
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci., 03 October 2023
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1198137
This article is part of the Research TopicEcocentric Fisheries Management in European Seas: Data Gaps, Base Models and Initial Assessments, Volume IView all 10 articles
 Luisa R. Abucay1
Luisa R. Abucay1 Patricia Sorongon-Yap1
Patricia Sorongon-Yap1 Kathleen Kesner-Reyes1
Kathleen Kesner-Reyes1 Emily C. Capuli1Rodolfo B. Reyes Jr1Eva Daskalaki2
Emily C. Capuli1Rodolfo B. Reyes Jr1Eva Daskalaki2 Carmen Ferrà3,4
Carmen Ferrà3,4 Giuseppe Scarcella3
Giuseppe Scarcella3 Gianpaolo Coro5
Gianpaolo Coro5 Francesc Ordines6
Francesc Ordines6 Paula Sánchez-Zulueta6
Paula Sánchez-Zulueta6 Georgi Dakalov7
Georgi Dakalov7 Stefania Klayn7
Stefania Klayn7 Liesa Celie8
Liesa Celie8 Marco Scotti9,10
Marco Scotti9,10 David Grémillet11,12Charlotte Lambert13
David Grémillet11,12Charlotte Lambert13 Gideon Gal14
Gideon Gal14 Maria Lourdes D. Palomares15,16Donna Dimarchopoulou17,18
Maria Lourdes D. Palomares15,16Donna Dimarchopoulou17,18 Athanassios C. Tsikliras2*
Athanassios C. Tsikliras2*Available information and potential data gaps for non-fish marine organisms (cnidarians, crustaceans, echinoderms, molluscs, sponges, mammals, reptiles, and seabirds) covered by the global database SeaLifeBase were reviewed for eight marine ecosystems (Adriatic Sea, Aegean Sea, Baltic Sea, Bay of Biscay/Celtic Sea/Iberian Coast, Black Sea, North Sea, western Mediterranean Sea, Levantine Sea) across European Seas. The review of the SeaLifeBase dataset, which is based on published literature, analyzed information coverage for eight biological characteristics (diet, fecundity, maturity, length-weight relationships, spawning, growth, lifespan, and natural mortality). These characteristics are required for the development of ecosystem and ecological models to evaluate the status of marine resources and related fisheries. Our analyses revealed that information regarding these biological characteristics in the literature was far from complete across all studied areas. The level of available information was nonetheless reasonably good for sea turtles and moderate for marine mammals in some areas (Baltic Sea, Bay of Biscay/Celtic Sea/Iberian Coast, Black Sea, North Sea and western Mediterranean Sea). Further, seven of the areas have well-studied species in terms of information coverage for biological characteristics of some commercial species whereas threatened species are generally not well studied. Across areas, the most well-studied species are the cephalopod common cuttlefish (Sepia officinalis) and the crustacean Norway lobster (Nephrops norvegicus). Overall, the information gap is narrowest for length-weight relationships followed by growth and maturity, and widest for fecundity and natural mortality. Based on these insights, we provide recommendations to prioritize species with insufficient or missing biological data that are common across the studied marine ecosystems and to address data deficiencies.
Ecocentric (=ecosystem centered) fisheries management requires detailed knowledge of the structure and functioning of the marine ecosystems, from abiotic data to the status of all ecosystem components (Dimarchopoulou, 2020). This includes the fishing pressure applied on commercial and non-commercial marine populations and their respective biomasses (Tsikliras et al., 2023) and biological information (growth, maturity, spawning, fecundity, mortality, lifespan and diet) of all organisms in an ecosystem because marine organisms respond differently to fishing pressure and population time to recovery depends upon their life-history strategy and ecological traits (Dimarchopoulou et al., 2017). Ecosystem structure is usually described using mass balance ecosystem models (Heymans et al., 2020) while the stock status is derived from age-based or surplus production stock assessments (Tsikliras and Froese, 2019). Ecosystem models and stock assessments are thereby required to examine fisheries management and marine policy scenarios (Piroddi et al., 2022) within the context of environmental (Piroddi et al., 2021), oceanographic (Coll et al., 2019) and climatic change factors (Corrales et al., 2018), whilst also incorporating economic and social parameters (Link, 2010).
The development of ecosystem models demands specific biological data, mainly growth parameters, natural mortality and diet composition per species or functional group of species (Christensen and Walters, 2004), as well as catch data that are available per fleet through official landings statistics (global and regional databases of the Food and Agricultural Organisation of the United Nations: FAO, 2020) and catch datasets (Sea Around Us: Pauly and Zeller, 2016). Similarly, the simpler age-based stock assessments require growth parameters, size at maturity, spawning and natural mortality data (Jardim et al., 2015) while some surplus production models use the maximum intrinsic population growth that is based on several biological characteristics including growth, fecundity, maturity and natural mortality (Froese et al., 2018a; Froese et al., 2018b; Froese et al., 2020).
Among European Seas, the North East (NE) Atlantic Ocean is a marine ecosystem with a long scientific history of investigations across all marine science disciplines (Lotze and Worm, 2009). As a result, long time-series of biological, oceanographic and fisheries data exist, most of which are publicly available. These datasets have supported many ecosystem models (Keramidas et al., 2023) and the official full stock assessments in most marine ecosystems of the NE Atlantic (ICES, 2022). In contrast, the Mediterranean and the Black Seas, despite their longer history of fisheries exploitation (Stergiou et al., 2016) and the early scientific work on biology and fisheries by Aristotle and Oppian, respectively (Deacon, 1997), lack long time series of biological, oceanographic and fisheries data (Fortibuoni et al., 2017, but see Ravier and Fromentin, 2004). Consequently, data-limitations have constrained ecosystem models to specific and well-studied areas of the northern and eastern Mediterranean coastline that are well studied (Adriatic Sea: Barausse et al., 2009; Libralato et al., 2015; Catalan coast: Coll et al., 2008; Coll et al., 2009; Aegean Sea: Dimarchopoulou et al., 2019; Dimarchopoulou et al., 2022; Keramidas et al., 2022; Levantine Sea: Corrales et al., 2017; Shabtay et al., 2018; Corrales et al., 2019; Ofir et al., 2023). This also restricted full analytical stock assessments to a proportionally low number of exploited stocks (Piroddi et al., 2020) despite the increasing efforts of the Expert Working Groups (EWG) of the Scientific, Technical and Economic Committee for Fisheries (STECF) of the European Union and the General Fisheries Commission for the Mediterranean Sea (GFCM) of the FAO. However, the most important issues in Mediterranean fisheries are the north-south gradient in marine research and data, with more scientific output along the northern Mediterranean coastline (Stergiou and Tsikliras, 2006), and that valuable datasets are not openly available (McManamay and Utz, 2014). This is because some people/institutions (and countries, which were historically amongst the most scientifically advanced) do not believe in open science – a policy priority for the European Commission – even when supported by public funds (Damalas et al., 2018).
Robust and adaptive fisheries management policies require understanding their key sources of uncertainty, such as knowledge gaps in biology of marine species (Link et al., 2012). A recent update on the gaps in the biological knowledge of Mediterranean marine fishes (Daskalaki et al., 2022) indicated that efforts were made to reduce these gaps in knowledge across the Mediterranean Sea compared to previous records (Dimarchopoulou et al., 2017). This is especially true for threatened species fishes such as sharks and rays (Tsikliras and Dimarchopoulou, 2021) as well as for alien species that rapidly colonized the Mediterranean during the last decades (Katsanevakis et al., 2014). Filling the gaps in ecological and biological knowledge and assessing anthropogenic impacts marine ecosystems are prerequisites for developing robust ecosystem models (Heymans et al., 2020) hence for promoting effective ecocentric management (Claudet et al., 2019).
The principal aim of the present work was to review available information on key biological characteristics (diet, fecundity, maturity, length-weight relationships, spawning, growth, lifespan, and natural mortality) of non-fish marine species across European Seas. This allowed a gap analysis and a comparison of the availability of biological data across areas and taxonomic groups, leading to recommendations to reduce knowledge gaps (if and where required). Thus, future research will have a baseline to prioritize species of special interest based on specific criteria such as conservation status. This review covers the non-fish marine organisms belonging to eight taxonomic groups occurring in the European Seas. Fish species will be covered in a separate publication that will follow the same methodology and spatial coverage and will expand the review of the Mediterranean marine fishes (Daskalaki et al., 2022) to European Seas.
This review was based on information that was extracted from the literature and captured in SeaLifeBase (www.sealifebase.org; Palomares and Pauly, 2021, consulted in December 2021) for eight marine ecosystems (Adriatic Sea, Aegean Sea, Baltic Sea, Bay of Biscay/Celtic Sea/Iberian Coast, Black Sea, Levantine Sea, North Sea, and western Mediterranean Sea) (Figure 1). SeaLifeBase is a global biodiversity information system on non-fishes that covers a wide range of information on taxonomy, biology, trophic ecology, life history and uses (Palomares and Pauly, 2021). The extensiveness of information in the database has catered to a diversity of stakeholders (scientists, researchers, policy-makers, fisheries managers, donors, conservationists, teachers, and students) for various applications targeting sustainable fisheries management (Froese et al., 2018a), ecosystem modelling (Grüss et al., 2019), biodiversity conservation (Stasolla et al., 2021) and environmental protection (Jâms et al., 2020).
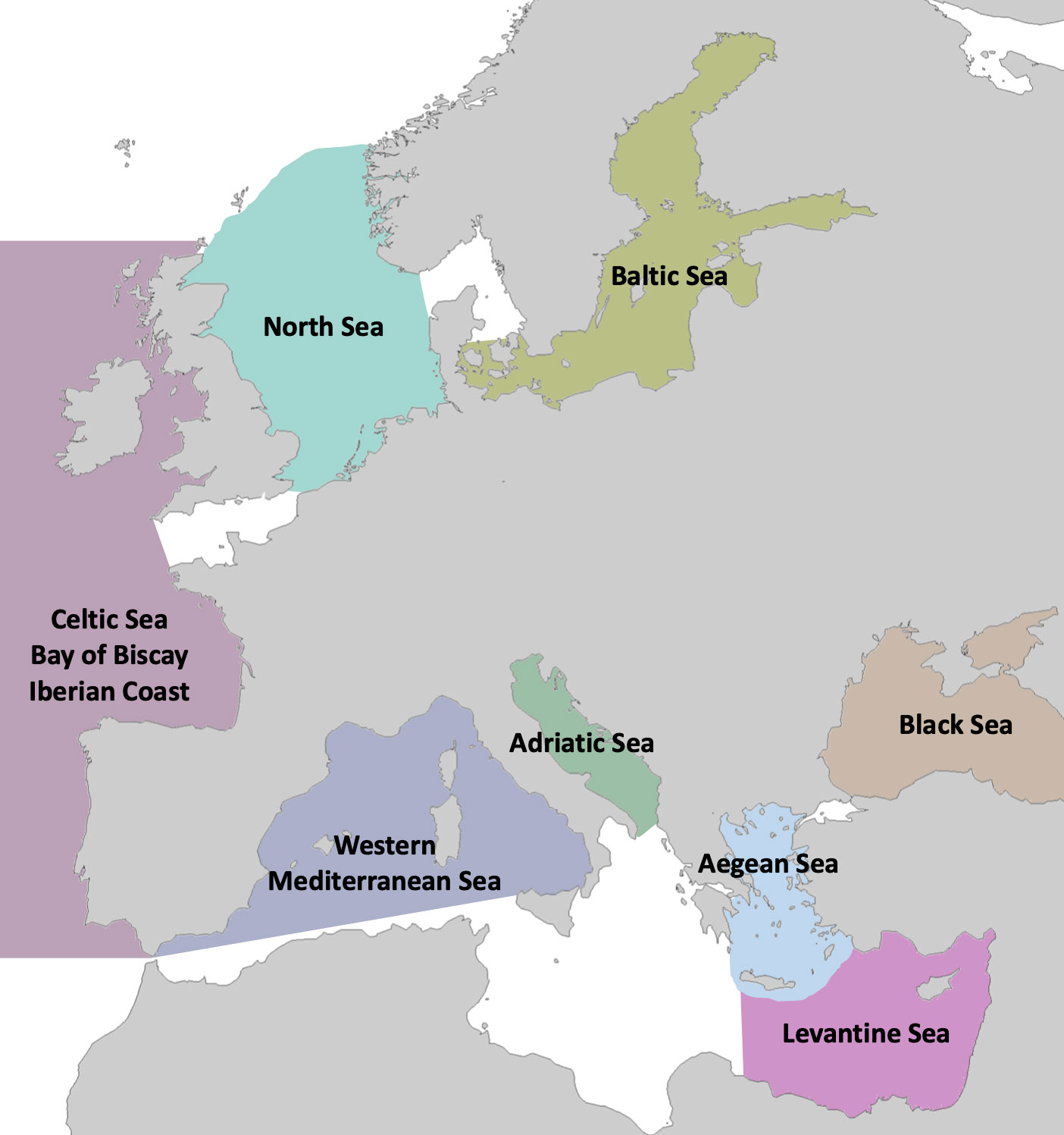
Figure 1 Map of the marine ecosystems that were reviewed.
Eight major groups of marine non-fish species were considered, including cnidarians (corals, jellyfishes, other cnidarians like hydrozoans, hydroids, anemones, and sea pens), crustaceans (decapods, other malacostraca like shrimps, amphipods, isopods, copepods and ostracods), echinoderms (sea cucumbers, sea urchins, starfishes, brittle stars, crinoids, basket stars), molluscs (bivalves, cephalopods, gastropods, chitons, solenogasters and tusk shells), sponges, mammals (dolphins, whales, seals), reptiles (sea turtles), and seabirds.
The working species lists for the review were drawn from a combination of ecosystem, country, and FAO area assignments in SeaLifeBase that approximate the areas covering each of the eight study areas (Palomares and Pauly, 2021). Thus, the species lists for the Adriatic Sea and the Aegean Sea come from the SeaLifeBase faunal records under the Adriatic Sea and Aegean Sea marine ecoregions, whereas those for the Baltic Sea, Black Sea, and North Sea come from the faunal records under the Large Marine Ecosystem (LME) units of the same name (Figure 1). The species list for the western Mediterranean Sea area combines faunal records for the Balearic Islands, Tyrrhenian Sea, Sardinia Island, Corsica Island, as well as marine records for the Mediterranean coasts of Spain and France, i.e., excluding the southern Mediterranean coastline (Figure 1). The list for the Levantine Sea consolidates records from the Levantine Sea ecosystem, marine records for Cyprus, Syria, Lebanon as well as from the side of Israel, Egypt, and Turkey (excluding the Aegean Sea) in FAO area 37. The Bay of Biscay/Celtic Sea/Iberian Coast combines faunal lists for two LMEs, namely, Celtic-Biscay Shelf and Iberian Coast, and thus includes species in an area that extends from the Gulf of Cadiz and north to the Outer Hebrides (Figure 1). An assessment of the representativeness of the species coverage in each site, however, was not within the scope of this work.
The review of biological information covered in SeaLifeBase largely follows the approach of two recent gap analyses reviews on the biology of fishes in the Mediterranean Sea (Dimarchopoulou et al., 2017; Daskalaki et al., 2022).
Eight categories of biological characteristics were examined and include corresponding records in SeaLifeBase: Diet (D) covered diet composition, prey items, and feeding preferences; fecundity (F) included absolute and relative number of oocytes produced per female; maturity covered length/size at first maturity (Lm); spawning (S) looked at onset and duration of spawning (i.e., spawning period); mortality (M) considered the rate of natural mortality regardless of the estimation method; life span (tmax) covered maximum age; growth (G) refers to the growth parameters asymptotic length (L∞) and the rate at which it is approached (K), while length-weight relationships (LWR) considered the slope and intercept of the LWR function (Dimarchopoulou et al., 2017; Daskalaki et al., 2022). Gaps arise from the difference between the level of current knowledge and that of desired knowledge. For the present review, well-studied species were identified as those with available information for at least six out of eight of the biological characteristics described above, and which have 30 or more records available in SeaLifeBase. The desired knowledge for an area was defined as the area with at least half of the reported species being well-studied. The least-studied species were those that do not meet the above criteria (Table 1). Overall, the number of unique references for data on biological characteristics from the literature captured in SeaLifeBase total 637 records (Palomares and Pauly, 2021).

Table 1 Criteria for comparing the knowledge level of areas based on the number of studied biological characteristics and the available number of records for each characteristic.
Species were categorized as threatened/non-threatened according to the species conservation status in SeaLifeBase which follows the IUCN Red List of Threatened Species (version 2021-1) considering the global classification of species (EX: Extinct; EW: Extinct in the Wild; CR: Critically Endangered; EN: Endangered; VU: Vulnerable; NT: Near Threatened; LC: Least Concern; DD: Data deficient; NE: Not Evaluated). For this review, species categorized as threatened included only those flagged as Critically Endangered (CR), Endangered (EN), Vulnerable (VU) and Near-Threatened (NT).
This review assesses how much information on biological traits is available for species, examining species with and without biological information, identifying where data gaps are smallest and widest, and identifying the most and least-studied species. Where a species list is short, full details for the species are included in the table, otherwise the list is summarized according to Order/Family and species count. An overall assessment of information that follows a basic traffic light classification of Good, Moderate and Poor information coverage and the criteria for each category are also provided (Table 1).
Recommendations for filling data gaps are provided in two levels. The first consists of specific recommendations on species and aims at addressing deficiencies or missing information on biological characteristics. The second focuses on filling the gaps with respect to biological characteristics, and particularly on prioritizing species with insufficient or missing biological data that are common to most areas.
A total of 359 non-fish species were recorded for the Adriatic Sea in SeaLifeBase, including 349 native, five endemic and five introduced species that belong to 168 Families, 68 Orders and 15 Classes. Regarding the number of biological characteristics studied, there is no information for 248 species (69%). There are 48 species (13%) with information for only one characteristic (mostly on length-weight relationships), while two species (1%) have studies for all eight biological characteristics (Figure 2).
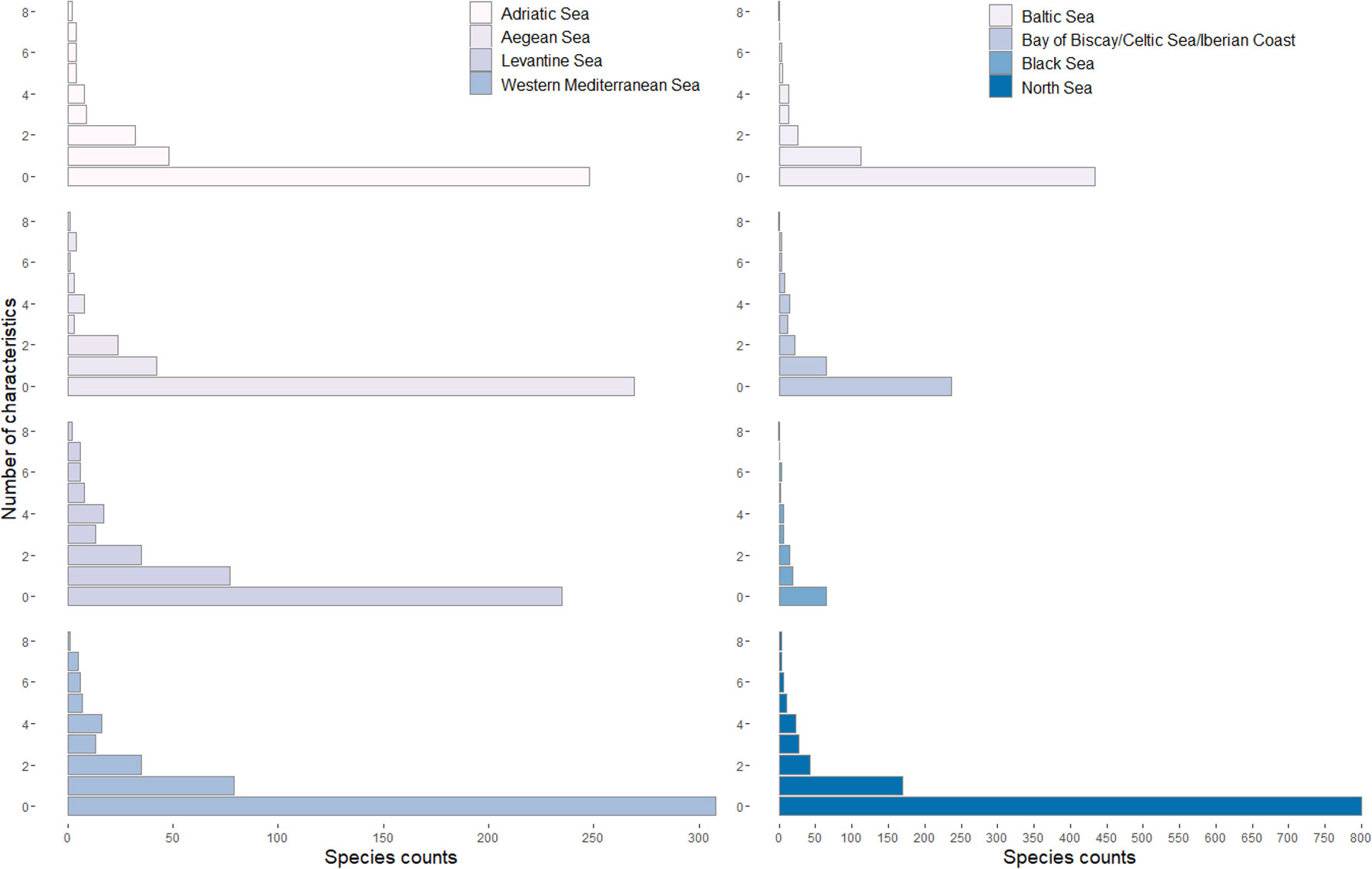
Figure 2 Species counts by number of biological characteristics (0 to 8) studied for non-fish marine organisms in the Adriatic Sea.
The individual biological characteristics of the non-fish species of the Adriatic Sea, from most-studied to least-studied, are: length-weight relationships (97 species, 27%), followed by growth (52 species, 14%), size at maturity (33 species, 9%), spawning (22 species, 6%), maximum age (22 species, 6%), natural mortality (16 species, 4%), fecundity (11 species, 3%), and diet (6 species, 2%) (Figure 3).

Figure 3 (top) Percentage of non-fish species in the Adriatic Sea, Aegean Sea, Black Sea, Levantine Sea Baltic Sea, Bay of Biscay/Celtic Sea/Iberian Coast, North Sea and Western Mediterranean Sea with (dark color) and without (light color) information on biological characteristics: feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR), (bottom) the same percentages calculated for species under IUCN Red List categories near threatened (NT), vulnerable (VU), endangered (EN) and critically endangered (CR).
Five species are included in the IUCN Red List and are listed under the categories CR [Noble pen shell (Pinna nobilis)], VU [Leatherback turtle (Dermochelys coriacea), Common spiny lobster (Palinurus elephas), Horned grebe (Podiceps auritus)], and NT [Dalmatian pelican (Pelecanus crispus)]. These five species have a relatively small gap for one biological characteristic, growth (3 species, 60% studied). This is followed by larger gaps regarding six biological characteristics: length-weight (2 species, 40% studied), maturity (2 species, 40% studied), lifespan (2 species, 40% studied), fecundity (1 species, 20% studied), spawning (1 species, 20% studied), and natural mortality (1 species, 20% studied). The largest data gap is for diet, where no information was available for any of the threatened species (Figure 3).
The most-studied species of the Adriatic Sea make up about 2% (6 species) of non-fish species reported from the area. These species belong to two Classes and six Families (Table 2). The six most-studied Adriatic species in terms of biological characteristics are the Blue crab (Callinectes sapidus), which is an introduced species, and Common cuttlefish (Sepia officinalis), each with available information on eight biological characteristics. Deep-water rose shrimp (Parapenaeus longirostris), Giant red shrimp (Aristaeomorpha foliacea), Spottail mantis shrimp (Squilla mantis) and Common spiny lobster (Palinurus elephas) have seven studied biological characteristics. Out of these six species, only the Common spiny lobster is included in the IUCN Red List as VU (Table 2).
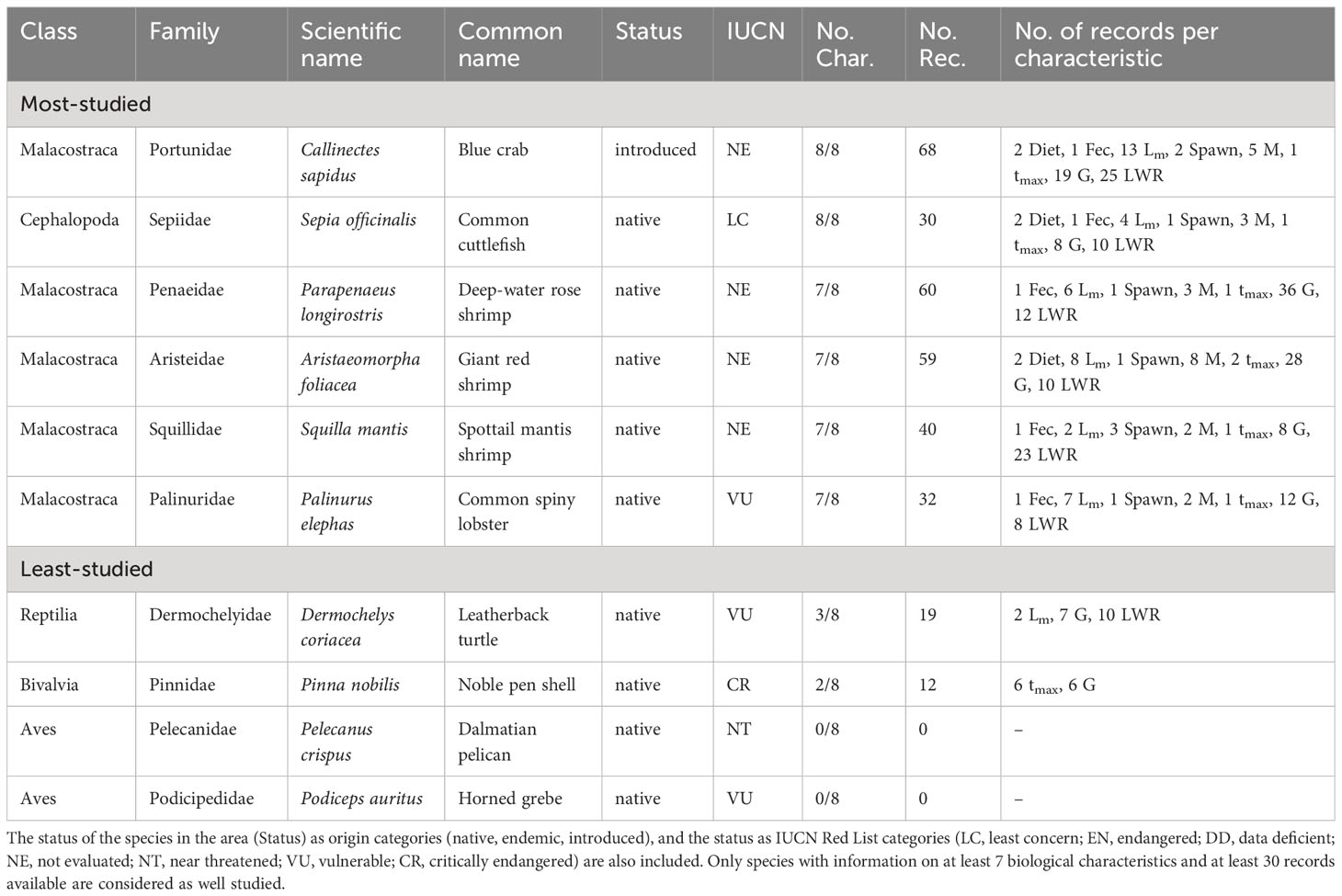
Table 2 List of the most- and least-studied non-fish species in the Adriatic Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec.) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least-studied species make up about 98% (353 species) of non-fish species reported in the Adriatic Sea (15 Classes and 164 Families) (Table S1) including four IUCN Red List species (Table 2).
A total of 355 non-fish species were recorded for the Aegean Sea in SeaLifeBase, including 347 native, six endemic and two introduced species, and belong to 166 Families, 61 Orders and 15 Classes. Regarding the number of biological characteristics studied, there is no information for 269 species (76%). There are 42 species (12%) with information for only one characteristic (mostly length-weight relationships), while for one species all eight biological characteristics are available (Figure 2).
The individual biological characteristics of the non-fish species of the Aegean Sea, from most-studied to least-studied, are length-weight relationships (77 species, 22%), growth (39 species, 11%), maturity (19 species, 5%), spawning (17 species, 5%), lifespan (13 species, 4%), natural mortality (11 species, 3%), fecundity (7 species, 2%), and diet (5 species, 1%) (Figure 2).
Eight species are threatened and listed under the IUCN Red List categories as endangered (EN) and vulnerable (VU), and have large gaps regarding all biological characteristics: length-weight (2 species, 25%), maturity (2 species, 25%), growth (2 species, 25%), fecundity (1 species, 13%), diet (1 species, 13%), spawning (1 species, 13%), natural mortality (1 species, 13%) and lifespan (1 species, 13%) (Figure 3).
The most-studied species of the Aegean Sea make up about 1% (5 species) of non-fish species reported from the area and cover two Classes and five Families (Table 3). The five most studied Aegean species in terms of biological characteristics are the Common cuttlefish (Sepia officinalis) with available information on eight biological characteristics, as well as the Deep-water rose shrimp (Parapenaeus longirostris), Giant red shrimp (Aristaeomorpha foliacea), Spottail mantis shrimp (Squilla mantis) and Common spiny lobster (Palinurus elephas) that have seven studied biological characteristics. Of these, only the Common spiny lobster is included in the IUCN Red List as VU (Table 3).

Table 3 List of the most- and least-studied non-fish species in the Aegean Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least-studied species make up about 99% (350 species) of the non-fish species reported in the Aegean Sea, covering 15 Classes and 162 Families (Table S2). Seven of the least-studied species are included in the IUCN Red List. These include the Mediterranean monk seal (Monachus monachus), listed as EN, with available information on one biological characteristic, the Leatherback turtle (Dermochelys coriacea), listed as VU, with three studied biological characteristics, as well as the coral (Crassophyllum thessalonicae) and the sea anemone (Paranemonia vouliagmeniensis) listed as CR. The Pink sea fan (Eunicella verrucosa), Horned grebe (Podiceps auritus), and, Levantine shearwater (Puffinus yelkouan) are listed as VU, and have no available biological information (Table 3).
A total of 606 non-fish species were retrieved for the Baltic Sea from SeaLifeBase, including 595 native and 11 introduced species. The species of the area belong to 263 Families, 75 Orders and 23 Classes. Regarding the number of biological characteristics studied, there is no information for 434 species (72%). There are 112 species (18%) with information on one biological characteristic (mostly on length-weight relationships) while one species has studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the Baltic Sea, from most-studied to least-studied, are length-weight relationships (140 species, 23%), growth (46 species, 8%), diet (29 species, 5%), lifespan (29 species, 5%), maturity (24 species, 4%), spawning (15 species, 2%), natural mortality (11 species, 2%) and fecundity (7 species, 1%) (Figure 3). Out of all 606 non-fish species reported from the Baltic Sea there are no species that could be considered as well-studied (Table S3). All the Baltic species range from having none to moderately sufficient information on their biological characteristics.
Eight species are listed in IUCN Red List, and have large gaps regarding four biological characteristics: growth (3 species, 33% studied), length-weight relationships (2 species, 22% studied), maturity, (2 species, 22% studied) and diet (2 species, 22% studied). Species listed as VU are Leatherback turtle (Dermochelys coriacea), Sperm whale (Physeter macrocephalus), Long-tailed duck (Clangula hyemalis), Velvet scoter (Melanitta fusca), Horned grebe (Podiceps auritus), and Steller’s eider (Polysticta stelleri)] all of which have no available biological information. Those listed as NT include Eurasian river otter (Lutra lutra) with information on one biological characteristic, and Common eider (Somateria mollissima) with no biological information available (Table 4). There is no available record on these species regarding fecundity, spawning, natural mortality, and lifespan (Figure 3).
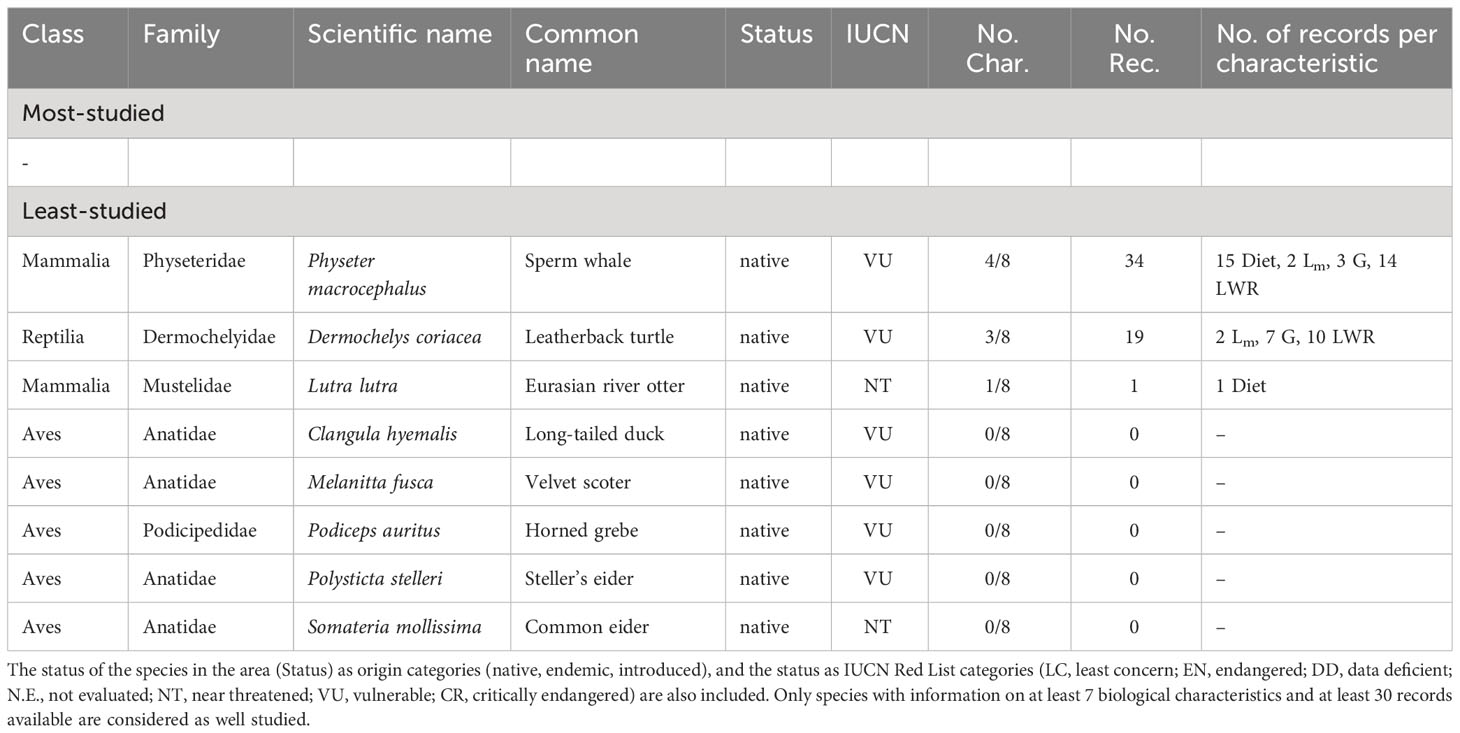
Table 4 List of the most- and least-studied non-fish species in the Baltic Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
A total of 362 non-fish species from the Bay of Biscay/Celtic Sea/Iberian Coast have records in SeaLifeBase, including 356 native and six introduced species. The species within this area belong to 206 Families, 78 Orders and 22 Classes. Regarding the number of biological characteristics studied, there is no information for 236 species (65%), whereas there are 65 species (18%) with information for only one characteristic (mostly on length-weight relationships), and one species has studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the Bay of Biscay/Celtic Sea/Iberian Coast, from most studied to least studied, are length-weight relationships (112 species, 31%), growth (53 species, 15%), maturity (28 species, 8%), diet (24 species, 7%), spawning (22 species, 6%), lifespan (21 species, 6%), natural mortality (11 species, 3%) and fecundity (10 species, 3%) (Figure 3).
Ten species listed under the IUCN Red List have relatively smaller gaps regarding four biological characteristics: length-weight (10 species, 83%), growth (10 species, 83%), diet (7 species, 58%), and maturity (7 species, 58%). Larger gaps were observed in fecundity (3 species, 25%), spawning (3 species, 25%), lifespan (2 species, 17%) and natural mortality (2 species, 17%) (Figure 3). These species are Hawksbill turtle (Eretmochelys imbricata; CR), Kemp’s ridley turtle (Lepidochelys kempii; CR), North Atlantic right whale (Eubalaena glacialis; CR), Sei whale (Balaenoptera borealis; EN), Blue whale (Balaenoptera musculus; EN), Loggerhead turtle (Caretta; VU), Leatherback turtle (Dermochelys coriacea; VU), Fin whale (Balaenoptera physalus; VU), Hooded seal (Cystophora cristata; VU), and Cape Verde petrel (Pterodroma feae; NT).
The most studied non-fish species in the Bay of Biscay/Celtic Sea/Iberian Coast make up about 1% (4 species) and cover three Classes and four Families (Table 5). The four most studied species from the area in terms of biological characteristics were the Common cuttlefish (Sepia officinalis) with information on eight biological characteristics, and the Green sea turtle (Chelonia mydas), Giant red shrimp (Aristaeomorpha foliacea) and Spottail mantis shrimp (Squilla mantis) which have seven studied biological characteristics. Out of these species, only the Green sea turtle is included in the endangered list of IUCN (Table 5).
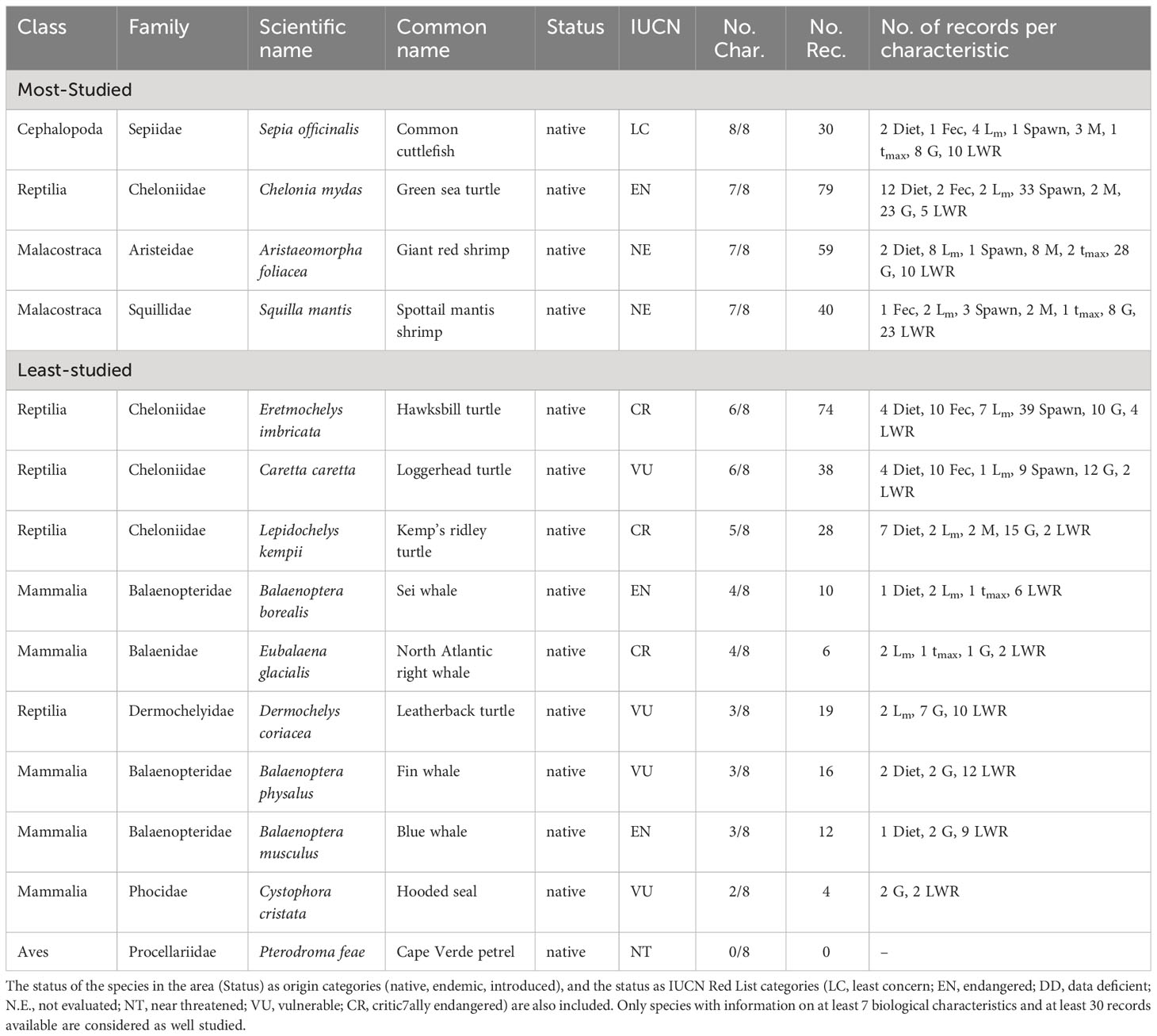
Table 5 List of the most- and least-studied non-fish species in the Bay of Biscay/Celtic Sea/Iberian Coast based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec.) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least studied species make up about 99% (358 species) of non-fish species reported in the Bay of Biscay/Celtic Sea/Iberian Coast, covering 22 Classes and 205 Families (Table S4) including the ten species of the IUCN Red List (Table 5).
A total of 97 non-fish species recorded from the Black Sea in SeaLifeBase, including seven introduced species. The species of the area belong to 57 Families, 40 Orders and 13 Classes. Regarding the number of biological characteristics studied, there is no information for 58 species (56%). There are 19 species (16%) with information on one biological characteristic (mostly on length-weight relationships), whereas one species (1%) has studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the Black Sea, from most studied to least studied, are length-weight relationships (41 species, 35%), growth (30 species, 26%), maturity (14 species, 12%), spawning (9 species, 8%), lifespan (9 species, 8%), diet (9 species, 8%), natural mortality (7 species, 6%), and fecundity (6 species, 5%) (Figure 3).
Five species are listed under the categories CR, endangered EN and vulnerable VU of the IUCN Red List and have large gaps regarding seven biological characteristics: diet (2 species, 33%), growth (2 species, 33%), fecundity (1 species, 17%), maturity (1 species, 17%), spawning (1 species, 17%), lifespan (1 species, 17%) and length-weight relationships (1 species, 17%). The widest information gap refers to natural mortality, where no biological information is available (Figure 3).
The only well-studied non-fish species in the Black Sea is the Blue crab (Callinectes sapidus), an introduced species, having information for all eight biological characteristics (Table 6). The least studied species make up about 96% (93 species) of non-fish species reported in the Black Sea, covering 13 Classes and 57 Families (Table S5). The least-studied species reported from the Black Sea are 28 species in total, covering seven Classes and 23 Families belonging to five taxonomic groups (Table 6). Amongst these species, the Mediterranean monk seal (Monachus monachus) with one biological characteristic and the Loggerhead turtle (Caretta caretta), with six biological characteristics, are listed as endangered (EN) and vulnerable (VU), respectively (Table 6). The former is considered extinct in the Black Sea and the latter is reported in occasional sightings, without an established population. Three species that are listed as VU [Velvet scoter (Melanitta fusca), Horned grebe (Podiceps auritus), and the Levantine shearwater (Puffinus yelkouan)], have no available biological information (Table 6).
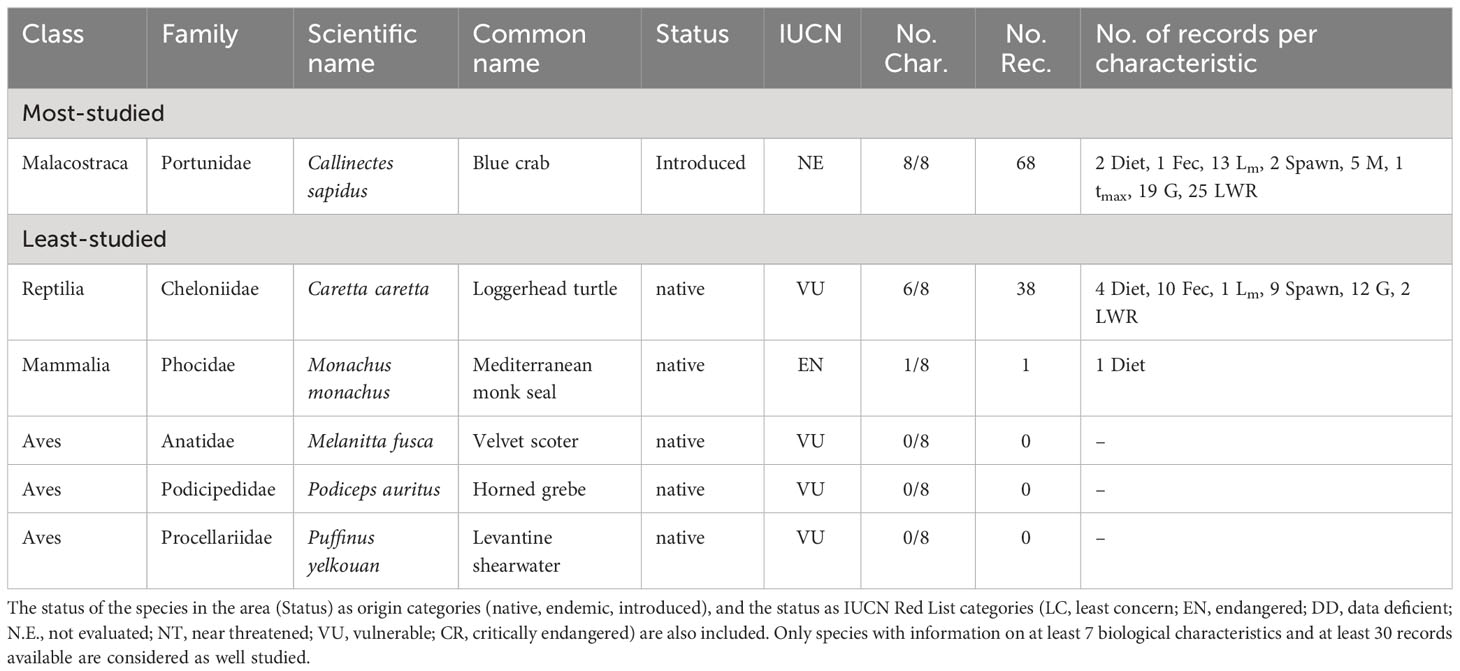
Table 6 List of the most- and least-studied non-fish species in the Black Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
A total of 401 non-fish species recorded from the Levantine Sea are found in SeaLifeBase, including 388 native, one endemic and 12 introduced species that belong to 172 Families, 60 Orders and 17 Classes. Regarding the number of biological characteristics studied, there is no information for 235 species (59%). There are 77 species (19%) with information on one biological characteristic (mostly on length-weight relationships), while two species have studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the Levantine Sea, from most studied to least studied, are length-weight relationships (136 species, 34%), growth (83 species, 21%), maturity (45 species, 11%), spawning (38 species, 9%), lifespan (33 species, 8%), diet (23 species, 6%), natural mortality (22 species, 5%) and fecundity (20 species, 5%) (Figure 3).
Eight species are listed under the IUCN Red List categories near threatened (NT), vulnerable (VU), endangered (EN) and critically endangered (CR) and have smaller gaps regarding four biological characteristics: growth (7 species, 87%), diet (6 species, 75%), maturity (6 species, 75%) and length-weight relationships (6 species, 75%). Larger gaps were observed in spawning (4 species, 50%), fecundity (3 species, 33%), lifespan (3 species, 33%) and natural mortality (2 species, 22%) (Figure 3). These species are the Mediterranean monk seal (Monachus monachus; EN), Loggerhead turtle (Caretta caretta; VU), Sperm whale (Physeter macrocephalus; VU), Leatherback turtle (Dermochelys coriacea; VU), Levantine shearwater (Puffinus yelkouan; VU), Armenian gull (Larus armenicus; NT), False killer whale (Pseudorca crassidens; NT), and Noble pen shell (Pinna nobilis; CR).
The well-studied species of the Levantine Sea make up about 2% (8 species) of non-fish species reported from the area and cover three Classes and seven Families (Table 7). The eight most studied Levantine species in terms of biological characteristics are: the alien Blue crab (Callinectes sapidus) and Common cuttlefish (Sepia officinalis), both having eight biological characteristics, the Speckled shrimp (Metapenaeus monoceros), which is another introduced species, Green sea turtle (Chelonia mydas), Deep-water rose shrimp (Parapenaeus longirostris), Giant red shrimp (Aristaeomorpha foliacea), Spottail mantis shrimp (Squilla mantis) and Common spiny lobster (Palinurus elephas), all of which have seven biological characteristics. Out of these eight species only two are included in the IUCN Red List, the Green sea turtle as EN and the Common spiny lobster as VU (Table 7).
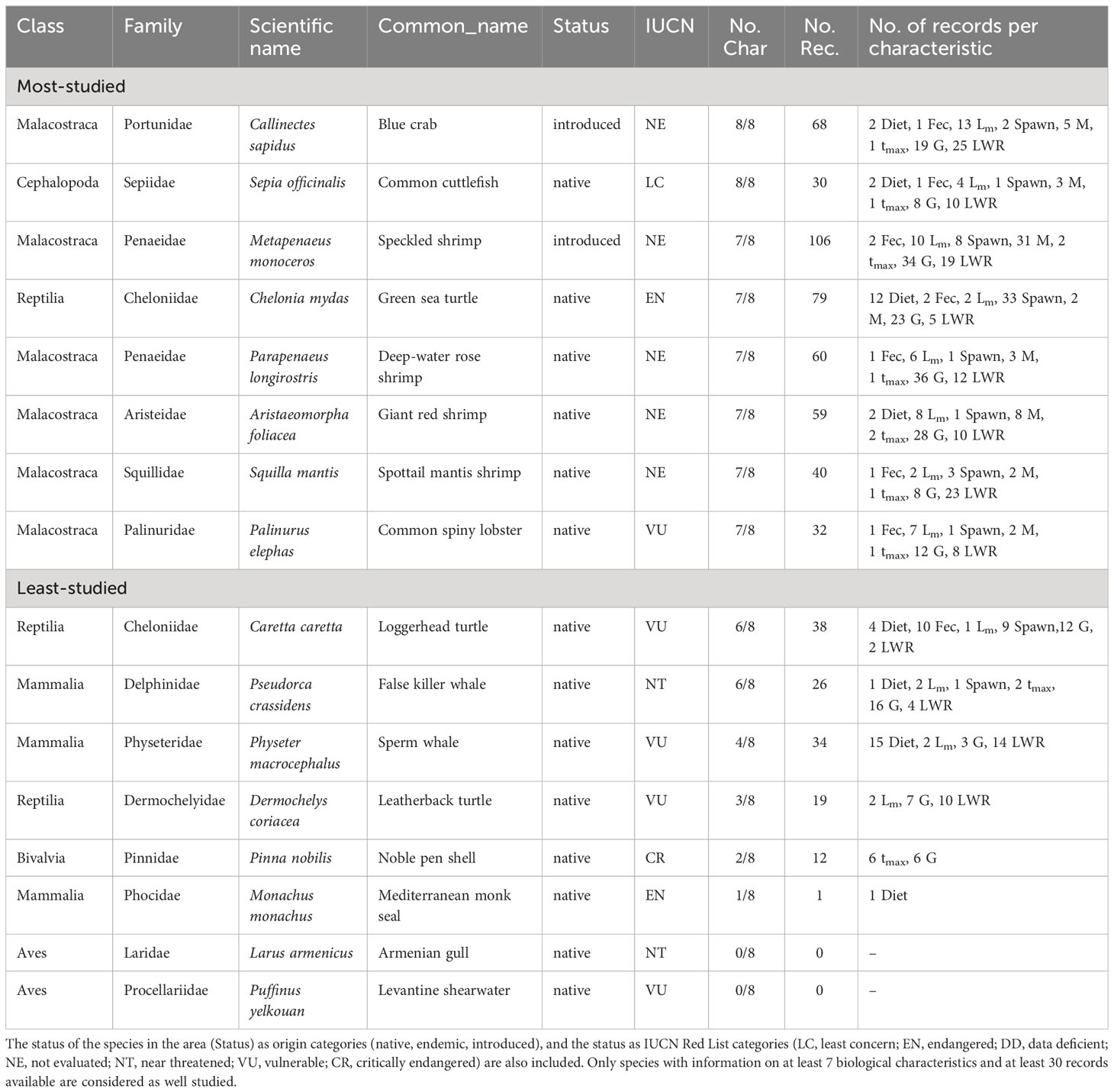
Table 7 List of the most- and least-studied non-fish species in the Levantine Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec.) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least studied species make up about 98% (393 species) of non-fish species reported in the Levantine Sea, covering 17 Classes and 170 Families (Table S6). Including the eight species reported in the IUCN Red List (Table 7).
A total of 1,084 non-fish species were recorded from the North Sea in SeaLifeBase, including 1043 native species and 41 introduced ones. These species belong to 389 Families, 101 Orders and 24 Classes. There is no information on biological characteristics for 800 species (74%). Furthermore, there are 170 species (16%) with information on one biological characteristic (mostly on length-weight relationships), and three species have studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the North Sea, from most studied to least studied, are length-weight relationships (216 species, 20%), growth (112 species, 10%), diet (55 species, 5%), maturity (49 species, 5%), lifespan (48 species, 4%), spawning (36 species, 3%), natural mortality (24 species, 2%) and fecundity (18 species, 2%) (Figure 3).
Twenty-two species are listed under the categories near threatened (NT), vulnerable (VU), endangered (EN) and critically endangered (CR) of the IUCN Red List. These species show smaller gaps regarding two biological characteristics: growth (16 species, 73%) and length-weight relationships (14 species, 64%), but larger gaps were observed in diet (10 species, 45%), maturity (9 species, 41%), fecundity (4 species, 18%), spawning (4 species, 18%), natural mortality (3 species, 14%) and lifespan (3 species, 14%) (Figure 3). The twenty-two species that are included in IUCN Red List are Hawksbill turtle (Eretmochelys imbricata; CR), Kemp’s ridley turtle (Lepidochelys kempii; CR), North Atlantic right whale (Eubalaena glacialis; CR), Sei whale (Balaenoptera borealis; EN), Blue whale (Balaenoptera musculus; EN), Loggerhead turtle (Caretta; VU), Sperm whale (Physeter macrocephalus; VU), Leatherback turtle (Dermochelys coriacea; VU), Fin whale (Balaenoptera physalus; VU), Walrus (Odobenus rosmarus; VU), Hooded seal (Cystophora cristata; VU), North Atlantic bottlenose whale (Hyperoodon ampullatus; NT), Balearic shearwater (Puffinus mauretanicus; CR), Black-legged kittiwake (Rissa tridactyla; VU), Atlantic puffin (Fratercula arctica; VU), Velvet scoter (Melanitta fusca; VU), Horned grebe (Podiceps auritus; VU), Steller’s eider (Polysticta stelleri; VU), Starlet anemone (Nematostella vectensis; VU), Sooty shearwater (Puffinus griseus; NT), Red knot (Calidris canutus; NT) and Common eider (Somateria mollissima; NT).
The most studied species of the North Sea make up about 0.5% (5 species) of non-fish species reported in the area and cover four Classes and five Families (Table 8). These five species are the alien Blue crab (Callinectes sapidus and Common cuttlefish (Sepia officinalis), with information on eight biological characteristics, the Japanese carpet shell (Ruditapes philippinarum), which is another introduced species, Green sea turtle (Chelonia mydas) and Common spiny lobster (Palinurus elephas) all have information on seven biological characteristics. Out of these five species, the Green sea turtle is listed as EN and the Common spiny lobster as VU (Table 8).
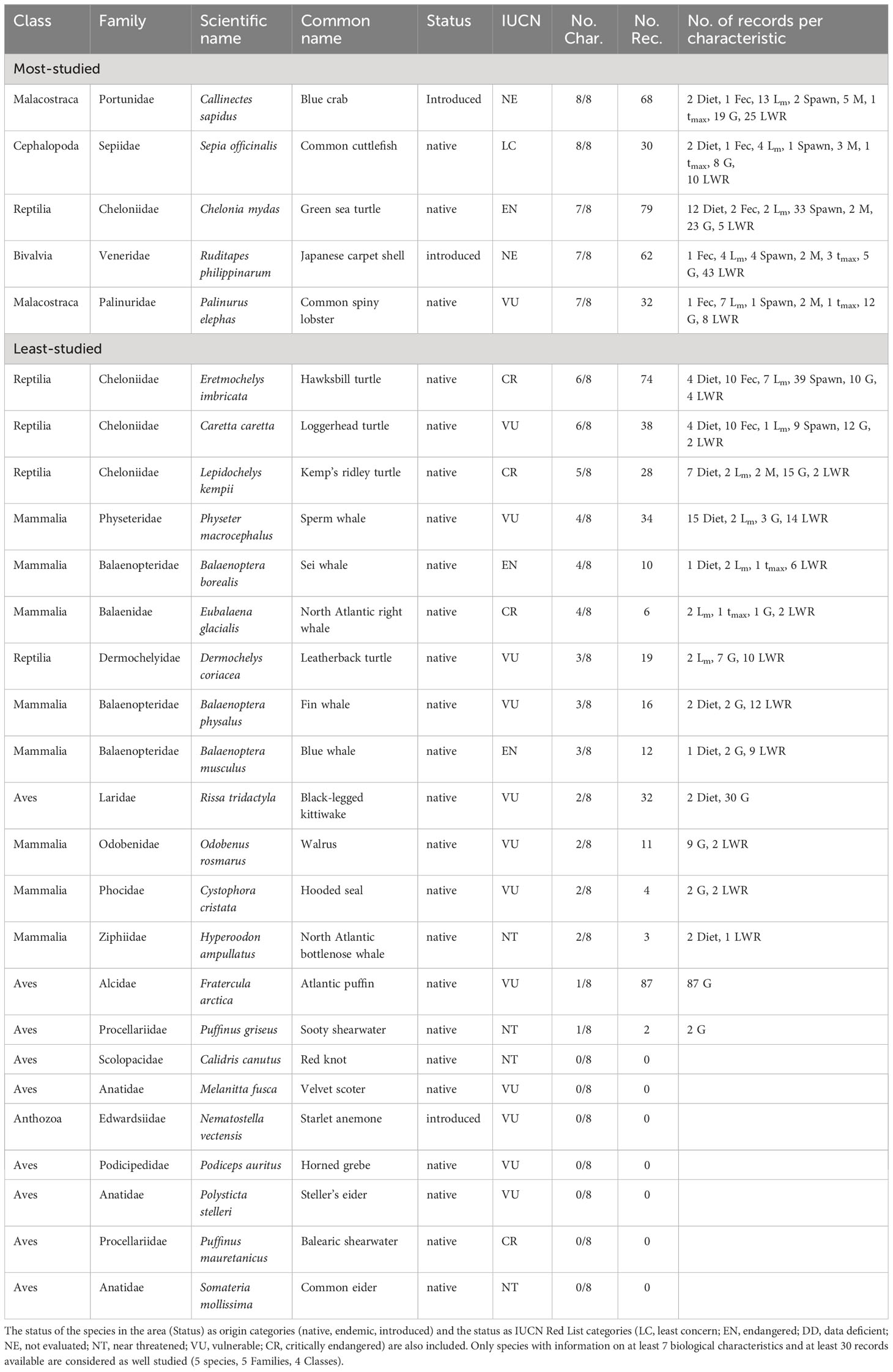
Table 8 List of the most- and least-studied non-fish species in the North Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least studied species make up about 99.5% (1079 species) of non-fish species reported in the North Sea and cover 24 Classes and 387 Families (Table S7) including the 22 species reported in the IUCN Red List (Table 8).
A total of 470 non-fish species are recorded from the Western Mediterranean Sea in SeaLifeBase, including 462 native, two endemic and six introduced species, belonging to 210 Families, 73 Orders and 19 Classes. Regarding the number of biological characteristics studied, there is no information for 308 species (66%). There are 79 species (17%) with information on one biological characteristic (mostly on length-weight relationships), while only one species has studies for all eight biological characteristics (Figure 2).
The individual biological characteristics of the non-fish species of the Western Mediterranean Sea, from most studied to least studied, are: length-weight relationships (136 species, 29%), followed by growth (80 species, 17%), maturity (42 species, 9%), spawning (29 species, 6%), lifespan (26 species, 6%), natural mortality (22 species, 5%), fecundity (16 species, 3%) and diet (15 species, 3%) (Figure 3).
Thirteen species are included in the IUCN Red List under the categories near threatened (NT), vulnerable (VU), endangered (EN) and critically endangered (CR). These species have a small gap regarding one biological characteristic (growth) with information available for 10 species (67%). Larger gaps were observed for the rest biological characteristics: length-weight relationships (7 species, 47%), maturity (7 species, 47%), diet (4 species, 27%), fecundity (4 species, 27%), spawning (4 species, 27% studied), natural mortality (3 species, 20%), and lifespan (2 species, 13%) (Figure 3). The thirteen included in the IUCN Red List are Hawksbill turtle (Eretmochelys imbricata; CR), Kemp’s ridley turtle (Lepidochelys kempii; CR), Loggerhead turtle (Caretta caretta; VU), Leatherback turtle (Dermochelys coriacea; VU), Olive ridley turtle (Lepidochelys olivacea; VU), Noble pen shell (Pinna nobilis; CR), Balearic shearwater (Puffinus mauretanicus; CR), Audouin’s gull (Larus audouinii; VU), Velvet scoter (Melanitta fusca; VU), Horned grebe (Podiceps auritus; VU), Levantine shearwater (Puffinus yelkouan; VU), Sooty shearwater (Puffinus griseus; NT) and Common eider (Somateria mollissima; NT).
The most studied species of the western Mediterranean Sea make up about 1% (6 species) of non-fish species reported from the area and cover three Classes and six Families (Table 9). The six most studied western Mediterranean species in terms of biological characteristics were the Common cuttlefish (Sepia officinalis), the only species with eight studied biological characteristics, while the Green sea turtle (Chelonia mydas), Deep-water rose shrimp (Parapenaeus longirostris), Giant red shrimp (Aristaeomorpha foliacea), Spottail mantis shrimp (Squilla mantis) and Common spiny lobster (Palinurus elephas) have seven biological characteristics studied. Out of these six species, only the Common spiny lobster is included in the list of IUCN as VU (Table 9).
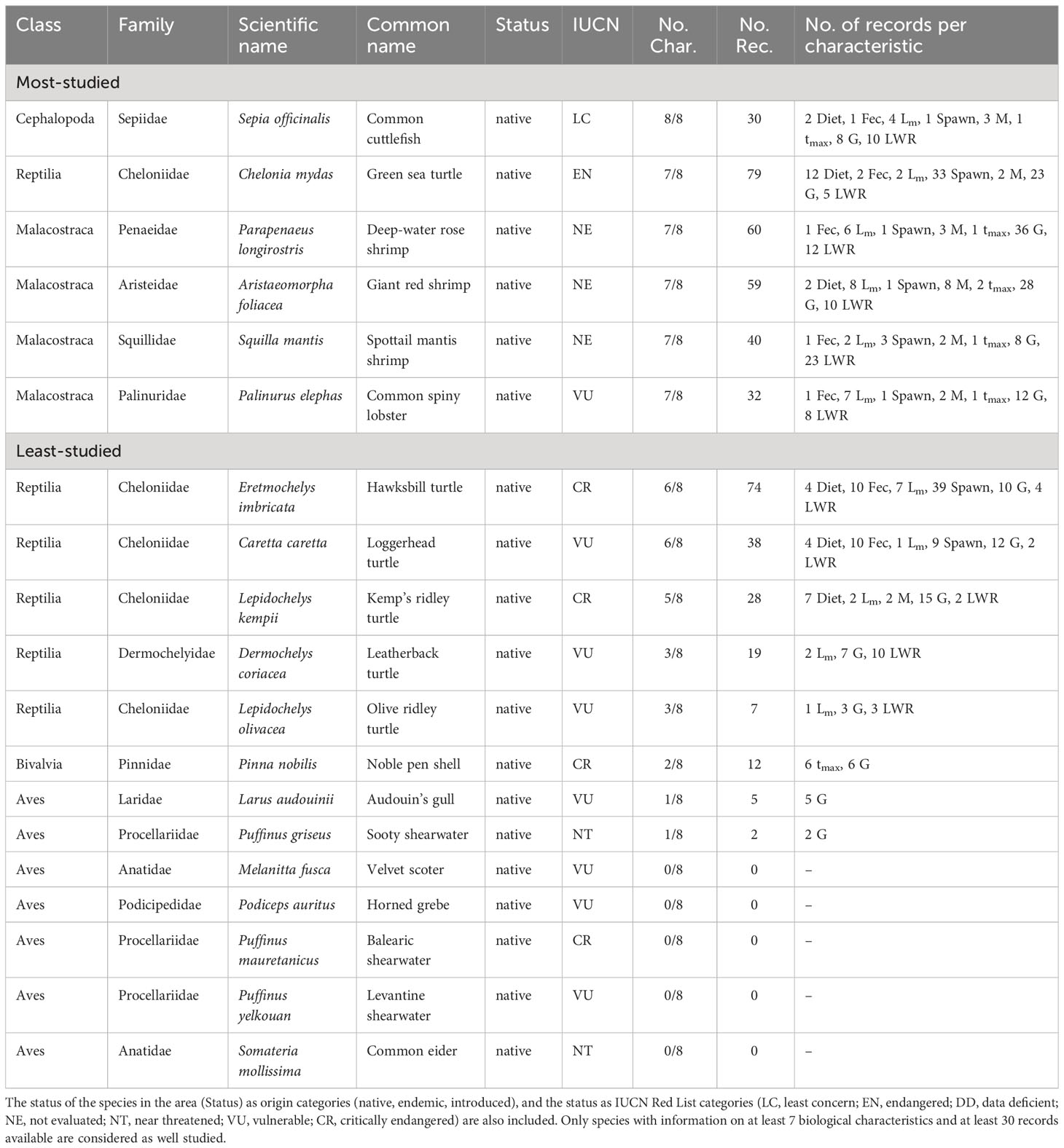
Table 9 List of the most- and least-studied non-fish species in the western Mediterranean Sea based on the number of studied biological characteristics (No. Char.) and the number of records (No. Rec.) per characteristic (feeding preferences (Diet), fecundity (Fec), maturity (Lm), spawning (Spawn), mortality (M), lifespan (tmax), growth (G), and length-weight relationships (LWR).
The least-studied species make up about 98% (464 species) of non-fishes reported in the western Mediterranean Sea, cover 19 Classes and 210 Families (Table S8). Of the least-studied species, there are thirteen species that are included in the IUCN Red List (Table 9).
The general pattern, observed across all studied ecosystems, is that data availability on biological characteristics of non-fish marine organisms are rather poor, with only two taxonomic groups (sea turtles and marine mammals) appearing to have been adequately studied across most study areas. There is moderately good information coverage for sea turtles in the Celtic Sea/Bay of Biscay/Iberian coast, Black Sea, Levantine Sea, North Sea and western Mediterranean Sea, whereas information coverage for marine mammals is moderate for the Baltic Sea, Celtic Sea/Bay of Biscay/Iberian coast, Black Sea, Levantine Sea, North Sea and western Mediterranean Sea (Table 10). Data on biological characteristics was lowest for cnidarians, whereas there were no available biological information on sponges; the latter group of organisms being globally understudied in terms of biological characteristics (Bell et al., 2015).

Table 10 Comparison regarding the status of the studied groups in European Seas and adjacent waters.
Because of the low total number of species recorded compared to the other areas, the Black Sea appears to be among the better studied areas together with the Bay of Biscay/Celtic Sea/Iberian Coast, the Levantine Sea and the western Mediterranean Sea. The Adriatic Sea is the area with the most data gaps compared to the other study areas. This area, for instance, does not have information available on any of the present marine mammal species (Lotze et al., 2011). Likewise, there are no data reported on the biological characteristics of sponges in the Black Sea, where their checklist has been recently updated, at least for part of this area (Topaloglu and Alper, 2014). The difference in species composition among areas has certainly contributed to the number of species studied and the extent of the available information.
There are seven well-studied species across the reviewed ecosystems: Common cuttlefish (Sepia officinalis), Giant red shrimp (Aristaeomorpha foliacea), Spottail mantis shrimp (Squilla mantis), Common spiny lobster (Palinurus elephas), Blue crab (Callinectes sapidus), Green sea turtle (Chelonia mydas) and Deep-water rose shrimp (Parapenaeus longirostris). These species have good coverage of biological information and sufficient data records for use in ecosystem assessments and modelling. Of these, the most common well-studied species include the Common cuttlefish (Sepia officinalis), which is reported within six sites and is highly commercial (Pereira et al., 2019). The current absence of data for the studied taxonomic groups may be due to actual absence of real data (i.e., lack of studies on non-fish marine organisms) or time-lagged entering of research publications in SeaLifeBase or that source of information has not been considered (for instance, grey literature or local journals).
In terms of biological characteristics, the information gap for all species is largest for fecundity, natural mortality and diet, with the better studied characteristics being length-weight relationships (LWR) followed by spawning, lifespan, maturity and growth. The most-studied characteristic (LWR) is common and well-studied across areas but the least-studied ones differ between the Atlantic and the Mediterranean areas. Consequently, research priorities and survey data availability often differ (e.g., Ugland, 1976), as well as to scientific tradition and historical data records (Lotze and Worm, 2009) that are generally scarce in the Mediterranean (Stergiou and Tsikliras, 2006; Fortibuoni et al., 2017). It is worth mentioning here that LWR is the most common even though it is not a trait that is measured for many non-fish taxonomic groups such as marine mammals, reptiles and seabirds. Contrary to LWR that are easier to collect and compute, the sample collection and laboratory work required to determine the diet and fecundity of specimens are costly and time-consuming and require technological equipment and advanced expertise (Dimarchopoulou et al., 2017). In contrast, natural mortality can be easily calculated using existing datasets based on the many known empirical equations that are available (constant across ages/sizes: Pauly, 1980; Then et al., 2015; size/age-based: Chen and Watanabe, 1989; Gislason et al., 2010) without any extra cost or sampling that would be required if other methods were selected (tagging: Krause et al., 2020; length-based and age structured models: Lorenzen, 2022). Therefore, it is strongly recommended, at least for decapod crustaceans and cephalopods, to report maximum age in every study in which growth parameters are determined and, if possible, to calculate and report natural mortality.
For the species that are exploited such as many crustaceans and cephalopods, commercially targeted species are indeed better-studied compared to by-catch and discarded ones that are generally neglected (Baran, 2002). The biological information of the former is more complete due to historically more intensive sampling effort across the studied marine ecosystems because of their economic importance to the fisheries and frequent assessments (Dimarchopoulou et al., 2017). For non-commercial groups, there is generally less information on threatened species compared to those with high commercial value, as it has recently reported for fishes (Dimarchopoulou et al., 2017; Daskalaki et al., 2022) due to the low accessibility of deep-water non-fish marine species that are generally less sampled in routine surveys that rarely extend to deep waters (Sardà et al., 2004). Therefore, the study of threatened, deep-water and non-indigenous species should be prioritized over the well-studied commercial species similar to recommendations for marine fishes (Daskalaki et al., 2022). When threatened species cannot be sampled with non-destructive methods, such as underwater censuses or tagging experiments, it is suggested that if dead after capture, the specimens should be exhaustively studied across their biological characteristics to ensure the maximum economy of sampling (Dimarchopoulou et al., 2017). The study of threatened species should be a priority as they are all good candidates for field data collection. However, the existing gaps on species that are routinely sampled during scientific surveys should also be considered by scientists. The basic characteristics of a species (measurement of length and weight) should always be recorded even from single individuals in the market (in the case of crustaceans and cephalopods) or stranded individuals in the case of marine mammals and reptiles (see the importance of single specimen characteristics for sharks in Tsikliras and Dimarchopoulou, 2021).
Better research coverage on the diets of all marine organisms would greatly benefit future ecosystem models and improve future versions of the current ones (Dimarchopoulou et al., 2017), while more studies on growth, mortality, maturity and spawning of exploited populations will improve the quality of stock assessments within the framework of STECF and GFCM. This, in turn, will reduce uncertainty on the outcome of stock assessment and ecosystem models and will eventually lead to improvements in ecosystem based fisheries management, especially in the Mediterranean and the Black Sea (Rodriguez-Perez et al., 2023). In many areas that experience an influx of non-indigenous species, such as the eastern Mediterranean Sea (Galil et al., 2015), the study of non-indigenous species biology should also be prioritized. Their biological characteristics in the new habitats/areas should be compared to those in native range, aiming to identify the potential effects of alien species on local populations, habitats and communities (Daskalaki et al., 2022).
In the Adriatic Sea, there is currently poor biological information coverage for non-fish species, with relatively more information being available for sea turtles compared to crustaceans, echinoderms and cnidarians, while no data exists for marine mammals (Table 10). Only sea turtles qualify as moderate data coverage with all other areas being data poor (Table 10). In general, the gap is widest for information on diet and fecundity. The information coverage for the Adriatic is good for only a few well-studied species of crustaceans and one cephalopod that are commercially important.
Compared to other Mediterranean areas, the Adriatic Sea is an overall well-studied ecosystem in terms of stock assessments (Froese et al., 2018b) and ecosystem models (Barausse et al., 2009), with important contributions on the effects of fishing (Coll et al., 2007), filling gaps in survey datasets (Coro et al., 2022), the effect of COVID-19 on fish stocks (Scarcella et al., 2022) and fisheries in general, especially in the western part of the sea (Lotze et al., 2011). There are even some historical data available for large marine animals (Lotze and Worm, 2009). Non-indigenous species have also been extensively studied in terms of their effect on the food web dynamics (Libralato et al., 2010; Libralato et al., 2015). It appears that the data collected from scientific surveys on non-fish marine organisms either remain unpublished, or do not include the biological characteristics covered in this review. Furthermore, they have potentially not yet been included in SeaLifeBase.
Within the Aegean Sea, Sea turtles have better coverage, in terms of biological characteristics, compared to all other groups, (Table 10). However, similarly to the Adriatic, only sea turtles qualify as moderately studied while in all other areas are poorly studied (Table 10). The knowledge gap is widest for information on diet followed by fecundity. Good biological information is available for a few well-studied species of crustaceans and one cephalopod, all being commercially exploited.
Official stock assessments are generally scarce in the Aegean Sea and cover only a handful of commercial stocks owing to several years missing from data collection framework (Tsikliras et al., 2021). Nevertheless, over 100 fish and invertebrate Aegean Sea stocks have been recently assessed using data-poor methods (Froese et al., 2018b; Tsikliras et al., 2021). Several EwE ecosystem models have been recently developed for parts of the Aegean Sea (Thracian Sea: Tsagarakis et al., 2010; Pagassitikos Gulf: Dimarchopoulou et al., 2019; Thermaikos Gulf: Dimarchopoulou et al., 2022) and a recent one for the entire Aegean Sea (Keramidas et al., 2022) along with temporal simulations (Papantoniou et al., 2021; Dimarchopoulou et al., 2022), while spatial models are still not available (Keramidas et al., 2023). Besides overfishing, which is considered the main driver of exploited marine populations in the Mediterranean Sea (Dimarchopoulou et al., 2021), the direct (sea warming and species distribution shifts) and indirect (entering and expansion of alien species) effects of climate change are major issues in the eastern Mediterranean Sea that concern the scientific community (Cherif et al., 2020).
Although there are no well-studied species in the Baltic Sea, marine mammals and echinoderms have higher counts of species with studied biological characteristics compared to molluscs, crustaceans, seabirds, and cnidarians (Table 10). The biological information coverage is moderate for sea turtles and marine mammals and poor for all other groups (Table 10). The knowledge gap is widest for information on fecundity and natural mortality and narrowest for length-weight relationships. An introduced species, Harris mud crab (Rhithropanopeus harrisii) is considered as a near well-studied species, having eight biological characteristics and 17 records available.
The Baltic Sea is a well-studied ecosystem (Feistel et al., 2008) with many stock assessments available (Froese et al., 2018b; Froese et al., 2021) and ecosystem models using various approaches (Österblom et al., 2007; Bauer et al., 2019) that geographically cover basins of the entire sea (see Scotti et al., 2022 and references in their Table S2). Status and dynamics of several ecosystem components have been studied in the Baltic Sea including hypotheses on alien species (Dobrzycka-Krahel and Medina-Villar, 2020) and their effect on ecosystem services (Ojaveer et al., 2023), eutrophication (Bauer et al., 2019), fisheries (Scotti et al., 2022) but also climate change (Niiranen et al., 2013) and grey seal (Halichoerus grypus) interaction with fisheries (Costalago et al., 2019). The study of non-indigenous species, which are numerous in the Baltic Sea (Reusch et al., 2018), and their effects on marine ecosystems should be prioritized. With such a wealth of biotic and abiotic information on the ecosystems of the Baltic Sea ecosystems, with long-term datasets of many marine groups available since the 1950s and some expeditions dating back to 1850s (Ojaveer et al., 2010), it is surprising that the basic biological characteristics for many marine organisms supporting ecosystem models and assessments have not been published. The possibility that this information is published but has not yet been scrutinized by SeaLifeBase is also a potential explanation especially in case of local or not yet digitized journals.
In the Bay of Biscay/Celtic Sea/Iberian Coast (combined), vertebrates (marine mammals, sea turtles and seabirds) have higher counts of species with biological information compared to invertebrates (echinoderms, molluscs, crustaceans and cnidarians) (Table 10). The biological information coverage is good for sea turtles, moderate for marine mammals and poor for all other groups (Table 10). The knowledge gap is widest for information on fecundity and natural mortality and narrowest for length-weight relationships. In the Bay of Biscay/Celtic Sea/Iberian Coast area, there is good biological information for a few well-studied species of crustaceans, one cephalopod (all commercial) and one species of sea turtle.
The three combined areas of the NE Atlantic (Celtic Sea, Bay of Biscay, Iberian Coast) are all high biodiversity areas with many habitats and marine species, including endangered and protected species like cetaceans (Laran et al., 2017; Spitz et al., 2018) and seabirds (Morley et al., 2016). The Celtic Sea and Bay of Biscay are often considered as a single ecosystem in modelling studies (Moullec et al., 2017). They are all rich in terms of scientific output (Borja et al., 2011), with many ecosystem models developed, simulated (Lassalle et al., 2011; Corrales et al., 2022) and compared (Moullec et al., 2017). Several ecological hypotheses have been examined based on ecosystem and ecological models (Le Marchand et al., 2020). The number of stocks that have been assessed covers the majority of commercial fisheries (Guénette and Gascuel, 2012; Froese et al., 2018a; Froese et al., 2021).
Marine mammals and molluscs have higher counts of species with biological information compared to crustaceans and cnidarians in the Black Sea (Table 10). The biological information coverage is good for sea turtles, moderate for marine mammals and poor for all other groups (Table 10). The biological knowledge gap is widest for information on fecundity and natural mortality and narrowest for length-weight relationships. An introduced crustacean, Blue crab (Callinectes sapidus) has sufficient information for eight biological characteristics and is considered the best studied organism in the Black Sea of those with records in SeaLifeBase.
The Black Sea together with the Mediterranean marine ecosystems are rather poorly studied compared to the NE Atlantic ones (Güneroğlu et al., 2019). However, the Black Sea ecosystem structure (Akoglu et al., 2014) and fisheries (Prodanov et al., 1997; Daskalov, 2002; Gucu, 2002) are relatively well studied in certain parts of the sea. The effect of non-indigenous species on the populations and ecosystem of the Black Sea (Shiganova, 1998) and the overall status of the ecosystem after anthropogenic impacts has been evaluated (Zaitsev, 1992; Kideys, 2002; Daskalov et al., 2017). Furthermore, some of the commercial fish and invertebrate Black Sea stocks have been assessed using catch-based (Tsikliras et al., 2015) and other data-limited methods (Froese et al., 2018a; Daskalov et al., 2020; Demirel et al., 2020).
In the Levantine Sea, marine mammals, sea turtles, seabirds and crustaceans have higher counts of species with information on biological characteristics compared to echinoderms, molluscs and cnidarians (Table 10). The biological information coverage is good for sea turtles, moderate for marine mammals and poor for all other groups (Table 10). In general, the knowledge gap is widest for information on fecundity and natural mortality. Overall, current coverage on biological information is good for a few well-studied species of crustaceans, one sea turtle and one cephalopod.
Due to its proximity to the Suez Canal, the Levantine Sea is the first to receive the non-indigenous species migrating into the Mediterranean Sea from the Red Sea, which have altered the biodiversity of the area (Galil et al., 2015). Despite the existence of local journals with long publishing history in the area (Israel Journal of Ecology & Evolution/Israel Journal of Zoology, published since 1963; Turkish Journal of Zoology, published since 1977; Egyptian Journal of Aquatic Biology and Fisheries, published since 1997) and some recent attempts (e.g., Syrian Journal of Agricultural Research since 2014), the amount of data on the biology of non-fish marine organisms is rather limited (Stergiou and Tsikliras, 2006) and the Syrian coast has been characterized as one of the least-studied areas for marine mammals in the Mediterranean Sea (Saad and Mahfoud, 2022). Although the extent of scientific surveys is rather limited and historical biological data are generally lacking or concentrated in specific countries (Tsikliras et al., 2010), there has been an increase of scientific output in the Levantine Sea during the last decades (Tsikliras and Stergiou, 2014). Despite the data deficiencies and the lack of long time series, EwE ecosystem models have been developed to examine the effect of non-indigenous species, climate change and other anthropogenic affects in Israel (Corrales et al., 2017; Corrales et al., 2018; Shabtay et al., 2018; Grossowicz et al., 2020; Ofir et al., 2023) in addition to bioeconomic models (Peled et al., 2020; Michael-Bitton et al., 2022) and non-indigenous species in Cyprus (Michailidis et al., 2019), including lionfish Pterois miles (Savva et al., 2020) and silver-cheeked toadfish Lagocephalus sceleratus (Ulman et al., 2021).
Similarly to the Bay of Biscay/Celtic Sea/Iberian Coast (combined), marine vertebrates (sea turtles, marine mammals and seabirds) have higher counts of species with information on biological characteristics compared to the marine invertebrates (echinoderms, molluscs, crustaceans and cnidarians) (Table 10). Biological coverage is good for sea turtles, moderate for marine mammals and poor for all other groups (Table 10). In general, the knowledge gap is widest for information on fecundity and natural mortality, and narrowest for length-weight relationships. Overall, the current coverage on biological information is good for a few well-studied species of crustaceans, sea turtle, cephalopod and Harris mud crab (Rhithropanopeus harrisii), an introduced non-commercial species, is considered a near well-studied species, having eight biological characteristics and 17 records available.
The North Sea marine ecosystem is one of the most biotically-rich and productive seas in Europe (Quante et al., 2016) and has been well studied for many decades with respect to ecosystem structure (Stäbler et al., 2018), effects of fishing and climate (Heath, 2005), system dynamics (Luczak et al., 2012), and regime shifts (Beaugrand, 2004), as well as ecological models (Fransz et al., 1991). There are several ecosystem models available for the North Sea (Burkhard et al., 2011; Mackinson et al., 2018), including temporal (Mackinson et al., 2009), spatial (Püts et al., 2020) and bioeconomic (Beattie et al., 2002) models, while the majority of commercial fish and invertebrate stocks are being regularly assessed (Froese et al., 2021).
Marine mammals and sea turtles have higher counts of species with studied biological characteristics compared to seabirds, crustaceans, molluscs, echinoderms and cnidarians in the western Mediterranean Sea with good biological information coverage for sea turtles, moderate for marine mammals and poor for all other groups (Table 10). In general, the knowledge gap is widest for information on diet followed by fecundity. Overall, the current coverage on biological information is better for a few well-studied species of crustaceans, one sea turtle and one cephalopod.
Parts of the western Mediterranean Sea, especially the northern coastline, have been well studied in terms of ecosystem modelling (Catalan Sea: Coll et al., 2006; Coll et al., 2008; Gulf of Lions: Vilas et al., 2021), even in deep waters (Tecchio et al., 2013), and invertebrate stock assessments (Froese et al., 2018a). Specific aspects of the biology of many marine taxonomic groups have been studied in various parts of the area (feeding/crustaceans: Cartes et al., 2002; maturity/cephalopods: Quetglas et al., 2010; cnidarians/growth and spawning: Rosa et al., 2013). This is partly due to the presence of scientific journals in the area with long publication history (e.g., Scientia Marina published since 1955 as Investigación Pesquera) devoted to the biology of marine organisms and of course due to the long and consistent scientific tradition of western Mediterranean countries in marine sciences. It should be noted here that the southern Mediterranean countries have a long scientific tradition in fisheries and marine biology (Stergiou and Tsikliras, 2006) and have produced significant scientific output on the biology of marine populations for over a century (Tsikliras et al., 2010).
In order to reduce knowledge gaps on the biology of non-fish marine species across European Seas, future research should focus on species with insufficient or missing biological data that are common to the majority of the studied areas such as sea turtles, monk seal and seabirds; more effort is generally required for the Adriatic and the Aegean Seas. Invertebrate species with low or no commercial value that are often collected in scientific surveys and/or as by-catch in commercial fisheries should not be overlooked. Long-lived species should be prioritized in order to understand their biology and potential threats to their populations other than fishing. In areas invaded by non-indigenous species, such as the eastern Mediterranean Sea, research should be focused on the study of life-history characteristics of these species in their new environment and a comparison with their habits in their native distribution. Besides overfishing and incidental fishing, climate change is one of the major threats to marine life and the response of marine populations to climate effects is directly related to their population characteristics and thermal preferences. Knowledge of the latter, which today is known only for a small proportion of marine species, will improve species distribution models and the understanding of climate effects. Threatened species that are listed under the IUCN categories should be prioritized through focused research and use of any possible data available, including strandings and incidental catches (without harming the animal if still alive). This approach offers an expedient strategy in addressing the gap between current and desired knowledge with respect to biological characteristics through focused field studies. Despite the number of scientific publications that investigate the welfare of charismatic rather than non-charismatic species (Hosey et al., 2020) the gaps of biological knowledge in charismatic species are still wide and should be addressed.
LA, PS-Y, KK-R, EC, and RR extracted and analyzed the data and drafted the manuscript with contributions from all other authors. All authors contributed to the article and approved the submitted version.
The present work was supported by the H2020 funded project “EcoScope” (Contract No. 101000302).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1198137/full#supplementary-material
Akoglu E., Salihoglu B., Libralato S., Oguz T., Solidoro C. (2014). An indicator-based evaluation of Black Sea food web dynamics during 1960–2000. J. Mar. Syst. 134, 113–125. doi: 10.1016/j.jmarsys.2014.02.010
Baran E. (2002). “The importance of non-commercial fish,” in Fisheries and Aquaculture: Towards Sustainable Aquatic Living Resources Management. UNESCO Encyclopedia of Life Support Systems. Ed. Safran P. (Oxford: EOLSS Publishers), 1±7.
Barausse A., Duci A., Mazzoldi C., Artioli Y., Palmeri L. (2009). Trophic network model of the Northern Adriatic Sea: Analysis of an exploited and eutrophic ecosystem. Estuarine Coast. Shelf Sci. 83, 577–590. doi: 10.1016/j.ecss.2009.05.003
Bauer B., Horbowy J., Rahikainen M., Kulatska N., Müller-Karulis B., Tomczak M. T., et al. (2019). Model uncertainty and simulated multispecies fisheries management advice in the Baltic Sea. PloS One 14, e0211320. doi: 10.1371/journal.pone.0211320
Beattie A., Sumaila U. R., Christensen V., Pauly D. (2002). A model for the bioeconomic evaluation of marine protected area size and placement in the North Sea. Nat. Resour. Model 15, 413–437. doi: 10.1111/j.1939-7445.2002.tb00096.x
Beaugrand G. (2004). The North Sea regime shift: Evidence, causes, mechanisms and consequences. Prog. Oceanogr. 60, 245–262. doi: 10.1016/j.pocean.2004.02.018
Bell J. J., McGrath E., Biggerstaff A., Bates T., Cárdenas C. A., Bennett H. (2015). Global conservation status of sponges. Conserv. Biol. 29, 42–53. doi: 10.1111/cobi.12447
Borja Á, Galparsoro I., Irigoien X., Iriondo A., Menchaca I., Muxika I., et al. (2011). Implementation of the European Marine Strategy Framework Directive: a methodological approach for the assessment of environmental status, from the Basque Country (Bay of Biscay). Mar. Pollut. Bull. 62, 889–904. doi: 10.1016/j.marpolbul.2011.03.031
Burkhard B., Opitz S., Lenhart H., Ahrendt K., Garthe S., Mendel B., et al. (2011). Ecosystem based modeling and indication of ecological integrity in the German North Sea—Case study offshore wind parks. Ecol. Indic. 11, 168–174. doi: 10.1016/j.ecolind.2009.07.004
Cartes J. E., Abelló P., Lloris D., Carbonell A., Torres P., Maynou F., et al. (2002). Feeding guilds of western Mediterranean demersal fish and crustaceans: an analysis based in a spring survey. Scientia Marina 66 (S2), 209–220. doi: 10.3989/scimar.2002.66s2209
Chen S., Watanabe S. (1989). Age dependence of natural mortality coefficient in fish population dynamics. Nippon Suisan Gakkaishi 55, 205–208. doi: 10.2331/suisan.55.205
Cherif S., Doblas-Miranda E., Lionello P., Borrego C., Giorgi F., Iglesias A., et al. (2020). “Drivers of change,” in Climate and Environmental Change in the Mediterranean Basin – Current Situation and Risks for the Future. First Mediterranean Assessment Report. Eds. Cramer W., Guiot J., Marini K. (Marseille, France: Union for the Mediterranean, Plan Bleu, UNEP/MAP), 59–180.
Christensen V., Walters C. (2004). Ecopath with Ecosim: methods, capabilities and limitations. Ecol. Model. 172, 109–139. doi: 10.1016/j.ecolmodel.2003.09.003
Claudet J., Bopp L., Cheung W. W. L., Devillers R., Escobar-Briones E., Haugan P., et al. (2019). A roadmap for using the UN decade of ocean science for sustainable development in support of science, policy, and action. One Earth 2, 34–42. doi: 10.1016/j.oneear.2019.10.012
Coll M., Grazia Pennino M., Steenbeek J., Sole J., Bellido J. M. (2019). Predicting marine species distributions: Complementarity of food-web and Bayesian hierarchical modelling approaches. Ecol. Model. 405, 86–101. doi: 10.1016/j.ecolmodel.2019.05.005
Coll M., Palomera I., Tudela S. (2009). Decadal changes in a NW Mediterranean Sea food web in relation to fishing exploitation. Ecol. Model. 220, 2088–2102. doi: 10.1016/j.ecolmodel.2009.04.049
Coll M., Palomera I., Tudela S., Dowd M. (2008). Food-web dynamics in the South Catalan Sea ecosystem (NW Mediterranean) for 1978–2003. Ecol. Model. 217, 95–116. doi: 10.1016/j.ecolmodel.2008.06.013
Coll M., Palomera I., Tudela S., Sardà F. (2006). Trophic flows, ecosystem structure and fishing impacts in the South Catalan Sea, Northwestern Mediterranean. J. Mar. Syst. 59, 63–96. doi: 10.1016/j.jmarsys.2005.09.001
Coll M., Santojanni A., Palomera I., Tudela S., Arneri E. (2007). An ecological model of the Northern and Central Adriatic Sea: Analysis of ecosystem structure and fishing impacts. J. Mar. Syst. 67, 119–154. doi: 10.1016/j.jmarsys.2006.10.002
Coro G., Bove P., Armelloni E. N., Masnadi F., Scanu M., Scarcella G. (2022). Filling Gaps in Trawl Surveys at Sea through Spatiotemporal and Environmental Modelling. Front. Mar. Sci. 9, 919339. doi: 10.3389/fmars.2022.919339
Corrales X., Coll M., Ofir E., Heymans J. J., Steenbeek J., Goren M., et al. (2018). Future scenarios of marine resources and ecosystem conditions in the Eastern Mediterranean under the impacts of fishing, alien species and sea warming. Sci. Rep. 8, 14284. doi: 10.1038/s41598-018-32666-x
Corrales X., Coll M, Ofir E, Piroddi C., Goren M., Edelist D., et al. (2017). Hindcasting the dynamics of an Eastern Mediterranean marine ecosystem under the impacts of multiple stressors. Mar. Ecol. Prog. Ser. 580, 17–36. doi: 10.3354/meps12271
Corrales X., Katsanevakis S., Coll M., Heymans J. J., Piroddi C., Ofir E., et al. (2019). Advances and challenges in modelling the impacts of invasive alien species on aquatic ecosystems. Biol. Invasions 22, 907–934. doi: 10.1007/s10530-019-02160-0
Corrales X., Preciado I., Gascuel D., Lopez de Gamiz-Zearra A., Hernvann P.-Y., Mugerza E., et al. (2022). Structure and functioning of the Bay of Biscay ecosystem: A trophic modelling approach. Estuarine Coast. Shelf Sci. 26, 107658. doi: 10.1016/j.ecss.2021.107658
Costalago D., Bauer B., Tomczak M. T., Lundström K., Winder M. (2019). The necessity of a holistic approach when managing marine mammal–fisheries interactions: Environment and fisheries impact are stronger than seal predation. Ambio 48, 552–564. doi: 10.1007/s13280-018-1131-y
Damalas D., Kalyvioti G., Sabatella E. C., Stergiou K. I. (2018). Open data in the life sciences: the ‘Selfish Scientist Paradox’. Ethics Sci. Environ. Politics 18, 27–36. doi: 10.3354/esep00182
Daskalaki E., Koufalis E., Dimarchopoulou D., Tsikliras A. C. (2022). Scientific progress made towards bridging the knowledge gap in the biology of Mediterranean marine fishes. PloS One 17, e0277383. doi: 10.1371/journal.pone.0277383
Daskalov G. (2002). Overfishing drives a trophic cascade in the Black Sea. MEPS 225, 53–63. doi: 10.3354/meps225053
Daskalov G. M., Boicenco L., Grishin A. N., Lazar L., Mihneva V., Shlyakhov V. A., et al. (2017). Architecture of collapse: regime shift and recovery in an hierarchically structured marine ecosystem. Global Change Biol. 23, 1486–1498. doi: 10.1111/gcb.13508
Daskalov G. M., Demirel N., Ulman A., Georgieva Y., Zengin M. (2020). Stock dynamics and predator–prey effects of Atlantic bonito and bluefish as top predators in the Black Sea. ICES J. Mar. Sci. 77, 2995–3005. doi: 10.1093/icesjms/fsaa182
Deacon M. (1997). Scientists and the Sea 1650–1900: A Study of Marine Science. 2nd ed (England: Routledge).
Demirel N., Zengin M., Ulman A. (2020). First large-scale eastern Mediterranean and Black Sea stock assessment reveals a dramatic decline. Front. Mar. Sci. 7, 103. doi: 10.3389/fmars.2020.00103
Dimarchopoulou D. (2020). Ecosystem approach to fisheries management in the Aegean Sea (School of Biology, Aristotle University of Thessaloniki, Thessaloniki, Greece).
Dimarchopoulou D., Keramidas I., Sylaios G., Tsikliras A. C. (2021). Eco-trophic effects of fishing across the Mediterranean Sea. Water 13, 482. doi: 10.3390/w13040482
Dimarchopoulou D., Stergiou K. I., Tsikliras A. C. (2017). Gap analysis on the biology of Mediterranean marine fishes. PloS One 12, e0175949. doi: 10.1371/journal.pone.0175949
Dimarchopoulou D., Tsagarakis K., Keramidas I, Tsikliras A. C. (2019). Ecosystem models and effort simulations of an untrawled gulf in the central Aegean Sea. Front. Mar. Sci. 6, 648. doi: 10.3389/fmars.2019.00648
Dimarchopoulou D., Tsagarakis K., Sylaios G., Tsikliras A. C. (2022). Ecosystem trophic structure and fishing effort simulations of a major fishing ground in the northeastern Mediterranean Sea (Thermaikos Gulf). Estuarine Coast. Shelf Sci. 264, 107667. doi: 10.1016/j.ecss.2021.107667
Dobrzycka-Krahel A., Medina-Villar S. (2020). Alien species of Mediterranean origin in the Baltic Sea Region: current state and risk assessment. Environ. Rev. 28 (3), 339–356. doi: 10.1139/er-2019-0074
Feistel R., Nausch G., Wasmund N. (2008). State and Evolution of the Baltic Sea. (Wiley Interscience), 1952–2005.
Fortibuoni T., Libralato S., Arneri E., Giovanardi O., Solidoro C., Raicevich S. (2017). Fish and fishery historical data since the 19th century in the Adriatic Sea, Mediterranean. Sci. Data 4, 170104. doi: 10.1038/sdata.2017.104
Fransz H. G., Mommaerts J. P., Radach G. (1991). Ecological modelling of the North sea. Netherlands J. Sea Res. 28, 67–140. doi: 10.1016/0077-7579(91)90005-L
Froese R., Tsikliras A. C., Scarcella G., Gascuel D. (2021). Progress towards ending overfishing in the Northeast Atlantic. Mar. Policy 125, 104282. doi: 10.1016/j.marpol.2020.104282
Froese R., Winker H., Coro G., Demirel N., Tsikliras A. C., Dimarchopoulou D., et al. (2020). Estimating stock status from relative abundance and resilience. ICES J. Mar. Sci. 77, 527–538. doi: 10.1093/icesjms/fsz230
Froese R., Winker H., Coro G., Demirel N., Tsikliras A. C., Dimarchopoulou D., et al. (2018b). A new approach for estimating stock status from length frequency data. ICES J. Mar. Sci. 75, 2004–2015. doi: 10.1093/icesjms/fsy078
Froese R., Winker H., Coro G., Demirel N., Tsikliras A. C., Dimarchopoulou D., et al. (2018a). Status and rebuilding of European fisheries. Mar. Policy 93, 159–170. doi: 10.1016/j.marpol.2018.04.018
Galil B. S., Boero F., Campbell M. L., Carlton J. T., Cook E., Fraschetti S., et al. (2015). ‘Double trouble’: the expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 17, 973–976. doi: 10.1007/s10530-014-0778-y
Gislason H., Daan N., Rice J. C., Pope J. G. (2010). Size, growth, temperature and the natural mortality of marine fish. Fish Fisheries 11, 149–158. doi: 10.1111/j.1467-2979.2009.00350.x
Grossowicz M., Ofir E., Shabtay A., Wood J., Biton E., Belkin N., et al. (2020). Modeling the effects of brine outflow from desalination plants on coastal food-webs of the Levantine basin (eastern Mediterranean Sea). Desalination 496, 114757. doi: 10.1016/j.desal.2020.114757
Grüss A., Palomares M. L. D., Poelen J. H., Barile J. R., Aldemita C. D., Ortiz S. R., et al. (2019). Building bridges between global information systems on marine organisms and ecosystem models. Ecol. Model. 398, 1–19. doi: 10.1016/j.ecolmodel.2019.01.023
Gucu A. C. (2002). Can Overfishing be Responsible for the Successful Establishment of Mnemiopsis leidyi in the Black Sea? Estuarine Coast. Shelf Sci. 54, 439–451. doi: 10.1006/ecss.2000.0657
Guénette S., Gascuel D. (2012). Shifting baselines in European fisheries: The case of the Celtic Sea and Bay of Biscay. Ocean Coast. Manage. 70, 10–21. doi: 10.1016/j.ocecoaman.2012.06.010
Güneroğlu A., Samsun O., Feyzioğlu M., Dihkan M. (2019). “Chapter 21 - The Black Sea—The Past, Present, and Future Status,” in Coasts and Estuaries, vol. 2019 . Eds. Wolanski E., Day J. W., Elliott M., Ramachandran. R., 363–375.
Heath M. R. (2005). Changes in the structure and function of the North Sea fish foodweb 1973–2000, and the impacts of fishing and climate. ICES J. Mar. Sci. 62, 847–868. doi: 10.1016/j.icesjms.2005.01.023
Heymans J. J., Bundy A., Christensen V., Coll M., de Mutsert K., Fulton E. A., et al. (2020). The Ocean Decade: A true ecosystem modelling challenge. Front. Mar. Sci. 7, 554573. doi: 10.3389/fmars.2020.554573
Hosey G., Melfi V., Ward S. J. (2020). “Problematic Animals in the Zoo: The Issue of Charismatic Megafauna,” in Problematic Wildlife II. Eds. Angelici F., Rossi L. (Cham: Springer).
ICES (2022). ICES Stock Information Database (Copenhagen, Denmark). Available at: https://sid.ices.dk (Accessed 17 Nov 2022).
Jâms I. B., Windsor F. M., Poudevigne-Durance T., Ormerod S. J., Durance I. (2020). Estimating the size distribution of plastics ingested by animals. Nat. Commun. 11, 1594. doi: 10.1038/s41467-020-15406-6
Jardim E., Millar C. P., Mosqueira I., Scott F., Osio G. C., Ferretti M., et al. (2015). What if stock assessment is as simple as a linear model? The a4a initiative. ICES J. Mar. Sci. 72, 232–236. doi: 10.1093/icesjms/fsu050
Katsanevakis S., Coll M., Piroddi C., Steenbeek S., Ben Rais Lasram F., Zenetos A., et al. (2014). Invading the Mediterranean Sea: biodiversity patterns shaped by human activities. Front. Mar. Sci. 1, 32. doi: 10.3389/fmars.2014.00032
Keramidas I., Dimarchopoulou D., Ofir E., Tsikliras A. C., Gal G. (2023). Ecotrophic perspective in fisheries management: A review on EwE models in European marine ecosystems. Front. Mar. Sci. 10, 1182921. doi: 10.3389/fmars.2023.1182921
Keramidas I., Dimarchopoulou D., Tsikliras A. C. (2022). Modelling and assessing the ecosystem of the Aegean Sea, a major hub of the eastern Mediterranean at the intersection of Europe and Asia. Regional Stud. Mar. Sci. 56, 102704. doi: 10.1016/j.rsma.2022.102704
Kideys A. E. (2002). Fall and rise of the Black Sea ecosystem. Science 297, 1482–1484. doi: 10.1126/science.1073002
Krause J. R., Hightower J. E., Poland S. J., Buckel J. A. (2020). An integrated tagging and catch-curve model reveals high and seasonally-varying natural mortality for a fish population at low stock biomass. Fisheries Res. 232, 105725. doi: 10.1016/j.fishres.2020.105725
Laran S., Authier M., Blanck A., Dorémus G., Falchetto H., Monestiez P., et al. (2017). Seasonal distribution and abundance of cetaceans within French waters – Part II: the Bay of Biscay and the English Channel. Deep Sea Res. Part II 141, 31–40. doi: 10.1016/j.dsr2.2016.12.012
Lassalle G., Lobry J., Le Lo’h F., Bustamante P., Certain G., Delmas D., et al. (2011). Lower trophic levels and detrital biomass control the Bay of Biscay continental shelf food web: implications for ecosystem management. Prog. Oceanogr. 91, 561–575. doi: 10.1016/j.pocean.2011.09.002
Le Marchand M., Hattab T., Niquil N., Albouy C., Le Loc’h F., Lasram F. B. R. (2020). Climate change in the Bay of Biscay: Changes in spatial biodiversity patterns could be driven by the arrivals of southern species. Mar. Ecol. Prog. Ser. 647, 17–31. doi: 10.3354/meps13401
Libralato S., Caccin A., Pranovi F. (2015). Modeling species invasions using thermal and trophic niche dynamics under climate change. Front. Mar. Sci. 2, 29. doi: 10.3389/fmars.2015.00029
Libralato S., Coll M., Tempesta M., Santojanni A., Spoto M., Palomera I., et al. (2010). Food-web traits of protected and exploited areas of the Adriatic Sea. Biol. Conserv. 143 (9), 2182–2194. doi: 10.1016/j.biocon.2010.06.002
Link J. S. (2010). Ecosystem-Based Fisheries Management: Confronting Tradeoffs (Cambridge, UK: Cambridge University Press).
Link J. S., Ihde T. F., Harvey C. J., Gaichas S. K., Field J. C., Brodziak J. K. T., et al. (2012). Dealing with uncertainty in ecosystem models: The paradox of use for living marine resource management. Prog. Oceanogr. 102, 102–114. doi: 10.1016/j.pocean.2012.03.008
Lorenzen K. (2022). Size- and age-dependent natural mortality in fish populations: Biology, models, implications, and a generalized length-inverse mortality paradigm. Fisheries Res. 255, 106454. doi: 10.1016/j.fishres.2022.106454
Lotze H. K., Coll M., Dunne J. A. (2011). Historical changes in marine resources, food-web structure and ecosystem functioning in the Adriatic sea, mediterranean. Ecosystems 14, 198–222. doi: 10.1007/s10021-010-9404-8
Lotze H. K., Worm B. (2009). Historical baselines for large marine animals. Trends Ecol. Evol. 24, 254–262. doi: 10.1016/j.tree.2008.12.004
Luczak C., Beaugrand G., Lindley J. A., Dewarumez J.-M., Dubois P. J., Kirby R. R. (2012). North Sea ecosystem change from swimming crabs to seagulls. Biol. Lett. 8, 821–824. doi: 10.1098/rsbl.2012.0474
Mackinson S., Deas B., Beveridge D., Casey J. (2009). Mixed-fishery or ecosystem conundrum? Multispecies considerations inform thinking on long-term management of North Sea demersal stocks. Can. J. Fish. Aquat. Sci. 66, 1107–1129. doi: 10.1139/F09-057
Mackinson S., Platts M., Garcia C., Lynam C. (2018). Evaluating the fishery and ecological consequences of the proposed North Sea multi-annual plan. PloS One 13 (1), e0190015. doi: 10.1371/journal.pone.0190015
McManamay R. A., Utz R. M. (2014). Open-Access databases as unprecedented resources and drivers of cultural change in fisheries science. Fisheries 39, 417–425. doi: 10.1080/03632415.2014.946128
Michael-Bitton G., Gal G., Corrales X., Ofir E., Mordechai S., Zemah-Shamir S. (2022). Economic aspects of fish stock accounting as a renewable marine natural capital: The Eastern Mediterranean continental shelf ecosystem as a case study. Ecol. Economics 200, 107539. doi: 10.1016/j.ecolecon.2022.107539
Michailidis N., Corrales X., Karachle P. K., Chartosia N., Katsanevakis S., Sfenthourakis S. (2019). Modelling the role of alien species and fisheries in an Eastern Mediterranean insular shelf ecosystem. Ocean Coast. Manage. 175, 152–171. doi: 10.1016/j.ocecoaman.2019.04.006
Morley T. I., Fayet A. L., Jessop H., Veron P., Veron M., Clark J., et al. (2016). The seabird wreck in the Bay of Biscay and South-Western Approaches in 2014: A review of reported mortality. Seabird 29, 22–38.
Moullec F., Gascuel D., Bentorcha K., Guénette S., Robert M. (2017). Trophic models: What do we learn about Celtic Sea and Bay of Biscay ecosystems? J. Mar. Syst. 172, 104–117. doi: 10.1016/j.jmarsys.2017.03.008
Niiranen S., Yletyinen J., Tomczak M., Blenckner T., Hjerne O., MacKenzie B. R., et al. (2013). Combined effects of global climate change and regional ecosystem drivers on an exploited marine food web. Global Change Biol. 19 (11), 3327–3342. doi: 10.1111/gcb.12309
Ofir E., Ribas X., Coll M., Heymans J. J., Goren M., Steenbeek J. G., et al. (2023). Evaluation of fisheries management policies in the alien species-rich Eastern Mediterranean under climate change. Front. Mar. Sci. 10, 1155480. doi: 10.3389/fmars.2023.1155480
Ojaveer H., Einberg H., Lehtiniemi M., Outinen O., Zaiko A., Jelmert A., et al. (2023). Quantifying impacts of human pressures on ecosystem services: Effects of widespread non-indigenous species in the Baltic Sea. Sci. Total Environ. 858, 159975. doi: 10.1016/j.scitotenv.2022.159975
Ojaveer H., Jaanus A., MacKenzie B. R., Martin G., Olenin S., Radziejewska T., et al. (2010). Status of biodiversity in the Baltic sea. PloS One 5, e12467. doi: 10.1371/journal.pone.0012467
Österblom H., Hansson S., Larsson U., Hjerne O., Wuff F., Elmgren R., et al. (2007). Human-induced trophic cascades and ecological regime shifts in the Baltic sea. Ecosystems 10, 877–889. doi: 10.1007/s10021-007-9069-0
Palomares M. L. D., Pauly D. (2021). SeaLifeBase (World Wide Web electronic publication). Available at: www.sealifebase.org.
Papantoniou G., Giannoulaki M., Stoumboudi M. T., Lefkaditou E., Tsagarakis K. (2021). Food web interactions in a human dominated Mediterranean coastal ecosystem. Mar. Environ. Res. 172, 105507. doi: 10.1016/j.marenvres.2021.105507
Pauly D. (1980). On the interrelationships between natural mortality, growth parameters, and mean environmental temperatures in 175 fish stocks. J. du Conseil Int. pour l’Exploration la Mer 39, 175–192. doi: 10.1093/icesjms/39.2.175
Pauly D., Zeller D. (2016). Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 7, 10244. doi: 10.1038/ncomms10244
Peled Y., Zemah-Shamir S., Israel A., Shechter M., Ofir E., Gal G. (2020). Incorporating insurance value into ecosystem services assessments: mitigation of ecosystem users’ welfare uncertainty through biological control. Ecosyst. Serv. 46, 101192. doi: 10.1016/j.ecoser.2020.101192
Pereira F., Vasconcelos P., Moreno A., Gaspar M. B. (2019). Catches of Sepia officinalis in the small-scale cuttlefish trap fishery off the Algarve coast (southern Portugal). Fisheries Res. 214, 117–125. doi: 10.1016/j.fishres.2019.01.022
Piroddi C., Akoglu E., Andonegi E., Bentley J. W., Celic I., Coll M., et al. (2021). Effects of nutrient management scenarios on marine food webs: a Pan-European Assessment in support of the Marine Strategy Framework Directive. Front. Mar. Sci. 7, 596797. doi: 10.3389/fmars.2021.596797
Piroddi C., Coll M., Macias D., Steenbeek J., Garcia-Gorriz E., Mannini A., et al. (2022). Modelling the Mediterranean Sea ecosystem at high spatial resolution to inform the ecosystem-based management in the region. Sci. Rep. 12, 19680. doi: 10.1038/s41598-022-18017-x
Piroddi C., Colloca F., Tsikliras A. C. (2020). The living marine resources in the Mediterranean Sea Large Marine Ecosystem. Environ. Dev. 36, 100555. doi: 10.1016/j.envdev.2020.100555
Prodanov K., Mikhailov K., Daskalov D., Maxim K., Ozdamar E., Shlyakhov V., et al. (1997). Environmental management of fish resources in the Black Sea and their rational exploitation. Stud. Reviews 68 GFCM 1997, 178.
Püts M., Taylor M., Núñez-Riboni I., Steenbeek J., Stäbler M., Möllmann C., et al. (2020). Insights on integrating habitat preferences in process-oriented ecological models – a case study of the southern North Sea. Ecol. Model. 431, 109189. doi: 10.1016/j.ecolmodel.2020.109189
Quante M., Colijn F., Bakker J. P., Härdtle W., Heinrich H., Lefebvre C., et al. (2016). Introduction to the assessment—Characteristics of the region. (Springer). doi: 10.1007/978-3-319-39745-0_1
Quetglas A., de Mesa A., Ordines F., Grau A. (2010). Life history of the deep-sea cephalopod family Histioteuthidae in the western Mediterranean. Deep Sea Res. Part I: Oceanographic Res. Papers 57, 999–1008. doi: 10.1016/j.dsr.2010.04.008
Ravier C., Fromentin J.-M. (2004). Are the long-term fluctuations in Atlantic bluefin tuna (Thunnus thynnus) population related to environmental changes? Fish. Oceanogr. 13, 145–160. doi: 10.1111/j.1365-2419.2004.00284.x
Reusch T. B. H., Dierking J., Andersson H. C., Bonsdorff E., Carstensen J., Casini M., et al. (2018). The baltic sea as a time machine for the future coastal ocean. Sci. Adv. 4, eaar8195. doi: 10.1126/sciadv.aar8195
Rodriguez-Perez A., Tsikliras A. C., Gal G., Steenbeek J., Falk-Andersson J., Heymans S. J. J. (2023). Using ecosystem models to inform ecosystem-based fisheries management in Europe: a review of the policy landscape and related stakeholder needs. Front. Mar. Sci. 10, 1196329. doi: 10.3389/fmars.2023.1196329
Rosa S., Pansera M., Granata A., Guglielmo L. (2013). Interannual variability, growth, reproduction and feeding of Pelagia noctiluca (Cnidaria: Scyphozoa) in the Straits of Messina (Central Mediterranean Sea): Linkages with temperature and diet. J. Mar. Syst. 111–112, 97–107. doi: 10.1016/j.jmarsys.2012.10.001
Saad A., Mahfoud I. (2022). “Marine mammals in Syria,” in Marine mammals. Ed. Kaoud H. A. E. (London: IntechOpen).
Sardà F., Calafat A., Flexas M. M., Tselepides A., Canals M., Espino M., et al. (2004). An introduction to Mediterranean deep-sea biology. Scientia Marina 68 (S3), 7–38. doi: 10.3989/scimar.2004.68s37
Savva I., Chartosia N., Antoniou C., Kleitou P., Georgiou A., Stern N., et al. (2020). They are here to stay: the biology and ecology of lionfish (Pterois miles) in the Mediterranean Sea. J. Fish Biol. 97, 148–162. doi: 10.1111/jfb.14340
Scarcella G., Angelini S., Armelloni E. N., Costantini I., De Felice A., Guicciardi S., et al. (2022). The potential effects of COVID-19 lockdown and the following restrictions on the status of eight target stocks in the Adriatic Sea. Front. Mar. Sci. 9, 920974. doi: 10.3389/fmars.2022.920974
Scotti M., Opitz S., MacNeil L., Kreutle A., Pusch C., Froese R. (2022). Ecosystem-based fisheries management increases catch and carbon sequestration through recovery of exploited stocks: The western Baltic Sea case study. Front. Mar. Sci. 9, 879998. doi: 10.3389/fmars.2022.879998
Shabtay A., Portman M., Ofir E., Carmel Y., Gal G. (2018). Using ecological modelling in marine spatial planning to enhance ecosystem-based management. Mar. Policy 95, 14–23. doi: 10.1016/j.marpol.2018.06.018
Shiganova T. A. (1998). Invasion of the Black Sea by the ctenophore Mnemiopsis leidyi and recent changes in pelagic community structure. Fish. Oceanogr. 7, 305–310. doi: 10.1046/j.1365-2419.1998.00080.x
Spitz J., Ridoux V., Trites A. W., Laran S., Authier M. (2018). Prey consumption by cetaceans reveals the importance of energy-rich food webs in the Bay of Biscay. Prog. Oceanogr. 166, 148–158. doi: 10.1016/j.pocean.2017.09.013
Stäbler M., Kempf A., Temming A. (2018). Assessing the structure and functioning of the southern North Sea ecosystem with a food-web model. Ocean Coast. Manage. 165, 280–297. doi: 10.1016/j.ocecoaman.2018.08.017
Stasolla G., Tricarico E., Vilizzi L. (2021). Risk screening of the potential invasiveness of non-native marine crustacean decapods and barnacles in the Mediterranean Sea. Hydrobiologia 848, 1997–2009. doi: 10.1007/s10750-020-04432-6
Stergiou K. I., Somarakis S., Triantafyllou G., Tsiaras K. P., Giannoulaki M., Petihakis G., et al. (2016). Trends in productivity and biomass yields in the Mediterranean Sea Large Marine Ecosystem during climate change. Environ. Dev. 17, 57–74. doi: 10.1016/j.envdev.2015.09.001
Stergiou K. I., Tsikliras A. C. (2006). Underrepresentation of regional ecological research output by bibliometric indices. Ethics Sci. Environ. Politics 2006, 15–17. doi: 10.3354/esep006015
Tecchio S., Coll M., Christensen V., Company J. B., Ramírez-Llodra E., Sardà F. (2013). Food web structure and vulnerability of a deep-sea ecosystem in the NW Mediterranean Sea. Deep Sea Res. Part I: Oceanographic Res. Papers 75, 1–15. doi: 10.1016/j.dsr.2013.01.003
Then A. Y., Hoenig J. M., Hall N. G., Hewitt D. A. (2015). Evaluating the predictive performance of empirical estimators of natural mortality rate using information on over 200 fish species. ICES J. Mar. Sci. 72, 82–92. doi: 10.1093/icesjms/fsu136
Topaloglu B., Alper E. (2014). Updated checklist of sponges (Porifera) along the coasts of Turkey. Turkish J. Zool. 38, 665–676. doi: 10.3906/zoo-1405-79
Tsagarakis K., Coll M., Giannoulaki M., Somarakis S., Papaconstantinou C., Machias A. (2010). Food web traits of the North Aegean Sea ecosystem (Eastern Mediterranean) and comparison with other Mediterranean ecosystems. Estuarine Coast. Shelf Sci. 88 (2), 233–248. doi: 10.1016/j.ecss.2010.04.007
Tsikliras A. C., Antonopoulou E., Stergiou K. I. (2010). Spawning period of Mediterranean marine fishes. Rev. Fish Biol. Fisheries 20, 499–538. doi: 10.1007/s11160-010-9158-6
Tsikliras A. C., Coro G., Daskalov G., Gremillet D., Scotti M., Sylaios G. (2023). A need for a paradigm shift from anthropocentric to ecocentric fisheries management. Front. Mar. Sci. 10
Tsikliras A. C., Dimarchopoulou D. (2021). Filling in knowledge gaps: length-weight relations of 46 uncommon sharks and rays in the Mediterranean Sea. Acta Ichthyologica Piscatoria 51, 249–255. doi: 10.3897/aiep.51.e65858
Tsikliras A. C., Dinouli A., Tsiros V.-Z., Tsalkou E. (2015). The Mediterranean and Black Sea fisheries at risk from overexploitation. PloS One 10, e0121188. doi: 10.1371/journal.pone.0121188
Tsikliras A. C., Froese R. (2019). “Maximum sustainable yield,” in Encyclopedia of ecology, 2nd ed., vol. 1 . Ed. Fath B. (Oxford: Elsevier), 108–115.
Tsikliras A. C., Stergiou K. I. (2014). Size at maturity of Mediterranean marine fishes. Rev. Fish Biol. Fisheries 24, 219–268. doi: 10.1007/s11160-013-9330-x
Tsikliras A. C., Touloumis K., Pardalou A., Adamidou A., Keramidas I., Orfanidis G., et al. (2021). Status of 74 non-commercial fish and invertebrate stocks in the Aegean Sea using abundance and resilience. Front. Mar. Sci. 7, 578601. doi: 10.3389/fmars.2020.578601
Ugland K. (1976). Population studies on eastern North Atlantic minke whales, Balaenoptera acutorostrata Lacepede. Rep. Int. Whaling Commission (Sci. Rep.) 26, 366–381.
Ulman A., Yildiz T., Demirel N., Canak O., Yemişken E., Pauly D. (2021). The biology and ecology of the invasive silver-cheeked toadfish (Lagocephalus sceleratus), with emphasis on the Eastern Mediterranean. NeoBiota 68, 145–175. doi: 10.3897/neobiota.68.71767
Vilas D., Coll M., Corrales X., Steenbeek J., Piroddi C., Macias D., et al. (2021). Current and potential contributions of the Gulf of Lion Fisheries Restricted Area to fisheries sustainability in the NW Mediterranean Sea. Mar. Policy 123, 104296. doi: 10.1016/j.marpol.2020.104296
Keywords: data gaps, marine biodiversity, marine mammals, seabirds, marine reptiles, marine invertebrates, SeaLifeBase, European waters
Citation: Abucay LR, Sorongon-Yap P, Kesner-Reyes K, Capuli EC, Reyes RB Jr, Daskalaki E, Ferrà C, Scarcella G, Coro G, Ordines F, Sánchez-Zulueta P, Dakalov G, Klayn S, Celie L, Scotti M, Grémillet D, Lambert C, Gal G, Palomares MLD, Dimarchopoulou D and Tsikliras AC (2023) Scientific knowledge gaps on the biology of non-fish marine species across European Seas. Front. Mar. Sci. 10:1198137. doi: 10.3389/fmars.2023.1198137
Received: 31 March 2023; Accepted: 15 September 2023;
Published: 03 October 2023.
Edited by:
Periklis Kleitou, Marine and Environmental Research Lab (MER), CyprusReviewed by:
Othman Jarboui, Institut National des Sciences et Technologies de la Mer, TunisiaCopyright © 2023 Abucay, Sorongon-Yap, Kesner-Reyes, Capuli, Reyes, Daskalaki, Ferrà, Scarcella, Coro, Ordines, Sánchez-Zulueta, Dakalov, Klayn, Celie, Scotti, Grémillet, Lambert, Gal, Palomares, Dimarchopoulou and Tsikliras. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Athanassios C. Tsikliras, YXRzaWtAYmlvLmF1dGguZ3I=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.