
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 15 June 2023
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1035448
This article is part of the Research TopicInnovations in Fishing Technology Aimed at Achieving Sustainable FishingView all 23 articles
 Francesca Ferragut-Perello1*
Francesca Ferragut-Perello1* Claudio Vasapollo2
Claudio Vasapollo2 Maria Valls1
Maria Valls1 Maria Teresa Farriols1Enric Massutí1
Maria Teresa Farriols1Enric Massutí1 Beatriz Guijarro1Sergi Joher1Marina Bibiloni-Socias1
Beatriz Guijarro1Sergi Joher1Marina Bibiloni-Socias1 Francesc Ordines1
Francesc Ordines1The Spanish bottom trawl fleet operating in the Mediterranean currently uses 40 mm square mesh codends. Its selectivity is still too low to overcome the overall problem of high proportions of immature specimens in the catches. Even for some target species such as the European hake (Merluccius merluccius), there are high proportions of individuals smaller than the minimum conservation reference size (MCRS). The aim of the present work is to assess the selectivity of three different configurations of the traditional net (TRA) used in the bottom trawl hake fishery off the northwest Iberian Peninsula, each including the following modifications: i) an extension piece made of 90° turned diamond mesh (T90), ii) a 52 mm square meshed codend (52S), and iii) both modifications combined (EXP). The experimental fishing survey applied two methodologies: alternate hauls to compare the different net configurations; and the covered codend method to calculate the selectivity parameters of 52 mm square meshed codend for the main commercial species. Catch comparisons showed no discard reduction using the T90 extension piece for any of the species analysed, nor any selectivity improvement. Conversely, the 52 mm square meshed codend showed a clear discard reduction for M. merluccius and a generalized improvement of selectivity for most commercial species. This improvement raised the 50% retention length (L50) for M. merluccius to 22.2 cm, well above its MCRS (20 cm), allowing to escape 90% of the undersized individuals. However, the implementation of the 52 mm square meshed codend would involve important economic losses for main target species, like M. merluccius and Mullus barbatus, representing up to 32 and 28% of the incomes, respectively. Considering all analysed species, economic losses using the 52 mm square meshed codend would represent 27% of the incomes obtained using the current 40 mm square meshed codend in force. Despite it, transition analyses showed that the yield per recruit of the main target species would recover after two years, and even increase up to 30% and 17% for M. merluccius and M. barbatus, respectively, after the fourth year of the implementation of the 52 mm square meshed codend.
The Mediterranean fisheries are characterized by high diversity of catches, which is particularly clear for the bottom trawl, highly multi-specific, with a large amount of by-catch of both commercial and non-commercial species (Stergiou et al., 1997; Moranta et al., 2000; Lleonart and Maynou, 2003). Currently, most of the assessed stocks in the area are overexploited. Even though exploitation state of stocks seems to have improved since the 2014 peak with 88% of overexploited stocks, still more than 70% of stocks are fished exceeding biologically sustainable limits (FAO, 2022), i.e. exploited beyond the maximum sustainable yield (MSY). This overexploitation results not only in reduced stock productivity but also in low economic yields. The Multiannual Plan for the Fisheries exploiting demersal stocks in the western Mediterranean Sea (EU-MAP; Regulation (EU), 2019/1022 of the European Parliament and the Council of 20 June 2019) is aiming to reverse this situation, by restoring exploited populations above levels which can produce the MSY. To do so, the EU-MAP will apply technical measures such as the reduction of fishing effort, the establishment of closure areas and the improvement of the selectivity of the gears. In this sense, Colloca et al. (2013) showed that for most overexploited stocks, a simple fishing effort reduction “will not allow neither to maximize the stock biomass nor the fisheries yield and revenue”, being imperative a selectivity improvement. However, this improvement is challenging in the Mediterranean multi-species demersal fishery, due to the different sizes, shapes, behaviors and other characteristics of the species, as well as their different minimum conservation reference sizes (MCRS) (Lucchetti et al., 2021). For the bottom trawl fishery, the General Fisheries Commission for the Mediterranean recommended a selectivity improvement (GFCM, 2001), which was also made compulsory by the European Parliament in order to eliminate discards of undersized specimens (EU, 2013). The main characteristics affecting the selectivity of the trawl catches are not only the mesh size but also its shape, because squared meshes increase the openness of the codend when compared to diamond shaped meshes (Robertson and Stewart, 1988; Reeves et al., 1992).
The most recent regulation aiming to improve the selectivity of the bottom trawl codend is the Council Regulation (EC) No 1967/2006 of 21 December 2006 concerning management measures for the sustainable exploitation of fishery resources in the Mediterranean Sea, which replaced the 40 mm diamond mesh in the codend by a 40 mm square mesh or, at the duly justified request of the ship owner, a diamond mesh of 50 mm. Currently, the Spanish bottom trawl fleet uses 40 mm square mesh codends. This selectivity improvement involved a reduction of catches of individuals below the Minimum Conservation Size (MCRS) although it still does not guarantee that the 50% retention length is above the MCRS for most of the species (Bahamon et al., 2006; Guijarro and Massutí, 2006; Ordines et al., 2006). In the case of the European hake (Merluccius merluccius Linnaeus, 1758), one of the main target species of this fleet, the 50% retention length (L50) using 40 mm square-meshed codends is well below its MCRS: 15.2-17.2 cm vs. 20 cm, respectively (Bahamon et al., 2006; Guijarro and Massutí, 2006; Ordines et al., 2006, Baro and Muñoz de los Reyes, 2007).
Modifying the design of the extension piece by turning 90° the diamond mesh netting (T90) can also affect the selectivity of trawl nets. However, the use of T90 extensions has shown contradictory results in the Mediterranean, even for the same species. Whereas Sola and Maynou (2018) detected a reduction of catches of small-sized M. merluccius and red mullet (Mullus barbatus Linnaeus, 1758) when using T90 in the western Mediterranean, Petetta et al. (2022) did not detect such selectivity improvement for these two species when using this gear modification in the Adriatic Sea.
According to official data from 2021 (Data Collection Framework), the FAO-GFCM geographical sub-area 6 (GSA 6), northern Spain, harbors the majority of bottom trawlers of the Spanish Mediterranean, up to 420 vessels, almost the triple of those in GSAs 1 (northern Alboran Sea) and 5 (Balearic Islands) together (153 vessels). Although the landings of bottom trawlers fleet represented 33% (ranking second in importance after purse seine corresponding to 55%) of the total fish biomass landed in GSA 6, it was the most important fleet in terms of economic benefit. Bottom trawlers’ landings accounted for up to 50% of total revenues in 2019, followed by purse seiners, small scale fishery and longliners (31, 15 and 4%, respectively) (Data Collection Framework). The most important landings of the bottom trawl fleet in the GSA 6 are those of M. merluccius, representing around the 7% of total landings, although being second in terms of economic income (12%) after red shrimp (Aristeus antennatus Risso, 1816) with 21% (Martín et al., 2019). Despite the importance of the M. merluccius fishery in GSA 6, this species has a long-term history of overexploitation, with a negative trend of both spawning stock biomass and recruitment. It was diagnosed in recruitment overfishing in 2019 from an assessment using Mediterranean International Trawl Survey (MEDITS) data (Ordines et al., 2019), although recruitment seemed to recover in 2020 (GFCM, 2022).
Therefore, the aim of this work was to assess the effects of both 52 mm square meshed codend and T90 extension on the selectivity, catches and economic yields in one of the most important ports exploiting M. merluccius in northwestern Mediterranean. We evaluated the potential economic consequences of such measure, discussing on its viability for the sustainability of trawl fishery and the consequent variation in the exploitation pattern of its most important target species.
The fishing trials were conducted off the Ebro River delta (northwestern Mediterranean), between September the 11th and the 25th, 2020, onboard the F/V ‘Vicent Gras’ (length 25 m; 44.5 grt; nominal engine power 323 kw) from La Ràpita harbor (Figure 1). The vessel is equipped with ITI systems allowing to monitor, in real time, the gear geometry during the fishing operations and to estimate the effective duration of the trawls. This vessel is a typical bottom trawler that operates in the continental shelf of the area, targeting M. merluccius. In fact, the previous year it was dedicated more than 97% of the fishing days to exploit depths between 60 and 200 m, where the most important catches in terms of both biomass and economic benefit were M. merluccius and M. barbatus.
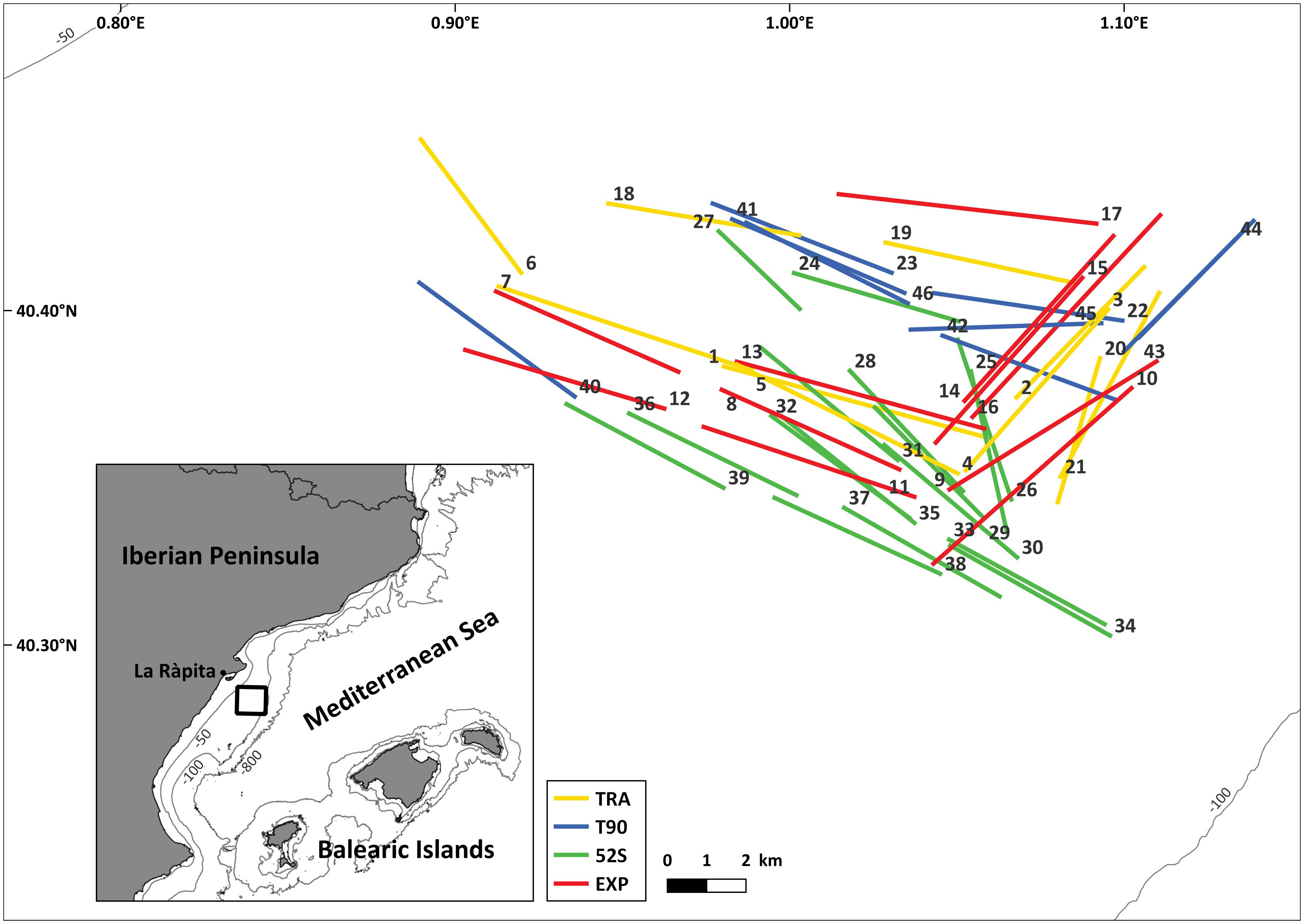
Figure 1 Study area and hauls’ positions by net configuration: traditional (TRA), traditional equipped with a 90° turned mesh in the extension (T90), traditional equipped with a 52 mm square meshed codend and a codend cover (52S), and T90 equipped with a 52 mm square meshed codend (EXP).
The traditional gear used by this fishing vessel is composed of: (i) Viking doors MX (2.6 m2 and 570 kg); (ii) wire and Nylon sweeps of 40 mm diameter and 220 m length; (iii) 50 m bridles (Nylon, and Nylon and wire 44 mm diameter in the headrope and footrope parts, respectively); and (iv) a four panel net “Quadrat” type, with 84.3 and 110.5 m headrope and footrope, respectively, made in polystyrene in the conical anterior section, with an extension piece (untapered section) of 14.10 m made of 44 mm diamond mesh (44.1 ± 0.7 mm ( ± SD)) built with 4 mm twine Nylon, and a 1.5 x 1.5 m codend made of 42 mm (42.1 ± 0.4 mm) knotted square mesh built with 3 mm twine Nylon. A schematic diagram of this net is shown in Supplementary Material Figure S1. Four different net configurations were used:
● the traditional gear equipped with a 42 mm square mesh codend (TRA);
● the traditional gear equipped with a T90 44 mm (44.1 ± 0.9 mm) diamond mesh extension (T90);
● the traditional gear equipped with a 52 mm (52.3 ± 0.5 mm) square mesh codend and a codend cover made of 18 mm diamond mesh (52S); and
● the T90 configuration equipped with the 52 mm square mesh codend (EXP).
The mesh size in the extensions and codends used in the different net configurations was checked using an OMEGA mesh gauge. The gauge was set at 50N and 20 consecutive measures were made on the wet net, selecting the meshes on the longitudinal axis of the net or perpendicularly to this axis depending on the measures were taken from diamond and square meshes or T90, respectively, following Commission Regulation (EC) No 517/2008.
An alternated hauls strategy was applied to compare catches and yields between different gear configurations. To do that, fishing trials were made under strictly commercial conditions. Hauls were alternated between control and test nets and carried out as close in time (repeated the day after or after two days) and space as possible in order to trawl the same fish populations. A total of 46 hauls between 64 and 85 m of depth with duration of 45-65 minutes were done: 10 alternated hauls with each gear type, plus 6 more hauls using the 52S net configuration. Hence, to determine selectivity parameters of the 52 mm square meshed codend, 16 hauls using the covered codend methodology were carried out. During these hauls, the 18 mm diamond mesh codend cover was attached directly to the funnel end of the net. To maintain a good flow of water and to avoid masking the codend meshes, the cover was 1.5 m wider and longer than the codend. This method has been considered appropriate where catches are not very large (Wileman et al., 1996), and it has been used in most of the trawl selectivity studies in the Mediterranean. For specifications regarding the performed hauls see Table S1.
After each haul, the crew sorted the catches between landings and discards, and further sorted landings according to commercial categories. All commercial categories and discards were counted and weighed separately by the scientific team, they also measured all the individuals of fish (total length at the lowest half cm; TL) and cephalopod (mantle length at the lowest half cm; ML) species, and commercial species of crustaceans (carapace length at the lowest mm; CL).
The biomass yields of the total catch, the main taxonomic groups and the most important commercial species (M. merluccius, Lophius budegassa, Eledone cirrhosa, Scomber scombrus, Illex coindetii, Parapenaeus longirostris, M. barbatus, Trisopterus minutus, Trachinus draco, Chelidonichthys lucerna, Squilla mantis, Citharus linguatula, Chelidonichthys gurnardus, Loligo vulgaris, Sepia elegans, Arnoglossus laterna, Scyliorhinus canicula, Lepidotrigla cavillone and Alloteuthis media, from most to less abundant) were compared among net configurations. The selected species altogether represented more than 96% of the total biomass yield in the hauls using TRA.
Catches were standardized based on the formulae:
where CPUEW is the catch per unit effort expressed as weight (W; kg) per hour of trawling, CPUEI is the catch expressed in terms of individuals (Ind) caught per hour. Trawl duration is the effective fishing time in minutes.
Mean biomass yield and its standard error were calculated for the landings and discards, considering total catch and main taxa by net configuration and pooling hauls carried out using 42 mm square meshed codend (TRA+T90) and using 52 mm square meshed codend (EXP+52S). Pooling was done after checking no differences attributable to selectivity were detected. For those species with MCRS in the study area, the percentage of individuals below the MCRS retained in the codend was calculated for each net configuration and also pooling data by codend mesh size (TRA+T90 and EXP+52S). For the 52S configuration, the data collected from the cover allowed to calculate the percentage of individuals below the MCRS in relation to the total number of individuals entering the net (retained and escaped), and the percentage of individuals below the MCRS escaped in relation to the total number of undersized individuals. The species included in these analyses and their MCRS were M. merluccius (20 cm TL), L. budegassa (30 cm TL), T. trachurus (15 cm TL), M. barbatus (11 cm TL) and P. longirostris (20 mm CL).
In order to compare the percentages of individuals below the MCRS retained in the codend between net configurations, the Student’s t-test was applied using R software (R Core Team, 2022). The comparisons included: TRA vs. T90, TRA vs. 52S, EXP vs. 52S and TRA+T90 vs. EXP+52S.
To assess the influence of the T90 extension on the size of catches, the standardized length frequency distributions (LFDs) of the main commercial species were analyzed. The catch efficiency (at length) was assessed by means of the polynomial regression generalized linear mixed model (GLMM, with hauls as random intercept to account for between haul variability) (Holst and Revill, 2009). According to Robert et al. (2020), observations outside 95 quantiles of the length distribution of each species were excluded from the analysis to reduce the influence of outliers on the fits. The probability of a fish being retained by the test net (the net with T90 extension) relative to the control net (net without T90 extension) follows from:
where and are species subsampling ratios in each test and control of the h haul (although in our case they have not been used as all fishes caught were measured), and k is the order of the polynomial to be tested. A binomial error distribution is used to calculate the probability of the number of fish caught in the test net given they enter both gears. A probability of 0.5 corresponds to equal catches on both gears to be tested. According to Holst and Revill (2009), the best model is the minimal degree polynomial curve that captures the main trends indicated by the observed proportions, while the third order polynomial followed by subsequent reductions until all terms showed significance would be adequate for most cases, although in some instances a 1st or 2nd order would be enough. The best model describing the retention probability was chosen based on the Akaike’s Information Criteria (AIC; Akaike, 1974). According to the parsimony rule, when two models showed AIC values difference less than 5 units, the best model was considered the one with less degree of freedom (i.e., the simpler model). We then estimated the 95% Efron’s Confidence Intervals (Efron, 1987), by accounting for within- and between-hauls variation (Millar, 1993). Confidence intervals were calculated by means of a double bootstrap, resampling first hauls, then resampling fish within hauls and fitting the chosen model to the resampled data. This procedure was repeated 1000 times. Catch comparison analysis was performed using the selfisher R package (Brooks et al., 2022; R Core Team, 2022).
Selectivity parameters of the 52 mm square meshed codend were calculated using the data collected during the hauls applying the covered codend method. The retention probability of individuals entering in the codend (SL) by length class (L), i.e. the number of individuals of a given length class retained in the codend divided by the total number of individuals of that size class retained in both codend and cover pooled together, was modeled using the logistic curve (testing different link functions: logit, probit and cloglog) or the Richard’s curve (Wileman et al., 1996). The best fitting curve and model for every selected species was chosen taking into account the lowest AIC value. Prior to apply the logistic or Richard’s curve, the rule by Krag et al. (2014) to not consider the hauls with less than 10 individuals between codend and cover was applied.
The 50% retention length (L50, length at which the probability of being retained in the codend is 50%; Wileman et al., 1996) and the selection range (SR: L75−L25, length range at which the retention probability increases from 0.25 to 0.75) were calculated from the expressions:
where S1 and S2 are the parameters derived from the selected model.
Selectivity parameters and uncertainty (calculated by means of double bootstrap 999 times) were estimated using the R package Selfisher v. 1.0 (Brooks et al., 2022).
We applied a theoretical approach to assess the economic consequences of the implementation of the 52 mm square meshed codend, as compared to the currently used 40 mm square meshed codends. To do that, only the 52S net configuration hauls were used (16 hauls). This net configuration allowed us to simulate, considering that codend and cover catches represent the total population of the studied species in the area swept by the net, the theoretical catches that would be obtained using 40 mm and the 52 mm square meshed codends. These theoretical catches were calculated for the most important species (representing 83% of the total catches, see Supplementary Material Tables S2, S3) by applying the probability of retention at a given size (selectivity curves) with the 40 mm and 52 mm square meshed codends, obtained from the bibliography (see Supplementary Material Tables S4) and the present work, respectively. To do so, the probability of retention at a given size was multiplied by the total number of caught individuals in that size including codend and cover pooled together. Then, taking into account that some species can have different prices depending on their size, individuals theoretically retained with each codend were sorted by size categories according to the size ranges corresponding to each of the size categories in which fishermen from La Ràpita distribute fish (this ranges were measured during their normal fishing activity, see Supplementary Material Tables S5). The theoretical weight of each size category in each haul by mesh size was determined applying the weight-length relationship to each individual within each category. Finally, the biomass yield of each commercial category was standardized to one hour of effective trawling and this was multiplied by the corresponding price per kg at the first sale of each category (see Supplementary Material Tables S5, S6 to obtain the economic yield (€/h), both by size category and by species (pooling €/h of all size categories of a particular species).
The Student-t test was used to assess significant differences in biomass and economic yields between both mesh sizes in the codend, both at species level and size category within a species.
Economic losses (€/h) were calculated as the difference between the theoretical economic yield obtained using 40 mm and 52 mm square meshed codends for every selected species. The percentage that this economic yield loss represented in relation to the economic benefits theoretically obtained using 40 mm square meshed codend was also calculated. This percentage was used to calculate the expectable economic loss resulting by a change from 40 mm to 52 mm square meshed codend in the M. merluccius fishery exploited by the trawl fleet from La Ràpita.
To do so, the fishing days dedicated to the fishing strategy exploiting the M. merluccius fishery were determined by means of cluster analysis of the official daily sales bills (one bill per day and boat, including the weight and price of all landed commercial categories) recorded in the fish market from La Ràpita in 2019. In this cluster analysis, the Bray-Curtis index was used as a between-daily sales bills similarity measure and the unweighted pair-group method with arithmetic mean was applied to link daily sales bills into clusters. Previously to the cluster analysis, those commercial categories (not taking into account size categories) representing less than 1% of the total biomass landed in 2019 were eliminated. Landings analyzed represented up to 91% of the landed biomass. The groups identified in the cluster analysis were assigned to a fishing strategy (named after the approximate bathymetric range) according to their composition, the bathymetric distribution of the main species, and the expert knowledge of the fishermen from La Ràpita. The sum of the number of daily sales bills included in each cluster were considered as the fishing days dedicated to each fishing strategy. For each fishing strategy we calculated total landed biomass and economic income by commercial category. The cluster analysis was performed with R software.
We used the transitional analysis implemented in the VIT software (Lleonart and Salat, 1992), widely applied for stock assessment in the Mediterranean, in order to assess the effect, at short and medium term, of a change from 40 mm to 52 mm square meshed codend on the yield per recruit (expected life time yield per fish recruited into a stock, Y/R) of the main target species of the M. merluccius fishery in the study area, sited in the GSA 6. First, using this software, we ran a Virtual population analysis (VPA) for the three target species currently assessed by the GFCM in the GSA 6: M. barbatus, M. merluccius and P. longirostris. The VIT software accepts length frequencies as input data to develop a length cohort analysis (LCA) working with pseudocohorts. Although the LCA assumes a steady state, it provides a more general and synthetic outlook of the state of the fishery (Lleonart, 1993). Two files are required to develop this model: (i) the length frequency distribution of the species (from official catch and discards data and data collected by the observers oboard the commercial fleet in the GSA6), which the software converts into age distribution; and (ii) its biological parameters (same data used as in GFCM for 2019 assessments; GFCM, 2021). Once the VPA had been run, transitional analyses were conducted. For this, the probability of retention of the 40 mm and 52 mm square meshed codends by age class was calculated as the mean between the minimum and the maximum probability of retention for the size classes included in every age class. Then, considering the current fishing mortality (F) using 40 mm square meshed condend for every age class, calculated in the VPA, we extrapolated a theoretical F for every age class if the fleet used 52 mm square meshed codends. To do so, we multiplied the current F by a factor calculated dividing the probability of retention using a 52 mm square meshed codend by that of the 40 mm square meshed codend. Based on these results, transition and yield per recruit analyses for the next 20 years were conducted for each of the three target species mentioned above.
In general, no significant differences were found for the biomass yields between net configurations with the same codend mesh size (i.e. TRA vs. T90 and EXP vs. 52S; Table 1 and Table S2). The only differences comparing TRA and T90 appeared comparing total catches (pooling commercial and discards) for S. elegans and A. media with higher yields using TRA than T90, and for the discards of L. cavillone, with higher yields with T90 than TRA. When comparing EXP with 52S, higher yields were observed using 52S in the case of M. barbatus. By contrast, higher yields were seen using EXP for commercial and discards of benthic species, total, commercial and discard yields of I. coindetii, and total and commercial yields of S. scombrus. In the case of pooled data by codend mesh size, all detected differences were due to higher yields when using the 42 mm square meshed codend (TRA+T90) than using the 52 mm square meshed codend (EXP+52S), including total catches and total discards, total catches of osteichthyes, total and commercial catches of L. cavillone, M. merluccius, T. minutus, I. coindetii, Alloteuthis media and Arnoglossus laterna, and discards of M. merluccius (Table 1).
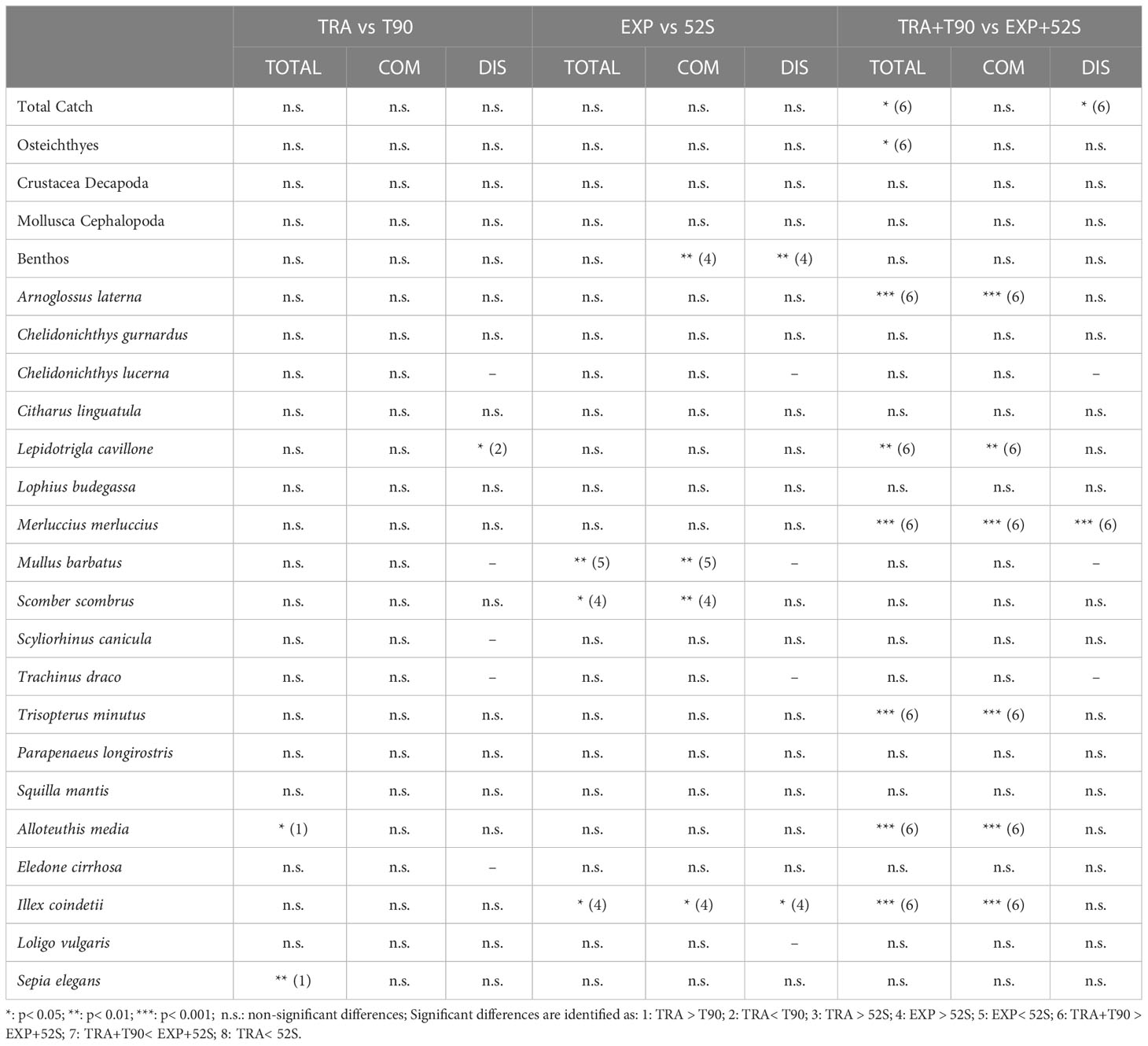
Table 1 t-Student analyses comparing the biomass yields of the main taxonomic groups and target and by-catch species, for the total, commercial (COM) and discarded (DIS) fractions obtained using the different net configurations: traditional (TRA), traditional equipped with a 90° turned mesh in the extension (T90), traditional equipped with a 52 mm square meshed codend and a codend cover (52S) and, T90 equipped with a 52 mm square meshed codend (EXP).
Trachurus was the species with the highest percentage of individuals below MCRS in the retained catch using all net configurations (ranging between 79 and 95%; Table 2). However, it ranks second after M. barbatus and P. longirostris (with 100% and 98% of individuals below the MCRS escaping) in the percentage of individuals below the MCRS that escape from the codend in the 52S net configuration, up to 95%. For this species the percentage of individuals below MCRS in the retained catch showed no significant differences either between TRA and T90 net configurations nor between EXP than 52S, although significant differences (p<0.01, t = -3.12) were found when pooling data according to codend mesh size, with higher percentages using TRA+T90 than 52S+EXP. In the case of M. merluccius, up to 90% of the individuals below MCRS escaped from the codend when using the 52S net configuration. For this species the percentage of retained individuals below MCRS showed no significant differences between TRA and T90 net configurations but values were significantly higher using EXP than 52S (p<0.001, t=3.75), and significantly higher values were observed for TRA+T90 (56%) than for 52S+EXP (36%) when pooling data according to codend mesh size (p<0.001, t=6.01). No individual of L. budegassa escaped from the codend of the 52S net configuration. No significant differences were detected for the percentage of retained individuals below MCRS among the different net configurations, with values of 56 and 54% for TRA+T90 and 52S+EXP, respectively. For M. barbatus, individuals below the MCRS only appeared in the retained catch of the TRA and T90 net configurations, representing very low percentages, 0.1 and 0.3, respectively. Finally, for P. longirostris, up to 98% of the individuals below MCRS escaped from the codend when using the 52S net configuration. For this species the percentage of individuals below MCRS in the retained catch showed no significant differences between TRA and T90 and between 52S and EXP, but significantly higher values (p<0.05, t=2.38) were detected using TRA+T90 (7.3%) than using 52S+EXP (3%) (Table 2).
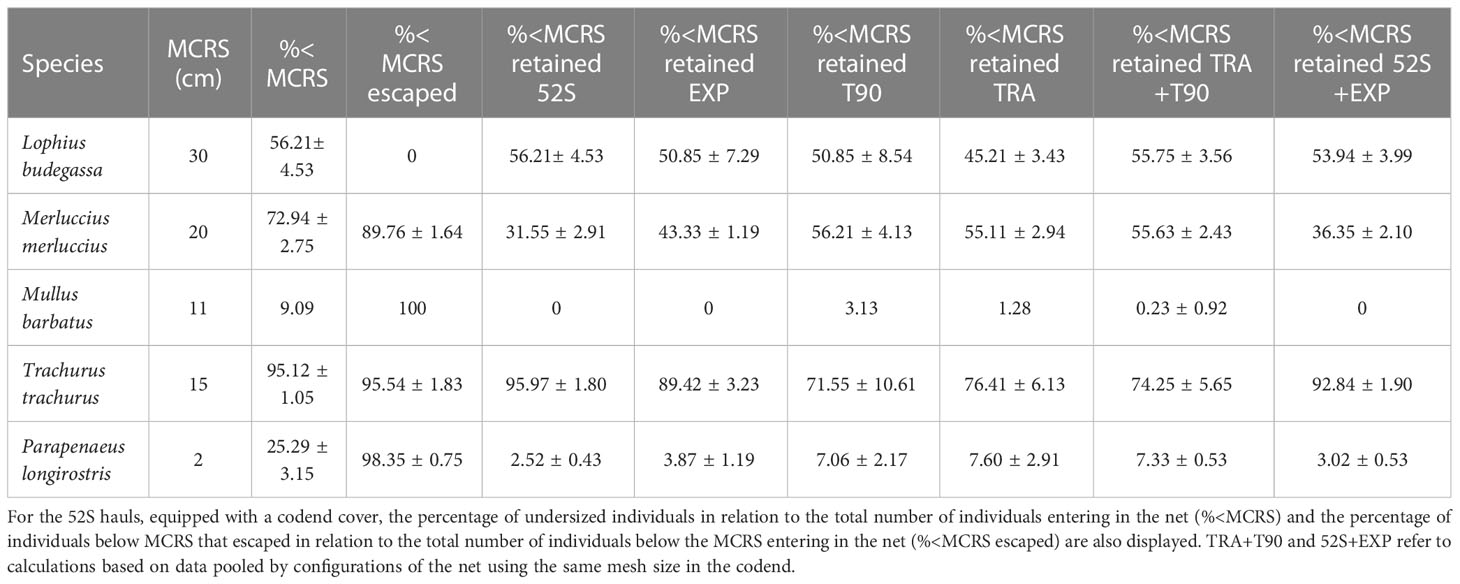
Table 2 Minimum conservation reference sizes by species and percentage of undersized retained individuals in relation to the total number of retained individuals (%<MCRS retained) by net configuration (traditional (TRA), traditional equipped with a 90° turned mesh in the extension (T90), traditional equipped with a 52 mm square meshed codend and a codend cover (52S) and, T90 equipped with a 52 mm square meshed codend (EXP)).
The catch comparison analysis to assess the effect of the T90 extension showed contrasting results between net configurations. The results obtained for M. merluccius, M. barbatus and P. longirostris, the three main species caught during the surveys, did not display differences in the selection of lengths between TRA and T90 net configurations (Figure 2, Table 3) as shown by the overlap of the 95% confidence intervals area with the 0.5 probability line. Only for P. longirostris a small size interval (2 mm among the whole length range) seems to be significantly different between the two nets, but this is a negligible difference in relation to the general pattern of the model and the generally large confidence intervals. Conversely, the comparison between EXP and 52S net configurations showed EXP appeared to be much more efficient than 52S at catching individuals below the MCRS in the case of M. merluccius. In the case of M. barbatus, with very few individuals below the MCRS, EXP caught 80% less of individuals than 52S throughout the whole size range caught during the survey, while for P. longirostris, no clear differences were detected between EXP and 52S for individuals below MCRS, although the EXP net configuration appeared less efficient than 52S for catching the largest ones. For I. coindetii, T90 was less efficient than TRA to catch large individuals, but this difference was not noticed when comparing EXP and 52S. The contrary was obtained in the case of T. minutus, with a higher efficiency to catch larger individuals when using EXP than 52S, but no such difference when comparing TRA and T90.

Figure 2 Catch comparison results for Merluccius merluccius, Mullus barbatus and Parapenaeus longirostris between the different net configurations (traditional (TRA), traditional equipped with a 90° turned mesh in the extension (T90), traditional equipped with a 52 mm square meshed codend and a codend cover (52S) and, T90 equipped with a 52 mm square meshed codend (EXP)) equipped with the same codend mesh size. Solid grey line corresponds to the length frequency distribution of the net mounting the T90 extension while the dashed grey line corresponds to the length frequency distribution of the control net (TRA on the left, and 52S on the right). For Merluccius merluccius and Parapeneaus longirostris, the point grey line corresponds to the minimum conservation reference size of the species. The grey area is the confidence interval at 95% of the modelled catch comparison (black solid line). Dots correspond to the individual proportions retained and the size is proportional to the total individual of each size class. The point-dashed horizontal line at 0.5 represents the point at which both gears have equal catch rates.
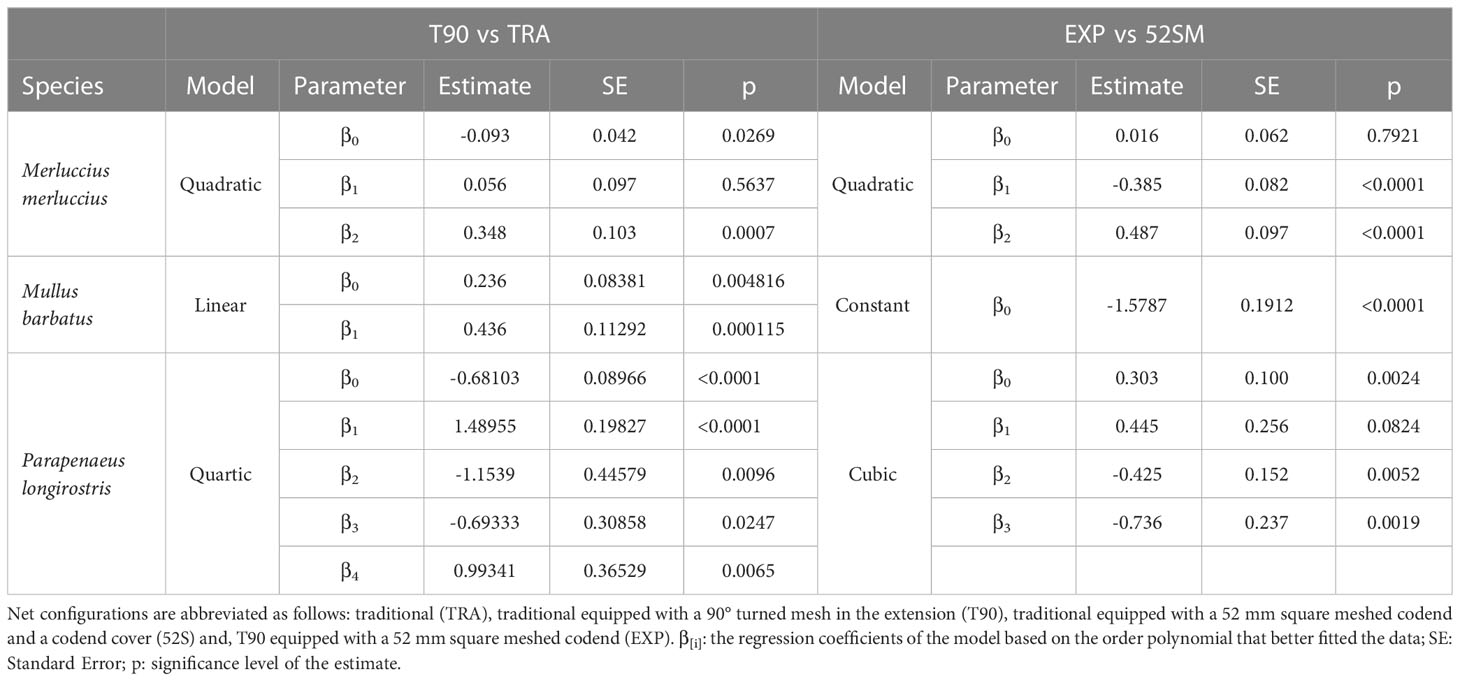
Table 3 Catch comparison of the Generalized Linear Mixed Model parameters from trials with nets mounting the T90 extension.
Selectivity parameters could be calculated for 6 fish species, 2 decapod crustaceans and 4 cephalopod mollusks (Table 4). Selection curves for these species can be found in Supplementary Figures S4, S5. The retention probability at size was better fitted using a general logistic function for most of the species, except for M. merluccius, M. barbatus and P. longisrostris, which were better fitted using Richard’s curve. The L50 calculated for the 52 mm square mesh (Table 4) was higher than the L50 reported for the 40 mm square mesh for all species analyzed (see Supplementary Material Table S4). Among the rounded fishes, T. draco and M. merluccius showed the largest values of L50, 24 and 22.2 cm in TL, respectively, whereas L. cavillone showed the smallest one, 11.3 cm TL (Table 4). The flatfish C. linguatula showed an L50 of 13 cm TL. The two crustacean species, P. longirostris and S. mantis, showed similar L50 values of 22.6 and 19.7 mm CL, respectively, whereas the octopus E. cirrhosa and the squid I. coindetii had L50 values of 8.5 and 10.4 cm ML, respectively (Table 4).
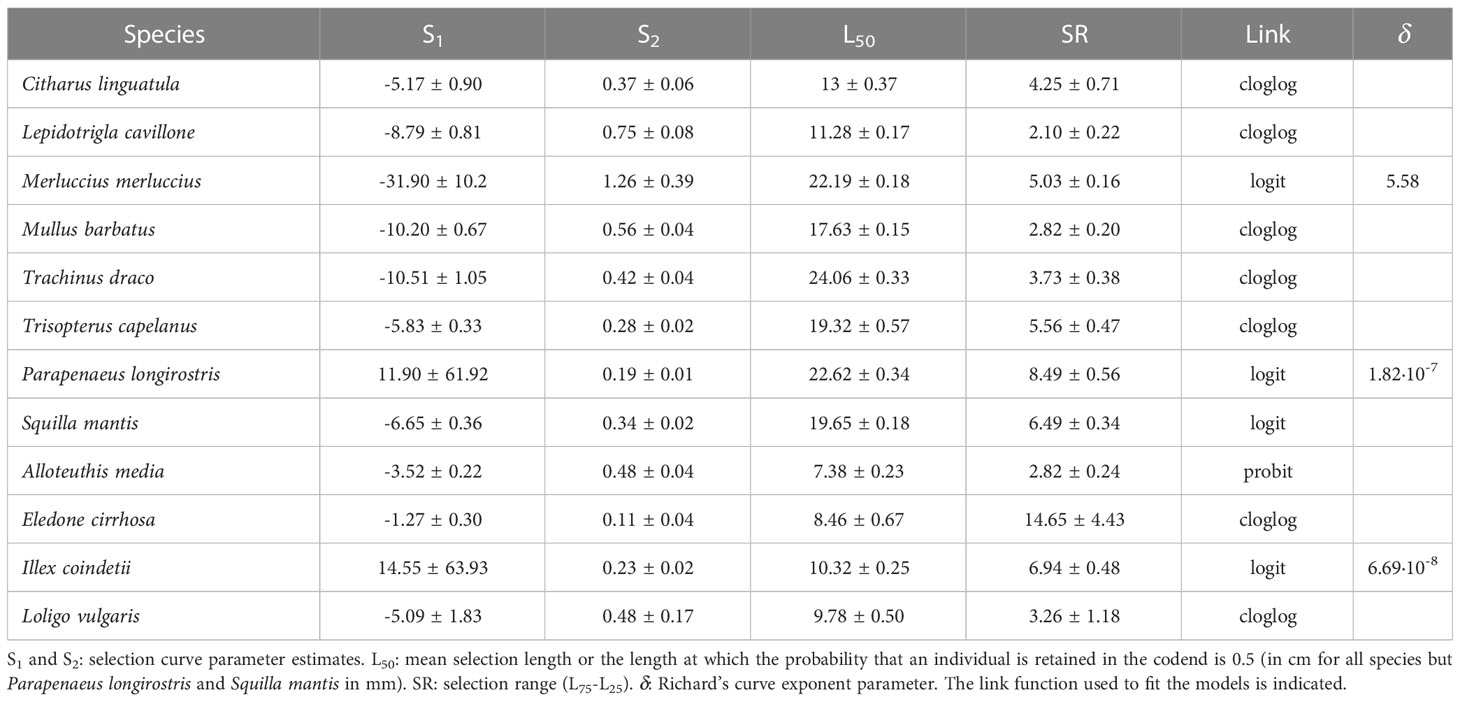
Table 4 Selectivity parameters (mean ± standard error) of the 52 mm square meshed codend.
The species selected for the comparison of economic yields between 40 and 52 mm square meshed codends represented altogether the 86.4% of the total theoretical economic yield obtained during the survey. In general, biomass and economic yields were higher using 40 mm square meshed codend than the 52 mm square meshed codend (Figure 3). These differences were significant for the smallest categories of M. merluccius and M. barbatus (Figure 3) and the two size categories of T. minutus and I. coindetii, whereas no significant differences were observed for any of the two size categories of P. longirostris. Regarding species that do not have size categories, all of them also showed higher biomass and economic yields using the 40 mm than the 52 mm square meshed codend, although significant differences were only found for A. media (Figure 3).
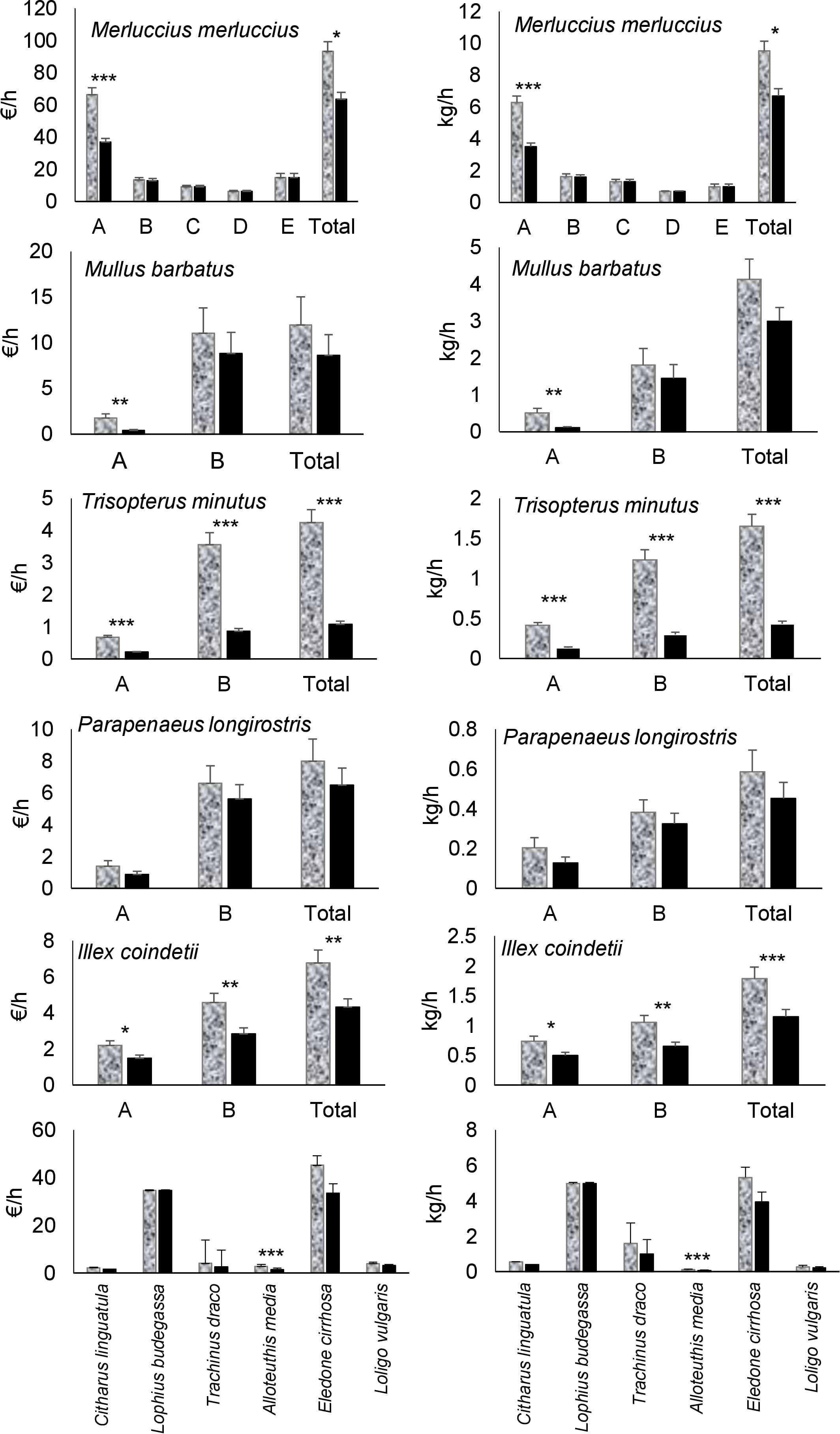
Figure 3 Mean (± standard error) economic and biomass yields of the 40 mm square meshed codend (in grey) and the 52 mm square meshed codend (in black), estimated from the hauls carried out using the 52S net configuration (traditional net equipped with a 52 mm square mesh codend and a codend cover) for the main species. Five of these species have size categories: for Merluccius merluccius A (20-25 cm), B (26-29 cm), C (30-37 cm), D (38-42 cm) and E (>42 cm); for Mullus barbatus A (11-16 cm) and B (>16 cm); for Trisopterus minutus A (11-14 cm) and B (>17 cm); for Parapenaeus longirostris A (<2.7 cm) and B (>2.7 cm); and for Illex coindetii A (<10 cm) and B (>10 cm). Results of the t-Student tests comparing the yields between the two mesh sizes are also presented. Significant differences identified as *: p< 0.05; **: p< 0.01; ***: p< 0.001.
Considering the total catch, the mean loss of economic yields with the implementation of the 52 mm square meshed codend was estimated in 56 €/h, which represented 26% of the total estimated economic yield when using 40 mm square meshed codend. Lophius budegassa was the only target species that did not show any economic loss when using 52 mm square meshed codend. The species representing the highest economic loss was M. merluccius (30 €/h, up to 32% of the benefit obtained with this species; Figure 4). Although the losses for T. minutus were low (3 €/h), they represented losing up to 74% of the benefit obtained with this species. Losses for E. cirrhosa ranked second after M. merluccius, up to 12 €/h, representing 26% of the benefit obtained with this species. The rest of the species represented lower losses, all below 5 €/h and with percentage losses related to each species ranging between 50 and 20% (Figure 4).
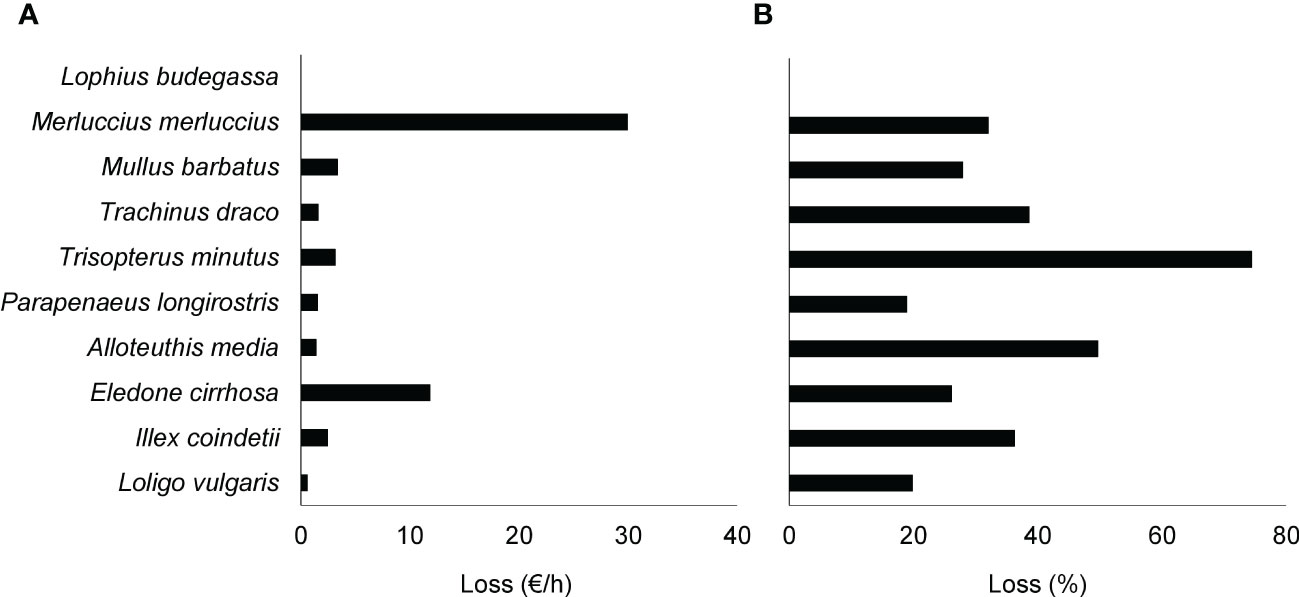
Figure 4 (A) Absolute and (B) percentage economic losses, expected for the main commercial species due to the change of mesh size in the codend from the current 40 mm square meshed codend to the 52 mm square meshed codend; estimated from hauls carried out using the 52S net configuration (traditional net equipped with a 52 mm square mesh codend and a codend cover).
Almost half of the fishing days in La Ràpita were dedicated to the fishing strategy (FS) that exploits the deep shelf (60-200 m depth; Figure S3 and Table 5). This FS also accounts for 56% of the biomass caught and for 57% of the earnings (Table 5) in this port. Within this FS, M. merluccius and M. barbatus were the species that represented the highest percentage of biomass, 10% and 9%, respectively, and the highest percentage of economic incomes, 17% and 12%, respectively (Table 6). For each of the taxa analyzed, the economic income from this FS represented at least half of the total incomes obtained from all FS conducted in La Ràpita (Table 6). Taking into account the composition of the annual catches of this FS in La Ràpita in 2019, economic losses would represent up to 27% of revenues obtained using the 40 mm square meshed codend.
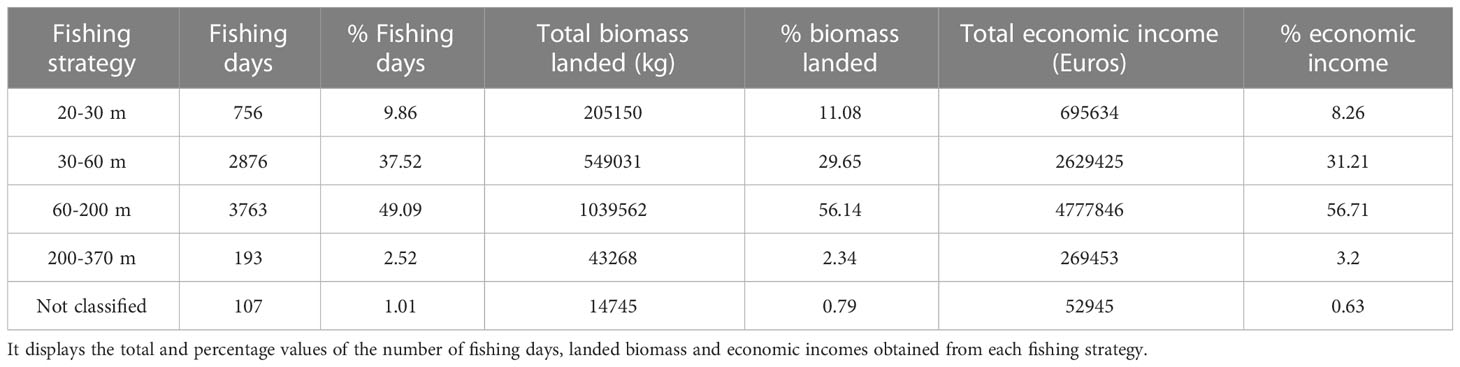
Table 5 Summary of the importance in terms of fishing days, landings and economic incomes of the different fishing strategies carried out by the bottom trawl fleet in La Ràpita in 2019.
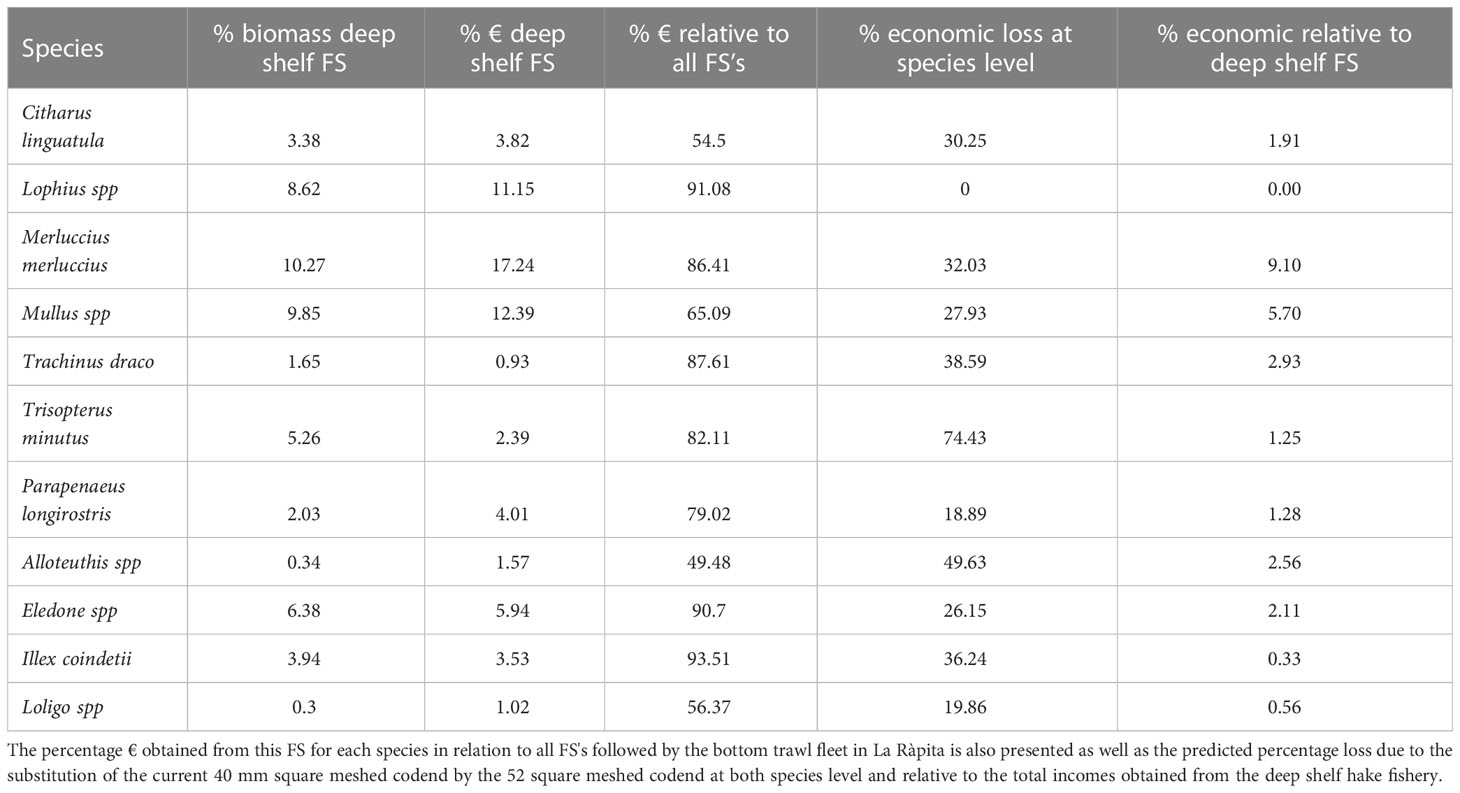
Table 6 Summary of the percentages represented by the most important commercial species in terms of both landed biomass and economic benefit (€) obtained in the fishing strategy (FS) exploiting the deep shelf (60-200 m depth) M. merluccius fishery from La Ràpita in 2019.
Transition analyses showed a pronounced initial decrease of the yield per recruit (Y/R), almost 2 g/individual, during the first year after the implementation of the 52 mm square meshed codend for both M. merluccius and M. barbatus, but an almost negligible effect for the Y/R of P. longirostris. Two years after this implementation, the current Y/R of M. merluccius and P. longirostris would be recovered (Figure 5), and half a year later it would also be recovered for M. barbatus. Moreover, after the fourth year, the (Y/R) would increase by 30% compared to the current value in the case of M. merluccius, from the current Y/R of 14.3 g/ind to 18.7 g/ind, whereas for M. barbatus, after four years and a half the Y/R would increase up to 20%, from the current Y/R of 10 g/ind to 12 g/ind. Regarding P. longirostris, its Y/R would not increase after the recovery, remaining at 2 g/ind. The estimated Y/Rmax with the currently used 40 mm square meshed codends is 27.5, 10 and 2 g/ind for M. merluccius, M. barbatus and P. longirostris, respectively.
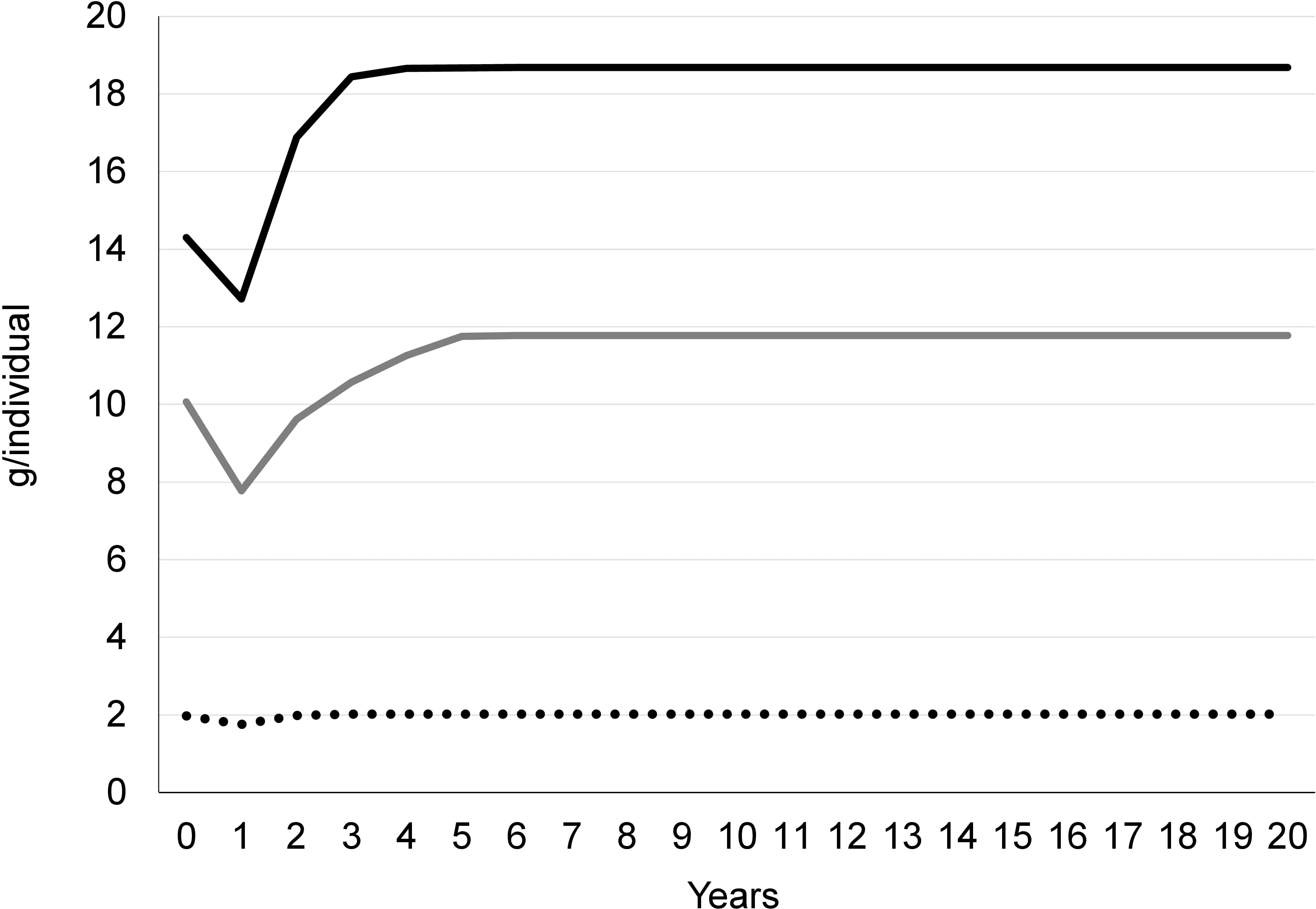
Figure 5 Transition analysis results showing the evolution of the yield per recruit (g/individual) in the northeastern Spanish Mediterranean (geographical sub-area 6; GSA 6) during the 20 years following the implementation of the 52 mm square meshed codend, assuming no significant changes in the fishing effort. Black solid line: Merluccius merluccius; grey solid line: Mullus barbatus; and black dotted line: Parapenaeus longirostris.
Improvement of the selectivity in the Mediterranean bottom trawling is the key to correct the current generalized overfishing and to ensure the conservation of marine fishing resources (Colloca et al., 2013). In this work we have assessed the effects on fishing yields and selectivity of two net modifications, the T90 extension and the 52 mm square mesh in the codend, in a fishery targeting M. merluccius in the northwestern Mediterranean.
The results have shown that the introduction of T90 did not contribute to improve the selectivity of any of the target species of the fishery. Moreover, in the case of M. merluccius, catch comparisons performed between TRA and T90 net configurations and between 52S and EXP showed the same trend, with higher efficiency at catching small individuals when using T90, although this trend was only significant when comparing 52S and EXP. For M. barbatus, results were contrasting when comparing TRA and T90, showing no significant effect of T90 on selectivity, but showing a lower efficiency to catch this species throughout the whole size range when using EXP (equipped with T90) than 52S. Mullus barbatus showed low catches, particularly using the 52 mm square meshed codend, which increased substantially its 50% retention length. These low catches, paired to the relatively high between-haul variability of the catch (see Supplementary Material Tables S2, S3), could be the reason for these contrasting results, perhaps reflecting more the variability of this species between hauls, despite both temporal and space proximity of the samples, than the effect of T90. In any case, our results for M. merluccius are similar to those reported by Petetta et al. (2022), who did not find any indication of improvement of selectivity of M. merluccius when comparing the standard extension of 44 mm diamond mesh with a T90 extension piece of the same netting in the Adriatic Sea bottom trawl fishery. Both works contrast with Sola and Maynou (2018), who found a significant reduction of individuals below the MCRS of M. merluccius and small individuals, although larger than the MCRS, of M. barbatus, when comparing a 53 mm diamond mesh extension piece with a T90 extension piece made of 50 mm diamond mesh, also in the north western Mediterranean M. merluccius fishery, close to our study area (off Blanes at the north of GSA 6). Perhaps one explanation for the contrasting results could be related to the 6 mm larger mesh size used in the extension piece tested in Sola and Maynou (2018), compared to the 44 mm diamond mesh used in the present study and Petetta et al. (2022). Maynou et al. (2021), in the same M. merluccius fishery than Sola and Maynou (2018), also reported a selectivity improvement for M. merluccius and M. barbatus using 50 mm diamond mesh mounted in T90 panels at two different positions, front and back of the extension piece, when compared to a standard extension piece made entirely of 53 mm diamond mesh, with more evident differences when the panel was placed closer to the codend. Although in Sola and Maynou (2018) and Maynou et al. (2021) the standard extension piece was also made of larger diamond meshes than in the present work and in Petetta et al. (2022), the diamond mesh stretches during the towing, due to the pressure exerted by the tow and the accumulation of the catch in the codend, reducing the openness of the mesh to a greater extent than using T90, an orientation of the net that allows to maintain meshes more open than the diamond shape (Wienbeck et al., 2011). This fact could have made negligible, or similarly low, the selection through the diamond mesh extension (standard) in all these studies, independently of the mesh size difference (44-53 mm), while the 6 mm larger meshes could have been enough to significantly improve the selectivity of the T90 in Sola and Maynou (2018) and Maynou et al. (2021). The T90 made of 44 mm tested in the present work and Petetta et al. (2022) simply is too small once stretched as to involve a significant selectivity improvement.
Nevertheless, Maynou et al. (2021) showed that the improvement in selectivity was not general, and that even for morphologically similar species, like M. barbatus and M. surmuletus, the results using T90 can be completely different, i.e. no selectivity improvement was detected for the latter species. On the other hand, when modifications of the net are done in the codend, the improvement is more generalized among the different species. The simple modification, introduced by the Regulation (EC) 1967/2006, of the mesh geometry from diamond to square mesh without changing the mesh size resulted in an increase of the 50% retention length of the most important target and bycatch commercial species in both continental shelf and slope fisheries (Bahamon et al., 2006; Guijarro and Massutí, 2006; Ordines et al., 2006). The larger effect on selectivity of codend modifications compared to modifications in the rest of the net is expectable given that the codend has long been known as the more relevant part of the bottom trawl nets where selectivity occurs. Studies conducted during early 1960’s already demonstrated that the great majority of small fish susceptible to escape from the net does it through the codend meshes (Clark, 1963). Moreover, most fish that escapes from the codend “do so from very near the end” (Beverton, 1963). Our results confirm the effectiveness of codend modifications for improving selectivity. For all the species for which the selectivity parameters could be calculated for the 52 mm square meshed codend, there was an improvement of the selectivity when compared to the currently used 40 mm square meshed codends (Table 4 and Table S4).
Whereas the 40 mm square meshed codend was already enough to achieve a 50% retention length (L50) higher than the MCRS for both M. barbatus and P. longirostris, it failed in the case of M. merluccius for which the L50 (13-16 cm; e.g. Bahamon et al., 2006; Guijarro and Massutí, 2006; Lucchetti, 2008) was still below its MCRS (20 cm). This contradiction in the management of the bottom trawl fishery that allows a minimum mesh size in the codend which L50 is much smaller than MCRS would be resolved using the 52 mm square meshed codend, whose L50 for M. merluccius was 22.2 cm and allowed to escape up to 90% of the individuals below the MCRS. These selectivity improvements also allowed to increase the L50 of M. barbatus (17.6 cm) above its size at firs maturity (12-13 cm; Cherif et al., 2007; Carbonara et al., 2015), although for M. merluccius and P. longirostris (L50 = 22.6 mm), with sizes at first maturity of 32.7 and 35.8 cm (Oliver, 1993; Recasens et al., 2008) and 25.6 and 28.5 mm (females; García-Rodríguez et al., 2009; Guijarro et al., 2009), respectively, this is still far to be reached.
However, the selectivity improvements obtained using 52 mm square meshed codend come along with important decreases in the landed biomass and the economic yields when compared to the 40 mm square meshed codend. The simulations of the theoretical catches obtained using 40 and 52 mm square meshed codends showed important and generalized losses in the landings and economic incomes obtained with the most important commercial species. These losses are particularly important for M. merluccius and M. barbatus, together representing up to 30% of the total incomes obtained from the M. merluccius fishery in the study area, for which up to 32 and 28% of the income obtained with each species, respectively, would be lost due to this increase of the mesh size in the codend. Overall, taking the most important commercial species into account, our simulations anticipate economic losses reaching up to 27% of the economic income currently obtained from the bottom trawl hake fishery in the study area using the 40 mm square meshed codend. These losses are even higher than those estimated in the GSA 6 as results of the previous change in the geometry of the 40 mm meshed codends from diamond to square, introduced by the Regulation (EC) No 1967/2006. That selectivity improvement was estimated to cause losses of around 10% of the yield per recruit for M. merluccius in the first year, with a sharp recovery and an increase up to 55% after the fifth year (Bahamon et al., 2007). In practice, this improvement of the yield per recruit never happened in the GSA 6, and after 2006 the exploitation state of M. merluccius got worse, showing a clear decreasing trend in both recruitment and spawning stock biomass in the successive annual assessments of the exploitation state of this species, until it was diagnosed in recruitment overexploitation (Ordines et al., 2019). Similarly to Bahamon et al. (2007), our results showed that yield per recruit for M. merluccius, M. barbatus and P. longirostris would be recovered, assuming fishing effort do not increase markedly, after 2-2.5 years from the implementation of the 52 mm square meshed codend, and increases by up to 30 and 20% are predicted after four years of implementation for M. merluccius and M. barbatus, respectively. This means that just considering the recovery of these three species, up to 62% of the economic losses would be restored, and it should be expected that other species could experience the same trend.
It seems not plausible achieving the MCRS for all species without compromising the economic sustainability of this fishery. This is the case of one of the target and most economically important species in the study area, L. budegassa, a species for which the increase in mesh size tested in the present work did not represent any possibility of escapement for the individuals caught below MCRS. Similarly, it seems complicated to increase selectivity of the nets to a level that could guarantee the L50 is above the size at first maturity, even for the most important target species (e.g. M. merluccius). However, at least an effort should be made to attempt that the majority of species are managed correctly, and the 52 mm square meshed codend could contribute to it. Nonetheless, the short term predicted decrease of catches and the economic losses involved are so important after an eventual implementation of this codend, that it seems to be unsustainable for the bottom trawl fleet, particularly in the current context of rising operational costs mainly due to fuel prices, unless a subsidy policy is put in place in order to ease losses until yields are recovered.
Clark (1963) concluded that “In terms of managing fisheries this means that control of the codend alone may be necessary, as the escapement within the selection range is, in fact, higher for the codend than for the total remaining area of the trawl. Fish prevented from escaping through the forward parts of the net would still be able to escape from the codend in all probability”. Our results agree with this conclusion and indicate that modifying the codend mesh size is a far more effective way to improve selectivity of the bottom trawl nets than modifying the extension piece.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
FO, EM and BG conceived the research. MV, MF, MB-S and FO collected data. CV, FF-P, SJ and FO analysed data. FF-P, EM and FO wrote the article with the support of CV, MV, MF, BG, SJ and MB-S. FO was the scientific responsible of the research. All authors contributed to the article and approved the submitted version.
This study was conducted with the financial support of the European Commission i.e. IMPLEMED project: “Improving the selectivity of trawl gears in the Mediterranean Sea to advance the sustainable exploitation pattern of trawl fisheries” (Specific contract: EASME/EMFF/2019/1.3.2.6/01/SI2.818717-SC04).
The authors would like to thank the crew of the F/V ‘Vicent Gras’, whose involvement was essential for carrying out the sea trials. The collection of official landings and effort data has been co-funded by the EU through the European Maritime and Fisheries Fund (EMFF) within the National Program of collection, management and use of data in the fisheries sector and support for scientific advice regarding the Common Fisheries Policy. Authors would also like to thank Pau Comí, for his work developing the schematic diagram of the net.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1035448/full#supplementary-material
Akaike H. (1974). A new look at the statistical model identification. IEEE Trans. Automat. Contr. 19, 716–723. doi: 10.1109/TAC.1974.1100705
Comission regulation (EC) no 517/2008 of 10 June 2008 laying down detailed rules for the implementation of council regulation (EC) no 850/98 as regards the determination of the mesh size and assessing the thickness of twine of fishing nets. Off. J. Eur. Union L 151/5.
Council regulation (EC) no 1967/2006 of 21 December 2006 concerning management measures for the sustainable exploitation of fishery resources in the Mediterranean Sea, amending regulation (EEC) no 2847/93 and repealing regulation (EC) no 1626/94. Off. J. Eur. Union L 409/11.
Regulation (EU) no 1380/2013 of the European parliament and of the council of 11 December 2013 on the common fisheries policy, amending council regulations (EC) no 1954/2003 and (EC) no 1224/2009 and repealing council regulations (EC) no 2371/2002 and (EC) no 639/2004 and council decision 2004/585/EC. Off. J. Eur. Union L 354/22.
Regulation (EU) 2019/1022 of the European parliament and of the council of 20 June 2019 establishing a multiannual plan for the fisheries exploiting demersal stocks in the western Mediterranean Sea and amending regulation (EU) no 508/2014. Off. J. Eur. Union L 172/1.
Bahamon N., Sardà F., Suuronen P. (2006). Improvement of trawl selectivity in the NW Mediterranean demersal fishery by using a 40 mm square mesh codend. Fish. Res. 81, 15–25. doi: 10.1016/j.fishres.2006.05.020
Bahamon N., Sardà F., Suuronen P. (2007). Potential benefits from improved selectivity in the northwest Mediterranean multispecies trawl fishery. ICES J. Mar. Sci. 64, 757–760. doi: 10.1093/icesjms/fsm052
Baro J., Muñoz de los Reyes I. (2007). Comparación de los rendimientos pesqueros y la selectividad del arte de arrastre empleando mallas cuadradas y rómbicas en el copo. Informes técnicos. Instituto Español Oceanografía 188), 1–23.
Beverton R. J. H. (1963). “Escape of fish through different parts of a codend,” in The selectivity of fishing gear (Lisbon, Portugal: ICNAF Spec. Publ), 9–11.
Brooks M. E., Melli V., Savina E., Santos J., Millar R., O’Neill F. G., et al. (2022). Introducing selfisher: open source software for statistical analyses of fishing gear selectivity. Can. J. Fish. Aquat. Sci. 9, 1–9. doi: 10.1139/cjfas-2021-0099
Carbonara P., Intini S., Modugno E., Maradonna F., Spedicato M. T., Lembo G., et al. (2015). Reproductive biology characteristics of red mullet (Mullus barbatus l. 1758) in southern Adriatic Sea and management implications. Aquat. Living Resour. 28, 21–31. doi: 10.1051/alr/2015005
Cherif M., Zarrad R., Gharbi H., Missaoui H., Jarboui O. (2007). Some biological parameters of the red mullet, Mullus barbatus l. 1758, from the gulf of Tunis. Acta Adriat. 48 (2), 131–144.
Clark J. R. (1963). “Size selection of fish by otter trawls. results of recent experiments in the Northwest Atlantic,” in The selectivity of fishing gear (Lisbon, Portugal: ICNAF Spec. Publ), 25–96.
Colloca F., Cardinale M., Maynou F., Giannoulaki M., Scarcella G., Jenko K., et al. (2013). Rebuilding Mediterranean fisheries: a new paradigm for ecological sustainability. Fish Fisheries 14 (1), 89–109. doi: 10.1111/j.1467-2979.2011.00453.x
Efron B. (1987). Better bootstrap confidence intervals. J. Am. Stat. Assoc. 82, 171–185. doi: 10.1080/01621459.1987.10478410
FAO (2022). The state of Mediterranean and black Sea fisheries 2022 (Rome: General Fisheries Commission for the Mediterranean). doi: 10.4060/cc3370en
García-Rodríguez M., Pérez Gil J. L., Barcala E. (2009). Some biological aspects of parapenaeus longirostris (Lucas 1846) (Decapoda, dendrobranchiata) in the gulf of alicante (S.E. Spain). Crustaceana 82 (3), 293–310. doi: 10.1163/156854009X409108
GFCM (2021). Working group on stock assessment of demersal species (WGSAD). scientific advisory committee (SAC) (Rome, Italy: central and eastern Mediterranean and Adriatic Sea), 115.
GFCM (2022). Working group on stock assessment of demersal species (WGSAD). scientific advisory committee (SAC) (central and eastern Mediterranean and Adriatic Sea), 98.
Guijarro B., Massutí E. (2006). Selectivity of diamond- and square-mesh codends in the deepwater crustacean trawl fishery off the Balearic islands (western Mediterranean). ICES J. Mar. Sci. 63, 52–67. doi: 10.1016/j.icesjms.2005.08.011
Guijarro B., Massutí E., Moranta J., Cartes J. E. (2009). Short spatio-temporal variations in the population dynamics and biology of the deep-water rose shrimp Parapenaeus longirostris (Decapoda: Crustacea) in the western Mediterranean. Scientia Marina 73 (1), 183–197. doi: 10.3989/scimar.2009.73n1183
Holst R., Revill A. (2009). A simple statistical method for catch comparison studies. Fish. Res. 95, 254–259. doi: 10.1016/j.fishres.2008.09.027
Krag L. A., Herrmann B., Iversen S., Engås A., Nordrum S., Krafft B. A. (2014). Size selection of Antarctic krill (Euphausia superba) in trawls. PloS One 9 (8), e102168. doi: 10.1371/journal.pone.0102168
Lleonart J. (1993). Methods to analyse the dynamics of exploited marine populations: use and development of models. Sci. Mar. 57 (2-3), 261–267.
Lleonart J., Maynou F. (2003). Fish stock assessments in the Mediterranean: state of the art. Scientia Marina 67 (S1), 37–49. doi: 10.3989/scimar.2003.67s137
Lucchetti A. (2008). Comparison of diamond- and square-mesh codends in the hake (Merluccius merluccius l. 1758) trawl fishery of the Adriatic Sea (central Mediterranean). Scientia Marina 72 (3), 451–460. doi: 10.3989/scimar.2008.72n3451
Lucchetti A., Virgili M., Vasapollo C., Petetta A., Bargione G., Li Veli D., et al. (2021). An overview of bottom trawl selectivity in the Mediterranean Sea. Mediterr. Mar. Sci. 22, 566–585. doi: 10.12681/mms.26969
Martín P., Maynou F., Garriga M., Ramírez J., Recasens L. (2019). Fishing effort alternatives for the management of demersal fisheries in the western Mediterranean. Sci. Mar. 83 (4), 293–304. doi: 10.3989/scimar.04937.29B
Maynou F., García de Vinuesa A. G., Martínez-Baños P., Sánchez P., Demestres M. (2021). Relative catch performance of two gear modifications used to reduce bycatch of undersized fish and shrimp in Mediterranean bottom trawl fisheries. Mar. Coast. Fisheries: Dynamics Management Ecosystem Sci. 13, 529–544. doi: 10.1002/mcf2.10178
Millar R. (1993). Incorporation of between-haul variation using bootstrapping and nonparametric estimation of selection curves. Fish. Bull. 91, 564–572.
Moranta J., Massutıí E., Morales-Nin B. (2000). Fish catch composition of the deep-sea decapod crustacean fisheries in the Balearic islands (western Mediterranean). Fisheries Res. 45 (3), 253–264. doi: 10.1016/S0165-7836(99)00119-8
Oliver P. (1993). Analysis of fluctuations observed in the trawl fleet landings of the Balearic islands. Scientia Marina 57, 219–227.
Ordines F., Lloret J., Tugores P., Manfredi C., Guijarro B., Jadaud A., et al. (2019). A new approach to recruitment overfishing diagnosis based on fish condition from survey data. Sci. Mar. 83 (S1), 223–233. doi: 10.3989/scimar.04950.03A
Ordines F., Massutí E., Guijarro B., Mas R. (2006). Diamond vs. square mesh codend in a multi-species trawl fishery of the western Mediterranean: effects on catch composition, yield, size selectivity and discards. Aquat. Living Resour 19, 329–338. doi: 10.1051/alr:2007003
Petetta A., Herrmann B., Virgili M., Li Veli D., Brinkhof J., Lucchetti A. (2022). Effect of extension piece design on catch patterns in a Mediterranean bottom trawl fishery. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.876569
R Core Team (2022). R: A language and environment for statistical computing. R foundation for statistical computing (Vienna, Austria). Available at: https://www.R-project.org/.
Recasens L., Chiericoni V., Belcari P. (2008). Spawning pattern and batch fecundity of the European hake (Merluccius merluccius (Linnaeus 1758)) in the western Mediterranean. Scientia Marina 72 (4), 721–732. doi: 10.3989/scimar.2008.72n4721
Reeves S. A., Armstrong D. W., Fryer R. J., Coull K. A. (1992). The effects of mesh size, cod-end extension length and cod-end diameter on the selectivity of Scottish trawls and seines. ICES J. Mar. Sci. 49, 279–288. doi: 10.1093/icesjms/49.3.279
Robert M., Morandeau F., Scavinner M., Fiche M., Larnaud P. (2020). Toward elimination of unwanted catches using a 100 mm T90 extension and codend in demersal mixed fisheries. PloS One 15, e0235368. doi: 10.1371/journal.pone.0235368
Robertson J. H. B., Stewart P. A. M. (1988). A comparison of size selection of haddock and whiting by square and diamond mesh codends. J. Cons. Int. Explor. Mer 44, 148–161. doi: 10.1093/icesjms/44.2.148
Sola I., Maynou F. (2018). Assessment of the relative catch performance of hake, red mullet and striped red mullet in a modified trawl extension with T90 netting. Sci. Mar. 82, 19–26. doi: 10.3989/scimar.04711.04A
Stergiou K. I., Politou C. Y., Christou E. D., Petrakis G. (1997). Selectivity experiments in the NE Mediterranean: the effect of trawl codend mesh size on species diversity and discards. ICES J. Mar. Sci. 54 (5), 774–786. doi: 10.1006/jmsc.1997.0231
Wienbeck H., Herrmann B., Moderhak W., Stepputtis D. (2011). Effect of netting direction and number of meshes around on size selection in the codend for Baltic cod (Gadus morhua). Fish. Res. 109, 80–88. doi: 10.1016/j.fishres.2011.01.019
Keywords: selectivity, European hake, red mullet, T90 extension, 52 mm square meshed codend, western Mediterranean, bottom trawling
Citation: Ferragut-Perello F, Vasapollo C, Valls M, Farriols MT, Massutí E, Guijarro B, Joher S, Bibiloni-Socias M and Ordines F (2023) Effects of the implementation of T90 extension and 52 mm square mesh codend on the bottom trawl hake fishery of the north western Mediterranean. Front. Mar. Sci. 10:1035448. doi: 10.3389/fmars.2023.1035448
Received: 02 September 2022; Accepted: 30 May 2023;
Published: 15 June 2023.
Edited by:
Konstantinos Tsagarakis, Hellenic Centre for Marine Research (HCMR), GreeceReviewed by:
Rosalia Ferreri, National Research Council (CNR), ItalyCopyright © 2023 Ferragut-Perello, Vasapollo, Valls, Farriols, Massutí, Guijarro, Joher, Bibiloni-Socias and Ordines. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francesca Ferragut-Perello, ZnJhbmNlc2NhLmZlcnJhZ3V0QGllby5jc2ljLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.