 Enrique González-Ortegón1*
Enrique González-Ortegón1* Selina Berger2
Selina Berger2 João Encarnação3Hicham Chairi4
João Encarnação3Hicham Chairi4 Pedro Morais3,5
Pedro Morais3,5 Maria Alexandra Teodósio3
Maria Alexandra Teodósio3 Francisco J. Oliva-Paterna6
Francisco J. Oliva-Paterna6 Christoph D. Schubart2
Christoph D. Schubart2 Jose A. Cuesta1
Jose A. Cuesta1- 1Instituto de Ciencias Marinas de Andalucía, CSIC, Puerto Real, Spain
- 2Zoology and Evolutionary Biology, University of Regensburg, Regensburg, Germany
- 3Centre of Marine Sciences (CCMAR), University of Algarve, Faro, Portugal
- 4Polydisciplinary Faculty of Larache, Abdelmalek Essaadi University, Tetouan, Morocco
- 5Institute of Environment, Florida International University, Miami, FL, United States
- 6Departamento de Zoología y Antropología Física, Universidad de Murcia, Murcia, Spain
The invasive Atlantic blue crab Callinectes sapidus has considerably extended its distribution along the Gulf of Cadiz and Moroccan coasts in the northeast Atlantic. This could indicate a new introduction event in the Gulf of Cadiz from the native area, in case of detectable genetic differences from the Mediterranean Sea populations. To test this and reconstruct the invasion, we assessed the genetic structure of crabs in the Gulf of Cadiz and Alboran Sea. We used sequences of the mitochondrial gene cytochrome c oxidase subunit I (COI), in almost its full length (1511 basepairs), of 149 individuals from three sites along the Spanish Mediterranean coast, two in the Alboran Sea, and two in the Gulf of Cadiz. Our data reveal low genetic variability, with only two haplotypes (here referred to as CSWM1 and CSWM2), compared to its known high genetic diversity in the native range, indicating a strong founder effect in the study region. Here we document an inversion of haplotype predominance between regions, haplotype CSWM2 being dominant in most of the Spanish Mediterranean coast, but less frequent in the Gulf of Cadiz and Alboran Sea. This suggests a secondary introduction of propagules into the investigated area, rather than a new introduction from the native area, nor a natural westward expansion with environmental selection of one haplotype over the other. Further studies with additional populations would help to better unveil the history of yet another invasive species in the Gulf of Cadiz.
Introduction
The range expansion of aquatic non-indigenous species (NIS) can be caused by natural phenomena (e.g., migrations, facilitated by other species, oceanographic currents) or be human-mediated (Galil et al., 2019). The most prominent human-mediated dispersal vectors in aquatic ecosystems are intentional introductions, aquaculture, and shipping/boating (e.g., ballast water, hull fouling) (Gollasch, 2006; González-Ortegón et al., 2007; Cuesta et al., 2016). The invasion of exotic species is being accelerated worldwide as a consequence of expanding transport and commerce (Mack et al., 2000); for instance, non-native species have been reported in the Gulf of Cadiz since 1980, but have increased rapidly in the last five years (González-Ortegón et al., 2020). The so-called Atlantic blue crab Callinectes sapidus Rathbun, 1896, a species native to the western Atlantic Ocean, is a good example for the introduction and fast growth of new non-native populations in the Euro-African area (Nehring, 2011; Mancinelli et al., 2021). Ballast water has been accepted as the most probable introduction vector of this species in numerous regions (Nehring, 2011).
C. sapidus has a disjunct distribution in the native range, from the coast of Massachusetts (United States) to Venezuela and then in the southwestern Atlantic from Alagoas (Brazil) to northern Argentina (dos Santos and D’Incao, 2004; Windsor et al., 2019). It was introduced in the Eastern Atlantic, including the Mediterranean, Adriatic, North and Baltic seas as well as waters of Hawaii and Japan, via human-mediated vectors (Windsor et al., 2019). This species often uses estuaries to complete its life cycle and is a particularly successful invader of marine and estuarine ecosystems, because of its high fecundity and larval survivability, broad environmental tolerance, large body size, aggressive behavior, omnivorous diet, and strong swimming ability (Nehring, 2011). The extended planktonic larval phase of this species maximizes its chances of introduction through ballast water (DiBacco et al., 2012), as well as natural dispersal (Encarnação et al., 2022; Png-Gonzalez et al., 2021).
In European Atlantic waters, C. sapidus was detected for the first time along the Atlantic coast of France in 1901 (Bouvier, 1901). In the Mediterranean Sea, it was reported in the Aegean Sea as early as the 1930s, and in the adjacent Turkish coast in 1949 (Enzenroß et al., 1997). Since then, this species has greatly expanded its distribution along the entire Mediterranean Sea and adjacent seas, but also in the North Sea and the Atlantic coast of the Iberian Peninsula and northern Africa (Encarnação et al., 2021; Mancinelli et al., 2021; et al., 2022; González-Ortegón and Chairi [in press)]. On the western coast of the Iberian Peninsula, the first record dates back to 1967 in the Tagus Estuary (Portugal), but only a few sporadic records were made since then in this region (Encarnação et al., 2022). Along the Mediterranean coast of the Iberian Peninsula, the first record was made in the Ebro Delta in 2012-2013 (Castejón & Guerao, 2013), and since then its distribution expanded rapidly towards the Strait of Gibraltar where it has been registered since 2017 (Cabal et al., 2006; Nehring, 2011; Mancinelli et al., 2017a; Mancinelli et al., 2017b; González-Ortegón et al., 2020; Mancinelli et al., 2021). In the Gulf of Cadiz, the first published record of Atlantic blue crab dates back to 2016 (Morais et al., 2019), however, a male adult specimen was already collected in the Guadalquivir Estuary in 2002 (Rodríguez & Cuesta, unpublished data). Since 2016, the species expanded rapidly westwards along the southern coast of Portugal and has now reached the southwestern coast of Portugal (Encarnação et al., 2022; Vasconcelos et al., 2019; Encarnação et al., 2021). Meanwhile, in the Alboran Sea, the first record was made in 2017 in Nador Lagoon (Morocco, Mediterranean Sea) (Chartosia et al., 2018), and the species later expanded to the east and west (Taybi and Mabrouki, 2020). Today, the species is also present in the Moroccan Atlantic coast ( et al., 2022; González-Ortegón and Chairi [in press)].
Complex oceanographic patterns exist in the Gulf of Cadiz and Strait of Gibraltar, which are influenced by surface currents and the Mediterranean outflow, all of which vary seasonally (García-Lafuente et al., 2006; Peliz et al., 2007; Sánchez-Leal et al., 2017). A recent study demonstrated that the westward expansion of the Atlantic blue crab along the southern Portuguese coast towards the western coast (north of Cape St. Vincent) could be due to a seasonal coastal counter-current that coincides with the species reproductive period (Encarnação et al., 2022), similar to the passive transport of particles in summer under a strong and persistent counter-current event in the Gulf of Cadiz (see Figure 1 in Casaucao et al., 2021). This observation prompted two hypotheses about the origin of C. sapidus in the study regions: 1) new and independent introduction into the Gulf of Cadiz from the native area in the Western Atlantic, or 2) a metapopulation expansion across the Gulf of Cadiz, Alboran Sea, and western Mediterranean Sea. To test these hypotheses, we used genetic tools to compare DNA-sequences of the gene cytochrome c oxidase subunit I (COI) of Atlantic blue crab specimens collected from the Mediterranean coast of eastern of Spain, Alboran Sea, and Gulf of Cadiz and compared them with sequences of individuals from native populations.
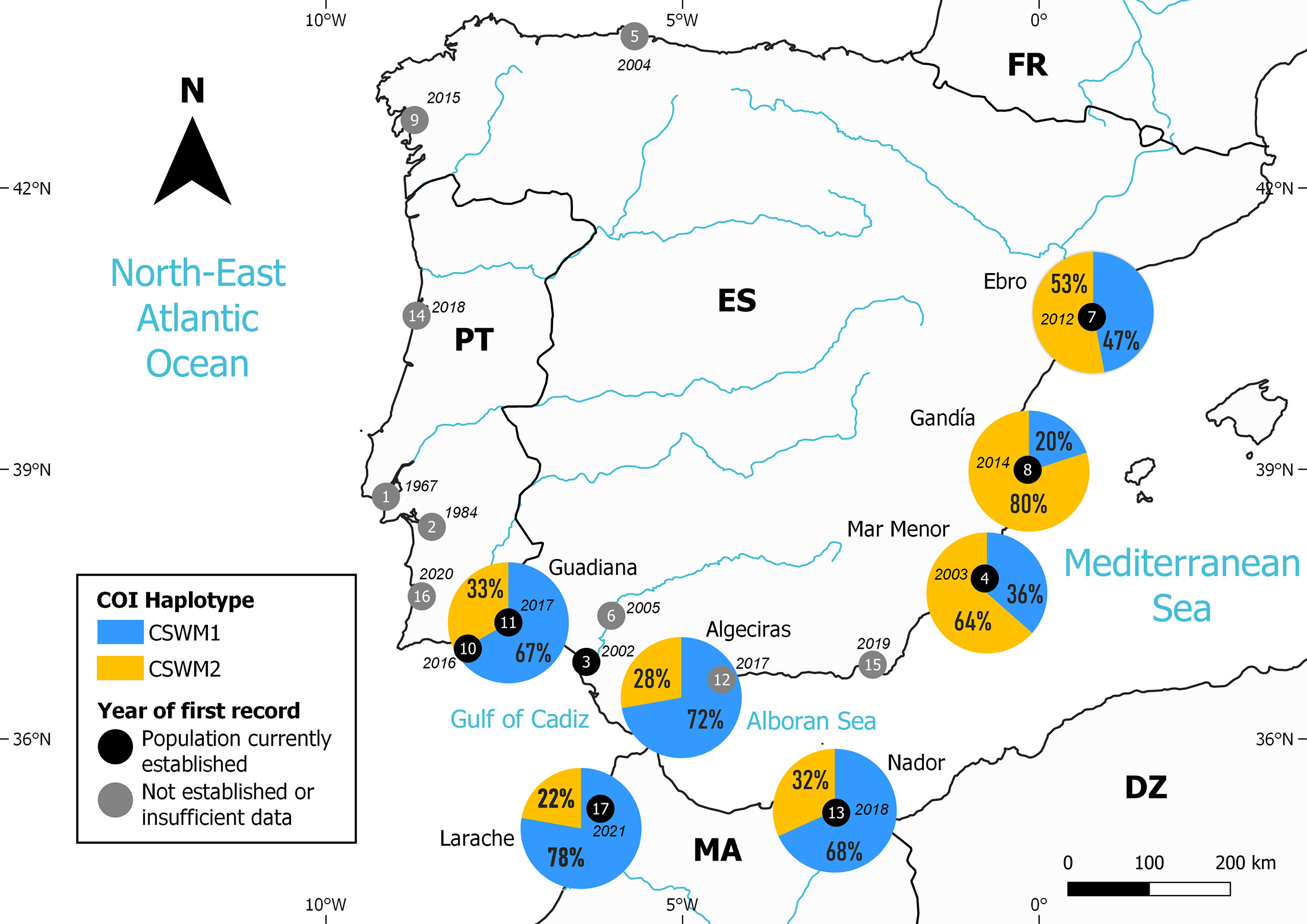
Figure 1 Location of the sampling sites and proportion of COI haplotypes – CSWM1 and CSWM2 – identified in the invasive populations of Atlantic blue crab Callinectes sapidus Rathbun, 1896 analysed for this study. Locations and year of first record are presented for the Atlantic and Mediterranean coasts of the Iberian Peninsula and Morocco. Populations were considered established if evidence of completion of life cycle exists (juveniles, males, females, ovigerous females) or recurrent reports for at least 2 years. The year represents the first known records. Colour grey or black represent the current-day status of populations based on available literature. 1, 2, 16: Encarnação et al. (2022); 3: Rodríguez & Cuesta, (unpublished data); 4: Chic and Garrabou (2020); 5: Cabal et al. (2006); 6: ICES (2007); 7: Castejón and Guerao (2013); 8: Izquierdo-Gómez and Izquierdo-Muñoz in Karachle et al. (2016); 9: Bañón et al. (2016); 10, 11: Morais et al. (2019); 12: González-Ortegón et al. (2020); 13: Oussellam and Bazairi in Chartosia et al. (2018); 14: Izquierdo-Gómez (2022); 15: de Vries and Lemmens (2022); 17: Chaouti et al. (2022); González-Ortegón and Chairi (in press).
Methods
Sampling Sites
Atlantic blue crab specimens were collected from two sites in the Gulf of Cadiz (Guadiana and Larache estuaries), two in the Sea of Alboran (Algeciras and Nador Lagoon), and three in the eastern Mediterranean Spanish coast (Mar Menor, Gandía, Ebro Delta) (Figure 1). Details about the samples and sampling sites are described in Table 1.
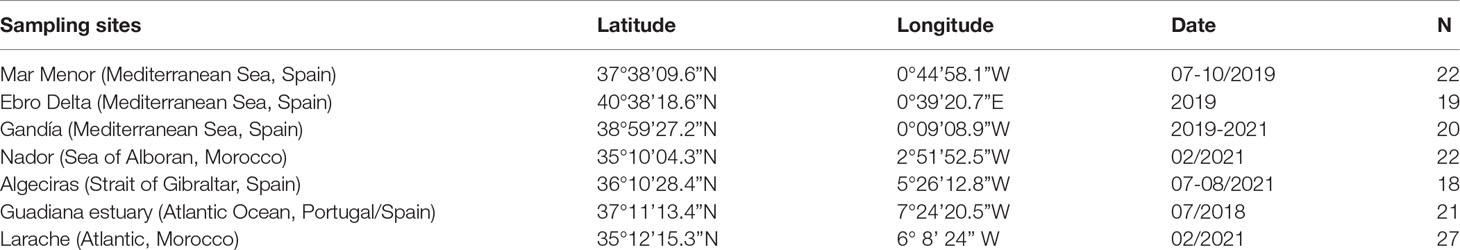
Table 1 Location of the sampling sites where Atlantic blue crab Callinectes sapidus Rathbun, 1896 specimens were collected for mtDNA analyses, date of capture, and number of sequenced specimens (N).
Genetic and Data Analyses
In total, 149 DNA sequences were analysed from individuals collected in these seven locations. In the frame of the present study, the pereiopod muscle tissue from 110 individuals were excised for DNA isolation using a modified Chelex 10% protocol (Estoup et al., 1996). The mitochondrial DNA (mtDNA) of up to 1511 basepairs encoding the enzyme COI were amplified to compare haplotype frequencies between populations. The amplification temperature profile was: i) initial denaturation for 4 min. at 94°C; ii) 40 cycles of 45 s at 95°C for denaturation; iii) 1 min. at 50°C for primer annealing; iv) 1 min. at 72°C for elongation; v) final extension for 4 min. at 72°C. The primers used were COL6a (5’-CAAATCATAAAGAYATTGG-3’) as forward primer and COH16 (5’-CATYWTTCTGCCATTTTAGA-3’) as reverse primers (Schubart, 2009), in addition to the newly designed internal primers COL4Cs (5’-TAGCAGGTGCTATTACTATACT-3’) and COH900Cs (5’-CTAAAAATTTTAATACCAGTGG-3’). PCR products were sent to Stab-Vida Laboratories to be purified and then bidirectionally sequenced. Sequence files were proofread using the trace viewer software Chromas Lite 2.6.4 (Technelysium Pty Ltd 2017), aligned with Clustal W, and implemented in BioEdit 7.2.6.1 (Hall, 1999).
The similarity of haplotypes of the populations of Callinectes sapidus from the Gulf of Cadiz and Alboran Sea and their possible geographic origin were investigated by ‘blasting’ the haplotype sequences in GenBank and searched in BOLD for the same sequences (100% fitting) deposited in these databases.
Results
Only two haplotypes of the mtDNA COI gene were identified in the seven Atlantic blue crab populations analysed in this study, herein referred to as CSWM1 and CSWM2. Sequences of these haplotypes were deposited in GenBank under the accession numbers ON248058 (CSWM1) and ON248059 (CSWM2)and corresponding voucher specimens were deposited in the Decapoda and Stomatopoda Crustacean Collection (Spanish Institute of Oceanography in Cadiz, IEO-CSIC, Spain) under catalogue numbers IEO-CD-ICMAN/2936 (CSWM1) and IEO-CD-ICMAN/2937 (CSWM2).The Gulf of Cadiz and Alboran Sea were predominantly CSWM1 (67-78%), while the Mediterranean Spanish coast was dominated by CSWM2 (53-80%) (Figure 1).
The Blast and BOLD search for the CSWM1 haplotype showed a 100% similarity with GenBank records Turkey-1 (MG462529) (origin: Turkey) from whole frozen uncooked crabs, and USA-2-5 (MG462571) (origin: United States of America) from commercial crabmeat products (Windsor et al., 2019). Two other sequences showed a 100% identity [not considering the primers area] with GenBank sequences KT282079 (Segura River, Mediterranean Sea coast, Spain) (Gonzalez-Wangüemert and Pujol, 2016) and MN759041 (Ordu, Black Sea coast, Turkey) (Öztürk et al., 2020). On the other hand, CSWM2 only fits 100% with the Genbank COI sequence KR030241, corresponding to voucher specimen USNM : IZ:1286742 from Rhode River (Maryland, USA) (Aguilar et al., unpublished).
Discussion
The Gulf of Cadiz is now the most southwestern point in the distribution of Callinectes sapidus along the European coast. The genetic data obtained in this study showed that the populations observed in recent years along the Gulf of Cadiz and Alboran Sea have a low genetic diversity and strong population connectivity. All sampled populations include two haplotypes (CSWM1, CSWM2) that differ in their frequencies between the Spanish Mediterranean coast metapopulation and the Gulf of Cadiz-Alboran Sea metapopulation.
New Introduction Event Versus Expansion or Secondary Introduction?
Invasive species often exhibit low genetic diversity due to a strong founder effect, while the absence of genetic variability between invasive populations is most likely the indication of a single introduction event, with a subsequent expansion along the non-native range or secondary introduction within the non-native range (Harrison et al., 2013). Although cryptic intraspecific invasions are more common than acknowledged, since most biological invasion studies rely on morphological identifications rather than on molecular analyses, a new introduction event into a distinct area would more likely imply that genetic diversity would be distinct between non-native populations (Morais and Reichard, 2018). For example, the northward expansion of the invasive European green crab Carcinus maenas (Linnaeus, 1758) along the northwest Atlantic coast was apparently due to the introduction of a new lineage and not to its expansion northward by climate change or its acclimatization to cold water (Roman, 2006). Thus, although changes in the distribution of an exotic species may become obvious, molecular analyses could warn us when an adaptation of the present genotypes or the introduction of new lineages occur, being able to further increase its invasive potential in the respective area.
The genetic diversity of Callinectes sapidus in its native range is substantially higher than in our study area. On the U.S. East Coast there are at least 67 haplotypes and three genetic lineages along the entire native area (Windsor et al., 2019). Lineage 1 is predominant along the Atlantic coast of North America and Gulf of Mexico, lineage 2 is predominant in the Caribbean Sea, while lineage 3 is predominant and exclusive to the southwestern Atlantic (Windsor et al., 2019). CSWM1 and CSWM2 are the only haplotypes observed in our study area and are rare in lineage 1 (Windsor et al., 2019). In the invaded range, haplotype CSWM1 has been found in the Eastern Mediterranean Sea, Black Sea, and Turkey (Windsor et al., 2019; Öztürk et al., 2020, Berger et al., unpublished), as well as in the Segura River (Gonzalez-Wangüemert and Pujol, 2016).
Considering these genetic data, and the fact that adult blue crabs have a considerable ability to swim and travel up to hundred kilometers (Kara and Chaoui, 2021) or the high fecundity of females, which would allow the passive transport of a huge amount of larvae by oceanic currents between the Mediterranean Sea and the Gulf of Cadiz, it is more likely that the populations of C. sapidus in the Gulf of Cadiz and Alboran Sea result from the expansion of the species along the Mediterranean Sea – single introduction – or to a secondary introduction event within the invaded range. For example, the timeline of the first reports along the southern coast of the Iberian Peninsula were made sequentially from the east to the west, between the Ebro Delta and the Gulf of Cadiz, in just seven years (Castejón & Guerao, 2013; Morais et al., 2019; González-Ortegón et al., 2020; Encarnação et al., 2021). Additionally, the haplotype diversity is extremely low between populations – only two haplotypes are present. The inversion in haplotypic frequency could be explained by two processes, a secondary introduction of few individuals from the Spanish Mediterranean coast to the Gulf of Cadiz or Alboran Sea, or to changes in abiotic conditions since estuarine ecosystems are absent between Mar Menor and the Strait of Gibraltar. In case of a new introduction into the Gulf of Cadiz and Alboran Sea, a different diversity of haplotypes would have been expected and, probably, one of the most common haplotypes in the Atlantic blue crab native area (e.g. H34, see Windsor et al., 2019) would have been present in these two adjacent non-native areas, but this was not the case.
Population Connectivity
The presence of only two haplotypes of Atlantic blue crabs collected at the west and the east of the Strait of Gibraltar suggests a lack of genetic structuring and high connectivity between the Gulf of Cadiz and Alboran Sea populations. This may be explained by two significant autecological traits, the species protracted pelagic larval duration and adult migration, which are known to favor the gene flow of marine species over long distances (Cordero et al., 2014; Lacerda et al., 2016). Besides these two traits, ship transport may contribute significantly to the gene flow in the southwestern coast of the Iberian Peninsula (Lacerda et al., 2016).
The haplotype frequency discontinuity between CSWM1 and CSWM2 coincides with the Almeria-Oran Front that is known to act as a gene flow barrier between the Alboran Sea and the Mediterranean Basin to the east, as observed for other marine crustaceans (Patarnello et al., 2007), such as the rockpool shrimp Palaemon elegans (Reuschel et al., 2010) and harbour crab Liocarcinus depurator (Pascual et al., 2016). If this marked haplotype cline between these two metapopulations subsists, then it will show that the Almeria-Oran Front can also act as a barrier to the widespread admixture between metapopulations of a migratory crustacean species (Reuschel et al., 2010).
Conclusions
The Atlantic blue crab populations present east and west of the Strait of Gibraltar currently consist of two COI haplotypes – CSWM1 and CSWM2. In the Gulf of Cadiz and Alboran Sea, haplotype CSWM1 is predominant, while haplotype CSWM2 is predominant in the Spanish Mediterranean coast populations. These data suggest that these populations are the product of a single invasion event into the Mediterranean Sea, with subsequent westward expansion or secondary introduction to other areas. Additional studies are needed to continue unveiling the details about the Atlantic blue crab invasion in the southwestern Iberian Peninsula coast.
Data Availability Statement
The data presented in the study are deposited in the https://www.ncbi.nlm.nih.gov/genbank/ repository, accession numbers ON248058 and ON248059.
Author Contributions
EGO, JE, HC, PM, MAT, CDS and JAC contributed to conception and design of the study. EGO, JE, HC, FOP and CS contributed to the collection of the crabs. JAC and SB analysed the samples. EGO wrote the first draft of the manuscript. EGO, PM, JE, CDS and JAC contributed to manuscript revision. All authors read and approved the submitted version.
Funding
This study received Portuguese national funds from FCT - Foundation for Science and Technology through projects UIDB/04326/2020, UIDP/04326/2020 and LA/P/0101/2020. This study also received partial financial support from the ATLAZUL (Poctep/Interreg 0755_ATLAZUL_6_E) and ALFCORAZUL projects (Fundo Ambiental, candidature n° 48). JE has a Ph.D. scholarship (SFRH/BD/140556/2018) funded by FCT, Portugal.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This paper was prepared within the framework of the EcoInvadiz project (P18-RT-5010) from the Regional government of Andalusia and InvBlue project (PID2019-105978RA-I00) from the Spanish “Ministerio de Economía y Competitividad (MINECO), Plan Nacional I+D”. This article is also contribution #1444 from the Institute of Environment at Florida International University. The haplotype frequencies from the Gandía population (generated for an ongoing independent trans-Mediterranean study) could be included thanks to previous efforts by Alberto Gil Fernández, Silvia Laura Falco Giaccaglia and Giorgio Mancinelli. Our thanks to the reviewers for their interesting and helpful corrections and comments. We are grateful to Carlos Sánchez (ICMAN) by laboratory work.
References
Bañón R., Cuesta J. A., Almón B., Pérez-Dieste J., Trigo J. E., Ríos M. B. (2016). First Record of Two Decapod Crustaceans, the Caramote Prawn Penaeus kerathurus and the Blue Crab Callinectes sapidus From Galician Waters (NE Atlantic). Cah. Biol. Mar. 57, 323–328.
Bouvier E. L. (1901). Sur Un Callinectes sapidus M. Rathbun Trouvé a Rochefort. Bull. du Muséum Natl. d’histoire naturelle Paris 7, 1617.
Cabal J., Millán A. P., Arronte J. C. (2006). A New Record of Callinectes sapidus Rathbun 1896 (Crustacea: Decapoda: Brachyura) From the Cantabrian Sea, Bay of Biscay, Spain. Aquat. Invasions 1, 186–187. doi: 10.3391/ai.2006.1.3.14
Casaucao A., González-Ortegón E., Jiménez M. P., Teles-Machado A., Plecha S., Peliz A. J., et al. (2021). Assessment of the Spawning Habitat, Spatial Distribution, and Lagrangian Dispersion of the European Anchovy (Engraulis Encrasicolus) Early Stages in the Gulf of Cadiz During an Apparent Anomalous Episode in 2016. Sci. Total Environ. 781, 146530. doi: 10.1016/j.scitotenv.2021.146530
Castejón D., Guerao G. (2013). A New Record of the American Blue Crab, Callinectes sapidus Rathbun 1896 (Decapoda: Brachyura: Portunidae), From the Mediterranean Coast of the Iberian Peninsula. BioInvasions Rec. 2 (2), 141–143. doi: 10.3391/bir.2013.2.2.08
Chaouti A., Belattmania Z., Nadri A., Serrão E., Encarnação J., Teodósio A., et al. (2022). The Invasive Atlantic Blue Crab Callinectes sapidus Rathbun 1896 Expands its Distributional Range Southward to Atlantic African Shores: First Records Along the Atlantic Coast of Morocco. BioInvasions Rec. 11 (1), 227–237. doi: 10.3391/bir.2022.11.1.23
Chartosia N., Anastasiadis D., Bazairi H., Crocetta F., Deidun A., Despalatovic M., et al. (2018). New Mediterranean Biodiversity Records. Mar. Biodivers. Rec. 19, 398–415. doi: 10.12681/mms.18099
Chic O, Garrabou J. and . (2020). Global Marine Biodiversity Data From Seawatchers Marine Citizen Science Platform Between 1980 and 2020. Blue crab. https://www.gbif.org/ [accessed April 11, 2022]. doi:10.14284/519
Cordero D., Peña J. B., Saavedra C. (2014). Phylogeographic Analysis of Introns and Mitochondrial DNA in the Clam Ruditapes decussatus Uncovers the Effects of Pleistocene Glaciations and Endogenous Barriers to Gene Flow. Mol. Phylogenet. Evol. 71, 274–287. doi: 10.1016/j.ympev.2013.11.003
Cuesta J. A., Almón B., Pérez-Dieste J., Trigo Je., Bañon R. (2016). Role of Ships´ Hull Fouling and Tropicalization Process on European Carcinofauna: New Records in Galician Waters (NW Spain). Biol. Invas.s 18 (3), 619–630. doi: 10.1007/s10530-015-1034-9
de Vries H., Lemmens M. (2022). Observation.org, Nature Data From Around the World. Blue Crab. https://observation.org/ [accessed June 12, 2022]. doi: 10.15468/5nilie
DiBacco C., Humphrey D. B., Nasmith L. E., Levings C. D. (2012). Ballast Water Transport of non-Indigenous Zooplankton to Canadian Ports. ICES J. Mar. Sci. 69, 483–491. doi: 10.1093/icesjms/fsr133
Encarnação J., Baptista V., Teodósio M. A., Morais P. (2021). Low-Cost Citizen Science Effectively Monitors the Rapid Expansion of a Marine Invasive Species. Front. Environ. Sci. 9, 752705. doi: 10.3389/fenvs.2021.752705
Encarnação J., Krug L. A., Teodósio M. A., Morais P. (2022) Coastal Counter-Currents Increase Propagule Pressure of an Aquatic Invasive Species to an Area Where Previous Introductions Failed. Estuaries Coast. doi: 10.1007/s12237-022-01092-8
Enzenroß R., Enzenroß L., Bingel F. (1997). Occurrence of Blue Crab, Callinectes sapidus (Rathbun 1896) (Crustacea, Brachyura) on the Turkish Mediterranean and the Adjacent Aegean Coast and its Size Distribution in the Bay of Iskenderun. Turk. J. Zool. 21, 113–122. doi: 10.55730/1300-0179.2971
Estoup A., Lagiardèr C. R., Perrot E., Chourrout D. (1996). Rapid One-Tube DNA Extraction for Reliable PCR Detection of Fish Polymorphic Markers and Transgenes. Mol. Mar. Biol. Biotechnol. 5, 295–298.
Galil B. S., McKenzie C., Bailey S., Campbell M., Davidson I., Drake L., et al. (2019). ICES Viewpoint Background Document: Evaluating and Mitigating Introduction of Marine non-Native Species via Vessel Bio-Fouling ( ICES Ad Hoc Report 2019). doi: 10.17895/ices.pub.4680
García-Lafuente J., Delgado J., Criado-Aldeanueva F., Bruno M., del Río J., Vargas J. M. (2006). Water Mass Circulation on the Continental Shelf of the Gulf of Cadiz. Deep Sea Res. Part II Top. Stud. Oceanogr. 53 (11-13), 1182–1197. doi: 10.1016/j.dsr2.2006.04.011
Gollasch S. (2006). Overview on Introduced Aquatic Species in European Navigational and Adjacent Waters. Helgol. Mar. Res. 60, 84–89. doi: 10.1007/s10152-006-0022-y
González-Ortegón E., Cuesta J. A., Schubart C. D. (2007). First Report of the Oriental Shrimp Palaemon macrodactylus Rathbun 1902 (Decapoda, Caridea, Palaemonidae) From German Waters. Helgol. Mar. Res. 61, 67. doi: 10.1007/s10152-006-0048-1
González-Ortegón E., Jenkins S., Galil B. S., Drake P., Cuesta J. A. (2020). Accelerated Invasion of Decapod Crustaceans in the Southernmost Point of the Atlantic Coast of Europe: A non-Natives’ Hot Spot? Biol. Inva. 22 (12), 3487–3492. doi: 10.1007/s10530-020-02345-y
González-Ortegón E., Chairi H. (in press). Additional Records of the Blue Crab Callinectes sapidus Rahtbun, 1896 in the Moroccan Sea. BioInvasions Rec.
Gonzalez-Wangüemert M., Pujol J. A. (2016). First Record of the Atlantic Blue Crab Callinectes sapidus (Crustacea: Brachyura: Portunidae) in the Segura River Mouth (Spain, Southwestern Mediterranean Sea). Turk. J. Zool. 40 (4), 615–619. doi: 10.3906/zoo-1511-23
Hall T. (1999). BioEdit. A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. 41, 95–98.
Harrison G. F., Kim K., Collins A. G. (2013). Low Genetic Diversity of the Putatively Introduced, Brackish Water Hydrozoan, Blackfordia virginica (Leptothecata: Blackfordiidae), Throughout the United States, With a New Record for Lake Pontchartrain, Louisiana. Proc. Biol. Soc Wash. 126 (2), 91–102. doi: 10.2988/0006-324X-126.2.91
ICES. (2007). Report of the Working Group on Introductions and Transfers of Marine Organisms (WGITMO). Copenhagen: ICES CM 2007/ACME:05. 160 pp.
Izquierdo-Gómez D., Izquierdo-Muñoz A. (2016). Evidence on the Establishment of the American Blue Crab, Callinectes sapidus Rathburn, 1896, in the Levantine Coast of Spain (Western Mediterranean Sea). In: PK Karachle . New Mediterranean Biodiversity Records (March 2016). Medit Mar Sci 17 (1): 232-233
Izquierdo-Gómez D. (2022). Synergistic Use of Facebook, Online Questionnaires and Local Ecological Knowledge to Detect and Reconstruct the Bioinvasion of the Iberian Peninsula by Callinectes sapidus Rathbun, 1896. Biol. Invas. 24, 1059–1082. doi: 10.1007/s10530-021-02696-0
Kara M. H., Chaoui L. (2021). Strong Invasion of Mellah Lagoon (South-Western Mediterranean) by the American Blue Crab Callinectes sapidus Rathbun 1896. Mar. Pollut. Bull. 164, 112089. doi: 10.1016/j.marpolbul.2021.112089
Lacerda A. L. F., Kersanach R., Cortinhas M. C. S., Prata P. F. S., Dumont L. F. C., Proietti M. C., et al. (2016). High Connectivity Among Blue Crab (Callinectes sapidus) Populations in the Western South Atlantic. PLoS One 11 (4), e0153124. doi: 10.1371/journal.pone.0153124
Mack R. N., Simberloff D., Lonsdale W. M., Evans H., Clout M., Bazzaz F. A. (2000). Biotic Invasions: Causes, Epidemiology, Global Consequences, and Control. Ecol. Appl. 10 (3), 689–710. doi: 10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2
Mancinelli G., Bardelli R., Zenetos A. (2021). A Global Occurrence Database of the Atlantic Blue Crab Callinectes sapidus. Sci. Data 8, 111. doi: 10.1038/s41597-021-00888-w
Mancinelli G., Chainho P., Cilenti L., Falco S., Kapiris K., Katselis G., et al. (2017a). The Atlantic Blue Crab Callinectes sapidus in Southern European Coastal Waters: Distribution, Impact and Prospective Invasion Management Strategies. Mar. Pollut. Bull. 119, 5–11. doi: 10.1016/j.marpolbul.2017.02.050
Mancinelli G., Chainho P., Cilenti L., Falco S., Kapiris K., Katselis G., et al. (2017b). On the Atlantic Blue Crab (Callinectes sapidus Rathbun 1896) in Southern European Coastal Waters: Time to Turn a Threat Into a Resource? Fish. Res. 194, 1–8. doi: 10.1016/j.fishres.2017.05.002
Morais P., Gaspar M., Garel E., Baptista V., Cruz J., Cerveira I., et al. (2019). The Atlantic Blue Crab Callinectes sapidus Rathbun 1896 Expands its non-Native Distribution Into the Ria Formosa Lagoon and the Guadiana Estuary (SW-Iberian Peninsula, Europe). Bioinvasions Rec. 8 (1), 123–133. doi: 10.3391/bir.2019.8.1.14
Morais P., Reichard M. (2018). Cryptic Invasions: A Review. Sci. Total Environ. 613, 1438–1448. doi: 10.1016/j.scitotenv.2017.06.133
Nehring S. (2011). “Invasion History and Success of the American Blue Crab Callinectes sapidus in European and Adjacent Waters,” in In the Wrong Place - Alien Marine Crustaceans: Distribution. Biology and Impacts. Eds. Galil B. S., Clark P. F., Carlton J. T. ( Dordrecht: Springer), 607–624. doi: 10.1007/978-94-007-0591-3_21
Öztürk R. Ç., Terzi Y., Feyzioğlu A. M., Şahin A., Aydın M.(2020). Genetic Characterization of the Invasive Blue Crab, Callinectes sapidus (Rathbun 1896), in the Black Sea. Reg. Stud. Mar. Sci. 39, 101412. doi: 10.1016/j.rsma.2020.101412
Pascual M., Palero F., García-Merchán V. H., Macpherson E., Robainas-Barcia A., Mestres F., et al. (2016). Temporal and Spatial Genetic Differentiation in the Crab Liocarcinus depurator Across the Atlantic-Mediterranean Transition. Sci. Rep. 6 (1), 1–10. doi: 10.1038/srep29892
Patarnello T., Volckaert F. A. M. J., Castilho R. (2007). Pillars of Hercules: Is the Atlantic-Mediterranean Transition a Phylogeographical Break? Mol. Ecol. 16 (21), 4426–4444. doi: 10.1111/j.1365-294X.2007.03477.x
Peliz A., Dubert J., Marchesiello P., Teles-Machado A. (2007). Surface Circulation in the Gulf of Cadiz: Model and Mean Flow Structure. J. Geophys. Res. Oceans 112. doi: 10.1029/2007JC004159
Png-Gonzalez L., Papiol V., Balbín R., Cartes J. E., Carbonell A. (2021). Larvae of the Blue Crab Callinectes sapidus Rathbun 1896 (Decapoda: Brachyura: Portunidae) in the Balearic Archipelago (NW Mediterranean Sea). Mar. Biodivers. Rec. 14 (21), 1–5. doi: 10.1186/s41200-021-00217-5
Reuschel S., Cuesta J. A., Schubart C. D. (2010). Marine Biogeographic Boundaries and Human Introduction Along the European Coast Revealed by Phylogeography of the Prawn Palaemon elegans. Mol. Phylogenet. Evol. 55 (3), 765–775. doi: 10.1016/j.ympev.2010.03.021
Roman J. (2006). Diluting the Founder Effect: Cryptic Invasions Expand a Marine Invader’s Range. Proc. R. Soc B: Biol. Sci. 273 (1600), 2453–2459. doi: 10.1098/rspb.2006.3597
Sánchez-Leal R. F., Bellanco M. J., Fernández-Salas L. M., García-Lafuente J., Gasser-Rubinat M., González-Pola C., et al. (2017). The Mediterranean Overflow in the Gulf of Cadiz: A Rugged Journey. Sci. Adv. 3 (11), eaao0609.doi: 10.1126/sciadv.aao0609
Santos C. R. M. D. S., D’Incao F. (2004). Crustáceos no Cerrito Ariano Souza, Rio Grande, Rio Grande do Sul e Distribuição de Callinectes sapidus (Brachyura, Portunidae). Zoologia 94 (1), 73–76. doi: 10.1590/S0073-47212004000100013
Schubart C. D. (2009). “Mitochondrial DNA and Decapod Phylogenies: The Importance of Pseudogenes and Primer Optimization,” in Crustacean Issues 18: Decapod Crustacean Phylogenetics. Eds. Martin J. W., Crandall K. A., Felder D. L. (Boca Raton, Florida: Taylor & Francis/CRC Press), 47–65.
Taybi A. F., Mabrouki Y. (2020). The American Blue Crab Callinectes sapidus Rathbun 1896 (Crustacea: Decapoda: Portunidae) Is Rapidly Expanding Through the Mediterranean Coast of Morocco. Thalassas 36, 267–271. doi: 10.1007/s41208-020-00204-0
Vasconcelos P., Carvalho A. N., Piló D., Pereira F., Encarnação J., Gaspar M. B., et al. (2019). Recent and Consecutive Records of the Atlantic Blue Crab (Callinectes sapidus Rathbun 1896): Rapid Westward Expansion and Confirmed Establishment Along the Southern Coast of Portugal. Thalassas 35, 485–494. doi: 10.1007/s41208-019-00163-1
Windsor A. M., Moore M. K., Warner K. A., Stadig S. R., Deeds J. R. (2019). Evaluation of Variation Within the Barcode Region of Cytochrome C Oxidase I (COI) for the Detection of Commercial Callinectes sapidus Rathbun 1896 (Blue Crab) Products of non-US Origin. PeerJ 7, e7827. doi: 10.7717/peerj.7827
Keywords: Atlantic blue crab, invasive species, founder effect, gene flow, COI, haplotype
Citation: González-Ortegón E, Berger S, Encarnação J, Chairi H, Morais P, Teodósio MA, Oliva-Paterna FJ, Schubart CD and Cuesta JA (2022) Free Pass Through the Pillars of Hercules? Genetic and Historical Insights Into the Recent Expansion of the Atlantic Blue Crab Callinectes sapidus to the West and the East of the Strait of Gibraltar. Front. Mar. Sci. 9:918026. doi: 10.3389/fmars.2022.918026
Received: 11 April 2022; Accepted: 02 June 2022;
Published: 12 July 2022.
Edited by:
Anne Chenuil, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Raven Delaney Blakeway, United States Army Corps of Engineers, United StatesVictor Surugiu, Alexandru Ioan Cuza University, Romania
Copyright © 2022 González-Ortegón, Berger, Encarnação, Chairi, Morais, Teodósio, Oliva-Paterna, Schubart and Cuesta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enrique González-Ortegón, ZS5nb256YWxlei5vcnRlZ29uQGNzaWMuZXM=