D., Blasselle C., Palm E. R., Redwan M., Bouteau F. (2020). Metabolism regulation during salt exposure in the halophyte cakile maritima. Environ. Exp. Bot. 177, 104075. doi: 10.1016/j.envexpbot.2020.104075
Benjamin J. J., Lucini L., Jothiramshekar S., Parida A. (2019). Metabolomic insights into the mechanisms underlying tolerance to salinity in different halophytes. Plant Physiol. Bioch. 135, 528–545. doi: 10.1016/j.plaphy.2018.11.006
Borzouei A., Shalmani M. A. M., Eskandari A. (2020). Effects of salt and nitrogen on physiological indices and carbon isotope discrimination of wheat cultivars in the northeast of Iran. J. Integr. Agr. 19 (3), 656–667. doi: 10.1016/S2095-3119(19)62629-8
Buffington K. J., Goodman A. C., Freeman C. M., Thorne K. M. (2020). Testing the interactive effects of flooding and salinity on tidal marsh plant productivity. Aquat. Bot. 164, 103231. doi: 10.1016/j.aquabot.2020.103231
Cong P., Chen K., Qu L., Han J. (2019). Dynamic changes in the wetland landscape pattern of the Yellow River Delta from 1976 to 2016 based on satellite data. Chin. Geogra. Sci. 29 (3), 372–381. doi: 10.1007/s11769-019-1039-x
Cui B., Yang Q., Yang Z., Zhang K. (2009). Evaluating the ecological performance of wetland restoration in the Yellow River Delta, China. Ecol. Eng. 35, 1090–1103. doi: 10.1016/j.ecoleng.2009.03.022
Cui G., Zhang Y., Zhang W., Lang D., Lang D., Zhang X., Li Z. (2019). Response of carbon and nitrogen metabolism and secondary metabolites to drought stress and salt stress in plants. J. Plant Biol. 62, 387–399. doi: 10.1007/s12374-019-0257-1
Dasgupta S., Laplante B., Meisner C., Wheeler D., Yan J. (2009). The impact of sealevel rise on developing countries: a comparative analysis. Clim. Change. 93, 379–388. doi: 10.1007/s10584-008-9499-5
da Silva H. A., , de Oliveira D. F. A., Avelino A. P., de Macêdo C. E. C., Barros-Galvão T., Voigt E.L. (2019). Salt stress differentially regulates mobilisation of carbon and nitrogen reserves during seedling establishment of Pityrocarpa moniliformis. Plant Biol. J. 21, 1110–1118. doi: 10.1111/plb.13017
Dietze M. C., Sala A., Carbone M. S., Czimczik C. I., Mantooth J. A., Richardson A. D., et al. (2014). Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 65, 667–687. doi: 10.1146/annurev-arplant-050213-040054
Farhat N., Kouas W., Braun H. P., Debez A. (2021). Stability of thylakoid protein complexes and preserving photosynthetic efficiency are crucial for the successful recovery of the halophyte Cakile maritima to high salinity. Plant Physiol. Bioch. 166, 177–190. doi: 10.1016/j.plaphy.2021.05.044
Gao Y., Sun Y., Ou Y., Zheng X., Feng Q., Zhang H., et al. (2021). Pretreating poplar cuttings with low nitrogen ameliorates salt stress responses by increasing stored carbohydrates and priming stress signaling pathways. Ecotox. Environ. Safe. 225, 112801. doi: 10.1016/j.ecoenv.2021.112801
Gao Y., Xia J., Chen Y., Zhao Y., Kong Q., Lang Y. (2017). Effects of extreme soil water stress on photosynthetic efficiency and water consumption characteristics of Tamarix chinensis in China’s Yellow River Delta. J. For. Res. 28 (3), 491–501. doi: 10.1007/s11676-016-0339-6
Ghaderi N., Hatami M., Mozafari A., Siosehmardeh A. (2018). Change in antioxidant enzymes activity and some morpho-physiological characteristics of strawberry under long-term salt stress. Physiol. Mol. Biol. Plants 24 (5), 833–843. doi: 10.1007/s12298-018-0535-2
Guan B., Yu J., Hou A., Han G., Wang G., Qu F., et al. (2017). The ecological adaptability of Phragmites australis to interactive effects of water level and salt stress in the Yellow River Delta. Aquat. Ecol. 51, 107–116. doi: 10.1007/s10452-016-9602-3
Guan B., Yu J., Wang X., Fu Y., Kan X., Lin Q., et al. (2011). Physiological responses of halophyte suaeda salsa to water table and salt stresses in coastal wetland of yellow river delta. Clean. – Soil. Air. Water 39 (12), 1029–1035. doi: 10.1002/clen.201000557
Guo L., Cao B., Mao P., Li Z., Hao M., Wang T., et al. (2021). Fine root vertical-seasonal distribution of Robinia pseudoacacia in relation to abiotic factors in a chronosequence in coastal saline alkali land of the Yellow River Delta, China. Turk. J. Agric. For. 45, 750–765. doi: 10.3906/tar-2105-100
Guo Q. X., Wu X. Y., Korpelainen H., Li C. Y. (2020). Stronger intraspecific competition aggravates negative effects of drought on the growth of Cunninghamia lanceolata. Environ. Exp. Bot. 175, 104042. doi: 10.1016/j.envexpbot.2020.104042
Hartmann H., Trumbore S. (2016). Understanding the roles of nonstructural carbohydrates in forest trees-From what we can measure to what we want to know. New Phytol. 211, 386–403. doi: 10.1111/nph.13955
He W., Yan K., Zhang Y., Bian L., Mei H., Han G. (2021). Contrasting photosynthesis, photoinhibition and oxidative damage in honeysuckle (Lonicera japonica Thunb.) under iso-osmotic salt and drought stresses. Environ. Exp. Bot. 182, 104313. doi: 10.1016/j.envexpbot.2020.104313
Hoch G., Richter A., Korner C. (2003). Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 26 (7), 1067–1081. doi: 10.1046/j.0016-8025.2003.01032.x
Hou W., Zhang R., Xi Y., Liang S., Sun Z. (2020). The role of waterlogging stress on the distribution of salt marsh plants in the Liao River estuary wetland. Glob. Ecol. Conserv. 23, e01100. doi: 10.1016/j.gecco.2020.e01100
Irshad A., Rehman R. N. U., Abrar M. M., Saeed Q., Sharif R., Hu T. (2021). Contribution of rhizobium-legume symbiosis in salt stress tolerance in Medicago truncatula evaluated through photosynthesis, antioxidant enzymes, and compatible solutes accumulation. Sustainability 13, 3369. doi: 10.3390/su13063369
Jia J., Huang C., Bai J., Zhang G., Zhao Q., Wen X. (2018). Effects of drought and salt stresses on growth characteristics of euhalophyte Suaeda salsa in coastal wetlands. Phys. Chem. Earth 103, 68–74. doi: 10.1016/j.pce.2017.01.002
Jiang D., Fu X., Wang K. (2013). Vegetation dynamics and their response to freshwater inflow and climate variables in the Yellow River Delta, China. Quat. Int. 304, 75–84. doi: 10.1016/j.quaint.2012.10.059
Jiao L., Zhang Y., Sun T., Yang W., Shao D., Zhang P., et al. (2021). Spatial analysis as a tool for plant population conservation: a case study of Tamarix chinensis in the Yellow River Delta, China. Sustainability 13, 8291. doi: 10.3390/su13158291
Khoshbakht D., Asghari M. R., Haghighi M. (2018). Influence of foliar application of polyamines on growth, gas-exchange characteristics, and chlorophyll fluorescence in Bakraii citrus under saline conditions. Photosynthetica 56 (2), 731–742. doi: 10.1007/s11099-017-0723-2
Kiarash J. G., Amin B., Mansour K., Foad F. (2020). Combined effects of salinity and drought on physiological and biochemical characteristics of pistachio rootstocks. Sci. Hortic-amsterdam. 261, 108970. doi: 10.1016/j.scienta.2019.108970
Kozlowski T. T. (1992). Carbohydrate sources and sinks in woody plants. Bot. Rev. 58 (2), 107–222. doi: 10.1007/BF02858600
Li X., Xia J., Zhao X., Chen Y. (2019). Effects of planting Tamarix chinensis on shallow soil water and salt content under different groundwater depths in the Yellow River Delta. Geoderma 335, 104–111. doi: 10.1016/j.geoderma.2018.08.017
Liu Z., Fagherazzi S., Ma X., Xie C., Cui B. (2020a). Efficient tidal channel networks alleviate the drought-induced die-off of salt marshes: Implications for coastal restoration and management. Sci. Total. Environ. 749, 141493. doi: 10.1016/j.scitotenv.2020.141493
Liu Z., Fagherazzi S., Ma X., Xie C., Li J., Cui B. (2020b). Consumer control and abiotic stresses constrain coastal saltmarsh restoration. J. Environ. Manage. 274, 111110. doi: 10.1016/j.jenvman.2020.111110
Liu J., Xia J., Fang Y., Li T., Liu J. T. (2014). Effects of salt-drought stress on growth and physiobiochemical characteristics of Tamarix chinensis seedlings. Sci. World J. 6, 765–840. doi: 10.1155/2014/765840
Lu Q., Kang L., Shao H., Zhao Z., Chen Q., Bi X., et al. (2016). Investigating marsh sediment dynamics and its driving factorsin Yellow River delta for wetland restoration. Ecol. Eng. 90, 307–313. doi: 10.1016/j.ecoleng.2016.01.059
Maaloul S., Abdellaoui R., Mahmoudi M., Bouhamda T., Bakhshandeh E., Boughalleb F. (2021). Seasonal environmental changes affect differently the physiological and biochemical responses of two limonium species in sabkha biotope. Physiol. Plantarum. 172, 2112–2128. doi: 10.1111/ppl.13446
Macneill G. J., Mehrpouyan S., Minow M. A. A., Patterson J. A., Tetlow I. J., Emes M. J. (2017). Starch as a source, starch as a sink: the bifunctional role of starch in carbon allocation. J. Exp. Bot. 68, 4433–4453. doi: 10.1093/jxb/erx291
Mao P., Zhang Y., Cao B., Guo L., Shao H., Cao Z., et al. (2016). Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total. Environ. 568, 118–123. doi: 10.1016/j.scitotenv.2016.06.012
McDowell N. G. (2011). Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 155, 1051–1059. doi: 10.1104/pp.110.170704
Middleton B. A., Kleinebecker T. (2012). “The effects of climate-change-induced drought and freshwater wetlands,” in Global change and the function and distribution of wetlands. Global change ecology and wetlands, vol. 1 . Ed. Middleton B. (Dordrecht: Springer).
Muchate N. S., Nikalje G. C., Rajurkar N. S., Suprasanna P., Nikam T. D. (2016). Physiological responses of the halophyte sesuvium portulacastrum to salt stress and their relevance for saline soil bio-reclamation. Flora 224, 96–105. doi: 10.1016/j.flora.2016.07.009
Mushtaq Z., Faizan S., Gulzar B. (2021). Inoculation of rhizobium alleviates salinity stress through modulation of growth characteristics, physiological and biochemical attributes, stomatal activities and antioxidant defence in Cicer arietinum L. J. Plant Growth Regul. 40, 2148–2163. doi: 10.1007/s00344-020-10267-1
Nxele X., Klein A., Ndimba B. K. (2017). Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 108, 261–266. doi: 10.1016/j.sajb.2016.11.003
O’Brien M., Leuzinger , S., Philipson C., Tay J., Hector A. (2014). Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nature Clim. Change 4, 710–714. doi: 10.1038/nclimate2281
Ou C., Zhang M., Yao X., Yang L., Liu Y., Qu C. (2019). Effects of melatonin on growth, ion absorption and photosynthesis of Toona sinensis seedlings under salt stress. Acta Bot. Boreal.–Occident. Sin. 39 (12), 2226–2234. doi: 10.7606/j.issn.1000-4025.2019.12.2226
Per T. S., Khan N. A., Reddy P. S., Masood A., Hasanuzzaman M., Khan M., et al. (2017). Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: phytohormones, mineral nutrients and transgenics. Plant Physiol. Bioch. 115, 126–140. doi: 10.1016/j.plaphy.2017.03.018
Rahmstorf S. (2007). A semi-empirical approach to projecting future sea-level rise. Science 315, 368–370. doi: 10.1126/science.1135456
Rangani J., Panda A., Patel M., Parida A. K. (2018). Regulation of ros through proficient modulations of antioxidative defense system maintains the structural and functional integrity of photosynthetic apparatus and confers drought tolerance in the facultative halophyte Salvadora persica L. J. Photoch. Photobio. B. 189, 214–233. doi: 10.1016/j.jphotobiol.2018.10.021
Sala A., Woodruff D. R., Meinzer F. C. (2012). Carbon dynamics in trees: feast or famine. Tree Physiol. 32 (6), 764–775. doi: 10.1093/treephys/tpr143
Schuerch M., Spencer T., Temmerman S., Kirwan M. L., Wolff C., Lincke D., et al. (2018). Future response of global coastal wetlands to sea-level rise. Nature 561, 231–234. doi: 10.1038/s41586-018-0476-5
Smith T., Boers N. (2023). Global vegetation resilience linked to water availability and variability. Nat. Commun. 14, 498. doi: 10.1038/s41467-023-36207-7
Smith S. M., Lee K. D. (2015). The influence of prolonged flooding on the growth of Spartina alterniflora in Cape Cod (Massachusetts, USA). Aquat. Bot. 127, 53–56. doi: 10.1016/j.aquabot.2015.08.002
Souid A., Bellani L., Magné C., Zorrig W., Smaoui A., Abdelly C., et al. (2018). Physiological and antioxidant responses of the sabkha biotope halophyte Limonium delicatulum to seasonal changes in environmental conditions. Plant Physiol. Bioch. 123, 180–191. doi: 10.1016/j.plaphy.2017.12.008
Sun B. Y., Jiang M., Han G., Zhang L., Zhou J., Bian C., et al. (2022). Experimental warming reduces ecosystem resistance and resilience to severe flooding in a wetland. Sci. Adv. 8 (4), eabl9526. doi: 10.1126/sciadv.abl9526
Taherkhani M., Vitousek S., Barnard P. L., Frazer N., Anderson T. R., Fletcher C. H. (2020). Sea-level rise exponentially increases coastal flood frequency. Sci. Rep. 10, 6466. doi: 10.1038/s41598-020-62188-4
Tomasella M., Casolo V., Aichner N., Petruzzellis F., Savi T., Trifilò P., et al. (2019). Non-structural carbohydrate and hydraulic dynamics during drought and recovery in Fraxinus ornus and Ostrya carpinifolia saplings. Plant Physiol. Bioch. 145, 1–9. doi: 10.1016/j.plaphy.2019.10.024
Watson E. B., Szura K., Wigand C., Raposa K. B., Blount K., Cencer M. (2016). Sea level rise, drought and the decline of Spartina patens in New England marshes. Biol. Conserv. 196, 173–181. doi: 10.1016/j.biocon.2016.02.011
Xia J., Zhang S. Y., Zhang G. C., Xu J. W., Liu J. T., Li C. R. (2013). Growth dynamics and soil water ecological characteristics of Tamarix Chinensis Lour. forests with two site types in coastal wetland of Bohai golf. J. Food Agric. Environ. 11, 1492–1498.
Xia J., Zhao X., Ren J., Lang Y., Qu F., Xu H. (2017). Photosynthetic and water physiological characteristics of Tamarix chinensis under different groundwater salinity conditions. Environ. Exp. Bot. 138, 173–183. doi: 10.1016/j.envexpbot.2017.03.015
Keywords: dominant species, relative growth rate, chlorophyll, malondialdehyde, nonstructural carbohydrates
Citation: Mao P, Lin Q, Pang Y, Wang K, Ni R, Han X and Cao B (2023) Eco-physiological response mechanism of Tamarix chinensis to soil water changes in coastal wetlands of the Yellow River Delta. Front. Mar. Sci. 10:1231928. doi: 10.3389/fmars.2023.1231928
Received: 31 May 2023; Accepted: 08 August 2023;
Published: 22 August 2023.
Edited by:
Laibin Huang, University of California, Davis, United StatesReviewed by:
Peng Zhao, Hainan University, ChinaCopyright © 2023 Mao, Lin, Pang, Wang, Ni, Han and Cao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Banghua Cao, Y2FvYmFuZ2h1YUAxMjYuY29t
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.