 Aylin Akkaya1Tim Awbery1,2
Aylin Akkaya1Tim Awbery1,2 Kirsty Medcalf1*Patrick Lyne1
Kirsty Medcalf1*Patrick Lyne1 Giulia Cipriano3
Giulia Cipriano3 Morgana Alvarenga1
Morgana Alvarenga1 Leyla İsrapilova1
Leyla İsrapilova1 Yağmur Atalan1
Yağmur Atalan1 Olivier Eikelenboom1
Olivier Eikelenboom1 Pasquale Ricci3Roberto Crugliano4
Pasquale Ricci3Roberto Crugliano4 Elena Papale5Carmelo Fanizza4
Elena Papale5Carmelo Fanizza4 Roberto Carlucci3
Roberto Carlucci3- 1Deniz Memelileri Araştırma Derneği (DMAD)- Marine Mammals Research Association, Istanbul, Türkiye
- 2Marine Mammal Research Team, Scottish Association for Marine Science (SAMS), Oban, United Kingdom
- 3Department of Bioscience, Biotechnologies and Environment, University of Bari, Bari, Italy
- 4Institute for the Study of Anthropic Impacts and Sustainability in the Marine Environment (IAS), National Research Council, unit of Capo Granitola, Torretta Granitola (TP), Italy
- 5Unit of Capo Granitola, National Research Council, Institute for the Study of Anthropic Impacts and Sustainability in the Marine Environment (IAS), Torretta, Italy
Bottlenose dolphins have a complex vocal repertoire that varies depending on behavioral context, social structure, group composition, and anthropogenic pressures. This current study describes the whistle characteristics of bottlenose dolphins for the first time in the South Adriatic Sea while assessing the potential differences between whistle characteristics of geographically separated dolphins within neighbouring waters of the North Ionian Sea. The results show that whistle characteristics were similar between Taranto Gulf (Italy) and Boka Bay (Montenegro), despite their spatial differences. The mean peak frequency was 10kHz for each study location while the mean minimum and maximum frequency ranged from 7 to 14kHz. The average duration of whistles was 500 milliseconds. These results share similarities with previous literature, although several studies reported slightly different mean peak frequencies, ranging up to 15kHz in the neighbouring waters of Croatia and Italy. Further, harmonics were produced and formed in 40% of the whistles in Taranto Gulf and 30% of the whistles in Boka Bay. A high incidence of harmonics has previously been associated with behavioral states (i.e., travelling) and with certain types of marine traffic (i.e., fishing vessels). Therefore, it is important to collect simultaneous data on the visual behavior of the focal group as well as document the type and density of marine traffic within the proximity of the dolphins to have an in-depth understanding of vocal behavior. Despite the similarities of whistle characteristics of Taranto and Boka Bay, the whistle contours showed notable variations. Upsweep whistles were the most regularly produced whistle type in each location, which coincides with previous studies in the Mediterranean Sea. However, the least produced whistle had a concave contour in Taranto and was flat in Boka Bay. Previous studies have confirmed that flat whistles account for the least produced whistle contour in the Mediterranean Basin. Examining the whistle characteristics and the variation in whistle contours provides an in-depth understanding of the behavioral complexity as well as its plasticity in the presence of pressure. Therefore, future studies need to include behavior, group composition, noise levels, and human presence to enable an effective understanding of variation in whistle characteristics of bottlenose dolphins.
1 Introduction
Cetaceans are recognized as key top-level controllers in marine ecosystems due to their roles in preserving the structure and function of marine communities (Heithaus et al., 2013; Ricci et al., 2021; Carlucci et al., 2021). The ecological health and population status of cetaceans can potentially be used as bio-indicators for the assessment of ecosystem health (Wells et al., 2004; Paudel et al., 2020). The implementation of effective strategies to conserve these species can spread downwards throughout the food chain and protect the entire ecosystem. Therefore, robust knowledge regarding population size, spatial-temporal habitat usage, and home ranges coupled with threat assessment are of critical importance not only for the species in question but also for the entire marine ecosystem (Panigada et al., 2017; Azzolin et al., 2018; Carlucci et al., 2021). The western and central Mediterranean Sea, including the northern and central Adriatic Sea have benefited from dedicated research effort on cetaceans since the 1980s (Notarbartolo di Sciara et al., 1993; Bearzi and Notarbartolo di Sciara, 1995; Bearzi et al., 1997; Miokovic et al., 1997; Bearzi et al., 1999; Pribanić et al., 2000; Bearzi et al., 2008; Bearzi et al., 2009). Conversely, the southern Adriatic region, and the eastern and southern Mediterranean Sea has limited baseline data, with dedicated research efforts starting in the early 2000s (Bearzi et al., 2009; Baş et al., 2017a; Akkaya et al., 2020).
Common bottlenose dolphins (Tursiops truncatus) are the most widely studied cetacean species, both visually and acoustically (Hill and Lackups, 2010), throughout the Mediterranean Sea due to their highly coastal distribution (Bearzi et al., 2008). According to the IUCN Red List, bottlenose dolphins have experienced a 30% decline in their population size since the 1940s in the Mediterranean Sea, while Adriatic populations declined by almost 50% in the latter half of the 20th century (Bearzi et al., 2004; Sackl et al., 2007; Bearzi et al., 2008; Bearzi et al., 2012). Due to the continuous decline in populations, this species was classified as “vulnerable” in the IUCN Red List in 2009 with its classification recently upgraded to “least concern” in 2021 (Bearzi et al., 2021). Basin-wide population estimates of bottlenose dolphins have historically relied on a handful of highly surveyed areas (e.g. Bearzi et al., 2008). Whilst an effort was made to address the disparities in survey effort between regions during the ACCOBAMS Survey Initiative (ACCOBAMS, 2021), smaller countries received only two or three transects and population estimates conducted during a single day of flying rendered their population sizes virtually unknown. Therefore, data-deficient regions (which may be at threat from uncontrolled and unregulated pressure) have the potential to considerably affect basin-wide estimates and year-round dedicated research at local scales remain integral to an accurate understanding of the status of the species in question (Awbery et al., 2022).
The majority of the effort in the Adriatic and Ionian Sea focuses on behavioural activity, individual identification, threat assessment and movement patterns (Bearzi et al., 1997; Fortuna et al., 2011; Baş et al., 2017a; Baş et al., 2017b; Awbery et al., 2019; Akkaya et al., 2021). Based on previous knowledge, bottlenose dolphins are generally known as a resident species with limited movement (Genov et al., 2009; Bearzi et al., 2016). Contrarily, a recent study documented an adult male bottlenose dolphin travelling between the Northern Adriatic Sea, passing through the Ionian and Tyrrhenian Sea, and finally reaching the Ligurian Sea with an estimated minimum distance of 1251 km across all three seas (Genov et al., 2022).
While traditional visual data collection can reveal critical information on species as highlighted in the example of an indivduals’ movement over a wider area as ever previously thought, it is limited by the duration that the species spends at the surface, and additionally, environmental conditions and daylight hours (Mellinger and Barlow, 2003; Mellinger et al., 2007). Considering that cetaceans spend the majority of their time under the surface and that sound emission is their primary source of information acquisition, acoustic monitoring as a research tool, increases our knowledge of underwater species and amplifies the potential of conservation strategies (Van Parijs et al., 2009; Davis et al., 2017). The marine soundscape has changed drastically since the beginning of the industrial revolution as humans have either deliberately added sound to the environment (e.g. when describing the bottom or sub-bottom of the seabed; Hildebrand, 2009; Estabrook et al., 2016) or as a by-product (e.g. the intensification of marine traffic; Duarte et al., 2021). As sound propagates further than light or chemicals in the marine environment, many marine animals have evolved to be sensitive to sound. Thus, the cumulative increase in anthropogenic sound may elicit many short and long-term effects, from behavioural and habitat alterations to area avoidance, to changes in population dynamics and even physical injuries and mortality (La Manna et al., 2013; Domit et al., 2016; Baş et al., 2017b; Fruet et al., 2017; Caruso et al., 2020).
Bottlenose dolphins are highly vocal animals with great plasticity and complexity in their vocal repertoire (Luís et al., 2021). The sounds emitted play a fundamental role in their social interactions, individual recognition, group coordination, foraging success, and recognition of an individual’s surroundings (Lind et al., 1996; Rendall et al., 1996; Janik and Slater, 1998; Azzolin et al., 2017; MacFarlane et al., 2017; La Manna et al., 2020). The acoustic repertoire of bottlenose dolphins includes echolocation clicks (broadband click trains), burst-pulsed sounds (closely spaced broadband click trains) and frequency modulated whistles (narrowband tonal sounds) (Caldwell et al., 1990; Janik, 2009; Herzing, 2014; La Manna et al., 2017; Luís et al., 2021; Pace et al., 2022). Echolocation clicks are known to be used primarily for navigation and foraging while whistles and pulsed sounds are considered to be used for individual recognition, social maintenance, group coordination, communication, as well as foraging activity (Au, 1993; Branstetter et al., 2012; Janik et al., 2012; MacFarlane et al., 2017). The vocal repertoires of dolphins are also known to vary significantly between populations at macro-and-micro geographic scales (Hawkins, 2010; Papale et al., 2013; La Manna et al., 2020). Within the Mediterranean Sea, acoustic studies on dolphins have mostly been assessed by categorisation of whistle type characteristics (Díaz López, 2011; La Manna et al., 2017; La Manna et al., 2020; Corrias et al., 2021; Terranova et al., 2021; La Manna et al., 2022; Terranova et al., 2022; Pace et al., 2022). These studies have mainly focused on the identification of species presence, geographical variation in vocalisation types, the impact of boat traffic, dolphin-fishery interactions, and whistle characteristics with bottlenose dolphins, being by far the most frequently studied species (Connor and Smolker, 1996; Boisseau, 2005; Díaz López and Shirai, 2010; La Manna et al., 2017).
This current study is the first attempt to understand the whistle characteristics of bottlenose dolphins in the Southern Adriatic Sea and provides additional information for the Northern Ionian Sea. This study assesses variation in their acoustic behaviour by examining the whistle characteristics of bottlenose dolphins and aims to evaluate the potential similarities or dissimilarities in regional repertoires between the two survey locations. The assessment of this baseline data aims to deepen our understanding of the vocal repertoire of bottlenose dolphins while reviewing the potential effect of geographical differences on their vocal behaviour.
2 Material and methods
The acoustic data was collected in two nearby regions: the Gulf of Taranto, Italy between 11.06.2018 and 20.06.2021 and Boka Bay, Montenegro between 01.04.2020 and 16.07.2022. The selected study locations are within a 400 km straight line distance from each other, and the presented data represents only part of a long-term research effort from each region.
2.1 Study locations
The Gulf of Taranto (Northern Ionian Sea, Central-Eastern Mediterranean Sea) covers an area of approximately 14000 km2 from Santa Maria di Leuca to Punta Alice reaching depths of over 2000 m (Figure 1). It is characterised by a narrow continental shelf with a steep slope cut by several channels in the western sector, while the eastern one shows descending terraces toward the submarine canyon system of Taranto Valley with no clear bathymetric connection to a major river system (Capezzuto et al., 2010; Pinardi et al., 2016; Carlucci et al., 2017). Both the complex bottom topography and the mix of environmental conditions makes this area suitable for the presence of different cetacean species such as the common bottlenose dolphin, usually occurring within the continental shelf, the striped dolphin (Stenella coeruleoalba), short-beaked common dolphin (Delphinus delphis), Risso’s dolphin (Grampus griseus), Cuvier’s beaked whale (Ziphius cavirostris), sperm whale (Physeter macrocephalus) and fin whale (Balaenoptera physalus) mainly distributed on the continental slope and offshore waters (Natoli et al., 2008; Carlucci et al., 2016; Carlucci et al., 2018a; Carlucci et al., 2018b; Carlucci et al., 2018c; Bellomo et al., 2019; Santacesaria et al., 2019; Carlucci et al., 2020a; Carlucci et al., 2020b; Azzolin et al., 2020; Papale et al., 2020; Cipriano et al., 2022). Moreover, the basin includes valuable habitats from a conservation perspective such as the Santa Maria di Leuca cold-water coral province and the Amendolara shoal (Maiorano et al., 2022). The survey area investigated in this study includes the head of the Taranto Valley canyon system covering 960 km2 including waters up to approximately 1200 m (Figure 1).
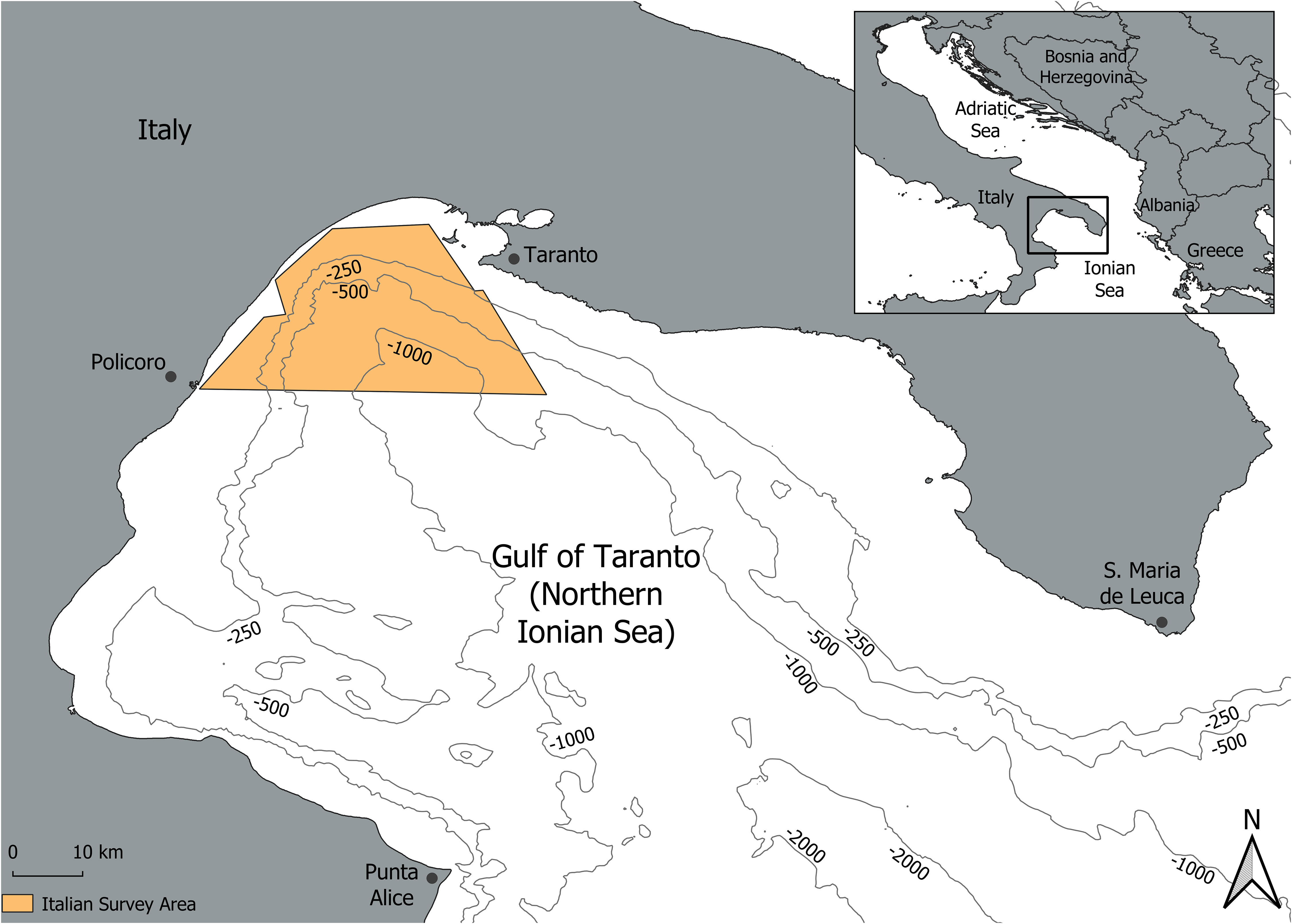
Figure 1 Map of the study location in the Taranto Gulf, Northern Ionian Sea, Central-Eastern Mediterranean Sea.
The semi-enclosed region of Boka Bay belongs to the Adriatic-Ionian subregion and is situated in the south-eastern Adriatic Sea forming the northernmost part of the Montenegro coastline. The bay extends 106 km and contains a surface area of 87.3 km² and consists of four smaller coves: the Bay of Kotor, the Bay of Risan, the Bay of Tivat, and the Bay of Herceg Novi. The sea surface temperature is highly influenced by the precipitation rate, and the currents are stronger during the colder months and spring, while they are weaker in summer months (Peraš et al., 2022). The general depth of the Boka Bay is below 50m (Figure 2). The specific environmental characteristics make the bay a unique area with ecological conditions that differ from the rest of the Adriatic coast (MAP-UNEP, 2015; Ðurovic et al., 2016). The bottlenose dolphins are the only commonly sighted cetacean species in this area (Ðurovic et al., 2016) and the survey area covers the entirety of Boka Bay (Figure 2).
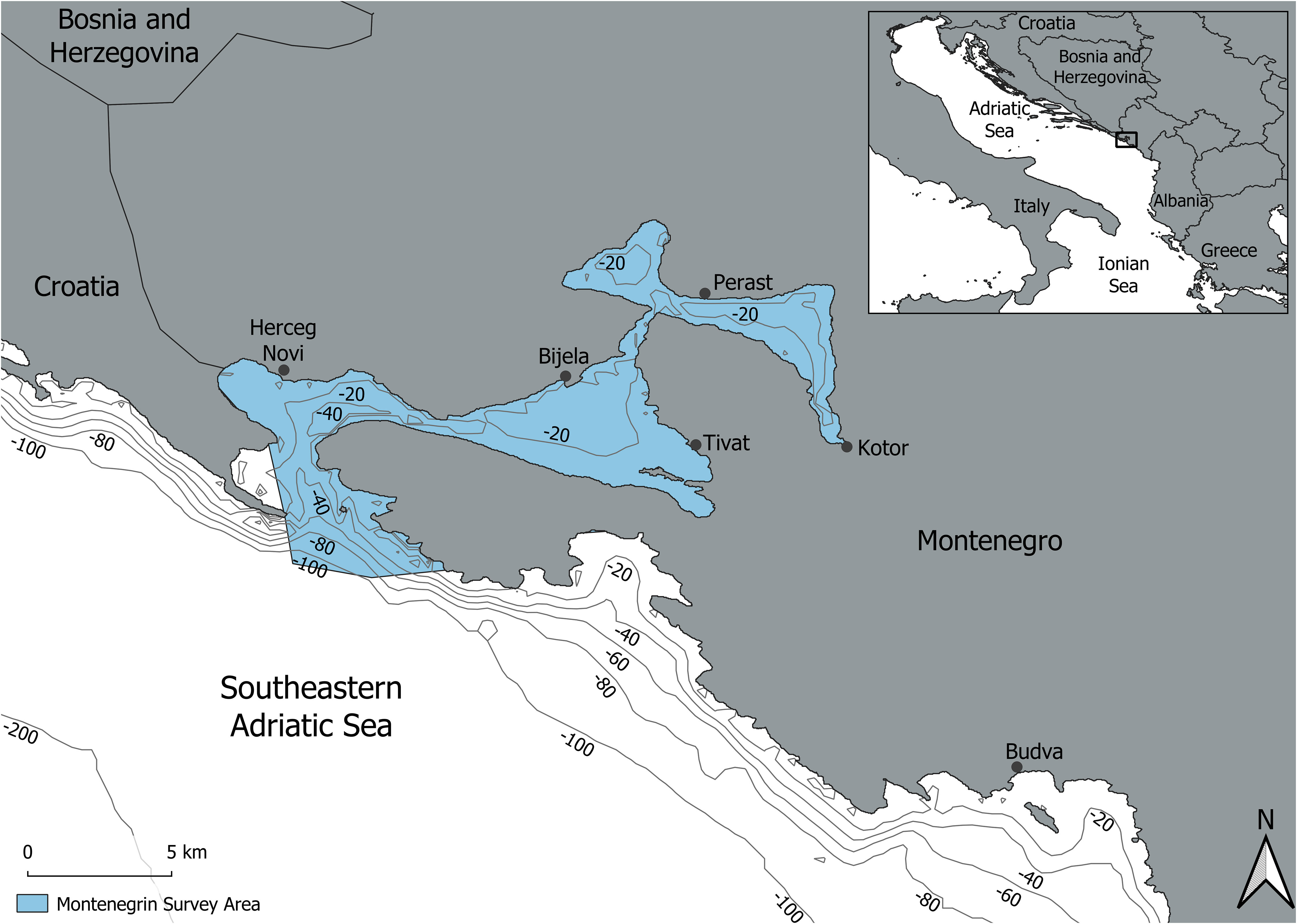
Figure 2 Map of the study location in the Boka Bay, South Adriatic Sea, Central-Eastern Mediterranean Sea.
2.2 Data collection
Acoustic data was collected during dedicated boat-based surveys, following random routes with stratified effort to increase the chance of sighting bottlenose dolphins within the study locations. The survey platforms were a 12 m catamaran in Italy while a 12 m rigid-hull inflatable boat (rib) with 500 hp inboard engine was used in the Montenegrin region. The geographic position of the survey boat was recorded every 30 s using a GNSS (global navigation satellite system) receiver. In each study location, the vessel followed an average speed of 4 knots during the surveys.
Visual surveys took place only during daylight hours with favourable weather conditions (Beaufort ≤3). When a focal group was visually encountered, the boat speed was reduced to idle, while ensuring the path of the focal group was not blocked and an observational distance of between 50 m and 400 m was maintained. The hydrophone was dropped within the 400 m radius of the focal group when the boat was either idling or the engine was turned off and returned to deck on completion of a focal group sighting. The focal group was defined as a minimum of two individuals with a maximum distance of 100m from the nearest individual. The position of the focal group relative to the position of the hydrophone remained varied during recordings between study locations. If a group was not sighted or heard for more than 20 min, the resighting would be considered a new group as it was not possible to ascertain if individuals belonged to the previous focal group. The focal group was followed for a maximum of 30 minutes in order to reduce any potential negative impact of the research boat presence.
The drop-down acoustic system varied between the two study locations. A pre-amplified omnidirectional hydrophone (Colmar GP0190) with a working band of 5 Hz to 170 kHz, a sensitivity of -175 ± 5 dB re 1V/µPa and a flat response of -171 dB re 1V/µPa under 12 kHz was employed in Italy. It was attached to a 20 m cable and connected to a laptop for data recording. A custom-built omnidirectional hydrophone (Vanishing Point) was used in Montenegro. The hydrophone recorded between 0 and 48 kHz and had a sensitivity to 201 dB without a preamplifier. The hydrophone was attached to a 20 m cable with a TASCAM DR-40x Linear PCM acoustic recorder.
2.3 Data analysis
The acoustic analysis was carried out using RAVEN Pro 1.6 software (Conservation Bioacoustics at the Cornell Lab of Ornithology, 2022). A 512-point Hamming window Brightness 48 and contrast 60 was used to visualize the dolphin vocalisations in a spectrogram. The time axis was kept at 5 seconds and the frequency axis between 0 to 48 kHz and any possible whistles extending the 48 kHz limit were discarded. Whistles were manually cropped from the spectrogram and were classified as good, medium, and poor quality based on their visual and aural patterns in the spectrogram. Additionally, the background of the selected whistle was scored between 0 to 4 (0 = No clear noise, 1 = There is a single click or whistle in the background, 2 = There is a click and whistle in the background, 3 = There are either multiple clicks and/or multiple whistles, 4 = The background is noisy with multiple clicks and whistles). Harmonics were also identified during the selection process. From the selected whistles, eleven parameters were measured: start frequency, end frequency, peak frequency, central frequency, high frequency, low frequency, delta frequency (change in frequency), whistle duration, number of inflection points, and whistle type (Table 1; Figure 3). The contour of each whistle was determined by visual inspection and was categorised into the following whistle types based on previously accepted contour shape categories designed by Hickey et al. (2009) (Figure 4). Annotation of whistles in Raven software was conducted independently by two experienced acousticians. Annotations were compared by a third researcher who chose the annotation that best fit the whistle in order to minimise human error when making whistle selections and during contour identification. It was not possible to assign whistles to individual dolphins and thus not possible to know whether whistles were produced by multiple individuals or just a single individual other than when whistles overlapped.
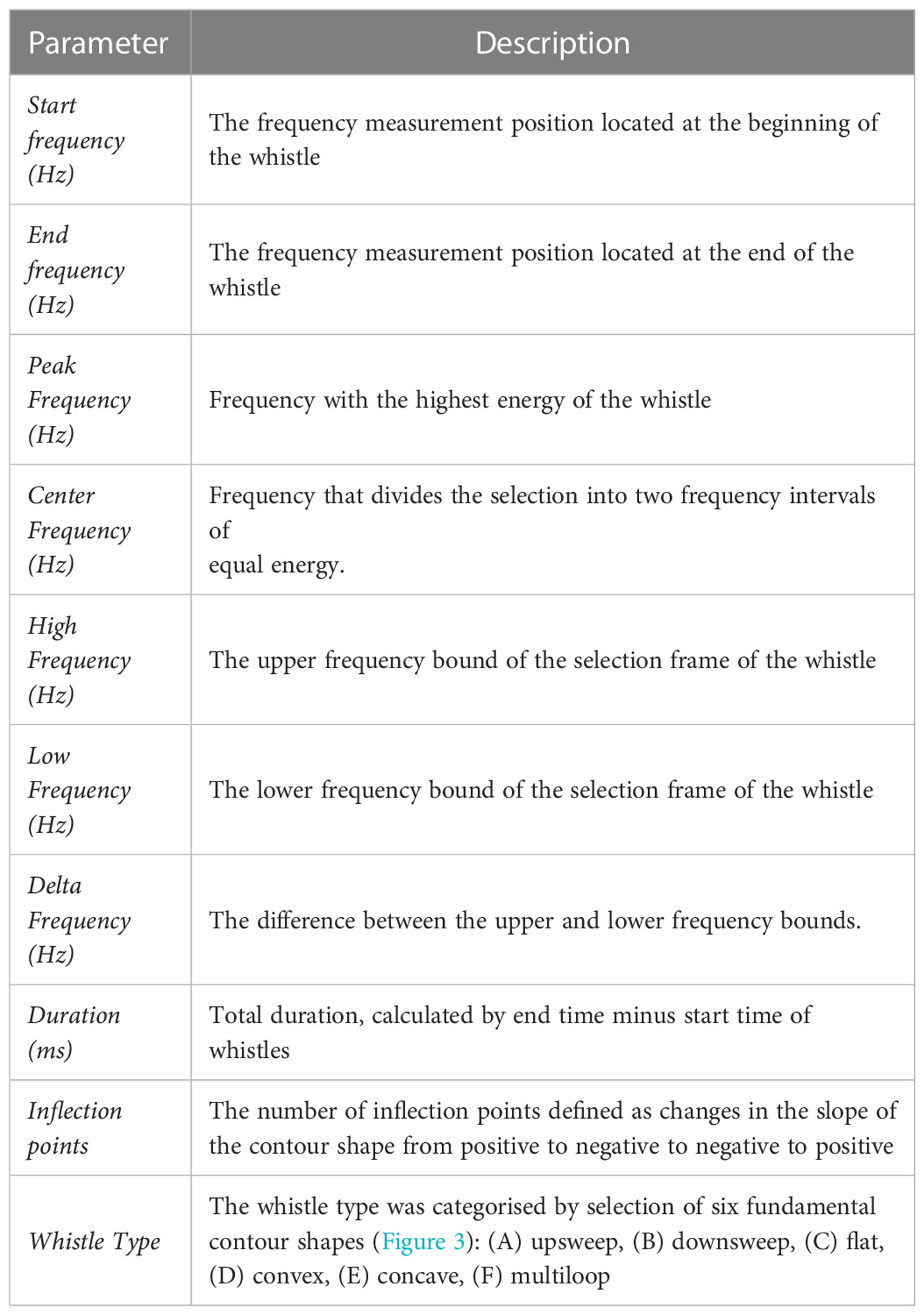
Table 1 Whistle parameters measured manually within spectrograms using Raven Pro 1.6.
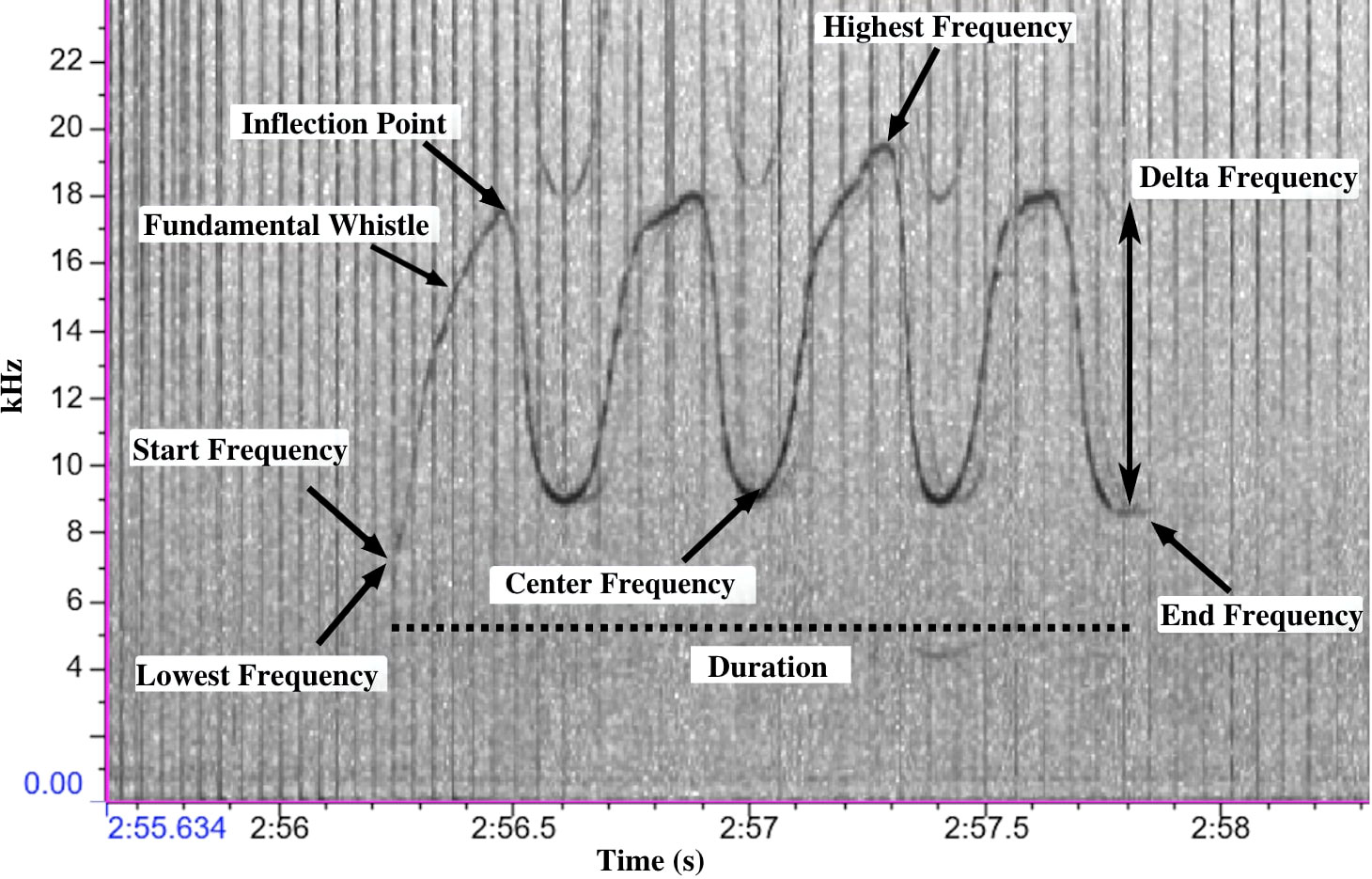
Figure 3 A spectrogram of whistle type F displaying the positions of the whistle parameters measured manually in Raven Pro 1.6.
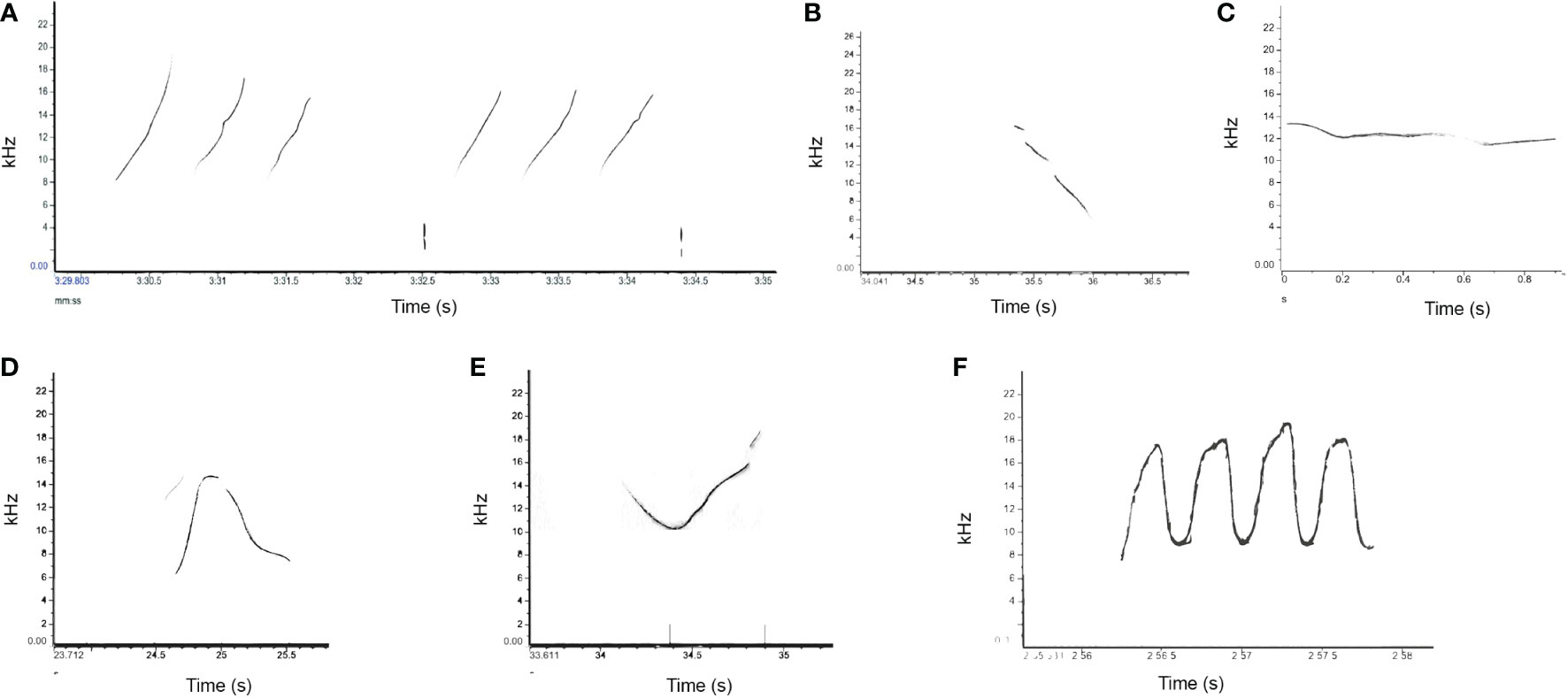
Figure 4 Examples of each whistle types: (A) whistle type A (upsweep), (B) whistle type B (downsweep), (C) whistle type C (flat), (D) whistle type D (convex), (E) whistle type E (concave), and (F) whistle type F (multiloop), recorded in Taranto Gulf and Boka Bay.
2.4 Statistical analysis
Individual whistles were evaluated according to their whistle quality and background noise before they were embedded into the analysis. Poor quality whistles as well as any whistle quality with a background score of 3 or more were discarded from further analysis. Harmonics were only considered if it was possible to isolate the fundamental frequency (lowest frequency component of the harmonic) and if the fundamental frequency did not overlap with any other whistle in the selected frame.
Summary statistics (minimum, maximum, mean, median, standard deviation, standard error, and the coefficient of variation) for each whistle variable of each whistle type (Figure 4) at each study location (Figures 1, 2). Before proceeding with the statistical analysis, histograms and quantile-quantile plots of each whistle parameter were tested for normality with the addition of the Shapiro-Wilks normality tests.
Principal Component Analysis (PCA) was conducted to reduce the dimensionality of the whistle variables while retaining as much of the information within the data as possible. The data for each parameter prior to the PCA analyses were centred to zero and with a standard deviation of one. The loading scores of each component were analysed to assess which variables made the greatest contribution to that component. To select a cut off point for the number of necessary principal components, the rule of 80% of explained variance of the dataset was followed and only these components were used in the subsequent statistical analysis (Jung et al., 2018).
Due to the non-normal distribution of most of the whistle variables, non-parametric tests were used for any further analyses. Permutational multivariate analysis of variance (PERMANOVA) was conducted to test if the selected principal components varied between the study locations. PERMANOVA was also used to check if the principal components significantly varied between the different whistle types. PERMANOVA conveys whether there are differences between all whistle types but does not calculate differences between pairs of whistle types, and thus, multilevel pairwise post hoc tests were employed using the pairwise Adonis package in R Studio (Martinez Arbizu, 2020). P-values for all PERMANOVA tests were calculated based on Euclidean distances using 999 permutations to estimate the probability of group differences and Bonferroni adjustments were made to the significance level to reduce the chances of type I errors when multiple pairwise tests are conducted. Finally, a chi-squared test was performed to identify if whistle type showed significant variation between the study locations. All the statistical tests were performed in RStudio (Version 4.1.2).
3 Results
The overall survey effort consisted of 346 days in the Taranto Gulf between 11.06.2018 and 20.06.2021 and 39 days in Boka Bay between 01.04.2020 and 16.07.2022. Bottlenose dolphin presence was recorded during 82 days in Taranto Gulf, of which the majority of survey days were conducted with visual surveys and an onboard hydrophone resulted in obtaining 10 acoustic recordings from bottlenose dolphins. The species was present for 21 days in Boka Bay, of which 14 days resulted in the acquisition of acoustic data. The acoustic data could only be collected when the vocalisations of the focal group were within detection range of the hydrophone. Regarding the total duration of recordings, Taranto Gulf had 01:44:38 hours of recording, where whistles were recorded during 01:04:16 hours. Whereas, Boka Bay had 08:08:05 hours of recording, of which whistles were present in 03:28:12 hours. Analysis of these audio files revealed 433 whistle detections in Taranto Gulf and 546 whistle detections in Boka Bay. Harmonics were present in 40% and 32% of tonal sounds in Taranto Gulf and Boka Bay, respectively. After evaluating the quality of each whistle and its background, a total of 423 whistles qualified as satisfactory for further analysis of which 238 whistles came from the Taranto Gulf and 185 whistles from Boka Bay.
Principal Component Analysis was performed on nine whistle variables. The first three principal components explained 87% of the variance with each component having eigenvalues greater than one (Table 2). Therefore, the first three components were used for further analysis while the following six components were considered redundant and were not used in subsequent analysis. Principal Component 1 explained 51.3% of the variance in the dataset with the center frequency and high frequency having the highest loading scores, suggesting that they are the main drivers of variation in component 1. Principal component 2 formed 24.1% of the variance and the duration of the whistle and the number of inflection points had the highest loading scores in component 2. Lastly, component 3 comprised 11.5% of the variance with the highest loading score attained by the number of inflection points (Table 2; Figure 5).
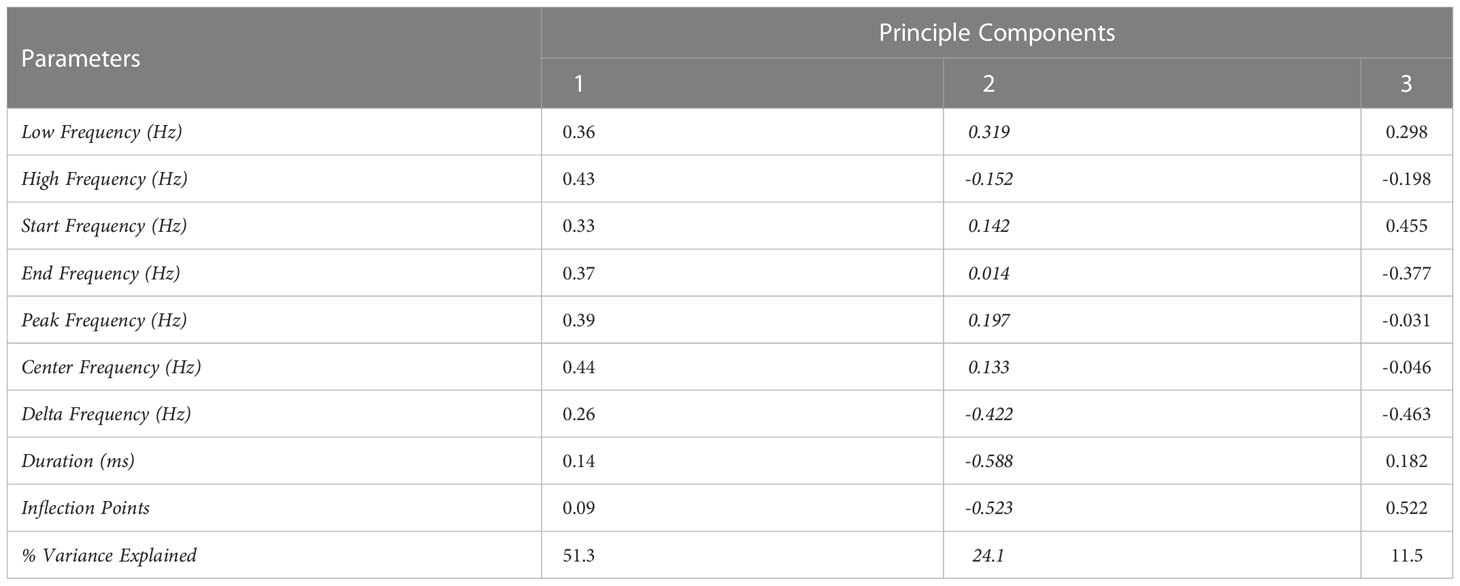
Table 2 The loading scores and the proportion of explained variance for each principal component for bottlenose dolphin whistles recorded in the Taranto Gulf and Boka Bay.
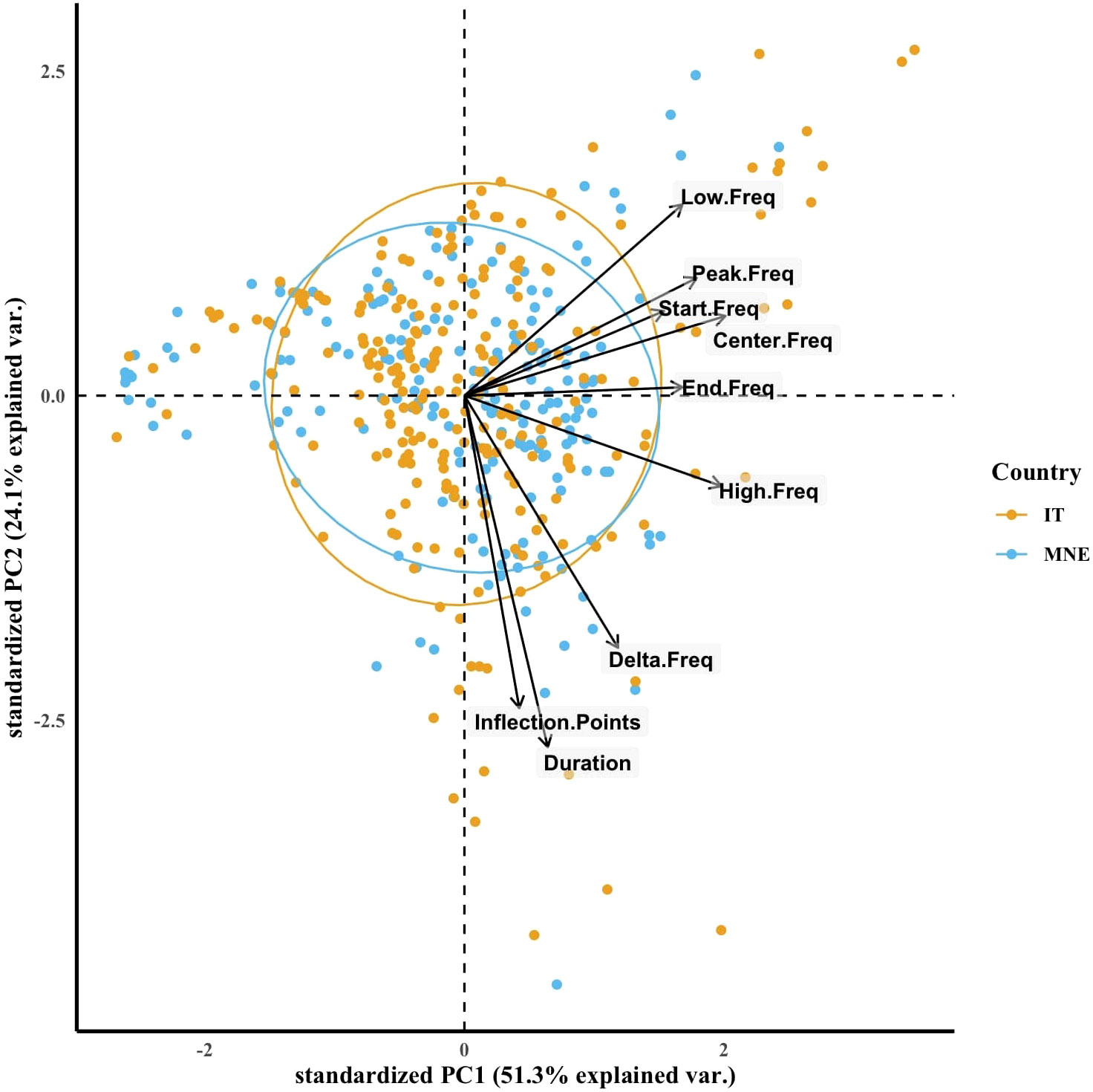
Figure 5 PCA correlation biplot of the whistle parameters grouped by the study location (IT = Taranto Gulf, MNE = Boka Bay) with 95% confidence interval ellipses.
Whistle parameters did not show significant differences between Taranto Gulf and Boka Bay (F=0.11, df=1, p=0.94). Detected whistles fell in the range of 659 Hz and 26,122 Hz in Taranto Gulf, while they were between 850 Hz and 24,000 Hz in Boka Bay. The peak frequency had a mean of 10.121 ± 282 Hz in Taranto Gulf and 9,310 ± 256 Hz in Boka Bay. The average duration of the detected whistles was 570 and 500 milliseconds with a maximum whistle duration of 3,160 and 3,530 milliseconds for Taranto Gulf and Boka Bay respectively. The maximum number of inflection points was 17 (mean of two) in Taranto Gulf and 12 inflection points (mean of one) in Montenegro (Table 3).
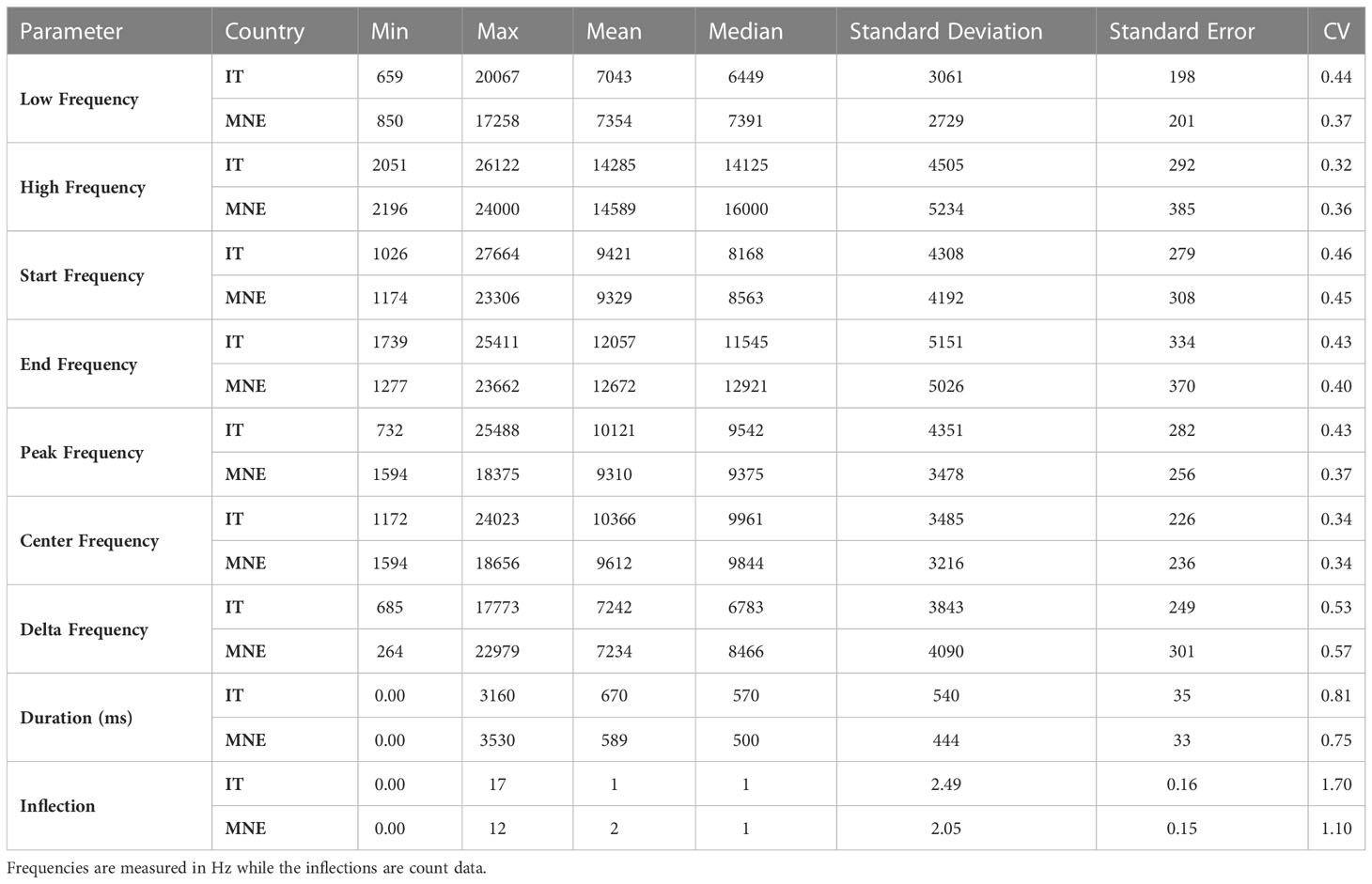
Table 3 Summary statistics of whistles produced in Taranto Gulf, Italy (denoted by IT) and Boka Bay, Montenegro (denoted by MNE).
In addition, whistle variables showed significant variation between whistle types (F=24.84, df=5, p=0.001). The whistle variables of whistle type F showed significant variation from the other whistle types. Additionally, the whistle variables of whistle type C were also significantly different from the other whistle types, except whistle type B. Conversely, the whistle variables for whistle type E showed similar variation between each of the whistle types, except for whistle type C and F (Table 4). Further, the whistles with highest frequencies were produced during whistle type F in both Taranto Gulf (median of 17,104 Hz) and Boka Bay (median of 17,832 Hz). The whistles with the highest peak frequencies were produced for whistle type D in both Taranto Gulf (median of 10,453 Hz) and in Boka Bay (median of 11,063 Hz). However, the whistle types with the highest central frequency showed variation between the study locations, with whistle types E and F possessing the highest medians (both of 10,986 Hz) in Taranto Gulf whilst whistle type D had the highest median (10,969 Hz) in Boka Bay. Finally, the duration of the whistle was longest for whistle type F in Taranto Gulf (median of 1,650 milliseconds) and Boka Bay (median of 1,160 milliseconds) (Table 5).
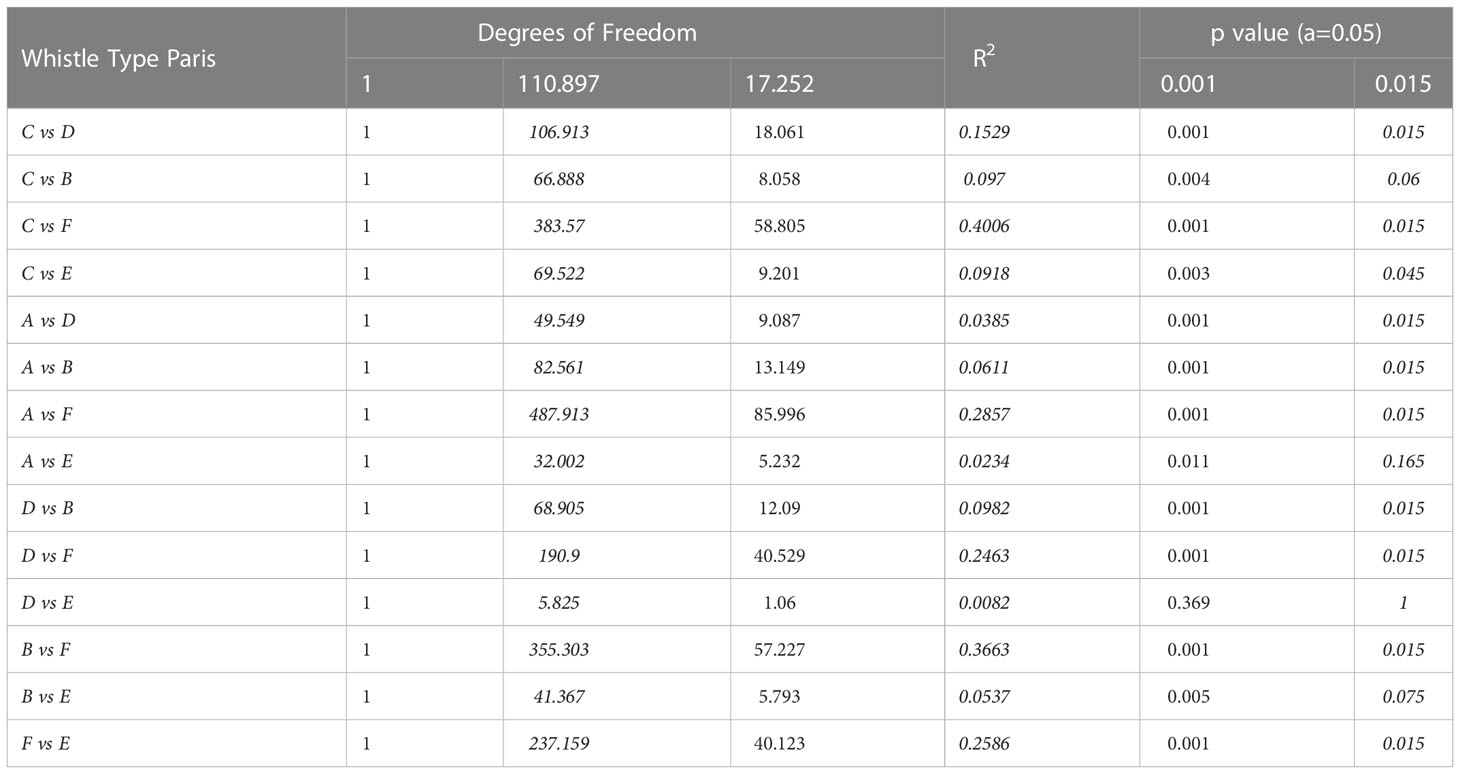
Table 4 Post hoc pairwise Adonis testing of the whistle types based on euclidean distances with 999 permutations and Bonferroni adjustments to the significance level.
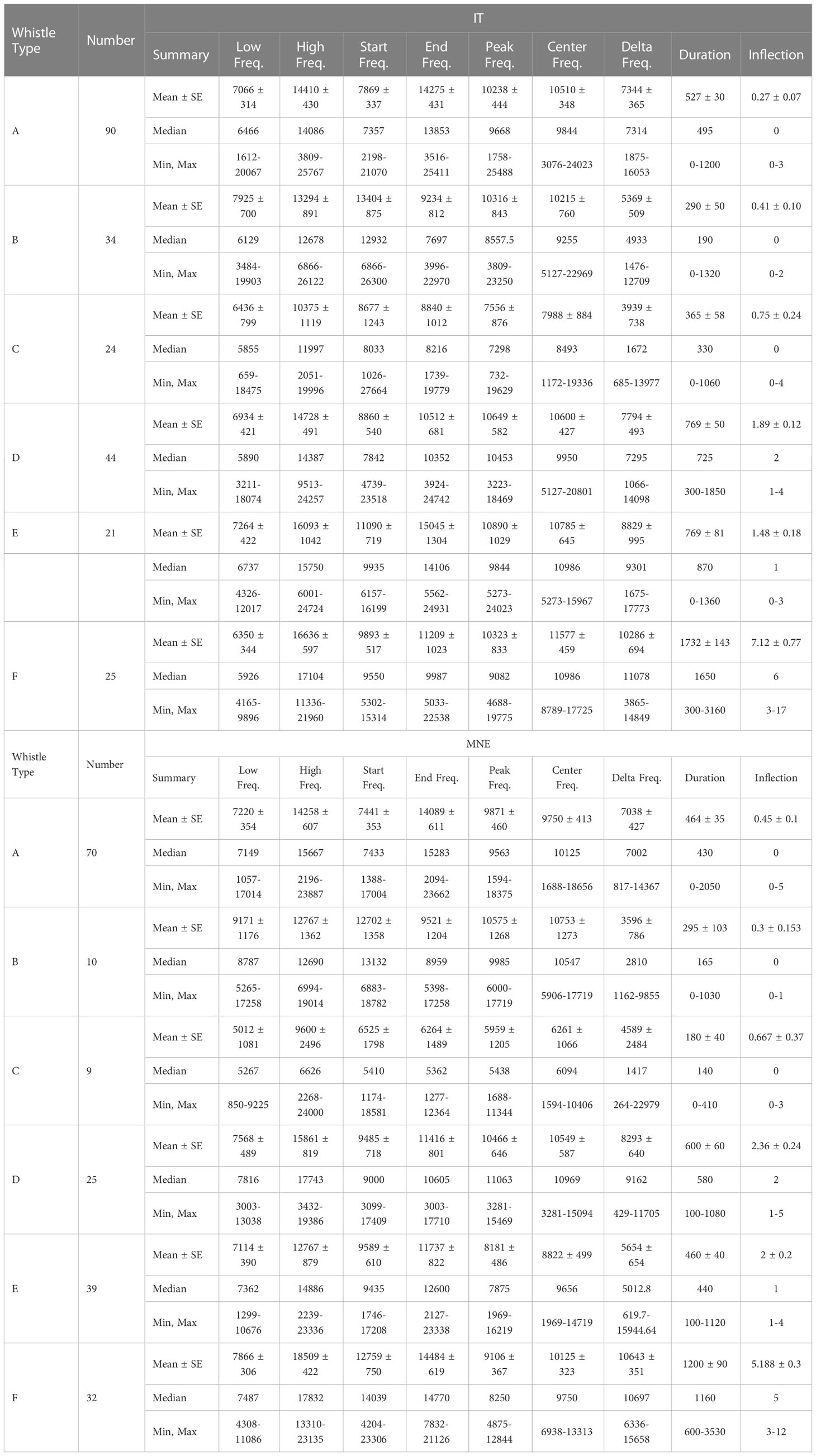
Table 5 Summary statistics of detected whistle types in the Taranto Gulf and in the Boka Bay.
When variation in whistle types between the study locations was assessed, there was a significant difference between Taranto Gulf and Boka Bay (x2 = 27.7, df=5, p=0.0004). Whistle type A was the most dominant whistle in each location and formed almost 40% of the overall produced whistles both for Taranto Gulf and Boka Bay. In contrast, the least detected whistles were type E for Taranto Gulf and type B and C whistles for Boka Bay (Figure 6).
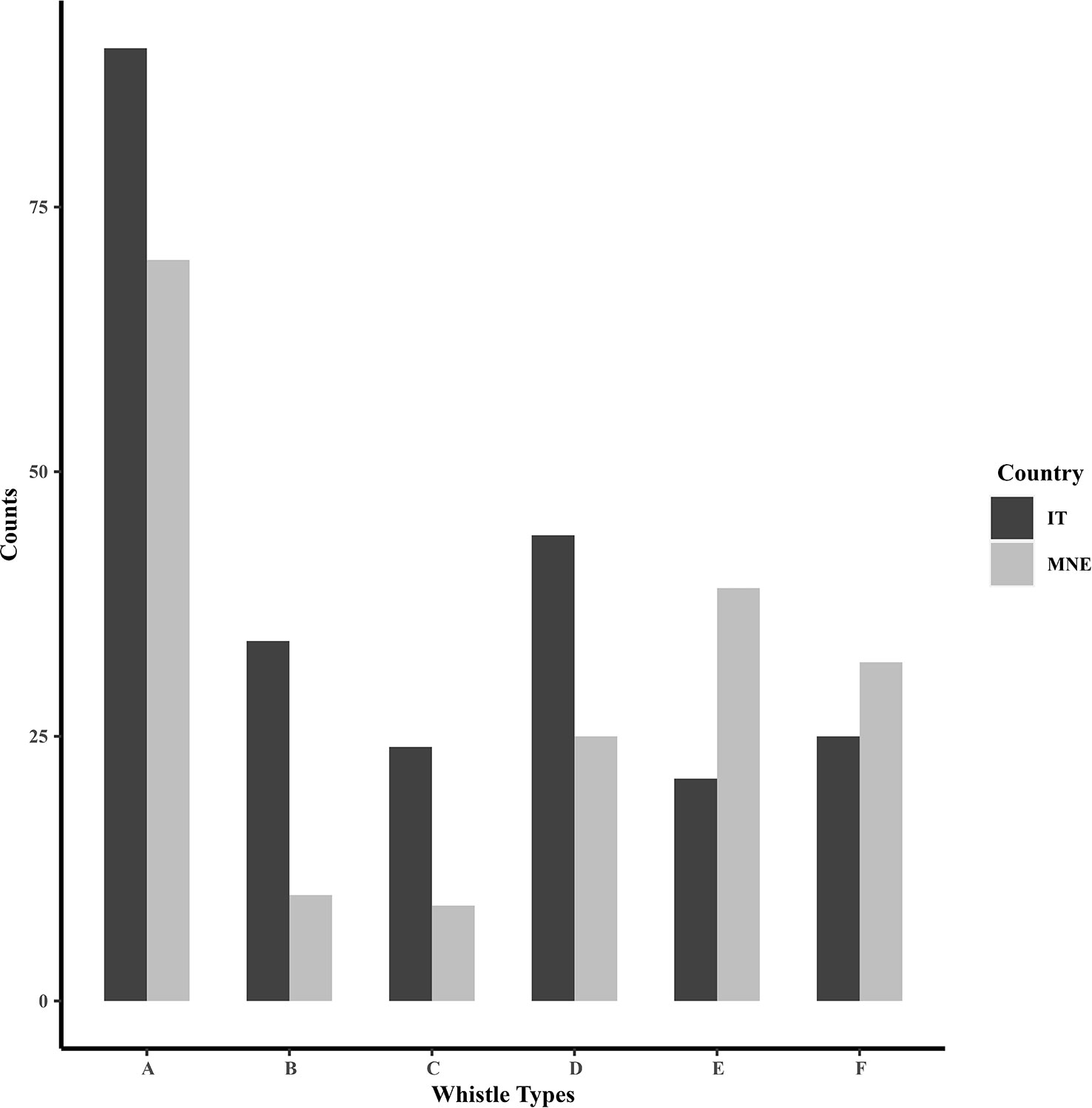
Figure 6 Counts of detected whistle types in the Taranto Gulf (IT) and Boka Bay (MNE).
4 Discussion
The results of this current study represent the first acoustic analysis of bottlenose dolphins in Montenegrin waters as well as providing additional information on the variation of whistle characteristics of two geographically separated populations residing in two neighbouring bodies of water: the Southern Adriatic and Northern Ionian Sea. The whistle characteristics of bottlenose dolphins reported here are consistent with those reported in the Mediterranean Sea (Rendell et al., 1999; Gannier et al., 2010; Díaz López, 2011; Papale et al., 2014a; Bolger et al., 2017; La Manna et al., 2019; Rako-Gospić et al., 2020; Papale et al., 2021) and elsewhere (Conner, 1982; Boisseau, 2005; May-Collado and Wartzok, 2008; Baron et al, 2008; Hawkins, 2010). The mean peak frequency was reported at 10kHz, with a frequency ranging between a mean low frequency of 7kHz and a mean maximum frequency of 14 kHz, with a mean duration of 500 ms in each of the locations. Although the frequency range, duration, and inflection points are similar to those reported for other populations, the mean peak frequency range was slightly higher in Croatian and other Italian studies with peak frequencies ranging from 13 to 15kHz (Díaz López, 2011; La Manna et al., 2013; La Manna et al., 2020). Previous studies have highlighted that bottlenose dolphins generally emit whistles ranging between 1 and 20 kHz, yet low frequency whistles reaching 200 Hz have also been detected (Richardson et al., 1995; Ding et al., 1995; Schultz et al., 1995; Lammers et al., 2003; Herzing, 2015). This study also recorded whistles below 1 kHz in each location, although these occurred rarely. On the other hand, the highest recorded fundamental whistles reached just over 25 kHz in each location with harmonics potentially extending beyond the 48 kHz window displayed in the analysed spectrograms, although these were discarded from the current analysis. Additionally, over 40% of whistles contained harmonics in Taranto Gulf while the ratio reached 30% in Boka Bay. Previously, it was noted that the occurrence of harmonics can be linked to behavioural activity and/or group composition (Henderson et al., 2011; La Manna et al., 2019). Henderson et al. (2011) reported that more complex whistles (harmonics) with increased number and duration were emitted when common dolphins (Delphinus sp.) were travelling, whilst whistles were less complex if they were engaged in milling activity. Therefore, it has been suggested that dolphins are likely to use these harmonics for directionality in order to maintain group cohesion and to convey changes in the orientation of the group (Miller, 2002; Rasmussen et al., 2006; Branstetter et al., 2013). It has been further suggested that increased underwater noise in the close proximity of boats may result in the production of harmonics, with the number of harmonic vocalisations having been observed to increase with calf presence and a larger group size (La Manna et al., 2019). However, it is important to consider that the production of harmonics may vary among species (Gannier et al., 2020).
It is important to understand the role of harmonics, specifically considering their high occurrence in each location during the current study. Although this study focuses on geographical differences of whistle variables between study locations, previous studies indicated that 50% of focal bottlenose dolphin groups in Montenegro were observed with the presence of subadults, with dolphins spending 70% of their time travelling, while milling and socialising behaviour only formed 10% or less of the total behavioural budget (Clarkson et al., 2020; Akkaya et al., 2021; Rudd et al., 2022). Above all, both of the locations have a significant amount of marine traffic; with approximately 4,130 vessels in the port of Taranto per year (Shipnext.com). Moreover, Boka Bay has faced a considerable annual increase in marine traffic between 2020-2021 and 2021-2022 respectively. A total of 579 vessels arrived in the Port of Kotor during the entirety of 2021, whilst 692 port calls were made within the first nine months of 2022 alone. It is critical to mention that both locations are utilised by large, environmentally disturbing marine vessels. Large merchant ships encompass approximately 57% of marine traffic in the port of Taranto (Marinetraffic.com), and an estimated 2,000 cruise ships were reported to visit the port of Kotor between 2013 and 2018 (Balkaninsight.com). It is plausible that the aforementioned behaviours, subadult presence, marine traffic presence, or the cumulative effects of these may explain the high occurrence of harmonics, but there is a clear need for future simultaneous visual and acoustic studies to understand the possible relationships influencing the production of harmonics by potentially extending this study to understand the potential role of behavioural activity as well as considering the importance of the direction of the hydrophone in relation to focal group the rate of frequencythat harnonics are recorded
When the whistle characteristics were investigated, the results of principal component analysis indicated that the centre frequency, high frequency, whistle duration, and the number of inflection points’ loading scores were the highest across principal components 1, 2 and 3. These acoustic features were also identified as the main contributors to whistle variation in the Mediterranean Sea by previous studies (Díaz López, 2011; Papale et al., 2014b; La Manna et al, 2020). Therefore, it is possible that these acoustic features play a critical role on the emitted whistles basin-wide, without a clear difference between groups or populations. Bottlenose dolphins are likely to adjust these features depending on the behavioural context and it is believed that these whistles carry information such as identification of an individual, group cohesion, stress level, the presence of food, or danger (Wells, 1991; Wang et al., 1995; Janik and Slater, 1998; Buckstaff, 2004; Esch et al., 2009a; Díaz López, 2011; Henderson et al., 2011). Previously, it has also been stated that an increased number of inflection points on a whistle could indicate the complexity of its contextuality (Steiner, 1981; Wang et al., 1995; Rendell et al., 1999; Azevedo et al., 2007; May-Collado and Wartzok, 2008; Amorim et al., 2016) and is directly linked to the duration of the whistle. It is important to take into account that multiloop whistles, which hold the highest inflection numbers, were responsible for only 11% of whistle types in Taranto Gulf and 17% of whistles in Boka Bay. Meanwhile, harmonics were responsible for almost half of the produced whistles of bottlenose dolphins in each location. The presence of harmonics vs multiloop whistles with the consideration of behavioural and environmental variation should be investigated to understand the role and differences of each acoustic feature.
The emitted whistle type did reveal considerable variation between Taranto Gulf and Boka Bay. While the dominant whistle type was the upsweep whistle (whistle type A) in each location comprising 40% of the whistles, the least emitted whistle was concave (whistle type E) in Taranto Gulf and flat (whistle type C) in Boka Bay. A neighbouring study revealed an absence of flat whistles in Croatia (Rako-Gospić et al., 2020), while the same whistle type was also identified as the least prominent whistle of bottlenose dolphins elsewhere in the Mediterranean Sea and its adjacent waters (Díaz López and Shirai, 2010; Bolger et al., 2017; Rako-Gospić et al., 2020). In the Mediterranean Sea, the dominant whistle type of bottlenose dolphins was reported to be either upsweep or multiloop whistles (Díaz López and Shirai, 2010; Gannier et al., 2010; Rako-Gospić et al., 2020). Adjacent Atlantic waters of the Mediterranean Sea also documented multiloop whistles as being the most produced whistle type (Bolger et al., 2017). Multiloop whistles were reported in moderate numbers at both locations with little difference between the two sites. Terranova et al. (2021) and Esch et al. (2009b) suggest dolphins often emit signature whistles in the form of a continuous connected multiloop whistle (an identical repeated unit in time without intervals) (Janik, 1999; Sayigh et al., 2007). Therefore, it may be likely that some of these multiloop whistles were representing signature whistles. Therefore, future studies should focus on the identification of signature whistles in the study locations. Previous studies suggested that produced whistle types are most likely to be linked with the behavioural activity that the dolphins are engaged in at the time, as well as the presence of marine traffic in the area (Quick and Janik, 2008; Díaz López and Shirai, 2010; Henderson et al., 2011; Jones et al., 2019; Rako-Gospić et al., 2020). Recently, Rako-Gospić et al. (2020) reported that whistle type does show significant variation during foraging activities, but also in the presence of trawlers or motorboats which also play a critical role in the dominant whistle type. Therefore, the number of anthropogenic activities that take place in the region should be considered in order to understand variation of whistle types under different behavioural activities and vice versa.
This current study represents the first attempt to understand whistle variability of bottlenose dolphins from the data deficient waters of the Southern Adriatic, while investigating potential differences in whistles produced in the relatively close region of the North Ionian Sea. Despite the small sample size and slightly variable survey methodology followed in each location, the acoustic features of whistles reported here are consistent with the results reported elsewhere in the Mediterranean Sea. Whistles are known to carry species-specific cues and it has been suggested that they carry information on social structure, behaviour, and the cohesion of the group (Miller, 2002; Lammers et al., 2003; Branstetter et al., 2013; Díaz López et al, 2017). Therefore, further research is needed to identify the role that behavioural, environmental, and anthropogenic states have on the acoustic features of whistles of bottlenose dolphins both in Taranto Gulf and Boka Bay.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by The Montenegrin Government.
Author contributions
AA, TA, PL, GC, MA, LI, OE, YA, PR, RCr, EP, and CF participated in the field surveys. Analysis was conducted by AA, TA, KM, and PL. AA and RCa supervised the manuscript. The manuscript was written by all the authors. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Rufford Small Grant Foundation, under the project (project number 35446-B) titled “From Research to Policy Implications; Minimizing Negative Human Impacts on Cetaceans and their Vulnerable Habitats in the South Adriatic”.
Acknowledgments
The authors would like to thank the Rufford Small Grant Foundation for their invaluable funding contributions which allowed this research to take place. Further, we would like to thank the Environmental Agency of Montenegro for providing us the necessary grant to run the current project. Finally, we would like to extend our gratitude to all of the volunteers and researchers, specifically crediting Yağmur Atalan for her time and contribution that helped to collect this data.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ðurovic M., Holcer D., Joksimovic A., Mandic M., Fortuna C., Ikica Z., et al. (2016). Cetaceans in the boka kotorska bay. Handb. Environ. Chem. 54), 411–437. doi: 10.1007/698_2016_46
ACCOBAMS (2021). Estimates of abundance and distribution of cetaceans, marine mega-fauna and marine litter in the Mediterranean Sea from 2018-2019 surveys Vol. 177. Eds. Panigada S., Boisseau O., Canadas A., Lambert C., Laran S., McLanaghan R., et al (Monaco: ACCOBAMS - ACCOBAMS Survey Initiative Project).
Akkaya A., Lyne P., Schulz X., Awbery T., Capitain S., Rosell B. F., et al. (2020). Preliminary results of cetacean sightings in the eastern Mediterranean Sea of Turkey. J. Black Sea/Mediterranean Environ. 26 (1), 26–47.
Akkaya A., Ruegg N., Awbery T., Velez N., Goossens C., Papadopoulo K., et al. (2021). “October. bottlenose dolphins of Montenegro: Abundance estimation and residency patterns,” in 2021 international workshop on metrology for the sea; learning to measure Sea health parameters (MetroSea) (Reggio Calabria, Italy: IEEE), vol. 100-104. , IEEE. doi: 10.1109/MetroSea52177.2021.9611636
Amorim T. O. S., Andriolo A., Reis S. S., Santos M. E. (2016). Vocalisations of Amazon river dolphins (Inia geoffrensis): Characterization, effect of physical environment and differences between populations. J. Acoustical Soc. America, 139 (3), 1285–1293. doi: 10.1121/1.4943556
Awbery T., Akkaya A., Lyne P., Rudd L., Hoogenstrijd G., Nedelcu M., et al. (2022). Spatial distribution and encounter rates of delphinids and deep diving cetaceans in the Eastern Mediterranean Sea of Turkey and the extent of overlap with areas of dense marine traffic. Front. Mar. Sci. 739. doi: 10.3389/fmars.2022.860242
Awbery T., Nikpaljevic N., Clarkson J., Abbiss L., van der Pouw Kraan D., Liebig P., et al. (2019). Bottlenose and striped dolphins of Montenegro: an insight into sighting variations, behavioural patterns, photo-identification, core habitats, marine traffic and conservation initiatives, (2017–2018). Annu. Rep. Montenegro Dolphin Research DMAD Antalya.
Azevedo F. A., Oliveira A. M., Dalla Rosa L., Lailson-Brito J. (2007). Characteristics of whistles from resident bottlenose dolphins (Tursiops truncatus) in southern Brazil. J. Acoust. Soc Am. 121 (5), 2978–2983. doi: 10.1121/1.2713726
Azzolin M., Arcangeli A., Cipriano G., Crosti R., Maglietta R., Pietroluongo G., et al. (2020). Spatial distribution modelling of striped dolphin (Stenella coeruleoalba) at different geographical scales within the EU Adriatic and Ionian Sea region, central-eastern Mediterranean Sea. Aquat. Conservation: Mar. Freshw. Ecosyst. 30 (6), 1194–1207. doi: 10.1002/aqc.3314
Azzolin M., Gannier A., Papale E., Buscaino G., Mussi B., Ardizzone G., et al. (2017). Whistle variability of the Mediterranean short beak common dolphin. Aquat. Conservation: Mar. Freshw. Ecosyst. 31 (May 2019), 36–50. doi: 10.1002/aqc.3168
Azzolin M., Saintingan S., Zampollo A., Carlucci R., Maglietta R., Cipriano G., et al. (2018). Multi source data analysis for improving striped dolphin distribution modelling at a regional scale. 2018 IEEE Int. Workshop Metrology Sea; Learn. to Measure Sea Health Parameters (MetroSea), 145–149. doi: 10.1109/MetroSea.2018.8657895
Balkaninsight (2019). Available at: https://balkaninsight.com/2019/03/20/crowded-with-cruise-ships-fears-for-montenegrin-bay/ (Accessed November 13, 2022).
Baron S. C., Martinez A., Garrison L. P., Keith E. O. (2008). Differences in acoustic signals from delphinids in the western north Atlantic and northern gulf of Mexico. Mar. Mammal Sci. 24, 42–56. doi: 10.1111/j.1748-7692.2007.00168.x
Baş A. A., Affinito F., Martin S., Vollmer A., Gansen C., Morris N., et al. (2017a). Bottlenose dolphins and striped dolphins: Species distribution, behavioural patterns, encounter rates, residency patterns and hotspots in Montenegro, south adriatic. Montenegro dolphin project annual report. Mar. Mammal Res. Assoc. Bar.
Baş A. A., Erdogan M. A., Morris N. R. C., Yeoman K., Humphrey O., Gaggioli E., et al. (2017b). Seasonal encounter rates and residency patterns of an unstudied population of bottlenose dolphin (Tursiops truncatus) in the northwestern levantine Sea, Turkey. Hyla: Herpetological Bull. 2016 (1), 1–13.
Bearzi G., Bonizzoni S., Santostasi N., Furey N., Eddy L., Valavanis V., et al. (2016). “Dolphins in a scaled-down Mediterranean: the gulf of corinth's odontocetes,” in Elsevier. Eds. Notarbartolo Sciara G., Podestà M., Curry B. E., p297–p331. (Cambridge, Massachusetts: Academic Press).
Bearzi G., Costa M., Politi E., Agazzi S., Pierantonio N., Tonini D., et al. (2009). Cetacean records and encounter rates in the northern Adriatic sea during the years 1988-2007. Annales Ser. Hist. Naturalis 19 (2), 145–150.
Bearzi G., Fortuna C., Reeves R. (2008). Ecology and conservation of common bottlenose dolphins Tursiops truncatus in the Mediterranean Sea. Mammal Rev. 39 (2), 92. doi: 10.1111/j.1365-2907.2008.00133.x
Bearzi G., Fortuna C., Reeves R. (2012)Tursiops truncatus. In: The IUCN red list of threatened species 2012: e.T22563A2782611. Available at: https://www.iucnredlist.org/species/22563/2782611 (Accessed 13 Nov 2022).
Bearzi G., Genov T., Natoli A., Gonzalvo J., Pierce G. J. (2021). Delphinus delphis (Inner Mediterranean subpopulation). IUCN Red List Threatened Species 2021. doi: 10.2305/IUCN.UK.2021-3.RLTS.T189865869A189865884.en
Bearzi G., Holcer D., Notarbartolo di Sciara G. (2004). The role of historical dolphin takes and habitat degradation in shaping the present status of northern Adriatic cetaceans. Aquat. Conservation: Mar. Freshw. Ecosyst. 14 (4), 363–379. doi: 10.1002/aqc.626
Bearzi G., Notarbartolo di Sciara G. (1995). A comparison of the present occurrence of bottlenose dolphins, Tursiops truncatus, and common dolphins, Delphinus delphis, in the kvarneric (northern Adriatic Sea). Ecology Status Conserv. cetaceans Adriatic Sea Project 7, 61–68.
Bearzi G., Politi E., Notarbartolo di Sciara G. N. (1999). Diurnal behavior of free-ranging bottlenose dolphins in the kvanerić northern Adriatic Sea. Mar. Mammal Sci. 15 (4), 1065–1097. doi: 10.1111/j.1748-7692.1999.tb00878.x
Bearzi G., Notarbartolo di Sciara G., Politi E. (1997). Social ecology of bottlenose dolphins in the kvarnerić (Northern Adriatic Sea). Mar. Mammal Sci. 13 (4), 650–668. doi: 10.1111/j.1748-7692.1997.tb00089.x
Bellomo S., Santacesaria F. C., Fanizza C., Cipriano G., Renò V., Carlucci R., et al. (2019). “Photo-identification of physeter macrocephalus in the gulf of taranto (Northern Ionian Sea, central-eastern Mediterranean Sea),” in Proceedings of the IMEKO metrology for the Sea, vol. 3–5 October 2019. (Italy: Genova), 33–37.
Boisseau O. (2005). Quantifying the acoustic repertoire of a population: the vocalizations of free-ranging bottlenose dolphins in fiordland, new Zealand. J.Acoust.Soc. Am. 117 (4), 2318–2329. doi: 10.1121/1.1861692
Bolger M. K., Díaz L. B., Hope V., Methion S. (2017). “Whistles characteristics of bottlenose dolphins in the ria of arousa, Galicia NW Spain,” in Conference: European Cetacean Society, Middelfart, Denmark Ordinal, Vol. 31s.
Branstetter B. K., Black A., Bakhtiari K. (2013). Discrimination of mixed-directional whistles by a bottlenose dolphin (Tursiops truncatus). J. Acoustical Soc. America 134, 2274. doi: 10.1121/1.4816404
Branstetter B. K., Moore P. W., Finneran J. J., Tormey M. N., Aihara H. (2012). Directional properties of bottlenose dolphin (Tursiops truncatus) clicks, burst-pulse, and whistle sounds. J. Acoustical Soc. America 131 (2), 1613. doi: 10.1121/1.3676694
Buckstaff K. C. (2004). Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota bay, Florida. Mar. Mammal Sci. 20, 709–725. doi: 10.1111/j.1748-7692.2004.tb01189.x
Caldwell M. C., Caldwell D. K., Tyack P. L. (1990). Review of the signature-whistle hypothesis for the Atlantic bottlenose dolphin. bottlenose dolphin, 199–235. doi: 10.1016/B978-0-12-440280-5.50014-7
Capezzuto F., Carlucci R., Maiorano P., Sion L., Battista D., Giove A., et al. (2010). The bathyal benthopelagic fauna in the north-western Ionian Sea: structure, patterns and interactions. Chem. Ecol. 26, 199–217. doi: 10.1080/02757541003639188
Carlucci R., Bandelj V., Ricci P., Capezzuto F., Sion L., Maiorano P., et al. (2018b). Exploring spatio-temporal changes in the demersal and benthopelagic assemblages of the north-western Ionian Sea central Mediterranean Sea. Mar. Ecol. Prog. Ser. 598, 1–19. doi: 10.3354/meps12613
Carlucci R., Baş A. A., Liebig P., Renò V., Santacesaria F. C., Bellomo S., et al. (2020b). Residency patterns and site fidelity of Grampus griseus cuvier 1812 in the gulf of taranto northern Ionian Sea, central-Eastern Mediterranean Sea. Mammal Res. doi: 10.1007/s13364-020-00485-z
Carlucci R., Baş A. A., Maglietta R., Reno V., Fanizza C., Rizzo A., et al. (2018c). Site fidelity, residency and habitat use of the risso’s dolphin Grampus griseus in the gulf of taranto northern Ionian Sea, central-eastern Mediterranean Sea by photo-identification. 2018 IEEE Int. Workshop Metrology Sea; Learn. to Measure Sea Health Parameters (MetroSea) 173–177. doi: 10.1109/MetroSea.2018.8657847
Carlucci R., Cipriano G., Paoli C., Ricci P., Fanizza C., Capezzuto F., et al. (2018a). Random forest population modelling of striped and common-bottlenose dolphins in the gulf of taranto northern Ionian Sea, central-eastern Mediterranean Sea. Estuarine Coast. Shelf Sci. 204, 177–192. doi: 10.1016/j.ecss.2018.02.034
Carlucci R., Cipriano G., Santacesaria F. C., Ricci P., Maglietta R., Petrella A., et al. (2020a). Exploring data from an individual stranding of a cuvier’s beaked whale in the gulf of taranto northern Ionian Sea, central-eastern Mediterranean Sea. J. Exp. Mar. Biol. Ecol. 65, 445–455. doi: 10.1016/j.jembe.2020.151473
Carlucci R., Fanizza C., Cipriano G., Paoli C., Russo T., Vassallo P. (2016). Modelling the spatial distribution of the striped dolphin (Stenella coeruleoalba) and common bottlenose dolphin (Tursiops truncatus) in the gulf of taranto (Northern Ionian Sea, central-eastern Mediterranean Sea). Ecol. Indic. 69, 707–721. doi: 10.1016/j.ecolind.2016.05.035
Carlucci R., Manea E., Ricci P., Cipriano G., Fanizza C., Maglietta R., et al. (2021). Managing multiple pressures for cetaceans’ conservation with an ecosystem-based marine spatial planning approach. J. Environ. Manage. 287, 112240. doi: 10.1016/j.jenvman.2021.112240
Carlucci R., Ricci P., Cipriano G., Fanizza C. (2017). Abundance, activity and critical habitat of the striped dolphin Stenella coeruleoalba in the gulf of taranto northern Ionian Sea, central Mediterranean Sea. Aquat. Conservation: Mar. Freshw. Ecosyst. 28 (2), 324–336. doi: 10.1002/aqc.2867
Caruso F., Dong L., Lin M., Liu M., Gong Z., Xu W., et al. (2020). Monitoring of a nearshore small dolphin species using passive acoustic platforms and supervised machine learning techniques. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00267
Cipriano G., Carlucci R., Bellomo S., Santacesaria F. C., Fanizza C., Ricci P., et al. (2022). Behavioural pattern of risso’s dolphin (Grampus griseus) in the gulf of taranto northern Ionian Sea, central-Eastern Mediterranean Sea. J. Mar. Sci. Eng. 10 (2), 175. doi: 10.3390/jmse10020175
Clarkson J., Christiansen F., Awbery T., Abbiss L., Nikpaljevic N., Akkaya A. (2020). Non-targeted tourism affects the behavioural budgets of bottlenose dolphins Tursiops truncatus in the south Adriatic Montenegro. Mar. Ecol.Prog. Ser. 638, 165–176. doi: 10.3354/meps13257
Conner D. A. (1982). Dialects versus geographic variation in mammalian vocalisations. Anim. Behav. 30, 297–298. doi: 10.1016/S0003-3472(82)80269-8
Connor C., Smolker A. (1996). ‘Pop’ goes the dolphin: a vocalization male bottlenose dolphins produce during courtship. Behaviour 133, 643–662. doi: 10.1163/156853996X00404
Corrias V., de Vincenzi G., Ceraulo M., Sciacca V., Sala A., de Lucia G. A., et al. (2021). Bottlenose dolphin (Tursiops truncatus) whistle modulation during a trawl bycatch event in the Adriatic Sea. Animals 11 (12), 3593. doi: 10.3390/ani11123593
Davis G. E., Baumgartner M. F., Bonnell J. M., Bell J., Berchok C., Bort Thornton J., et al. (2017). Long-term passive acoustic recordings track the changing distribution of north Atlantic right whales (Eubalaena glacialis) from 2004 to 2014. Sci. Rep. 7 (1), 1–12. doi: 10.1038/s41598-017-13359-3
Díaz López B. (2011). Whistle characteristics in free-ranging bottlenose dolphins (Tursiops truncatus) in the Mediterranean Sea: Influence of behaviour. Mamm. Biol. 76 (2), 180–189. doi: 10.1016/j.mambio.2010.06.006
Díaz López B., López A., Methion S., Covelo P. (2017). Infanticide attacks and associated epimeletic behaviour in free-ranging common bottlenose dolphins (Tursiops truncatus). J. Mar. Biol. Assoc. United Kingdom 98 (05), 1159–1167. doi: 10.1017/S0025315417001266
Díaz López B., Shirai J. A. B. (2010). “Mediterranean Common bottlenose dolphin’s repertoire and communication use,” in Dolphins: Anatomy, behavior, and threats. Eds. Pearce A. G., Correa L. M. (New York: Nova Science Publishers, Inc.), 129–148.
Ding W., Wursing B., Evans W. E. (1995). Whistles of bottlenose dolphins: Comparisons among populations. Aquat. Mammals 21, 65–77.
Domit C., Laporta P., Zappes C. A., Lodi L., Hoffmann L. S., Genoves R., et al. (2016). Report of the working group on the behavioral ecology of bottlenose dolphins in the southwest Atlantic ocean. Latin Am. J. Aquat. Mammals 11 (1-2), 106–120. doi: 10.5597/lajam00220
Duarte C. M., Chapuis L., Collin S. P., Costa D. P., Devassy R. P., Eguiluz V. M., et al. (2021). The soundscape of the anthropocene ocean. Science 371 (6529), 4658. doi: 10.1126/science.aba4658
Esch H. C., Sayigh L. S., Blum J. E., Wells R. S. (2009a). Whistles as potential indicators of stress in bottlenose dolphins (Tursiops truncatus). J. Mammalogy 90 (3), 638–650. doi: 10.1644/08-MAMM-A-069R.1
Esch H. C., Sayigh L. S., Wells R. S. (2009b). Quantifying parameters of bottlenose dolphin signature whistles. Mar. Mammal Sci. 25 (4), 976–986. doi: 10.1111/j.1748-7692.2009.00289.x
Estabrook B. J., Ponirakis D. W., Clark C. W., Rice A. N. (2016). Widespread spatial and temporal extent of anthropogenic noise across the north-eastern gulf of Mexico shelf ecosystem. Endangered Species Res. 30, 267–282. doi: 10.3354/esr00743
Fleetmon (2022). Available at: https://www.fleetmon.com/ports/taranto_ittar_7053/?language=en (Accessed November 13, 2022).
Fortuna C. M., Holcer D., Filidei E., Donovan G., Tunesi L. (2011). “March. the first cetacean aerial survey in the Adriatic Sea: summer 2010,” in 7th meeting of the ACCOBAMS scientific committee, p16.
Fruet P. F., Zappes C. A., Bisi T. L., Simões-Lopes P. C., Laporta P., Loureiro J. D., et al. (2017). Report of the working group on interactions between humans and tursiops truncatus in the southwest Atlantic ocean. Latin Am. J. Aquat. Mammals 11, 79–98. doi: 10.5597/lajam00218
Gannier A., Fuchs S., Gannier A., Fernandez M., Azevedo J. M. N. (2020). Dolphin whistle repertoires around são Miguel (Azores): Are you common or spotted? Appl. Acoustics 161, 107169. doi: 10.1016/j.apacoust.2019.107169
Gannier A., Fuchs S., Quèbre P., Oswald J. N. (2010). Performance of a contour-based classification method for whistles of Mediterranean delphinids. Appl. Acoustics 71 (11), 1063–1069. doi: 10.1016/j.apacoust.2010.05.019
Genov T., Wiemann A., Fortuna C. M. (2009). Towards identification of the bottlenose dolphin (Tursiops truncatus) population structure in the north-eastern Adriatic Sea: preliminary results. Varstvo Narave 22, 73–80.
Genov T., Železnik J., Bruno C., Ascheri D., Fontanesi E., Blasi M. F. (2022). The longest recorded movement of an inshore common bottlenose dolphin (Tursiops truncatus). Mamm. Biol., 1–13. doi: 10.1007/s42991-022-00316-5
Hawkins E. R. (2010). Geographic variations in the whistles of bottlenose dolphins (Tursiops aduncus) along the east and west coasts of Australia. J. Acoustical Soc. America 128 (2), 924–935. doi: 10.1121/1.3459837
Heithaus M. R., Vaudo J. J., Kreicker S., Layman C. A., Krützen M., Burkholder D. A., et al. (2013). Apparent resource partitioning and trophic structure of large-bodied marine predators in a relatively pristine seagrass ecosystem. Mar. Ecol. Prog. Ser. 481, 225–237. doi: 10.3354/meps10235
Henderson E. E., Hildebrand J. A., Smith M. H., Falcone E. A. (2011). The behavioral context of common dolphin (Delphinus sp.) vocalizations. Mar. Mammal Sci. 28 (3), 439–460. doi: 10.1111/j.1748-7692.2011.00498.x
Herzing D. L. (2014). Clicks, whistles and pulses: Passive and active signal use in dolphin communication. Acta Astronautica 105 (2), 534–537. doi: 10.1016/j.actaastro.2014.07.003
Herzing D. L. (2015). Synchronous and rhythmic vocalizations and correlated underwater behavior of free-ranging Atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in the Bahamas. Anim. Behav. Cogn. 2 (1), 14–29. doi: 10.12966/abc.02.02.2015
Hickey R., Berrow S., Goold J. (2009). Towards a bottlenose dolphin whistle ethogram from the Shannon estuary, Ireland. Biol. Environment: Proc. R. Irish Acad. 109B (2), 89–94. doi: 10.3318/BIOE.2009.109.2.89
Hildebrand J. A. (2009). Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 395, 5–20. doi: 10.3354/meps08353
Hill H., Lackups M. (2010). Journal publication trends regarding cetaceans found in both wild and captive environments: What do we study and where do we publish? Int. J. Comp. Psychol. 23 (3), 414–534. doi: 10.46867/IJCP.2010.23.03.01
Janik V. M. (1999). Pitfalls in the categorization of behaviour: A comparison of dolphin whistle classification methods. Anim. Behav. 57 (1), 133–143. doi: 10.1006/anbe.1998.0923
Janik V. M. (2009). Acoustic communication in delphinids. Adv. Study Behav. 40, 123–157. doi: 10.1016/S0065-3454(09)40004-4
Janik V. M., King S. L., Sayigh L. S., Wells R. S. (2012). Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 29 (1), 109–122. doi: 10.1111/j.1748-7692.2011.00549.x
Janik V. M., Slater P. J. (1998). Context-specific use suggests that bottlenose dolphin signature whistles are cohesion calls. Anim. Behav. 56 (4), 829–838. doi: 10.1006/anbe.1998.0881
Jones B., Zapetis M., Samuelson M. M., Ridgway S. (2019). Sounds produced by bottlenose dolphins (Tursiops): a review of the defining characteristics and acoustic criteria of the dolphin vocal repertoire. Bioacoustics 29, 1–42. doi: 10.1080/09524622.2019.1613265
Jung S., Lee M. H., Ahn J. (2018). On the number of principal components in high dimensions. Biometrika 105 (2), 389–402. doi: 10.1093/biomet/asy010
La Manna G., Manghi M., Pavan G., Lo Mascolo F., Sarà G. (2013). Behavioural strategy of common bottlenose dolphins (Tursiops truncatus) in response to different kinds of boats in the waters of lampedusa island (Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 23, 745–757. doi: 10.1002/aqc.2355
La Manna G., Rako Gospić N., Manghi M., Picciulin M., Sarà G. (2017). Assessing geographical variation on whistle acoustic structure of three Mediterranean populations of common bottlenose dolphin (Tursiops truncatus). Behaviour 154 (5), 583–607. doi: 10.1163/1568539X-00003435
La Manna G., Rako-Gospić N., Pace D. S., Bonizzoni S., Di Iorio L., Polimeno L., et al. (2022). Determinants of variability in signature whistles of the Mediterranean common bottlenose dolphin. Sci. Rep. 12 (1), 1–16. doi: 10.1038/s41598-022-10920-7
La Manna G., Rako-Gospić N., Sarà G., Gatti F., Bonizzoni S., Ceccherelli G. (2020). Whistle variation in Mediterranean common bottlenose dolphin: The role of geographical, anthropogenic, social, and behavioural factors. Ecol. Evol. 10, 1971–1987. doi: 10.1002/ece3.6029
La Manna G., Rako-Gòspic N., Manghi M., Ceccherelli G. (2019). Influence of environmental, social and behavioural variables on the whistling of the common bottlenose dolphin (Tursiops truncatus). Behav. Ecol. Sociobiology 73 (9), 1–15. doi: 10.1007/s00265-019-2736-2
Lammers M. O., Au W. W. L., Herzing D. L. (2003). The broadband social acoustic signaling behavior of spinner and spotted dolphins. J. Acoustical Soc. America 114 (3), 1629–1639. doi: 10.1121/1.1596173
Lind H., Dabelsteen T., McGregor P. K. (1996). Female great tits can identify mates by song. Anim. Behav. 52, 667–671. doi: 10.1006/anbe.1996.0211
Luís A. R., May-Collado L. J., Rako-Gospić N., Gridley T., Papale E., Azevedo A., et al. (2021). Vocal universals and geographic variations in the acoustic repertoire of the common bottlenose dolphin. Sci. Rep. 11, 11847. doi: 10.1038/s41598-021-90710-9
MacFarlane N., Janik V., Jensen F. H., McHugh K., Sayigh L., Wells R., et al. (2017). Signature whistles facilitate reunions and/or advertise identity in bottlenose dolphins. J. Acoustical Soc. America 141, 3543. doi: 10.1121/1.4987492
Maiorano P., Capezzuto F., Carluccio A., Calculli C., Cipriano G., Carlucci R., et al. (2022). Food from the depths of the Mediterranean: The role of habitats, changes in the Sea-bottom temperature and fishing pressure. Foods 11, 1420. doi: 10.3390/foods11101420
MAP-UNEP (2015). Progress report on activities carried out during the 2014-2015 biennium and draft decision on the action plans concerning cetaceans, corralligenous and other calcareous bioconcretions, and species introductions and invasive species under the protocol concerning specially protected areas and biological diversity in the mediterranean. Athens, Greece.
MarineTraffic (2022). Available at: https://www.marinetraffic.com/en/ais/home/centerx:3.8/centery:30.2/zoom:4 (Accessed November 13, 2022).
Martinez Arbizu P. (2020). PairwiseAdonis: Pairwise multilevel comparison using adonis. r package version 0.4. Available at: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 1 September 2022).
May-Collado L. J., Wartzok D. (2008). A comparison of bottlenose dolphin whistles in the Atlantic ocean: Factors promoting whistle variation. J. Mammalogy 89 (5), 1229–1240. doi: 10.1644/07-MAMM-A-310.1
Mellinger D., Barlow J. (2003). Future directions for acoustic marine mammal surveys: stock assessment and habitat use 20–22. NOAA OAR Special Report. Report of a workshop held in La Jolla, California.
Mellinger D. K., Stafford K. M., Moore S. E., Dziak R. P., Matsumoto H. (2007). An overview of fixed passive acoustic observation methods for cetaceans. Oceanography 20 (4), 36–45. doi: 10.5670/oceanog.2007.03
Miller P. J. O. (2002). Mixed-directionality of killer whale stereotyped calls: A direction of movement cue? Behav. Ecol. Sociobiol. 52, 262–270. doi: 10.1007/s00265-002-0508-9
Miokovic D., Kovacic D., Pribanic S. (1997). “Stomach content analysis of a bottlenose dolphin (Tursiops truncatus) from the Adriatic sea. eur. res. cetaceans 11-149,” in eleventh annual conference of the European Cetacean Society, Stralsund, Germany. Nat. Croat 8,61-65. Zagreb 10–12.
Natoli A., Cañadas A., Vaquero C., Politi E., Fernandez-Navarro P., Hoelzel A. R. (2008). Conservation genetics of the short-beaked common dolphin (Delphinus delphis) in the Mediterranean Sea and in the eastern north Atlantic ocean. Conserv. Genet. 9 (6), 1479–1487. doi: 10.1007/s10592-007-9481-1
Notarbartolo di Sciara G., Venturino M. C., Zanardelli M., Bearzi G., Borsani F. J. (1993). Cetaceans in the central Mediterranean Sea:Distribution and sighting frequencies. Bolletino di Zoologia 60, 131–138. doi: 10.1080/11250009309355800
Pace D. S., Tumino C., Silvestri M., Giacomini G., Pedrazzi G., Pavan G., et al. (2022). Bray-call sequences in the Mediterranean common bottlenose dolphin (Tursiops truncatus) acoustic repertoire. Biology 11 (3), 367. doi: 10.3390/biology11030367
Panigada S., Lauriano G., Donovan G., Pierantonio N., Cañadas A., Vázquez J. A., et al. (2017). Estimating cetacean density and abundance in the central and Western Mediterranean Sea through aerial surveys: implications for management. Deep Sea Res. Part II: Topical Stud. Oceanography 141, 41–58. doi: 10.1016/j.dsr2.2017.04.018
Papale E., Azzolin M., Cascão I., Gannier A., Lammers M. O., Martin V. M., et al. (2013). Geographic variability in the acoustic parameters of striped dolphin’s (Stenella coeruleoalba) whistles. J. Acoustical Soc. America 133 (2), 1126–1134. doi: 10.1121/1.4774274
Papale E., Azzolin M., Cascão I., Gannier A., Lammers M. O., Martin V. M., et al. (2014a). Acoustic divergence between bottlenose dolphin whistles from the central–Eastern north Atlantic and Mediterranean Sea. Acta Ethologica 17 (3), 155–165. doi: 10.1007/s10211-013-0172-2
Papale E., Azzolin M., Cascão I., Gannier A., Lammers M. O., Martin V. M., et al. (2014b). Macro- and micro-geographic variation of short-beaked common dolphins whistles in the mediterranean sea and atlantic ocean. Ethology Ecol. Evol. 26 (4), 392–404. doi: 10.1080/03949370.2013.851122
Papale E. B., Azzolin M. A., Cascão I., Gannier A., Lammers M. O., Martin V. M., et al. (2021). Dolphin whistles can be useful tools in identifying units of conservation. BMC Zoology 6 (1), 1–13. doi: 10.1186/s40850-021-00085-7
Papale E., Fanizza C., Buscaino G., Maria C., Cipriano G., Crugliano R., et al. (2020). The social role of vocal complexity in striped dolphins. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.584301
Paudel S., Koprowski J. L., Thakuri U., Sigdel R., Gautam R. C. (2020). Ecological responses to flow variation inform river dolphin conservation. Sci. Rep. 10 (1), 1–13. doi: 10.1038/s41598-020-79532-3
Peraš I., Mandić M., Nikolić S. (2022). Settlement patterns of molluscs, with particular reference on great Mediterranean scallop (Pecten jacobaeus, linnaeus) and biofouling organisms on different type of collectors and locations in boka kotorska bay. Acta Adriatica: Int. J. Mar. Sci. 63 (1), 9–26. doi: 10.32582/aa.63.1.1
Pinardi N., Lyubartsev V., Cardellicchio N., Caporale C., Ciliberti S., Coppini G., et al. (2016). Marine rapid envrionemntal assessment in the gulf of taranto a multiscale approach. Nat. Hazaards Earth Syst. Sci. 16, 2623–2639. doi: 10.5194/nhess-16-2623-2016
Pribanić S., Mioković D., Kovačić D. (2000). Preliminary growth rate and body lengths of the bottlenose dolphins Tursiops truncatus (Montagu 1821) from the Adriatic Sea. Natura Croatica: Periodicum Musei Historiae Naturalis Croatici 9 (3), 179–188.
Quick N. J., Janik V. M. (2008). Whistle rates of wild bottlenose dolphins (Tursiops truncatus): Influences of group size and behavior. J. Comp. Psychol. 122 (3), 305–311. doi: 10.1037/0735-7036.122.3.305
Rako-Gospić N., La Manna G., Picciulin M., Ceccherelli G. (2020). Influence of foraging context on the whistle structure of the common bottlenose dolphin. Behav. Processes 182, 104281. doi: 10.1016/j.beproc.2020.104281
Rasmussen M. H., Lammers M., Beedholm K., Miller A. (2006). Source levels and harmonic content of whistles in white-beaked dolphins (Lagenorhynchus albirostris). J. Acoust Soc. Am. 120, 510–517. doi: 10.1121/1.2202865
Rendall D., Rodman P. S., Emond R. E. (1996). Vocal recognition of individuals and kin in free-ranging rhesus monkeys. Anim. Behav. 51, 1007–1015. doi: 10.1006/anbe.1996.0103
Rendell L., Matthews J., Gill A., Gordon J., Macdonald D. (1999). Quantitative analysis of tonal calls from five odontocete species, examining interspecific and intraspecific variation. J. Zoology 249 (4), 403–410. doi: 10.1111/j.1469-7998.1999.tb01209.x
Ricci P., Manea E., Cipriano G., Cascione D., D’Onghia G., Ingrosso M., et al. (2021). Addressing cetacean–fishery interactions to inform a deep-Sea ecosystem-based management in the gulf of taranto northern Ionian Sea, central Mediterranean Sea. J. Mar. Sci. Eng. 9 (8), 872. doi: 10.3390/jmse9080872
Richardson W. J., Greene C. R. Jr., Malme C. I., Thomson D. H. (1995). Marine mammals and noise (San Diego, CA: Academic Press), 159–204. doi: 10.1016/C2009-0-02253-3
Rudd L. E., Awbery T., Waller R., Crowe M., Bauer J., Jacquemart A., et al. (2022). The effect of commercial and artisanal fishing practices on the behavioral budget of bottlenose dolphins off the coast of Montenegro, south Adriatic Sea. Mar. Mammal Sci. 38, 990–1006. doi: 10.1111/mms.12913
Sackl P., Smole J., Saveljić D., Štumberger B. (2007). Inland observations of common bottlenose dolphins tursiops truncatus montagu 1821 in the delta of the Bojana/Buna river, Albania and Montenegro. Annales. Ser. Hist. naturalis 17 (2), 191–194.
Santacesaria F. C., Bellomo S., Fanizza C., Maglietta R., Renò V., Cipriano G., et al. (2019). Long-term residency of Tursiops truncatus in the gulf of taranto northern Ionian Sea, central-eastern Mediterranean sea. international workshop on metrology for the Sea: Learning to measure Sea health parameters, MetroSea 2019. Genoa Italy 2019, 28–32.
Sayigh L. S., Esch H. C., Wells R. S., Janik V. M. (2007). Facts about signature whistles of bottlenose dolphins, tursiops truncatus. Anim. Behav. 74 (6), 1631–1642. doi: 10.1016/j.anbehav.2007.02.018
Schultz K. W., Cato D. H., Corkeron P. J., Bryden M. M. (1995). ). low frequency narrow-band sounds produced by bottlenose dolphins. Mar. Mammal Sci. 11 (4), 503–509. doi: 10.1111/j.1748-7692.1995.tb00673.x
Shipnext (2022). Available at: https://shipnext.com/solution-shipnext-fleet-monitor (Accessed November 13, 2022).
Steiner W. (1981). Species-specific differences in pure tonal whistle vocalizations of five western north Atlantic dolphin species. Behav. Ecol. Sociobiology 9, 241–246. doi: 10.1007/BF00299878
Terranova F., Gnone G., Friard O., Bellingeri M., Giacoma C., Favaro L. (2021). Signature whistles of the demographic unit of bottlenose dolphins (Tursiops truncatus) inhabiting the Eastern ligurian Sea: characterisation and comparison with the literature. Eur. Zoological J. 88 (1), 771–781. doi: 10.1080/24750263.2021.1936225
Terranova F., Raffa A., Floridia S., Monaco C., Favaro L. (2022). Vocal behaviour of a bottlenose dolphin pod during a deadly bycatch event in the gulf of catania, Ionian Sea. J. Mar. Sci. Eng. 10, 616. doi: 10.3390/jmse10050616
Van Parijs S. M., Clark C. W., Sousa-Lima R. S., Parks S. E., Rankin S., Risch D., et al. (2009). Management and research applications of real-time and archival passive acoustic sensors over varying temporal and spatial scales. Mar. Ecol. Prog. Ser. 395, 21–36. doi: 10.3354/meps08123
Wang D., Bernd W., William E. E. (1995). Whistles of bottlenose dolphins: comparisons among populations. Aquat. Mammals 21 (1), 65–77.
Wells R. S. (1991). “The role of long-term study in understanding the social structure of a bottlenose dolphins community,” in Dolphin societies: discoveries and puzzles (Berkeley: University of California Press), 199–225. doi: 10.1017/S0025315409991251
Keywords: cetacean, data-deficient, delphinids, geographical variation, Mediterranean Sea, vocalisation, whistles
Citation: Akkaya A, Awbery T, Medcalf K, Lyne P, Cipriano G, Alvarenga M, İsrapilova L, Atalan Y, Eikelenboom O, Ricci P, Crugliano R, Papale E, Fanizza C and Carlucci R (2023) Initial results on the variation of whistle characteristics of bottlenose dolphins from two neighbouring regions of the Mediterranean Sea: Northern Ionian and Southern Adriatic Sea. Front. Mar. Sci. 10:1099576. doi: 10.3389/fmars.2023.1099576
Received: 15 November 2022; Accepted: 09 January 2023;
Published: 23 January 2023.
Edited by:
Leslie New, Ursinus College, United StatesReviewed by:
Salvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), BrazilDenise Lore Herzing, Florida Atlantic University, United States
Copyright © 2023 Akkaya, Awbery, Medcalf, Lyne, Cipriano, Alvarenga, İsrapilova, Atalan, Eikelenboom, Ricci, Crugliano, Papale, Fanizza and Carlucci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kirsty Medcalf, a2lyc3R5Lm0ubWVkY2FsZkBnbWFpbC5jb20=