 Jingjing Zhang1
Jingjing Zhang1 Hongliang Li1,2,3*
Hongliang Li1,2,3* Martin G. Wiesner1,4
Martin G. Wiesner1,4 Lihua Ran1
Lihua Ran1 Xingju He1
Xingju He1 Guangxi Chi1,5
Guangxi Chi1,5 Xinyang Wang1,6
Xinyang Wang1,6 Jinping Yu1
Jinping Yu1 Jianfang Chen1,7*
Jianfang Chen1,7*- 1Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou, China
- 2Southern Marine Science and Engineering Guangdong Laboratory, Zhuhai, China
- 3Observation and Research Station of Yangtze River Delta Marine Ecosystems, Ministry of Natural Resources, Zhoushan, China
- 4Institute of Geology, University of Hamburg, Hamburg, Germany
- 5College of Oceanography, Hohai University, Nanjing, China
- 6School of Oceanography, Shanghai Jiao Tong University, Shanghai, China
- 7State Key Laboratory of Satellite Ocean Environment Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou, China
Accurately assessing the capacity of the modern ocean to photosynthetically fix and sequester atmospheric CO2, termed the biological carbon pump (BCP), is a key component in studies on the marine carbon cycle and the global climate system. Particulate organic carbon (POC) flux into the ocean interior is an important indicator of the BCP strength, and it can be directly measured by sediment traps on time scales from days to years. This study has been conducted in the South China Sea (SCS) for over three decades. The SCS is one of the largest tropical marginal seas, located in the Asian monsoon region with frequent occurrence of dynamic physical processes and anthropogenic perturbations. It hosts an ideal natural laboratory to investigate the response of the BCP to multiscale physical processes. In this mini review, we briefly introduce the study history of mooring sediment traps in the SCS, synthesize the processes that regulate the temporal variability in mesopelagic POC flux, and how it is sensitive to climate changes. The time-series characteristics of the POC flux are clearly linked to primary production, as well as the key physical processes in the upper layer. The seasonal East Asian monsoon, intraseasonal eddies, aerosol deposition and interannual El Niño Southern Oscillation (ENSO) events are the main controlling factors over weekly to yearly timescales. Together, they suggest that the multiscale physical forcing in the upper layer regulates the mesopelagic POC export flux by controlling nutrient supplementation and subsequent POC production.
Introduction
The biological carbon pump (BCP) is a suite of processes collectively leading to the net uptake of atmospheric CO2. It transfers dissolved inorganic carbon to organic carbon through photosynthesis by phytoplankton in the sunlit surface ocean at a rate of ~50 GtC/yr (Chavez et al., 2011; Johnson and Bif, 2021), and then delivers part of the produced particulate organic carbon (POC) into the ocean interior (Sigman and Boyle, 2000). When POC settles to depths below the mesopelagic zone (i.e., beyond about 1000 m), it can be stored and isolated from the atmospheric carbon pool for time scales of hundreds of years or even longer (Kwon et al., 2009). Without these processes, the atmospheric CO2 content would be 200 ppm higher than it is today (Parekh et al., 2006; Henson et al., 2019). The biogenic particles that sink out of the surface layer not only fuel the dark ocean and benthic ecosystem but also play a vital role in the distributions of oxygen, carbon and nutrients. Thus, it is important to develop an understanding of the layout of global deep ocean POC fluxes over a range of spatial and temporal scales.
In this context, a prevalent mooring sediment trap was invented in the 1970s to intercept sinking particles settling into the ocean interior for up to a year based on a consecutive time series sequence to better understand its composition, transport and transformation in the oceans (Honjo et al., 1980; Deuser et al., 1981). This device not merely provides a direct measurement of sinking particle flux over a range of time scales but also supplies intact time-series samples for further multidisciplinary analysis, such as chemical components (Lee, 2019) and plankton taxonomy (Takahashi and Honjo, 1981; Takahashi, 1986) in sinking particles. Therefore, an internationally coordinated global array of sediment traps was proposed as a major undertaking of the Joint Global Ocean Flux Study (JGOFS) project, which aimed to more accurately assess and better understand the processes controlling regional to global and seasonal to interannual carbon fluxes among the atmosphere, surface ocean and ocean interior, as well as their sensitivities to climate changes. Overall, hundreds of sediment traps have been widely deployed in different ocean regions worldwide for multidisciplinary objectives since 1983. A comprehensive understanding of global POC fluxes to the ocean interior as well as associated biogeochemical parameters has benefited from these projects. Clear imparities have been discerned in POC fluxes between regions and basins, much greater emphasis was placed on marginal seas for the disproportionate importance of POC flux within their limited sea surface area (Muller-Karger et al., 2005; Honjo et al., 2008).
The South China Sea (SCS) is one of the largest world’s semi-enclosed marginal seas, situated in the East Asian monsoon system, between the Western Pacific Warm Pool and Tibet Plateau, with only one major deep channel, the Luzon Strait (>2200 m), to effectively exchange with the western Pacific. It consists vast shelves on the north and south sides, steep slopes on the west and east sides, and deep basins with a maximum depth reaching 4700 m in the center (Ning et al., 2004). The basin scale circulation (Wyrtki, 1961) and physical-biogeochemical condition of the SCS are mainly governed by the strong forcing of alternating monsoons and sensitive to climate variations (Liu et al., 2002; Liu et al., 2013; Ning et al., 2004). Besides, randomly occurring mesoscale eddies (Xiu et al., 2010; Xiu and Chai, 2011), dust deposition (Wang et al., 2012), as well as the inter-annual El Niño (Liu et al., 2013) would also affect its biogeochemistry. In order to tease out BCP characteristics and responses to these multiscale processes, the sediment trap studies have been conducted in the SCS for over 40 years. In September 1987, the first successful mooring deployment of sediment traps in the SCS was executed during the joint Chinese-German R/V Sonne cruise 50. A project known as “Modern Marine sedimentary processes in the South China Sea” was proposed as one of the main purposes of the cruise within a scientific cooperation framework between the State Oceanic Administration (China) and the Federal Ministry of Research and Technology in the field of marine geosciences and technology (Germany). This particle flux-oriented project lasted for ten years, subsequently, eight joint cruises were conducted, and a total of 12 sediment traps were deployed and retrieved in the northern, central and western (upwelling area off Vietnam) SCS. This project followed the forefront of international marine science research and was considered to be part of the JGOFS program at that time (Wiesner et al., 1996). In the following decades, more sediment traps were deployed by several universities and institutes to extend the long-term observations over the SCS basin, broaden the research area and simultaneously enrich multiparameter investigations to determine the sinking particle components and fluxes (Lahajnar et al., 2007; Gaye et al., 2009; Ho et al., 2010; Schröeder et al., 2015; Dong et al., 2016; Li J. et al., 2022).
So far, the researches move forward on the mechanisms that regulate the dynamics of vertically settling particle fluxes in the deep SCS (Ran et al., 2015; Li et al., 2017; Zhang et al., 2019; Tan et al., 2020; Li H. et al., 2022), but a comprehensive understanding is still lacking. In this context, we firstly synthesize the reported results to introduce the general characteristics of BCP in the mesopelagic SCS, then highlight how key physical processes occurring from intraseasonal to interannual time scales control the mesopelagic POC flux, and finally examine the prospects for future research in this area.
BCP characteristics in the mesopelagic layer
Coupled trends between the mesopelagic and upper layers
Literature data derived from sediment trap studies (Details of data source and compilation can be found in Supplementary material) have been complied to show a general layout for total mass flux (TMF) and POC flux at mesopelagic SCS of current stage. On average, the TMF at ~1000 m depth ranges from 66.3 ± 16.2 to 558.9 ± 446.0 mg m−2 d−1, while the POC flux varies from 3.4 ± 1 to 14.6 ± 10.3 mg m−2 d−1. Both the TMF and POC flux decrease from the slope to the deep basin, with relatively higher values in the western boundary and the lowest values in the central basin, showing a consistent spatial pattern with the annual mean Chl-a derived from satellite sensors (Figure 1A). Compared with the global synthetic data of POC flux at 2000 m from JGOFS (Honjo et al., 2008), the POC flux in SCS is obviously higher than those in the Pacific Warm Pool (0.8 mg m−2 d−1), but lower than those in the divergent Arabian Sea (19.9 mg m−2 d−1). Temporally, one-year consecutive records (Zhang et al., 2019; Tan et al., 2020), multiyear time series (Lui et al., 2018) and climatological results (Li et al., 2017) have all confirmed that the variability in sinking POC in the mesopelagic layer is synchronized with the overlying Chl-a changes.
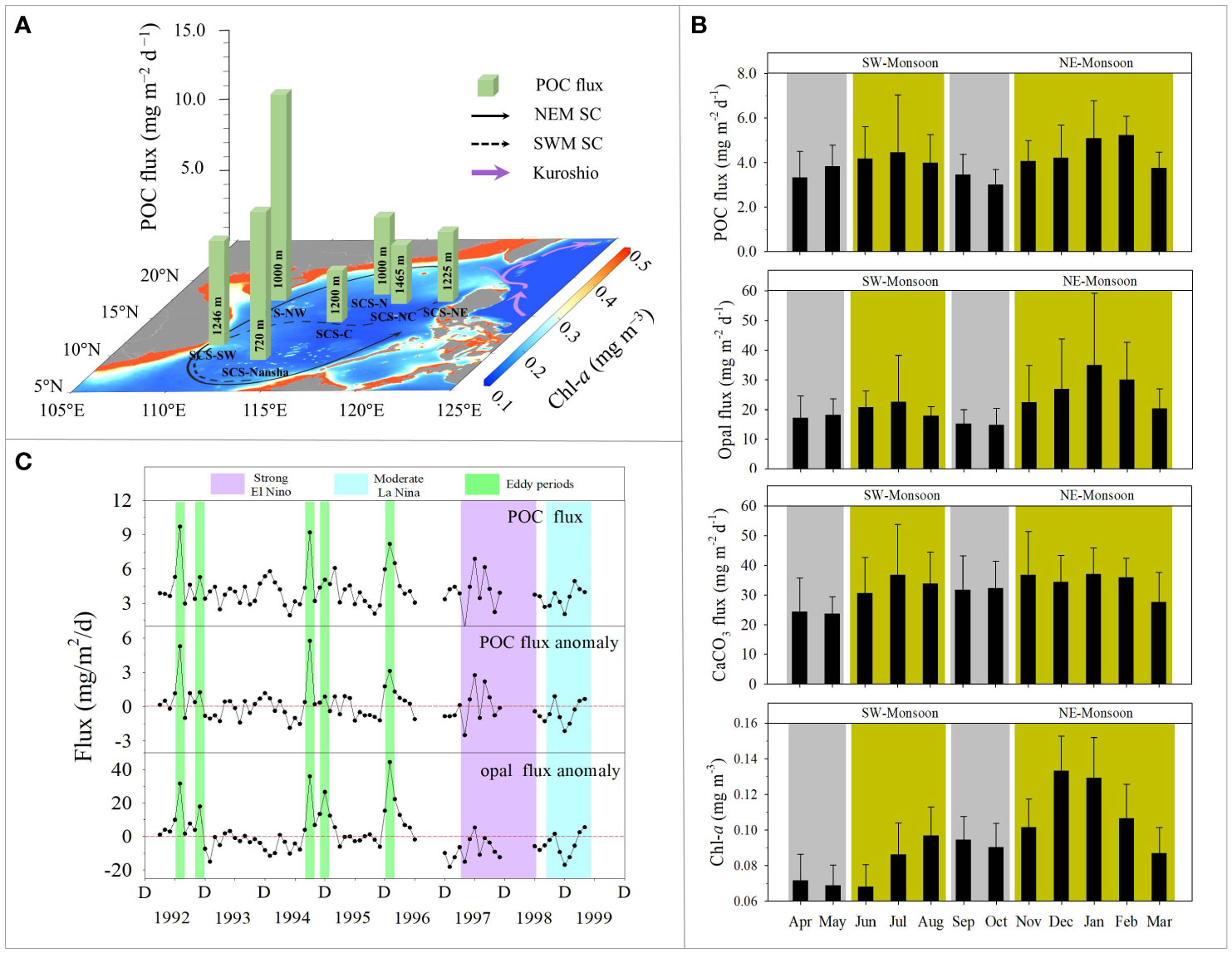
Figure 1 Characteristics of mesopelagic particulate organic carbon (POC) flux from the sediment trap observations of South China Sea. (A) illustrates the spatial distribution of mesopelagic POC flux and surface chlorophyll a (Chl-a) content. The green column height represents annual average POC flux in mg m−2 d−1 at mesopelagic layer, the sampling depths labelled in the columns. The POC data are derived from individual published papers with detailed values and station information listed in Supplementary Table S1. The Chl-a distribution is the multi-year averaged result (from 2002 to 2022) of the annual mean surface data in mg m−3, which derived from Moderate-resolution Imaging Spectroradiometer (MODIS) Aqua satellite (https://oceancolor.gsfc.nasa.gov/l3/). The black solid arrow represents the surface circulation during northeast monsoon period (NEM SC), the black dashed arrow represents the surface circulation during southwest monsoon period (SWM SC), and purple arrows represent the Kuroshio Current. (B) Climatology of POC flux, opal flux, CaCO3 flux, and surface Chl-a at station SCS-C (Li et al., 2017). Yellow and gray shades indicate monsoon and inter-monsoon periods, respectively. (C) shows time series results of monthly mean POC flux, POC flux anomaly and opal flux anomaly during 1992-1999 at station SCS-C (Li et al., 2017). The green indicates eddy periods with large flux anomalies, whilst purple and cyan indicate 1997/1998 El Niño and 1998/1999 La Niña events, respectively.
In addition to POC, calcium carbonate (CaCO3) and opal are other two main biogenic components, which refer to biogenic minerals, and these are mainly sourced from the skeletons of calcareous and siliceous plankton, respectively. Significant relationships between biogenic minerals, POC and TMF have been observed in the mesopelagic SCS (Li et al., 2017; Zhang et al., 2019; Tan et al., 2020). In general, the CaCO3 and opal fluxes exhibit a similar temporal pattern to that of POC flux at a certain depth for a given station (Honjo et al., 2008 and references therein); in turn, they act as vehicles and protection shielding for organic matter, increasing the settling rates and potentially decreasing degradation rates (Armstrong et al., 2002; Francois et al., 2002). Microscopic inspections of siliceous and calcareous plankton in sediment trap samples exhibited higher abundance during productive seasons with larger export fluxes and less abundance during periods of low productivity and fluxes (Chen et al., 2007; Ran et al., 2015; Priyadarshani et al., 2019; Ladigbolu et al., 2020). This biological information remaining in the mesopelagic particles is consistent with the previously reported significant temporal variations in phytoplankton biomass and community structures in the euphotic layer (Ning et al., 2004).
The impact of lateral transport on BCP export components
Notably, lithogenic matter occupies a large proportion of sinking particles (Jennerjahn et al., 1992; Zhang et al., 2019; Tan et al., 2020; Li H. et al., 2022), which raises the question of whether there exists strong advection of heterochthonous material in the water column of SCS basin. Micropaleontological investigations have provided some evidences. For example, extinct coccoliths were inspected as a perennial component in the mesopelagic sinking particles from the northern SCS basin, with average proportions of 3.3-5.1% in the annual total coccolith flux. These extinct coccoliths probably sourced from the Miocene and remaining in the sediment cover to the west and south of the Dongsha Islands, which somehow lateral transported to the northern basin (Priyadarshani et al., 2019). Again groups of benthic and freshwater diatoms were also found in the trap samples from the northwestern SCS, with the abundances of 1.5-6.9% (Ran et al., 2022). However, the lateral contribution is currently hard to quantify due to the lack of information as to the abundance of benthic diatoms and reworked coccoliths in the surface sediment in the northern and northwestern SCS.
Based on the analysis of POC source indicators, the C/N mole ratio and dual carbon isotope compositions (δ13Corg and radiocarbon content), the sinking POC characteristics in the mesopelagic layer were relatively homogeneous and comparable with the POC collected from the overlying surface water (Liu et al., 2007; Zhang et al., 2019; Zhang et al., 2022). Estimates of binary mixing end-member models demonstrated that POC incorporated in sinking particles was predominantly derived from primary productivity in the overlying surface ocean, no matter in the slope area (Zhang et al., 2019) or at the basin station (Zhang et al., 2022). The relatively high proportion of lithogenic matter, as well as the covarying trend between lithogenic matter and POC flux, is likely due to scavenging by sinking organic matter, and in turn, the lithogenic matter acts as ballast material to accelerate the settling process, especially during the highly productive seasons (Zhang et al., 2019). The contribution of lateral transported organic carbon is reported to be much more important in the offshore sediment or in the sinking particles collected from the near bottom layers (Kao et al., 2014; Blattmann et al., 2018; Wei et al., 2020), but the export flux of sinking biogenic matter in the mesopelagic layer of SCS basin is overall mainly controlled by the productive processes in the overlying layer with the nonsignificant impact of lateral advection.
Multiscale mechanisms of high-flux events
The SCS is a typical well-stratified oligotrophic and unproductive sea, except for its coastal areas (Wong et al., 2007; Du et al., 2017), thereby the nutrient influx into the euphotic zone introduced by multiple physical forcings will determine the magnitude of primary production and the subsequent export flux. These physical forcings have been distinguished by an axis of time scales, ranging from intraseasonal episodic events to seasonal monsoon and interannual El Niño Southern Oscillation (ENSO) events (Figure 2), which mediate the dynamic temporal variability in mesopelagic particle export flux.
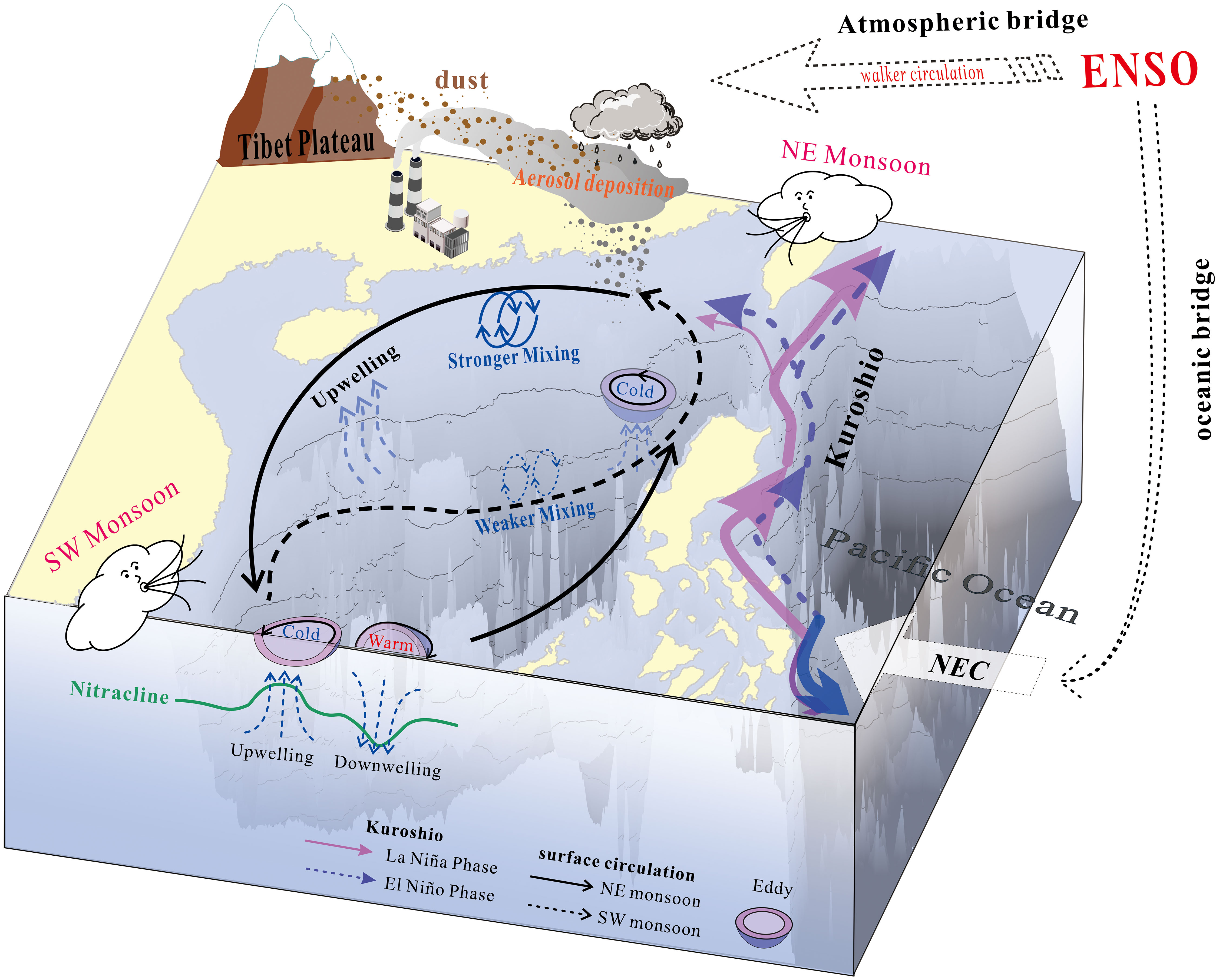
Figure 2 Schematic diagram of the multi-scale physical processes on particle export flux in the SCS basin: from interannual ENSO, to seasonal monsoon, then to intra-seasonal processes, such as upwelling, dust events, and mesoscale eddies. NE represents northeast, SW represents southwest, NEC represents north equatorial current.
Seasonal transition
Overall, the East Asian monsoon has been regarded as a first-order control on BCP strength in the SCS. It not only drives the basin-scale background circulation (Wyrtki, 1961; Hu et al., 2000) but also profoundly influences biogeochemical processes and particle export (Ning et al., 2004). Clear seasonal variabilities in total mass and main component (POC, CaCO3, opal and amino acid) fluxes of trap-collected particles correspond to monsoon transition. Generally, the values of POC flux in the northern SCS basin are always highest in winter but remain at a relatively low level during summer and the intermonsoon seasons (Wiesner et al., 1996; Lui et al., 2018). The prevailing, strong northeast wind coupled with extensive cooling in winter enhances vertical mixing (i.e., r2 = 0.61, N=19, p<0.001, Tseng et al., 2009a), which entrains more subsurface nutrient into the upper layer (Tseng et al., 2005; 2009) and then fuels photosynthetic productivity (Chen, 2005; Liu et al., 2013; Wong et al., 2015; Du et al., 2017). Such controlling mechanism of mixed layer depth has also been quantitatively validated in further studies, and corresponding biogenic parameters including Chl-a, net primary production as well as all components of mesopelagic particles show significant correlations with it (Table 3 in Tan et al. (2020); Table 1 in Li et al. (2017). Statistical result indicates that the biogenic particles in winter accounts for nearly half of the annual measured fluxes (41.5-48.9%). In the central SCS, except for the winter peak, there is a secondary peak during the southwest monsoon period, showing a distinct bimodal seasonality in the monthly multiyear average POC and opal fluxes (Li et al., 2017; Figure 1B). The fluxes of total mass, POC and foraminifera at a station located further south also demonstrate a covaried bimodal pattern, with maximum values during monsoon seasons, and the values in summer seem to be slightly higher than those in winter (Wan et al., 2010). The spatial differences in the seasonal transition pattern of the particle export flux might be highly correlated with the strength discrepancy of the East Asian monsoon from north to south across the SCS.
Significantly, several extremely high-flux peaks coupled with cyclonic eddies have been observed in the long-term records of a central station of the SCS (Li et al., 2017; Figure 1C). Cyclonic eddies, typically upwelling nutrient-rich subsurface water and stimulating phytoplankton blooms (Tang et al., 1999; Ning et al., 2004), are mainly generated during the northeastern monsoon period (Wang et al., 2003). Their contributions to biogenic particle flux may overlap the regular winter flux peak, with higher mesopelagic POC and opal fluxes of 41% and 116% respectively, compared with fluxes during non–cyclonic eddy periods (Li et al., 2017). Therefore, the seasonal pattern of biogeochemical responses in the SCS cannot be solely attributed to the influence of the monsoon transition, winter cyclonic eddies would amplify the seasonal particle export cycle.
Intraseasonal episodic events
The temporal variability in biogenic particle flux does not always follow its climatological seasonal pattern in the SCS. In addition to the northeast monsoon maxima, some discrete high-flux events have been observed in both the northwestern slope and the northern basin (Ran et al., 2015; Zhang et al., 2019; Tan et al., 2020). These episodic flux pulses are likely to be driven by randomly occurring intraseasonal physical processes, such as subsurface upwelling, aerosol deposition or mesoscale eddies. Local upwelling along the western boundary of the SCS (e.g., the northern shelf and the Vietnam coast) is probably generated by southwest winds during the summer monsoon. These intraseasonal processes may serve to provide “new” nutrients from underlying water or atmospherically deposition into the euphotic zone, thereby stimulating phytoplankton growth and a subsequent particle rainout. In contrast, the occasional passage of anticyclonic eddies may likely suppress the subsurface nutrient supply, which would substantially reduce primary production and subsequent export flux (Tan et al., 2020). As statistically estimated, three discrete high-flux events in the northwestern SCS determined by upwelling, aerosol deposition, and the northeast monsoon occupied 40% of the total sampling period but contributed nearly 80% of the biogenic particles (Zhang et al., 2019). Notably, the events contributed nearly equally to the total POC flux with their own unique biogeochemical overprints, implying that in the SCS, the intraseasonal physical processes might be as important locally as the seasonal monsoon in modulating the strength of the BCP.
Interannual ENSO
ENSO is a recurring ocean-atmosphere coupled phenomenon that occurs across the tropical Pacific, with extreme warmer (El Niño) and cooler phases (La Niña) that dominate the global weather and climate patterns at the interannual time scale. In 2009, Tseng et al. proposed that winter monthly Chl-a (and integrated primary production) in the northern SCS decreased by 42% (and 42%) and 13% (and 10%), respectively, during two El Niño events (1997/1998 and 2002/2003). The reduction was attributed to the diminished vertical mixing and strengthened stratification by statistical analysis (Tseng et al., 2009a). Later studies have further elucidated that ENSO could impact the SCS biogeochemical conditions not only via abnormal changes in atmospheric forcing (wind speed and heat flux), but also through basin-scale circulation (Kuroshio and SCS throughflow) (Xiao et al., 2017), ultimately resulting in an asymmetric response in both surface Chl-a and mesopelagic flux during the 1997-1999 ENSO cycle (Liu et al., 2013; Li H. et al., 2022). This response is different from the general responses in the equatorial Pacific Ocean: the BCP weakens during El Niño events and strengthens during La Niña events (Chavez et al., 1999; Gierach et al., 2012; Brainard et al., 2018).
By extension, during the 1997/1998 super El Niño event, weak wind and intense solar radiation strengthened the upper water stratification, the Luzon Strait transport was therefore enhanced and intensified the oligotrophic Kuroshio intrusion to dilute the nutrient inventory in the SCS basin (Du et al., 2013). Together, compared with the climatological means, these processes led to impoverished bioavailable nutrients for photosynthetic productivity (decreased by 17%), and in particular, they inhibited diatom growth (opal flux decreased by 31.7%), thus resulting in an inefficient BCP with low particle export flux (Li H. et al., 2022). When the climate conditions oscillated to the La Niña phase in 1998/1999, the atmospheric forcing and the ongoing water mixing rapidly recovered to the normal state, but no obvious biological activity occurred. The net primary production and mesopelagic POC flux were still lower (13% and 10.9%, respectively) than the climatological means (Li H. et al., 2022). Although the Kuroshio intrusion was weakened, the thermocline/nutricline deepened due to basin-scale downwelling circulation or episodic warm eddies (Wang et al., 2002), which debilitated the nutrient supply, subsequently restraining the rebound of primary production and deep biogenic export flux (Liu et al., 2013; Li H. et al., 2022). However, the studies explained only the extreme event within a whole ENSO cycle, from El Niño to La Niña, while dynamic ENSO events with individual phases may present processes that respond differently. For example, the interannual variation in Chl is modulated by the intensity of the monsoon and mesoscale eddies which exhibits a close relationship with ENSO (Wang et al., 2006; Tseng et al., 2009b; He et al., 2016; Xiu et al., 2018). As extreme El Niño events are predicted to occur more often due to anthropogenically forced global warming (Cai et al., 2014), systematic studies are needed to evaluate the alterations of marine ecosystems and carbon cycles in response to the combined multiscale physical forcings in the SCS.
Synthesis and perspectives
The current research suggests that temporal variabilities in mesopelagic biogenic particle export are tightly coupled with the biological responses to nutrient influx into the euphotic zone, which is mediated by multiple physical forcings in the overlying SCS. However, sinking organic matter in the mesopelagic layer integrates an overlay of multiple physical and biological processes, which are intertwined from production to sedimentation. The underlying mechanisms and processes delivering these particles to the mesopelagic zone are complicated and have rarely been explored in the SCS, particularly the biogeochemical controls, which hinder mechanistic reproductions and predictions of the magnitude and efficiency of downward POC export. To better understand BCP processes and provide reliable parameterized conditions for coupled physical/biogeochemical models, the following aspects still need to be understood. First, systematic studies on coupled physical and biological processes based on high-resolution sampling, e.g., multidisciplinary integrated stationary moorings, could provide more refined insights into the underlying mechanisms of ecosystem processes and POC dynamics, as reviewed in this study. Second, research on biogeochemical controls on sinking particle dynamics should be conducted, particularly in the under-sampled twilight zone, with processes, such as aggregation and disaggregation, respiration and remineralization, the role of zooplankton, and food web interactions. Finally, the characteristics of sinking particles with particle sizes, phytoplankton structures, organic compounds, fecal pellets, biominerals and clay minerals, must be determined in future studies since they all appear to affect the POC transfer efficiency to the deep ocean, especially the minerals which subsequently act as vehicles for particle transport.
Author contributions
The manuscript was drafted by JZ. All authors contributed to the article and approved the submitted version.
Funding
The authors acknowledge funding from the Scientific Research Fund of the Second Institute of Oceanography, MNR (Grant Nos. JG2213, JZ2001, JG1913), the Project supported by Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai) (No. SML2021SP207), the Natural Science Foundation of China (Grant Nos. 42176039, 42106045), and the National Program on Global Change and Air-Sea Interaction II (Grant Nos. GASI-04-HYST-01).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1209686/full#supplementary-material
References
Armstrong R. A., Lee C., Hedges J. I., Honjo S., Wakeham S. G. (2002). A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals. Deep Sea Res. Part II: Topical Stud. Oceanography 49 (1–3), 219–236. doi: 10.1016/S0967-0645(01)00101-1
Blattmann T. M., Zhang Y., Zhao Y., Wen K., Lin S., Li J., et al. (2018). Contrasting fates of petrogenic and biospheric carbon in the South China Sea. Geophysical Res. Lett. 45, 9077–9086. doi: 10.1029/2018GL079222
Brainard R. E., Oliver T. A., Mcphaden M. J., Cohen A. L., Venegas R., Heenan A., et al. (2018). Ecological impacts of the 2015/16 el niño in the central equatorial pacific. Bull. Am. Meteorological Soc. 99(1), 21–26. doi: 10.1175/BAMS-D-17-0128.1
Cai W., Borlace S., Lengaigne M., Rensch P. V., Collins M., Vecchi G. A., et al. (2014). Increasing frequency of extreme El Niño events due to greenhouse warming. Nat. Climate Change 4, 111–116. doi: 10.1038/nclimate2100
Chavez F. P., Messié M., Pennington J. T. (2011). Marine primary production in relation to climate variability and change. Annu. Rev. Mar. Sci. 3, 227–260. doi: 10.1146/annurev.marine.010908.163917
Chavez F. P., Strutton P. G., Friederich G. E., Feely R. A., Feldman G. C., Foley D. G., et al. (1999). Biological and chemical response of the equatorial pacific ocean to the 1997-98 El Niño. Science 286, 2126–2131. doi: 10.1126/science.286.5447.2126
Chen Y. L. (2005). Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea. Deep Sea Res. Part I: Topical Stud. Oceanography 52, 319–340. doi: 10.1016/j.dsr.2004.11.001
Chen R., Wiesner G. M., Zheng Y., Cheng X., Jin H., Zhao Q., et al. (2007). Seasonal and annual variations of marine sinking particulate flux during 1993–1996 in the central South China Sea. Acta Oceanologica Sin. 26, 33–43.
Deuser W. G., Ross E. H., Anderson R. F. (1981). Seasonality in the supply of sediment to the deep Sargasso Sea and implications for the rapid transfer of matter to the deep ocean. Deep Sea Res. Part A: Oceanography Res. Papers 28(5), 495–505. doi: 10.1016/0198-0149(81)90140-0
Dong Y., Li Q. P., Wu Z., Zhang J. (2016). Variability in sinking fluxes and composition of particle-bound phosphorus in the Xisha area of the northern South China Sea. Deep Sea Res. Part I: Topical Stud. Oceanography 118, 1–9. doi: 10.1016/j.dsr.2016.10.007
Du C., Liu Z., Dai M., Kao S., Cao Z., Zhang Y., et al. (2013). Impact of the Kuroshio intrusion on the nutrient inventory in the upper northern South China Sea: insights from an isopycnal mixing model. Biogeosciences 10, 6419–6432. doi: 10.5194/bg-10-6419-2013
Du C., Liu Z., Kao S., Dai M. (2017). Diapycnal fluxes of nutrients in an oligotrophic oceanic regime: the south China sea. Geophysical Res. Lett. 44 (11), 510–518. doi: 10.1002/2017GL074921
Francois R., Honjo S., Krishfield R. A., Manganini S. J. (2002). Factors controlling the flux of organic carbon to the bathypelagic zone of the ocean. Global Biogeochemical Cycles 16(4), 1087. doi: 10.1029/2001GB001722
Gaye B., Wiesner M. G., Lahajnar N. (2009). Nitrogen sources in the South China Sea, as discerned from stable nitrogen isotopic ratios in rivers, sinking particles, and sediments. Mar. Chem. 114, 72–85. doi: 10.1016/j.marchem.2009.04.003
Gierach M. M., Lee T., Turk D., Mcphaden M. J. (2012). Biological response to the 1997–98 and 2009–10 El Niño events in the equatorial Pacific Ocean. Geophysical Res. Lett. 39, L10602. doi: 10.1029/2012GL051103
He Q., Zhan H., Cai S., Li Z. (2016). Eddy effects on surface chlorophyll in the northern South China Sea: Mechanism investigation and temporal variability analysis. Deep Sea Res. Part I: Topical Stud. Oceanography 112, 25–36. doi: 10.1016/j.dsr.2016.03.004
Henson S. A., Le Moigne F. A., Giering S. L. (2019). Drivers of carbon export efficiency in the global ocean. Global Biogeochemical Cycles 33 (9), 891–903. doi: 10.1029/2018GB006158
Ho T., Chou W., Wei C., Lin F., Wong G. T., Line H. (2010). Trace metal cycling in the surface water of the South China Sea: Vertical fluxes, composition, and sources. Limnology Oceanography 55 (5), 1807–1820. doi: 10.4319/lo.2010.55.5.1807
Honjo S., Connell J. F., Sachs P. L. (1980). Deep-ocean sediment trap; design and function of PARFLUX Mark II. Deep Sea Res. Part A: Oceanographic Res. Papers 27 (9), 745–753. doi: 10.1016/0198-0149(80)90026-6
Honjo S., Manganini S. J., Krishfield R. A., Francois R. (2008). Particulate organic carbon fluxes to the ocean interior and factors controlling the biological pump: A synthesis of global sediment trap programs since 1983. Prog. Oceanography 76, 217–285. doi: 10.1016/j.pocean.2007.11.003
Hu J., Kawamura H., Hong H., Qi Y. (2000). A review on the currents in the south China sea: seasonal circulation, south China sea warm current and kuroshio intrusion. J. Oceanography 56, 607–624. doi: 10.1023/A:1011117531252
Jennerjahn T. C., Liebezeit G., Kempe S. (1992). “Particle flux in the north ern South China Sea,” in Marine geology and geophysics of South China Sea. Eds. Jin X., Kudrass H. R., Pautot G. (Bei-jing: China Ocean Press), 228–235.
Johnson K. S., Bif M. B. (2021). Constraint on net primary productivity of the global ocean by Argo oxygen measurements. Nat. Geosci. 14769, 774. doi: 10.1038/s41561-021-00807-z
Kao S. J., Hilton R. G., Selvaraj K., Dai M., Zehetner F., Huang J. C., et al. (2014). Preservation of terrestrial organic carbon in marine sediments offshore Taiwan: Mountain building and atmospheric carbon dioxide sequestration. Earth Surface Dynamics 2 (1), 127–139. doi: 10.5194/esurf-2-127-2014
Kwon E. Y., Primeau F., Sarmiento J. L. (2009). The impact of remineralization depth on the air-sea carbon balance. Nat. Geoscience 2, 630–635. doi: 10.1038/ngeo612
Ladigbolu I. A., Li H., Li B., Wiesner M. G., Zhang J., Sun L., et al. (2020). Calcification depths and temperatures of planktonic foraminifera off southwest Hainan Island and their paleoceanographic implications. Mar. Micropaleontol. 158, 101878. doi: 10.1016/j.marmicro.2020.101878
Lahajnar N., Wiesner M. G., Gaye B. (2007). Fluxes of amino acids and hexosamines to the deep South China Sea. Deep Sea Res. Part I: Oceanographic Res. Papers 54 (12), 2120–2144. doi: 10.1016/j.dsr.2007.08.009
Lee C. (2019). Passing the baton to the next generation: A few problems that need solving. Annu. Rev. Mar. Sci. 11, 1–13. doi: 10.1146/annurev-marine-010318-095342
Li J., Liu Z., Lin B., Zhao Y., Cao J., Zhang X., et al. (2022). Zooplankton fecal pellet characteristics and contribution to the deep-sea carbon export in the southern south China sea. J. Geophysical Research: Oceans 127, e2022JC019412. doi: 10.1029/2022JC019412
Li H., Wiesner M. G., Chen J., Ling Z., Zhang J., Ran L. (2017). Long-term variation of mesopelagic biogenic flux in the central South China Sea: Impact of monsoonal seasonality and mesoscale eddy. Deep Sea Res. Part I: Topical Stud. Oceanography 126, 62–72. doi: 10.1016/j.dsr.2017.05.012
Li H., Zhang J., Xuan J., Wu Z., Ran L., Wiesner M. G., et al. (2022). Asymmetric response of the biological carbon pump to the ENSO in the South China Sea. Geophysical Res. Lett. 49, e2021GL095254. doi: 10.1029/2021GL095254
Liu K. K., Kao S. J., Hu H. C., Chou W. C., Hung G. W., Tseng C. M. (2007). Carbon isotopic composition of suspended and sinking particulate organic matter in the northern South China Sea—From production to deposition. Deep Sea Res. Part II: Topical Stud. Oceanography 54 (14-15), 1504–1527. doi: 10.1016/j.dsr2.2007.05.010
Liu K. K., Chao S., Shaw P., Gong G. C., Chen C. C., Tang T. Y. (2002). Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study. Deep Sea Res. Part I: Topical Stud. Oceanography 49, 1387–1412. doi: 10.1016/S0967-0637(02)00035-3
Liu K. K., Wang L. W., Dai M., Tseng C. M., Yang Y., Sui C. H., et al. (2013). Inter-annual variation of chlorophyll in the northern South China Sea observed at the SEATS station and its asymmetric responses to climate oscillation. Biogeosciences 10 (11), 7449–7462. doi: 10.5194/bg-10-7449-2013
Lui H., Chen K., Chen C. A., Wang B., Lin H., Ho S., et al. (2018). Physical forcing-driven productivity and sediment flux to the deep basin of northern south China sea: A decadal time series study. Sustainability 10, 971. doi: 10.3390/su10040971
Muller-Karger F. E., Varela R., Thunell R. C., Luerssen R., Hu C., Walsh J. J. (2005). The importance of continental margins in the global carbon cycle. Geophysical Res. Lett. 32, L01602. doi: 10.1029/2004GL021346
Ning X., Chai F., Xue H., Cai Y., Liu C., Shi J. (2004). Physical-biological oceanographic coupling influencing phytoplankton and primary production in the south China sea. J. Geophysical Res. 109, C10005. doi: 10.1029/2004JC002365
Parekh P., Follows M. J., Dutkiewicz S., Ito T. (2006). Physical and biological regulation of the soft tissue carbon pump. Paleoceanography 21, PA3001. doi: 10.1029/2005PA001258
Priyadarshani W., Ran L., Wiesner M. G., Chen J., Ling Z., Yu S., et al. (2019). Seasonal and interannual variability of coccolithophore flux in the northern South China Sea. Deep Sea Res. Part I: Oceanographic Res. Papers 145, 13–30.
Ran L., Chen J., Wiesner M. G., Ling Z., Lahajnar N., Yang Z., et al. (2015). Variability in the abundance and species composition of diatoms in sinking particles in the northern South China Sea: Results from time-series moored sediment traps. Deep-sea Res. Part Ii-topical Stud. Oceanography 122, 15–24. doi: 10.1016/j.dsr2.2015.07.004
Ran L., Ma W., Wiesner M. G., Wang Y., Chen J., Zhang L., et al. (2022). Sediment resuspension as a major contributor to sinking particles in the northwestern south China sea: evidence from observations and modeling. Front. Mar. Sci. 9, 819340. doi: 10.3389/fmars.2022.819340
Schröeder A., Wiesner M. G., Liu Z. (2015). Fluxes of clay minerals in the South China Sea. Earth Planetary Sci. Lett. 430, 30–42. doi: 10.1016/j.epsl.2015.08.001
Sigman D. M., Boyle E. A. (2000). Glacial/interglacial variations in atmospheric carbon dioxide. Nature 407 (6806), 859–869. doi: 10.1038/35038000
Takahashi K. (1986). Seasonal fluxes of pelagic diatoms in the subarctic Pacific 1982-1983. Deep Sea Res. Part A:Oceanographic Res. Papers 33, 1225–1251.
Takahashi K., Honjo S. (1981). Vertical flux of Radiolaria: A taxon-quantitative sediment trap study from the western tropical Atlantic. Micropaleontology 2, 140–190. doi: 10.2307/1485284
Tan S. Y., Zhang J., Li H., Sun L., Wu Z., Wiesner M. G., et al. (2020). Deep ocean particle flux in the northern south China sea: variability on intra-seasonal to seasonal timescales. Front. Earth Sci. 8, 74. doi: 10.3389/feart.2020.00074
Tang D., Ni I., Kester D., Müllerkarger F. (1999). Remote sensing observations of winter phytoplankton blooms southwest of the Luzon Strait in the South China Sea. Mar. Ecol. Prog. Ser. 191, 43–51. doi: 10.3354/meps191043
Tseng C., Gong G. C., Wang L., Liu K., Yang Y. (2009a). Anomalous biogeochemical conditions in the northern South China Sea during the El-Nin˜o events between 1997 and 2003. Geophysical Res. Lett. 36, L14611. doi: 10.1029/2009GL038252
Tseng C., Liu K., Wang L., Gong G. C. (2009b). Anomalous hydrographic and biological conditions in the northern South China Sea during the 1997-1998 El Niño and comparisons with the equatorial Pacific. Deep Sea Res. Part I: Oceanographic Res. Papers 56, 2129–2143. doi: 10.1016/j.dsr.2009.09.004
Tseng C., Wong G. T., Lin I., Wu C., Liu K. (2005). A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea. Geophysical Res. Letters. 32, L08608. doi: 10.1029/2004GL022111
Wang S. H., Hsu N. C., Tsay S. C., Lin N. H., Sayer A. M., Huang S. J., et al. (2012). Can Asian dust trigger phytoplankton blooms in the oligotrophic northern South China Sea? Geophysical Res. Lett. 39 (5), L05811. doi: 10.1029/2011GL050415
Wan S., Jian Z., Cheng X., Qiao P., Wang R. (2010). Seasonal variations in planktonic foraminiferal flux and the chemical properties of their shells in the southern South China Sea. Sci. China Earth Sci. 40 (7), 881–892. doi: 10.1007/s11430-010-4039-3
Wang C., Wang W., Wang D., Wang Q. (2006). Interannual variability of the South China Sea associated with El Niño. J. Geophysical Res. 111, C03023. doi: 10.1029/2005JC003333
Wang G., Su J., Chu P. C. (2003). Mesoscale eddies in the South China Sea observed with altimeter data. Geophysical Res. Lett. 30 (21), 2121. doi: 10.1029/2003GL018532
Wang D., Xie Q., Du Y., Wang W., Chen J. (2002). The 1997–1998 warm event in the South China Sea. Chin. Sci. Bulletin 47, 1221–1227. doi: 10.1007/BF02907614
Wiesner M. G., Zheng L., Wong H. K., Wang Y., Chen W. (1996). “Fluxes of particulate matter in the South China Sea,” in Particle flux in the ocean. Eds. Ittekkot. V., Honjo S. (New York: John wiley & Sons), 293–312.
Wong G. T., Pan X., Li K., Shiah F., Ho T., Guo X. (2015). Hydrography and nutrient dynamics in the Northern South China Sea Shelf-sea (NoSoCS). Deep-sea Res. Part II: Topical Stud. Oceanography 117, 23–40.
Wong G. T. F., Tseng C. M., Wen L. S., Chung S. W. (2007). Nutrient dynamics and N-anomaly at the SEATS station. Deep Sea Res. Part II: Topical Stud. Oceanography 54 (14–15), 1528–1545. doi: 10.1016/j.dsr2.2007.05.011
Wyrtki K. (1961). Scientific results of marine investigations of the South China Sea and the Gulf of Thailand 1959–1961. NAGA Rep. 2.
Xiao F., Zeng L., Liu Q., Zhou W., Wang D. (2017). Extreme subsurface warm events in the South China Sea during 1998/99 and 2006/07: observations and mechanisms. Climate Dynamics 50, 115–128. doi: 10.1007/s00382-017-3588-y
Xiu P., Chai F. (2011). Modeled biogeochemical responses to mesoscale eddies in the South China Sea. J. Geophysical Res. 116, C10006. doi: 10.1029/2010jc006800
Xiu P., Chai F., Shi L., Xue H. J., Chao Y. (2010). A census of eddy activities in the South China Sea during 1993–2007. J. Geophysical Res. 115, C03012. doi: 10.1029/2009jc005657
Xiu P., Dai M., Chai F., Zhou K., Zeng L., Du C. (2018). On contributions by wind-induced mixing and eddy pumping to interannual chlorophyll variability during different ENSO phases in the northern South China Sea. Limnology Oceanography 64, 503–514. doi: 10.1002/lno.11055
Zhang J., Li H., Wiesner M. G., Eglinton T. I., Haghipour N., Jian Z., et al. (2022). Carbon isotopic constraints on basin-scale vertical and lateral particulate organic carbon dynamics in the northern South China Sea. J. Geophysical Research: Oceans 127, e2022JC018830. doi: 10.1029/2022JC018830
Keywords: biological carbon pump, POC flux, sediment trap, physical forcing, South China Sea
Citation: Zhang J, Li H, Wiesner MG, Ran L, He X, Chi G, Wang X, Yu J and Chen J (2023) Biological carbon pump responses to multiscale physical processes: a review of sediment trap studies in the South China Sea. Front. Mar. Sci. 10:1209686. doi: 10.3389/fmars.2023.1209686
Received: 21 April 2023; Accepted: 21 August 2023;
Published: 06 September 2023.
Edited by:
Selvaraj Kandasamy, Central University of Kerala, IndiaReviewed by:
Jeomshik Hwang, Seoul National University, Republic of KoreaPeng Yao, Ocean University of China, China
Copyright © 2023 Zhang, Li, Wiesner, Ran, He, Chi, Wang, Yu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongliang Li, bGlob25nbGlhbmdAc2lvLm9yZy5jbg==; Jianfang Chen, amZjaGVuQHNpby5vcmcuY24=