 Martha O’Keefe1†
Martha O’Keefe1† Elizabeth G. T. Bengil2,3†
Elizabeth G. T. Bengil2,3† Josie L. Palmer1†
Josie L. Palmer1† Damla Beton2Çiğdem Çağlar2
Damla Beton2Çiğdem Çağlar2 Brendan J. Godley1Meryem Özkan2Robin T. E. Snape1,2*
Brendan J. Godley1Meryem Özkan2Robin T. E. Snape1,2* Annette C. Broderick1*
Annette C. Broderick1*- 1Centre for Ecology and Conservation, University of Exeter, Penryn, United Kingdom
- 2Society for the Protection of Turtles, Gönyeli, Cyprus
- 3Cukurova University, Fisheries Faculty, Adana, Türkiye
Introduction: Loss of biodiversity in marine ecosystems is a globally acknowledged problem. To address this and avoid extinctions, improved conservation is required to protect and restore our oceans. Elasmobranch species are considered the second most threatened vertebrate lineage, with overfishing considered the most important driver of declines. The Mediterranean Sea is a hotspot for elasmobranchs, with the eastern basin considered a data-poor area.
Methods: This study sought to address that deficiency, using bycatch data to assess the diversity and distribution of elasmobranch species caught as a result of commercial fishing in Northern Cyprus from 2018 to 2022.
Results: Thirty-six elasmobranch species were recorded, including 24 new records for Northern Cyprus, representing 41% of all species recorded in the Mediterranean. Of these 36 species, 61% are listed on the IUCN Red List as globally threatened (Critically Endangered, Endangered or Vulnerable). Of the species recorded, 71% of sharks (n=17), and 53% of batoids (skates and rays; n=19) are listed as threatened. We present data on the distribution of captures and size classes and infer life stage from these data.
Discussion: Our study demonstrates the importance of the coastal waters of Cyprus for multiple life stages of a broad range of elasmobranch species. As most elasmobranchs caught are retained for bait, subsistence or trade, our study has highlighted the need to update the regulations for this small-scale fishery, and led to a recent amendment to prohibit trade in all species listed under the Barcelona Convention.
1 Introduction
Anthropogenic impacts continue to drive loss of function and degradation in marine ecosystems. Biodiversity within these systems has declined over time, and we are now facing the extinction of some of the world’s most important and charismatic species. Elasmobranchs (sharks, rays, skates and chimeras) are showing population declines globally, with more than 30% of species globally threatened with extinction (IUCN, 2022), and oceanic elasmobranch populations are thought to have decreased by at least two-thirds between 1971 and 2020 (Pacoureau et al., 2021). Elasmobranchs overall have a higher extinction risk than most other vertebrates, in part owing to their low fecundity, late maturation and long generation times, with large shallow water-dwelling species most at risk (Dulvy et al., 2014).
Although these declines are due to a cumulative range of anthropogenic impacts – habitat degradation, pollution and climate change, for example – overfishing, resulting in both targeted and incidental catch, is considered the most important driver (Dulvy et al., 2014; Dulvy et al., 2021). Across the world, elasmobranchs are fished by artisanal, recreational and commercial fisheries for their fins, livers and meat (Oliver et al., 2015; Tiralongo et al., 2018a) and it has been estimated from reported landings, unreported landings and discards that the global catch of sharks in 2010 was 1.41 million tonnes (Worm et al., 2013). As well as targeted fishing, a major problem facing elasmobranch populations is bycatch, the incidental capture of non-target species (Oliver et al., 2015). It has been estimated that bycatch makes up 40% of annual global marine catch (Davies et al., 2009). Bycaught elasmobranchs are increasingly being retained by fishers, due to their market value and decreases in target species catch, but a lack of management and legislation persists (Oliver et al., 2015; Pacoureau et al., 2021). Even if species are released alive, there can be sub-lethal impacts on individuals that could, in turn, lead to population impacts, as well as post-release mortalities (Wilson et al., 2014; Pacoureau et al., 2021).
Within the Mediterranean Sea, 88 species of elasmobranchs have been recorded (Serena et al., 2020), and many are facing declines in population size and range (Dulvy et al., 2016). Here, due to increased relative fishing pressure, 50% of skates and rays (hereafter referred to as batoids) and 56% of sharks, have regional International Union for Conservation of Nature (IUCN) Red List conservation status assessments that are elevated compared to their global populations (Dulvy et al., 2016). Historically, elasmobranch species diversity and richness were considered greatest in the western Mediterranean Sea, but in recent years there have been a greater number of local extinctions in the west and an increase in threats throughout the Mediterranean (Dulvy et al., 2016; Serena et al., 2020). There have been fewer studies of elasmobranch diversity in the eastern Mediterranean, although Türkiye alone has over three quarters of all Mediterranean elasmobranch species (Bengil and Basusta, 2018), with known nursery areas for Carcharhinus plumbeus (Nardo 1827, sandbar sharks), Rhinobatos rhinobatos (Linnaeus 1758, common guitarfish) and Glaucostegus cemiculus (Geoffroy St. Hilaire 1817, blackchin guitarfish) in the coastal waters of Türkiye (Bengil et al., 2020; Basusta et al., 2021).
Elasmobranchs make up just 1% of the annual fisheries catch in the Mediterranean (Bradai et al., 2018), however, fishing mortality due to bycatch is a major threat for this group (Bengil and Bengil, 2018). In most Mediterranean countries, elasmobranch bycatch is not reported by species, leading to underestimations of catches and poorly informed conservation efforts (Cashion et al., 2019; Giovos et al., 2021a). Many elasmobranch studies use data from large industrial fishing vessels such as trawlers and longliners (Molina and Cooke, 2012). Given that at least eighty percent of Mediterranean fishing vessels are small-scale vessels (<12m total length; FAO, 2020), and small-scale fisheries (SSF) have recently been identified as the second greatest contributor to bycatch of threatened elasmobranchs in the Mediterranean (after longline vessels; Tiralongo et al., 2018a; Carpentieri et al., 2021), understanding bycatch within the Mediterranean SSF fleet is urgently needed.
A checklist of 60 chondrichthyans was recently reported by a study which reviewed available data, studies and citizen science reports for the island of Cyprus (Giovos et al., 2021b). A published record of Hexanchus nakamurai (Teng 1962, bigeyed sixgill shark, now H. vitulus, Springer and Waller 1969, Atlantic sixgill shark) has since been retracted (Bengil et al., 2021), while a Cetorhinus maximus (Gunnerus 1785, basking shark) previously reported for Northern Cyprus (Kabasakal, 2013), and for the whole island (Giovos et al., 2021b) was based on a newspaper report which was later found to have been inaccurate, instead being Alopias superciliosus (Lowe 1841, bigeye thresher shark, Hakan Kabasakal pers. comm.). This brings the total species count for the entire island to 58 (30 sharks and 28 batoids). Most of the contributing records, however, are from studies focused in the area of the island under the effective control of the Republic of Cyprus (RoC), and until our current study, just fourteen species had been documented in Northern Cyprus (Turkish Republic of Northern Cyprus (TRNC); Oray and Karakulak, 2005; Çoker and Akyol, 2014; Akbora et al., 2019) a self-declared state recognised only by Türkiye and considered by the international community to be part of the Republic of Cyprus (hereafter referred to as Northern Cyprus).
The commercial fishing fleet of Northern Cyprus includes 300-400 SSF vessels [Northern Cyprus Department of Animal Husbandry (DAH)], predominantly fishing with gill nets, trammel nets and longlines. Interactions with marine vertebrate species, including elasmobranchs, marine turtles, Mediterranean monk seals (Monachus monachus, Hermann 1779) and dolphins are common, with an estimated 1,000 marine turtles caught in this fishery each year (Snape et al., 2013; Snape et al., 2018b; Beton et al., 2021). Trawl fishing is not permitted and there are no industrialised longline vessels. Although, marine protected areas (MPAs) have been established to protect sea turtles, monk seals, seabirds and some key habitats across more than a quarter of the coast of Northern Cyprus, (Snape et al., 2018a), few fisheries restrictions are implemented within or outside of these MPAs. Until May 2023, there were no formal protection measures for any elasmobranch species from commercial trade, other than for C. plumbeus and C. maximus of which catching or landing were prohibited (Fisheries Law 27-2000, 2022). This was partly as a result of the lack of data available for policy makers, however, as a result of data from this current study, in May 2023 the fisheries regulations were updated to prohibit trade of all species listed on Annex I of the Barcelona Convention. The authorities have also maintained a blanket ban on industrialised fishing, with no trawling permitted since the 1990s (Ulman et al., 2015), thus, habitats may be in favourable condition compared with other parts of the Mediterranean, where bottom trawling persists.
This study aimed to describe the diversity, distribution and life stages of elasmobranch species caught in the fisheries of Northern Cyprus, through deploying onboard observers and engaging small-scale fishers in data collection, to inform fisheries management and conservation action in line with global, regional and national biodiversity conservation strategies.
2 Materials and methods
2.1 Study area
Cyprus is the third largest island in the Mediterranean Sea, divided into two geopolitical subregions by the United Nations-monitored buffer zone, the Green Line (Sabri and Sakallı, 2021). This study focuses on the coastal waters of Northern Cyprus (Figure 1), where small-scale coastal fisheries operate using wooden boats of <12m length with inboard diesel engines (Ulman et al., 2015), using bottom-set gillnets and trammel nets, benthic longlines, with a small number of pelagic longlines (Snape et al., 2013). Fourteen fishing harbours are maintained and managed by the DAH and all commercial fishing vessels are registered to these ports (Figure 1). The informal shelter at Apostolos Andreas (Ap Andreas; Figure 1) was also included in our study as it is frequently used by fishers during the summer months. Active fishing vessel data were available from the DAH for coastal zones 1, 2 and 3 (Zone 3 was subdivided into three areas (a, b and c) for finer spatial resolution resulting in a total of five areas; Figure 1) and were used to plan observer trips and recruit self-reporting fishers to ensure our data were representative of the fleet.
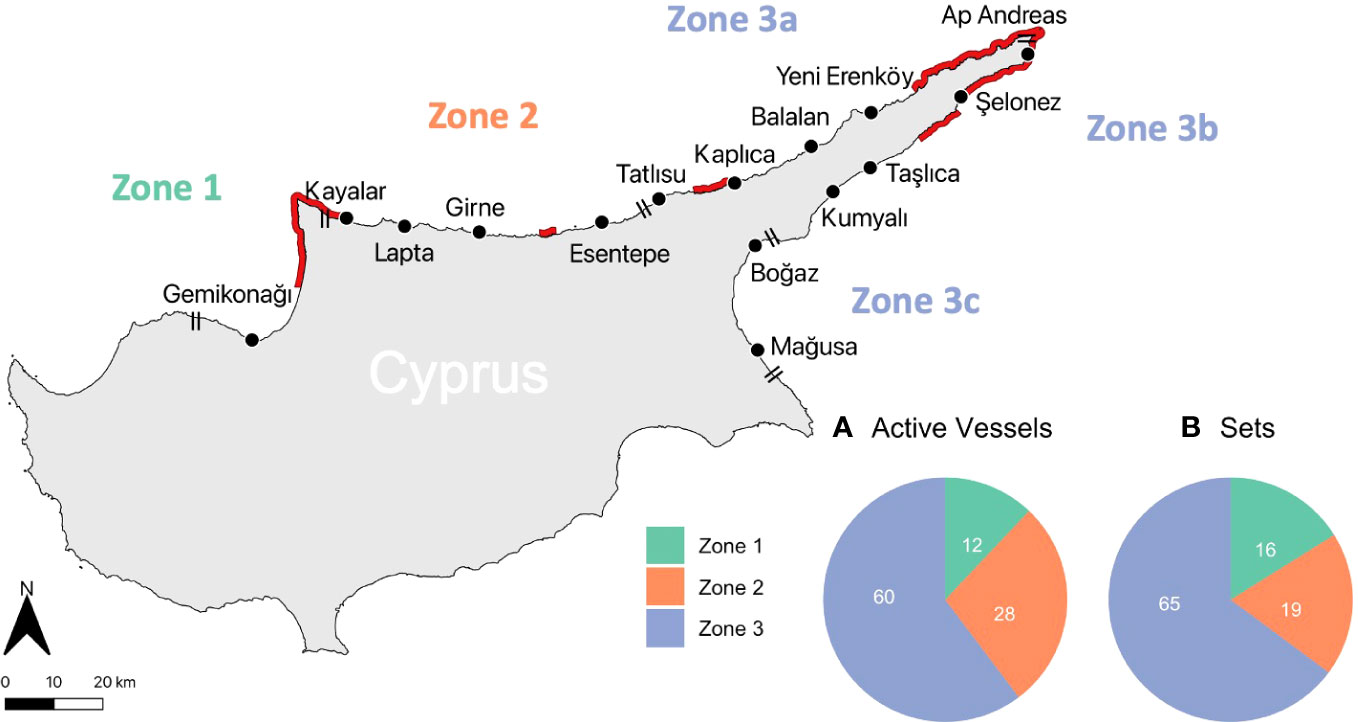
Figure 1 Study area, with 15 harbours, in three coastal zones, regularly used by fishers in Northern Cyprus. Double black lines show the division of harbours into coastal zones used by the authorities 1, 2 and 3; single black lines show further division of Zone 3 into three separate areas (a, b and c) in this study. Inset: Pie chart A shows proportion of the total active fishing vessels operating from each coastal zone in 2018/2019 (n = 340), with data provided by Department for Animal Husbandry for 2019 (the most recent available data); pie chart B, the proportion of all sets (n=1,899) from which departure port was recorded during 2018-2022, divided into coastal zone. Location of designated Marine Protected Areas (MPAs) is also shown. Greek place names (west to east) are Karavostasi, Orga, Lapithos, Kyrenia, Agios Amvrosios, Akanthou, Davlos, Platanissos, Gialousa, Apostolos Andreas, Chelones, Neta, Koma tou Gialou, Trikomo, Famagusta.
2.2 Data collection
Data on elasmobranch bycatch were collected between 04/01/2018 to 19/06/2022 through the Cyprus Bycatch Project (an island-wide project to understand and mitigate bycatch of vulnerable species) in the following three ways.
2.2.1 Onboard observers
Trained onboard observers broadly followed the General Fisheries Commission for the Mediterranean (GFCM) protocol (FAO, 2020). For every set observed, the time of deployment and GPS coordinates at deployment and hauling were taken. When elasmobranch bycatch occurred, individual specimens were removed from the nets or hooks by either the fishers or onboard observers. Where possible, individuals were measured using a flexible tape measure; for batoids, disc width and total length were taken and for sharks total length (FAO, 2020). Tails of Dasyatidae were often removed by fishers prior to handling; instances of tail removal were noted where possible and these individuals were removed from total length distributions. Photographs were taken for records and to confirm identification. Identification was made to the lowest possible taxonomic group using Otero et al. (2019), and other literature and expert knowledge as required. Observers were not always able to assess maturity from external characteristics, therefore, maturity was estimated using L50 values – the total length at which 50% of the population are sexually mature – found in the peer-reviewed literature and using data from the Mediterranean where available or the lowest L50 estimations available globally (Supplemental Tables 1, 2; Supplemental References). The conservation status and common names for species were taken from the IUCN RedList (IUCN, 2022) for both global and regional status. For species endemic to the Mediterranean, that only have a global Redlist status, this was also included for the Mediterranean status. We recorded whether animals were alive or dead at hauling and from this calculated at-vessel mortality score for each species (AVM; percentage of individuals dead on hauling). Observers always encouraged and trained fishers to release live individuals where possible.
2.2.2 Self-reporting
In 2018 and 2019, prior to the COVID-19 pandemic, commercial fishers were trained to report catch and bycatch, using data sheets provided, in return for a subsidy per fishing trip. Fishers shared photographs of species caught via telephone applications and completed a simplified form which was the same as the onboard observer forms except individual fish measurements and the status of individuals on hauling were not recorded. A fisheries liaison officer was employed to visit fishers regularly during this period, to collect and replenish forms every two months during active fishing. In some cases where specimens were retained for trade, observers attended the port to take measurements and maturity was estimated using L50 values.
2.2.3 Opportunistic data
Further data were collected opportunistically using local newspaper, social media posts and direct information from fishers who were not part of the self-reporting study, including amateur and sports fishers who represent a much broader demographic and for whom no data on fishing effort and distribution are estimated. Photographs were provided for identification, and where possible, measurements and locations of capture were collected from follow up conversations with the fisheries liaison officer. Fishers could also make contact regarding elasmobranch bycatch, and when able, observers met the fishers to measure the individual, and take photographs, samples or the whole specimen, and again, maturity was estimated using L50 values.
2.3 Spatial analysis
Using the median location calculated from the start and end coordinates of onboard observer sets, the spatial distribution of (1) number of observed sets, (2) number of sets with one or more individuals of each species, and (3) bycatch per unit effort (BPUE), were mapped and presented using a 100 km2 tessellating hexagonal polygon matrix. BPUE was calculated as the number of individuals bycaught per 1,000 m for set nets or per 1,000 hooks for longlines.
3 Results
A total of 1,702 sets were recorded, 730 by onboard observers and 972 by self-reporting fishers. These included 1,367 demersal net sets, 282 demersal longline sets and 2 pelagic (surface/epipelagic) longline sets, with average set depths of 34.2 m (SD: 23.7 m, range: 1.0-250.0 m), 53.5 m (SD: 39.7 m, range: 6.0-296.0 m) and 162.2 m (SD: 92.4 m, range: 96.8-227.5 m), respectively. The number of observed sets by year and month together with the proportion of these that had elasmobranch bycatch is shown in Supplementary Figure 1.
3.1 Species composition
A total of 1,205 individual elasmobranchs were recorded (2018: 294, 2019: 591, 2020: 140, 2021: 63, 2022: 117), of 36 different species (batoid n=19; shark n=17; Figure 2), including two of the three endemic to the Mediterranean (Raja polystigma, Regan 1923, speckled skate and Raja radula, Delaroche 1809, rough skate). During the study period, 160 (13%) individuals of 28 species were recorded opportunistically. Of these, seven were only recorded through opportunistic observations (R. rhinobatos; Prionace glauca, Linnaeus 1758, blue shark; Mobula mobular, Bonnaterre 1788, spinetail devil ray; Oxynotus centrina, Linnaeus 1758, angular roughshark; A. superciliosus; Hexanchus griseus, Bonnaterre 1788, bluntnose sixgill shar; Odontaspis ferox, Risso 1810, smalltooth sand tiger). In addition, for completeness, we have included a further two unique species records from opportunistic data from outside the study period (Carcharhinus brachyurus, Günther 1870, copper shark; Carcharodon carcharias, Linnaeus 1758, white shark), recorded in 2015 and 2017 respectively.
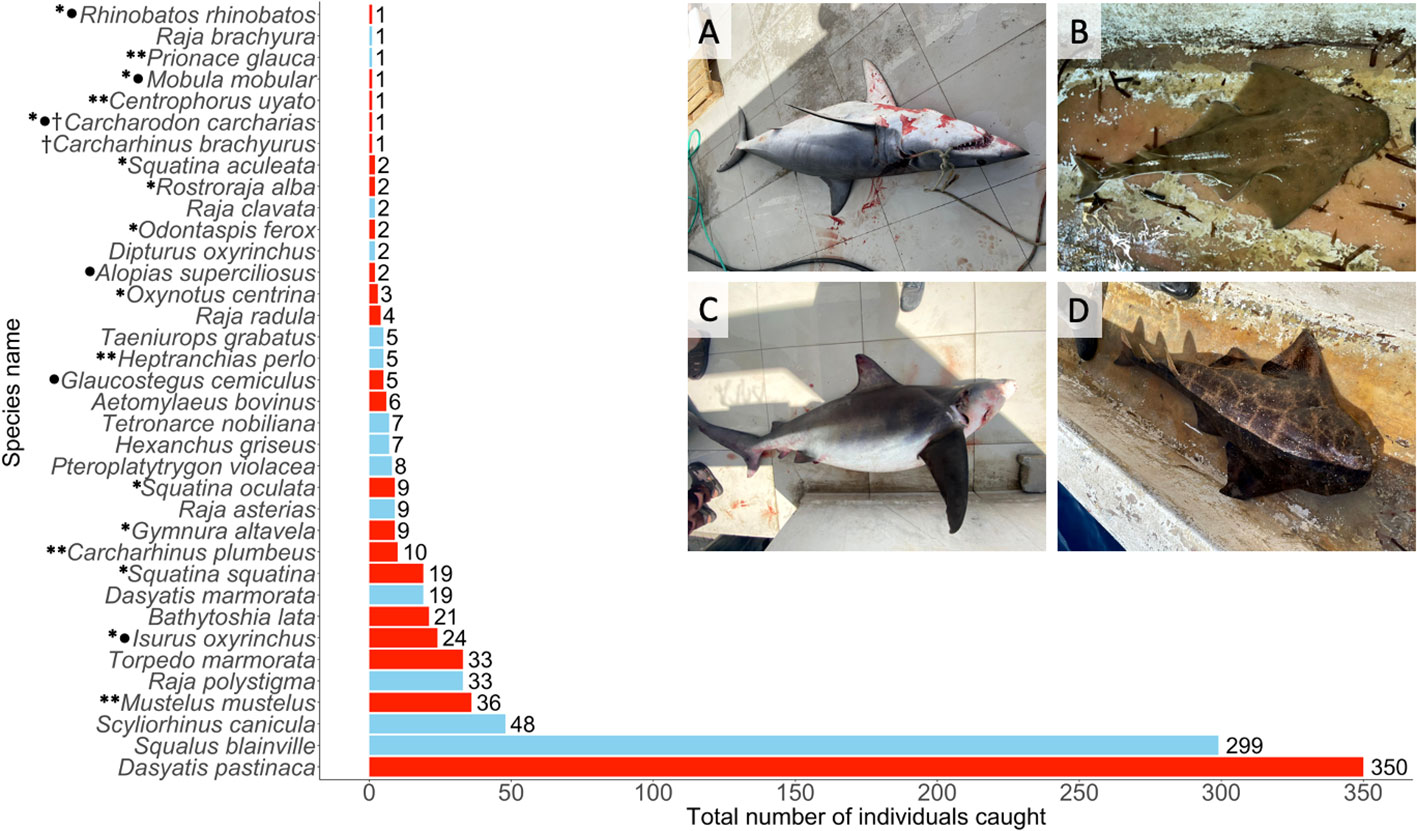
Figure 2 Total individuals (n = 988) that could be identified to species level caught as bycatch. Numbers on bars show individual count per species, red bars indicate species with critically endangered, endangered and vulnerable IUCN Red List status. * denotes species included in Annex II and ** denotes species in Annex III of the SPA/BD Protocol covered by GFCM/36/2012/3 and GFCM/42/2018/2, • denotes species listed in Appendix II of the CITES, † denotes species recorded outside of 2018-2022 data collection. Inset pictures show examples of critically endangered species observed as bycatch: (A) I. oxyrinchus, (B) S. oculata, (C) C. plumbeus, (D) S. squatina.
Of the species encountered, 61% are listed on the IUCN RedList as globally threatened (17% Critically Endangered; 25% Endangered; 19% Vulnerable; Figure 3) whereas 50% of recorded species are listed as regionally threatened in the Mediterranean [28% Critically Endangered; 17% Endangered; 6% Vulnerable (Supplementary Figure 2, note values rounded up within categories)]. Twenty-four species had not previously been recorded in Northern Cyprus (Tables 1, 2).
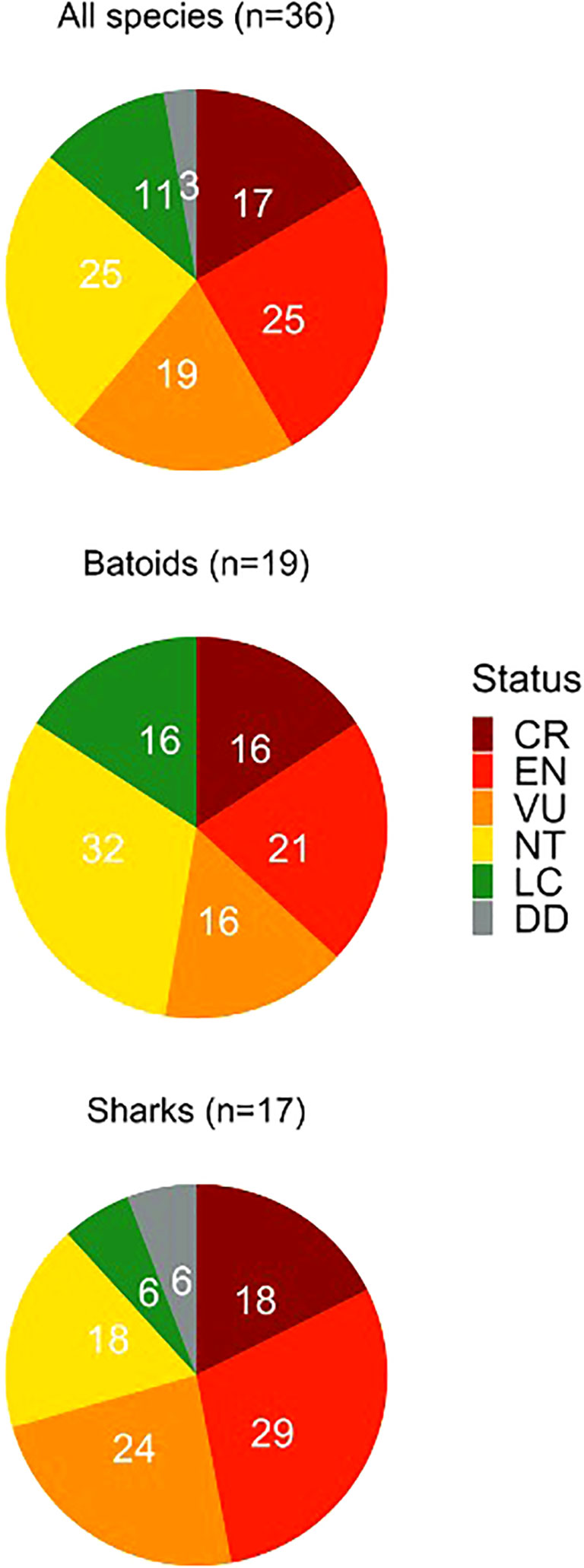
Figure 3 IUCN global Red List status for all species caught as bycatch across study period, subdivided into batoid and shark species. The categories for classifying a species risk of extinction: CR, critically endangered; EN, endangered; VU, vulnerable; NT, near threatened; LC, least concern; and DD, data deficient.
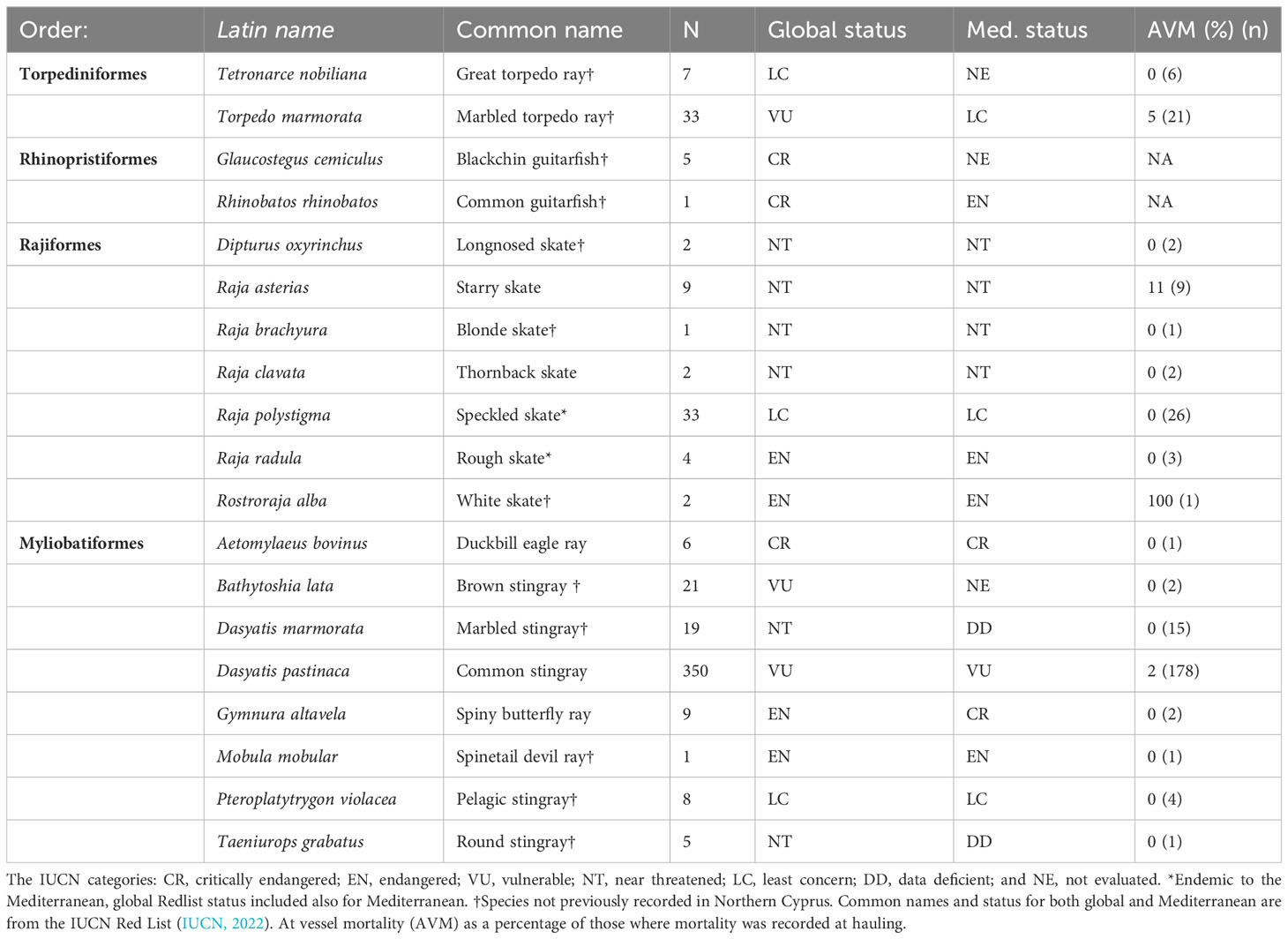
Table 1 Batoid species caught across this study.
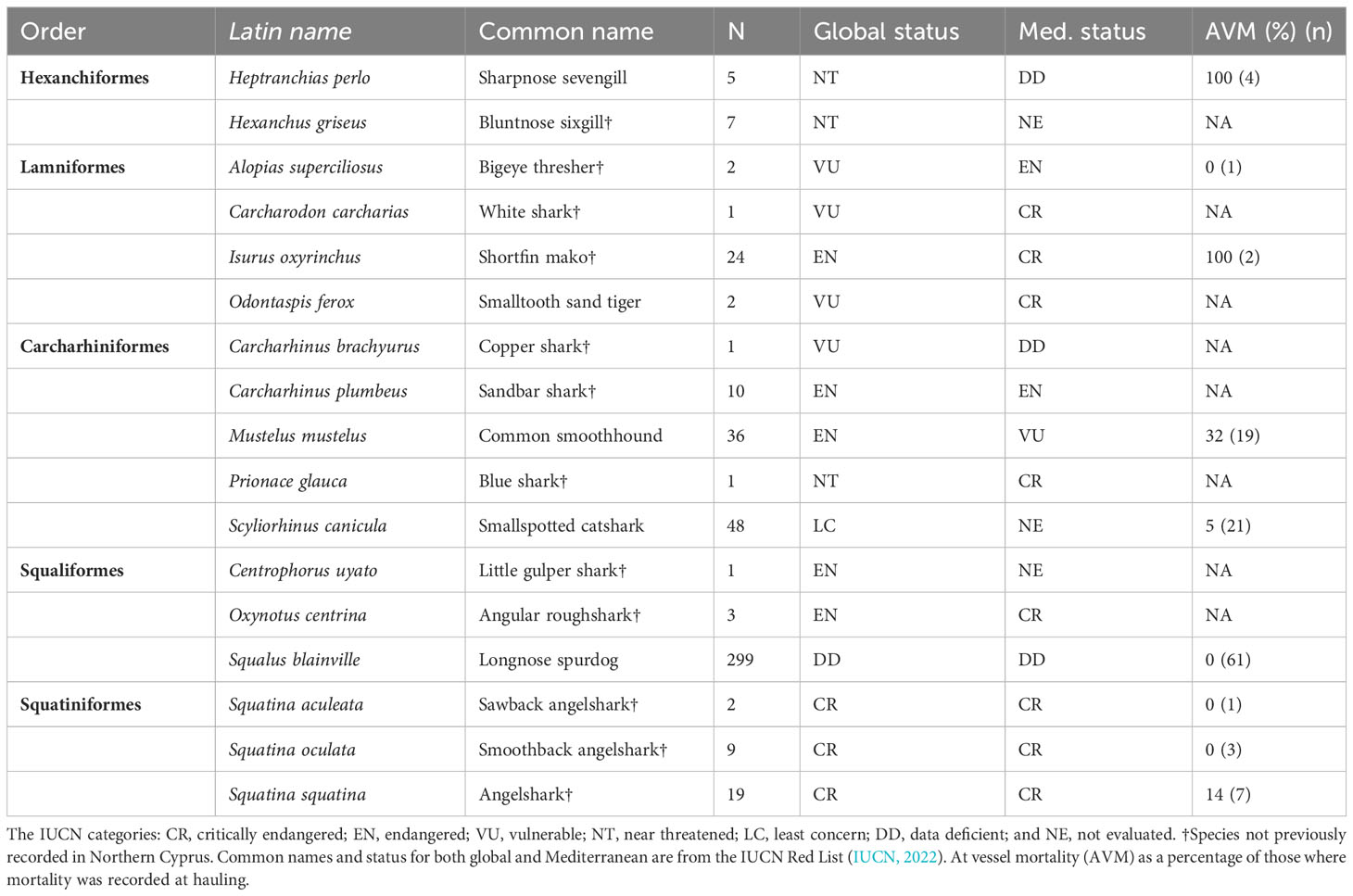
Table 2 Shark species caught across this study.
The most abundant species recorded was Dasyatis pastinaca (Linnaeus 1758, common stingray, n=350), followed by Squalus blainville (Risso 1827, longnose spurdog, n=299) and Scyliorhinus canicula (Linnaeus 1758, smallspotted catshark, n=48; Figure 2). Fifteen species caught throughout this study are listed by the Barcelona Convention, and banned from fishing and retention in the Mediterranean by the GFCM (Carpentieri et al., 2021; Figure 2). Six species are listed on Appendix II (international trade monitored and controlled) of CITES (Convention on the Trade in Endangered Species of wild fauna and flora): A. superciliosus, C. carcharias, M. mobular, Isurus oxyrinchus, (Rafinesque 1810, shortfin mako), G. cemiculus and R. rhinobatos (Figure 2). All extant Mediterranean representatives of the critically endangered Squatina spp. (angelsharks) were recorded in this study, namely Squatina squatina (Linnaeus 1758, angelshark), Squatina oculata (Bonaparte 1840, smoothback angelshark) and Squatina aculeata (Cuvier 1829, sawback angelshark; Figure 2).
3.2 Life stages and mortality
Size class distributions of a selective group of threatened batoid and shark species/families are presented in Figure 4, and by individual species, where available, in Supplemental Figures 3, 4. The largest individual measured was from an opportunistic record of I. oxyrinchus at 270 cm in total length, however, some individuals that were opportunistically recorded but were not measured, appeared larger than the maximum recorded from photographs, such as a A. superciliosus caught on the west coast estimated at >4 m. The smallest individuals was a D. pastinaca that had a total length of 16 cm (Supplementary Table 1). Most species with total length measurements and L50 estimates were likely dominated by juveniles, except for Torpedo marmorata (Risso 1810, marbled torpedo ray), D. pastinaca and S. blainville for which 66.7%, 65.9% and 56.7% of females and 85.7%, 66.7% and 66.7% of males exceeded their sex specific L50s, respectively.
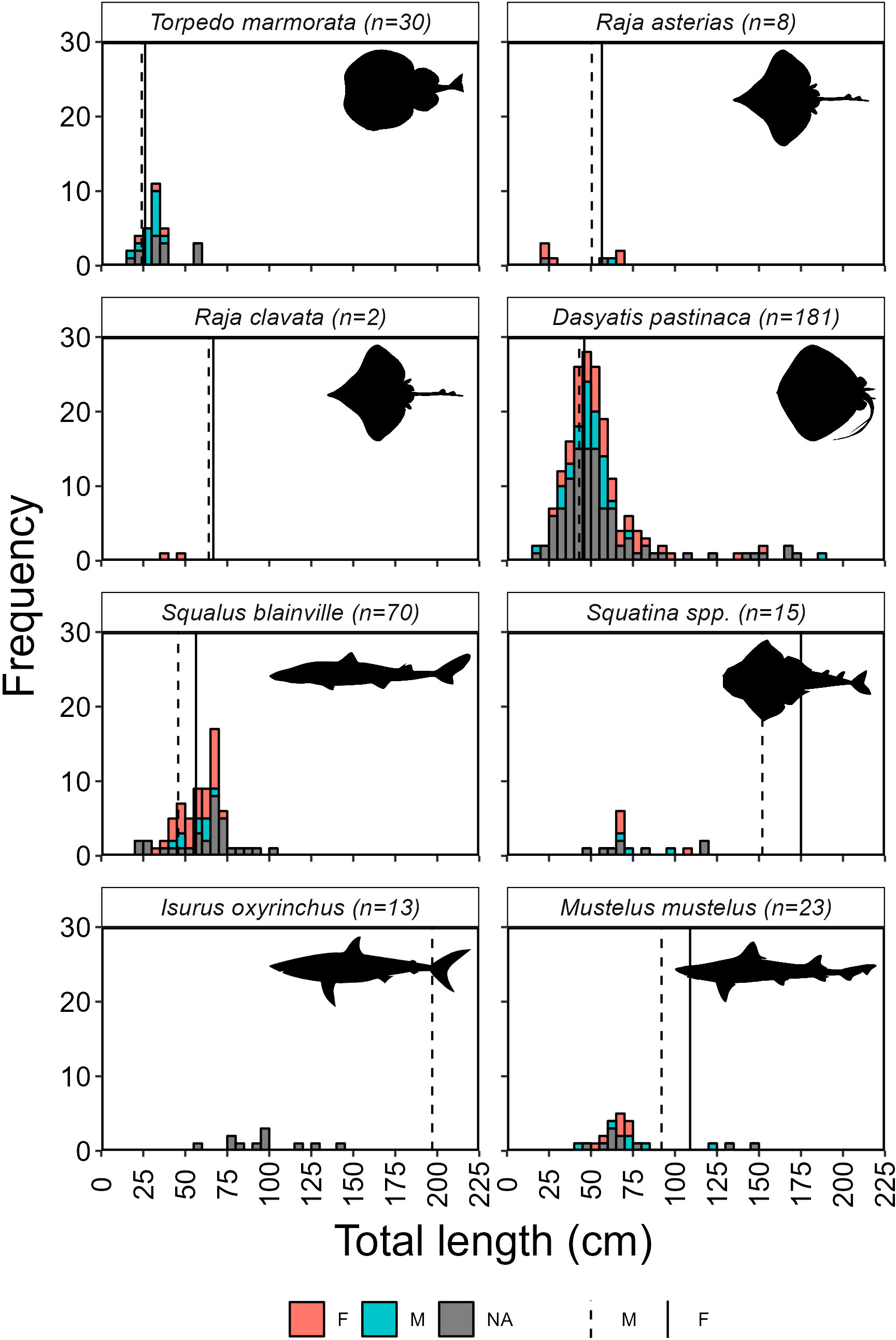
Figure 4 Size distribution of bycaught ray (T. marmorata, R. asterias; Raja clavata thornback skate; D. pastinaca) and shark species (S. blainville, Squatina spp.*, I. oxyrinchus and M. mustelus) recorded during onboard observations and self-reported trips between 2018 and 2022. *Squatina sizes are derived from all three species recorded in this study as well as any unidentified Squatina individuals. Total length at 50% maturity (L50) are given for females (solid line) and males (dashed line) of each species; L50 for Squatina is taken from S. aculeata estimates; L50 of female I. oxyrinchus (282.0 cm) exceeded the range of data. Illustrations from https://www.phylopic.org/.
The large number of S.blainville (n=137) caught in one set, were mostly of juvenile size, but included pregnant females. In addition, one fisher provided video footage of a S. oculata birthing on the west coast, while a large Aetomylaeus bovinus (Geoffroy Saint-Hilaire 1817, duckbill eagle ray) was filmed birthing on the deck of a boat on the north coast. Also worthy of note is that 10 out of the 25 I. oxyrinchus recorded were newborn or 1 - 2 year old and two were sexually mature, possibly pregnant, females (according to obtained length information and photos of the individuals).
At vessel mortality was higher for sharks (28%) than batoids (7%) with some species such as Heptranchias perlo (Bonnaterre 1788, sharpnose sevengill shark), I. oxyrinchus and Rostroraja alba (Lacepède 1803,white skate) having 100% mortality on hauling, although sample sizes are low (Tables 1, 2).
3.3 Spatial distribution
Figure 5 shows the spatial distribution of BPUE for sharks (Figures 5A, B) and batoids per 100 km2 (Figures 5C, D); location of all observer sets are shown by the presence of shaded hex cells in Figure 5, and the number of observed fishing operations per hex cell in Supplementary Figure 5 (Observed sets panel). High-diversity areas of elasmobranch catch were at the eastern (Karpaz (Karpas)) and western (Koruçam (Kormakitis)) capes, and in Famagusta and Güzelyurt (Morphou) Bays, although, onboard observer effort was higher in Famagusta Bay due to favourable weather conditions in this area (Supplementary Figure 5). Dasyatis spp. were the most abundant genus caught across the study (n = 401), followed by Squalus spp. (n = 299) and Raja spp. (n = 115). Dasyatis spp. was one of the most abundant genus caught in all five of the coastal areas across the three zones, and Squalus spp. in areas 1, 2, 3a and 3c (Supplementary Figure 5). If the mass bycatch event that occurred in 2019 in Zone 3a were removed, Squalus spp. would no longer be one of the top three genus in this area; the order would be Dasyatis spp., Torpedo spp. and Raja spp.

Figure 5 Spatial distribution of mean bycatch per unit effort (BPUE) derived from onboard observations with location data (n=664 fishing operations) of sharks (A, B) and batoids (C, D) in set nets (A, C) and longlines (B, D) per 100 km2. Cells shown indicate those where observer trips occurred; NULL cells are those where BPUE could not be calculated with the available data. Greek names for coastal areas indicated are (A) Cape Kormakitis, (B) Cape Karpas, (C) Morphou Bay, (D) Famagusta Bay.
4 Discussion
Using novel methods in an under-studied, yet regionally important fleet, this study helps further our understanding of elasmobranch diversity and distribution in Cyprus, and the contribution of this coast to Mediterranean biodiversity. It also provides detailed insight into the interaction of elasmobranchs with small-scale fisheries and their wider ecology in the study area. We recorded 36 elasmobranch species in the coastal waters of Northern Cyprus, 61% of which are considered globally threatened and 50% regionally threatened in the Mediterranean (IUCN, 2022). Of these species, two of the three Mediterranean endemic species (R. polystigma and R. radula) were recorded and 24 species (65%) were previously unpublished records for Northern Cyprus, bringing the total number of species recorded from 14 (Çoker and Akyol, 2014; Akbora et al., 2019) to 39. Nearly half (47%) of the 36 species we report are priority species under the GFCM or listed on Appendix II of CITES (Figure 2), meaning that international trade should be controlled.
The most abundant threatened elasmobranch species (Figure 2) caught in our study were D. pastinaca (VU), Mustelus mustelus (Linnaeus 1758, common smoothhound EN) and T. marmorata (VU), differing from the top three previously reported by Carpentieri et al. (2021) for small-scale fisheries in the eastern Mediterranean (C. plumbeus, M. mustelus and M. mobular). They also differ from the top three most abundant threatened species in both the western Mediterranean (Alopias spp., P. glauca and I. oxyrinchus) and central Mediterranean (C. plumbeus, M. mustelus and G. cemiculus). This regional variation in elasmobranch species composition may be driven by habitat or prey availability, or intensity of threats such as coastal development or fisheries activity, especially the lack of trawling activity in Northern Cyprus.
Three species that had previously been reported in Northern Cyprus, but were not found in this study were: Etmopterus spinax (Linnaeus 1758, velvet belly lanternshark), Galeus melastomus (Rafinesque 1810, blackmouth catshark) and Mustelus punctulatus (Risso 1827, blackspotted smoothhound; Oray and Karakulak, 2005; Çoker and Akyol, 2014). E. spinax and G. melastomus have typically been recorded in deep water trawls (Bengil and Basusta, 2018), and thus may be unlikely to have been caught in this SSF fleet where trawling is prohibited. M. punctulatus has apparently been shifting periodically in the Mediterranean since the 1920s (Colloca et al., 2017), and although it had previously been reported in the area of Cyprus under the effective control of the RoC Authorities (Hadjichrisophorou, 2006) there have been no reports since 1984. It is not surprising that our study did not find all 58 species considered to be present in waters of the whole island, because, compared to Northern Cyprus, the fisheries in the RoC controlled area are more diverse (with active trawl and pelagic longline fleets), because data availability for the latter region is far greater (partly due to the isolation of Northern Cyprus from many data collection frameworks), and because our study was restricted to include systematic methods and opportunistic observations during only recent years. In addition, it is worthy of note that fishermen from the Karpaz (Zone 3) region talk about the rare presence and capture of Sphyrna spp. (hammerhead sharks) but there have been no recent records in the area.
Although there were few species for which we had multiple measurements, clearly both adult and juvenile life stages of many species are present in coastal waters of Northern Cyprus. Many coastal elasmobranch species are known to use inshore nursery grounds with high productivity that offer protection from predators (Heupel and Simpfendorfer, 2002; Tiralongo et al., 2018b), and the large bycatch event of S. blainville of both juvenile and adult pregnant females, suggests there is a nursery ground for this species at least in the Karpaz region, with previous records supporting this (Bengil, 2022). From the Squatina size classes and birthing events reported, and the presence of new-born I. oxyrinchus it appears that Northern Cyprus may also be an important breeding and nursery area for these Critically Endangered and Endangered species.
A large proportion of the species found in Northern Cyprus are coastal – A. superciliosus, Bathytoshia lata (Garman 1880, brown stingray), Dasyatis marmorata (Steindachner 1892, marbled stingray), Taeniurops grabatus (Geoffroy St. Hilaire 1817, round stingray), D. pastinaca, A. bovinus and Raja asterias (Delaroche 1809, starry skate) – or found on the continental shelf and upper slopes, making them vulnerable to interactions with small-scale fisheries that are concentrated along the coast (Carpentieri et al., 2021). The majority of elasmobranch species caught by small-scale fisheries in the Mediterranean are demersal species (Carpentieri et al., 2021) caught in trammel nets or trawls, however, in Northern Cyprus trawling is not permitted, and the majority of fishers use trammel nets. Gear modifications or bycatch reduction technology could be considered to mitigate the impact of this fishery, however, a more detailed analysis of the drivers of bycatch is required to allow for greater planning and prioritisation of mitigation measures to more discrete métiers. Greater temporal analysis of catches could also help focus conservation efforts to seasonal métiers. With only Türkiye recognizing Northern Cyprus, it often falls outside international conventions, research and conservation networks which creates a challenging environment for funding the monitoring and management of the fishery.
Since the decision of fishers to retain (for trade or bait) or release was influenced by the presence of onboard observers and interaction with the authors, we have not presented data on survivorship and release rates after hauling. However, there is currently little incentive to release specimens. The relatively low vessel mortality rates (% dead on hauling) for many species illustrate the opportunity to drastically reduce bycatch mortality rates through a successful release programme in this fishery. While momentum is being gained from an ongoing education and awareness raising programme in collaboration with local authorities, the current legal trade in endangered elasmobranch species must be tackled. Recent legislative changes are welcomed but enforcement remains a challenge. Meanwhile, it is commendable and highly positive for marine conservation, that the authorities have been able to eliminate bottom trawling. However, small-scale fishers in Northern Cyprus are currently permitted to use drift nets which are banned across the rest of the Mediterranean because of their high impacts on large pelagic species including elasmobranchs. To our knowledge, drift nets are not in use and have never been noted on any vessels or in ports, yet, their use should be prohibited to avoid their possible future use.
A larger number of individuals were caught in coastal Zone 3, indicative of the higher number of active vessels and greatest observer coverage compared to other zones (Figure 1). When looking at BPUE however, several hotspots around Mağusa (Famagusta Bay), west of Cape Koruçam and the eastern Karpaz region can be seen (Figure 5). Existing MPAs (Figure 1) appear to overlap with these areas of high elasmobranch BPUE and so may be well-placed for continued work to designate strictly protected areas within them. Indeed, two candidate Important Shark and Ray Areas have been identified in Northern Cyprus which also overlap with these hotspots and the Akdeniz MPA on the west coast, and the two Karpaz MPAs (Figure 1; https://sharkrayareas.org/e-atlas/). In the management plans for these MPAs, that were primarily put in place to protect marine turtles, monk seals, seabirds and seagrass beds, fishing with set nets is prohibited to 30 m deep and to 1.5 km offshore, although this has never been enforced. Further analysis with larger observed effort will be useful in confirming such hotspots, refining MPA management plans, effectively integrating no-take zones to protect multiple taxa and developing new MPAs. Further studies should aim to increase onboard observation and fill the spatial gaps in our survey coverage, especially along the west and central north coasts.
The results of this study have been shared with the Northern Cyprus authorities and a proposal to expand the list of protected species and introduce the first no-take zones was accepted and policy came into effect in May 2023. Meanwhile, the strong reaction of some fishers participating in the project, sharing videos of themselves releasing threatened elasmobranch species, are an encouraging reaction to a programme of education delivered during this project. The recognition of the value and diversity of elasmobranchs among fishers is critical, so that retention moves from being the norm, to becoming a taboo, since enforcement of elasmobranch trade bans also has inherent challenges. Bycatch mitigation gear for dolphins (Snape et al., 2018b) and turtles has been trialled in this fishery, and could be expanded to elasmobranchs in certain métiers, or at least for fishers operating in MPAs.
This study has underlined the value of collaborating with SSFs to identify marine biodiversity. Results support the existence of important elasmobranch species which should be protected from expanding fishing pressure and trade, through the development of strictly protected areas/no-take zones, development of mitigation, education, and most importantly, implementing and policing the recent ban on elasmobranch trade and no-take zones. Although management of small-scale fisheries is extremely difficult, high community engagement, education and engaging fishers through SSF co-management, can effect change (Piovano et al., 2012). As data in this location continues to grow, species-specific analysis of critically endangered elasmobranchs must be undertaken to further inform conservation planning and management of fisheries. More fine-scale distribution data would also be of benefit, to improve the accuracy and identification of species richness hotspots, and reliability of management actions. With nearly half (44%) of Mediterranean elasmobranch diversity across less than 1% of its coastline, Northern Cyprus should be considered a regional conservation priority for this taxonomic group.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The animal study was approved by University of Exeter Ethics Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RTES, DB, BJG and ACB contributed to the conception and design of the study. MaO, EGTB, JLP, DB, CC, MeO, RTES, collected data. MaO, EGTB, JLP, RTES and ACB developed the first draft of the manuscript and all authors contributed to further drafts. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by the MAVA Foundation through the Cyprus Bycatch Project to the Society for the Protection of Turtles (Project 20146-20123) and the University of Exeter (Projects 17149 and 20148-20123) and the Cyprus Elasmobranch Research and Conservation Network (Project 190710). JLP was funded through a PhD studentship from SPOT and the University of Exeter.
Acknowledgments
The Cyprus Bycatch Project was funded by the MAVA foundation, coordinated by BirdLife International and was a collaboration between BirdLife Cyprus and Enalia Physis who managed work in the Republic of Cyprus controlled area, and the Society for the Protection of Turtles (SPOT) and University of Exeter who manage work in Northern Cyprus. We thank all those onboard observers who helped with data collection and the fishers who took part in this study. Specific thanks to the director and all staff at the Department of Animal Husbandry of the Northern Cyprus Ministry for Agriculture and Natural Resources, who have been fundamental in securing permits for observers to participate in fishing activities and supportive of the work and aims of this study. We thank Fabrizio Serena for help with identification of some specimens.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1181437/full#supplementary-material
References
Akbora H. D., Bayri E., Ayas D., Çiçek B. A. (2019). Recent record of Odontaspis ferox (Risso 1810) in Northern Cyprus (Eastern Mediterranean Sea). J. Black Sea/Mediterranean Environ. 25 (3) 315–320.
Basusta N., Basuta A., Ozyurt C. E. (2021). Evidence of a second nursery area of the sandbar shark, Carcharhinus plumbeus (Nardo 1827) in the Eastern Mediterranean Sea. Mediterr. Mar. Sci. 22 (1), 20–26. doi: 10.12681/mms.24490
Bengil E. G. T. (2022). Biology and ecology of squalus Blainville (Risso, 1827) from the eastern Mediterranean. Thalassas:Int. J. Mar. Sci., 38(2):1423–1432. doi: 10.1007/s41208-022-00482-w
Bengil E. G. T., Akbora H. D., Hadjioannou L., Papageorgiou M., Snape R. T. E. (2021). A new species in town: New record of Hexanchus nakamurai Teng 1962 from the Levantine Sea. J. Appl. Ichthyology 37, 113–119. doi: 10.1111/jai.14125
Bengil E. G. T., Basusta N. (2018). Chondrichthyan species as by-catch: A review on species inhabiting Turkish waters. J. Black Sea/Mediterranean Environ. 24 (3), 288–305.
Bengil E. G. T., Bengil F. (2018). Less known species of North Cyprus: Chondrichthyans. II. Int. Fisheries Symposium- 2018.
Bengil E. G. T., Bengil F., Ozaydin O. (2020). Feeding ecology and reproduction biology of Glaucostegus cemiculus (Geoffroy Saint-Hilaire 1817) from the eastern Aegean Sea. Regional Stud. Mar. Sci. 33, 100952. doi: 10.1016/j.rsma.2019.100952
Beton D., Broderick A. C., Godley B. J., Kolaç E., Ok M., Snape R. T. E. (2021). New monitoring confirms regular breeding of the Mediterranean monk seal in Northern Cyprus. Oryx 55 (4), 522–525. doi: 10.1017/S0030605320000848
Bradai M. N., Saidi B., Enajjar S. (2018). “Overview on mediterranean shark’s fisheries: impact on the biodiversity,” in Marine ecology - biotic and abiotic interactions (London, UK: IntechOpen).
Carpentieri P., Nastasi A., Sessa M., Srour A. (2021). “Incidental catch of vulnerable species in Mediterranean and Black Seafisheries – A review,” in General fisheries commission for the mediterranean. Studies and reviews. No. 101 (Rome: FAO).
Cashion M. S., Bailly N., Pauly D. (2019). Official catch data underrepresent shark and ray taxa caught in Mediterranean and Black Sea fisheries. Mar. Policy 105, 1–9. doi: 10.1016/j.marpol.2019.02.041
Çoker T., Akyol O. (2014). An overview on the fish diversity in coasts of Turkish Republic of Northern Cyprus (Mediterranean). Ege J. Fisheries Aquat. Sci. 31, 113–118. doi: 10.12714/egejfas.2014.31.2.08
Colloca F., Enea M., Ragonese S., Di Lorenzo M. (2017). A century of fishery data documenting the collapse of smooth-hounds (Mustelus spp.) in the Mediterranean Sea. Aquat. Conservation: Mar. Freshw. Ecosyst. 27 (6), 1145–1155. doi: 10.1002/aqc.2789
Davies R. W. D., Cripps S. J., Nickson A., Porter G. (2009). Defining and estimating global marine fisheries bycatch. Mar. Policy 33, 661–672. doi: 10.1016/j.marpol.2009.01.003
Dulvy N. K., Allen D. J., Ralph G. M., Walls R. H. L. (2016). The conservation status of sharks, rays and chimaeras in the mediterranean sea (Malaga, Spain: IUCN).
Dulvy N. K., Fowler S. L., Musick J. A., Cavanagh R. D., Kyne P. M., Harrison L. R., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. elife 3, e00590. doi: 10.7554/eLife.00590
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4787. doi: 10.1016/j.cub.2021.08.062
FAO. (2020). The state of world fisheries and aquaculture 2020. Sustainability in action. (Rome). doi: 10.4060/ca9229en
Fisheries Law 27-2000. (2022). Turkish republic of north Cyprus ministry of agriculture and natural resources.
Giovos I., Aga Spyridopoulou R. N., Doumpas N., Glaus K., Kleitou P., Kazlari Z., et al. (2021a). Approaching the “real“ state of elasmobranch fisheries and trade: A case study from the Mediterranean. Ocean Coast. Manag 211, 105743. doi: 10.1016/j.ocecoaman.2021.105743
Giovos I., Serena F., Katsada D., Anastasiadis A., Barash A., Charilaou C., et al. (2021b). Integrating literature, biodiversity databases, and citizen-science to reconstruct the checklist of chondrichthyans in Cyprus (Eastern mediterranean sea). Fishes 6, 24. doi: 10.3390/fishes6030024
Hadjichrisophorou M. (2006). “Chondrichthyes in Cyprus,” in The proceedings of the international workshop on mediterranean cartilaginous fish with emphasis on southern and eastern mediterranean. Eds. Basusta N., Keskin C., Serena F., Seret B. (Istanbul, Turkey: Turkish Marine Research Foundation (TUDAV).
Heupel M. R., Simpfendorfer C. A. (2002). Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can. J. Fisheries Aquat. Sci. 59, 624–632. doi: 10.1139/f02-036
IUCN. (2022). The IUCN red list of threatened species. Version 2022-2. Available at: https://www.iucnredlist.org (Accessed 01/08/2023).
Kabasakal H. (2013). Rare but present: Status of basking shark, Cetorhinus maximus (Gunneru) in eastern mediterranean. Ann. Ser. Hist. Nat. 23, 127–132.
Molina J. M., Cooke S. J. (2012). Trends in shark bycatch research: current status and research needs. Rev. Fish Biol. Fisheries 22, 719–737. doi: 10.1007/s11160-012-9269-3
Oliver S., Braccini M., Newman S. J., Harvey E. S. (2015). Global patterns in the bycatch of sharks and rays. Mar. Policy 54, 86–97. doi: 10.1016/j.marpol.2014.12.017
Oray I. K., Karakulak F. S. (2005). Investigations on the artisanal fishery of Güzelyurt Bay (Northern Cyprus).Journal of Fisheries and Aquatic Sciences. İstanbul Üniversitesi Su Ürünleri Dergisi. 19, 57–63.
Otero M., Serena F., Gerovasileiou V., Barone M., Bo M., Arcos J. M., et al. (2019). Identification guide of vulnerable species incidentally caught in Mediterranean fisheries (Malaga, Spain: IUCN).
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589, 567–571. doi: 10.1038/s41586-020-03173-9
Piovano S., Basciano G., Swimmer Y., Giacoma C. (2012). Evaluation of a bycatch reduction technology by fishermen: A case study from Sicily. Mar. Policy 36, 272–277. doi: 10.1016/j.marpol.2011.06.004
Sabri R., Sakallı B. (2021). The politics and ethical dilemmas of architectural conservation in an unrecognised state: insights from Northern Cyprus. Int. J. Heritage Stud. 27, 1245–1263. doi: 10.1080/13527258.2021.1950031
Serena F., Abella A. J., Bargnesi F., Barone M., Colloca F., Ferretti F., et al. (2020). Species diversity, taxonomy and distribution of Chondrichthyes in the Mediterranean and Black Sea. Eur. Zoological J. 87, 497–536. doi: 10.1080/24750263.2020.1805518
Snape R. T. E., Beton D., Broderick A. C., Çiçek B. A., Fuller W. J., Özden Ö., et al. (2013). Strand monitoring and anthropological surveys provide insight into marine turtle bycatch in small-scale fisheries of the eastern mediterranean. Chelonian Conserv. Biol. 12, 44–55. doi: 10.2744/CCB-1008.1
Snape R. T., Bradshaw P. J., Broderick A. C., Fuller W. J., Stokes K. L., Godley B. J. (2018a). Off-the-shelf GPS technology to inform marine protected areas for marine turtles. Biol. Conserv. 1;227, 301–309. doi: 10.1016/j.biocon.2018.09.029
Snape R. T. E., Broderick A. C., Çiçek B. A., Fuller W. J., Tregenza N., Witt M. J., et al. (2018b). Conflict between dolphins and a data-scarce fishery of the european union. Hum. Ecol. 46, 423–433. doi: 10.1007/s10745-018-9989-7
Tiralongo F., Messina G., Gatti R. C., Tibullo D., Lombardo B. M. (2018b). Some biological aspects of juveniles of the rough ray, Raja radula Delaroch in Eastern Sicily (central Mediterranean Sea). J. Sea Res. 142, 174–179. doi: 10.1016/j.seares.2018.10.001
Tiralongo F., Messina G., Lombardo B. M. (2018a). Discards of elasmobranchs in a trammel net fishery targeting cuttlefish, Sepia officinalis Linnaeus 1758, along the coast of Sicily (central Mediterranean Sea). Regional Stud. Mar. Sci. 20, 60–63. doi: 10.1016/j.rsma.2018.04.002
Ulman A., Çiçek B. A., Salihoglu I., Petrou A., Patsalidou M., Pauly D., et al. (2015). Unifying the catch data of a divided island: Cyprus’s marine fisheries catches 1950–2010. Environ. Dev. Sustain 17, 801–821. doi: 10.1007/s10668-014-9576-z
Wilson S. M., Raby G. D., Burnett N. J., Hinch S. G., Cooke S. J. (2014). Looking beyond the mortality of bycatch: sublethal effects of incidental capture on marine animals. Biol. Conserv. 171, 61–72. doi: 10.1016/j.biocon.2014.01.020
Keywords: shark, batoid, skate, ray, mediterranean, fisheries
Citation: O’Keefe M, Bengil EGT, Palmer JL, Beton D, Çağlar Ç, Godley BJ, Özkan M, Snape RTE and Broderick AC (2023) Diversity and distribution of elasmobranchs in the coastal waters of Cyprus: using bycatch data to inform management and conservation. Front. Mar. Sci. 10:1181437. doi: 10.3389/fmars.2023.1181437
Received: 07 March 2023; Accepted: 27 September 2023;
Published: 31 October 2023.
Edited by:
Fabio Fiorentino, National Research Council (CNR), ItalyReviewed by:
Fabrizio Serena, National Research Council (CNR), ItalyFrancesco Tiralongo, University of Catania, Italy
Copyright © 2023 O’Keefe, Bengil, Palmer, Beton, Çağlar, Godley, Özkan, Snape and Broderick. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Robin T. E. Snape, Ui5ULkUuU25hcGVAZXhldGVyLmFjLnVr; Annette C. Broderick, QS5DLkJyb2Rlcmlja0BleGV0ZXIuYWMudWs=
†These authors share first authorship