 Signe Gunborg Bentzrød Svensson1,2,4*
Signe Gunborg Bentzrød Svensson1,2,4* Tore Strohmeier1Helen Rastrick1†Antonio Aguera Garcia1†Erik-Jan Lock1‡Harald Sveier2‡Henrice M. Jansen3
Tore Strohmeier1Helen Rastrick1†Antonio Aguera Garcia1†Erik-Jan Lock1‡Harald Sveier2‡Henrice M. Jansen3- 1Institute of Marine Research, Bergen, Norway
- 2Lerøy Seafood Group, Bergen, Norway
- 3Aquaculture and Fisheries Group, Wageningen University and Research, Wageningen, Netherlands
- 4Department of Biological Sciences, Bergen University, Bergen, Norway
Ophryotrocha craigsmithi (Wiklund, Glover & Dahlgren, 2009) has repeatedly been observed in high densities at salmonid fish farms in Norway and has been shown to feed on the organic waste released during fish production. This species has therefore been pointed out as a candidate for the benthic component of integrated multitrophic aquaculture (IMTA). Little is known about the life history traits of Ophryotrocha species found at greater depths in temperate waters. Insight into life traits is of key importance in evaluating the bio-mitigation and harvest potential of (benthic) species within IMTA systems. In this study, we aimed to describe key life history traits for O. craigsmithi, such as growth, reproductive pattern, and offset of reproduction. The main findings are as follows: (I) O. craigsmithi exhibits early sexual maturation and rapid growth, going from 5 to 16 mm in length in 8 weeks; (II) their estimated lifetime is approximately 15 weeks with a generation time of 4 weeks; (III) there are strong indications that they are gonochoric semicontinuous spawners; (IV) the majority of mature eggs ranged from 120 to 150 µm; and (V) the larvae developed into a free-swimming stage after approximately 1 week. To this end, we present a framework for assessing the suitability of deposit-feeding polychaetes as candidate species in IMTA, considering environmental adaptiveness, production potential, as well as carbon and nitrogen mitigation capacity. On the basis of this, O. craigsmithi shows a high suitability as an extractive farmed species in benthic IMTA.
1 Introduction
In open-water salmonoid farming, a substantial amount of particulate organic waste, consisting of uneaten fish feed and feces, generally settles in the close vicinity of fish farms (Strain and Hargrave, 2005; Broch et al., 2017) and is of ecological concern for the benthic ecosystems (Kutti et al., 2007b; Keeley et al., 2020). For utilizing these waste streams, integrated multitrophic aquaculture (IMTA) offers a strategy for improving system sustainability by taking a circular approach. In IMTA, waste streams from fed species are used to cultivate other extractive species occupying lower trophic levels. This enables diversification and higher utilization of inputs to fish farming while, at the same time, reducing environmental pressure (Troell et al., 2009; Chopin et al., 2012). The capacity of bivalves to mitigate organic particulate waste in open-water IMTA has been questioned (Cranford et al., 2013), and the focus has now shifted toward the benthic part of IMTA. The high amounts of organic waste and its nutritional composition offer an opportunity for utilizing this, today, neglected resource in open-cage farming systems by developing benthic IMTA integrating benthic deposit feeders (Wang et al., 2012; Cranford et al., 2013; Filgueira et al., 2017; Jansen et al., 2019).
A variety of polychaetes are identified as candidates for bioremediation of organic fish waste (Tsutsumi et al., 1990; Fang et al., 2017; Marques et al., 2017). Within the polychaetae genus Ophryotrocha, there have been several reports of these species in high densities under commercial fish farms. In Canada, O. cyclops is frequently observed in polychaete complexes covering the epibenthos under fish farms (Salvo et al., 2014; Armstrong et al., 2020); and in Tasmania, O. shieldsi has been estimated in significant densities under fish farms, up to 100,000 individuals per square meter (Paxton and Davey, 2010). In Norwegian waters, O. craigsmithi has recently gained attention, as it has been frequently observed to cover and dominate the benthic fauna under fish farms, especially in areas dominated by hard bottom. This has led to the hypothesis that these have a competitive advantage on organically enriched hard substrates (Hansen et al., 2011; Eikje, 2013; Jansen et al., 2019). The hypothesized advantage of O. craigsmithi on hard bottom has led to the idea to develop harvesting structures that offer artificial hard bottom to facilitate the establishment of high Ophryotrocha sp. populations (Jansen et al., 2019). The first large-scale harvest structure is under development by a Norwegian salmon company, and the first pilot harvests have been performed on commercial fish farms (unpublished data).
Furthermore, it has been estimated that, a mixed polychaetae community consisting of Vigtorinella spp. and Ophryotrocha spp. with densities between 8,000 and 71,000 individuals per square meter can be supported under a commercial Norwegian salmon farm (Eikje, 2013). In addition to Ophryotrochas sp. high densities at fish farms, they have shown high carbon turnover rates and favorable protein and fatty acid profiles when fed with fish waste (Nederlof et al., 2019; Nederlof et al., 2020). Their nutritional value makes them attractive for the aquaculture feed industry as an ingredient in future feed formulations (Nederlof et al., 2019). The demand for high-quality feed ingredients is increasing as the industry grow, and the need for these ingredients to be sustainably sourced is crucial for further expansion of the industry (Albrektsen et al., 2022). Combining the traits described above has led to the suggestion of farming O. craigsmithi in benthic IMTA systems (Fang et al., 2018; Jansen et al., 2019; Nederlof et al., 2020).
Today, we lack information on population dynamics for O. craigsmithi. Information regarding growth and reproduction is a key element in understanding population dynamics, and such life history traits are essential when assessing bioremediation potentials for specific species in IMTA (Ramskov and Forbes, 2008; Galasso et al., 2020). Knowledge on growth and reproduction is further valuable for the establishment of future harvesting strategies of this species in IMTA. Ophryotrocha is a well-studied genus with several Mediterranean, shallow, and warm water species held in laboratory cultures (Thornhill et al., 2009). In general, the genus is characterized by opportunistic small worms (< 3 mm in body length) with short life cycles, but the genus shows great variation in their reproductive strategies with species displaying both sequential and simultaneously hermaphroditism as well as gonochoric species (Dahlgren et al., 2001; Kvalø Heggøy et al., 2007). Several Ophryotrocha species are described as sequential spawners laying multiple batches of eggs after reaching sexual maturity, and the number of eggs at each spawning and the number of spawning events vary both within and between species (Cassai and Prevedelli, 1999; Thornhill et al., 2009). Norwegian salmon farms are often located over great depths (50–300 m) with a stable temperature of approximately 8°C (Kutti et al., 2007a), representing a very different environment from where the majority of information on Ophryotrocha spp. comes from. Life history has been described. Therefore, knowledge regarding the life cycles of Ophryotrocha species in temperate waters is limited, and, to the best of our knowledge, only one deep-sea species has successfully been fully reared under laboratory conditions (Mercier et al., 2014). O. craigsmithi was first identified and described from a whale carcass on the Swedish west coast in 2009 (Wiklund et al., 2009). Growth rates for this species exist for adult members of the population (Nederlof et al., 2019), but measurements from juvenile to adult stages are lacking. Knowledge regarding reproduction mode and larvae development is also unknown. Because of the limited knowledge for life history traits on Ophryotrocha spp. found at greater depths and in temperate waters, it is important to describe these traits for O. craigsmithi to enable further assessment of this species bioremediation and harvest potential in IMTA.
The objective of this study is to gain knowledge on key life history traits for O. craigsmithi, which are of importance for understanding the population dynamics, nutrient bioremediation, and harvest potential of this species in the context of IMTA farming. By this means, we (I) determined growth rates from juveniles to adults and (II) characterized their reproduction in terms of sexual differentiation, fecundity, and age at onset of reproduction. Finally, we present a framework that evaluates the suitability of deposit feeding polychaetes as extractive components in IMTA. In addition to the key life history traits as described in our study, this framework considers environmental adaptiveness, production and market potential, as well as nutrient mitigation capacity.
2 Materials and methods
2.1 Field collection
Field collection of polychaetes took place at a salmonid fish farm in Hjeltefjorden (60°30′37.0″N 4°57′10.9″E) situated on the west coast of Norway in December 2021. For the collection, we used perforated metal trays adapted from Jansen et al. (2019). These trays were equipped with boxes with lids (18 × 26 × 8 cm, n = 4 boxes per tray). Eight trays were attached to ropes and gently submerged and positioned on the seafloor at depths ranging between 100 and 160 m. Ropes were then attached directly on to the fish cages and left for 4 and 10 weeks. On the day of sampling, the trays were lifted of the sea floor, and lids gently sealed the boxes, reducing the possibility of losing the content of the boxes. Upon retrieval, water quality in the boxes was measured. Temperature of ~8.5°C and salinity of ~34.7 within the boxes confirmed that these had been closed upon retrieval from the seafloor. The boxes were selected on the basis of visual confirmation of successful collection of polychaetes. Boxes were removed from the frames, and, to replace any water that was lost during collection of trays, fresh deep water was added in boxes, kept cool, and transported to the laboratory within 2.5 h after collection.
2.2 Laboratory holding conditions
In the laboratory facility, the boxes were placed into three 40-L flumes provided with a continuous flow (250–400 ml min−1) of unfiltered deep-sea water (8.9°C ± 0.4°C, 34.9–35.1). Flumes were left in the dark. Red light was used during visual inspection of flumes. Given the collection method, the boxes contained complexes consisting of a variety of polychaetes together with organic fish farm waste that had accumulated during the deployment period. Visual observations of boxes showed a clear dominance of O. craigsmithi. Throughout this study, polychaetes were fed ad libitum with salmon fish pellets (3–7 mm; 40%–47% protein and 24%–32% fat) to simulate fish farm conditions, where food is not assumed to be a limiting factor for growth and reproduction. Before the initiation of the main experiments, polychaetes were allowed to acclimatize for a minimum of 8 weeks. During this time period, pilot studies were performed to establish experimental protocols.
2.3 Length-weight conversions
These polychaetes are mobile and normally contract during disturbance or movement. Thus, these require sedation to obtain actual length measurements. Growth measurements (see Section 2.4) were collected on the basis of non-sedated individuals as multiple measurements in time were collected for the same population and interference was thereby minimized. Length measurements for individuals used for fecundity estimates (see Section 2.5) were defined on the basis of sedated individuals. To allow for conversion between sedated and unsedated polychaete body length, a separate batch of individual polychaetes (n = 158, smaller polychaetes< 7 mm represented by pooled samples) was sampled from holding flumes and photographed in unsedated state prior to being sedated using isotonic magnesium chloride (75 g of MgCl hexahydrate per liter of purified fresh water) and photographed again. Subsequently, these were dried at 60°C until constant dry weight (~24–48 h) and combusted at 450°C for 6 h to obtain ash-free dry weights (AFDWs; in milligrams). State linear regression was used to define the sedated to unsedated length conversions, and a power function was used to describe the relationship between polychaete length and weight.
Polychaete length is always presented as non-sedated length in millimeters, and conversions by means of linear regression (see Section 2.3) were used when measurements were performed on sedated animals.
2.4 Growth and survival
From holding flumes, a small subset of individuals was selected and measured during an 8-week-long growth trial. Polychaetes were sorted according to total body length measured with the use of millimeter paper from the anterior to last segment posteriorly. Individuals were sorted in three size groups: large = 13.3 ± 0.6 mm, medium = 7.9 ± 0.7 mm, and small = 4.8 ± 0.5 mm (mean ± SD). The smallest size group was set to the given size due to practical limitations for working with smaller individuals. The largest size group represented the largest individuals sampled in the maintained population. To ensure correct species collection, the morphological traits described by Wiklund et al. (2009) were used during sorting. Groups of polychaetes were kept isolated in 8.5-L flow-through chambers (n = 50 individuals per chamber; n = 3 chambers per size group) with a flow rate of 23 ± 0.3 ml min−1 and temperature of 8.3°C ± 0.3°C. Polychaetes were fed in excess with salmon feed (4.5 mm; 43%–46% protein and 25%–28% fat) twice a week with ~105 mg per feeding event. This was to simulate fish farm conditions, where food is not assumed to limit growth of polychaetes. On a weekly basis, images of all chambers were collected using a SLR camera (Canon EOS 600D, EFS 18-55 mm). To obtain a high-quality image of all individuals at once, polychaetes were gently directed toward the bottom of each chamber, and, when normal behavior was resumed, after being disturbed, images were collected. Images were taken with a scale (millimeter paper placed on the bottom of each chamber), and images were later imported, scaled, and analyzed with ImageJ software Java 1.8.0191 (Schneider et al., 2012) to measure polychaete total body length. To obtain survival data, all individuals observed on images were counted on a weekly basis. The method of using images reduced handling of polychaetes to a minimum.
Growth rates were calculated as length-specific growth rates (LSGRs) by the equation given below:
where Lt is the total body length (in millimeters) at time of measurement, Li is the initial total body length (in millimeters), and T is the number of days passed since the first measurement.
2.5 Fecundity
Fecundity in this experiment is defined as the number of mature eggs spawned per spawning and individual; hence, we are not including immature eggs to consider potential fecundity or following specific individuals to estimate lifetime fecundity (total number of eggs laid throughout a lifetime). Two separate experiments were performed because isolated individuals did not spawn naturally but were observed to spawn in larger groups. First, the size variation, measured as egg diameter, of naturally spawned eggs was investigated to define a size range for mature eggs. Second, the number of mature eggs was counted and measured after induced spawning by heat-shock treatment, for an estimate of individual fecundity.
2.5.1 Definition of the size for mature eggs
To obtain naturally spawned eggs, polychaetes were sorted out in three size groups (n = 3 per group; n = 20 per chamber) represented by the sizes of 15.8 ± 0.4 mm (large), 8.1 ± 0.6 mm (medium) and 5.1 ± 0.2 (small) in length (mean ± SD). Spawning chambers consisted of 0.8-L containers provided with a flow rate of 11 ± 2 ml min−1 and temperature of 8.3°C ± 0.2°C. Every morning, containers were visually inspected for eggs. Spawned eggs were collected and put on ice. Each egg sample was sub-sampled (n = 3) for eggs with the aim of collecting a minimum of 10 eggs per sub-sample. Eggs were fixed on a microscopic slide with Metcel (Methyl cellulose, M7027) and photographed (Canon DS126201). Images were analyzed in ImageJ software where images were scaled using a microscale slide. From images, we measured egg diameter and categorized the first stages of egg development (Figure 1). The remaining eggs from each sample were diluted in filtered seawater (0.2 µm) and measured by a laser particle counter (PAMAS S4031GO) in triplicates. Every evening, chambers were cleaned from leftover food and eggs before provided with a fresh feed pellet. This trial was done over the course of 7 days.
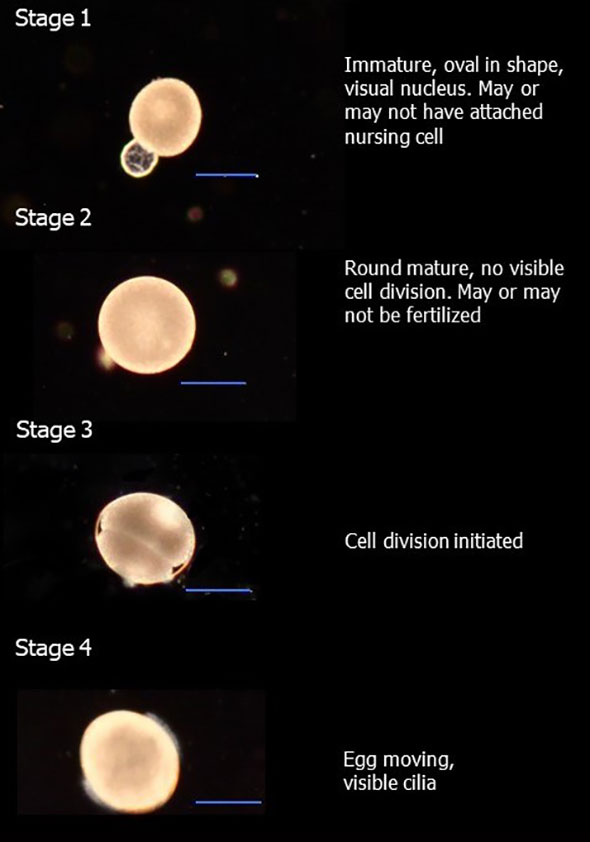
Figure 1 Developmental stages in eggs used for categorization during natural spawning’s. Scale bars represent 100 µm.
2.5.2 Individual fecundity
Prior to choosing the heat-shock method, pilot studies were performed and evaluated mechanical opening of individuals, isolation of separate animals/pairs, and heat-shock treatment at different temperature elevations. The final protocol (see below) provided predictable spawning events and comparison of manual counting with PAMAS measurements ensured satisfying precision between the two methods.
Polychaetes were collected from holding tanks on the basis of the total body length, and females were then identified on the basis of coloration and visual eggs through the body wall. Each individual was placed in a separate well in a well plate together with 10 ml of filtered (0.2 µm) sea water. The sea water temperature was 3.5°C higher compared with that in the holding-flume conditions. Plates were left in room temperature (19.1°C ± 0.9°C) for 2–2.5 h. This time was sufficient for polychaetes to spawn without compromising egg quality, because it was observed that eggs left for more than 3 h could become fragile and easily break during handling. After spawning, polychaetes were gently removed from each well, and eggs were put on ice, whereas polychaetes were placed in a fridge until further handling (2°C). Each egg sample was diluted with 50 ml of filtered (0.2 µm) sea water. A magnet stirrer provided gentle stirring, to suspend the eggs, during measurements by the PAMAS, as described in Section 2.5.1. Afterward, each individual was sedated in isotonic MgCl (see Section 2.3), and the total body length was measured. Subsequently, individuals were dissected and checked if spawning had been successful. For smaller individuals where sex is hard to distinguish, dissected polychaetes were checked under microscope at ×40 magnification. On the basis of results derived from Section 2.5.1, counting of eggs started at egg diameter > 120 µm.
2.6 Histology
Histology was used to determine the onset of gamete production in O. craigsmithi. Polychaetes (n = 30) were collected on the basis of their total body length and included the range of 4–16 mm from the stock population. Differentiation between assumed sexes was based on coloration, visible eggs through the body, and the width of the back-part, as it was observed that males appeared to be broader toward the end compared with females (personal observation, see Figure 2). Individuals were sedated with isotonic MgCl solution, measured, and washed in filtered seawater before being fixed in Davidsons fix with 5% acetic acid for 24–48 h. Samples were dehydrated and casted in paraffin (Histowax, 60°C–63°C) before being sliced (3 µm) with a microtome (Thermo Scientific, HM355S), stained with Instant hematoxylin (Thermo Scientific), and photographed with NanoZoomer S60 Digital (HAMAMATSU). Images were analyzed in NDP.view2 U12388-01 (HAMAMATSU).

Figure 2 Male polychaete of ~17 mm in length (upper) and female polychaete of ~13 mm in length (lower).
2.7 Courtship behavior and larvae development
On a few occasions, spawned eggs were collected and isolated in cups or well plates (20–100 ml) before placed in a climate room of ~10°C, without aeration. These were checked intermittently to record new developmental stages. Courtship behavior was observed in the holding tanks on multiple occasions and caught on video.
2.8 Statistics
Statistics were computed using Rstudio (Rstudio, Team 2022). All data are presented as means ± SD unless stated otherwise. To describe the weight–to–total body length relationship, a non-linear least squares model was used to estimate the power-parameter b and the coefficient a by the equation: weight ~a × (sedated length (in millimeters)b). Growth data were used in a von Bertalanffy growth model to estimate the following parameters: the asymptotic length (Li) and the intrinsic growth rate (rb) of the population by the equation L ~ Li − (Li − L0) × exp(−rb × (time)). von Bertalanffy parameters were used to calculate the approximate age of the polychaetes at the start size of the different size groups using the following von Bertalanffy formulation: t(time) = log((Li − L0)/Li − L))/rb, where time is in days since reaching L0, L0 is the mean initial size of the small groups of O. craigsmithi, and L is the mean size at measuring event for the other measurements event and groups (n = 3) during the growth trial. To investigate the potential correlation between egg size and total polychaete body length, a linear correlation test was used with p< 0.05 set as the significance level.
3 Results
3.1 Relationship between polychaetae length and weight
The relationship between unsedated and sedated length of polychaetes is given by the equation y = 0.6024x + 1.1314 (R2 = 0.85, p = 2.2*10-16 with x = unsedated length (in millimeters) and y = sedated length of polychaete (in millimeters). The parameters provided by the non-linear least squares model were a = 0.0092, p = 0.0236, and b = 2.3038 (p =<2 × 10−16), resulting in the following equation for describing the weight to sedated polychaete length relationship: AFDW mg = 0.0092 × y(mm)2.3038 (R2 = 0.78).
3.2 Growth and survival
The calibration of the von Bertalanffy growth model gave the intrinsic growth rate of rb = 0.0237 mm and asymptotic length of Li = 20.588 mm for O. craigsmithi. This further resulted in start age for the medium and large polychaete groups at 9 and 32.5 days, respectively (Figure 3B). The model also showed that the time for a 0.3-mm polychaete to reach the starting size of the small polychaetes used in our experiment was 11 days. Furthermore, they showed an estimated lifetime of ~15 weeks and a generation time of 4 weeks. Growth data revealed that the smallest size group of polychaetes had the highest LSGR of 4.2% ± 0.5% day−1 (Figure 3D). This group also doubled in length from 4.8 ± 0.5 mm to 11.0 ± 0.3 mm in length in 3 weeks before reaching their close to maximum size of 16 mm after 8 weeks (Figure 3A). The medium-sized polychaetes showed a maximum LSGR of 2.9% ± 0.6% day−1 during the first week of the experiment, which decreased to 1.3% ± 0.2% day−1 when polychaetes had reached a size of approximately 16 mm. The largest size group showed a slight increase in length and had a maximum growth rate of 1.7% ± 0.1% day−1 in the second week of the growth trial. Three weeks into the experiment, the growth decreased slightly for all three size groups. After the second week, the large group growth rates were stabilized, and their survival was down to 49% ± 22% after 3 weeks. The large groups continued to steadily decrease in numbers and survival was 23% ± 7% by week 6. In the last week of the experiment, only 17% ± 4% of the individuals were reaming. Medium- and small-sized groups showed high survival throughout the experiment, and, by the end of the growth trial, the medium group still maintained a survival of 91% ± 3%. Survival data (Figure 3C) show a variation in the number of polychaetes counted over time for the medium- and small-sized groups; this is explained by the difficulty to distinguish the smallest polychaetes in the beginning of the growth trial.
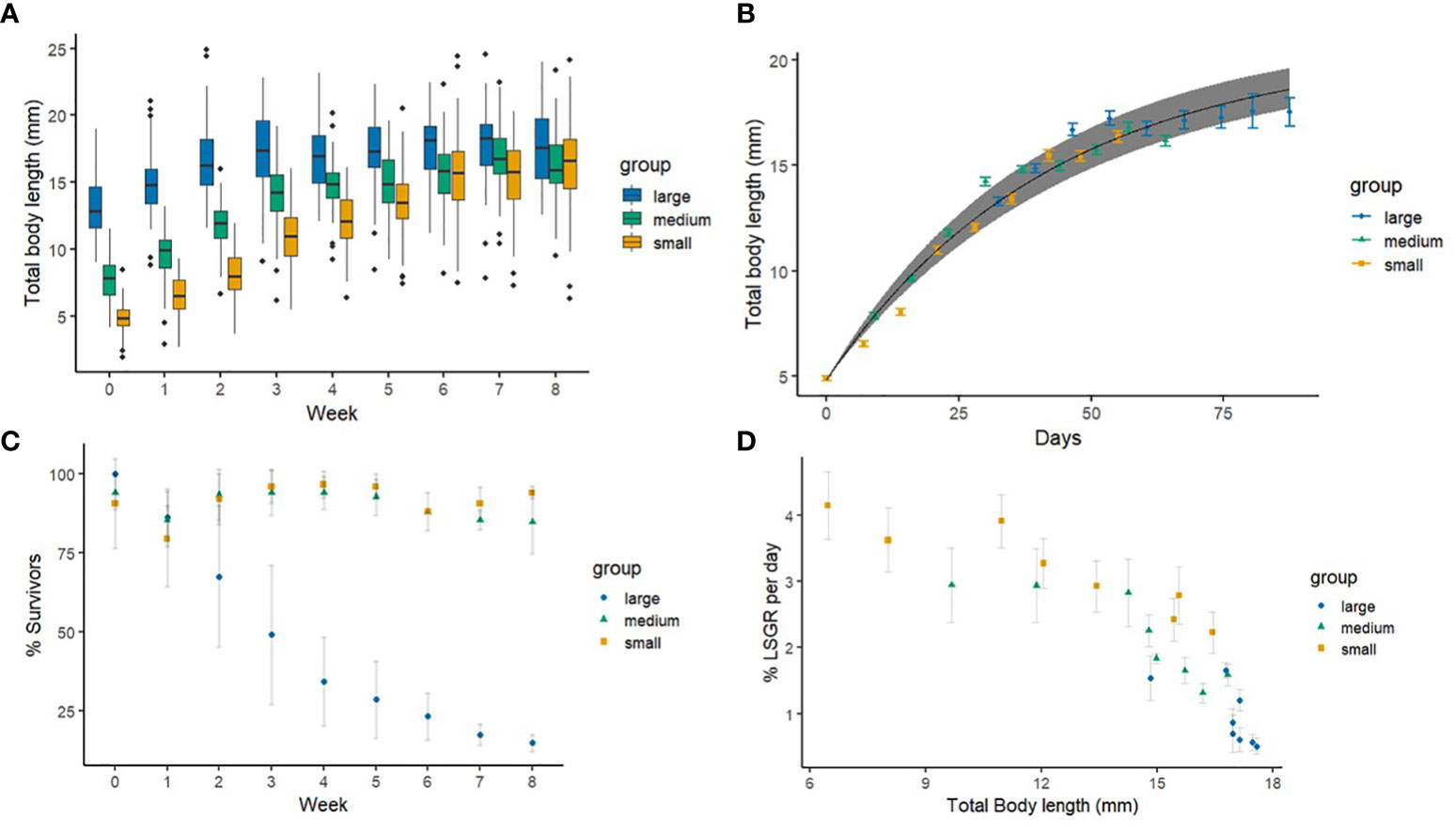
Figure 3 (A) Body lengths (in millimeters) for pooled polychaete groups over time with black horizontal line showing the median and vertical lines, illustrating minimum and maximum values. (B) The von Bertalanffy growth curve plotted together with measured data for mean total body length (mean ± SE) with the line for predicted mean values and 95% confidence interval. Polychaetae groups are plotted according to their calculated age (days). (C) Total survival (%) over time (n = 3 per group, mean ± SD). (D) Length-specific growth rate (LSGR % day−1) (n = 3 per group, mean ± SD). Size groups are separated by shape and color where blue circles = large, green triangles = medium, and yellow squares = small.
3.3 Histology
The sampled population consisted of 43% females and 53% males, and 4% of the individuals were not able to be determined. The smallest female sampled, 6 mm in unsedated length, contained oocytes in different stages of maturation. This trend for eggs in different maturation stages was consistent for the females sampled in the total body length range of 6–21 mm (Figure 4). All males longer than 7 mm contained a combination of mature/immature sperm, whereas half of the small males (5–7 mm; n = 6) showed only immature sperm. Egg and sperm were never observed in the same individual.

Figure 4 Histological section of (A) 21-mm-long female and (B) 25-mm-long male. IM, immature gametes; M, mature/maturing gametes. Scale bar, 100 µm.
3.3.1 Natural spawning
Eggs collected from natural spawning events were dominated by the sizes between 130 and 140 µm (Figure 5). The development stages (see Section 2.5.1 for definitions of stages) of all eggs, laid by the large group of O. craigsmithi (n = 635), showed 80% as stage 2 and 17% as stage 3, as determined by microscopy. Eggs within stage 2 displayed a large range in egg diameter from 115 to 202 µm. During the collection of naturally spawned eggs, both medium and large groups spawned, but only the large group laid sufficient amounts of eggs for measurements with the laser counter. However, eggs that were collected from the medium-sized groups followed similar trends in the egg size distribution. The eggs at sizes defined as mature ranged from 120 up to >190 µm with a clear peak at size of 140 µm in diameter (Figure 5). Eggs larger than 200 µm in diameter were hypothesized to swell due to change in osmotic pressure or have gone through multiple stages of cell division. Eggs smaller than 120 µm in diameter corresponded to stage 2 (see Figure 1) and were therefore not categorized as mature.
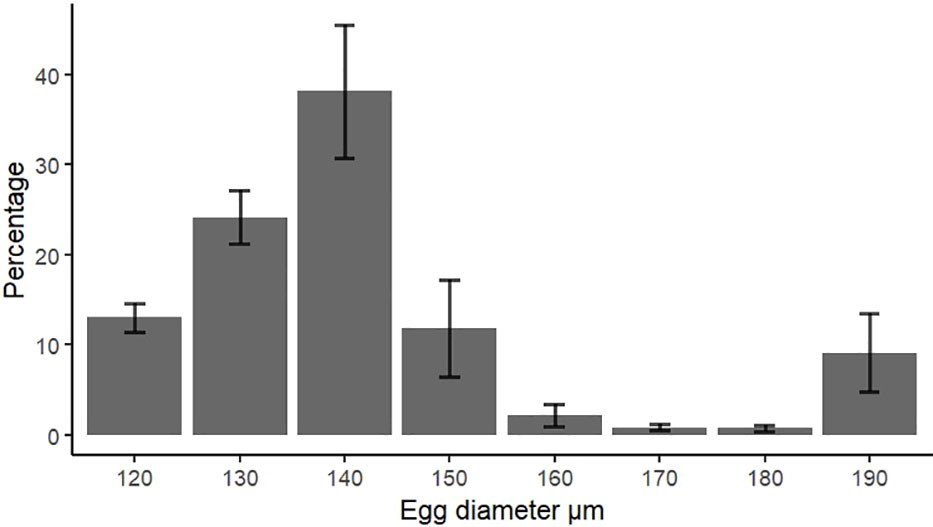
Figure 5 Size distribution of mature eggs (mean ± SD) collected during natural spawning events from the large polychaetes (N = 3 chambers, n = 20 polychaetes per chamber).”Mature” eggs defined by microscopy (Figure 1).
Courtship behavior was frequently observed in the holding tanks. Male and female would attach, and one individual appeared to twist itself around the other. Subsequent vigorous “wiggling” took place before a small white cloud was emitted into the water and settled on the bottom (see Supplementary Material for video). Pipetting of this material confirmed that there were eggs that had been fertilized, and a couple showing courtship behavior was also included in histology, which confirmed that the couple consisted of male and female.
3.3.2 Individual fecundity
There was an apparent variation in the number of eggs spawned by individuals of a similar length (Figure 6). There was no linear correlation between the total body length and the number of spawned eggs (p = 0.63). The majority of polychaetes spawned less than 250 eggs, whereas the maximum number of spawned eggs was 1,063 spawned by an individual of 13.1 mm. Mature eggs were not released by individuals smaller than ~6 mm in length, and neither were immature eggs observed during dissections on the same individuals. Seventeen large (> 10 mm in length) individuals also did not spawn after heat-shock treatment, but dissection revealed that these individuals sometimes contained a vast number of immature eggs.
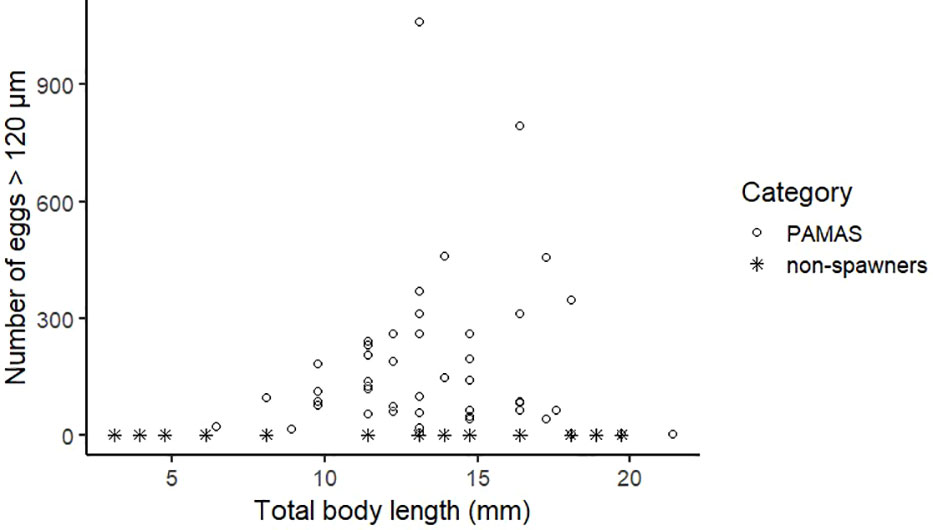
Figure 6 Individual fecundity as number of eggs > 120 µm released per individual after heat-shock treatment (n = 73). Circles represent spawning individuals, and asterisks represent non-spawning individuals (34% of individuals included did not spawn).
3.4 Larvae development, mucus production, and general behavior
Determination of developmental stages showed that fertilized eggs had started and gone through early cleavage within 6 h after collection (Figure 7). After ~15 h, we observed slow circular movements in the eggs. A free-swimming trochophore-like stage appeared after 70 h. This has developed later into a metatrochophore with clearly visible ciliated bands. The first segment appeared after 114 h and grew into three- to four-segment-long free-swimming larvae without chaeta after 1 week. Although there were several feeding attempts of larvae, offering live microalgae and different types of aquarium fish feed, there was no successful rearing of larvae beyond this point. This timeline for development of larvae should be interpreted with care due to few successful replicates.
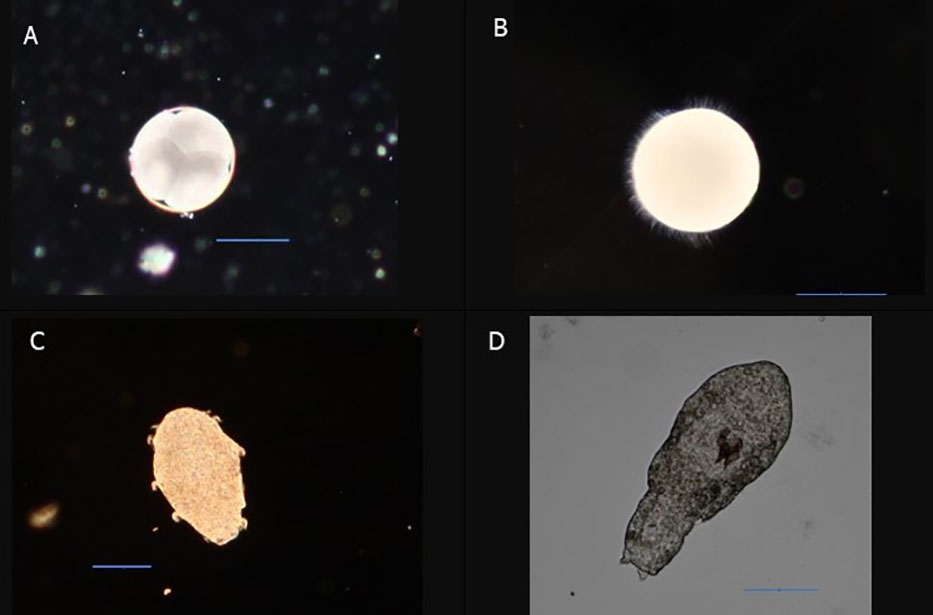
Figure 7 Developmental stages of fertilized eggs. (A) Egg with cell division. (B) Circular rotating stage prior to free-swimming gastrula. (C) Metatrochophore stage with ciliated bands. (D) Free-swimming larvae stage without chaetigers but with clearly visible jaws. Scale bar represents 100 µm.
Colonies of polychaetes formed three-dimensional–like structures with mucus along the edges of the holding tanks. Mucus was frequently sampled to investigate whether mucus were used for the development of larvae; however, larvae was never observed inside mucus. Individuals were very mobile and could quickly (hours to minutes) localize feed introduced to the flumes.
4 Discussion
In this study, we investigate key life history traits necessary for assessing O. craigsmithi as a candidate species in IMTA. We find that these polychaetes fed on a diet of fish pellets obtained high growth rates and have a short generation and lifetime. Furthermore, this species is gonochoristic and, most likely, a semicontinuous spawner. They have a free-swimming larvae stage that develops after approximately 1 week after fertilization at ~10°C. After assessing life history traits, nutrient mitigation, and production potential, the high potential for O. craigsmithi as an extractive species in (benthic) IMTA is strengthened, and more knowledge regarding the population growth and biomass under farms is the next step for understanding their future role in IMTA systems.
4.1 Growth, survival, and lifetime
Small individuals doubled in length in the first 3 weeks of the growth experiment, and, during this time frame, polychaetes went from non-spawning members of the population to spawning adults. The decreased growth measured for the adults coincided with increased mortality. At this stage, polychaetes were estimated to be 50 days old, indicating that this species has a short lifespan. The estimated lifespan for this polychaete is ~15 weeks, and the generation time is ~4 weeks. Their lifetime and generation time are comparable with those Ophryotrocha sp. measured at temperature rates of 15°C and 24°C (Åkesson, 1973; Simonini and Prevedelli, 2003). However, their lifetime is significantly shorter than the approximately 34 weeks observed for a temperate deep-sea Ophryotrocha sp. reported by Mercier et al. (2014). The high mortality rate measured for the large groups in the growth experiment was not observed in the two other size groups, even when their estimated age and size were comparable. The reason for this high survival remains unknown, and future growth studies should last until at least a 50% mortality rate is observed to further assess their lifetimes. The growth rates in this study (1.5%–2.9% day−1 for individuals between 12 and 14 mm in size) are comparable with previous results for similar sized individuals of the same species reared on a diet of freshly collected fish feces (Nederlof et al., 2019). These combined results indicate a high capacity for growth when reared on different waste fraction from fish farms. Care should be taken when comparing growth rates between species as the specific growth rate calculations include the starting size and the time interval between measurements, which are generally study and species specific. Nevertheless, these rates can be seen as an indication of the ability of a species to grow on a specific diet in a given environment. Wang et al. (2020) not only demonstrated the high growth rates of juvenile H. diversicolour reared in the laboratory with a commercial fish feed diet but also highlighted that this could vary based on the type of diet and the substrate provided. We assume that optimal growth rates were measured in our study given the high food quality provided in the growth trial. Furthermore, growth rates measured for young adults in our study (8–12 mm in length) were in the same order of magnitude to what has been observed for other polychaete species reared on aquaculture waste such as H. diversicolor (Wang et al., 2019) and Capitella sp (Nederlof et al., 2019). It can be hypothesized that the decreasing growth observed approximately 15 mm in size is linked to energy allocation to reproduction. For O. labronica, such investment into reproductive tissue has been proven to be significant (Cassai and Prevedelli, 1999).
4.2 Reproduction
We further conclude that this is a gonochoristic species on the basis of histology and the morphological characteristics distinct for individuals carrying eggs or sperm. The genus is represented by species with different modes of reproduction, but gonochoric is the most common one (Thornhill et al., 2009).
Several observations in our study support that O. craigsmithi is a semicontinuous spawner. First, the variation in the number of spawned eggs was large during spawning by individuals of a similar size. For example, for the polychaetes between 13 and 16 mm in length, the number of eggs varied between 0 and >1,000 for a single spawning event, suggesting that several batches of mature eggs could be produced after reaching reproductive age. Despite the large variability across sizes, it seems that the number of released eggs is related to polychaete size or age. A similar pattern has been described for O. puerelis, where the number of eggs increased from 30 eggs during first spawning to 300–400 eggs in older individuals (Åkesson, 1967). Second, dissections of spawned polychaetes revealed the presence of remaining, mostly immature, eggs. This, in combination with histology showing that all females (7–20 mm) contained eggs in several stages of maturation, builds on the first argument for semicontinuous spawning. Finally, they appear to have the ability to spawn naturally already at half of their maximum size (4 weeks of age), and, at this point, survival was high. Semicontinuous spawning is a common trait within Ophryotrocha (Åkesson, 1973; Thornhill et al., 2009) and is thought to be an important trait for opportunistic species as a way of quickly responding to changing environments (Tsutsumi, 1987). Given that the number of spawning’s per individual remains unknown, the lifetime fecundity could not be determined in our study. To do so, it requires investigation of individuals over time, which was not possible in our study as isolated individuals/couples did not spawn. The resistance to spawn spontaneously in isolation has also been was observed for the temperate O. cosmetandra (Oug, 1990). It is reasonable to assume that the high nutritional value provided in this study may have maximized the reproductive output in terms of number of eggs. The degree of importance of food quality and its potential influence on fecundity has been demonstrated for Dinophilus gyrociliatus, where a high-quality feed resulted in a two- to three-fold higher fecundity (Prevedelli and Simonini, 2000).
The size of mature eggs (120–160 um) was within ranges as reported in other Ophryotrocha spp (Paxton and Akesson, 2010; Mercier et al., 2014). However, the range in size for mature eggs seems to be large. The egg size and number are important parameters, indicating the amount of energy invested in reproduction by the individual that affects the population growth of the species (Simonini and Prevedelli, 2003). Spawning behavior revealed that eggs were released freely into the water before gently sinking to the bottom. This is unusual among Ophryotrocha because most species prepare some type of mucus cocoon or loose jelly where they deposit their eggs (Wilson, 1991; Thornhill et al., 2009), showing that this species does not put any effort in brood care. On the other hand, the timeline of larval development (~10°C) converged with patterns observed within Ophryotrocha spp. reared at 15°C–25°C (Åkesson, 1973), and the first stages of development were similar to that in O. shieldsi collected under a Tasmanian fish farm (Paxton and Davey, 2010). However, their development was faster than what has been reported for the temperate O. cosmetandra whose early development at 5°C–8°C took 30–50 days (Oug, 1990), implying that O. craigsmithi is highly adapted to rapid colonization and spreads and grows in temperate environments.
4.3 Implications for benthic IMTA
Insight into life history traits is of key importance when evaluating the suitability of a species for IMTA. To fully assess their potential, multiple aspects need to be considered. We therefore present a framework (Table 1), identifying the requirements for benthic polychaetaes to be in IMTA. The following factors are considered: environmental and biological suitability as well as nutrient mitigation capacity and market potential.
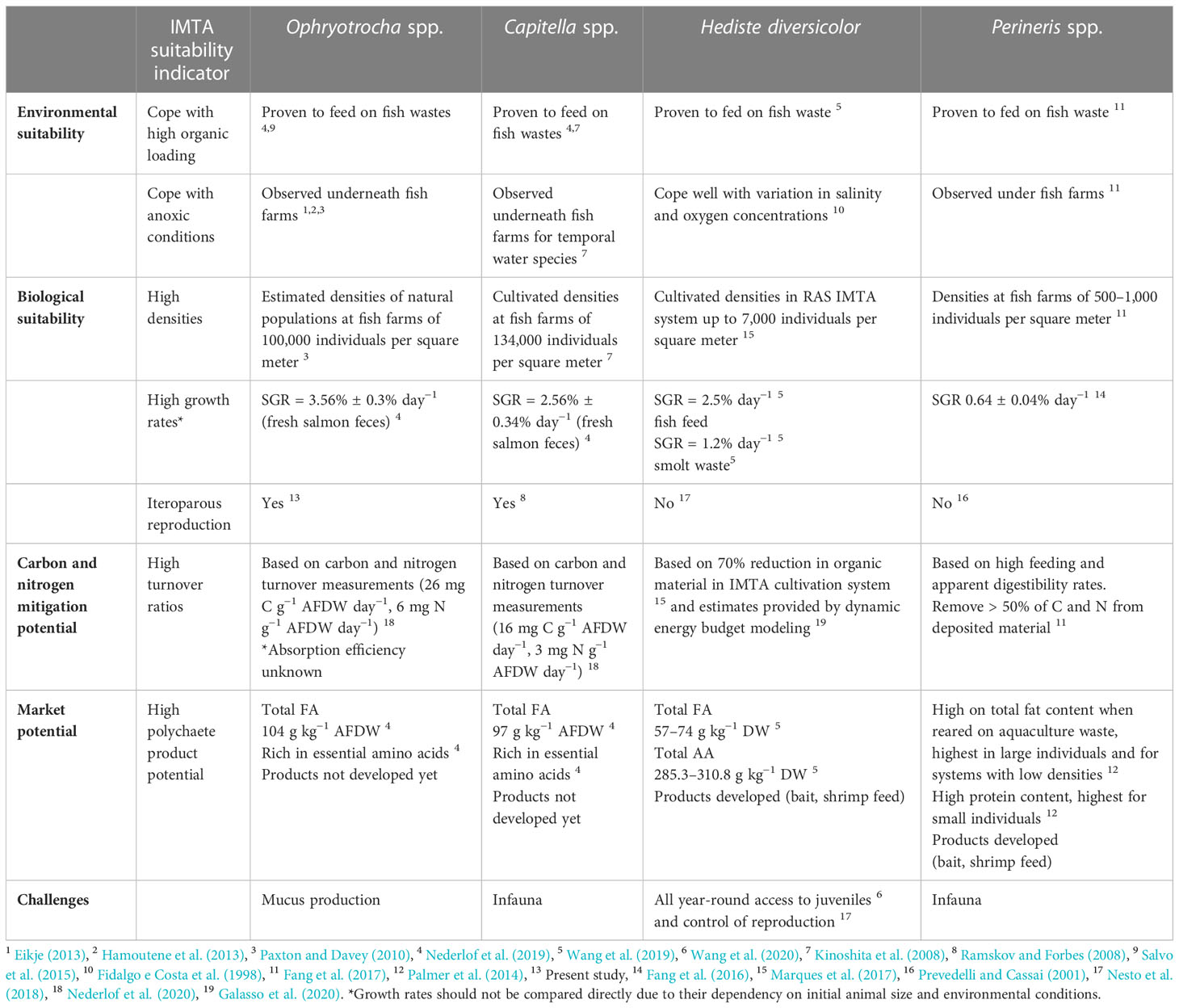
Table 1 Framework for evaluating the suitability of deposit feeding polychaetes as candidate species in IMTA, based on a set of IMTA suitability indicators.
Several polychaete species show potential as candidates in IMTA, all with their pros and cons (Table 1). The major challenges for successful incorporation of polychaetes in IMTA are often connected to controlling reproduction to have continuous access to polychaetes and to optimize the trade-off between biomass increase and rearing densities (Nesto et al., 2012; Palmer et al., 2014; Pombo et al., 2020; Wang et al., 2020). O. craigsmithi is not only among the smallest polychaetae candidate in our comparison between species in IMTA settings but also one of those reported to reach some of the highest densities under fish farms (Paxton and Davey, 2010; Eikje, 2013; Hamoutene et al., 2013). Furthermore, O. craigsmithi shows high carbon and nitrogen mitigation potential and high growth rates (Nederlof et al., 2020). Short generation time accompanied with semicontinuous reproduction would result in high population growth and allow for multiple harvests during a single fish production cycle that could increase the total yield for harvested polychaetes. Capitella sp. has been successfully seeded in organically enriched sediment under fish farms and has reached densities as high as 134,000 individuals per square meter (Kinoshita et al., 2008), showing its potential for being applied at sites situated over soft bottom substrate. Pereneris aibuthiensis has also shown promising nutrient mitigation potential when combining their organic removal efficiency and required densities for successful bio-mitigation under fish farms (Fang et al., 2017). Furthermore, H. diversicolor is a promising candidate for closed systems. Marques et al. (2017) reported a 70% reduction in organic material when H. diversicolor was integrated in sand filter systems and provided with aquaculture waste. Both H. diversicolor and Pereneris sp. are generally reared in systems with temperature rates of approximately 16°C to 20°C, whereas Capitella sp. and O. craigsmithi show higher suitability for open-water IMTA systems at greater depths where temperature is stable at ~8°C. High abundance observed for craigsmithi on hard substrates in Norwegian waters indicates a potential competitive advantage in these environments (Hansen et al., 2011; Brennan, 2018). This is opposite to Capitella sp., which is mainly found in soft sediment. Aspects such as substrate preference are crucial when considering future technical development to the chosen polychaete in open-water IMTA. Both Pereneris sp. and H. diversicolor, belonging to the Neridae family, have a semelparous reproduction strategy, which has been pointed out as a challenge for maintaining stable stocking densities within IMTA system (Prevedelli and Cassai, 2001; Wang et al., 2020). In contrast, O. craigsmithi and Capitella sp. have iteroparity and provide the advantage of continues recruitment. All polychaetes considered in the framework comprise valuable contents of fatty acids and essential ammino acids when feed with aquaculture waste, making them attractive as a raw material in aquatic feed. However, today, only Perineris sp. and H. diversicolor have a market potential due to their use as bait or as a component in shrimp feed (Olive, 1999; Palmer et al., 2014).
5 Conclusion
This study shows that O. craigsmithi displays multiple life history traits that are of benefit for farming and harvesting populations in benthic IMTA. They obtained high growth rates when fed with fish feed and have short a short generation time with multiple spawning events throughout their lifetime. This implies a high population growth and opens up for the possibility of multiple polychaete harvests during a single fish production cycle. Furthermore, these are gonochoric, and eggs are typically 120–150 µm in diameter. This species can initiate reproduction at half of their adult length, and fertilized eggs develop into free-swimming larvae after approximately 1 week, but further studies are needed to confirm this and to provide details for development past the free-swimming larvae stage. Life history traits are in line with what has been reported by studies on other members of the genus occupying shallower and warmer waters, but O. craigsmithi stands out as a large Ophryotrocha with broadcasting spawning. The described life history traits together with the presented framework, assessing different polychaete species suitability in a benthic IMTA, suggest that O. craigsmithi is an efficient, and well-suited extractive species for this purpose. Market potential remains to be realized, and future research should focus on further assessing the biomass and harvest potential in an open-water IMTA setting.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SS: First authorship with main responsibility for planning for experiments, collection and processing of data; main author on writing the current manuscript. TS: second author; key role in fieldwork and planning of the experiment; has contributed with guidance and editing in the manuscript. AG: third author, assisting in analyses of the data, especially for applying the von-bertalanffy model; provided feedback on the current manuscript. HR: third author, assisting in laboratory work, especially during growth trial and larvae development; feedback in the current manuscript. E-JL: Fourth author, provided feedback during the editing manuscript. HS: Fifth author, provided feedback during the editing manuscript. HJ: Senior authorship, she has made a great contribution to editing this manuscript and was involved during the planning and at some data sampling points during data collection in the laboratory. All authors contributed to the article and approved the submitted version.
Funding
This work was founded by the Norwegian Research Council, through project 323824.
Acknowledgments
We wish to thank Natalyia Budaeva, a professor at the University Museum in Bergen, for her assistance and advice in histology. We also want to thank the workers from Lerøy Sjøtroll at the fish farms Rongøy and Matløyso in Bergen for their assistance during field collection of polychaetes. Finally, we wish to thank Helena Wiklund and Thomas Dahlgren at the University of Gothenburg for generously sharing their expertise on the Ophryotrocha spp. genus.
Conflict of interest
Authors SS and HS are employed in Lerøy seafood group.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1116765/full#supplementary-material
References
Åkesson B. (1967). On the biology and larval morphology of ophryotrocha puerilis claparede & metschnikov (Polychaeta). Ophelia 4 (1), 111–119. doi: 10.1080/00785326.1967.10409615
Åkesson B. (1973). Reproduction and larval morphology of five ophryotrocha species (Poly cha eta, dorvilleidae). Zoologica Scripta 2 (4), 145–155.
Albrektsen S., Kortet R., Skov P. V., Ytteborg E., Gitlesen S., Kleinegris D., et al. (2022). Future feed resources in sustainable salmonid production: A review. Rev. Aquaculture. 14(4). doi: 10.1111/raq.12673
Armstrong E. G., Mersereau J., Salvo F., Hamoutene D., Dufour S. C. (2020). Temporal change in the spatial distribution of visual organic enrichment indicators at aquaculture sites in Newfoundland, Canada. Aquaculture Int. 28 (2), 569–586. doi: 10.1007/s10499-019-00478-z
Brennan N. B. (2018). Studying the waste recycling potential of naturally occurring polychaetes on benthic trays under a Norwegian fish farm (The University of Bergen).
Broch O. J., Daae R. L., Ellingsen I. H., Nepstad R., Bendiksen E.Å., Reed J. L., et al. (2017). Spatiotemporal dispersal and deposition of fish farm wastes: a model study from central Norway. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00199
Cassai C., Prevedelli D. (1999). Fecundity and reproductive effort in ophryotrocha labronica (Polychaeta : Dorvilleidae). Mar. Biol. 133 (3), 489–494. doi: 10.1007/s002270050488
Chopin T., Cooper J. A., Reid G., Cross S., Moore C. (2012). Open-water integrated multi-trophic aquaculture: environmental biomitigation and economic diversification of fed aquaculture by extractive aquaculture. Rev. Aquaculture 4 (4), 209–220. doi: 10.1111/j.1753-5131.2012.01074.x
Cranford P. J., Reid G. K., Robinson S. M. (2013). Open water integrated multi-trophic aquaculture: constraints on the effectiveness of mussels as an organic extractive component. Aquaculture Environ. Interact. 4 (2), 163–173.
Dahlgren T. G., Akesson B., Schander C., Halanych K. M., Sundberg P. (2001). Molecular phylogeny of the model annelid ophryotrocha. Biol. Bull. 201 (2), 193–203.
Eikje E. (2013). Benthic impacts of fish-farm waste on hard bottom habitats, the ecology of opportunistic epifauna polychaetes. Institutt Biologi 61.
Fang J., Jiang Z., Jansen H. M., Hu F., Fang J., Liu Y., et al. (2017). Applicability of perinereis aibuhitensis grube for fish waste removal from fish cages in sanggou bay, PR China. J. Ocean Univ. China 16 (2), 294–304. doi: 10.1007/s11802-017-3256-1
Fang J., Zhang J., Jiang Z., Du M., Liu Y., Mao Y., et al. (2016). Environmental remediation potential of perinereis aibuhitensis (Polychaeta) based on the effects of temperature and feed types on its carbon and nitrogen budgets. Mar. Biol. Res. 12 (6), 583–594. doi: 10.1080/17451000.2016.1177653
Fang J. H., Samuelsen O. B., Strand O., Jansen H. (2018). Acute toxic effects of hydrogen peroxide, used for salmon lice treatment, on the survival of polychaetes capitella sp and ophryotrocha spp. Aquaculture Environ. Interact. 10, 363–368.
Fidalgo e Costa P. F., Sarda R., da Fonseca L. C. (1998). Life cycle, growth and production of the polychaete nereis diversicolor OF müller in three lagoonal estuarine systems of the southwestern Portuguese coast (Odeceixe, aljezur and carrapateira). Ecologie 29 (4), 523.
Filgueira R., Guyondet T., Reid G., Grant J., Cranford P. (2017). Vertical particle fluxes dominate integrated multi−trophic aquaculture (IMTA) sites: implications for shellfish-finfish synergy. Aquaculture Environ. Interact. 9, 127–143.
Galasso H. L., Lefebvre S., Aliaume C., Sadoul B., Callier M. D. (2020). Using the dynamic energy budget theory to evaluate the bioremediation potential of the polychaete hediste diversicolor in an integrated multi-trophic aquaculture system. Ecol. Model. 437, 10. doi: 10.1016/j.ecolmodel.2020.109296
Hamoutene D., Mabrouk G., Sheppard L., MacSween C., Coughlan E., Grant C. (2013). Validating the use of beggiatoa sp. and opportunistic polychaete worm complex (OPC) as indicators of benthic habitat condition at finfish aquaculture sites in Newfoundland (Science Branch, Fisheries and Oceans Canada).
Hansen P. K., Bannister R., Husa V. (2011). Utslipp fra matfiskanlegg: Påvirkning på grunne og dype hardbunnslokaliteter. The Institute of Marine Research Bergen, Norway.
Jansen H. M., Hansen P. K., Brennan N., Dahlgren T. G., Fang J., Nederlof M., et al. (2019). Enhancing opportunistic polychaete communities under fish farms: an alternative concept for integrated aquaculture. Aquaculture Environ. Interact. 11, 331–336. doi: 10.3354/aei00318
Keeley N., Valdemarsen T., Strohmeier T., Pochon X., Dahlgren T., Bannister R. (2020). Mixed-habitat assimilation of organic waste in coastal environments–it's all about synergy! Sci. Total Environ. 699, 134281. doi: 10.1016/j.scitotenv.2019.134281
Kinoshita K., Tamaki S., Yoshioka M., Srithonguthai S., Kunihiro T., Ohwada K., et al. (2008). Bioremediation of organically enriched sediment deposited below fish farms with artificially mass-cultured colonies of a deposit-feeding polychaete capitella sp. I. Fisheries Sci. 74 (1), 77–87. doi: 10.1111/j.1444-2906.2007.01498.x
Kutti T., Ervik A., Hansen P. K. (2007a). Effects of organic effluents from a salmon farm on a fjord system. i. vertical export and dispersal processes. Aquaculture 262 (2-4), 367–381. doi: 10.1016/j.aquaculture.2006.10.010
Kutti T., Hansen P. K., Ervik A., Høisæter T., Johannessen P. (2007b). Effects of organic effluents from a salmon farm on a fjord system. II. temporal and spatial patterns in infauna community composition. Aquaculture 262 (2-4), 355–366. doi: 10.1016/j.aquaculture.2006.10.008
Kvalø Heggøy K., Schander C., Åkesson B. (2007). The phylogeny of the annelid genus ophryotrocha (Dorvilleidae). Mar. Biol. Res. 3 (6), 412–420. doi: 10.1080/17451000701695361
Marques B., Lillebo A. I., Ricardo F., Nunes C., Coimbra M. A., Calado R. (2017). Adding value to ragworms (Hediste diversicolor) through the bioremediation of a super-intensive marine fish farm. Aquaculture Environ. Interact. 10, 79–88. doi: 10.3354/aei00255
Mercier A., Baillon S., Hamel J.-F. (2014). Life history and seasonal breeding of the deep-sea annelid ophryotrocha sp.(Polychaeta: Dorvelleidae). Deep Sea Res. Part I: Oceanographic Res. Papers 91, 27–35. doi: 10.1016/j.dsr.2014.05.007
Nederlof M. A., Fang J., Dahlgren T. G., Rastrick S. P., Smaal A. C., Strand Ø., et al. (2020). Application of polychaetes in (de) coupled integrated aquaculture: an approach for fish waste bioremediation. Aquaculture Environ. Interact. 12, 385–399. doi: 10.3354/aei00371
Nederlof M. A., Jansen H. M., Dahlgren T. G., Fang J., Meier S., ivind Strand Ã., et al. (2019). Application of polychaetes in (de) coupled integrated aquaculture: production of a high-quality marine resource. Aquaculture Environ. Interact. 11, 221–237. doi: 10.3354/aei00309
Nesto N., Simonini R., Prevedelli D., Da Ros L. (2012). Effects of diet and density on growth, survival and gametogenesis of hediste diversicolor (OF müller 1776)(Nereididae, polychaeta). Aquaculture 362, 1–9. doi: 10.1016/j.aquaculture.2012.07.025
Nesto N., Simonini R., Prevedelli D., Da Ros L. (2018). Evaluation of different procedures for fertilization and larvae production in hediste diversicolor (O.F. müller 1776) (Nereididae, polychaeta). Aquaculture Res. 49 (4), 1396–1406. doi: 10.1111/are.13589
Olive P. J. (1999). Polychaete aquaculture and polychaete science: A mutual synergism. Hydrobiologia 402, 175–183. doi: 10.1023/A:1003744610012
Oug E. (1990). Morphology, reproduction, and development of a new species of ophryotrocha (Polychaeta, dorvilleidae) with strong sexual dimorphism. Sarsia 75 (3), 191–201.
Palmer P. J., Wang S., Houlihan A., Brock I. (2014). Nutritional status of a nereidid polychaete cultured in sand filters of mariculture wastewater. Aquaculture Nutr. 20 (6), 675–691. doi: 10.1111/anu.12129
Paxton H., Akesson B. (2010). The ophryotrocha labronica group (Annelida: Dorvilleidae) - with the description of seven new species. Zootaxa 2713), 1–24. doi: 10.1016/j.aquaculture.2012.07.025
Paxton H., Davey A. (2010). A new species of ophryotrocha (Annelida: Dorvilleidae) associated with fish farming at macquarie harbour, Tasmania, Australia. Zootaxa 2509 (1), 53–61.
Pombo A., Baptista T., Granada L., Ferreira S. M. F., Goncalves S. C., Anjos C., et al. (2020). Insight into aquaculture's potential of marine annelid worms and ecological concerns: a review. Rev. Aquaculture 12 (1), 107–121. doi: 10.1111/raq.12307
Prevedelli D., Cassai C. (2001). Reproduction and larval development of perinereis rullieri pilato in the Mediterranean Sea (Polychaeta: Nereididae). Ophelia 54 (2), 133–142. doi: 10.1080/00785236.2001.10409461
Prevedelli D., Simonini R. (2000). Effects of salinity and two food regimes on survival, fecundity and sex ratio in two groups of dinophilus gyrociliatus (Polychaeta: Dinophilidae). Mar. Biol. 137 (1), 23–29. doi: 10.1007/s002270000327
Ramskov T., Forbes V. E. (2008). Life history and population dynamics of the opportunistic polychaete capitella sp. I in relation to sediment organic matter. Mar. Ecol. Prog. Ser. 369, 181–192. doi: 10.3354/meps07584
Salvo F., Dufour S. C., Hamoutene D., Parrish C. C. (2015). Lipid classes and fatty acids in ophryotrocha cyclops, a dorvilleid from Newfoundland aquaculture sites. PLoS One 10 (8). doi: 10.1371/journal.pone.0136772
Salvo F., Wiklund H., Dufour S. C., Hamoutene D., Pohle G., Worsaae K. (2014). A new annelid species from whalebones in Greenland and aquaculture sites in Newfoundland: Ophryotrocha cyclops, sp. nov.(Eunicida: Dorvilleidae). Zootaxa 3887 (5), 555–568. doi: 10.11646/zootaxa.3887.5.3
Schneider C. A., Rasband W. S., Eliceiri K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9 (7), 671–675.
Simonini R., Prevedelli D. (2003). Life history and demography of three populations of ophryotrocha japonica (Polychaeta: Dorvilleidae). Mar. Ecol. Prog. Ser. 258, 171–180. doi: 10.3354/meps258171
Strain P., Hargrave B. (2005). Salmon aquaculture, nutrient fluxes and ecosystem processes in southwestern new Brunswick. Environ. Effects Mar. Finfish Aquaculture, 29–57. doi: 10.1007/b136003
Thornhill D. J., Dahlgren T. G., Halanych K. M. (2009). Evolution and ecology of ophryotrocha (Dorvilleidae, eunicida). Annelids modern Biol., 242–256.
Troell M., Joyce A., Chopin T., Neori A., Buschmann A. H., Fang J.-G. (2009). Ecological engineering in aquaculture–potential for integrated multi-trophic aquaculture (IMTA) in marine offshore systems. Aquaculture 297 (1-4), 1–9. doi: 10.1016/j.aquaculture.2009.09.010
Tsutsumi H. (1987). Population dynamics of capitella capitata (Polychaeta; capitellidae) in an organically polluted cove. Mar. Ecol. Prog. Ser. 36, 139–149.
Tsutsumi H., Fukunaga S., Fujita N., Sumida M. (1990). Relationship between growth of capitella sp. and organic enrichment of the sediment. Mar. Ecol. Prog. series. Oldendorf 63 (2), 157–162.
Wang H., Seekamp I., Malzahn A., Hagemann A., Carvajal A. K., Slizyte R., et al. (2019). Growth and nutritional composition of the polychaete hediste diversicolor (OF müller 1776) cultivated on waste from land-based salmon smolt aquaculture. Aquaculture 502, 232–241. doi: 10.1016/j.aquaculture.2018.12.047
Wang H. Q., Hagemann A., Reitan K. I., Handa A., Uhre M., Malzahn A. M. (2020). Embryonic and larval development in the semelparous nereid polychaeteHediste diversicolor(OF muller 1776) in Norway: Challenges and perspectives. Aquaculture Res. 51 (10), 4135–4151. doi: 10.1111/are.14756
Wang X., Olsen L. M., Reitan K. I., Olsen Y. (2012). Discharge of nutrient wastes from salmon farms: environmental effects, and potential for integrated multi-trophic aquaculture. Aquaculture Environ. Interact. 2 (3), 267–283.
Wiklund H., Glover A. G., Dahlgren T. G. (2009). Three new species of ophryotrocha (Annelida: Dorvilleidae) from a whale-fall in the north-East Atlantic. Zootaxa 2228 (1), 43–56.
Keywords: deposit feeders, polychaete, circular production, mitigation, salmon farming
Citation: Svensson SGB, Strohmeier T, Rastrick H, Garcia AA, Lock E-J, Sveier H and Jansen HM (2023) Life history traits for Ophryotrocha craigsmithi (Wiklund, Glover & Dahlgren, 2009), a candidate species in integrated multitrophic aquaculture. Front. Mar. Sci. 10:1116765. doi: 10.3389/fmars.2023.1116765
Received: 05 December 2022; Accepted: 18 January 2023;
Published: 07 February 2023.
Edited by:
Marcel Martinez-Porchas, Centro de Investigación en Alimentación y Desarrollo, Consejo Nacional de Ciencia y Tecnología (CONACYT), MexicoReviewed by:
Jianguang Fang, Yellow Sea Fisheries Research Institute (CAFS), ChinaJinghui Fang, Yellow Sea Fisheries Research Institute (CAFS), China
Stefano Carboni, Fondazione IMC, Italy
José Lino Vieira De Oliveira Costa, University of Lisbon, Portugal
Ana Pombo, Center for Marine and Environmental Sciences (MARE- IPLeiria), Portugal
Copyright © 2023 Svensson, Strohmeier, Rastrick, Garcia, Lock, Sveier and Jansen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Signe Gunborg Bentzrød Svensson, c2lnbmUuc3ZlbnNzb25AdWliLm5v
†These authors have contributed equally to this work and share third authorship
‡These authors have contributed equally to this work and share fifth authorship