 Daphne Wrobel Goldberg1*
Daphne Wrobel Goldberg1* Marina Molinas Alcala2
Marina Molinas Alcala2 Daniela Farias da Nóbrega3
Daniela Farias da Nóbrega3 Fábio Parra Sellera4,5Solange Fonseca6
Fábio Parra Sellera4,5Solange Fonseca6 José Soares Ferreira Neto7Denise Batista Nogueira7
José Soares Ferreira Neto7Denise Batista Nogueira7 Marcos Bryan Heinemann7
Marcos Bryan Heinemann7- 1Instituto Albatroz, Cabo Frio, RJ, Brazil
- 2Econservation Estudos e Projetos Ambientais Ltda, Rio de Janeiro, RJ, Brazil
- 3Pat Animal Laboratory, São José do Rio Preto, São Paulo, Brazil
- 4School of Veterinary Medicine, Metropolitan University of Santos, Santos, SP, Brazil
- 5Department of Internal Medicine, School of Veterinary Medicine and Animal Science, University of São Paulo, São Paulo, SP, Brazil
- 6SM Laboratory & Clinical Microbiology, Rio de Janeiro, RJ, Brazil
- 7Department of Preventive Veterinary Medicine and Animal Health, School of Veterinary Medicine and Animal Sciences, University of São Paulo, São Paulo, Brazil
Mycobacteriosis is a well-known disease that has been reported in a variety of reptiles species. However, the occurrence of mycobacteriosis in sea turtles has been rarely documented, and most reports are from captive-reared animals. A juvenile green turtle (Chelonia mydas) was found stranded in March 2020, in Guanabara Bay, one of the most urbanized and polluted coastal areas in Rio de Janeiro, Southeastern Brazil. The turtle was in fair body condition and exhibited multiple cutaneous fibropapillomas, covering almost 60% of its body. Despite continued medical therapy, the animal died 60 days after initial supportive care. Necropsy revealed granulomatous pneumonia, hydropericardium, small granulomas in the myocardium, liver, spleen, thyroid and brain and caseous exudate in the meninges of the brain. The histopathological analysis showed disseminated granulomatous inflammation in various organs examined during necropsy. Intrahistiocytic acid-fast bacteria were observed in the liver, heart, lungs, kidneys, spleen, brain and adrenals. The blood culture collected shortly after the animal was admitted tested positive for Mycobacterium gordonae, after an 8-week incubation period; however, the animal had already died by the time the results were obtained. Postmortem examination confirmed systemic mycobacteriosis, identified as M. gordonae in blood, liver and lung cultures and by PCR amplification of DNA extracts from frozen tissue samples. This is the first report of a disseminated infection caused by M. gordonae in a free-ranging sea turtle. Although the source of this Mycobacterium remains unknown, it could potentially be related to anthropogenic activities (e.g., waste dumping or leakage from landfill sites). Increasing surveillance of Mycobacterium in wild sea turtles is of paramount importance to better understand the role of mycobacterial species as causative agents of infections in marine species.
Introduction
Even though Mycobacterium species in the Mycobacterium tuberculosis complex are responsible for most mycobacterial infections worldwide, opportunistic infections due to nontuberculous mycobacteria (NTM) are rapidly increasing, becoming an additional public health hazard (Zulu et al., 2021). Nontuberculous mycobacteria are found in the environment, primarily in water and soil, and can be transmitted through direct contact with contaminated materials or through inhalation (Kazda et al., 2009). Clinical presentations are diverse, ranging from asymptomatic to severe life-threatening disease.
Nontuberculous mycobacteria are endemic worldwide and have been associated with infections in a large number of organisms, including mammals, birds, reptiles, amphibians, and fishes (Lacasse et al., 2007; Suykerbuyk et al., 2007; Slany et al., 2010; Palmieri et al., 2012; Willson et al., 2013; Woodhouse et al., 2014; Donnelly et al., 2016; Ullmann et al., 2016; Barrows et al., 2017; Gcebe and Hlokwe, 2017; Gcebe et al., 2018; Gcebe et al., 2018; Schmidt et al., 2022). Spontaneous mycobacteriosis in reptiles was first reported in a snake in 1889 (Sibley, 1889), but has since been documented in a wide variety of reptiles, including snakes (Ullman et al., 2016), lizards (Friend and Russell, 1979), crocodiles (Slany et al., 2010) and sea turtles (Brock et al., 1976; Glazebrook and Campbell, 1990; Nardini et al., 2014; Donnelly et al., 2016; Gcebe et al, 2018). In reptiles, gross lesions may include grayish-white nodules in different organs and in the subcutis. Histopathologic examination of tissue samples from affected animals often shows typical granulomatous inflammation with multinucleated giant cells. Unlike mammalian granulomas, calcification has not been observed (Soldati et al., 2004).
Specifically in chelonians, studies have identified plastral ulceration and pulmonary miliary nodules as common findings (Brock et al., 1976; Glazebrook and Campbell, 1990). In contrast, in immunocompromised turtles, this disease could rapidly progress to systemic infections, leading to severe damages to other organs, including but not limited to liver, spleen, kidneys, heart and brain (Nardini et al., 2014; Donnelly et al., 2016). To date, there is scant information regarding the prevalence, histopathological and microbiological findings, and clinical outcomes of NTM-related infections in sea turtles. In this study, we report a disseminated and fatal infection caused by Mycobacterium gordonae in a stranded green turtle (Chelonia mydas) in Rio de Janeiro, Brazil.
Material and methods
An 11 kg juvenile green turtle (C. mydas) (curved carapace length of 44.0 cm, and curved carapace width of 40.2 cm) was found stranded on March 16, 2020, in the city of Rio de Janeiro (lat: -22.91972; long: -43.16995), Southeastern Brazil. The green turtle was rescued in Guanabara Bay, by the Santos Basin Beach Monitoring Project (Projeto de Monitoramento de Praias - Bacia de Santos - PMP-BS). PMP-BS is a monitoring program required by Brazil’s Federal Environmental Agency, IBAMA, for the environmental licensing process for the oil production and transport by Petrobras at the Santos Basin area (http://www.comunicabaciadesantos.com.br). The turtle was immediately taken to the PMP-BS stabilization unit at Universidade Santa Ursula, Rio de Janeiro, where it was kept throughout the treatment. Physical examination revealed signs of weakness, anemia (PCV 17%) and hypoproteinemia (total protein 0.60 g/dL). The turtle was in fair body condition (i.e., 2/5) and had multiple cutaneous fibropapillomas covering almost 60% of its body. Respiratory rate was 3-5 breaths per minute and no abnormal breath sounds were heard. Heart rate was 22 beats/min, and it was obtained with the support of a veterinary Doppler. Blood samples were collected from the dorsal cervical sinus for haematology, biochemistry (i.e., glucose, total protein, albumin, globulin, AST, CK, uric acid, calcium, phosphorus, sodium, potassium), and microbiological analysis (i.e., blood culture).
The animal was initially dry-docked on a padded surface to prevent drowning and protect the turtle from damage. Colloid IV fluids (20 mL/kg) were administered to maintain a high osmotic pressure in the blood vessels. Then, an intravenous amino acid solution (Aminoven®) (13 mL/kg) was administered to correct hypoproteinemia. Two days after arrival, the turtle was placed in a 3000 L tank one quarter-filled with fresh water at 27°C, where it remained for 24 hours. After that, the animal was moved to an artificial saltwater tank, where the water level was gradually increased; however, the turtle remained lethargic. A variety of food items (e.g., fish, shrimp, and squid) were offered, however the turtle did not eat on its own and, despite that, a slight clinical improvement was observed. Despite continued medical therapy, the animal died 60 days after initial supportive care.
Postmortem examination was performed using standard techniques immediately after death. Swabs were taken from the liver and the lungs after disinfection with chlorhexidine (Asmar & Drancourt, 2015), and then placed in a sterile container for bacterial culture. Additionally, blood samples were collected from the heart and stored in blood culture bottles (Hemoprov - NewProv®). All samples were sent to SM Laboratory & Clinical Microbiology, in the city of Niterói, Rio de Janeiro. The material was inoculated aseptically onto general and selective (i.e., Lowenstein-Jensen) isolation media for Mycobacterium spp. The isolated bacteria was identified by phenotypic tests and by standard biochemical methods.
Duplicate blood and frozen tissue samples were sent to the Department of Preventive Veterinary Medicine and Animal Health, in the University of São Paulo, for biomolecular assays. For mycobacteria isolation the material was decontaminated by the HPC (0.75% hexadecylpyridinium chloride) method (Ikuta et al., 2016), and the supernatant plated on Stonebrink and Lowenstein-Jensen media and incubated at 37°C for up to 90 days. DNA was extracted from the isolated bacteria, using the DNeasy Blood & Tissue kit® (Qiagen©) following the manufacturer’s recommendations. The genus Mycobacterium was confirmed by TB Multiplex PCR (Wilton & Cousins, 1992) and species classification was based on PCR of an hsp65 gene fragment, followed by restriction analysis with BstE II and Hae III enzymes (Telenti et al., 1993, http://app.chuv.ch/prasite/index.html).
Tissue samples were collected and fixed in 10% neutral buffered formalin solution for histopathology and replicate samples were flash frozen in liquid nitrogen and stored at -80°C for PCR. The formalin-fixed samples were routinely processed, paraffin wax-embedded, sliced into 4-μm-thick sections and stained with haematoxylin and eosin (HE), for subsequent histopathologic examination by light microscopy. Additionally, for histochemical analysis, periodic acid–Schiff (PAS) for identification of fungi and Ziehl-Neelsen for identification of acid-fast bacteria stainings were used.
In order to assess the age of the turtle, both humeri were collected for skeletochronology. 3–5 mm transverse sections were taken from each humerus, fixed, decalcified in a commercial decalcifying solution and cut in 25 μm thick sections. The sections were then stained with haematoxylin and mounted on slides in 100% glycerin. Identification, counting and measurements of lines of arrested growth (LAGs) were performed utilizing digital photographs, according to the method proposed by Goshe et al. (2010). Sequential images were taken of each section under 40x magnification with light microscopy (Leica DM750).
Results
The blood culture collected shortly after the animal was admitted tested positive for Mycobacterium gordonae, after an 8-week incubation period; however, the animal had already died by the time the results were obtained. Postmortem examination confirmed systemic mycobacteriosis, identified as M. gordonae in blood, liver and lung cultures and by PCR amplification of DNA extracts from frozen tissue samples including liver, heart, lungs, kidneys, spleen, pancreas, brain, adrenals, thyroid, thymus, and salt glands.
The primary gross findings were multiple tumors arising from the turtle´s skin, plastron and carapace (Figure 1A); marked cachexia, with pronounced concavity in plastron and evident bone demineralization in the cranial carapace, over the scapular region, bilaterally; moderate amount of serosanguineous fluid accumulation in the coelomic cavity; marked muscle wasting with diffuse discoloration and gelatinous texture; pulmonary edema (Figure 1B), characterized by foamy fluid within the lungs; numerous yellowish nodules of 0.5-1 mm diameter in lungs (Figure 1B), heart (Figure 1C), liver (Figures 1D–E), spleen (Figure 1F), thyroid and brain; and a caseous layer of approximately 4 mm thickness on the ventral surface of the brain.
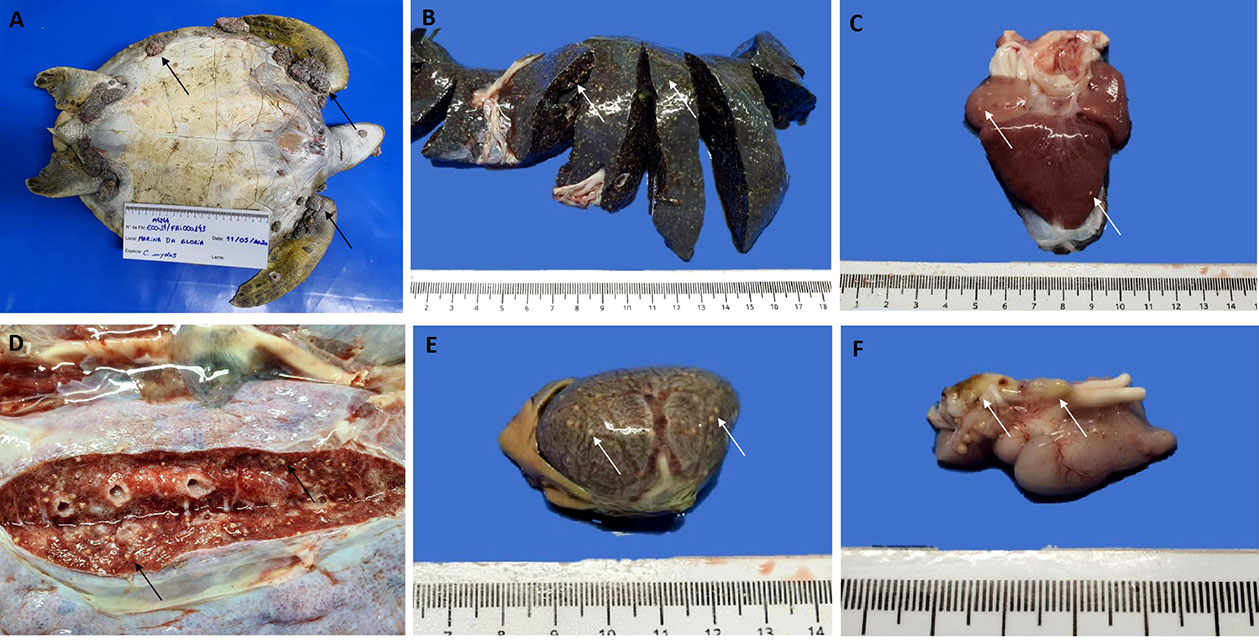
Figure 1 Macroscopic findings in (C) mydas - (A) Multiple fibropapillomas arising from the turtle´s skin and plastron (arrows). Yellowish, multifocal nodules (arrows), randomly distributed, indicating a granulomatous inflammatory process in liver surface and parenchyma (B), heart (C), lung (D), and spleen (E, F) Caseous layer of approximately 4 mm thickness on the ventral surface of the brain (arrows).
Histopathological analysis revealed multiple cutaneous fibropapillomas, histiocytic granulomatous pneumonia, hepatitis, splenitis (Figure 2A), myocarditis, nephritis, meningitis and adrenalitis with intrahistiocytic acid-fast bacteria (Figure 2B), and kidney necrosis with tubular degeneration. The mycobacterial granulomas were characterized by aggregates of epithelioid macrophages and multinucleated giant cells. Additionally, small amounts of trematode eggs surrounded by few macrophages were observed in lung, heart, spleen and stomach and heterophilic granulomas with abundant septate hyphae (stained by PAS) were observed in lung, liver (Figure 2C) and meninges.
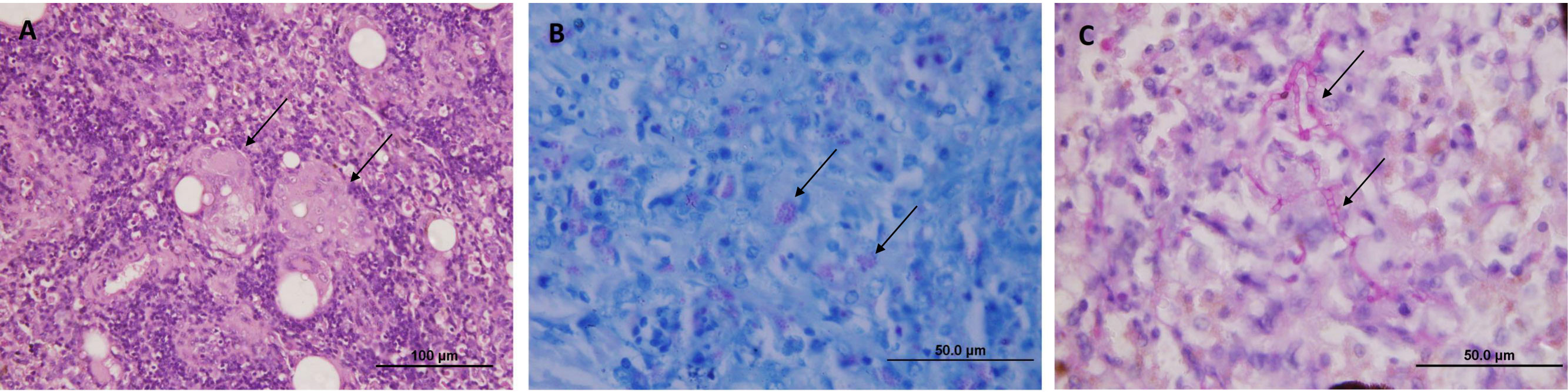
Figure 2 Microscopic findings in (C) mydas - (A) Spleen, aggregates of epithelioid macrophages and multinucleated giant cells (arrows) (HE). (B) Spleen, multiple small acid-fast bacteria in the cytoplasm of macrophages (arrows) (Ziehl-Neelsen stain), (C) Liver, parenchyma with septate hyphae (arrows) surrounded by inflammatory cells (PAS stain).
The inflammatory process in various organs associated with mycobacterial infection, presumably resulted in septic shock. The presence of trematode eggs in multiple organs, compatible with spirorchidiasis as well as the fungal infection, may have been contributory to mortality of the turtle.
Skeletochronology analysis estimated the turtle was 11 years of age.
Discussion
Disseminated mycobacteriosis was confirmed through gross, microscopic, microbiological and molecular analysis. Histiocytic granulomas, considered the predominant pathological finding in mycobacterial infections, as well as co-infections involving fungi, bacteria and/or parasites were observed in different reports of mycobacteriosis in sea turtles (Mitchell et al., 2012; Nardini et al., 2014; Donnelly et al., 2016).
In most cases of mycobacteriosis, sea turtles were presented for chronic disease, and granulomatous lesions were found postmortem (Brock et al., 1976; Greer et al., 2003). Mitchell, 2012 suggested that, due to the hematogenous spread of the disease, the bacteria could potentially reach diverse tissues in sea turtles. However, it is generally thought that the Mycobacterium sp. gain systemic access through cutaneous routes and this is especially important in animals undergoing rehabilitation because they tend to be considered immunocompromised and more susceptible to opportunistic infections.
Greer et al. (2003) isolated a non-tuberculosis Mycobacterium chelonae from a wild Kemp’s ridley (Lepidochelys Kempii) with osteoarthritis, in captivity. Mycobacterium chelonae was also reported in a stranded loggerhead turtle (Caretta caretta) with disseminated mycobacteriosis (Nardini et al., 2014). More recently, Donnelly et al. (2016) identified a nontuberculous Mycobacterium haemophilum in a juvenile leatherback turtle (Dermochelys coriacea) found stranded on the Atlantic coast of Florida. In southern Brazil, acid-fast bacteria were detected in the large intestine of a green turtle (Domiciano et al., 2022), however, the species was not identified. To the best of the authors’ knowledge, this is the first report of M. gordonae in sea turtles, which highlights the novelty of this investigation.
NTM are environmental opportunistic pathogens among humans and animals that have been associated with increased morbidity and mortality rates, especially in immunocompromised individuals (Henkle and Winthrop, 2015). Interestingly, M. gordonae is one of the least virulent NTM for humans, being considered pathogenic only to immunocompromised patients (Bennet et al., 2019). However, cases of severe mycobacteriosis in immunocompetent individuals are not uncommon. Whether the increase in cases is real or whether more cases are being recognized remains unclear (Piersimoni & Scarparo, 2009). In this case, sea turtle fibropapillomatosis, fungal infection and spirorchidiasis may have compromised the animal immune system, increasing susceptibility to opportunistic infections, and could have been a contributing factor to mycobacteriosis. Furthermore, clinical signs such as cachexia, hypoproteinemia, and muscle atrophy reinforce the existence of a chronic debilitating condition.
Despite a considerable increase in knowledge about NTM infections, they still represent a diagnostic and therapeutic challenge, as they can be difficult to diagnose because early signs and symptoms are non-specific and are often very similar to those of other diseases. In addition, NTM are often resistant to routinely used antibiotics or can lead to long-term antibiotic use (i.e., several months or even years). Recurrence of NTM infections after antibiotic treatment can also occur, requiring further rounds of antibiotics administration (Piersimoni & Scarparo, 2009).
In this study, M. gordonae was isolated from a free-living green turtle rescued in Guanabara Bay (GB), which is the second largest bay on the coast of Brazil. It is possible that the infection could have been acquired in GB, being related to the poor water quality of the area. Guanabara Bay has suffered extensive ecological degradation associated but not limited to poor urban planning, over-development, deforestation, unsustainable farming practices, and untreated domestic and industrial effluents. The situation is further aggravated by the lack of sewerage infrastructure in the urban areas surrounding GB, which contributes to bacterial contamination, eutrophication, and litter (Fries et al., 2019). These activities have led to a decrease in water quality, an increase in sedimentation, and an imbalance of the bay’s delicate ecosystem (Fistarol et al., 2015).
Water pollution has been linked to an increase of certain pathogenic bacteria (Fistarol et al., 2015). In GB, particularly, many genera that include potentially pathogenic bacteria were detected, e.g., Vibrio, Klebsiella, Pseudomonas, Clostridium, and Bacillus (Vieira et al., 2008; Coutinho et al., 2014). Many studies have demonstrated that high levels of pollutants in water sources can favor the increase in the number and diversity of pathogenic bacteria, which can further jeopardize the health of humans and free-living organisms (Smith and Schindler, 2009).
Coastal zones are highly used by neritic juvenile green turtles, like the one from this study, and are considered an important developmental area for the species. However, these regions have also been disproportionately affected by human activities. Recently, Williams et al. (2022) estimated that almost half of the coastal areas around the world have been heavily affected by human actions and have been showing signs of ecological degradation. Similar to other coastal areas, GB has been exposed to cumulative impacts and environmental changes, mainly caused by anthropogenic activities. Degradation of GB can be perceived simply by looking at its shores and the color of the water. As a result, not only sea turtles, but all living organisms in GB may be exposed to life-threatening pathogens directly related to pollution.
In summary, we present a case report of M. gordonae infection in a free-ranging sea turtle from an anthropogenically-impacted coastal area of Brazil. Our findings are remarkably important because they shed light on the appearance of M. gordonae in marine animals, which can result in infections and subsequent death. Additionally, considering that wild animals are an important source of emerging infectious diseases, improving surveillance of Mycobacterium in marine animals is of paramount importance for better understanding the role of mycobacterial species as causative agents of infections in marine species. Finally, considering mycobacterial infections can spread between humans and animals, these diseases should be monitored using a One Health approach.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The authors confirm that the ethical policies of the journal, as noted on the journal’s author guidelines page, have been adhered to. No ethical approval was required for this specific study. The licences and research permit for monitoring programme and the biological sampling were issued by the Brazilian government (IBAMA-ABIO 624/2015); all animal handling procedures and protocols followed the required ethics and welfare practices.
Author contributions
The authors confirm contribution to the paper as follows: study conception and design: DG, MA, DFN, FS, MH. Data collection: DG, MA. Analysis and interpretation of results: SF, JN, DBN, MH. Draft manuscript preparation: DG, MA, DFN, FS, MH. All authors contributed to the article and approved the submitted version.
Acknowledgments
We gratefully acknowledge IBAMA and Petrobras for the permission to use these data. The Santos Basin Beach Monitoring Project (Projeto de Monitoramento de Praias da Bacia de Santos—PMP‐BS), conducted from Laguna/SC to Saquarema/RJ, is a requirement set by the Brazilian Institute of the Environment (IBAMA) for the environmental licensing of the oil and natural gas production and transport by Petrobras.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Asmar S., Drancourt M. (2015). Chlorhexidine decontamination of sputum for culturing. Mycobacterium tuberculosis. BMC Microbiol. 15, 155. doi: 10.1186/s12866-015-0479-4
Barrows M., Koeppel K., Michel A., Mitchell E. (2017). Mycobacterial arthritis and synovitis in painted reed frogs (Hyperolius marmoratus). J. Comp. Pathol. 156:2-3, 275–280. doi: 10.1016/j.jcpa.2017.01.005
Bennet J. E., Dolin R., Blaser M. J. (2019). Principles and practice of infectious diseases. 9th ed (Amsterdam, Netherlands: Elsevier).
Brock J. A., Nakamura R. M., Miyahara A. Y., Chang E. M. L. (1976). Tuberculosis in pacific green sea turtles. Chelonia mydas Trans. Am. Fish. Soc 105, 4, 564–4, 566. doi: 10.1577/1548-8659(1976)105<564:TIPGST>2.0.CO;2
Coutinho F. H., Silveira C. B., Pinto L. H., Salloto G. R., Cardoso A. M., Martins O. B., et al. (2014). Antibiotic resistance is widespread in urban aquatic environments of Rio de Janeiro, Brazil. Microb. Ecol. 68 (3), 441–452. doi: 10.1007/s00248-014-0422-5
Domiciano I. G., da Silva Gagliotti G. F. P., Domit C., Lorenzetti E., Bracarense A. P. F. R. L. (2022). Bacterial and fungal pathogens in granulomatous lesions of Chelonia mydas in a significant foraging ground off southern Brazil. Vet. Res. Commun. 46, 859–870. doi: 10.1007/s11259-022-09911-w
Donnelly K., Waltzek T. B., Wellehan J. F. X., Stacy N. I., Chadam M., Stacy B. A. (2016). Mycobacterium haemophilum infection in a juvenile leatherback sea turtle (Dermochelys coriacea). J. Vet. Diagn. 28, 6, 718–6, 721. doi: 10.1177/1040638716661746
Fistarol G., Coutinho F., Moreira A., Venas T., Cánovas A., De Paula JR S., et al. (2015). Environmental and sanitary conditions of guanabara bay, Rio de Janeiro. Front. Microbiol. 6. doi: 10.3389/fmicb.2015.01232
Fries A. S., Coimbra J. P., Nemazie D. A., Summers R. M., Azevedo J. P. S., Filoso S., et al. (2019). Guanabara bay ecosystem health report card: science, management, and governance implications. Reg. Stud. Mar. Sci. 25, 100474. doi: 10.1016/j.rsma.2018.100474
Gcebe N., Hlokwe T. M. (2017). Non-tuberculous mycobacteria in south African wildlife: neglected pathogens and potential impediments for bovine tuberculosis diagnosis. Front. Cell Infect. Microbiol. 7. doi: 10.3389/fcimb.2017.00015
Gcebe N., Michel A. L., Hlokwe T. M. (2018) Non-tuberculous Mycobacterium species causing mycobacteriosis in farmed aquatic animals of south AfricaBMC Microbiol. 18, 1, 1–1,11. doi: 10.1186/s12866-018-1177-9
Glazebrook J. S., Campbell R. S. F. (1990). A survey of the diseases of marine turtles in northern Australia. i. farmed turtles. Dis. Aquat. Organ. 9, 83–95. doi: 10.3354/dao009083
Goshe L. R., Avens L., Scharf F. S., Southwood A. L. (2010). Estimation of age at maturation and growth of Atlantic green turtles (Chelonia mydas) using skeletochronology. Mar. Biol. 157, 1725–1740. doi: 10.1007/s00227-010-1446-0
Greer L. L., Strandberg J. D., Whitaker B. R. (2003). Mycobacterium chelonae osteoarthritis in a kemp’s ridley sea turtle (Lepidochelys kempii). J. Wildl. Dis. 39, 3, 736–3, 741. doi: 10.7589/0090-3558-39.3.736
Henkle E., Winthrop K. L. (2015). Nontuberculous mycobacteria infections in immunosuppressed hosts. Clin. Chest Med. 36, 1, 91–1, 99. doi: 10.1016/j.ccm.2014.11.002
Ikuta C. Y., Morato F., Souza G. O., Heinemann M. B., Amaku M., Ferreira Neto J. S. (2016). Influence of the incubation conditions on culture media to optimize primary isolation of. Mycobacterium bovis. Semin. Cienc. Agrar. 37, 5, 3693–3700. doi: 10.5433/1679-0359.2016v37n5Supl2p3693
Kazda J., Falkinham J. O., Pavlik I., Hruska K. (2009). The ecology of mycobacteria: impact on animal’s and human’s health (Dordrecht, Netherlands: Springer).
Lacasse C., Terio K., Kinsel M. J., Farina L. L., Travis D. A., Greenwald R. (2007). Two cases of atypical mycobacteriosis caused by Mycobacterium szulgai associated with mortality in captive african elephants (Loxodonta africana). J. Zoo Wildl. Med. 38, 1, 101–1, 107. doi: 10.1638/06-051.1
Mitchell M. A. (2012). Mycobacterial infections in reptiles. Vet. Clin. Exot. Anim. 15, 101–111. doi: 10.1016/j.cvex.2011.10.002
Nardini G., Florio D., Di Girolamo N., Gustinelli A., Quaglio F., Fiorentini L., et al. (2014). Disseminated mycobacteriosis in a stranded loggerhead sea turtle (Caretta caretta). J. Zoo Wildl. Med. 45, 2, 357–2, 360. doi: 10.1638/2013-0252r1.1
Palmieri C., Roy P., Dhillon A. S., Shivaprasad H. L. (2012). Avian mycobacteriosis in psittacines: a retrospective study of 123 cases. J. Comp. Pathol. 148, 2–3, 126-138. doi: 10.1016/j.jcpa.2012.06.005
Piersimoni C., Scarparo C. (2009). Extrapulmonary infections associated with nontuberculous mycobacteria in immunocompetent persons. Emerg. Infect. Dis. 15, 9, 1351–1358. doi: 10.3201/eid1509.081259
Schmidt V., Köhler H., Heenemann K., Moebius P. (2022). Mycobacteriosis in various pet and wild birds from Germany: pathological findings, coinfections, and characterization of causative mycobacteria. Microbiol. Spectr. 10 (4), e0045222. doi: 10.1128/spectrum.00452-22
Sibley W. K. (1889). Uber tuberkulose bei wirbelthieren. Virchows Arch. 116, 104–115. doi: 10.1007/BF02282103
Slany M., Knotek Z., Skoric M., Knotkova Z., Svobodova J., Mrlik V., et al. (2010). Systemic mixed infection in a brown caiman (Caiman crocodilus fuscus) caused by Mycobacterium szulgai and M. chelonae: a case report. Vet. Med. 55, 2, 91–2, 96. doi: 10.17221/26/2010-VETMED
Smith V. H., Schindler D. W. (2009). Eutrophication science: where do we go from here? Trends Ecol. Evol. 24, 201–207. doi: 10.1016/j.tree.2008.11.009
Soldati G., Lu Z. H., Vaughan L., Polkinghorne A., Zimmermann D. R., Huder J. B., et al. (2004). Detection of Mycobacteria and Chlamydiae in granulomatous inflammation of reptiles: a retrospective study. Vet. Pathol. 41, 4, 388–4, 397. doi: 10.1354/vp.41-4-388
Suykerbuyk P., Vleminckx K., Pasmans F., Stragier P., Ablordey A., Tran H. T., et al. (2007). Mycobacterium liflandii infection in european colony of Silurana tropicalis. Emerg. Infect. Dis. 13, 743–746. doi: 10.3201/eid1305.060625
Telenti A., Marchesi F., Bally M., Bottger E. G., Bodmer T. (1993). Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analyses. J. Clin. Microbiol. 31, 2, 175–2, 178. doi: 10.1128/jcm.31.2.175-178.1993
Ullmann L. S., das Neves Dias-Neto R., Cagnini D. Q., Yamatogi R. S., Oliveira-Filho J. P., Nemer V., et al. (2016). Mycobacterium genavense infection in two species of captive snakes. JVATiTD 22, 1. doi: 10.1186/s40409-016-0082-7
Vieira R. P., Gonzalez A. M., Cardoso A. M., Oliveira D. N., Albano R. M., Clementino M. M. (2008). Relationships between bacterial diversity high levels of resistance and gene transfer elements. PloS One 6, e17038. doi: 10.1371/journal.pone.0017038
Williams B. A., Watson J. E. M., Beyer H. L., Klein C. J., Montgomery J., Runting R. K., et al. (2022). Global rarity of intact coastal regions. Conserv. Biol. 36:4, e13874. doi: 10.1111/cobi.13874
Willson S. J., Kaufman M. G., Merritt R. W., Williamson H. R., Malakauskas D. M., Benbow M. E. (2013). Fish and amphibians as potential reservoirs of Mycobacterium ulcerans, the causative agent of buruli ulcer disease. Infect. Ecol. Epidemiol. 3, 19946. doi: 10.3402/iee.v3i0.19946
Wilton S., Cousins D. (1992). Detection and identification of multiple mycobacterial pathogens by DNA amplification in single tube. Gen. Res. 1: 269–273. doi: 10.1101/gr.1.4.269
Woodhouse S. J., Fitzgerald S. D., Lim A., Bolin S. R. (2014). Disseminated Mycobacterium haemophilum infection in an Assam trinket snake (Elaphe frenata). J. Zoo Wildl. Med. 45, 4, 966–4, 969. doi: 10.1638/2014-0078.1
Keywords: chelonian conservation, mycobacteriosis, sea turtles, immunosuppression, uncommon infectious diseases
Citation: Goldberg DW, Alcala MM, Nóbrega DFd, Sellera FP, Fonseca S, Neto JSF, Nogueira DB and Heinemann MB (2023) Mycobacterium gordonae infection in a free-ranging green turtle (Chelonia mydas), Brazil. Front. Mar. Sci. 10:1197731. doi: 10.3389/fmars.2023.1197731
Received: 31 March 2023; Accepted: 23 May 2023;
Published: 02 June 2023.
Edited by:
Andrea D. Phillott, Flame University, IndiaReviewed by:
Yasunari Kiryu, Florida Fish and Wildlife Research Institute, United StatesErina J. Young, Murdoch University, Australia
Copyright © 2023 Goldberg, Alcala, Nóbrega, Sellera, Fonseca, Neto, Nogueira and Heinemann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daphne Wrobel Goldberg, ZGFwaHdyb2JlbEBnbWFpbC5jb20=