 Li Jiang1†
Li Jiang1† Xinhong Zhou
Xinhong Zhou Bo Liu
Bo Liu- 1College of Life Science and Engineering, Southwest University of Science and Technology, Mianyang, China
- 2Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture and Rural Affairs, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi, China
The study aimed to evaluate the effects of fermented tea residue (FT) on growth performance, intestinal morphology, liver antioxidant capacity and Aeromonas hydrophila infection in juvenile Largemouth bass. A total of 240 fish were randomly distributed in 12 tanks with 20 fish per tank (4 treatments with 3 replications) and fed with diets FT at the rate of 0 (control), 2, 4 and 6%. The weight gain rate (WGR), specific growth rate (SGR) and intestinal villi height (VH) of juvenile largemouth bass were significantly higher than those of the control group after feeding FT (P< 0.05); meanwhile, the liver superoxide dismutase (SOD), glutathione peroxidase (GSH-PX) and catalase (CAT) activities of juvenile largemouth bass were significantly higher and the malondialdehyde (MDA) levels were significantly lower than those of the control group after feeding FT (P< 0.05). Mortality occurred in all groups of largemouth bass after the injection of A.hydrophila, but feeding FT reduced the cumulative mortality compared with the control group (P< 0.05). In juvenile largemouth bass infected with A.hydrophila, the relative mRNA expression of the intestinal anti-inflammatory factors IL-10 and TGF-α was significantly higher and that of the pro-inflammatory factors IL-1, IL-15, IL-8, and TNF-α was significantly lower (P< 0.05). In summary, it can be seen that a 2% FT addition can improve the liver antioxidant capacity of juvenile largemouth bass, enhance the resistance to A.hydrophila and increase the growth of largemouth bass.
1 Introduction
Largemouth bass (Micropterus salmoides) is a carnivorous warm-water fish from California, U.S.A. It has become one of the main species of freshwater aquaculture in China because of its strong adaptability, fast growth rate and tender flesh (Bureau 2021). In 2020, the farmed production reached 619,519 tons, an increase of 29.66% compared with 2019; (Bureau, 2021). However, under intensive farming, disease outbreaks are more frequent in largemouth bass, which are mainly caused by parasites, bacteria and fungi, The producers need to use a lot of antibiotics and disinfectants to treat these diseases, which leads to economic losses (ValladãO et al., 2015). However, the misuse of antibiotics or chemicals can cause many negative effects on the environment, animals and humans (Cabello, 2010; Rico and Brink, 2014). Therefore, the use of antibiotics is slowly being restricted. It has become a trend in the global aquaculture industry to use positive immune stimulants to enhance the innate immune mechanism of fish to increase their disease resistance (Fuchs et al., 2015; ValladãO et al., 2015).
Studies have shown that preventive treatments promote innate immune responses in fish and reduce disease outbreaks in aquaculture (Ma et al., 2020; Tadese et al., 2020). The use of micro-ecological preparations, such as fermented feed, and plant-based additives can better enhance the immunity of aquaculture animals (Burr et al., 2010; Newaj‐Fyzul and Austin, 2015; Dawood et al., 2018; Niu et al., 2020). The addition of Aqualase® (a yeast-based commercial probiotic composed of Saccharomyces cerevisiae and Saccharomyces ellipsoidal) in the feed can moderate the intestinal microbiota of rainbow trout and improve immunity and growth (Adel et al., 2017). The inclusion of 2.5 - 2.61*107 CFU/kg of two probiotics (Lactococcus lactis and weissella confuse) in the diet can improve the growth performance of fingerling great sturgeon, improve the immune index, and increase the height of intestinal villi (Yeganeh Rastekenari et al., 2021). The fermented dragon fruit in the diet improves the growth performance and feed utilization of Platax pinnatus and increases antioxidant enzyme activity to some extent (Chu et al., 2021). Feeding fermented feed by Bacillus subtilis to Penaeus monodon is shown to improve the growth performance, feed digestibility, survival rate and immunity of spot prawns (De et al., 2018).
After soaking goldfish infected with A. hydrophila for a certain period of days with 1% compound herbal water, the damaged primary gill flaps, liver, heart and muscle tissue structure of goldfish are restored (Ramasamy et al., 2010). Immunostimulant (mixture of Chinese herbs and Bacillus), could significantly upregulate the expression of NADPH oxidase genes and antioxidant genes in tilapia spleen neutrophils, thus improving the immunity of tilapia (Abarike et al., 2019).
Tea residue is a by-product of tea refining, which is rich in tea polyphenols, theanine and other active ingredients, with natural antioxidants, immune promotion and other functions (Rietveld and Wiseman, 2003; Hamer, 2007). There are more types of fermented feed, but the result of synergistic fermentation of bacteria and enzymes is better than the result of bacteria and enzymes alone (Xie et al., 2015). Because the combination of bacteria and enzymes can make the degradation of macromolecules more complete, the fermentation efficiency is higher (Sun et al., 2021). The addition of green tea powder to the diet could increase the serum TP content and SOD activity of rainbow trout, upregulate the mRNA expression levels of anti-inflammatory factors and downregulate the mRNA expression levels of pro-inflammatory factors in the spleen and kidney (Nootash et al., 2013). Green tea could improve the growth performance and feed utilization of Paralichthys olivaceus, and effectively reduce serum glutathione transaminase (GPT) and low-density lipoprotein (LDL) in fish (Cho et al., 2007). And it improves the growth performance and health of Nile Tilapia against infection by A.hydrophila (Abdel et al., 2010). Tea and fermented feed have positive effects on growth performance, immune system, pathogen protection and immunity in different fish species. The application of fermented tea residue on largemouth bass is limited. In this experiment, we investigated the effects of FT with different addition ratios on the growth performance, intestinal tissue structure, antioxidant capacity and infection of A.hydrophila of largemouth bass.
2 Materials and methods
2.1 Ethical statement
This study was conducted in strict accordance with the Experimental Animal Management Regulations of Southwest University of Science and Technology. All of the procedures were performed following the Declaration of Helsinki and relevant policies in China.
2.2 Experimental diets and design
Four test diets were prepared. Basal diets and rations with 2%, 4% and 6% fermented tea residue (FT) were added to the basal diet. The composition of the base diet is shown in Table 1, and the nutritional composition of the diets is shown in Table 2. After the base diet was prepared, we added different proportions of FT to the base diet and then mixed and kneaded them into soft pellets (1.2-mm diameter) for feeding. FT is made by crushing tea residue and adding 4% corn flour, 0.8% glucose, 35% water, 0.1% probiotics and enzyme preparations to ferment thoroughly at 36°C for 3 days. FT is the tea residue produced after Fuxuan 9 processing, from Sichuan, China. Probiotics and enzyme preparations were purchased from a biological company; the product label number is Q/12JX 4450-2019. The fermentation bacteria include Lactobacillus Plantarum ≥ 1.0×109 CFU/g, Bacillus subtilis ≥ 1.5×109 CFU/g, Saccharomyces cerevisiae ≥ 1.0×109 CFU/g; the enzyme preparation contains cellulase ≥ 1000 U/g, xylanase ≥ 500 U/g, β-glucanase ≥ 3000 U/g, β-mannanase ≥ 50 U/g.

Table 1 Composition and nutrient levels of the basal diet (air-dry basis).
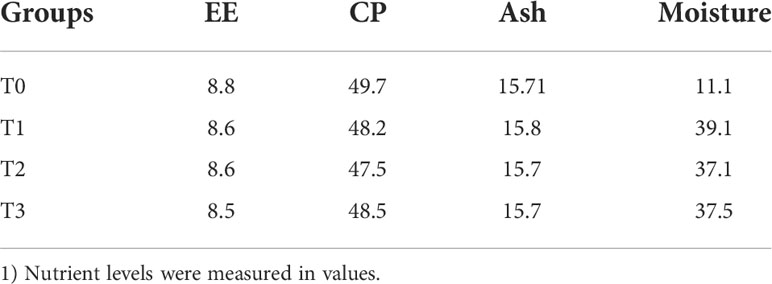
Table 2 Nutrient composition of the basal and experimental diets.
2.3 Experimental fish and breeding management
Healthy juvenile largemouth bass (average weight of about 5 g) were obtained from Meishan City, Sichuan Province, China. Upon arrival, all fish were tamed in a test environment for one week. In the formal trial, after all fish were starved for 24 hours, 240 largemouth bass were randomly divided into 4 treatments (groups T0 - T3). T0 was the control group receiving the basal diet, and T1-T3 were fed with FT added to the basal diet at 2%, 4% and 6%, respectively. Each treatment was stocked with three replicates stocked with 60 fish (20 fish/tank) in the tank (1m * 50cm * 1m). During the experiment, fish were hand-fed with experimental diets twice a day at a rate of 3% of body weight (8:30 am and 4:30 pm). We changed 1/5 - 3/5 of the water every 2 - 3 days and used the pump to remove the bottom feces. The water temperature was natural and the dissolved oxygen content was greater than or equal to 6.0 mg/L, pH 7.0 ± 0.2, ammonia nitrogen ≤ 0.02 mg/L. The rearing experiment lasted for 56 days.
2.4 A.hydrophila challenge test
At the end of the feeding trial, 30 fish of each group (10 fish/tank) were selected to be injected with A.hydrophila for infection, and previously we derived from pre-experiment that the LC50 of A.hydrophila on juvenile largemouth bass was 1.65×106 CFU/ml at an injection dose of 0.2 ml/tail. The trial fish were anesthetized with an appropriate amount of 50 ppm MS-222 for 3-5 min, and then 0.2 ml of A.hydrophila liquid with a concentration of 1.65×106 CFU/ml was slowly injected from the abdominal cavity using a 1 ml injector, and the breeding environment was kept unchanged.
2.5 Sample collection
For statistical analysis of growth performance, we fasted for 24 h at the end of the breeding test. We weighed the total weight of each test tank and counted the number of surviving fish for statistical analysis. Nine fish were randomly selected from each replicate, anesthetized with 100 ppm MS-222, and their body weight and length were measured. Subsequently, the midgut of three fish was randomly selected from each group of nine fish and fixed with 4% paraformaldehyde for intestinal histological observation. Take the remaining fish viscera to measure the body index. collected livers and intestines were snap frozen in liquid nitrogen (-196°C) and then transferred to -80°C refrigerator for storage. On the third day after the A.hydrophila infection treatment, all fish were fasted for 24 h, anesthetized with 100 ppm MS-222, and the whole intestines were taken into sterile tubes, snap-frozen in liquid nitrogen (-196°C), and then transferred to -80°C refrigerator storage for intestinal inflammatory factor expression assay. Growth indicators and cumulative mortality were calculated as follows:
Where Nt and No represent the total number of fish samples at the beginning and end of the experiment, respectively; Wt and W0 are the initial and final weight data at the beginning and end of the experiment (g); W is the body weight per fish, Wh is the liver weight of per fish. t represents the number of experimental days (day); L is the length of the fish (cm). Nd is the cumulative number of fish dead; Na is the initial number of fish after injection of A.hydrophila.
2.6 Sample measurement
2.6.1 Histomorphology of the intestine
The midgut of juvenile largemouth bass was rinsed with saline and fixed with 4% paraformaldehyde. After processing, sections (5 μm) were sectioned using a paraffin slicer, followed by hematoxylin-eosin (HE) staining and image acquisition by light microscopy, with the observed sections first observed under low magnification and the appropriate areas selected for image acquisition under high magnification. The data for villi height, width, and thickness of the muscular layer were measured by Image-Pro Plus software.
2.6.2 Analysis of antioxidant enzyme activities and immune enzyme activities in liver
The liver tissues stored at -80 °C were thawed on ice, and the tissues were homogenized with saline 1:9 according to the kit instructions, centrifuged at 4 °C and 2,500 r/min for 10 min, and then the supernatant was taken as the tissue homogenate. Total protein (TP), superoxide dismutase (SOD), total antioxidant capacity(T-AOC), glutathione peroxidase (GSH-Px), malondialdehyde (MDA) and catalase (CAT) were measured in the liver using a spectrophotometer or enzyme marker according to the steps of the kit (Nanjing Jiancheng Institute of Biological Engineering) instructions.
2.6.3 Measurement of intestinal inflammatory factor expression after A.hydrophila infection
Intestinal tissues stored at -80°C were placed in RNAase-free centrifuge tubes and ground using a microtissue homogenizer. Total RNA was extracted from the intestinal tissues by the Trizol method (TaKaRa, Japan). The concentration of RNA was measured using a micro ultraviolet spectrophotometer. The first-strand cDNA was synthesized using the kit (product number RR047A, TaKaRa) according to the instructions. Protocol for reverse transcription: 37°C for 15 minutes; 85°C for 5 seconds. The primers were used to refer to the study of Xv (Xv et al., 2021). Table 3 shows the PCR primers used in this study for the coding sequences of IL-1β, IL-8, IL-10, IL-15, TNF-α and TGF-β genes in the largemouth bass genome. And β-actin was used as an internal reference gene, the specific primers for β-actin and target genes were synthesized by Tsingke Biotechnology Co., Ltd. Quantitative real-time PCR (qPCR) was performed using NovoStart SYBR qPCR SuperMix Plus (Novoprotein) on Bio-Rad CFX96 (Bio-Rad) in a total volume of 20μL.

Table 3 Primer sequences for real-time PCR.
2.7 Calculations and statistical methods
All data are expressed as mean± SD. Significance levels were determined by one-way analysis of variance (ANOVA) with IBM SPSS Statistics 23. Multiple comparisons were performed using the Tukey multiple range test. The statistical significance level was set at p<0.05. Graphs were drawn using GraphPad Prism6 (GraphPad Software, Inc., USA).
3 Results
3.1 Growth performance and morphometric parameters
As the growth performance and morphological indices are shown in Table 4, FT can significantly improve the weight gain rate and specific growth rate of juvenile largemouth bass compared with T0 (P< 0.05). Compared with T3, T1 and T2 also significantly increased the weight gain and specific growth rate of juvenile largemouth bass (P< 0.05). The condition factor of T1 was significantly higher than the rest of the other groups, and FT could significantly reduce the HSI index of juvenile largemouth bass (P< 0.05). However, there was no significant difference in the mortality rate among all groups (P > 0.05).
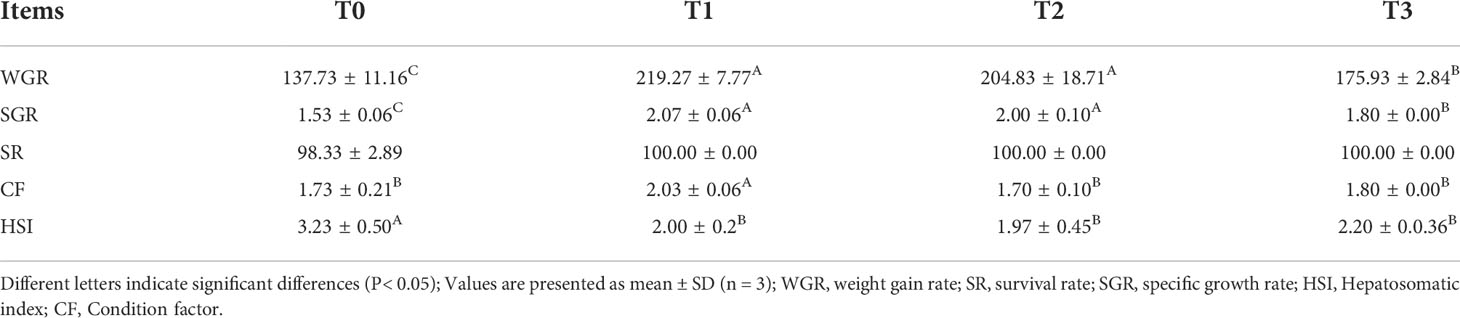
Table 4 Growth performance and morphological indicators.
3.2 Cumulative mortality of juvenile largemouth bass after injection of A.hydrophila
In this trial, largemouth bass were infected with A.hydrophila and mortality was counted for 3 consecutive days, as shown in Figure 1. After the injection of A.hydrophila, all groups showed mortality on the first day, but the cumulative mortality rate of the control group was higher than that of the other groups (P< 0.05), and the cumulative mortality rate rapidly increased with time. The cumulative mortality rate for T0 was 43.33%, while T1 was 23.33%, T2 was 20.00%, and T3 was 26.67%, respectively.
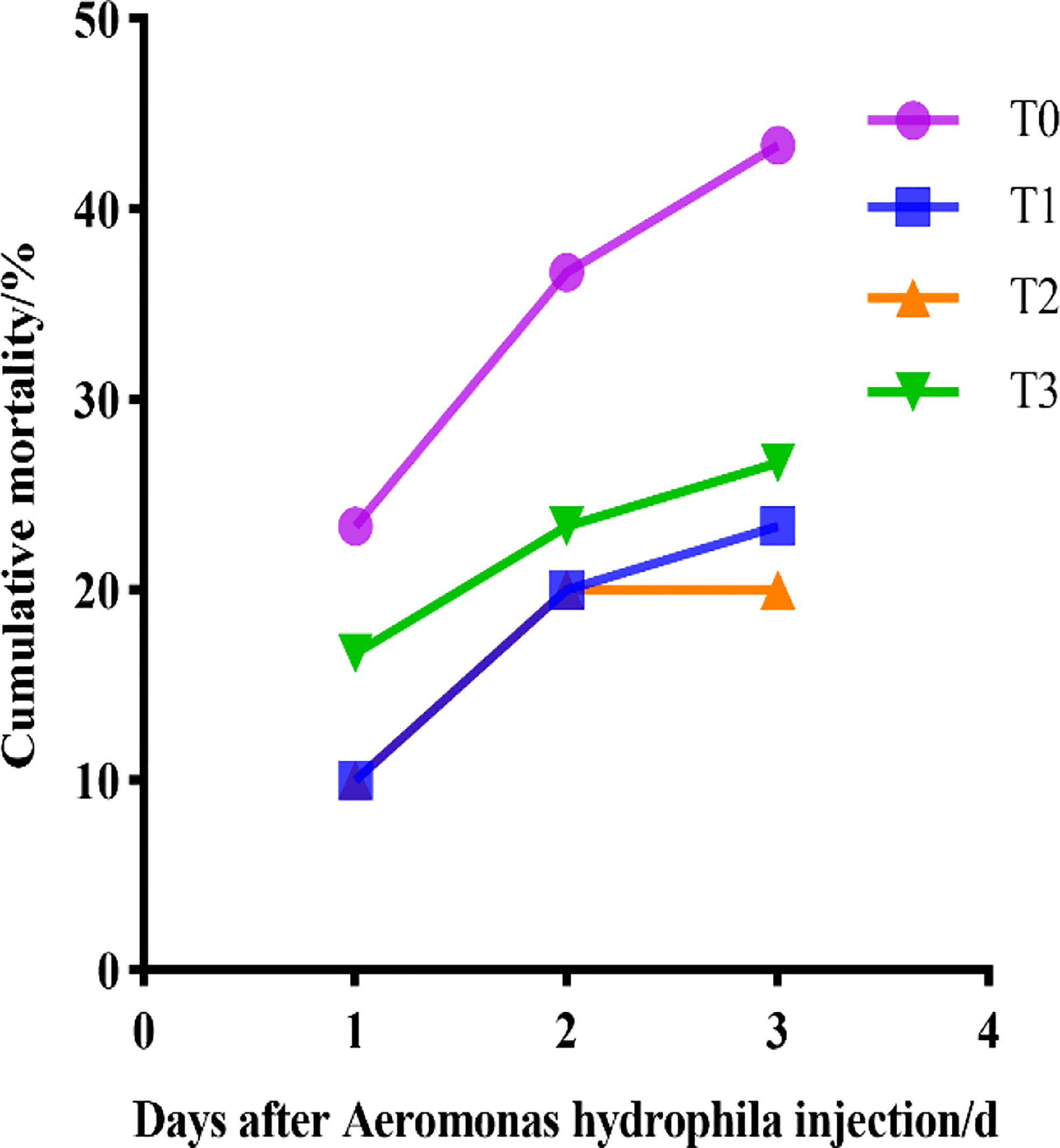
Figure 1 Cumulative mortality of juvenile largemouth bass after injected with A.hydrophila.
3.3 Morphological observation of the intestinal tract of juvenile largemouth bass
The tissue structure of the midgut of juvenile largemouth bass is shown in Figure 2, and the characteristics of the midgut villi are shown in Table 5. The intestinal villi of juvenile largemouth bass in the control group were shorter and sparser, with fewer villi and goblet cells (P< 0.05). The height of intestinal villi was significantly higher in the test group than in the control group (P< 0.05).
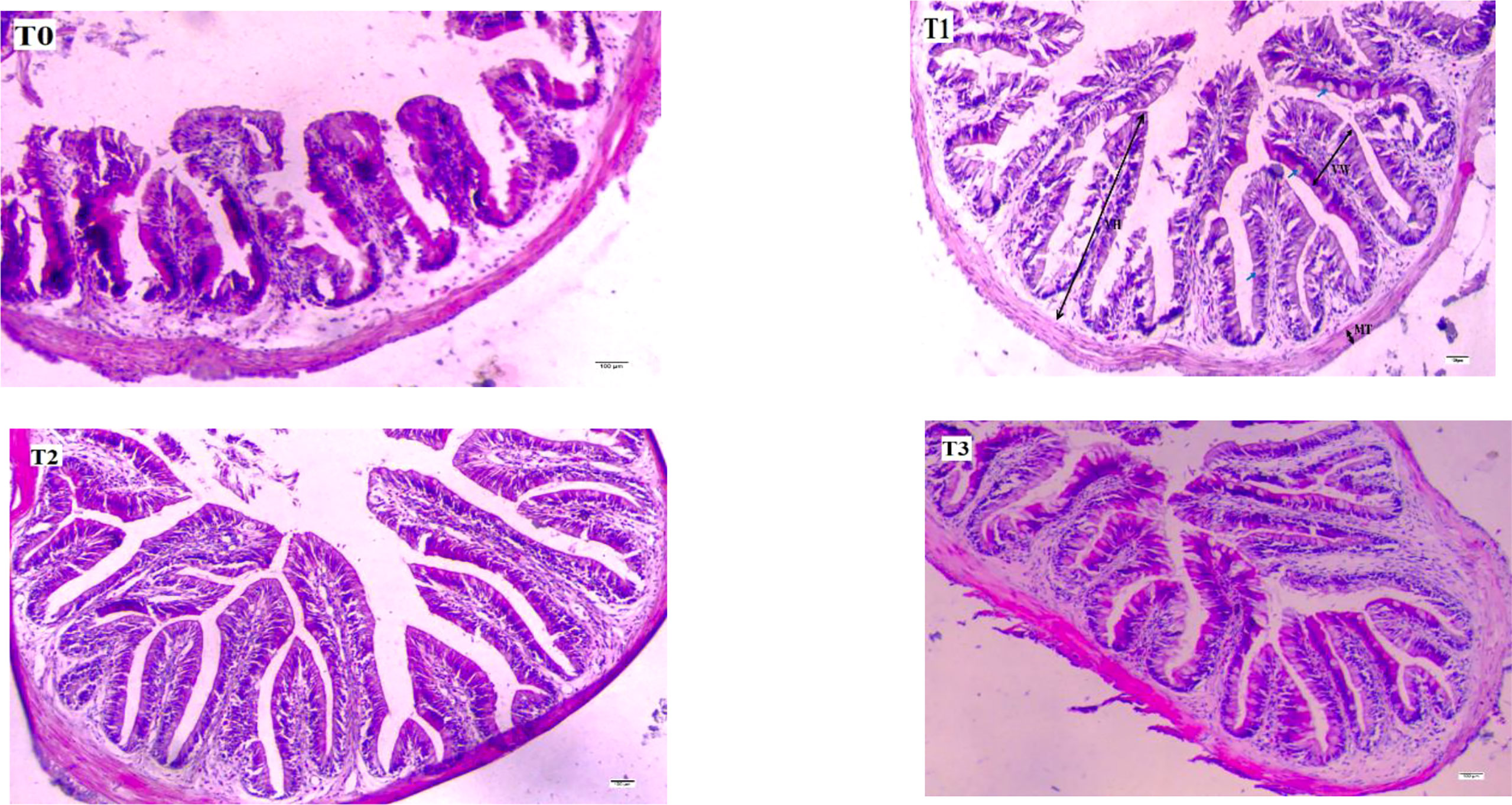
Figure 2 Effect of different scales of FT on the morphology of the mid-gut of the juvenile largemouth bass (X100, H&E staining, scale bar = 100(μm). Villi height (VH), villi width (VW), muscular layer thickness (MT), and the blue arrows indicated are goblet cells.

Table 5 Characteristics of midgut villi of largemouth bass.
3.4 Largemouth bass liver antioxidant index
The antioxidant indices in the livers of juvenile largemouth bass are shown in Table 6. Compared with the control group, the activities of SOD, GSH-PX, CAT and the level of T-AOC in the livers showed a significant increase as well as a significant decrease in MDA content in juvenile largemouth bass after 8 weeks of FT feeding (P< 0.05).
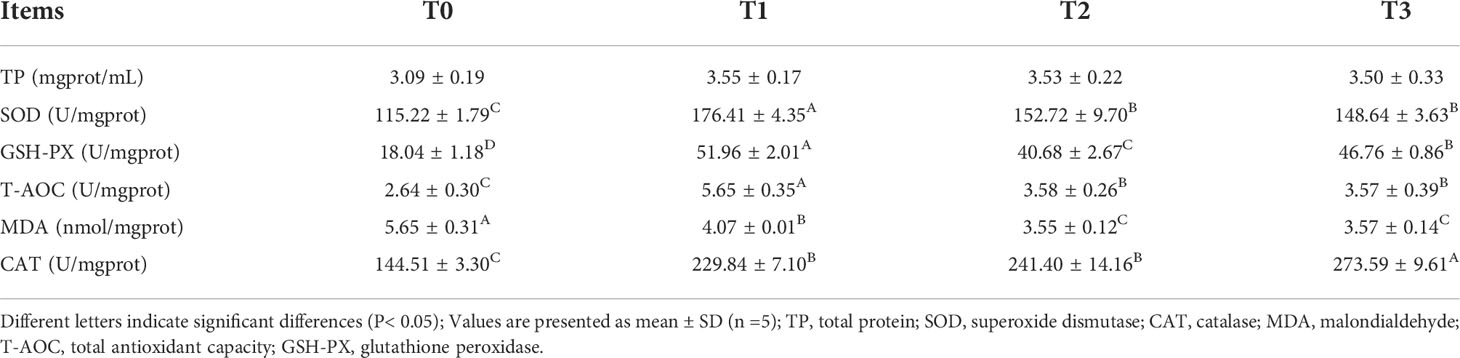
Table 6 Analysis of antioxidant enzyme activities and immune enzyme activities in liver.
3.5 Expression of intestinal inflammatory factors in juvenile largemouth bass after A.hydrophila infection
Infection with A.hydrophila caused an inflammatory response in the intestine characterized by increased expression of pro-inflammatory factors and decreased expression of anti-inflammatory factors. In this experiment, we measured the expression of intestinal inflammatory factors in largemouth bass after infected by A.hydrophila. And the results are shown in Figure 3. In juvenile largemouth bass fed FT for 8 weeks, the relative mRNA expressions of intestinal pro-inflammatory factors IL-1β, IL-15, IL-8 and TNF-α decreased significantly, while the relative mRNA expressions of anti-inflammatory factors IL-10 and TGF-β increased significantly (P< 0.05). The relative mRNA expression of anti-inflammatory factors IL-10 and TGF-β1 was significantly higher in T2 compared with those of T1 and T3. In contrast, the relative mRNA expression of pro-inflammatory factors IL-1β and IL-15 was significantly lower in T2 compared with T1 and T3 (P<0.05).
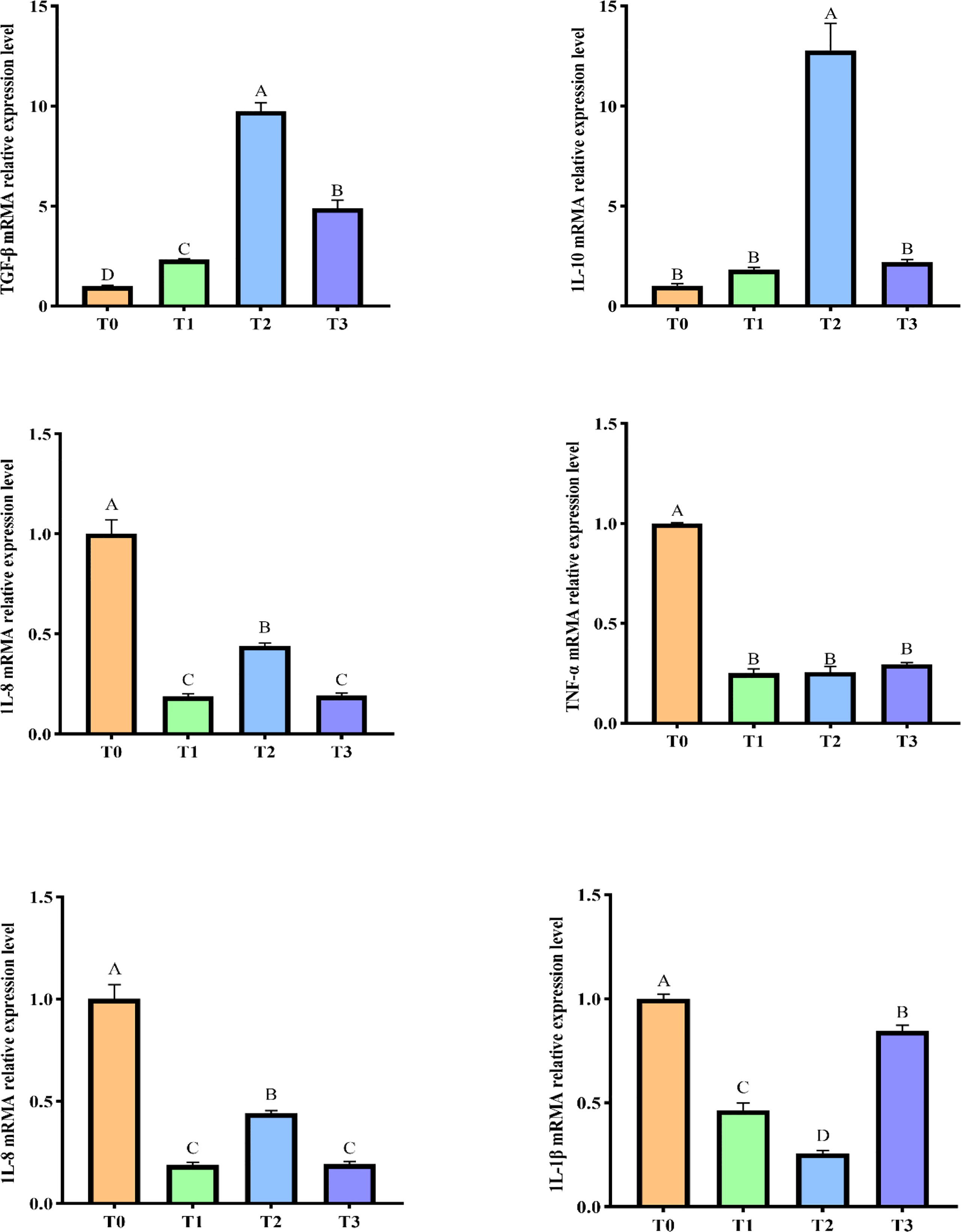
Figure 3 Effect of feeding FT on the expression of inflammatory factors in the intestine of largemouth bass after infection with A.hydrophila. Different letters indicate significant differences (P < 0.05); Values are presented as mean ± SD (n =5). IL-1β: interleukin-1β; IL-8: interleukin-8; IL-10: interleukin-10; IL-15: interleukin-15; TNF-α: tumor necrosis factor-α; TG F-β1: transforming growth factor-β.
4 Discussion
Tea residue is rich in tea polyphenols, theanine, tea saponin and other active substances, which can improve the production performance of livestock and meat quality (Hamer, 2007). It has been shown that the addition of appropriate amounts of tea to diets can improve the activity of digestive enzymes, reduce anti-nutritional factors, and improve the use of nutrients, thus promoting the growth of fish (Zhang et al., 2015; Zheng et al., 2017). Similar to this study, plant extracts such as Common Sage (Salvia officinalis), Coneflower (Echinacea angustifolia), Cornelian cherry (Cornus mas L.), Rose hip and Safflower can stimulate the innate immune response and feed intake, thus improving fish growth performance (Dadras et al., 2016; Dadras et al., 2019; Ahmadifar et al., 2022). Fermented feeds can reduce the anti-nutritional factors in feeds and increase the digestion and absorption capacity of feeds, thus promoting the growth of the organism (Ilha et al., 2017; Wang et al., 2017). The addition of fermented tea residue to the diet can improve the fattening performance and digestive performance of fattening pigs (Ding et al., 2020). In this study, FT was able to improve the weight gain rate and specific growth rate of juvenile largemouth bass, but the weight gain rate and specific growth rate were lower than the actual production values, which was probably because the experiment was conducted during the seasonal change in autumn and winter, the temperature may have affected the growth performance of largemouth bass.
The intestine is an important site for digestion and absorption of nutrients as well as protection against pathogens in fish. Intestinal histological assessment is an effective method to assess the effect of dietary components on the intestinal health of fish (Chauhan and Singh, 2018; Ding et al., 2020). Some studies found that tea polyphenols and fermented feed could increase the height of intestinal villi and thickness of the muscle layer in fish, and improve the intestinal histology of fish (Mamauag et al, 2019; Ma et al., 2021; Zhuo et al., 2021). It was found that the height and width of intestinal villi of juvenile largemouth bass were higher than those of the control group after feeding FT, which indicated that the integrity and stability of the intestinal tract were enhanced, thus strengthening the digestion and absorption of nutrients by juvenile largemouth bass and promoting their growth, this might be one of the reasons for their better growth performance compared to the control group.
Oxidative stress is a state in which there is an imbalance between oxidation and antioxidant action in the body, a negative effect produced by free radicals in the body, which predisposes the body to age and disease (Bai et al., 2017). The most common enzymatic antioxidants present in animals are CAT, SOD and GSH-Px, which mainly serve to scavenge peroxides in the body to protect it from damage caused by oxidative stress (Yuan et al., 2019; Chen et al., 2020). The antioxidant enzyme activity in animals determines the antioxidant capacity of their bodies, which can be used to assess the health of fish (Tovar-Ramírez et al., 2010). The level of T-AOC is one of the indicators of antioxidant capacity in animals, which is an important indicator of the antioxidant capacity of fish directly (Cui et al., 2014; Yu et al, 2021). This study found that FT could enhance the activity of SOD, GSH-Px and CAT as well as the level of T-AOC in the liver of juvenile largemouth bass, which indicated that FT could enhance the antioxidant capacity of juvenile largemouth bass and slow down the oxidative damage to the organism. MDA is a peroxidation metabolite generated by lipids in the body under the influence of free radicals. The level of MDA content can directly reflect the damage to the body by free radicals, and higher levels of MDA reflect higher peroxidation reactions in the body (Janero, 1990; Yu et al., 2019). In this study, we found that feeding FT could reduce the liver MDA content of juvenile largemouth bass. The results were generally consistent with other studies that dietary medical plants or fermented tea residue could enhance the activity of SOD, GSH-Px and CAT as well as the level of T-AOC and reduce the content of MDA in rainbow trout (Ghafarifarsani et al., 2022), Holstein heifers (Xie et al., 2020), common carp (Ahmadifar et al., 2022), Siberian sturgeon (Hasanpour et al., 2019), Tilapia (Qian et al., 2021), Sea bream (Pérez-Jiménez et al., 2012) and juvenile Wuchang bream (Guo et al., 2020).
Bacterial infection is often used as a final indicator of fish health status after nutrient analysis (Wang et al., 2015; Li et al., 2020). Bacterial enteritis is the most common intestinal disease of freshwater fish. Among many pathogenic bacteria, A.hydrophila is usually considered one of the main pathogens causing intestinal inflammation in fish (Cascón et al., 2000; Macpherson et al., 2012). In our study, the test fish fed FT showed a higher survival rate after infected with A.hydrophila, which may be due to the probiotic bacteria used in tea residue with fermentation to promote the immune response of largemouth bass. This is in agreement with previous studies that found green tea or probiotics can enhance the resistance of Nile tilapia (Abdel et al., 2010; Cavalcante et al., 2020), Cyprinus carpio (Chandravanshi et al., 2020), Lates calcarifer (Lin et al., 2017) and Labeo rohita (Rai et al., 2015) to A.hydrophila. It showed that feeding FT improved the survival of juvenile largemouth bass and protected the intestine from damage by A.hydrophila.
Intestinal immune regulatory molecules (cytokines) expression is positively correlated with the immune status of fish (Gil, 2002; Reda et al., 2018). Inflammation occurs as an important component of the innate immune responses. Therefore, inflammatory cytokines are often used as biomarkers of immune regulation (Safari et al., 2016). Fish cytokines can be classified into anti-inflammatory factors (such as IL-10 and TGF-β1) and pro-inflammatory factors (such as TNF-α, IL-1β, IL-15 and IL-8), which have important functions in the immune response. Enterocolitis decreases the expression of anti-inflammatory factors and increases the expression of pro-inflammatory factors, so they can indicate inflammatory damage at the molecular level (Song et al., 2014; Fcab et al., 2019). Dietary medical plants and probiotics can regulate fish intestinal innate immunity by promoting anti-inflammatory factors and reducing the expression of pro-inflammatory factors, thus strengthening the resistance of fish to disease-causing agents and thus slowing down inflammation (Panigrahi et al., 2007; Nootash et al., 2013; Feng et al., 2019; Vazirzadeh et al., 2019; Ahmadifar et al., 2022). In this study, it was found that feeding FT can reduce the mRNA relative expression levels of pro-inflammatory factors IL-1β, IL-15, TNF-α and IL-8, and increase the mRNA relative expression levels of anti-inflammatory factors IL-10 and TGF-β1 in juvenile largemouth bass. Therefore, the appropriate amount of FT can regulate the inflammatory state in the intestine of juvenile Largemouth bass after infection with A.hydrophila, thereby reducing the intestinal damage caused by A.hydrophila.
5 Conclusion
In this research, we found that the addition of a certain amount of FT could improve the growth performance and antioxidant capacity of juvenile largemouth bass, improve intestinal health, and increase resistance to A.hydrophila. A comprehensive analysis of this experiment showed that 2% FT addition was more effective.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Ethics statement
This trial was approved by the Southwest University of Science and Technology in China, Institutional Animal Care and Use Committee. All of the procedures were performed by the Declaration of Helsinki and relevant policies in China.
Author contributions
LJ: Conceptualization, Methodology, Formal analysis, Investigation, Writing – original draft. XZ: Conceptualization, Methodology, Formal analysis, Investigation, Writing – original draft. JY: Conceptualization, Methodology, Investigation, Writing – review and editing. SB: Formal analysis, Data curation, Investigation. JL: Data curation, Investigation. QW: Data curation, Investigation. MW: Investigation. YW: Investigation, Supervision, Funding acquisition. BL: Investigation, Writing – review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Project of National Key R&D Program of China (2019YFD09002), Central Publicinterest Scientific Institution Basal Research Fund, CAFS (2020TD59).
Acknowledgments
The authors would like to express their sincere thanks to the personnel of these teams for their kind assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abarike E. D., Jia C., Jian J., Tang J., Lu Y., Yu H. (2019). Characterization and expression of NADPH oxidase genes in stimulated splenic neutrophils of tilapia, oreochromis niloticus by a mix of traditional Chinese medicine and bacillus species (TCMBS). Aquacult. Res. 50 (1), 93–105. doi: 10.1111/are.13871
Abdel M., Ahmad M., Seden M., Sakr S. (2010). Use of green tea, camellia sinensis l., in practical diet for growth and protection of Nile tilapia, oreochromis niloticus (L.), against aeromonas hydrophila infection. J. World Aquacult. Soc. 41 (s2), 203–213. doi: 10.1111/j.1749-7345.2010.00360.x
Adel M., Lazado C. C., Safaric R., Yeganeh S., Zorriehzahra M. J. (2017). Aqualase, a yeast-based in-feed probiotic, modulates intestinal microbiota, immunity and growth of rainbow trout oncorhynchus mykiss. Aquacult. Res. 48 (4), 1815–1826. doi: 10.1111/are.13019
Ahmadifar E., Mohammadzadeh S., Kalhor N., Yousefi M., Moghadam M. S., Naraballobh W., et al. (2022). Cornelian cherry (Cornus mas l.) fruit extract improves growth performance, disease resistance, and serum immune-and antioxidant-related gene expression of common carp (Cyprinus carpio). Aquaculture 558 (15), 738372. doi: 10.1016/j.aquaculture.2022.738372
Bai K., Qiang H., Zhang J., He J., Tian W. (2017). Supplemental effects of probiotic bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. 96 (1), pew246. doi: 10.3382/ps/pew246
Burr G., Gatlin D., Ricke S. (2010). Microbial ecology of the gastrointestinal tract of fish and the potential application of prebiotics and probiotics in finfish aquaculture. J. World Aquacult. Soc. 36 (4), 425–36. doi: 10.1111/j.1749-7345.2005.tb00390.x
Cabello F. C. (2010). Heavy use of prophylactic antibiotics in aquaculture: a growing problem for human and animal health and for the environment. Environ. Microbiol. 8 (7), 1137–1144. doi: 10.1111/j.1462-2920.2006.01054.x
Cascón A., Yugueros J., Temprano A., Sánchez M., Naharro G. A. (2000). A major secreted elastase is essential for pathogenicity of aeromonas hydrophila. Infect. Immun. 68 (6), 3233–3241. doi: 10.1128/IAI.68.6.3233-3241.2000
Cavalcante R. B., Telli G. S., Tachibana L., Dias D., Ranzani-Paiva M. J. (2020). Probiotics, prebiotics and synbiotics for Nile tilapia: Growth performance and protection against aeromonas hydrophila infection. Aquacult. Rep. 17, 100343. doi: 10.1016/j.aqrep.2020.100343
Chandravanshi A., Naik M. G., Chandravanshi P., Rathore S., Keer N. R. (2020). Camellia sinensis (green tea) as feed additive enhanced immune response and disease resistance of cyprinus carpio (common carp) against aeromonas hydrophila infection. J. Exp. Zool. 23 (2), 1383–1390.
Chauhan A., Singh R. (2018). Probiotics in aquaculture: a promising emerging alternative approach. Symbiosis 77 (23), 99–113. doi: 10.1007/s13199-018-0580-1
Chen C., Xu C., Yang X., Qian D., Gu Z., Jia Y., et al. (2020). Growth, antioxidant capacity, intestine histology and lipid metabolism of juvenile red claw crayfish, cherax quadricarinatus , fed different lipid sources. Aquacult. Nutr. 27 (1), 261–273. doi: 10.1111/anu.13183
Cho S. H., Lee S. M., Park B. H., Ji S. C., Lee J., Oh B. (2007). Effect of dietary inclusion of various sources of green tea on growth, body composition and blood chemistry of the juvenile olive flounder, paralichthys olivaceus. Fish Physiol. Biochem. 33 (1), 49–57. doi: 10.1007/s10695-006-9116-3
Chu J. H., Liu K. J., Wu T. M. (2021). Effects of pitaya (Hylocereus polyrhizus) fermentation waste dietary supplement on growth performance, and anti-oxidation of pinnate batfish, platax pinnatus. Aquacult. Res. 52 (12):6689–6698. doi: 10.1111/are.15538
Cui Y. T., Liu B., Xie J., Xu P., Habte-Tsion H. M., Zhang Y. Y, et al. (2014). The effect of emodin on cytotoxicity, apoptosis and antioxidant capacity in the hepatic cells of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 38(1), 74–79. doi: 10.1016/j.fsi.2014.02.018
Dadras H., Hayatbakhsh M. R., Golpour A. (2019). Dietary administration of common sage (Salvia officinalis) and coneflower (Echinacea angustifolia) extracts affects growth, blood parameters and immune responses of beluga, huso huso. Turkish J. Fish. Aquat. Sci. 20 (5), 367–374. doi: 10.4194/1303-2712-v20_5_05
Dadras H., Hayatbakhsh M. R., Shelton W. L., Golpour A. (2016). Effects of dietary administration of rose hip and safflower on growth performance, haematological, biochemical parameters and innate immune response of beluga, huso huso (Linnaeus 1758). Fish Shellfish Immunol. 59, 109–114. doi: 10.1016/j.fsi.2016.10.033
Dawood M., Koshio S., Esteban M. (2018). Beneficial roles of feed additives as immunostimulants in aquaculture: a review. Rev. Aquacult. 10 (4), 950–974. doi: 10.1111/raq.12209
De D., Ananda Raja R., Ghoshal T. K., Mukherjee S., Vijayan K. K. (2018). Evaluation of growth, feed utilization efficiency and immune parameters in tiger shrimp ( penaeus monodon ) fed diets supplemented with or diet fermented with gut bacterium bacillus sp. DDKRC1. isolated from gut of Asian seabass ( lates calcarifer ). Aquacult. Res 49(6), 2147–2155. doi: 10.1111/are.13669
Ding X., Li H., Wen Z., Hou Y., Wang G., Fan J., et al. (2020). Effects of fermented tea residue on fattening performance, meat quality, digestive performance, serum antioxidant capacity, and intestinal morphology in fatteners. Anim. Open Access J. MDPI 10 (2), 185. doi: 10.3390/ani10020185
Fcab C., Mc A., Ajh C., Mla D., Cgf A. (2019). Anti-inflammatory effects of aloe vera on soy meal-induced intestinal inflammation in zebrafish. Fish Shellfish Immunol. 95, 564–573. doi: 10.1016/j.fsi.2019.10.075
Feng J., Chang X., Zhang Y., Yan X., Nie G. (2019). Effects of lactococcus lactis from cyprinus carpio l. as probiotics on growth performance, innate immune response and disease resistance against aeromonas hydrophila. Fish Shellfish Immunol. 93, 73–81. doi: 10.1016/j.fsi.2019.07.028
Fuchs V. I., Buck B. H., Steinhagen D., Schmidt J., Zentek J., Slater M. J. (2015). The effect of supplementation with polysaccharides, nucleotides, acidifiers and bacillus strains in fish meal and soy bean based diets on growth performance in juvenile turbot (Scophthalmus maximus). Aquaculture 437, 243–251. doi: 10.1016/j.aquaculture.2014.12.007
Ghafarifarsani H., Yousefi M., Hoseinifar S. H., Paolucci M., Lumsangkul C., Jaturasitha S., et al. (2022). Beneficial effects of Persian shallot (Allium hirtifolium) extract on growth performance, biochemical, immunological and antioxidant responses of rainbow trout oncorhynchus mykiss fingerlings. Aquaculture 555, 738162. doi: 10.1016/j.aquaculture.2022.738162
Gil A. (2002). Modulation of the immune response mediated by dietary nucleotides. Eur. J. Clin. Nutr. 56 Suppl 3, S1–S4. doi: 10.1038/sj.ejcn.1601475
Guo H., Lin W., Wang L., Zhang D., Wu X., Li L., et al. (2020). The supplementation of dietary selenium yeast and green tea-derived polyphenols improves antioxidant capacity and immune response in juvenile wuchang bream under ammonia stress. Aquacult. Res. 51 (9), 3790–3803. doi: 10.1111/are.14724
Hamer M. (2007). The beneficial effects of tea on immune function and inflammation: a review of evidence from in vitro, animal, and human research. Nutr. Res. 27 (7), 373–379. doi: 10.1016/j.nutres.2007.05.008
Hasanpour S., Sheikhzadeh N., Jamali H., Farsani M. N., Mardani K. (2019). Growth performance, antioxidant and immune status of Siberian sturgeon (Acipenser baeri) fed diets containing green tea extract and oxidized fish oil. J. Appl. Ichthyol. 35 (12), 1–10. doi: 10.1111/jai.13960
Ilha I., Hapsari F., Fotedar R. (2017). Growth, enzymatic glutathione peroxidase activity and biochemical status of juvenile barramundi (Lates calcarifer) fed dietary fermented lupin meal supplemented with organic selenium. Aquacult. Res. 43 (3), 775–790. doi: 10.1111/are.13444
Janero D. R. (1990). Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radical Biol. Med. 9 (6), 515–540. doi: 10.1016/0891-5849(90)90131-2
Lin H. L., Shiu Y. L., Chiu C. S., Huang S. L., Liu C. H. (2017). Screening probiotic candidates for a mixture of probiotics to enhance the growth performance, immunity, and disease resistance of Asian seabass, lates calcarifer (Bloch), against aeromonas hydrophila. Fish Shellfish Immunol. 60, 474–482. doi: 10.1016/j.fsi.2016.11.026
Li J., Wang C., Wang L., Xu Q., Zhao Z., Luo L., et al. (2020) Effects of glutamate in low-phosphorus diets on growth performance, antioxidant enzyme activity, immune-related gene expression and resistance to aeromonas hydrophila of juvenile mirror carp (Cyprinus carpio). Aquacult. Nutr. 26 (4), 1329–1339. doi: 10.1111/anu.13155
Li X., Wei X., Gu O X., Mi S., Hua X., Li N., et al. (2020). Enhanced growth performance, muscle quality and liver health of largemouth bass (Micropterus salmoides) were related to dietary small peptides supplementation. Aquacult. Nutr. 26 (6), 2169–2177. doi: 10.1111/anu.13155
Macpherson H. L., Bergh Ø, Birkbeck T. H. (2012). An aerolysin-like enterotoxin from vibrio splendidus may be involved in intestinal tract damage and mortalities in turbot, scophthalmus maximus (L.), and cod, gadus morhua l., larvae. J. Fish Dis. 35 (2), 153–167. doi: 10.1111/j.1365-2761.2011.01331.x
Ma A., Dawood M., Shafiei S., Sakhaie F., Seyed E., Shekarabi P. H. (2020) Dietary polygonum minus extract ameliorated the growth performance, humoral immune parameters, immune-related gene expression and resistance against yersinia ruckeri in rainbow trout (Oncorhynchus mykiss) - ScienceDirect. Aquaculture 519, 734–738. doi: 10.1016/j.aquaculture.2019.734738
Ma Y. B., Jiang W. D., Wu P., Liu Y., Feng L. (2021). Tea polyphenol alleviate aeromonas hydrophila - induced intestinal physical barrier damage in grass carp (Ctenopharyngodon idella). Aquaculture 544, 737067. doi: 10.1016/j.aquaculture.2021.737067
Mamauag R., Ragaza J. A., Nacionales T. (2019) Fish performance, nutrient digestibilities, and hepatic and intestinal morphologies in grouper epinephelus fuscoguttatus fed fermented copra meal. Aquacult. Rep. 14, 100202. doi: 10.1016/j.aqrep.2019.100202
Newaj-Fyzul A., Austin B. (2015). Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J. Fish Dis. 38 (11), 937–955. doi: 10.1111/jfd.12313
Niu K., Khosravi S., Kothari D., Lee W., Kim S. i. (2020). Potential of indigenous bacillus spp. as probiotic feed supplements in an extruded low-fish_._eal diet for juvenile olive flounder, paralichthys olivaceus. J. World Aquacult. Soc. 52 (1), 244–261. doi: 10.1111/jwas.12724
Nootash S., Sheikhzadeh N., Baradaran B., Oushani A. K., Moghadam M. M., Nofouzi K., et al. (2013). Green tea (Camellia sinensis) administration induces expression of immune relevant genes and biochemical parameters in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 35 (6), 1916–1923. doi: 10.1016/j.fsi.2013.09.030
Panigrahi A., Kiron V., Satoh S., Hirono I., Kobayashi T., Sugita H., et al. (2007). Immune modulation and expression of cytokine genes in rainbow trout oncorhynchus mykiss upon probiotic feeding. Dev. Comp. Immunol. 31 (4), 372–382. doi: 10.1016/j.dci.2006.07.004
Pérez-Jiménez A., Peres H., Rubio V. C., Oliva-Teles A. (2012). The effect of dietary methionine and white tea on oxidative status of gilthead sea bream (Sparus aurata). Br. J. Nutr. 108 (7), 1202–1209. doi: 10.1017/S0007114511006556
Qian Y.-C., Wang X., Ren J., Wang J., Limbu S. M., Li R.-X., et al. (2021). Different effects of two dietary levels of tea polyphenols on the lipid deposition, immunity and antioxidant capacity of juvenile GIFT tilapia (Oreochromis niloticus) fed a high-fat diet. Aquaculture 542, 736896. doi: 10.1016/j.aquaculture.2021.736896
Rai A. K., Vignesh V. S., Vinothkanna A., Ramesh D. (2015). Isolation of potential probiotic bacillus spp. and assessment of their subcellular components to induce immune responses in labeo rohita against aeromonas hydrophila. Fish Shellfish Immunol. 45 (2), 268–276. doi: 10.1016/j.fsi.2015.04.018
Ramasamy H., Moon Y.-G., Kim M.-C., Kim J.-S., Heo M.-S. (2010). Phytotherapy of aeromonas hydrophila-infected goldfish, carassius auratus. J. World Aquacult. Soc. 41 (3), 391–401. doi: 10.1111/j.1749-7345.2010.00380.x
Reda R. M., Selim K. M., Rania M., El-Araby I. E. (2018). Effect of dietary yeast nucleotide on antioxidant activity, non-specific immunity, intestinal cytokines, and disease resistance in Nile tilapia. Fish Shellfish Immunol. 80, 281–290. doi: 10.1016/j.fsi.2018.06.016
Rico A., Brink P. (2014). Probabilistic risk assessment of veterinary medicines applied to four major aquaculture species produced in Asia. Sci. Total Environ. 468, 630–641. doi: 10.1016/j.scitotenv.2013.08.063
Rietveld A., Wiseman S. (2003). Antioxidant effects of tea: Evidence from human clinical Trials1. J. Nutr 33 (10), 3285S–3292S. doi: 10.1093/jn/133.10.3285S
Safari R., Hoseinifar S. H., Nejadmoghadam S., Jafar A. (2016). Transciptomic study of mucosal immune, antioxidant and growth related genes and non-specific immune response of common carp ( cyprinus carpio ) fed dietary ferula ( ferula assafoetida ). Fish Shellfish Immunol. 55, 242–248. doi: 10.1016/j.fsi.2016.05.038
Sun F., Gu Z., Zhou Q., Sun H., Luo J., Liu Z., et al. (2021). Highly efficient microbial lipid synthesis from co-fermentation of enzymatic hydrolysate of sugarcane bagasse by a Trichosporon dermatis mutant. IND CROP PROD 171, 113975. doi: 10.1016/j.indcrop.2021.113975
Song X., Zhao J., Bo Y., Liu Z., Wu K., Gong C. (2014). Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): An experimental model. Aquaculture 434, 171–178. doi: 10.1016/j.aquaculture.2014.08.015
Tadese D. A., Sun C., Liu B., Muritu R. W., Xu H. (2020). Combined effects of emodin and clostridium butyricum on growth and non-specific immunity of giant freshwater prawns, macrobrachium rosenbergii. Aquaculture 525, 735281. doi: 10.1016/j.aquaculture.2020.735281
Tovar-Ramírez D., Mazurais D., Gatesoupe J. F., Quazuguel P., Cahu C. L., Zambonino-Infante J. L. (2010). Dietary probiotic live yeast modulates antioxidant enzyme activities and gene expression of sea bass (Dicentrarchus labrax) larvae. Aquaculture 300 (1-4), 142–147. doi: 10.1016/j.aquaculture.2009.12.015
ValladãO G., Gallani S. U., Pilarski F. (2015). Phytotherapy as an alternative for treating fish disease. J. Veterinary Pharmacol. Ther. 38 (5), 417–428. doi: 10.1111/jvp.12202
Vazirzadeh A., Roosta H., Masoumi H., Farhadi A., Jeffs A. (2019). Long-term effects of three probiotics, singular or combined, on serum innate immune parameters and expressions of cytokine genes in rainbow trout during grow-out. Fish Shellfish Immunol. 98, 748–757. doi: 10.1016/j.fsi.2019.11.023
Wang Y., Chen L., Huang Z., Zhou C., Yu W., Tan X. (2015). The effects of dietary soybean isoflavones on growth, innate immune responses, hepatic antioxidant abilities and disease resistance of juvenile golden pompano trachinotus ovatus. Fish Shellfish Immunol. 43 (1), 158–166. doi: 10.1016/j.fsi.2014.12.014
Wang J. H., Guo H., Zhang T. R., Wang H., Liu B. N., Xiao S. (2017). Growth performance and digestion improvement of juvenile sea cucumber apostichopus japonicus fed by solid-state fermentation diet. Aquacult. Nutr 23 (6), 1312–1318. doi: 10.1111/anu.12506
Xie Y., Chen Z., Wang D., Chen G., Sun J. (2020). Effects of fermented herbal tea residues on the intestinal microbiota characteristics of Holstein heifers under heat stress. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01014
Xie H., Wang Y., Zhang J., Chen J., Wu D., Wang L. (2015). Study of the fermentation conditions and the antiproliferative activity of rapeseed peptides by bacterial and enzymatic cooperation. Int. J. Food Sci. Technol. 50 (3), 619–625. doi: 10.1111/ijfs.12682
Xv Z., Zhong Y., Wei Y., Zhang T., Zhou W., Jiang Y., et al. (2021). Yeast culture supplementation alters the performance and health status of juvenile largemouth bass (Micropterus salmoides) fed a high-plant protein diet. Aquacult. Nutr. 27 (6), 2637–2650. doi: 10.1111/anu.13391
Xxab C., Xlab C., Zhen X., Wyab C., Xlab C. (2021). Dietary azomite, a natural trace mineral complex, improved the growth, immunity response, intestine health and resistance against bacterial infection in largemouth bass ( micropterus salmoides ). Fish Shellfish Immunol. 108, 53–62. doi: 10.1016/j.fsi.2020.11.016
Yeganeh Rastekenari H., Kazami R., Shenavar Masouleh A., Banavreh A., Najjar Lashgari S., Sayed Hassani M. H., et al. (2021). Autochthonous probiotics lactococcus lactis and weissella confusa in the diet of fingerlings great sturgeon, huso huso : effects on growth performance, feed efficiency, haematological parameters, immune status and intestinal morphology. Aquacult. Res. 52 (8), 3687–3695. doi: 10.1111/are.15213
Yuan Y., Wang X., Jin M., Sun P., Zhou Q. (2019). Influence of different lipid sources on growth performance, oxidation resistance and fatty acid profiles of juvenile swimming crab, portunus trituberculatus - ScienceDirect. Aquaculture 508, 147–158. doi: 10.1016/j.aquaculture.2019.04.068
Yu H., Yang Q., Liang H., Ren M., Ge X., Ji K. (2021) Effects of stocking density and dietary phosphorus levels on the growth performance, antioxidant capacity, and nitrogen and phosphorus emissions of juvenile blunt snout bream (Megalobrama amblycephala). Aquacult. Nutr 27 (2), 581–591. doi: 10.1111/anu.13208
Yu H., Zhang C., Zhang X., Wang C., Ji H. (2019). Dietary nano: elenium enhances antioxidant capacity and hypoxia tolerance of grass carp ctenopharyngodon idella fed with high-fat diet. Aquacult. Nutr. 26 (2), 545–557. doi: 10.1111/anu.13016
Zhang Y., Zhou Y. B., Sang B. Y., Wan X. C., Yang Y. O., Zhang J. L., et al. (2015). Effect of dietary Chinese tea on growth performance, disease resistance and muscle fatty acid profile of channel catfish (Ictalurus punctatus). Aquacult. Int 23 (2), 683–698 doi: 10.1007/s10499-014-9846-7
Zheng Q., Han C., Zhong Y., Wen R., Zhong M. (2017). Effects of dietary supplementation with green tea waste on growth, digestive enzyme and lipid metabolism of juvenile hybrid tilapia, oreochromis niloticus x o. aureus. Fish Physiol. Biochem. 43 (2), 361–371. doi: 10.1007/s10695-016-0292-5
Keywords: tea residue, largemouth bass, growth performance, immune capacity, A. hydrophila
Citation: Jiang L, Zhou X, Yu J, Bao S, Li J, Wu Q, Wu M, Wang Y and Liu B (2022) Fermented tea residue improved growth performance, liver antioxidant capacity, intestinal morphology and resistance to Aeromonas hydrophila infection in juvenile largemouth bass (Micropterus salmoides). Front. Mar. Sci. 9:999947. doi: 10.3389/fmars.2022.999947
Received: 21 July 2022; Accepted: 09 August 2022;
Published: 30 August 2022.
Edited by:
Lei Wang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Chuanpeng Zhou, Chinese Academy of Fishery Sciences (CAFS), ChinaHadiseh Dadras, South Bohemia Research Center of Aquaculture and Biodiversity of Hydrocenoses, Czechia
Copyright © 2022 Jiang, Zhou, Yu, Bao, Li, Wu, Wu, Wang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yachao Wang, d2FuZ3lhY2hhbzE2ODhAMTI2LmNvbQ==; Bo Liu, bGl1YkBmZnJjLmNu
†These authors have contributed equally to this work and share first authorship