 Bingzhong Yang1,2*
Bingzhong Yang1,2* Bent Herrmann3,4,5
Bent Herrmann3,4,5- 1South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou, China
- 2Key Laboratory of Open-Sea Fishery Development, Ministry of Agriculture and Rural Affairs, Guangzhou, China
- 3SINTEF Ocean, Fishing Gear Technology, Hirtshals, Denmark
- 4UiT, The Arctic University of Norway, Tromsø, Norway
- 5DTU Aqua, Technical University of Denmark, Hirtshals, Denmark
Size selectivity and catch efficiency of four codends were tested and compared for mantis shrimp (Oratosquilla oratoria) in demersal trawl fisheries of the South China Sea (SCS). These codends were differing in mesh shapes, diamond mesh (T0) and diamond-mesh turned 90° (T90) with mesh sizes of 30 and 35 mm, respectively. The results demonstrated that the T0 codend with a lower mesh size, 30 mm, presented poorer selective properties for the target species, while size selectivity would be significantly improved with the mesh size increasing to 35 mm, or substituting the T0 codend with the T90 codend. For the T90 codend with a larger mesh size, 35 mm, the size selectivity was the highest, whereas the loss of the legal individuals was also significantly considerable. Considering the trade-off between releasing undersized individuals and maintaining the legal ones, the T0 codend with 35-mm mesh size or the T90 codend with 30-mm mesh size might be a better choice to target mantis shrimp in demersal trawl fisheries of the SCS.
Introduction
China is the largest contributor to marine capture fisheries all around the world. However, the biomass of traditional commercially important fish species, such as small yellow croaker (Larimichthys polyactis), Spanish mackerel (Scomberomorus niphonius), and hairtail (Trichiurus spp.), has dramatically declined (Jin and Tang, 1998; Kang et al., 2018). In contrast, crustaceans have already been the major target species in marine fisheries due to low trophic levels and highly inherent productivity. It has been shown that crustacean fisheries are rising and playing an ever-increasing role in seafood security globally (Boenish et al., 2021). The situation is the same in China, and one of its most socio-economically important crustacean species is mantis shrimp (Oratosquilla oratoria). Widely distributed in all coastal waters and with relatively abundant resources, mantis shrimp has been a major target species for many marine fisheries in China, especially for demersal trawls (Wang and Xu, 1996; Liu et al., 2020). Moreover, mantis shrimp is so capable of staying alive, even when it is captured from the sea, that it greatly satisfies the consumers’ demand for luxury seafood that is actively moving when it is purchased.
Although with merits mentioned above, annual landing statistics show that the resources of mantis shrimp in China may have been overexploited. For instance, the national landing of mantis shrimp was 315,400 tonnes in 2010; this landing dropped to 206,000 tonnes in 2020 (MOA, 2011; MARA, 2021). A decade passed, and the loss in the landing was 109,400 tonnes. These statistics demonstrate that the resources of mantis shrimp have been suffering huge fishing pressure, and its fisheries need to be strictly regulated. Some studies indicated that bycatch and discard issues are attributed to the decline of stock for mantis shrimp (Ishii and Kitahara, 2002; Yang et al., 2017).
Like many shrimp species, mantis shrimp are benthic organisms and intensively fished by demersal trawls. Due to poor selectivity, especially by the codend in which most selection takes place (Glass, 2000), demersal trawling often induces serious bycatch and discard problems (Cashion et al., 2018). To address these issues, numerous gear modifications have been designed, tested, and evaluated. The simplest option to improve size selectivity is just increasing the mesh size used in the diamond-mesh codend (Fryer et al., 2016; O’Neill et al., 2020). Due to the inherent characteristics of diamond-mesh codends (Herrmann, 2005a; Herrmann, 2005b), only increasing mesh sizes may not obtain the intended improvement in selectivity. To further improve size selectivity, another simple and effective gear modification is just turning the direction of the diamond-mesh netting by 90° (termed as T90 codend). Compared with the traditional diamond-mesh codend (hereafter referred to as T0 codend), T90 codends usually exhibit more mesh opening and result in better selective properties. As a result, applying T90 codends could improve the size selectivity of trawl fisheries for many fishing species. This has been widely demonstrated by using computer simulation (Herrmann et al., 2007), flume tank test (Madsen et al., 2015; Cheng et al., 2022), and numerous sea trial experiments (Madsen et al., 2012; Bayse et al., 2016; Cheng et al., 2020; Kennelly and Broadhurst, 2021; Brinkhof et al., 2022).
As its economic and ecologic relevance, reproductive biology, growth, and age estimation of mantis shrimp have been widely investigated (Kodama et al., 2004; Kodama et al., 2005; Kodama et al., 2006; Kodama et al., 2009; Nakajima et al., 2010; Kim et al., 2017). Some of these studies suggested that the size selectivity of the trawl should be improved to recover the stock abundance of mantis shrimp (Kodama et al., 2006; Nakajima et al., 2010). However, few studies have been published to address the size selectivity of trawl for this important target specie (Tokai et al., 1990; Tokai et al., 1996; Wang et al., 2017; Petetta et al., 2020). Specifically, no selective experiment has been carried out to investigate the size selectivity of trawl codend for mantis shrimp in the South China Sea (SCS). In China, all fishing fleets targeting mantis shrimp and/or other shrimp species are required to follow a minimum mesh size (MMS) regulation of 25 mm in the codends. This MMS regulation is simple, but its effectiveness is questionable. Recently, Yang et al. (2021) conducted a selectivity experiment using six diamond-mesh codends, with mesh sizes ranging from 25 to 54 mm, in the SCS. The target species was Southern velvet shrimp (Metapenaeopsis palmensis), whose body size is often smaller than that of mantis shrimp. Their results demonstrated that the legal codend with a 25-mm mesh size was not proper for protecting juvenile shrimp and that a mesh size increased to 35 mm selectivity would be better (Yang et al., 2021). Will increasing the mesh sizes of diamond-mesh codend improve size selectivity for mantis shrimp in the SCS and substituting the T0 codend with T90 result in a positive outcome? These research questions need to be strictly addressed.
In addition, there is no legal minimum landing size (MLS) formulated to supplement the MMS regulation to protect juvenile individuals of mantis shrimp in the SCS. Some studies from aquaculture showed that the minimum length at maturity was 8.0 cm for mantis shrimp (Xu et al., 1996; Yang et al., 2004). Another study based on sea-trial surveys suggested that the minimum conservation reference size (MCRS) of mantis shrimp should be 9.5 cm in the Yellow Sea of China (Liu et al., 2020). The legal formulation of MLS for mantis shrimp should also take the selective properties of trawl codends into account.
In order to address the issues mentioned above, we tested and compared the size selectivity and catch efficiency of four codends, T0 and T90 with mesh sizes of 30 and 35 mm, respectively, and focused on the following research questions:
1) How did the size selectivity and catch efficiency change when the mesh sizes of the T0 codend increased?
2) How did the size selectivity and catch efficiency change when the T0 codend was substituted with the T90 codend?
(3) Are the potential changes length-dependent or not?
Materials and methods
Sea trials
Sea trials were conducted onboard a commercial demersal trawler, named Guibeiyu 96899 (38 m, 280 kW) in November 2020. The fishing grounds were located in the Beibu Gulf of the SCS, with a latitudinal and longitudinal range of N20°53′–21°10′ and E 108°33′–109°09′. The water depth in the fishing grounds varied between 18 and 39 m. All fishing procedures were identical to the ones of commercial fishing. Prior to the experimental fishing in 2020, there was a similar codend selectivity experiment, in which mantis shrimp served as a bycatch species, carried out by the same vessel on the same grounds (for detailed information, please refer to Yang et al., 2021). The experimental data of this earlier sea trial can be used to supplement the present study by generating another fishing population scenario of mantis shrimp.
Experimental setup
In order to focus on our research questions, we applied the gear components of the commercial trawler except for the codends, in which modifications were made. The selected trawler provided us with an ideal platform for selectivity experiments since it operated a double-rigged trawl system, in which two identical trawls were hauled simultaneously (Figure 1). These trawls had a fishing circumference of 860 meshes, in which the mesh size was 45 mm, and the stretched length was about 33 m in total. They were spread by two identical sets of otter board (250 kg and 1.6 m2), and the headline height was mainly 1.5 m, and the wing-end was spaced about 15 m during the commercial fishing.
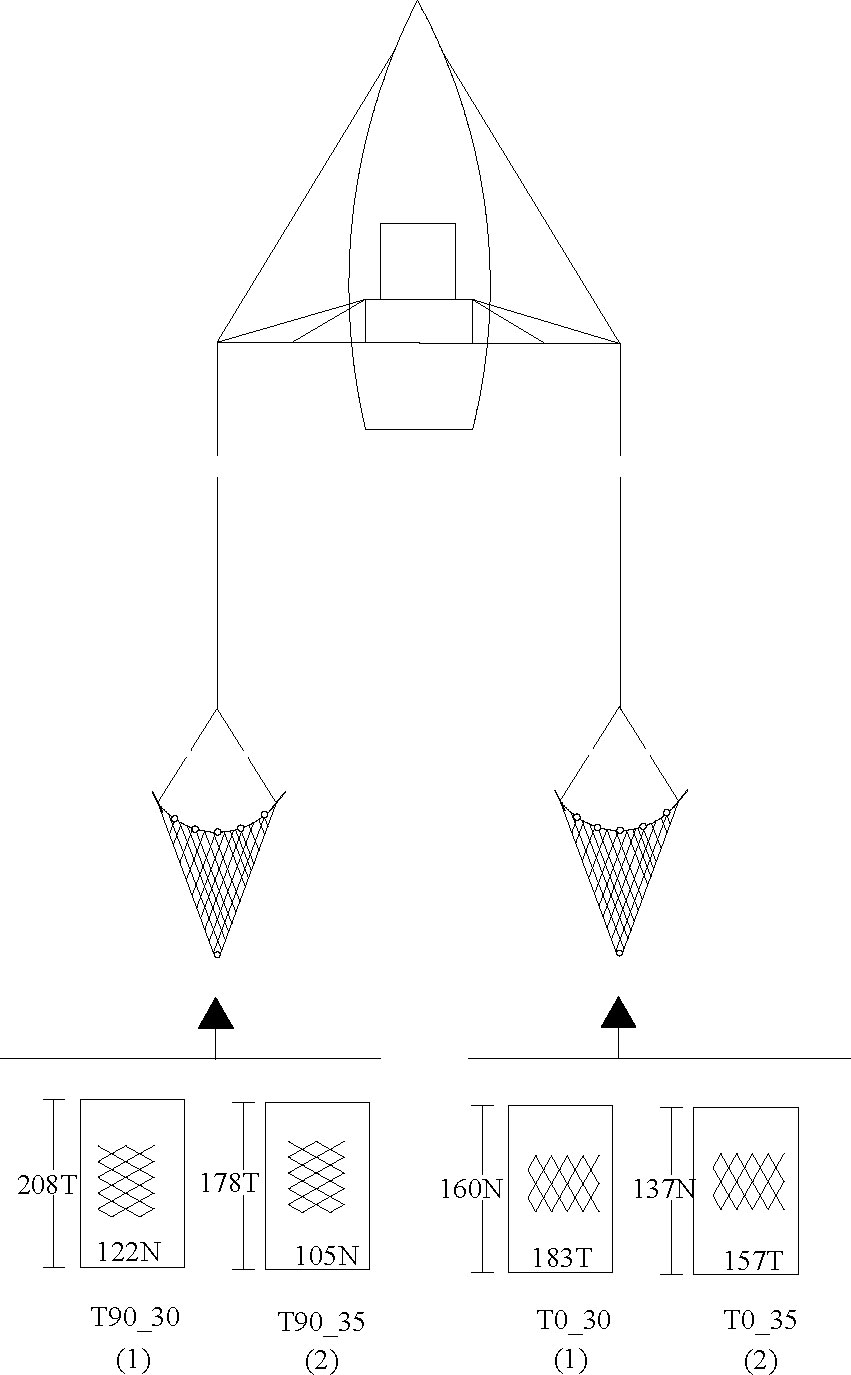
Figure 1 Schematic view of the experimental codends tested. (1) represents the first comparison test, and (2) represents the second comparison test. T0_30 and T0_35 represent the diamond-mesh codend with mesh sizes of 30 and 35 mm, whereas T90_30 and T90_35 represent the turned mesh codend with mesh sizes of 30 and 35 mm, respectively.
With the use of the dimension of the commercial codend, which is made of diamond mesh with a mesh size of 25 mm and 220 and 192 meshes in the circumference and vertical direction, respectively, four codends were designed and tested. The modifications in these codends are focused on mesh sizes and mesh shapes. Two mesh sizes, 30 and 35 mm, and two mesh shapes, T0 and T90, were applied. We identified the codends according to their mesh shape and mesh size, as follows: T0_30, T0_35, T90_30, and T90_35. For instance, T0_30 represents the codend with a diamond mesh, and its mesh size was 30 mm, while T90_30 represents the T90 codend with a 30-mm mesh size. Differences between the codends were mainly the mesh sizes, the mesh numbers in circumference, and the length direction reduced as mesh size increased to keep their stretched circumference and length constant. The T90 codends were manufactured using the same netting of the T0 codends with the same mesh size, but the netting direction turned 90°, and their mesh numbers in circumference and length direction were generated based on some previous related studies (Bayse et al., 2016; Robert et al., 2020). In our practice, the mesh number of the T90 codend in circumference was about 33% less and 30% more in the vertical direction compared with that of the T0 codend with the same mesh size (Table 1).

Table 1 Overview of specification of the experimental codends and cover.
To collect mantis shrimp escaping from the tested codends, the covered codend method was applied. Following the recommendation of Wileman et al. (1996), the cover net was 1.5 larger and longer than the tested codends, and its mesh opening was about 12 mm (Table 1). In order to remove the potential covered effect, a total of 12 flexible kites were mounted to the covers (He, 2007; Grimaldo et al., 2009; Yang et al., 2021). Additionally, underwater video recordings systems, composed of GoPro HERO4 and a framework, were applied to check how these kites work during the experiments. If a covered effect existed, adjustments were made, and the data of the specific haul were excluded. To facilitate access to the codend catch and avoid the potential “wash-out” effect, a zipper with a length of 1.1 m was mounted to the cover net.
As the double-rigged trawl systems were applied, we were able to test two codends at a time. Codends with the same mesh sizes but different mesh shapes were arranged as pairwise tests: T0_30 vs. T90_30 and T0_35 vs. T90_35 (Figure 1). Once the haul-back process finished, catch in the tested codends were firstly handled through the zippers. The catch of mantis shrimp was sorted and sub-sampled if the number of individuals was large. All mantis shrimp samples were kept in marked containers, frozen, and taken back to the laboratory for biological measurement.
Estimation of size selectivity
Catch data of the counted number of mantis shrimp in length were applied to estimate the size selectivity for the tested codends. For a given matins shrimp with an l length in a specific j haul, the fishing data were binominal, as they were either caught by the codend or covered, and the catch probability (proportion) can be expressed as rj(l). The actual value of rj(l) can be easily calculated with the number of mantis shrimp caught by the tested codend and the total number; however, this value has been proved to be varied among different hauls for the same codend (Fryer, 1991). The variations are often due to uncertainties in both within- and between-haul. To account for these uncertainties, we estimated the average retention probability of the tested codends for mantis shrimp with a specific length by pooling all catch data and used it to represent the size selectivity of codends for mantis shrimp in commercial fishing. This average retention probability can be further described as rav(l), in which v is a vector composed of selectivity parameters in some models. In order to estimate rav(l), we minimized the following expression:
in which the outer summation is over the m hauls carried out and the inner summation is over length class l; nRjl and nEjl represent the catch number of mantis shrimp by the tested codend and cover, respectively, while qRj and qEj are the sub-sampled ratios from the tested codends and cover, respectively. Minimizing expression (1) is equivalent to maximizing the likelihood for the observed experimental data combined over hauls based on a binominal distribution.
To describe rav(l), four commonly used models, Logit, Probit, Gompertz, and Richards, were used, as candidates. Two selective parameters, L50 (50% retention length) and SR (selection range = L75–L25), can fully represent the first three candidate models; for the last one, another parameter, 1/δ, should be added (Wileman et al., 1996).
The estimation of size selectivity was conducted using a two-step procedure. First of all, the four candidate models mentioned above were initially fitted to Equation (1) to generate their Akaike information criterion (AIC) values (Akaike, 1974), and the model with the lowest AIC value was considered the best-fitting model. Second, using these best models, a double-bootstrapping technique was applied to calculate the Efron percentile (Efron, 1982) 95% confidence interval (CI) for the selective parameters and selectivity curves (Millar, 1993; Herrmann et al., 2012; Herrmann et al., 2018; Yang et al., 2021). Specifically, this bootstrapping method accounts for uncertainty due to between-haul variation by selecting m hauls with replacement from the m hauls available during each bootstrap repetition (Equation 1). Within each resampled haul, the data for each length class were resampled in an inner bootstrap to account for the uncertainty in the estimation of the size selection in the haul resulting from only a limited number of mantis shrimp being caught, and length was measured in the specific haul. The inner resampling of the data in each length class was performed prior to the raising of the data with subsampling factors qRj and qEj to account for the additional uncertainty due to the subsampling (Eigaard et al., 2012). After the two-step procedure, we can evaluate how the selected models fit the experimental data through their p-values. Normally, when these models were sufficiently able to describe the fishing data, their p-values should be larger than 0.05 (Wileman et al., 1996). If they obtained p-values<0.05, the residuals should be checked to determine whether this was due to structural problems of the selected models or simply due to overdispersion in the fishing data (Wileman et al., 1996).
Our experimental designs enable us to compare the size selectivity of codends with different mesh sizes and mesh shapes for mantis shrimp. To quantify the differences in the size selectivity between different codends, delta selectivity, Δr(l), was calculated with the following expression:
where ra (l) is the size selectivity of codend a, while rb(l) represents the size selectivity of codend b. The Efron percentile 95% CIs for delta selectivity can be calculated with the double-bootstrapping approach mentioned above. These CIs of delta selectivity can enable us to determine a significant difference if they do not overlap 0.0 in some length classes.
Exploitation pattern indicators
In addition to the size selectivity, it is also fundamental to estimate how the mesh sizes and mesh shapes used in codends affect the exploitation pattern of demersal trawl fishing for mantis shrimp in different population scenarios. The exploitation pattern can be reflected with four indicators: nP−, nP+, nRatio, and dnRatio. To estimate these indicators, we first generated two different fishing population scenarios by pooling all catch data (both cover and codend) in two selectivity experiments, and then we combined the size selectivity result from the previous section as the following expression:
where rcodend(l) is the size selectivity of the codend tested, while nPopl represents the fishing population scenarios of mantis shrimp. Again, the Efron percentile 95% CIs for the exploitation pattern indicators can also be calculated with the double-bootstrapping approach mentioned above. For a tested codend to have ideal selective properties, the lower the nP−, the better; the higher the nP+, the better; and the lower the nRatio and dnRatio, the better. To calculate the exploitation pattern indicators, we applied the minimum length at maturity of mantis shrimp, 8.0 cm (Yang et al., 2004), as the MCRS.
All the data analyses were carried out using the selectivity software SELNET (Herrmann et al., 2012; Herrmann et al., 2018; Herrmann et al., 2019; Yang et al., 2021). The double-bootstrapping technique was implemented in this software. Information on how to obtain a copy of SELNET and instructions on how to use it can be obtained from the second author of this study. Additionally, a short description of SELNET is provided in the supplementary material.
Results
Experimental data
In total, 34 valid hauls were conducted; seven hauls for the T0_30 codend and nine hauls for the T0_35, T90_30, and T90_35 codends. During these hauls, the average duration was about 132 min, with a range of 115 to 158 min, and the average water depth was about 25 m, ranging from 18 to 39 m, in the fishing grounds (Table 2). Mantis shrimp was present in all these hauls and represented as one of the most dominant species in terms of quantity, and the sub-sampling ratios of mantis shrimp ranged from 0.25 to 1.00 (Table 2). Bycatch species included white croaker (Pennahia argentata), finespot goby (Chaeturichthys stigmatias), burrowing goby (Trypauchen vagina), and southern velvet shrimp (M. palmensis). A total of 1,364 individuals of mantis shrimp were collected, and their length was measured; 866 individuals were caught by the tested codends and 498 by their relative covers. By pooling all data from both codends and covers, we obtained two average populations of mantis shrimp, in terms of the relative frequency in length, based on the experimental data collected in 2019 and 2020 (Figure 2). Different length distribution was presented, as the length distribution ranged from 6 to 23 cm, with a mode at the range of 11.5 to 12.0 cm, and a very small fraction of mantis shrimp with length less than the MCRS was caught in the data of 2019, while the length range was 3 to 23 cm, with a mode at the length of 6 cm, and a larger fraction of mantis shrimp smaller than the MCRS was present in 2020 (Figure 2).
Table 2 Basic information on the experimental fishing and catch.
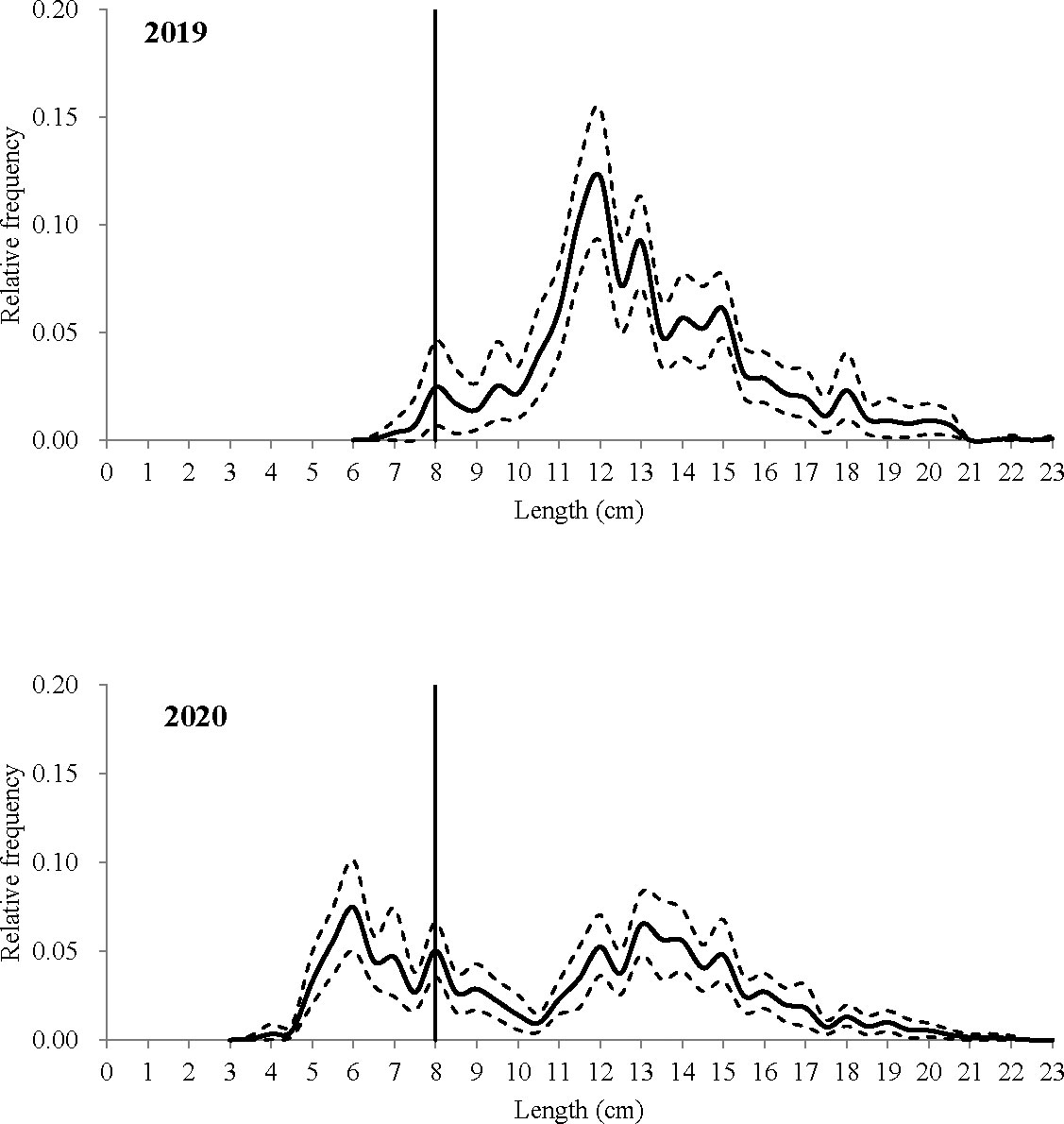
Figure 2 Estimated average population of mantis shrimp from all hauls during the sea trials in 2019 and 2020. Stipple lines show the 95% Efron confidence intervals, and the vertical line represents the MCRS (minimum conservation reference size) of mantis shrimp derived from the minimum length at maturity in the South China Sea (SCS).
Size selectivity
By comparing the AIC values from four candidate models, the Gompertz model was selected as the best model for all tested codends (Table 3). According to the p-values, the selected model was able to represent the tested codends sufficiently well, except the T90_30 codend for which a p-value<0.05 was obtained (Table 4). After checking the length-dependent residuals, we concluded that this low p-value could be due to overdispersion in the experimental data since no systematic patterns were indicated in the residuals. This overdispersion was probably due to the amount of subsampling in the data collection process (Larsen et al., 2018).
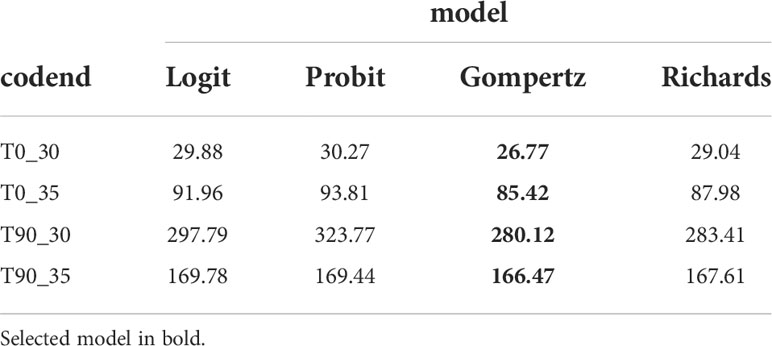
Table 3 Akaike’s information criterion (AIC) values for each model of the tested codends.
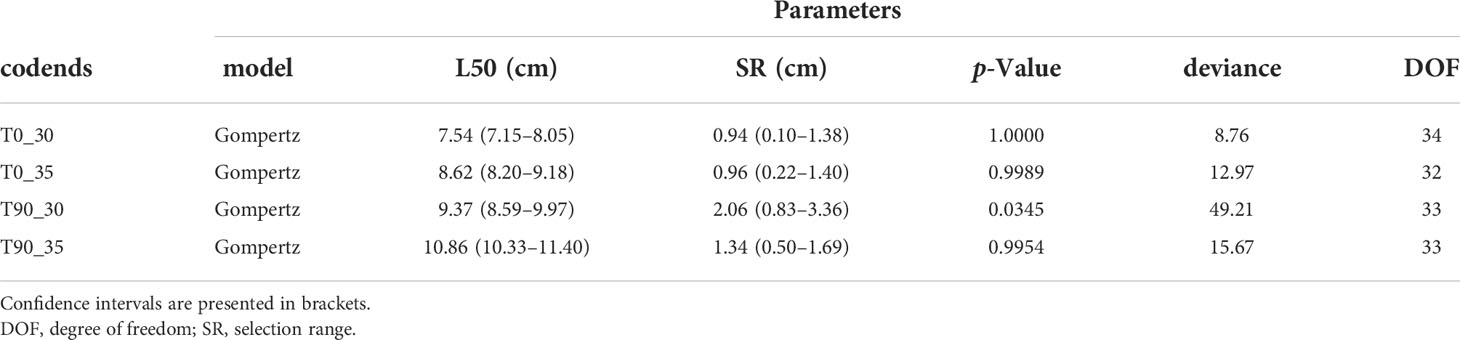
Table 4 Selective parameters and fit statistics obtained from the selected models for the tested codends.
The selective parameters showed that the values of L50 would significantly increase when the mesh size increased in the codends with the same mesh shape, while for the codends with the same mesh sizes, the T90 codends had larger values in L50. For instance, L50 of T0_35 was 8.62 (CI: 8.20–9.18) cm, which was significantly larger than that of the T0_30 codend, 7.54 (CI: 7.15–8.05) cm. A similar result was obtained for the T90_35 codend compared with the T90_30 codend. Compared with the T0_30 codend, the L50 of the T90_30 codend was significantly larger, and the same for the T90_35 vs. T0_35 comparison. There were some differences in values of SR among the tested codends, but they were not statistically significant.
The retention probability of a mantis shrimp with a length of MCRS would be reduced by modifying the mesh sizes and shape of the codends used. For instance, the retention probability for an individual with the MCRS length was 72.62% (CI: 47.66%–100.00%) for the T0_30 codend, the probability would drop to 14.81% (CI: 0.60%–27.96%) and 13.83 (0.00%–33.88%) when the mesh size increased to 35 mm (T0_35) and applying the T90 mesh shape (T90_30), and if both mesh size increment and T90 shape are applied, T90_35, the retention probability would be 0 (Figure 3).
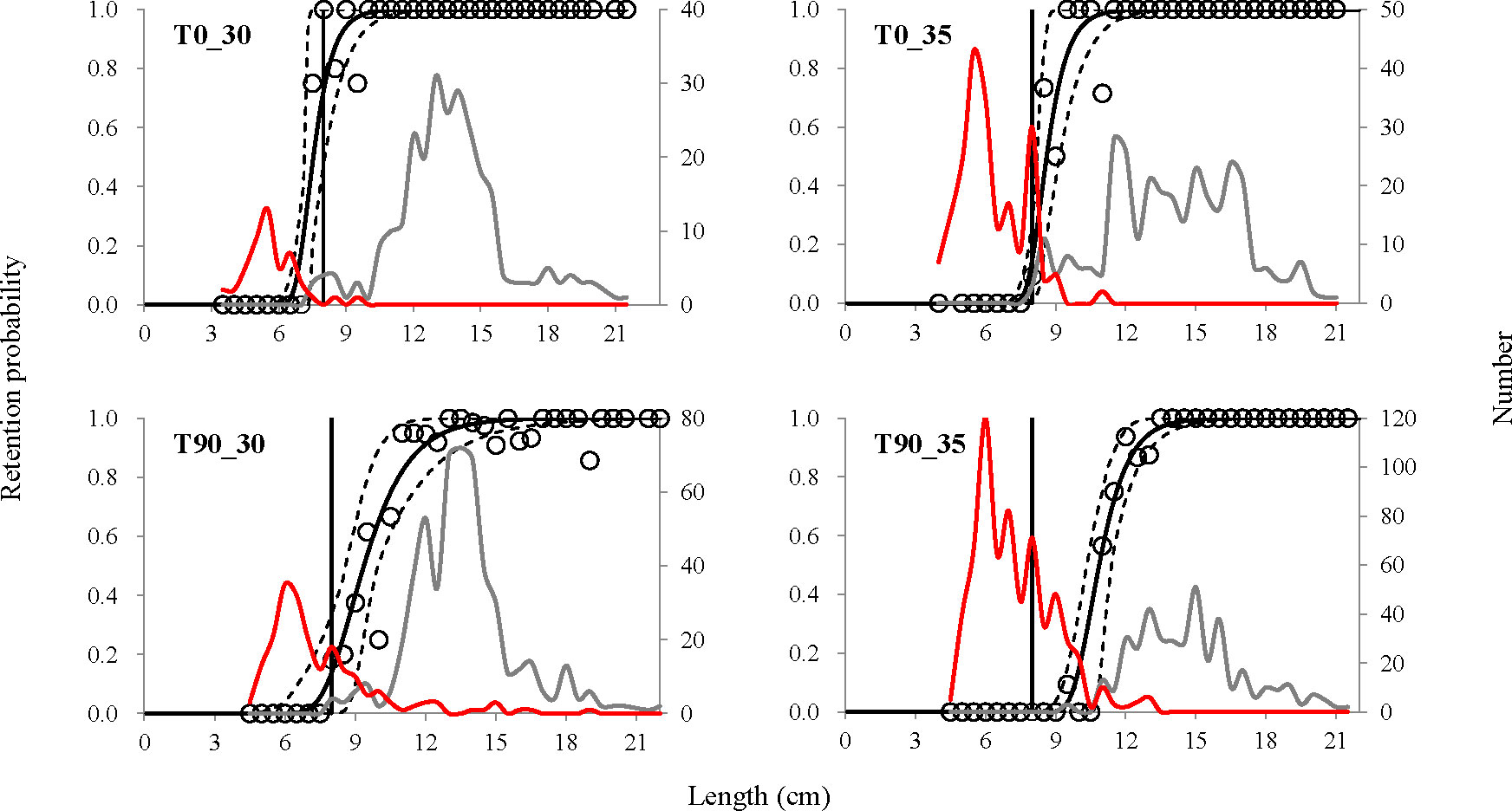
Figure 3 Experimental catch proportion and selectivity curves obtained for the T0 and T90 codends tested. Circle marks represent experimental catch proportion. Red curves represent the size distribution of fish caught by the cover, and gray curves represent the ones caught by the tested codend. Solid black curves represent selectivity curves, and stippled curves describe the 95% confidence intervals. Vertical lines represent the MCRS (minimum conservation reference size) of mantis shrimp derived from the minimum length at maturity in the South China Sea (SCS).
Delta selectivity
With the use of the traditional diamond-mesh codend with a 30-mm mesh size as a starting point, the differences in size selectivity between codends tested could be further demonstrated by the delta selectivity in terms of length-dependent retention probability. Compared with the baseline codend (T0_30), all codends had significantly lower retention probability for mantis shrimp at the following length range: 7.4–8.4 cm for the T0_35 codend, 7.5–15.6 cm for the T90_30 codend, and 7.3–15.3 cm for the T90_35 codend (Figure 4). The T90_35 codend would significantly have less retention probability for mantis shrimp at the length range of 7.9–13.5 and 7.7–11.2 cm when compared with the T0_35 and T90_30 codends, respectively (Figure 4). Only no significant difference was obtained for the comparison between the T90_30 and T0_35 codends (Figure 4).
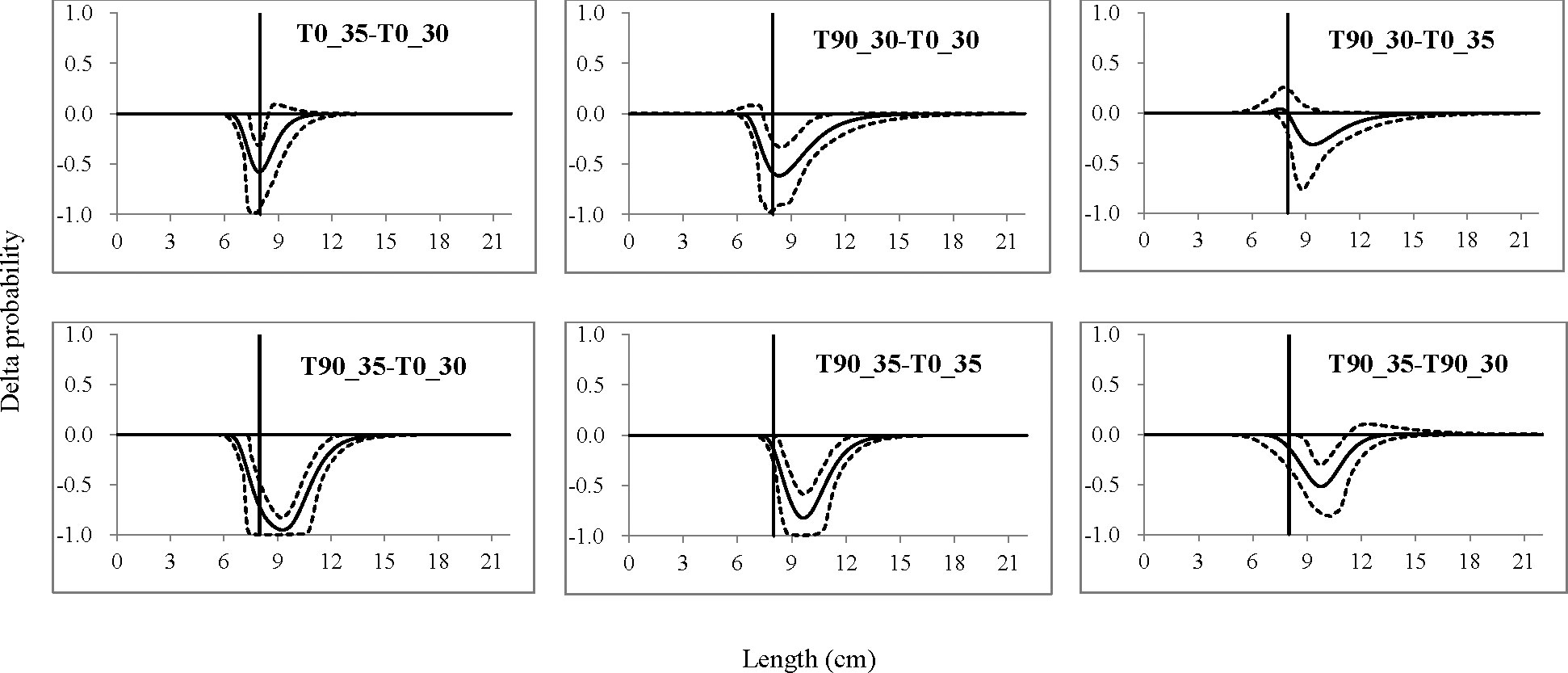
Figure 4 Delta selectivity of comparison between the tested codends. The black curves represent the delta selectivity for comparison, and the dotted curves are the 95% confidence intervals (CIs). Vertical lines are the MCRS (minimum conservation reference size) of mantis shrimp derived from the minimum length at maturity in the South China Sea (SCS). If the CIs of delta selectivity do not overlap 0.0 in some length classes, the difference is significant.
Exploitation pattern indicators
The exploitation pattern indicators showed that modifying the mesh sizes and shape would reduce the retention of mantis shrimp both for the undersized (nP−) and legal-size (nP+) individuals, although the values were fishing population scenario dependent. In the fishing population of 2019, for instance, the values nP− and nP+ were 35.67% (CI: 5.09–66.52) and 98.88 (CI: 96.69–100.00%) for the T0_30 codend; when increasing the mesh size or substituting with a T90 mesh shape, the values would be somewhat reduced; and if both mesh size increment and T90 mesh shape were applied (T90_35), the figures significantly dropped to 0.00 (CI: 0.00–0.00) and 78.20% (CI: 69.61%–86.18%) (Table 5). A similar trend was obtained for the fishing population in 2020. Relatively small values of nRatio and dnRatio were presented for the tested codends in these fishing populations.
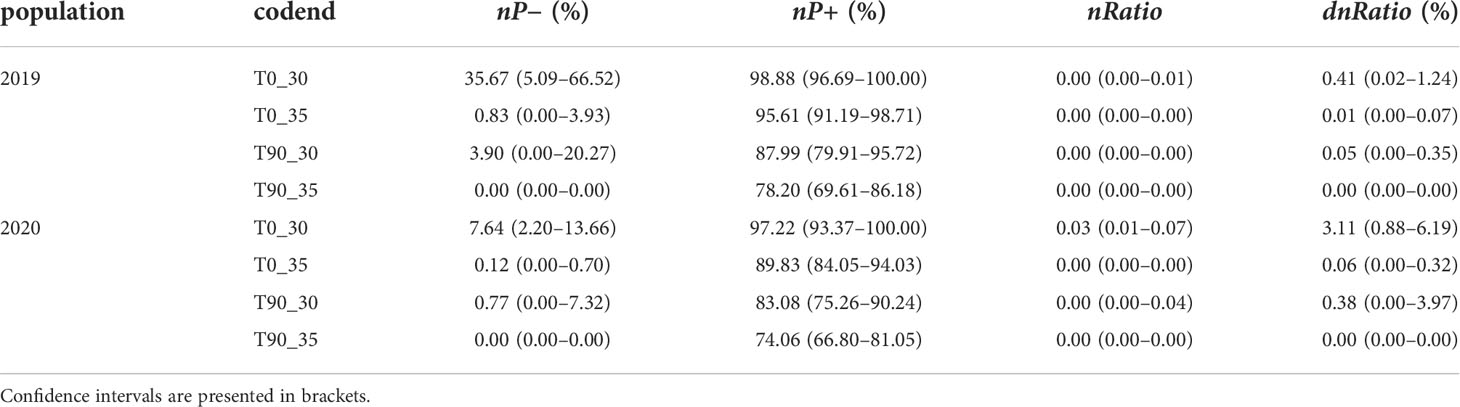
Table 5 Performance indicators obtained for the tested codends in fishing population of 2019 and 2020, respectively.
Discussion
To the best of our knowledge, we presented the first scientific study concerning the size selectivity of trawl codends for mantis shrimp in the SCS. Our results demonstrated that increasing the mesh size in the diamond-mesh codend or applying the T90 codend could improve the size selectivity for mantis shrimp. The improvement would be greatly achieved by using a T90 codend with a larger mesh size. As a starting point, we tested a diamond-mesh codend (T0_30) with a mesh size larger than the current MMS regulations’ requirement (25 mm). Its selective properties were not good enough to release undersized individuals of mantis shrimp. Because its L50 was less than the MCRS of mantis shrimp, the retention probability at the MCRS length was above 72%, and more than 35% of undersized mantis shrimp were retained in the fishing population scenarios of 2019. By comparison, the size selectivity could be improved through modification in mesh size or mesh shape. For instance, the T0_35 and T90_30 codends had larger L50 values than the MCRS of mantis shrimp and lower retention risk for undersized individuals. If modifications in mesh size and mesh shape were both applied, the T90_35 codend, the greatest size selectivity would be obtained. Applying this T90 codend with a larger mesh size, however, might compromise the catch of marketable-size individuals. For instance, the T90_35 codend had lower nP+ values than the other three codends, and some of these differences were statistically significant. Considering the trade-off between releasing undersized individuals and maintaining the legal ones, the T0_35 and T90_30 codends might be better choices to target mantis shrimp in demersal trawl fisheries of the SCS.
Petetta et al. (2020) compared the size selectivity of the T0 codend with the T90 codend, both with a mesh size of 54 mm, for another mantis shrimp species (Squilla mantis) in Mediterranean bottom trawl fisheries. Their results demonstrated that the T90 codend obtained a significantly higher L50 value than that of the T0 codend, 20.78 (CI: 18.79–22.27) vs. 13.35 (7.53–17.62) cm in carapace length, and the T90 codend had significantly lower retention probability for undersized individuals (<26 mm carapace length) (Petetta et al., 2020). Although the fishing gears used and codends tested might have some differences between the present study and those of Petetta et al. (2020), the data collection and analytical methods were the same, and both studies had similar results that the T90 codends had better size selectivity than the T0 codends with the same mesh sizes for mantis shrimp. However, because Petetta et al. (2020) used carapace length in their results while we applied total length, it might be difficult to further compare their selectivity parameters with ours. There are also two selectivity studies about the separator grid for mantis shrimp. One was conducted for shrimp beam trawl fisheries by Tokai et al. (1996) and the other for stow-net fisheries by Wang et al. (2017). Though the results of these studies might be hard to compare with ours, as the working principle of separator grid and codends differ, their results also have some implications for future work to further improve the size selectivity of fishing gears for mantis shrimp.
T90 codends have been widely tested around the world, and many of these studies proved that the selective properties would be improved for some species. For instance, Madsen et al. (2012) demonstrated that the T90 codend improved size selectivity for Norway lobster when compared with the standard diamond-mesh codend. More recently, Robert et al. (2020) and Cheng et al. (2020) reported that applying the T90 codends would improve selective properties from relative and absolute selectivity perspectives. To be consistent with these previous studies, our study also demonstrated that a positive improvement in size selectivity for mantis shrimp would be obtained by applying the T90 codends. These significant effects can be due to 1) more open meshes in the T90 codends and 2) the active swimming capacity and behavior of mantis shrimp. It has been demonstrated that the T90 codend would be more open than the traditional diamond-mesh codend in some previous studies (Madsen et al., 2012; Tokaç et al., 2014; Bayse et al., 2016). After the experiments, we put the two pairwise codends together, and the meshes of the T90 codend were more open than those of the diamond-mesh codend with the same mesh size. When the meshes of the codends were open, mantis shrimp would have more chance to escape. It has been reported that the swimming capacity of mantis shrimp was high, and they can squeeze their bodies through some small holes (Wang and Xu, 1996). Additionally, Jian (2016) estimated the swimming speed of mantis shrimp from an aquaculture tank and found that the speed of undersized individuals was mainly 1 to 3.5 cm/s, and 3 to 6 cm/s for adult individuals.
Recently, Cheng et al. (2022) tested the hydrodynamic performance of a T0 codend and three T90 codends using a flume tank in Canada. Their results showed that by applying the covered codend method, the water velocity between the cover and the T90 codend was significantly lower than that inside the T0 codend (Cheng et al., 2022). What is the situation in an at-sea experiment? If this difference exists, would it affect the escape behavior of mantis shrimp and then further impact the size selectivity of the tested codends? Unfortunately, however, we did not have detailed information to address these questions, and future studies are strictly needed.
At present, there is no official MLS enforced to supplement the MMS regulation for mantis shrimp in the SCS. Our study used its first matured length (8.0 cm) (Xu et al., 1996; Yang et al., 2004) as the MCRS to calculate the exploitation pattern indicators. In order to protect the undersized individuals, however, some studies suggested that the MLS of mantis shrimp should be larger than 8.0 cm. For instance, Liu et al. (2020) recommended that the minimum landing size of mantis shrimp would be larger than 9.5 cm in the Yellow Sea of China. Recently, Xu et al. (2022) suggested that the MLS of mantis shrimp should be larger than 9.42 cm in the Bohai Sea. If the MCRS is set to be 9.5 cm, the benefit of using the T90 codend will be further manifested. Because as demonstrated in Figure 3 the retention probability of the two diamond-mesh codends for mantis shrimp at the length of 9.5 cm was higher than 84%, in comparison, this value is about 53.38% and 3.14% for the T90_30 and T90_35 codends, respectively. Correspondingly, our results demonstrate that if the MCRS of mantis shrimp increase to 9.5 cm, for instance, the T90 codend would be an option to improve the size selectivity and to better match the MCRS regulation.
Facing the overexploited fisheries resources, the large size of the fishing fleets, and the complicated role of marine fisheries in China, the technical regulations in fisheries management are suggested to be simple (Shen and Heino, 2014; Cao et al., 2015; Cao et al., 2017; Su et al., 2020). Moreover, simple modification through codend configuration has been recently highlighted as an excellent starting point to address bycatch issues of trawl fisheries by Kennelly and Broadhurst (2021). Specifically, our study provides a case of gear modification to improve the size selectivity and catch efficiency for mantis shrimp in demersal trawl fishery. There are many simple options, however, that need to be tested and considered. For instance, using the square mesh panels in the diamond-mesh codends (Graham and Kynoch, 2001; Cuende et al., 2020; Kennelly and Broadhurst, 2021), shortening the lastridge ropes (Sistiaga et al., 2021), and adding artificial stimuli (Krag et al., 2017; Grimaldo et al., 2018; Melli et al., 2018; Ingólfsson et al., 2021) have been demonstrated to be simple and effective modifications to improve the size selectivity. It is suggested that future investigations should test and evaluate these modifications to further improve the size selectivity and catch efficiency of trawl fisheries for mantis shrimp in the SCS.
In addition, although our results demonstrated that modifications in the trawl codend could improve the size selectivity of mantis shrimp and provide mitigation measures for the bycatch issues, another major concern is the fate of escaping mantis shrimp. The mortality of animals escaping from the fishing gears is becoming an ever-increasingly important issue in fisheries management decision-making processes (Broadhurst et al., 2006; Suuronen and Erickson, 2010). Despite the fact that we did not have detailed data to address the mortality of mantis shrimp escaping from the codends, the commercial fishing practice indicates, to some extent, that mantis shrimp escaping from the gear might not suffer high mortality. As we witnessed onboard, after the fishing process, most mantis shrimp were kept alive in some water tanks and landed several days later. With this evidence, we assume that mantis shrimp will have low fishing mortality from the trawl codend. This assumption, however, needs to be validated by strict field experiments to truly understand the mortality of mantis shrimp escaping from the codends.
Conclusion
Our study demonstrated that increasing the mesh size of the T0 codend or applying the T90 codend would improve the size selectivity of the trawl codend for mantis shrimp. Considering the trade-off between releasing undersized individuals and maintaining the legal ones, the T0 codend with 35-mm mesh size or the T90 codend with 30-mm mesh size might be a better choice to target mantis shrimp in demersal trawl fisheries of the SCS.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
BY: conception, investigation, writing original draft, funding acquisition, review and editing of the original draft, data curation, and validation. BH: software, formal analysis, review, and editing of the original draft, and supervision. All authors contributed to the article and approved the submitted version.
Funding
This work was funded by the National Key R&D Program of China (2020YFD0901205) and Central Public-interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS (NO. 2020YJ01) and financially supported by the Ministry of Agriculture and Rural Affair “Standard and management regime of fishing gears”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.939269/full#supplementary-material
References
Akaike H. (1974). A new look at the statistical model identification. IEE. Trans. Automat. Contr. 19, 716–723. doi: 10.1109/TAC.1974.1100705
Bayse S. M., Herrmann B., Lenoir H., Depestele J., Polet H., Vanderperren E., et al. (2016). Could a T90 mesh codend improve selectivity in the Belgian beam trawl fishery? Fish. Res. 174, 201–209. doi: 10.1016/j.fishres.2015.10.012
Boenish R., Kritzer J., Kleisner K., Steneck R. S., Werner K. M., Zhu W., et al. (2021). The global rise of crustacean fisheries. Front. Ecol. Environ. 20 (2), 102–110. doi: 10.1002/fee.2431
Brinkhof J., Larsen R. B., Herrmann B. (2022). Make it simpler and better: T90 codend improves size selectivity and catch efficiency compared with the grid-and-diamond mesh codend in the northeast Atlantic bottom trawl fishery for gadoids. Ocean. Coast. Manage. 217, 106002. doi: 10.1016/j.ocecoaman.2021.106002
Broadhurst M. K., Suuronen P., Hulme A. (2006). Estimating collateral mortality from towed fishing gear. Fish. Fish. 7, 180–218. doi: 10.1111/j.1467-2979.2006.00213.x
Cao L., Chen Y., Dong S., Hanson A., Huang B., Leadbitter D., et al. (2017). Opportunity for marine fisheries reform in China. PNAS 114 (3), 435–442. doi: 10.1073/pnas.1616583114
Cao L., Naylor R., Henriksson P., Leadbitter D., Metian M., Troell M., et al. (2015). China’s aquaculture and the world’s fisheries. Science 347 (6218), 133–135. doi: 10.1126/science.1260149
Cashion T., Al-Abdulrazzk D., Belhabib D., Derrick B., Divovich E., Moutopoulos D. K., et al. (2018). Reconstructing global marine fishing gear use: Catches and landed values by gear type and sector. Fish. Res. 206, 57–64. doi: 10.1016/j.fishres.2018.04.010
Cheng Z., Winger P. D., Bayse S. M., Kebede G. E., DeLouche H., Einarsson H. A., et al. (2020). Out with the old and in with the new: T90 codends improve size selectivity in the Canadian redfish (Sebastes mentella) trawl fishery. Can. J. Fish. Sci. 00, 1–10. doi: 10.1139/ajfas-2020-0063
Cheng Z., Winger P. D., Bayse S. M., Kelly D. (2022). Hydrodynamic performance of full-scale T0 and T90 codends with and without a codend cover. J. Mar. Sci. Eng. 10, 440. doi: 10.3390/jmse10030440
Cuende E., Arregi L., Herrmann B., Sistiaga M., Onandia I. (2020). Stimulating release of undersized fish through a square mesh panel in the Basque trawl fishery. Fish. Res. 224, 105431. doi: 10.1016/j.fishres.2019.105431
Efron B. (1982). “SIAM monograph no. 38, CBSM-NSF regional conference series,” in The jackknife, the bootstrap and other resampling plans (Philadelphia: Applied Mathematics).
Eigaard O., Herrmann B., Nielsen J. R. (2012). Influence of grid orientaion and time of day on grid sorting in a small-meshed trawl fishery for Norway pout (Trisopterus esmarkii). Aqut. Living. Resour. 25, 15–26. doi: 10.1051/alr/2011152
Fryer R. J. (1991). A model of between-haul variation in selectivity. ICES J. Mar. Sci. 48, 281–290. doi: 10.1093/icesjms/48.3.281
Fryer R. J., O'Neill F. G., Edridge A. (2016). A meta-analysis of haddock size-selection data. Fish. Fishe. 17 (2), 358–374. doi: 10.1111/faf.12107
Glass G. W. (2000). Conversation of fish stocks through bycatch reduction: a review. Northeast. Nat. 7, 395–410. doi: 10.1656/1092-6194(2000)007[0395:COFSTB]2.0.CO;2
Graham N., Kynoch R. J. (2001). Square mesh panels in demersal trawls: some data on haddock selectivity in relation to mesh size and position. Fish. Res. 49, 207–218. doi: 10.1016/S0165-7836(00)00211-3
Grimaldo E., Larsen R. B., Sistiaga M., Madesen N., Breen M. (2009). Selectivity and escape percentage during three phases of the towing process for codends fitted with different selection systems. Fish. Res. 95, 198–205. doi: 10.1016/j.fishres.2008.08.019
Grimaldo E., Sistiaga M., Herrmann B., Larsen R. B., Brinkhof J., Tatone I. (2018). Improving release efficiency of cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) in the barents Sea demersal trawl fishery by stimulating escape behaviour. Can. J. Fish. Aquat. Sci. 75, 402–416. doi: 10.1139/cjfas-2017-0002
He P. (2007). Selectivity of large mesh trawl codends in the gulf of Maine: I. Comparison of square and diamond mesh. Fish. Res. 83, 44–59. doi: 10.1016/j.fishres.2006.08.019
Herrmann B. (2005a). Effect of catch size and shape on the selectivity of diamond mesh cod-ends I. Model development. Fish. Res. 71 (1), 1–13. doi: 10.1016/j.fishres.2004.08.024
Herrmann B. (2005b). Effect of catch size and shape on the selectivity of diamond mesh cod-ends II. Theoretical study of haddock selection. Fish. Res. 71 (2), 15–26. doi: 10.1016/j.fishres.2004.08.021
Herrmann B., Krag L. A., Krafft B. A. (2018). Size selection of Antarctic krill (Euphausia superba) in a commercial codend and trawl body. Fish. Rese. 207, 49–54. doi: 10.1016/j.fishres.2018.05.028
Herrmann B., Priour D., Krag L. A. (2007). Simulation-based study of the combined effect on cod-end size selection of turning meshes by 90 ° and reducing the number of meshes in the circumference for round fish. Fish. Res. 84 (2), 222–232. doi: 10.1016/j.fishres.2006.10.020
Herrmann B., Sistiaga M., Nielsen K. N., Larsen R. B. (2012). Understanding the size selectivity of redfish (Sebastes spp.) in north Atlantic trawl codends. J. Northwest. Atl. Fish. Sci. 44, 1–13. doi: 10.2960/J.v44.m680
Herrmann B., Sitiaga M., Larsen R. B., Brinkhof J. (2019). Effect of three different codend designs on the size selectivity of junvenile cod in the barents Sea shrimp trawl fishery. Fish. Res. 219, 1–8. doi: 10.1016/j.fishres.2019.105337
Ingólfsson Ó.A., Jørgensen T., Sistiaga M., Kvalvik L. (2021). Artificial light improves size selection for northern shrimp (Pandalus borealis) in trawls. Can. J. Fish. Aquati. Sci. 78, 1910–1917. doi: 10.1139/cjfas-2020-0458
Ishii H., Kitahara T. (2002). A discarding problem of mantis shrimp in Tokyo bay. Fish. Sci. 68 (sup1), 166–169. doi: 10.2331/fishsci.68.sup1_166
Jian T. (2016). A preliminary study on the behavoir in oratosqilla oratoria (Dalian, China: Dalian Ocean University).
Jin X. S., Tang Q. S. (1998). The structure , distribution and variation of the fishery resources in the bohai Sea. J. Fish. Sci. China 5 (3), 18–24.
Kang B., Liu M., Huang X. X., Li J., Yan Y. R., Han C. C., et al. (2018). Fisheries in Chinese seas: What can we learn from controversial official fisheries statistics? Rev. Fish. Biol. Fisher. 28, 503–519. doi: 10.1007/s11160-018-9518-1
Kennelly S. J., Broadhurst M. K. (2021). A review of bycatch reduction in demersal fish trawls. Rev. Fish. Biol. Fish. 31, 289–318. doi: 10.1007/s11160-021-09644-0
Kim S. E., Kim H. J., Bae H. J., Kim H. G., Oh C. W. (2017). Growth and reproduction of the Japanese mantis shrimp, Oratosquilla oratoria (De haan 1844) in the coastal area of tongyeong, Korea. Ocean. Sci. J. 52, 257–265. doi: 10.1007/s12601-017-0027-2
Kodama K., Shimizu T., Yamakawa T., Aoki I. (2006). Changes in reproductive patterns in relation to decline in stock abundance of the Japanese mantis shrimp oratosquilla oratoria in Tokyo bay. Fish. Sci. 72, 568–577. doi: 10.1111/j.1444-2906.2006.01185.x
Kodama K., Shimizu T., Yamakaw T., Aoki I. (2004). Reproductive biology of the female Japanese mantis shrimp Oratosquilla oratoria (Stomatopoda) in relation to changes in the seasonal pattern of larval occurrence in Tokyo bay, Japan. Fish. Sci. 70, 734–745. doi: 10.1111/j.1444-2906.2004.00866.x
Kodama K., Shiraishi H., Morita M., Horiguchi T. (2009). Reproductive biology of the Japanese mantis shrimp oratosquilla oratoria (Crustacea stomatopoda): Annual cycle of gonadal development and copulation. Mar. Biol. Res. 5 (5), 415–426. doi: 10.1080/17451000802644714
Kodama K., Yamakawa T., Shimizu T., Aoki I. (2005). Age estimation of the wild population of Japanese mantis shrimp Oratosquilla oratoria (Crustacea: Stomatopoda) in Tokyo bay, Japan, using lipofuscin as an age marker. Fish. Sci. 71, 141–150. doi: 10.1111/j.1444-2906.2005.00941.x
Krag L. A., Herrmann B., Feekings J., Lund. H. S., Karlsen J. D. (2017). Improving escape panel selectivity in nephrops-directed fisheries by actively stimulating fish behavior. Can. J. Fish. Aquat. Sci. 74, 486–493. doi: 10.1139/cjfas-2015-0568
Larsen R. B., Herrmann B., Sistiaga M., Brčić J., Brinkhof J., Tatone I. (2018). Could green artificial light reduce bycatch during barents Sea deep-water trawling? Fish. Res. 204, 441–447. doi: 10.1016/j.fishres.2018.03.023
Liu Y., Zhang C., Liu S., Wang S., Ren Y. (2020). Yield per recruitment evaluation of oratosquilla oratoria in cosatal waters of shangdong. J. Fish. China 44 (2), 213–221. doi: 10.11964/jfc.20181211595
Madsen N., Hansen K., Madsen N. A. H. (2015). Behavior of different trawl codend concepts. Ocean. Eng. 108, 571–577. doi: 10.1016/j.oceaneng.2015.08.047
Madsen N., Herrmann B., Frandsen R. P., Krag L. A. (2012). Comparing selectivity of a standard and turned mesh T90 codend during towing and haul-back. Aquat. Living. Resour. 25 (3), 231–240. doi: 10.1051/alr/2012021
Melli V., Krag L. A., Herrmann B., Karlsen J. D. (2018). Investigating fish behavioural responses to LED lights in trawls and potential applications for bycatch reduction in the nephrops-directed fishery. ICES J. Mar. Sci. 75 (5), 1682–1692. doi: 10.1093/icesjms/fsy048
Millar R. B. (1993). Incorporation of between-haul variation using bootstrapping and nonparametric estimation of selection curves. Fish. Bull. 91, 564–572.
Nakajima M., Kodama K., Horiguchi T., Tanaka Y., Shiraishi H. (2010). Impacts of shifts in spawning seasonality and size at maturation on the population growth of mantis shrimp in Tokyo bay. Mar. Ecol. Prog. Ser. 418, 179–188. doi: 10.3354/meps08824
O’Neill F. G., Fryer R. J., Frandsen R. P., Herrmann B., Madsen N., Mieske B. (2020). A meta-analysis of plaice size-selection data in otter trawl codends. Fish. Res. 227, 105558. doi: 10.1016/j.fishres.2020.105558
Petetta A., Herrmann B., Virgili M., De Marco R., Canduci G., Veli D. L., et al. (2020). Estimating selectivity of experimental diamond (T0) and turned mesh (T90) codends in multi-species Mediterranean bottom trawl. Mediterr. Mar. Sci. 21 (3), 545–557. doi: 10.12681/mms.22789
Robert M., Morandeau F., Scavinner M., Fiche M., Larnaud P. (2020). Toward elimination of unwanted catches using a 100 mm T90 extension and codend in demersal mixed fisheries. PloS One 15 (7), e0235368. doi: 10.1371/journal.pone.0235368
Shen G., Heino M. (2014). An overview of marine fisheries management in China. Mar. Policy 44, 265–272. doi: 10.1016/j.marpol.2013.09.012
Sistiaga M., Brinkhof J., Herrmann B., Larsen R. B., Grimaldo E., Cerbule K., et al. (2021). Potential for codends with shortened lastridge ropes to replace mandated selection devices in demersal trawl fisheries. Can. J. Fish. Aquat. Sci. 00, 1–16. doi: 10.1139/cjfas-2021-0178
Su S., Tang Y., Chang B., Zhu W., Chen Y. (2020). Evolution of marine fisheries management in China from 1949 to 2019: How did China get here and where does China go next? Fish. Fish. 21, 435–452. doi: 10.1111/faf.12439
Suuronen P., Erickson D. L. (2010). “Mortality of animals that escape fishing gears or are discarded after capture: Approaches to reduce mortality,” in Behavior of marine fishes: Capture processes and conservation challenges. Ed. He P. (Ames: Wiley-Blackwell), 265–293.
Tokaç A., Herrmann B., Aydin C., Özbilgin H., Kaykaç H., Ünlüler A., et al. (2014). Predictive models and comparison of the selectivity of standard (T0) and turned mesh (T90) codends for three species in the Eastern Mediterranean. Fish. Res. 150, 76–88. doi: 10.1016/j.fishres.2013.10.015
Tokai T., Ito H., Masaki Y., Kitahara T. (1990). Mesh selectivity curves of a shrimp beam trawl for southern rough shrimp trachypenaeus curvirostris and mantis shrimp Oratosquilla oratoria. Nippon. Suisan Gakkaishi 56 (8), 1231–1237. doi: 10.2331/suisan.56.1231
Tokai T., Omoto S., Sato R., Matuda R. (1996). A method of determining selectivity curve of separator grid. Fish. Res. 27 (1–3), 51–60. doi: 10.1016/0165-7836(95)00471-8
Wang Z., Gao Z., Zhang J. (2017). Selectivity study on y-shaped exclusion device for mantis shrimp and swimming crab in Chinese stow net. South China Fish. Sci. 13 (5), 117–123. doi: 10.3969/j.issn.2095-0780.2017.05.016
Wang C., Xu S. (1996). A biological basic character of oratosquilla oratoria. J. Zhejiang Coll. Fish. 15 (1), 60–62.
Wileman D., Ferro R. S. T., Fonteyne R., Millar R. B. (1996). Manual of methods of measuring the selectivity of towed fishing gear. ICES Coop. Res. Rep. 215, 1–126. doi: 10.17895/ices.pub.4628
Xu S., Wang C., Mei W., Bao B. (1996). Preliminary studies on propagation and feeding habits of oratosquilla oratoria in northern zhejiang Sea areas. J. Zhejiang Coll. Fisher. 15 (1), 30–36.
Xu H., Zhao R., Liang Q., Gu D., Xu L., Zhang D. (2022). Length at first sexual maturity of female mantis shrimp (Oratosquilla oratoria) in bohai bay based on gonadal development. J. Shanghai Ocean. Univ. 31 (3), 721–728. doi: 10.12024/jsou.20220303748
Yang B., Herrmann B., Yan L., Li J., Wang T. (2021). Comparing size selectivity and exploitation pattern of diamond-mesh codends for southern velvet shrimp (Metapenaeopsis palmensis) in shrimp trawl fishery of the south China Sea. PeerJ 9, e12436. doi: 10.7717/peerj.12436
Yang C., Su X., Li T. (2004). Aquaculture and artificial breeding of mantis shrimp (Oratosquilla oratoria). Shangdong Fish. 21 (3), 9–11.
Keywords: demersal trawl, codend selectivity, catch efficiency, T0, T90
Citation: Yang B and Herrmann B (2022) Simple and effective: T90 codends improve size selectivity and catch efficiency compared with diamond-mesh codends for mantis shrimp (Oratosquilla oratoria) in demersal trawl fishery of the South China Sea. Front. Mar. Sci. 9:939269. doi: 10.3389/fmars.2022.939269
Received: 09 May 2022; Accepted: 13 September 2022;
Published: 30 September 2022.
Edited by:
Alessandro Lucchetti, National Research Council (CNR), ItalyReviewed by:
Claudio Vasapollo, National Research Council (CNR), ItalyTadashi Tokai, Tokyo University of Marine Science and Technology, Japan
Russell Millar, The University of Auckland, New Zealand
Copyright © 2022 Yang and Herrmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bingzhong Yang, eWFuZ2Jpbmd6aG9uZ0BzY3NmcmkuYWMuY24=