
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
DATA REPORT article
Front. Mar. Sci. , 21 April 2023
Sec. Discoveries
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1165207
 Austin J. Gallagher1*
Austin J. Gallagher1* Oliver N. Shipley1
Oliver N. Shipley1 Christine De Silva1Johanna K. Kohler2
Christine De Silva1Johanna K. Kohler2 Teresa F. Fernandes3
Teresa F. Fernandes3 Timothy Austin2
Timothy Austin2 Rupert F. Ormond3,4
Rupert F. Ormond3,4 Mauvis A. Gore3,4
Mauvis A. Gore3,4The genus Etmopterus is the most speciose group of small bodied deep-sea sharks found throughout the tropical and subtropical Western Atlantic. Despite exhibiting a global distribution at the genus-level, the blurred lantern shark (Etempoterus bigelowi) is known only from a few records in the Western and Southern Atlantic Ocean. Through in-situ video observations using deep-sea landers, we provide two new locality records of the blurred lantern shark from the deep waters off the Cayman Islands, Caribbean Sea. Three unique individuals were recorded across two separate deployments between 653m – 668m. These observations provide the first records of this species in the Caribbean Sea, adding to the minimal knowledge of the species’ distribution throughout the Western Atlantic Ocean.
Despite their potentially significant contribution to the global biodiversity of fishes, deep-sea sharks remain difficult to observe and study, largely due to the logistical and operational challenges of surveying the deep ocean (Howell et al., 2020). Lantern sharks (Family Etmopteridae) are a small-bodied family of squaliform sharks that are found throughout all major ocean basins and are typically distinguished by light-producing photophores distributed across their body surfaces (Claes and Mallefet, 2011; Claes and Mallefet, 2015). The family includes four extant genera, Anculeola, Centroscyllium, Trigonognathus, and Etmopterus, the latter being by far the most speciose, comprising 44 described species (Pollerspöck and Straube, 2022). Species of the genus Etmopterus are typically associated with deep-sea environments (i.e., depths exceeding 200m), and as with most species of deep-sea shark, there is an inherent scarcity of basic biological information pertaining to most aspects of their biology and distribution (e.g., Finucci et al., 2021). This largely reflects the logistical challenges of targeted surveying and the absence of individuals in accessible fish markets, due to their low commercial value (Paramo et al., 2021).
Deep-sea ecosystems of the subtropical and tropical Western Atlantic are believed to support at least eight species of Etmopterus (Mejía-Falla et al., 2019). However, most biological information pertaining to these individuals is rudimentary, reflecting morphological and catch data gathered from deep-sea trawl surveys (Springer and Burgess, 1985; Paramo et al., 2012; Paramo et al., 2021). Existing information from waters of the South American Caribbean (Colombia) revealed that catches were largely dominated by a single species, the dwarf lantern shark E. perryi (Paramo et al., 2012; Paramo et al., 2021), suggesting comparatively high abundance relative to sympatric species such as E. robinsi and E. hillianus (Schofield and Burgess, 1997; Bunkley-Williams and Williams, 2004). In comparison, existing information on Etmopterus in the Caribbean is very limited, posing questions on the spatial distribution of species composition throughout the broader regions of the Western Atlantic.
The blurred lantern shark Etmopterus bigelowi (Shirai and Tachikawa, 1993) is patchily distributed through insular shelves and slopes of the tropical and subtropical Atlantic, Pacific, and Indian Oceans (Finucci et al., 2020a). Individuals are typically associated with pelagic and benthic deep waters, having been recorded between depths of approximately 100m-1000m (Ebert et al., 2013; Finucci et al., 2020a). Individuals have been found to reach a maximum total length (TL) of at least 73cm; size (TL) at sexual maturity is 40cm - 67cm for males and 50cm – 65cm for females (Ebert et al., 2013). Though populations of E. bigelowi are expected to be relatively stable, recent summaries outline a scarcity of information about any aspect of the species’ biology (Finucci et al., 2020a). Here, we report the first records of the blurred lantern shark from deep waters adjacent to Grand Cayman, Caribbean Sea, Western Atlantic Ocean, via deployment of deep-sea landers equipped with video cameras. This information provides the first known record of this species in the Caribbean Sea, with previous records only establishing a localized range within South America and the southern United States.
The Cayman Islands (Grand Cayman, Cayman Brac, and Little Cayman) are situated in the central Caribbean Sea adjacent to the most central portion of Cayman Trench. The region houses a suite of unique oceanographic and geological characteristics due to the proximity of the Oriente transform fault and Cayman Rise, which promote high levels of tectonic activity and hydrothermal vent fields in relatively shallow depths of less than 2,000m (Jones, 1994). Despite the unique oceanographic and marine geological conditions in the region, general trends in biodiversity, especially those related to deep-sea habitats, are poorly described, with the Cayman Islands receiving significantly less attention than similar regions such as the Lucayan Archipelago, Greater Antilles, and Jamaica (Burgess et al., 1994; McLaughlin and Morrissey, 2004; McLaughlin and Morrissey, 2005; Brooks et al., 2015). Further, there have been no systematic survey attempts to quantify patterns of deep-sea biodiversity around the Cayman Islands, despite the potential for housing a suite of unique ecosystems.
As part of a multi-year study to survey the biodiversity of sharks and fishes in the deep-sea territorial waters of the Cayman Islands, we conducted benthic video surveys between March and October 2022 off the shelf in deep waters approximately 3km – 15km offshore spanning the perimeter of Grand Cayman. Surveys were conducted using a custom free-falling, deep baited remote underwater lander system (hereafter dBRUV, Phillips et al., 2019).
The dBRUV system (Figure 1) consisted of a vertically oriented, carbon-fiber frame with pressure-tolerant flotation (G2200, McLane Research Laboratories Inc., East Falmouth, MA) and an acoustic weight-release system (CAT, EdgeTech, Massachusetts, US). This system was equipped with a depth-rated radio transmitter (F1845B, Advanced Telemetry Systems, Minnesota, USA) and an orange identification flag to facilitate recovery at the surface. A single GoPro camera (GoPro Hero 5/6/7), set to record 1080p video at 60 frames per second, was secured within a deep-water housing (GoBenthic, GroupB Incorporated, USA) and attached to the frame ~1.5m above the bottom of the unit. Two LED lights were used to illuminate the seafloor (SiteLite, Juice Robotics, Rhode Island, USA), and the cameras and lights powered with a custom lithium-ion battery pack. Temperature and depth were monitored using a calibrated Starmon TD stand-alone logger (Star Oddi, Iceland) or LAT1400 temperature and depth loggers (LOTEK). Approximately 500g of sardines were attached to a pole-mounted bait cage in front of the camera. The acoustic release system was used to release the drop weight, allowing for the entire system to return to the surface upon command, resulting in deployments of 6-7 hours. All dBRUVs were then located and retrieved at the surface using boat-based GPS unit, YAGI antenna, and VHF radio receiver (R410, Advanced Telemetry Systems, Minnesota, USA). Upon retrieval of the dBRUVs, videos were downloaded from micro-SD cards and reviewed at 0.5x regular speed. All research was conducted under scientific collection permits acquired from the Cayman Islands Department of Environment (DOE).

Figure 1 Diagram of the deep-sea BRUV (dBRUV) video camera/lander system used in the present study, with its associated components.
On July 26th 2022 a dBRUV was deployed 3km off Southwest Point, Grand Cayman (19.251°N, -81.416°W) at a depth of 653m, which recorded 4.5 hours of video between 10:00 and 16:00. A single male E. bigelowi was observed on a single video file and interacted with the bait crate for approximately one minute (Figure 2A). On September 19th, 2022, a dBRUV unit was deployed 15 km south of Breakers Bay, Grand Cayman (19.151°N, -81.191°W) recording 4.5 hours of video at a depth of 668m between 12:00 and 17:00. During the deployment, a single male (Figure 2B) and female (Figure 2C) E. bigelowi were observed and interacted with the bait crate for several minutes. Though positive identification of lantern shark species can be inherently challenging (Schofield and Burgess, 1997), the position of the first dorsal fin and snout length distinguishes this species from the sympatric E. pusillus (D. Ebert Pers. Comms). These behavioral observations suggest patterns of diel vertical migration in lantern sharks, as seen in other mesopelagic fishes (Abramov and Lipskaya, 1989). As our study suggests, lantern sharks occurring in the Caribbean Sea are likely to be recorded in-between periods of feeding in shallower waters during the night and resting at depth during the day, as documented elsewhere (e.g., Paramo et al., 2021)

Figure 2 In-situ images of Etmopterus bigelowi from deep waters of the Cayman Islands collected using deep-sea video landers (Phillips et al., 2019). One male individual was observed on July 26th 2022 (A) followed by observations of a male (B) and female (C) on September 19th 2022. The individuals made several passes across the field of view and were observed interacting with the bait cage.
These observations support other known localities for E. bigelowi from the central Western Atlantic, and represents the first record of this species from the Caribbean Sea (Table 1). Historical distributions of E. bigelowi in Western Atlantic are known from several observations in South America, including Panama, Brazil, Argentina, Suriname, and Uruguay (Table 1), and by a single observation from the USA (Froese and Pauly, 2000). Combined, this suggests that the species displays a moderate latitudinal distribution throughout the tropical and subtropical Western Atlantic, similar to other small and medium-bodied deep-sea sharks such as the Cuban dogfish (Squalus cubensis, Cotton et al., 2020) and gulper sharks (Centrophorus spp., Finucci et al., 2020b). However, given the very low number of locality records for E. bigelowi, it is possible that the species’ true latitudinal range remains significantly underestimated.
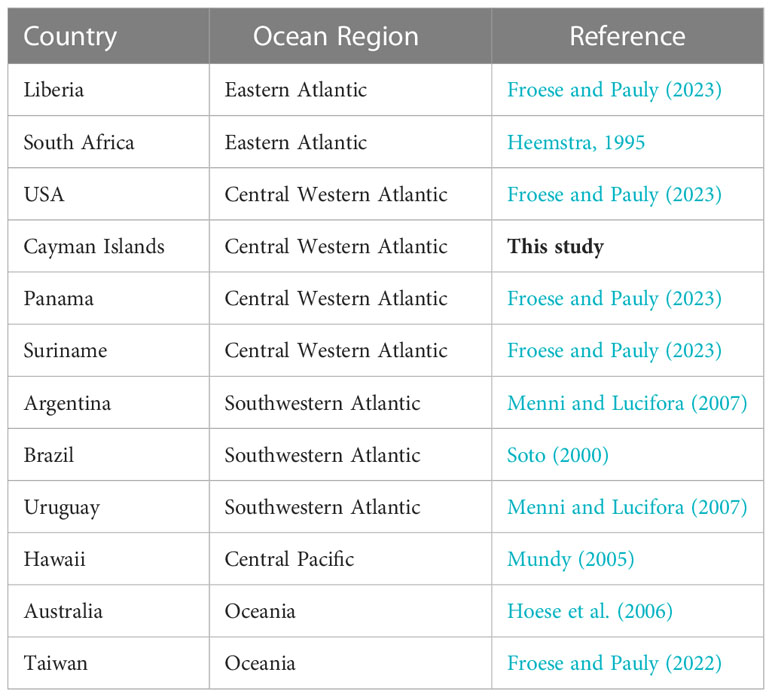
Table 1 Known localities of Etmopterus bigelowi throughout Atlantic, Pacific, and Indian Oceans.
While the waters of the Cayman Islands are known to support a relatively small, yet diverse population of sharks in its neritic waters (Gore et al., 2020), these new records suggest that its deep waters may have been an overlooked regional hotspot for the biodiversity of deep-sea sharks. As the Cayman Islands have designated 17.8% of their shelf area as Marine Protected Area (MPA) to manage the well-developed and economically important fringing coral reefs, mangrove stands and seagrass beds (Dromard et al., 2011), these MPAs extend only 1-2km offshore. However, the seabed extends to 2000m within Cayman’s territorial waters (12 nm), and to over 6000m within Cayman’s Exclusive Economic Zone, suggesting that deep-reef habitats may warrant inclusion in future plans for protected area expansion, especially given growing threats of unsustainable, unregulated fishing of deep-water species and of sea-bed mining for metalliferous nodules adjacent to ocean trenches (Irigoien et al., 2014; Churchyard et al., 2016).
These new locality records also highlight a broader need to determine patterns of deep-sea biodiversity throughout the Greater Caribbean (Howell et al., 2021). This is particularly pertinent, given recent calls to scale the size and volume of regional MPAs to facilitate effective conservation of biodiversity (Gallagher et al., 2020). This requires improved efforts to increase region-wide monitoring that will ensure the integration of deep-sea habitats into future MPA expansion (Paramo et al., 2012). To be successful, scaling efforts will require deliberate, international collaboration and improved mechanisms for support and empowerment of many developing countries (Howell et al., 2020; Amon et al., 2022). While providing the critical information required to determine hotspots of deep-sea biodiversity, these efforts are also likely to yield exciting new discoveries across the Greater Caribbean.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by Heriot-Watt University.
AG, TF, TA and MG led and administered the study. AG, RO, TA and MG conceived the study. All authors performed the field work. OS and JK analyzed the data. OS and AG wrote the first draft of the paper. All authors provided edits on the paper. All authors contributed to the article and approved the submitted version.
This work was funded through a Darwin Plus Grant (DPLUS140), part of the Darwin Initiative, Overseas Territories Environment and Climate Fund, Department of Environment Food and Rural Affairs of the UK Government, with support from the Department of Environment (DOE) of the Cayman Island Government.
We thank staff from the DOE for their assistance with field work and David Ebert for providing key insight on lantern fish biology and positive identification of E. bigelowi.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abramov A., Lipskaya H. (1989). On the feeding of epigonus elegans in the region of the nazca ridge. J. Ichthyology 29, 129–134.
Amon D. J., Rotjan R. D., Kennedy B. R., Alleng G., Anta R., Aram E., et al. (2022). My deep Sea, my backyard: a pilot study to build capacity for global deep-ocean exploration and research. Philos. Trans. R. Soc. B 377 (1854), 20210121. doi: 10.1098/rstb.2021.0121
Brooks E. J., Brooks A. M., Williams S., Jordan L. K., Abercrombie D., Chapman D. D., et al. (2015). First description of deep-water elasmobranch assemblages in the exuma sound, the Bahamas. Deep Sea Res. Part II: Topical Stud. Oceanography 115, 81–91.
Bunkley-Williams L., Williams E. H. (2004). New locality, depth, and size records and species character modifications of some Caribbean deep-reef/shallow slope fishes and a new host and locality record for the chimaera cestodarian. Caribbean J. Sci. 40 (1), 88–119.
Burgess G. H., Smith S. H., Lane E. D. (1994). “11. fishes of the Cayman islands,” in The Cayman islands: natural history and biogeography, (Dordrecht: Kluwer Academic Publishers) vol. 71. , 199.
Churchyard T., Eaton M. A., Havery S., Hall J., Millett J., Farr A., et al. (2016). The biodiversity of the united kingdom’s overseas territories: a stock take of species occurrence and assessment of key knowledge gaps. Biodiversity Conserv. 25, 1677–1694. doi: 10.1007/s10531-016-1149-z
Claes J. M., Mallefet J. (2011). Control of luminescence from lantern shark (Etmopterus spinax) photophores. Communicative Integr. Biol. 4 (3), 251–253. doi: 10.4161/cib.4.3.14888
Claes J. M., Mallefet J. (2015). Comparative control of luminescence in sharks: new insights from the slendertail lanternshark (Etmopterus molleri). J. Exp. Mar. Biol. Ecol. 467, 87–94. doi: 10.1016/j.jembe.2015.03.008
Cotton C. F., Derrick D., Herman K., Pacoureau N., Dulvy N. K. (2020). Squalus cubensis. The IUCN red list of threatened species 2020. doi: 10.2305/IUCN.UK.2020-3.RLTS.T61416A3104105.en
Dromard C. R., Mccoy C. M., Turner J. R. (2011). “Measuring the performance of marine protected areas: the case of little Cayman and Cayman brac, Cayman islands,” in Proceedings of the 63rd Gulf and Caribbean Fisheries Institute, San Juan, Puerto Rico. 246–253.
Finucci B., Bineesh K. K., Cheok J., Cotton C. F., Dharmadi D., Kulka D. W., et al. (2020b). Centrophorus granulosus. The IUCN red list of threatened species 2020. doi: 10.2305/IUCN.UK.2020-3.RLTS.T162293947A2897883.en
Finucci B., Cheok J., Cotton C. F., Kulka D. W., Neat F. C., Rigby C. L., et al. (2020a). Etmopterus bigelowi. The IUCN red list of threatened species 2020. doi: 10.2305/IUCN.UK.2020-3.RLTS.T60236A3096618.en
Finucci B., Cheok J., Ebert D. A., Herman K., Kyne P. M., Dulvy N. K. (2021). Ghosts of the deep–biodiversity, fisheries, and extinction risk of ghost sharks. Fish Fisheries 22 (2), 391–412. doi: 10.1111/faf.12526
Gallagher A. J., Amon D. J., Bervoets T., Shipley O. N., Hammerschlag N., Sims D. W. (2020). The Caribbean needs big marine protected areas. Science 367 (6479), 749–749.
Gore M., Ormond R., Clarke C., Kohler J., Millar C., Brooks E. (2020). Application of photo-identification and lengthened deployment periods to baited remote underwater video stations (BRUVS) abundance estimates of coral reef sharks. Oceans 1 (4), 274–299). doi: 10.3390/oceans1040019
Heemstra P. C. (1995). “Additions and corrections for the 1995 impression,” in Revised edition of smiths' Sea fishes. Eds. Smith M. M., Heemstra P. C. (Berlin: Springer-Verlag). p. v-xv.
Hoese D. F., Bray D. J., Paxton J. R., Allen G. R. (2006). “Fishes,” in Zoological catalogue of Australia, vol. Volume 35 . Eds. Beasley O. L., Wells A. (Australia: ABRS & CSIRO Publishing), 2178.
Howell K. L., Hilário A., Allcock A. L., Bailey D. M., Baker M., Clark M. R., et al. (2020). A blueprint for an inclusive, global deep-sea ocean decade field program. Front. Mar. Sci. 999. doi: 10.3389/fmars.2020.584861
Howell K. L., Hilário A., Allcock A. L., Bailey D., Baker M., Clark M. R., et al. (2021). A decade to study deep-sea life. Nat. Ecol. Evol. 5 (3), 265–267.
Irigoien X., Klevjer T. A., Røstad A., Martinez U., Boyra G., Acuña J. L., et al. (2014). Large Mesopelagic fishes biomass and trophic efficiency in the open ocean. Nat. Commun. 5 (1), 3271. doi: 10.1038/ncomms4271
Jones B. (1994). “Geology of the Cayman islands,” in The Cayman islands. monographiae biologicae, vol. vol 71 . Eds. Brunt M. A., Davies J. E. (Dordrecht: Springer). doi: 10.1007/978-94-011-0904-8_2
McLaughlin D. M., Morrissey J. F. (2004). New records of elasmobranchs from the Cayman trench, Jamaica. Bull. Mar. Sci. 75 (3), 481–485.
McLaughlin D. M., Morrissey J. F. (2005). Reproductive biology of centrophorus cf. uyato from the Cayman trench, Jamaica. J. Mar. Biol. Assoc. United Kingdom 85 (5), 1185–1192.
Mejía-Falla P. A., Navia A. F., Salcedo-Reyes J. C. (2019). Checklist of marine elasmobranchs of Colombia. Universitas Scientiarum 24 (1), 164–199.
Menni R. C., Lucifora L. O. (2007). Condrictios de la Argentina y Uruguay. ProBiota FCNyM UNLP Serie Técnica-Didáctica La Plata Argent. 11, 1–15.
Mundy B. C. (2005). Checklist of the fishes of the Hawaiian archipelago. Bishop Mus. Bull. Zool. 6), 1–704.
Paramo J., Wiff R., González R. (2021). A matter of size: the population structure of the smallest known living shark, Etmopterus perryi (Springer & burgess 1985), from deep waters off the Colombian Caribbean coast. J. Fish Biol. 99 (3), 755–764. doi: 10.1111/jfb.14755
Paramo J., Wolff M., Saint-Paul U. (2012). Deep-sea fish assemblages in the Colombian Caribbean Sea. Fisheries Res. 125, 87–98. doi: 10.1016/j.fishres.2012.02.011
Phillips B. T., Shipley O. N., Halvorson J., Sternlicht J. K., Gallagher A. J. (2019). First in situ observations of the sharpnose sevengill shark (Heptranchias perlo), from the tongue of the ocean, Bahamas. J. Ocean Sci. Foundation 32, 17–22.
Pollerspöck J., Straube N. (2022) Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichthyes: elasmobranchii, holocephali). Available at: https://shark-references.com/ (Accessed 23 Mar 2023).
Froese R., Pauly D. (Eds.) (2000). FishBase 2000: concepts, design and data sources (Los Baños, Laguna, Philippines: ICLARM), 344.
Froese R., Pauly D. (Eds.) (2022). FishBase. version (06/2022). Available at: https://www.fishbase.org.
Froese R., Pauly D. (Eds.) (2023). FishBase. version (06/2023). Available at: https://www.fishbase.org.
Schofield P. J., Burgess G. H. (1997). Etmopterus robinsi (Elasmobranchii, etmopteridae), a new species of deepwater lantern shark from the Caribbean Sea and western north Atlantic, with a redescription of Etmopterus hillianus. Bull. Mar. Sci. 60 (3), 1060–1073.
Shirai S., Tachikawa H. (1993). Taxonomic resolution of the Etmopterus pusillus species group (Elasmobranchii, etmopteridae), with description of E. bigelowi, n. sp. Copeia, 483–495. doi: 10.2307/1447149
Soto J. M. R. (2000). Revisão da ocorrência de tubarões etmopteríneos na costa brasileira, com dois novos registros de Etmopterus bigelowi shirai & tachikawa 1993 (Chondrichthyes, datatiidae, etmopterinae). Biotemas 13 (2), 167–178.
Keywords: biodiversity, Caribbean, deep-sea, lantern shark, lander, locality, shark
Citation: Gallagher AJ, Shipley ON, De Silva C, Kohler JK, Fernandes TF, Austin T, Ormond RF and Gore MA (2023) First records of the blurred lantern shark Etmopterus bigelowi from the Cayman Islands, Western Atlantic. Front. Mar. Sci. 10:1165207. doi: 10.3389/fmars.2023.1165207
Received: 13 February 2023; Accepted: 03 April 2023;
Published: 21 April 2023.
Edited by:
Carlotta Mazzoldi, University of Padua, ItalyReviewed by:
Emilio Sperone, University of Calabria, ItalyCopyright © 2023 Gallagher, Shipley, De Silva, Kohler, Fernandes, Austin, Ormond and Gore. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Austin J. Gallagher, YXVzdGluQGJlbmVhdGh0aGV3YXZlcy5vcmc=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.