 Brigitte Gavio
Brigitte Gavio Julián Prato
Julián Prato Mariana Gnecco4
Mariana Gnecco4 María Fernanda Maya
María Fernanda Maya José Ernesto Mancera-Pineda
José Ernesto Mancera-Pineda- 1Biology Department, Universidad Nacional de Colombia, Bogotá, Colombia
- 2Universidad Nacional de Colombia, Sede Caribe, San Luis Free Town, Colombia
- 3Corporation Center of Excellence in Marine Science-CEMarin, Bogotá, Colombia
- 4Blue Indigo Foundation, San Andrés isla, Colombia
Hurricanes and tropical storms are major climatic disturbances with potentially catastrophic effects on coastal and marine ecosystems. The impact of these climatic events on coastal communities may vary greatly and depends on the hurricane’s severity, trajectory, and duration. Furthermore, the impact of hurricanes can be patchy, causing extensive damage in some locations, while leaving other subjacent areas intact. In coral reef areas, breakage, dislodgement, and sandblasting may cause extensive coral mortality, and loss of coral cover may increase due to sedimentation and freshwater runoff. After the impact of the climatic event, successional stages in the reef community have been observed, where blooms of ephemeral algae may occupy space made available by the storm impact. We report a bloom of the red alga Trichogloeopsis pedicellata on the west coast of the island of San Andrés, following Hurricane Iota. The survey was carried out in seven locations at different depths: shallow (2–5 m) and deep (8–15 m). Three video transects were recorded at each sampling site, and 15 photo quadrats were analyzed for benthic cover. For the biomass estimate, three to five quadrats of 25 × 25 cm were laid on the substrate, all the algae inside were collected, and their wet biomass was determined. The algal cover reached 100% in some locations, with wet biomass up to 5,264 g/m2, and persisted for several weeks. The bloom was more severe at shallow depths. Apparently, the availability of the substrate due to the detachment of foliose algae triggered the bloom. After a few months, the bloom receded naturally.
Introduction
Hurricanes and tropical storms are major climatic disturbances with potentially catastrophic effects on both marine and terrestrial ecosystems. Damages are the result of hurricane-associated strong winds (which can exceed 250 km/h), storm surges, heavy rainfall, flooding, and stormwater runoff. On average, hurricanes are getting stronger, with a recent trend that is related to an increase in ocean temperatures (Emanuel, 2005; Webster et al., 2005). An intensification in both hurricane frequency and intensity due to global warming is expected in the near future (Knutson et al., 2010; Sobel et al., 2016). The impact of these climatic events on coastal communities may vary greatly and depends on the hurricane itself and the location (Anton et al., 2009). Furthermore, the impact of hurricanes can be patchy, causing extensive damage in some locations but leaving others intact (Edmunds and Witman, 1991; Anton et al., 2009).
In coral reef areas, breakage, dislodgement, and sandblasting may cause extensive coral mortality, and loss of coral cover may increase due to sedimentation and freshwater runoff (Edmunds, 2019). After the impact of the climatic event, successional stages in the reef community have been observed, where blooms of ephemeral algae may occupy space made available by the storm’s impact (e.g., Hughes, 1994). These blooms have been documented in the Caribbean in the 1980s and 1990s, consisting mainly of the green alga Trichosolen (Woodley et al., 1981) or red algae belonging to the genus Liagora (Rogers et al., 1982; Edmunds and Witman, 1991). More recently, similar blooms, composed of species belonging to the same family of algae, were reported for the Pacific and Indian Oceans (Littler and Littler, 1999; Jupp, 2007; Doropoulos et al., 2014), following typhoon events. These events tend to share the following characteristics (Pauly et al., 2011): they are generally 1) monospecific, 2) widespread in space, 3) following severe physical impact, 4) persisting for weeks regardless of herbivore abundance, and 5) previously unknown in the sites despite elaborate prior floristic work.
The Caribbean coast of Colombia is located south of the hurricane belt of the region, and it is normally spared such climatic events. From 1900 to 2010, only 12 storms significantly affected the coast of the country, and of these, just four hit land in Colombian territory (Ortiz Royero, 2012). The year 2020 had an exceptionally active hurricane season in the Caribbean Sea, with 30 named storms, 13 of which became hurricanes (Probst et al., 2021). At the very end of the storm season, Hurricane Eta (category 4) and Hurricane Iota (category 5) passed close to the Archipelago of San Andrés, Old Providence, and Santa Catalina, causing considerable damage (Gómez et al., 2022).
A few weeks after these climatic events, a large bloom of the red alga Trichogloeopsis pedicellata (M.Howe) I. A. Abbott and Doty was observed in San Andrés. It is the first time that the species has been observed at the site, and it is the first report of such a bloom for San Andrés. The objective of this work was to determine the extension, cover, and wet biomass of the bloom along the coast of San Andrés island.
Materials and methods
Study site
San Andrés (12°28′N; 81°40′W) is an oceanic island of coral origin located in the southwestern Caribbean, Colombia (Figure 1). On the eastern side, the island has a barrier reef running parallel to the coast, which encloses a shallow lagoon. The western side is characterized by two submerged terraces, parallel to the coastline: the first terrace is shallow (4–10-m depth), while the second is deeper (10–20-m depth) (Chaves-Fonnegra et al., 2007).
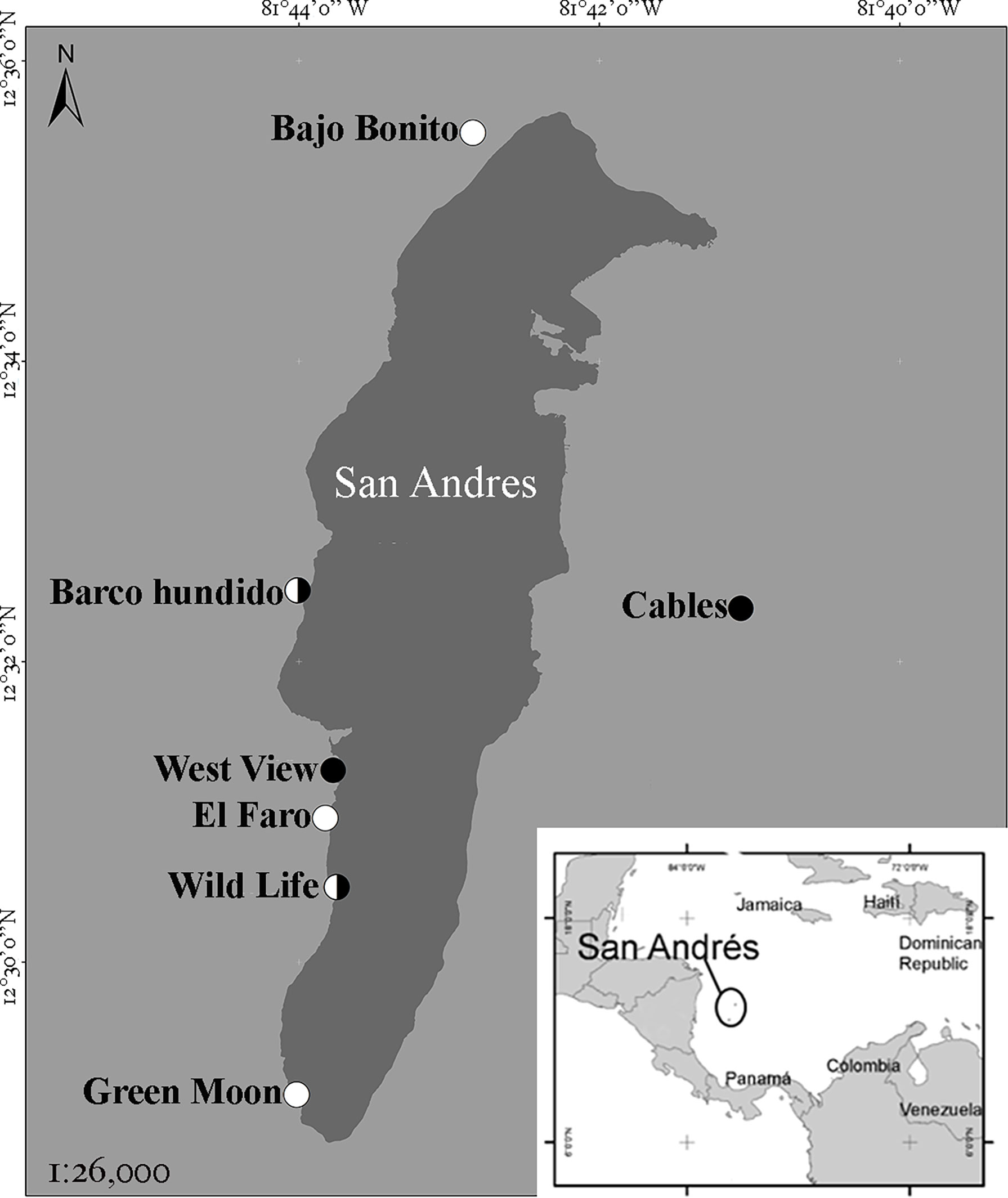
Figure 1 Sample sites. White dots are shallow sampling, black dots are deep sampling, and the half-white-and-half-black dots are both shallow and deep sampling.
The island lies south of the hurricane belt of the Caribbean Sea. However, at least 12 hurricanes or tropical storms have affected the island since 1900 (Ortiz Royero, 2012), with the strongest being category 3 Hurricane Hattie in 1961.
Hurricane Eta hit the island on 2 November 2020 as a category 4. Two weeks after Eta, Hurricane Iota, category 5, passed about 130 km north of the island on 16 November (Gómez et al., 2022). Both events caused very strong winds, rain, and storm waves on the island, especially on the western side, which was the most affected.
The immediate effects on the coastal ecosystems on the west coast of the island were visible a few days after the storm: broken coral and removed benthic species, especially fleshy and foliose algae, which dominated the rocky bottom before the storms. A few months after the hurricane’s passage, a bloom of a gelatinous red alga was observed.
Bloom survey
Between 26 January and 2 February 2021, the western coast of the island was surveyed (Figure 1). Both terraces (shallow and deep) were sampled. The eastern coast of the island was also surveyed, but no bloom was present. At each site, three 10-m-long video transects were realized with a GoPro Hero8 Black to determine the cover of the main categories of benthos, as well as the blooming alga (methodology modified from Castaño et al., 2021). The camera was mounted on a holder for greater stability, and the video was filmed at a constant distance (50 cm) from the bottom. The duration of each recording was about 4 min. Along the same transect, three to five 25 × 25 cm quadrants were placed to collect the algal biomass inside it. Additional specimens were collected separately for identification. The species identification was performed in the laboratory with specialized literature (Littler and Littler, 2000; Dawes and Mathieson, 2008). The video transects were analyzed in 15 photo quadrats per transect using ImageJ software (Sánchez et al., 2019) for the main benthic categories (algae, dead coral, live coral, sponges, sand, other algae, and rocks). Data were tested for normality with the Shapiro–Wilk test. Since the data were not normal, Kruskal–Wallis and Duncan tests were performed to determine differences among sites. The Wilcoxon test was performed to determine whether there were differences between depths. A regression analysis was performed between cover and biomass.
Results
Taxonomical identification
Trichogloeopsis pedicellata (M. Howe) I. A. Abbott and Doty
Thallus erect, attached to a hard substrate by a small holdfast; alga soft, gelatinous, 10–15-cm tall, pyramidal in outline, light pink to white in color (Figure 2A). Calcification mainly in outer mucus. Branching irregularly alternate. Cortex formed by dichotomously branched filaments of small cylindrical cells, round and moniliform near tips (Figure 2B); apical cells 12–20-μm diameter, 15–26 μm long, may bear colorless hairs. Medullary filaments longitudinal, cylindrical, 30–100-μm diameter (Figure 2C). Carposporophytes on outer cortical cells, 90–220-μm diameter. Cystocarps are round, without involucre, with descending rhizoids at the base (Figure 2D).
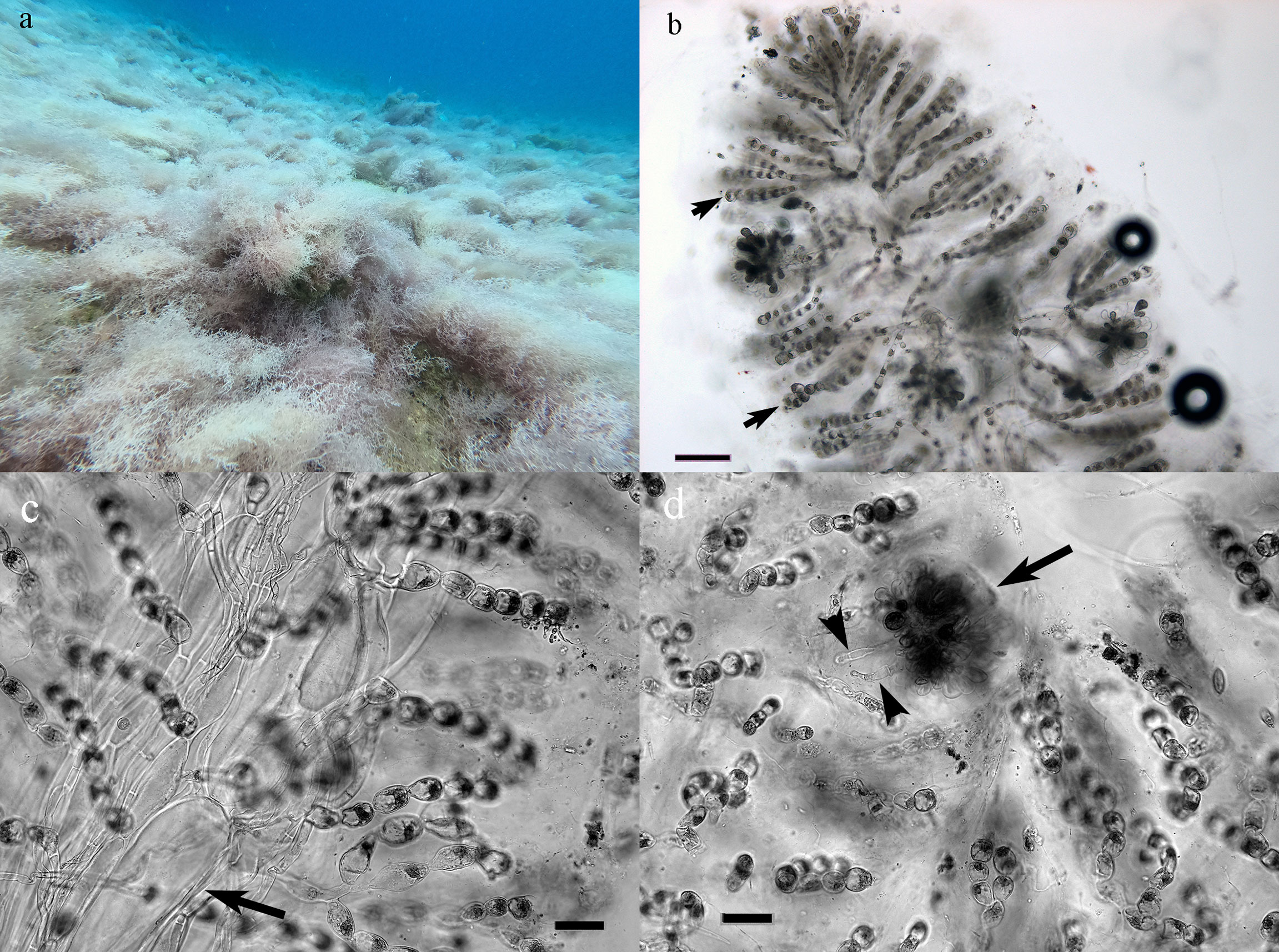
Figure 2 Trichogloeopsis pedicellata. (A) Bloom with almost 100% cover. (B) Squash mounting showing cortical cells (arrows). Scale bar: 100 μm. (C) Detail of medullary filament (arrow). Scale bar: 50 μm. (D) Cystocarp (arrow) with descending filaments (arrowheads) at the base. Scale bar: 50 μm.
Remarks: The species has not previously been reported for the Archipelago of San Andrés, Old Providence, and Santa Catalina. However, specimens of similar-looking species have been collected long before this blooming event. The family Liagoraceae includes many calcified species with similar vegetative morphologies, which often causes taxonomic confusion and makes field and laboratory identification difficult (Lin et al., 2013). Due to these difficulties, it is highly probable that this alga has been overlooked and not identified in the past.
The main characters used to separate the genera in the family are based largely on differences in postfertilization stages, such as the aspect of the carposporophyte (compact or diffuse), whether the cells of carpogonial branches fuse or remain discrete during carposporophyte development, and the presence or absence of involucral filaments, which may surround the developing carposporophyte densely or laxly (de Castro Nunes, 2005; Lin et al., 2015). In 1960, Abbott and Doty proposed a new genus, Trichogloeopsis, based on the following characteristics: the production of gonimoblastic rhizoids, the presence of compact naked carposporophytes, owing to the absence of sterile postfertilization filaments, and the particular architecture of carpogonial branches that occurs frequently in the terminal few cells of a cortical filament (Abbott and Doty, 1960; Kraft, 1989). Abbott (1970) mentioned that in Trichogloeopsis, the filaments in the center of the core of axial filaments are larger in diameter than those toward the periphery. All these characteristics are consistent with those observed in our specimens, which fit the description of the species T. pedicellata.
This taxon is the only species of Trichogloeopsis present in the Caribbean Sea (Wynne, 2017), being reported in Florida, the Bahamas, the Greater Antilles, the Lesser Antilles, the Gulf of Mexico, and Western Caribbean (Littler and Littler, 2000).
Cover and wet biomass
The cover of T. pedicellata ranged from 5% to 100% (Figure 2A), with means (per transect) ranging from 13% to 93%. The most affected site was Wild Life, while Green Moon showed the lowest cover (Figure 3A, Table 1). From north to south, there was an increase to the maximum cover at the shallow terrace in Wild Life and then a decrease to a minimum at Green Moon (Figure 3A, Table 1). The shallow terrace was more affected than the deep one (p = 0.0001) (Figure 3B).
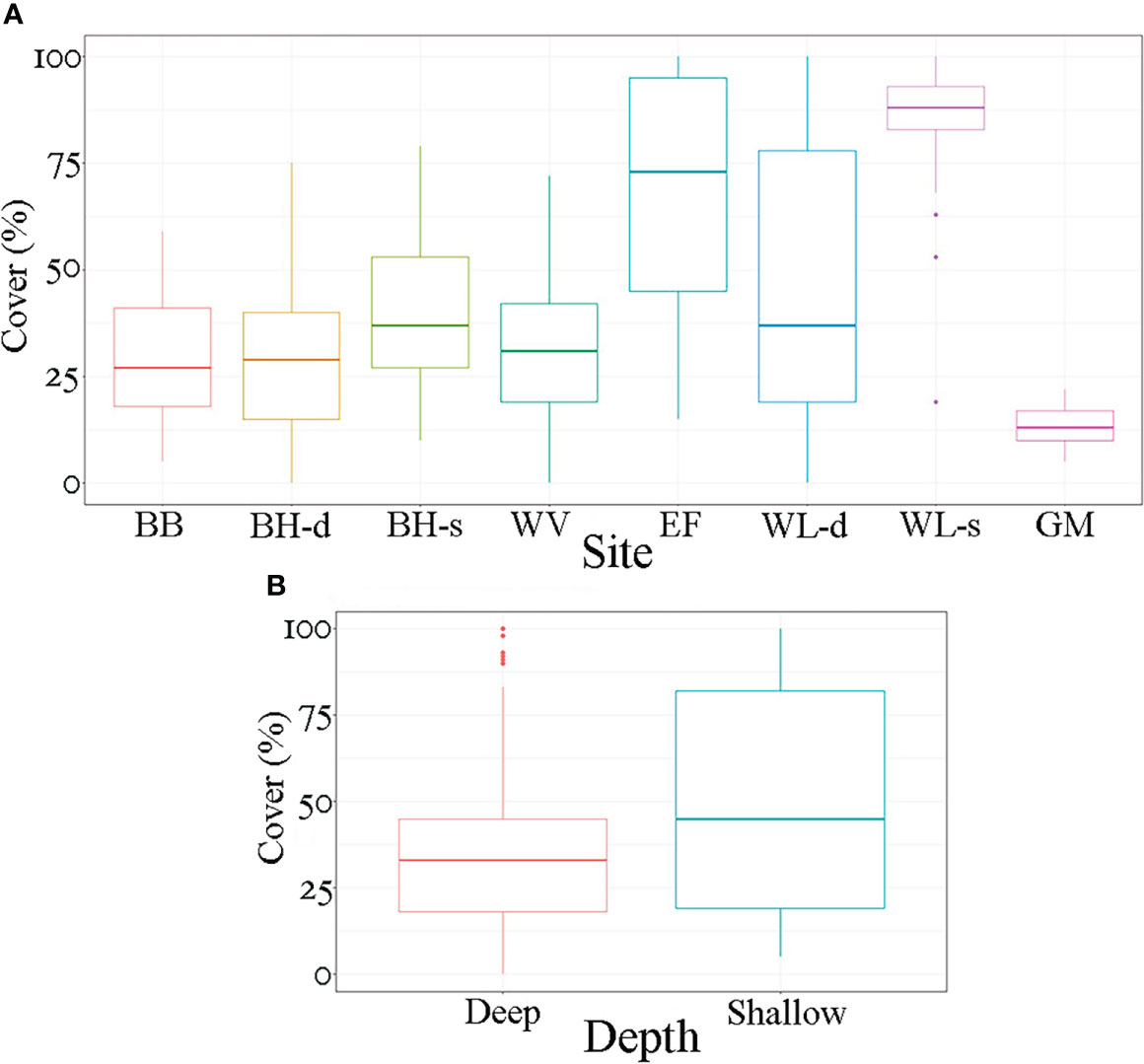
Figure 3 (A) Box plot of cover (%) of Trichogloeopsis pedicellata at each sampling site. Line inside the box represents median; box represents 25th and 75th percentile (top and bottom range, respectively); whiskers represent 10th and 90th percentile (top and bottom whisker, respectively). Points outside the whiskers represent outliers. BB, Bajo Bonito; BH-d, Barco Hundido Deep; BH-s, Barco Hundido Shallow; WV, West View; EF, El Faro; WL-d, Wild Life Deep; WL-s, Wild Life Shallow; GM, Green Moon. (B) Box plot of cover (%) of T. pedicellata at shallow and deep sites. Line inside the box represents median; box represents 25th and 75th percentile (top and bottom range, respectively); whiskers represent 10th and 90th percentile (top and bottom whisker, respectively). Points outside the whiskers represent outliers.
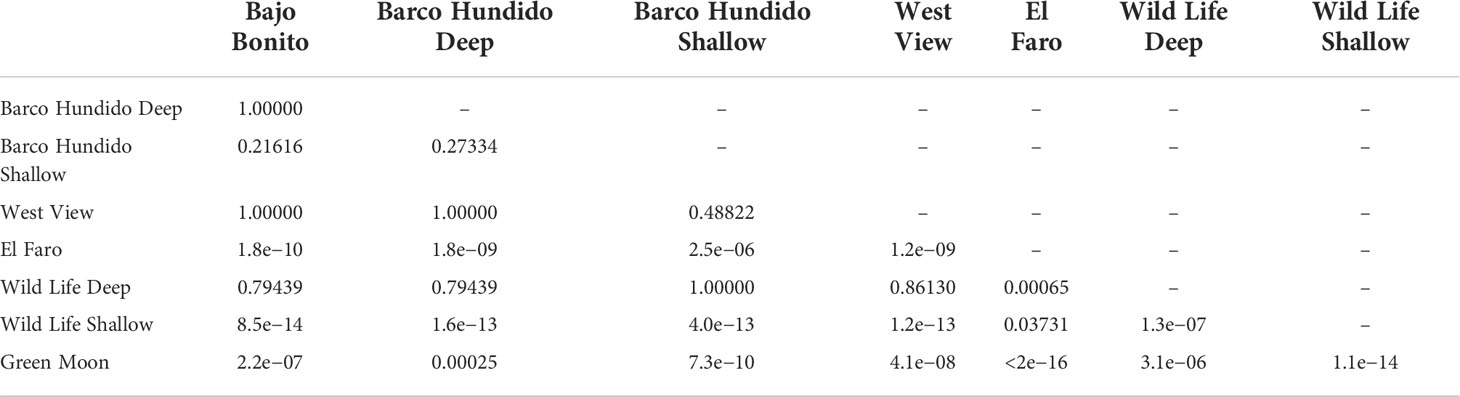
Table 1 p-Value (Duncan test) of cover percentage among sites.
Wet biomass (g/m2) ranged from a mean of 475 g/m2 in Green Moon to 3,387 g/m2 in Wild Life (shallow platform), with a maximum of 5,264 g/m2.
Discussion
Hurricane Iota was a major climatic event when it hit San Andrés. In coastal environments, the west coast of the island was the most affected, while the reef lagoon on the eastern side did not present visible damage. On the west coast, the waves hit with particular strength and caused the detachment of benthic organisms, especially in the shallow terrace, which was dominated by foliose macroalgae before the climatic event. After the hurricane, the free space available was gradually occupied by T. pedicellata, where its cover increased from 0% to up to 100% at some sites (Figure 2A). This phenomenon has been widely reported in the past in the Caribbean Sea. One of the first observations of such successional events was made by Woodley et al. (1981): shortly after the passage of Hurricane Allen north of Jamaica, they reported a great bloom of the green alga Trichosolen duchassaingii in shallow waters. The bloom persisted for about a month, and the species was replaced by the red algae Crouania pleonospora and Liagora sp. Similarly, Rogers et al. (1982) reported a bloom of Gloiocallis dendroidea (cited as Liagora mucosa) 1 month after the passage of Hurricanes David and Frederic in the Virgin Islands. Five months later, Gloiocallis was replaced by Dictyopteris delicatula. After the passage of Hurricane Hugo in the Virgin Islands, Edmunds and Witman (1991) reported a cover increase of Liagora spp., from 0% to 11%. The authors observed that the algae were not growing on living corals but occupied the free space made available by the hurricane.
More recently, the same phenomenon has been observed in the Pacific Ocean. Roff et al. (2015) reported an extensive bloom of Liagora spp. following Typhoon Bopha at Palau, reaching covers up to 43% at wave-exposed sites. The authors linked the explosive growth of Liagora with wave exposure, discarding other hypotheses such as lower herbivory pressure or a nutrient input from terrestrial sources (Hughes, 1994; Scheffer et al., 2008).
San Andrés has not been hit by major hurricanes frequently in the past, and this is the first time such a bloom has been observed. The bloom was more pronounced at shallow sites, and toward the center of the island, while it was less extensive at both the extreme north and south of the island.
Data availability statement
The datasets presented in this article are not readily available because only the ID of the alga was generated, no molecular analyses have been processed. Requests to access the datasets should be directed to YmdhdmlvQHVuYWwuZWR1LmNv.
Author contributions
BG designed the methodology, identified the species, analyzed the data, and wrote the manuscript. JP and MG designed the methodology, did the fieldwork, and processed and analyzed the data. MFM designed the methodology and analyzed the data. JEMP analyzed the data and wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was partially funded by Blue Indigo Foundation. Additional funding included Colciencias Ph.D. scholarship (Conv. 757) and Corporation Center of Excellence in Marine Science-CEMarin by the CEMarin Call 14, 2018, which funded the maintenance of the CEMarin young researcher Julián Prato and the project “Relationships between coral reef complexity and ecosystem services at Caribbean oceanic islands, Seaflower Biosphere Reserve, Colombia”, which contributed to this manuscript.
Acknowledgments
We wish to thank John Carvajal for his help in the field. Manuela Corrales weighed the algal biomass, for which she is acknowledged. We thank Dr John Huisman, who confirmed the identification of the alga.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbott I. A. (1970). Yamadaella, a new genus in the Nemaliales (Rhodophyta). Phycologia 9 (2), 115–123. doi: 10.2216/i0031-8884-9-2-115.1
Abbott I. A., Doty M. S. (1960). Studies in the Helminthocladiaceae. II. Trichogloeopsis. Amer. J. Bot. 47, 632–640. doi: 10.1002/j.1537-2197.1960.tb07146.x
Anton A., Cebrian J., Duarte C. M., Heck K. L. Jr., Goff J. (2009). Low impact of hurricane Katrina on seagrass community structure and functioning in the Northern Gulf of Mexico. Bull. Mar. Sci. 85 (1), 45–59. http://hdl.handle.net/10261/88908
Castaño D., Morales-de-Anda D., Prato J., Cupul-Magaña A. L., Echeverry J. P., Santos-Martínez A. (2021). Reef structural complexity influences fish community metrics on a remote oceanic island: Serranilla island, Seaflower Biosphere Reserve, Colombia. Oceans 2, 611–623. doi: 10.3390/oceans2030034
Chaves-Fonnegra A., Zea S., Gómez M. L. (2007). Abundance of the excavating sponge Cliona delitrix in relation to sewage discharge at San Andrés island, SW Caribbean, Colombia. Boletín Investigaciones Marinas y Costeras 36 (1), 63–78. doi: 10.25268/bimc.invemar.2007.36.0.201
Dawes C. J., Mathieson A. C. (2008). The seaweeds of Florida (Gainesville, United States of America: University Press of Florida), 592.
de Castro Nunes J. M. (2005). A familia Liagoraceae (Rhodophyta, Nemaliales) no estado da Bahia, Brasil. Hoehnea 32 (3), 429–444.
Doropoulos C., Roff G., Zupan M., Nestor V., Isechal A. L., Mumby P. J. (2014). Reef-scale failure of coral settlement following typhoon disturbance and macroalgal bloom in Palau, Western Pacific. Coral Reefs 33 (3), 613–623. doi: 10.1007/s00338-014-1149-y
Edmunds P. J. (2019). Three decades of degradation lead to diminished impacts of severe hurricanes on Caribbean reefs. Ecology 100 (3), e02587. doi: 10.1002/ecy.2587
Edmunds P. J., Witman J. D. (1991). Effect of hurricane Hugo on the primary framework of a reef along the south shore of St. John, US Virgin Islands. Mar. Ecol. Prog. Ser. 78, 201–204. doi: 10.3354/meps078201
Emanuel K. A. (2005). Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436 (7051), 686–688. doi: 10.1038/nature03906
Gómez C. E., Millán S., Acosta-Chaparro A., Abril-Howard A., Navas-Camacho R. (2022). Preliminary assessment of the impact of hurricane Iota on coral reefs in Providencia and Santa Catalina islands. Boletín Investigaciones Marinas y Costeras 51 (1), 193–200. doi: 10.25268/bimc.invemar.2022.51.1.1164
Hughes T. P. (1994). Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265, 1547–1551. doi: 10.1126/science.265.5178.1547
Jupp B. P. (2007). Report on destruction of corals on reef ball site, Fahal island, Muscat. Rapid assessment of impacts from cyclone Gonu. Ministry Environ. Climate Affairs Muscat.
Knutson T. R., McBride J. L., Chan J., Emanuel K., Holland G., Landsea C., et al. (2010). Tropical cyclones and climate change. Nat. Geosci. 3, 157–163. doi: 10.1038/ngeo779
Kraft G. T. (1989). Cylindraxis rotundatus gen. et sp. nov. and its generic relationships within the Liagoraceae (Nemaliales, Rhodophyta). Phycologia 28, 275–304. doi: 10.2216/i0031-8884-28-3-275.1
Lin S. M., Huisman J. M., Payri C. (2013). Characterization of Liagora ceranoides (Liagoraceae, Rhodophyta) on the basis of rbcL sequence analyses and carposporophyte development, including Yoshizakia indopacifica gen. et sp. nov. from the indo-pacific region. Phycologia 52 (2), 161–170. doi: 10.2216/12-101.1
Lin S. M., Rodríguez-Prieto C., Huisman J. M., Guiry M. D., Payri C., Nelson W. A., et al. (2015). A phylogenetic re-appraisal of the family Liagoraceae sensu lato (Nemaliales, Rhodophyta) based on sequence analyses of two plastid genes and postfertilization development. J. Phycol. 51, 546–559. doi: 10.1111/jpy.12301
Littler M. M., Littler D. S. (1999). Disturbances due to cyclone Gavin parallel those caused by a ship grounding. Coral Reefs 18, 146. doi: 10.1007/s003380050169
Littler D. S., Littler M. M. (2000). Caribbean Reef plants. An identification guide to the reef plants of the Caribbean, Bahamas, Florida and gulf of Mexico (Washington D.C., USA: OffShore Graphics, Inc), 543.
Ortiz Royero J. C. (2012). Exposure of the Colombian Caribbean coast, including San Andrés island, to tropical storms and hurricane, 1900–2010. Nat. Hazards 61, 815–827. doi: 10.1007/s11069-011-0069-1
Pauly K., Jupp B. P., De Clerck O. (2011). Modelling the distribution and ecology of Trichosolen blooms on coral reefs worldwide. Mar. Biol. 158, 2239–2246. doi: 10.1007/s00227-011-1729-0
Probst P., Annunziato A., Proietti C., Paris S. (2021). 2020 - Atlantic hurricane season: A record-breaking season, EUR 30635 EN. Publications Office Eur. Union Luxembourg. 33pp. doi: 10.2760/00114
Roff G., Chollett I., Doropoulos C., Golbuu Y., Steneck R. S., Isechal A. L., et al. (2015). Exposure-driven macroalgal phase shift following catastrophic disturbance on coral reefs. Coral Reefs 34, 715–725. doi: 10.1007/s00338-015-1305-z
Rogers C. S., Suchanek T. H., Pecora F. A. (1982). Effects of hurricanes David and Frederic (1979) on shallow Acropora palmata reef communities: St. Croix, U.S. Virgin Islands. Bull. Mar. Sci. 32 (2), 532–548.
Sánchez J. A., Gómez-Corrales M., Gutierrez-Cala L., Vergara D. C., Roa P., González-Zapata F. L., et al. (2019). Steady decline of corals and other benthic organisms in the SeaFlower Biosphere Reserve (Southwestern Caribbean). Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00073. article 73.
Scheffer M., van Nes E. H., Holmgren M., Hughes T. (2008). Pulse-driven loss of top-down control: the critical-rate hypothesis. Ecosystems 11, 226–237. doi: 10.1007/s10021-007-9118-8
Sobel A. H., Camargo S. J., Hall T. M., Lee C. Y., Tippett M. K., Wing A. A. (2016). Human influence on tropical cyclone intensity. Science 353 (6296), 242–246. doi: 10.1126/science.aaf6574
Webster P. J., Holland G. J., Curry J. A., Chang H. R. (2005). Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309, 1844–1846. doi: 10.1126/science.1116448
Woodley J. D., Chornesky E. A., Clifford P. A., Jackson J. B. C., Kaufman L. S., Knowlton N., et al. (1981). Hurricane Allen's impact on Jamaican coral reefs. Science 214 (4522), 749–755. doi: 10.1126/science.214.4522.749
Keywords: macroalgal bloom, ephemeral algae, disturbance, available substrate, climatic event
Citation: Gavio B, Prato J, Gnecco M, Maya MF and Mancera-Pineda JE (2022) Bloom of Trichogloeopsis pedicellata (Rhodophyta, Nemaliales) following hurricane Iota in San Andrés, Southwestern Caribbean Sea. Front. Mar. Sci. 9:978869. doi: 10.3389/fmars.2022.978869
Received: 26 June 2022; Accepted: 01 September 2022;
Published: 27 September 2022.
Edited by:
Ricardo Bermejo, University of Malaga, SpainReviewed by:
Ricardo A. Scrosati, St. Francis Xavier University, CanadaAlejandra Piñón-Gimate, Centro Interdisciplinario de Ciencias Marinas (IPN), Mexico
Copyright © 2022 Gavio, Prato, Gnecco, Maya and Mancera-Pineda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Brigitte Gavio, YmdhdmlvQHVuYWwuZWR1LmNv