 Ricardo Cardoso Neves
Ricardo Cardoso Neves Reinhardt Møbjerg Kristensen
Reinhardt Møbjerg Kristensen Axayacatl Rocha-Olivares
Axayacatl Rocha-Olivares Gerardo Rivas
Gerardo Rivas- 1Section for Biosystematics, Natural History Museum of Denmark, Copenhagen, Denmark
- 2Departamento de Oceanografía Biológica, Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), Ensenada, Mexico
- 3Departamento de Biología Comparada, Facultad de Ciencias, Universidad Nacional Autónoma de México (UNAM), Ciudad de México, Mexico
Loricifera is a group of marine, meiobenthic invertebrates of microscopic size that are found worldwide. The Loriciferan fauna of the Southern Gulf of Mexico (SGM) remains unknown. In the present study we report on the first exploration of the Loriciferans from the SGM, which is a strategic area for oil exploitation. The Loriciferan specimens were collected during deep-sea cruises XIXIMI-4 (August-September 2015) and XIXIMI-6 (August 2017), in which six out of 22 stations yielded specimens of Loricifera. A total of six specimens were found (although one of them was lost), namely two adults and four larval stages. Preliminary analyses based on the peculiarity of their morphological features indicate that the specimens include representatives of new species of Pliciloricus and Rugiloricus, an undescribed genus of Pliciloricidae and, possibly, an undescribed genus of Nanaloricidae. None of the specimens found could be assigned to previously described species. Since all specimens are either intermediate life cycle stages or poorly preserved, only a short description based on their morphological traits is provided here. Our findings are discussed from a comparative perspective with known Loriciferan species.
Introduction
Loricifera is a phylum of exclusively marine, meiobenthic invertebrates of microscopic size (Kristensen, 1983; Bang-Berthelsen et al., 2013; Neves et al., 2016). These free-living animals are found worldwide in shell gravel, coarse sand and muddy sediments at a wide range of depths (for a review see Neves et al., 2016). Despite an increasing research effort, the patterns of spatial distribution of Loriciferan species remain scarcely known (Fujimoto and Kristensen, 2020; Neves et al., 2021). Described in 1983, Loricifera encompasses at present 44 species that are accommodated into one order, Nanaloricida Kristensen, 1983, and three families, Nanaloricidae Kristensen, 1983, Pliciloricidae Higgins and Kristensen, 1986, and Urnaloricidae Heiner and Kristensen, 2009 (Kristensen, 1983; Higgins and Kristensen, 1986; Heiner and Kristensen, 2009; Neves et al., 2016). In addition, Tenuiloricus shirayamai Neves and Kristensen, 2014 and Patuloricus tangaroa Sørensen et al., 2022 are listed as incertae sedis within Loricifera (Neves and Kristensen, 2014; Sørensen et al., 2022).
The Loriciferan life cycle involves a succession through several larval and post-larval (= juvenile) stages leading to the adult forms (Kristensen and Brooke, 2002; Neves et al., 2016). Adults and post-larvae possess a bilateral body divided into a head (mouth cone and introvert), neck, thorax and abdomen (Kristensen, 1983; Kristensen 1991a; Kristensen 1991b; Neves et al., 2016). Their introvert is characterized by several rings of leg- or spine-like appendages (termed clavoscalids and spinoscalids), while the neck possesses flattened appendages called trichoscalids. The abdomen is encased in a lorica composed of several cuticular plates (Nanaloricidae) or plicae (Pliciloricidae). The most prominent larval stage – named the Higgins larva (which includes the Shira larva type) – is anatomically similar to the adult but possesses a pair of “toes” and two groups of lorical setae arranged into 2-3 pairs situated at the posteriormost region of the body (Neves et al., 2016; Sørensen et al., 2022).
The Loriciferan fauna of the Gulf of Mexico (GM) remains poorly explored Hubbard et al. 1988 analyzed samples taken from various depths (298 - 2959 m) during five cruises (1983 and 1985) in the northern GM, which yielded a total of 506 Loriciferan specimens. This material has not been thoroughly investigated nor resulted in formal descriptions of new species. In 2005, twenty of these Loriciferan specimens were loaned to the Zoological Museum of Copenhagen for observations under a light microscope (RK, pers. obs.). However, none of them was thoroughly investigated and these preliminary observations did not result in formal descriptions of new species.
In the present study, we show results from the first Loriciferans found in the Southern Gulf of Mexico (SGM) an economically important zone for hydrocarbon exploration and exploitation (Peters et al., 2021). Ensuing the Deep-Water horizon oil spill, the risks associated with exploration and production of oil in this area have recently attracted greater attention from scientists (Kujawinski et al., 2020), among other aspects due to the risk of biodiversity loss due to the strong anthropogenic activity. As a result, the Gulf of Mexico Research Consortium (CIGOM: https://cigom.org/), was created to carry out an interdisciplinary and multi-institutional project “Implementation of networks of physical, geochemical, and ecological oceanographic observations to generate scenarios for possible contingencies related to the exploration and production of hydrocarbons in the deep waters of the Gulf of Mexico”. This project (2015-2020) included the XIXIMI oceanographic campaigns in the Mexican exclusive economic zone (EEZ) of the SGM, which aimed at sampling the deep-sea benthos (> 500 m) and included seven cruises and a total of 68 stations.
Materials and methods
The Loriciferan specimens shown here were collected during deep-sea cruises (1200–3524 m; Supplementary Figure 1) within the Mexican EEZ of the SGM, led by the Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE). Sediment cores were obtained on board the R/V Justo Sierra (Universidad Nacional Autónoma de México) during XIXIMI-4 (August-September 2015) and XIXIMI-6 (August 2017). Sampling was carried out along horizontal transects (named with letters A, B, C, D, E, F and G) to cover different latitudinal degrees (25°, 24°, 23°, 22°, 21°, 20° and 19° latitude north respectively). Samples (12 cm-deep cores) were taken using boxcorer (sediment core subsampled with an acrylic core of internal diameter 8.1 cm) or multicorer (internal diameter 10 cm) which were preserved in 10% buffered formalin. Meiofauna was extracted from the sediment matrix by decantation and flotation with colloidal silica (Ludox ™, specific gravity 1.15 g cm−3 (de Jonge and Bouwman, 1977) followed by sieving through a 1000 μm mesh, and retained on a 45 μm mesh (Somerfield and Warwick, 1996). The process was repeated three times to maximize the number of extracted organisms. Sieved organisms were fixed and stored in 10% buffered formalin. Loriciferan specimens were sorted with the help of a stereomicroscope, transferred to glass slides in distilled water and photographed under cover slips using a Zeiss AxioCam 506 camera mounted on a ZeissAxio Scope A1 compound microscope. Subsequently, the specimens were mounted in Fluoromount G (SouthernBiotech) on glass slides and investigated with an Olympus BX51 microscope fitted with phase contrast (PC) and DIC optics. Photographs were taken with an Olympus DP20 Cell zoom digital camera.
The Loriciferans investigated in this study – a total of 5 specimens, though a sixth specimen was lost during the investigation – are deposited at the Zoological Museum, Natural History Museum of Denmark under accession numbers NHMD-621042 to NHMD-621046. General information on the various stations containing Loriciferans as well as information on the specimens are summarized in Supplementary Table 1.
Results
Loriciferans were found in five out of the 14 deep-sea stations inspected during XIXIMI-4 (August-September 2015) and in one out of eight stations during XIXIMI-6 (August 2017). A total of six specimens were recorded (see Supplementary Table 1) – two adults and four larval stages, none of which could be assigned to previously described species. Analyses based on the peculiarity of their morphological features indicate that these specimens include representatives of new species belonging to genera Pliciloricus and Rugiloricus, as well as an undescribed genus of Pliciloricidae and, possibly, an undescribed genus of Nanaloricidae. Since the specimens found during this survey are either poorly preserved or intermediate life cycle stages, a short description of their morphological traits is provided here, waiting for the formal description of these putative new species when additional material is collected.
One of the adult specimens could clearly be assigned to the family Nanaloricidae given the combination of morphological traits of this family (Supplementary Figure 2); for instance, a tripartite mouth cone with a long mouth tube and a lorica consisting of various (ca. 20) strongly sclerotized longitudinal plates. Internally, the specimen is characterized by a small pharyngeal bulb located within the basal region of the mouth cone. This specimen was unfortunately lost during manipulations.
The other adult specimen inspected during this study is assigned to Rugiloricus (Figures 1A-D; catalogue number NHMD-621046). As typically observed in all species belonging to this genus, the specimen found in the SGM possesses a short mouth cone with a ruff (Figure 1B), a neck region with single trichoscalids of identical structure and length (double trichoscalids are absent; Figures 1C, D) and a lorica with many plicae (>20; Figure 1A). Moreover, the neck has two rows of closely set basal plates (Figures 1C, D): (i) the anterior most row in which some of the plates possess a medial, posteriorly oriented spike and (ii) the posterior most row from which the trichoscalids protrude. In the introvert, the eighth row spinoscalids appear long and whip-like, while the ninth row spinsocalids are short and beak-like (Figure 1D).
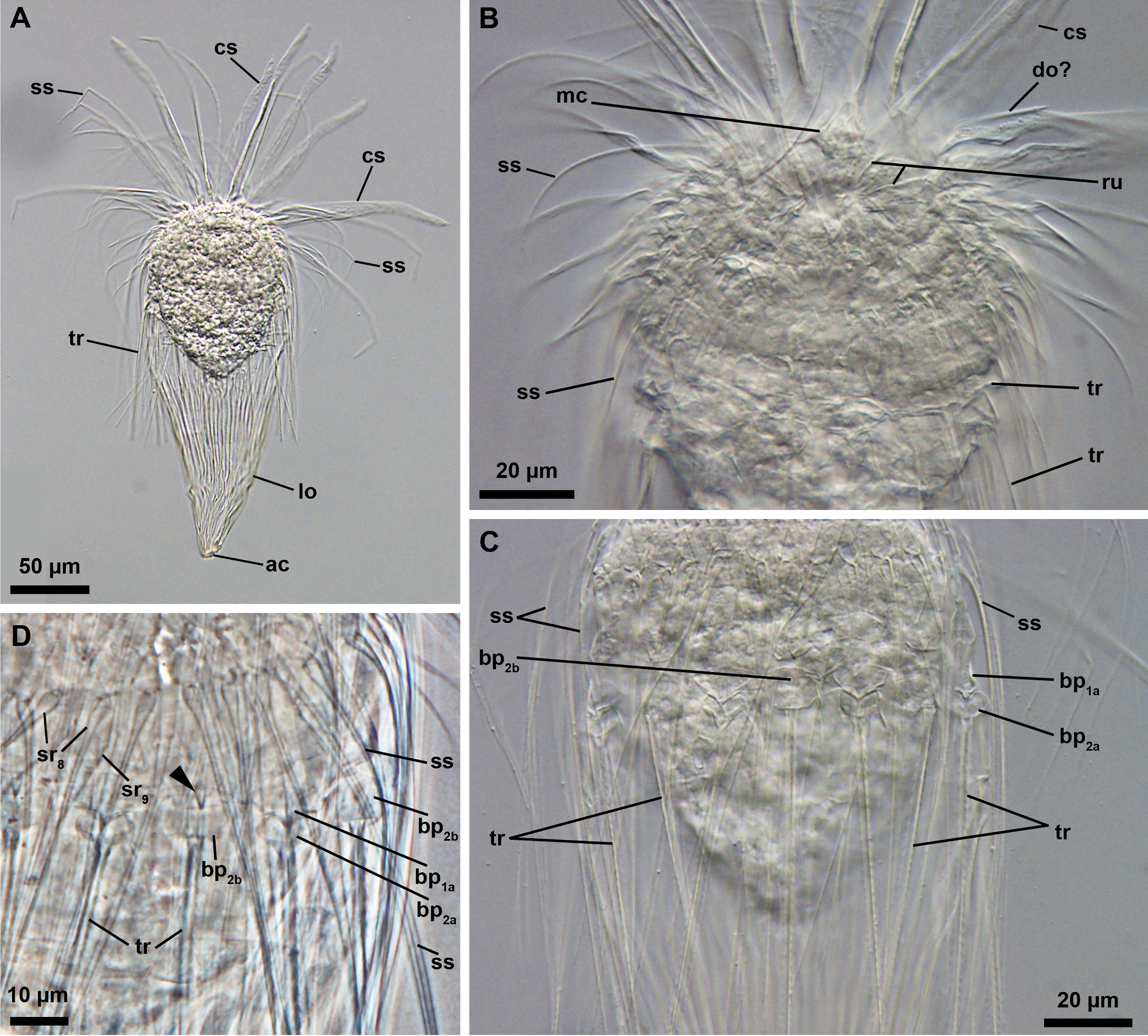
Figure 1 Light micrographs of the adult female of an undescribed species, Rugiloricus sp. 1 (NHMD-621046). (A) Overview of the specimen. Lateral view, anterior faces up. Note that the mouth cone is retracted inside the introvert. (B, C) Anterior region of the body. (D) Close-up of several basal plates of the neck region. Arrowhead points to the medial, posteriorly oriented spike of a basal plate of the anteriormost row. bp1a/b, anterior row basal plates of type a and b; bp2a/b, posterior row basal plates of type a and b; cs, clavoscalids; lo, lorica; mc, mouth cone; sr8,9, spinoscalids of 8th or 9th row; ss spinoscalids; ru, ruff; tr, trichoscalids.
Besides the two adult specimens, all the other Loriciferans found during this survey are larvae at different developmental stages. Among them, one Higgins larval stage could clearly be assigned to Pliciloricus (Figures 2A, B; catalogue number NHMD-621045). Although this larva is seemingly metamorphosing into a later larval instar or a postlarva (i.e., in an intermediate state), an unequivocal combination of characters clearly allows its generic determination. For example, possession of relatively long, thin toes and presence of three pairs of setae located in the posterior region of the body – namely the posterodorsal, posterolateral and posteroterminal pair –, is a combination of characters found only in Pliciloricus (Figure 2B). An additional Higgins larva, partially destroyed, was initially assigned to Pliciloricus based on the overall morphology of the clavoscalids and the toes (Figures 2C–E). However, a closer examination of the toes revealed the presence of a small bulge near their distal end (Figure 2E). Moreover, only two pairs of posterior setae were found – the posterodorsal and the posterolateral (i.e., the posteroterminal pair seems to be absent). Such a combination of characters was never observed before in specimens of Pliciloricus and the specimen appears, therefore, to belong to a yet undescribed genus of Loricifera.
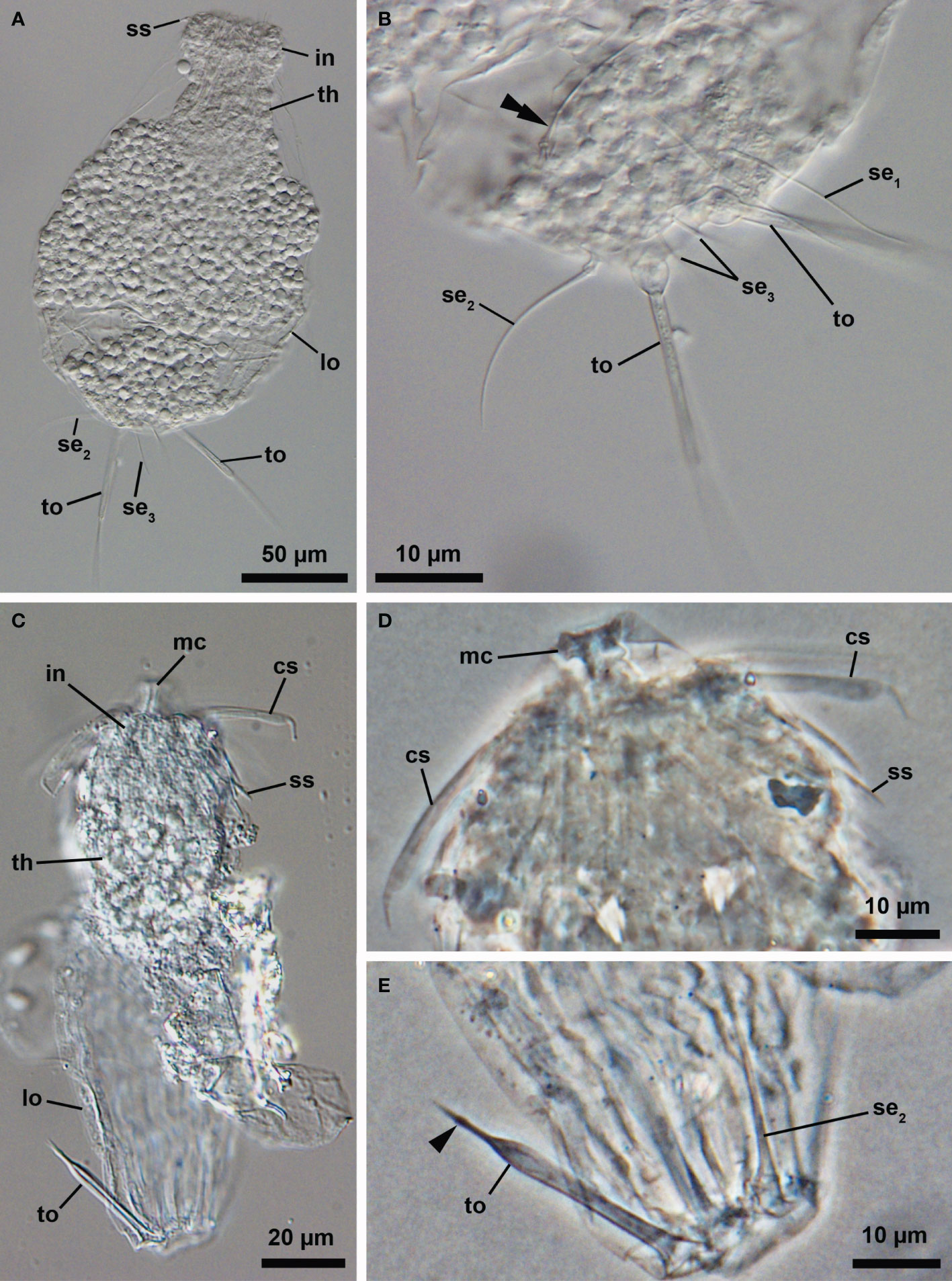
Figure 2 (A, B) Light micrographs of the molting Higgins larva of an undescribed species, Pliciloricus sp. 1 (NHMD-621045) (A) Overview. Ventrolateral view, anterior faces up. (B) Close-up of the posterior region of the body. (C–E) Light micrographs of the Higgins larva of an undescribed genus, (NHMD-621044) (C) Overview. Ventrolateral view, anterior faces up. (D) Close-up of the head. (E) Close-up of the posterior region of the body. Arrowhead points to a small bulge near the distal end of the toe. cs, clavoscalids; in, introvert; lo, lorica; mc, mouth cone; se1, posterodorsal seta; se2, posterolateral seta; se3, posteroterminal seta; ss, spinoscalid; th, thorax; to, toe.
Two of the analyzed specimens belong to an undescribed genus (or genera)? of Pliciloricidae. One is a Higgins larva characterized by three-segmented clavoscalids with a spine-like distal segment, a thoracic region not well separated from the abdominal region with many plicae present in both regions, and relatively short toes with a small bulge near their distal end (Figures 3A–D; catalogue number NHMD-621043). Two pairs of setae are present posteriorly in the larval body: one posterolateral (Figure 3D) and one posterodorsal (a posteroterminal pair was not observed). The additional specimen is seemingly a last instar Higgins larva carrying a postlarva and a young adult inside (Figures 4A–E; catalogue number NHMD-621042). This specimen possesses a combination of morphological traits similar to that described for the preceding larval specimen; however, a small bulge near the distal end of their toes was not observed (Figure 4E). In addition, the last instar larva has two pairs of setae located posteriorly of the body – one posterodorsal and one posterolateral (the posteroterminal pair was not found). A careful examination of the young adult enclosed inside revealed the presence of a ruff, i.e., a cuticular formation of the introvert that surrounds the mouth cone (Figure 4D).
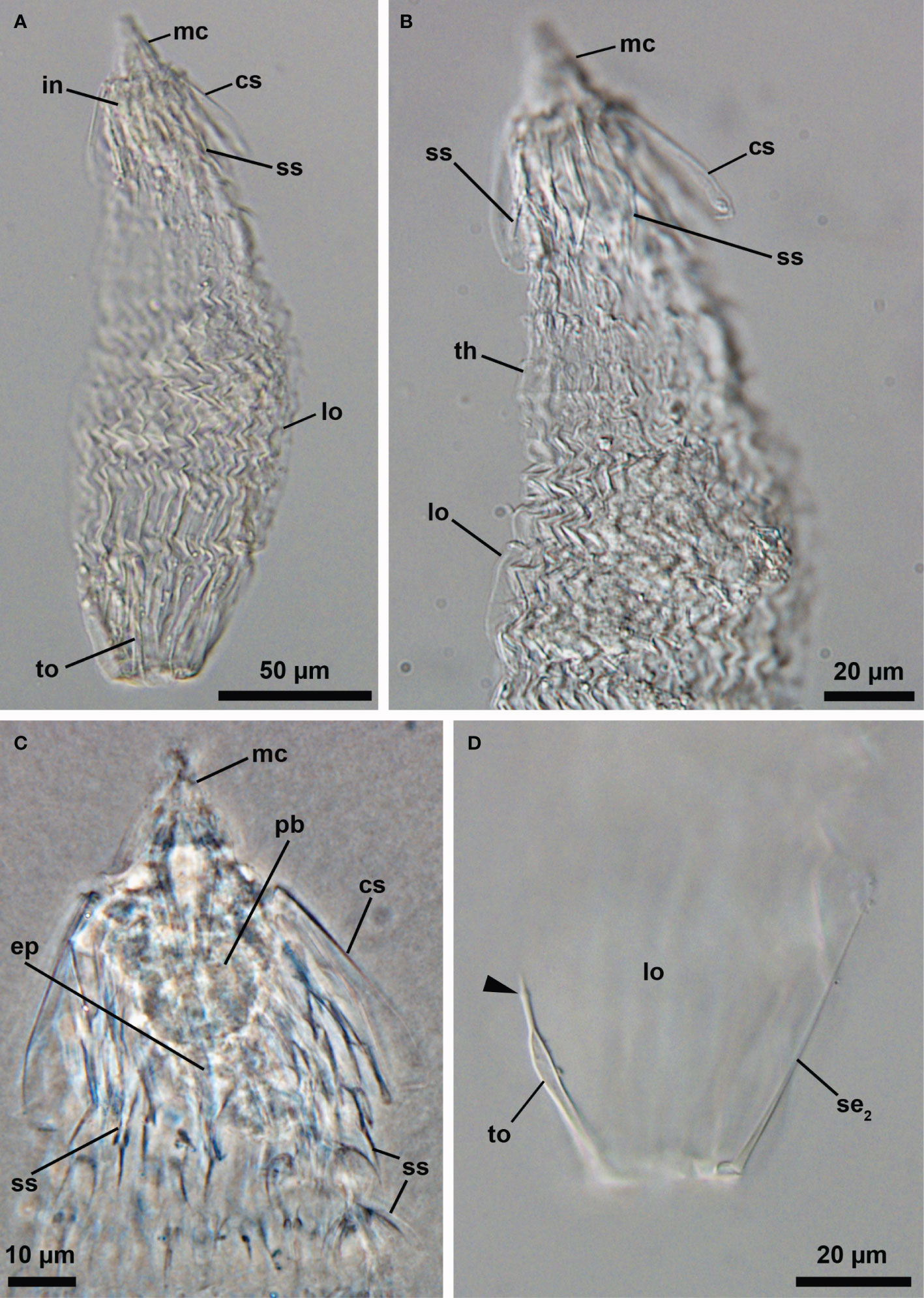
Figure 3 Light micrographs of the Higgins larva of an undescribed genus (NHMD-621043). (A) Overview. Ventral view, anterior faces up. (B) Posterior region of the body. (C) Close-up of the anterior region of the body. (D) Posterior region of the body. Note the small bulge near the distal end of the toe (arrowhead). Abbreviations: cs, clavoscalids; in, introvert; lo, lorica; mc, mouth cone; pb, pharyngeal bulb; se2, posterolateral seta; ss, spinoscalid; th, thorax; to, toe.
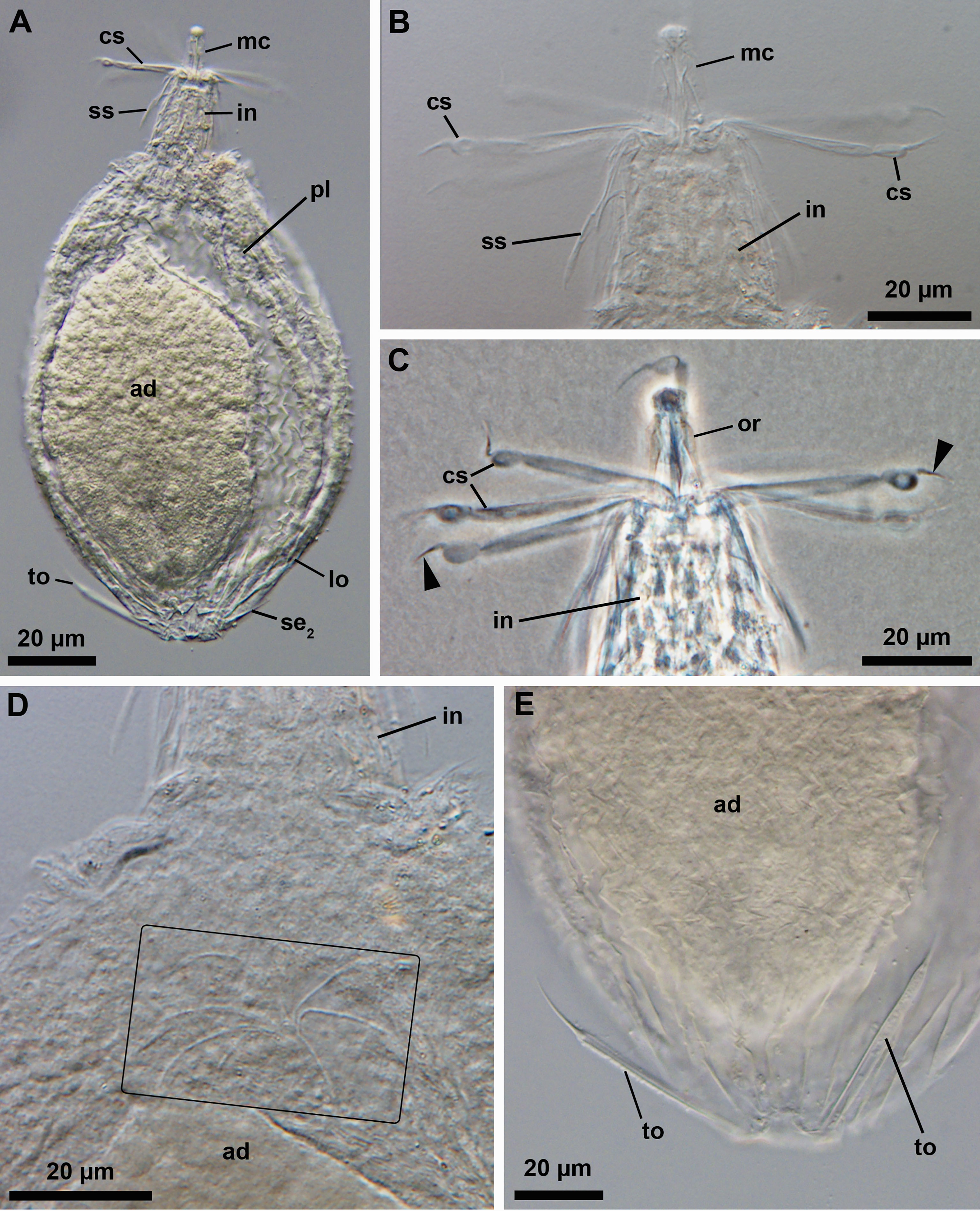
Figure 4 Light micrographs of the last instar Higgins larva (carrying a postlarva and a young adult inside) of an undescribed genus (NHMD-621042). (A) Overview. Ventral view, anterior faces up. (B, C) Anterior region of the body. The arrowhead points to the spine-like distal segment of the three-segmented clavoscalids. (D) Close-up of the abdominal region of the body. The rectangle outlines the ruff of the adult enclosed within the larval and postlarval body. (E) Close-up of the posterior region of the body. Abbreviations: ad, adult; cs, clavoscalids; in, introvert; lo, lorica; mc, mouth cone; or, oral ridge; pl, postlarva; se2, posterolateral seta; ss, spinoscalid; to, toe.
Discussion
Loricifera was not among the most abundant invertebrate groups in the samples collected during the course of XIXIMI Cruises in the SGM (see Supplementary Table 1). However, the six specimens taken from the 22 deep-sea stations sampled during this survey could represent a high richness of Loriciferan genera and species. Two of the genera represented have already been described, Pliciloricus and Rugiloricus, whereas at least two others are new to science, and each of these specimens may actually belong to species that are new to science. Moreover, two out of the three families described for Loricifera are represented, i.e., Pliciloricidae and Nanaloricidae (only Urnaloricidae is missing). Such pattern of low density and high diversity of Loricifera has been observed in several other deep-sea localities around the globe. For instance, a study on the abundance of meiofaunal communities off the coasts of California and Oregon revealed great diversity of genera and species of Loricifera (Rohal et al., 2014). After a careful inspection of a total of only 15 specimens found during this survey, two new species were described in a follow-up study – Rugiloricus californiensis and Urnaloricus ibenae – representing two out of the three families described for Loricifera, respectively – Pliciloricidae and Urnaloricidae (Neves et al., 2019). In addition, at least two putative new species were suggested – one belonging to Pliciloricus and one to Rugiloricus, and the presence of an undescribed genus was recognized.
The adult specimen assigned to family Nanaloricidae has similarities with a new genus recently described from the Clarion-Clipperton Fracture Zone in the Pacific Ocean, Fafnirloricus (Fujimoto and Murakami, 2020). Indeed, the specimen found in the SGM is characterized by a lorica with ca. 20 sclerotized plates, which is a condition found only in Fafnirloricus (all the other nanaloricids described so far have only 6-10 lorical plates). An additional character of interest in the species studied here is the presence of the pharyngeal bulb inside the mouth cone. Interestingly, such a feature was observed only once in a nanaloricid specimen collected from the Northern Gulf of Mexico (RK, pers. obs.). However, this specimen was never examined in detail and further comparisons are not possible at present. Since the nanaloricid specimen investigated here was lost during the observations under the microscope, the genus and species to which it belongs remains undetermined until more specimens are found and studied.
The adult specimen assigned to the genus Rugiloricus has some resemblances with Rugiloricus californiensis, the species recently described from the North East Pacific Region (Neves et al., 2019). For instance, both animals possess (i) neck plates of the anterior row with a medial, posteriorly oriented spike; (ii) a conical lorica; (iii) a relatively medium size end cone located at the posteriormost region of the lorica. However, the head of the specimen studied here is retracted and, therefore, the morphology of the clavoscalids and spinoscalids could not be assessed and thoroughly compared with that of R. californiensis. The similarities between the two Rugiloricus might then be limited to merely a few traits and should be treated with caution until a more comprehensive study is possible.
Among the larval specimens discovered, only one could be assigned to a described genus. Indeed, the Higgins larva metamorphosing into a late larva instar or a postlarva clearly belongs to Pliciloricus. As the mouth cone and the anteriormost region of the introvert of this specimen are fully retracted, important diagnostic characters (e.g., the internal armature and the clavoscalids) are not discernible. Therefore, it is not possible to compare it with any of the Pliciloricus described species and to determine whether it represents a new species or not.
The additional three Higgins larvae belong to an undetermined genus or more genera. Interestingly, they all possess at least one of the morphological traits that characterize a peculiar genus that has been reported primarily in an undergraduate thesis (see Gad, 2000) and briefly mentioned in a short article (see Gad, 2003 and Figure 1A therein). The undescribed genus is characterized by three-segmented clavoscalids with a spine-like distal segment, thorax and abdomen not well distinguishable from each other and composed of many plates, and toes with a small bulge near their distal end. Except for the latter, these features are also found in the Shira larva, a type of Loriciferan larval stage described solely in 2014 from a deep-sea oceanic plateau located in the Pacific Ocean (Neves and Kristensen, 2014). Moreover, specimens displaying the combination of characters that characterize the undescribed genus have also been found during the oceanographic expedition off the coast of California and Oregon and reported by Neves et al. (2019).
Finally, the young adult enclosed inside the last instar Higgins larva investigated here provides a new insight into the phylogenetic relationships of the undescribed genus it belongs to. The presence of a ruff (i.e., a cuticular formation of the introvert that surrounds the mouth cone) is a condition shared solely with the three pliciloricid genera, namely Pliciloricus, Rugiloricus and Titaniloricus (Higgins and Kristensen, 1986; Gad, 2005). Accordingly, we hypothesize that this undescribed genus – which is seemingly the genus described extensively in an undergraduate thesis (see Gad, 2000) – is phylogenetically close to Pliciloricidae, if not a member of this family. In the future, a comprehensive investigation of the distinctive morphological features that characterize the undetermined genus should provide a sufficient amount of information to help understand its evolutionary relationships.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
GR, RN, and RK contributed to conception of the taxonomic study. AR-O contributed to conception and design of the faunistic and ecological study. GR processed specimens. RN and RK performed the taxonomic description. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This research was funded by the Centro de Investigación Científica y Educación Superior de Ensenada (CICESE) and from the Consejo Nacional de Ciencia y Tecnología de México (CONACYT) - Secretaría de Energía (SE) - Hydrocarbon Trust, project 201441. This is a contribution of the Gulf of Mexico Research Consortium (CIGoM).
Acknowledgments
We are grateful to Vicente Ferreira and Ivonne Martínez Mendoza for their hard work collecting deep-sea sediments at sea, to Celia Bisbal, Samantha Guerrero, Ivette Lara, Marcela Ovalle and Viridiana Tapia for sample processing, to Martin V. Sørensen for specimens mounting, as well as to the crew of R/V Justo Sierra (UNAM) and scientific staff of cruises Xiximi-4 and Xiximi-6.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.944795/full#supplementary-material
Supplementary Figure 1 | Map showing the sampling localities in the Southern Gulf of Mexico that yielded Loriciferans. The yellow line represents countries limits, whereas the gray line outlines the Mexican Exclusive Economic Zone. Dotted lines represent transects of general sampling of XIXIMI´s cruises. Stations from cruise XIXIMI-6 are represented by black symbols: ■ = Rugiloricus sp., ▲= Undetermined genus of Pliciloricidae, ● = Undetermined genus of Nanaloricidae. The green symbol represents a station from the cruise XIXIMI-4: ♦ = Pliciloricus sp.
Supplementary Figure 2 | Light micrographs of the adult (of uncertain genus) of an undescribed species of Nanaloricidae. (A) Overview of the specimen. Dorsal view, anterior faces up. (B) Anterior region of the body. Note the location of the pharyngeal bulb inside the mouth cone.
References
Bang-Berthelsen I. H., Schmidt-Rhaesa A., Kristensen R. M. (2013). “6. Loricifera,” in Handbook of Zoology. Gastrotricha, Cycloneuralia And Gnathifera. vol. 1: Nematomorpha, Priapulida, Kinorhyncha, Loricifera. Ed. Schmidt-Rhaesa A. (Berlin: De Gruyter), 307–328.
de Jonge V. N., Bouwman L. A. (1977). A simple density separation technique for quantitative isolation of meiobenthos using the colloidal silica ludox-TM. Mar. Biol. 42, 143–148. doi: 10.1007/BF00391564
Fujimoto S., Kristensen R. M. (2020). “Loricifera,” in Guide to the identification of marine meiofauna. Ed. Schmidt-Rhaesa A. (München: Verlag Dr. Friedrich Pfeil), 415–427.
Fujimoto S., Murakami C. (2020). A new genus and species of Nanaloricidae (Loricifera: Nanaloricida) From the Clarion-Clipperton fracture zone. Zool. Anz. 289, 177–188. doi: 10.1016/j.jcz.2020.10.009
Gad G. (2000). Studies of Systematic, Morphology and Life Cycles of Deep-sea Loricifera (Oldenburg: Carl von Ossietzky Universität).
Gad G. (2005). Giant Higgins-larvae with paedogenetic reproduction from the deep sea of the Angola basin–evidence for a new life cycle and for abyssal gigantism in Loricifera? Org. Divers. Evol. 5, 44–59. doi: 10.1016/j.ode.2004.10.005
Heiner I., Kristensen R. M. (2009). Urnaloricus gadi nov. gen. et nov. sp. (Loricifera, Urnaloricidae nov. fam.), an aberrant Loricifera with a viviparous pedogenetic life cycle. J. Morphol. 270, 129–153. doi: 10.1002/jmor.10671
Higgins R. P., Kristensen R. M. (1986). New Loricifera from southeastern United States coastal waters. Smithson. Contrib. Zool. 438, 1–70. doi: 10.5479/si.00810282.438
Hubbard G. F., Howard R. L., Gallaway B. J. (1988). Loricifera, a recently described phylum, occurring in the northern gulf of Mexico. North. Gulf Sc. 10 (1), 49–50. doi: 10.18785/negs.1001.05
Kristensen R. M. (1983). Loricifera, a new phylum with Aschelminthes characters from the meiobenthos. Z. Zool. Syst. Evolut. -forsch. 21, 163–180.
Kristensen R. M. (1991a). “Loricifera,” in Microscopic anatomy of invertebrates, vol. 4, Aschelminthes. Eds. Harrison F. W., Ruppert E. E. (New York: Wiley-Liss), 351–375. doi: 10.1111/j.1439-0469.1983.tb00285.x
Kristensen R. M. (1991b). Loricifera - a general biological and phylogenetic overview. Verh. Dtsch. Zool. Ges. 84, 231–246.
Kristensen R. M., Brooke S. (2002). “Phylum Loricifera,” in Atlas of marine invertebrate larvae. Eds. Young C. M., Sewell M. A., Rice M. E. (London: Academic Press), 179–187.
Kujawinski E. B., Reddy C. M., Rodgers R. P., Cameron Thrash J., Valentine D. L., White H. K. (2020). The first decade of scientific insights from the deepwater horizon oil release. Nat. Rev. Earth Environ. 1, 237–250. doi: 10.1038/s43017-020-0046-x
Neves R. C., Kristensen R. M. (2014). A new type of Loriciferan larva (Shira larva) from the deep sea of shatsky rise, pacific ocean. Org. Divers. Evol. 14, 163–117. doi: 10.1007/s13127-013-0160-4
Neves R. C., Kristensen R. M., Møbjerg N. (2021). New records on the rich Loriciferan fauna of Trezen ar Skoden (Roscoff, France): Description of two new species of Nanaloricus and the new genus Scutiloricus. PLoS One 16 (5), e0250403. doi: 10.1371/journal.pone.0250403
Neves R. C., Kristensen R. M., Rohal M., Thistle D., Sørensen M. V. (2019). First report of Loricifera from the north East pacific region, with the description of two new species. Mar. Biodivers. 49, 1151–1168. doi: 10.1007/s12526-018-0898-1
Neves R. C., Reichert H., Sørensen M. V., Kristensen R. M. (2016). Systematics of phylum Loricifera: identification keys of families, genera and species. Zool. Anz. 265, 141–170. doi: 10.1016/j.jcz.2016.06.002
Peters E. M., Zaragoza-Álvarez R. A., Herzka S. Z., Herguera-García J. C. (Eds.) (2021). Atlas de línea base ambiental del golfo de México, tomo introducción (Ensenada: Consorcio de Investigación del Golfo de México).
Rohal M., Thistle D., Easton E. E. (2014). Meiofaunal abundances and faunal similarity on the continental rise off the coast of California. Deep-Sea Res. I 93, 131–144. doi: 10.1016/j.dsr.2014.07.004
Sørensen M. V., Grzelak K., Kristensena R. M., Herranz M. (2022). First account on Loricifera from New Zealand: A new species of Pliciloricus, and a shira larva with postlarva representing the new genus and species Patuloricus tangaroa gen. et sp. nov. Zool. Anz. 299, 207–220. doi: 10.1016/j.jcz.2022.06.004
Keywords: meiofauna, CIGOM, Ecdysozoa, Nanaloricus, Rugiloricus, Pliciloricus
Citation: Neves RC, Kristensen RM, Rocha-Olivares A and Rivas G (2022) A first look at the biodiversity of Loricifera in the southern Gulf of Mexico. Front. Mar. Sci. 9:944795. doi: 10.3389/fmars.2022.944795
Received: 15 May 2022; Accepted: 30 June 2022;
Published: 29 July 2022.
Edited by:
Clara F. Rodrigues, University of Aveiro, PortugalReviewed by:
Gonzalo Giribet, Harvard University, United StatesElisa Baldrighi, Institute for Biological Resources and Marine Biotechnology (CNR), Italy
Copyright © 2022 Neves, Kristensen, Rocha-Olivares and Rivas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gerardo Rivas, Z2VyYXJkb3JpdmFzQGNpZW5jaWFzLnVuYW0ubXg=