Abstract
Codium tomentosum is a recently domesticated green macroalga, being currently cultured as an extractive species in integrated multi-trophic aquaculture (IMTA). Optimization of light requirements in outdoor systems must be achieved to increase the market value of cultivated algal biomass. The present study addresses the seasonal effects of light intensity and wavelength on productivity, pigment composition and epiphyte overgrowth in C. tomentosum cultured in a land-based IMTA system. Exposure to high light (non-filtered sun light) lead to higher net productivities in spring. However, non-filtered sun light caused significantly reduced productivities during summer when compared to filtered sun light (~ 4x lower irradiance levels). Furthermore, lower photosynthetic capacity (Fv/Fm) was observed in macroalgae cultured under high light during summer, indicating photoinhibition. Treatments with filtered sun light (low and red light) showed intermediate and more stable productivities. Epiphyte biomass was higher under high light and the lowest epiphyte overgrowth was recorded under red light. Concentrations of light-harvesting pigments were lower in summer than in spring, indicating a seasonal photoacclimation of macroalgae. An opposite seasonal trend was observed for accessory xanthophylls, as the main role of these pigments is photoprotection. Higher all-trans-neoxanthin and violaxanthin concentrations were found in high light than in low or red light treatments, confirming the important role of these biomolecules in the photoprotection of C. tomentosum. This study underlines the importance of controlling light to optimize algal growth outdoors and enhance the production of high-value compounds (i.e., pigments). Additionally, this practice can also reduce epiphyte overgrowth, thus enhancing the valorization of macroalgal biomass derived from C. tomentosum aquaculture.
Introduction
Macroalgae production currently accounts for more than half of global marine aquaculture landings and is the fastest growing sector in aquaculture (> 6% per annum), providing food and natural products across a range of industries, while supporting a variety of ecosystem services (Duarte et al., 2022). Traditionally valued in Southeast Asia, cultivated macroalgal biomass has gained increased attention in developing markets of western countries, not only given the growing perception of algae as healthy and sustainable foodstuffs, but also as important sources of bioactive compounds (Kim et al., 2017; Banach et al., 2020; van den Burg et al., 2021; Moreira et al., 2022). However, to supply dietary, cosmetic, and pharmaceutical markets, high levels of standardization and quality control of cultivated macroalgal biomass are required. This is particularly true when targeting high value biomass for high-end applications, given that the concentration and composition of bioactive compounds in macroalgae may fluctuate with season, location, and genetic background (Hafting et al., 2015). Fundamental research is thus required to understand how macroalgae perform under different cultivation conditions (e.g. light and nutrient availability), and how to mitigate competitors and pests (e.g. pathogens, grazers, epiphytes) (Hafting et al., 2015; Buschmann and Camus, 2019).
Codium tomentosum (Bryopsidales) is among the most recently domesticated green macroalgal species in aquaculture, having earned increased attention given its potential for a variety of applications. This algal species is highly valued as a gourmet ingredient in high-end cuisine (Pérez-Lloréns et al., 2018), as well as raw material in the cosmetic industry, in which algal extracts are used as the main moisturising agents in cosmetic products (Wang et al., 2015; Leandro et al., 2019). Other studies suggested C. tomentosum to be a potential source of essential polyunsaturated fatty acids (Rey et al., 2020), and valuable pigments, such as siphonaxanthin, that presents high antiangiogenic and antitumoral activities (Ganesan et al., 2010; Ganesan et al., 2011; Sugawara et al., 2014). This species is currently produced in a land-based integrated multitrophic system (ALGAplus Lda) where the effluent from a semi-intensive marine fish farm is used to cultivate organic certified macroalgal biomass. The company developed proprietary protocols for year-round production of C. tomentosum, involving an indoor hatchery phase under controlled temperature and light conditions and an outdoor phase during which juvenile thalli are grown in tumble culture (Marques et al., 2021).
Notwithstanding, optimization of C. tomentosum aquaculture is still ongoing to reduce operational costs, enhance the scaling-up and add value to cultured algal biomass. During the outdoor grow-out phase, C. tomentosum can be negatively affected by suboptimal light and unsuitable temperature conditions, while epiphytic algae (e.g. Ceramium sp., Ectocarpus sp., Ulva sp.) tend to overgrow Codium thalli, consequently decreasing product quality and increasing operational costs associated to daily husbandry and processing practices. In a previous study, Marques et al. (2021) observed significant differences in relative growth rates and pigment content of C. tomentosum propagules grown indoors when exposed to different photoperiod and light spectral regimes. These findings suggested that light requirement optimization should follow during grow-out phase outdoors to improve quality and enhance the standardization of cultivated biomass.
Light is arguably the most dynamic variable affecting land-based macroalgal aquaculture, being strongly influenced by the interplay between external (e.g. geographical location, photoperiod, season, weather) and internal factors (e.g. stocking density, biomass growth dynamics and culture method) (Magnusson et al., 2015). Light regimes affect the physiology of cultured macroalgae in different ways, with downstream implications that may include shifts in algal growth rates and/or bioactive molecule concentrations (Magnusson et al., 2015; Huang et al., 2021; Marques et al., 2021), as well as epiphyte overgrowth dynamics (Fletcher, 1995; Ward et al., 2020). In outdoor production systems, common techniques employed to control the amount of light reaching the algal biomass include the shading of tanks or the manipulation of cultivation depth and stocking densities. In turn, light quality can be manipulated artificially using coverings, or by adding external light sources with different spectral outputs (Hafting et al., 2015).
Here, we investigate how manipulating incident solar light for different irradiance levels (high-light and low-light) and light spectra (full spectra and red light) affect the growth, pigment composition and epiphyte overgrowth in C. tomentosum cultivated outdoors in a land-based IMTA system. To achieve this, experiments were performed over different seasons (spring and summer) using custom-built filter screens placed on top of culture tanks. Parameters such as growth rates, maximum quantum yields of photosynthesis, pigment content, and epiphyte overgrowth were assessed and discussed in line with the current efforts to refine existing cultivation techniques for this green macroalga.
Materials and Methods
Biological Material and Culturing Conditions
Codium tomentosum Stackhouse, 1797 biomass was purchased from ALGAplus. For acclimation purposes, macroalgae were maintained for 2 weeks on a dedicated experimental setup of 100-L cylindrical polyethylene tanks with constant water inflow, for a total of eight daily volume renovations (33.33 L h-1), in which algae were kept under free-floating (tumble culture) conditions through strong aeration. The system was provided with seawater from ALGAplus earthen ponds for fish production, after a series of mechanical filtration steps (a drum filter, a sand filter, and a double lycra fabric layer) that reduce suspended solid particles (> 40 µm) in inflowing water.
Light Manipulation
The experiment was designed to test the effects of light quantity (irradiance) and quality (spectra) in the performance of C. tomentosum in outdoor culture conditions. A control and two light treatments were used: control high-light (HL – consisting of full spectrum unfiltered sunlight), low-light (LL – sunlight filtered through a neutral filter – approximately ¼ of total irradiance was obtained without compromising light spectrum), and red-light (RL – filtered sunlight for the red band region of the light spectrum – obtained by filtering sun light through a red filter). Neutral (209 0.3ND) and red (106 Primary Red) filters were obtained from Lee Filters (Hampshire, UK). Light spectra were measured using a Flame model Ocean Optics spectrometer (wavelength range 190–1100 nm) (Ocean Insight, Florida, USA) (Supplementary Figure S1).
The experimental setup consisted of a total of 15 round polyethylene 100-L tanks, with 5 replicate tanks per light condition (Supplementary Figure S2). The filters were fixed between two circular crystal acrylic screens (3 mm width), with 0.8 m diameter, supported by 5 equally distributed metal rods. The screens with the filters were placed 0.16 m above each tank to facilitate airflow and minimize eventual greenhouse effects. The light filtration structures for LL and RL conditions, sustained 2 sheets of Lee 0.3 ND neutral filters, and 1 sheet of Lee primary red filter respectively, fixed between the two crystal acrylic screens. In control HL, the structure consisted of the acrylic screens devoid of any filter screen.
Abiotic Parameters
Temperature, salinity, pH and quantum irradiance were measured 2 days before a designated weekly sampling day for C. tomentosum biomass. Measurements of abiotic parameters took place at solar noon over the course of the experiments. Water parameter measurements were performed in all the culture tanks, using a SenTix® pH electrode for pH and temperature, and a TetraCon® 325 conductivity measuring cell for salinity, both connected to an universal multi-parameter portable meter ProfiLine pH/Cond 3320 (WTW, Weilheim, Germany). Irradiance was quantified underwater, inside the culture tanks at surface and at the bottom (~ 0.35 m depth) of each tank, using a LI-193 Spherical Quantum Sensor connected to LI-250A Light Meter (LI-COR biosciences, Lincoln, USA). Irradiance measurements were taken at the center of each tank for all the replicates and values registered represent a 15 s average irradiance of approximately 60 instant readings.
Macroalgal Growth
The experiment ran for 5 weeks in each season during summer (July–August 2020) and spring (March–April 2021). At the beginning of each experiment, C. tomentosum was distributed among the polyethylene tanks at a density of ~ 1 kg fresh weight (FW) per tank. The cultures remained under free-floating, tumble culture conditions with strong aeration, keeping the algae in constant movement through air diffusion to assure homogeneous exposure of the seaweed to incident irradiance.
Changes in macroalgal biomass were quantified on a weekly basis. The macroalgae from each tank were transferred to separate plastic baskets and centrifuged, being drained to constant weight and re-stocked in the respective tanks to the initial density. Every week the entire experimental setup was thoroughly cleaned and reassembled. Macroalgae FW was quantified using a scale (sensibility ± 1 g). Before the calculation of growth parameters, FW was corrected for the weight of overgrowing epiphytes:
where, FWt is the fresh weight of Codium after centrifugation, rFWEpi is the ratio of fresh weight of epiphytes estimated for each tank (see below for details on the quantification of epiphyte biomass).
Then, net productivity (P) of C. tomentosum was calculated according to the equation:
where FWf is the final fresh weight, FWi is initial fresh weight (grams), V is culture tank volume (liters), and t is the time in culture (weeks).
Before returning C. tomentosum to culture tanks, a quick and general cleaning process took place. The algae were stirred and rinsed with seawater twice, to remove detached epiphytes, particulate matter, and biofilms from the algal surface.
Epiphyte Overgrowth
During C. tomentosum weekly harvest and processing, 30 ± 1 g were sampled from each replicate tank for epiphyte biomass quantification. Thereafter, one at a time, Codium samples were transferred to white plastic trays with seawater in which epiphytes were removed by hand. As the process of epiphyte removal was concluded (Supplementary Figure S3), C. tomentosum biomass was discarded, and the remaining epiphytes were recovered by filtering the sample through a nylon net (500 µm), and by transferring to 50 mL Falcon tubes with seawater. In the laboratory, samples were recovered by filtering the epiphyte suspensions through a nylon mesh (500 µm), and by successively pressing samples between two dry sheets of paper towel four times to remove excess water. Epiphytes were quantified and expressed as both fresh weight (FW) and dry weight (DW), after drying at 60°C for 72 h.
Chlorophyll Fluorescence
Variable chlorophyll (Chl) a fluorescence was measured the day before macroalgal growth quantification, every week during both seasons, using an Imaging-PAM fluorometer Mini version (Walz, Effeltrich, Germany). Maximum quantum yield of photosystem (PS) II (Fv/Fm) was determined by calculating (Fm – Fo)/Fm, where Fm and Fo are the maximum and the minimum fluorescence of 15 min dark-adapted samples, respectively (Murchie and Lawson, 2013). Numerical values of variable Chl fluorescence parameters were extracted from the digital images using the imaging system software (Imaging Win, Heinz Walz GmbH, Germany) by selecting areas of interest (AOI). Three circular AOI per replicate were chosen, defining regions of good camera focus close to the center of the display.
Photosynthetic Pigments Analysis
At the end of summer and spring experimental periods, macroalgal material was flash frozen in liquid nitrogen and freeze-dried prior to photosynthetic pigment analysis. Pigment quantification by High Performance Liquid Chromatography (HPLC) followed the C18 method described by Mendes et al. (2007). For every extraction, the amount of lyophilized sample used followed the ratio of 5 ± 0.1 mg per 1 mL of extraction solution (95% cold buffered methanol with 2% ammonium acetate). Samples were ground with a plastic rod and sonicated for 1 min. Samples were then transferred to –20°C for 20 min in the dark. Extracts were filtered through 0.2 µm PTFE membrane filters and immediately injected into a Prominence i –LC 2030C HPLC system (Shimadzu, Kyoto, Japan) with a photodiode array detector. Chromatographic separation was carried out using a Supelcosil C18 column (250 mm length; 4.6 mm diameter; 5 µm particles; Sigma-Aldrich, St. Louis, MO, USA) for reverse phase chromatography and a 35 min elution program. The solvent gradient followed Kraay et al. (1992), with an injection volume of 50 µL and a flow rate of 0.6 mL min-1. Pigments were identified from absorbance spectra and retention times and concentrations calculated from the signals in the photodiode array detector. Calibration curves were constructed with pure crystalline standards from DHI (Hørsolm, Denmark). Pigments were expressed in µmol g dw-1.
Statistical Analysis
Significant differences in productivity, epiphyte biomass, pigment concentrations, Fv/Fm and all the abiotic parameters for effects of light treatment and season were tested using two-way ANOVA, after checking for homogeneity of variances (Levene’s Test) and normality (Shapiro-Wilk’s test). Multiple comparisons within each season were performed with Tukey’s test. Statistical analysis was performed using SigmaPlot 14 for Windows (Systat Software, Germany).
Results
Several water parameters were monitored in C. tomentosum production tanks: pH, temperature, and salinity (Table 1). Regarding both temperature and salinity, significantly higher values (in all cases p < 0.001) were observed during summer, but no significant differences were observed between any of the light conditions (HL, LL and RL). Average water temperature was around 25°C in summer and 19°C in spring (Table 1). Values for pH were significantly higher (in all cases p < 0.001) under HL in both summer and spring (8.66 and 8.31, respectively) when compared to LL and RL treatments (values ranging from 8.41 and 8.05). Regarding LL and RL, no statistical differences were observed for pH. Surface and bottom irradiance values recorded in C. tomentosum production tanks are shown in Table 2. As projected, a significantly higher irradiance (in all cases p < 0.001) was observed under the non-filtered HL, when compared to LL and RL treatments. Under control HL, surface irradiance at solar noon reached average values close to 2,000 µmol photons m-2 s-1 in summer and 1,600 µmol photons m-2 s-1 in spring. Surface and bottom irradiances were not significantly different between LL and RL in either summer or spring, with average values at noon ranging from 568 to 525 µmol photons m-2 s-1 (Table 2).
Table 1
| Light treatment | ||||
|---|---|---|---|---|
| HL | LL | RL | ||
| pH | Summer | 8.66 ± 0.06a | 8.41 ± 0.02b | 8.40 ± 0.06b |
| Spring | 8.31 ± 0.05a | 8.05 ± 0.06b | 8.05 ± 0.08b | |
| Temperature | Summer | 25.87 ± 0.34a | 25.19 ± 0.47a | 25.21 ± 0.54a |
| (°C) | Spring | 19.68 ± 0.34a | 19.33 ± 0.58a | 19.23 ± 0.32a |
| Salinity | Summer | 34.27 ± 0.02a | 34.28 ± 0.01a | 34.28 ± 0.01a |
| (‰) | Spring | 29.32 ± 0.03a | 29.31 ± 0.05a | 29.31 ± 0.02a |
Water parameters measured at solar noon (mean ± SD, n = 5) over a 5-week culture period during summer and spring, under control high-light (HL), low-light (LL) and red-light (RL).
Different superscript letters indicate significant differences between light treatments in each season, obtained from Tukey’s multiple comparison tests (p < 0.05).
Table 2
| Light treatment | ||||
|---|---|---|---|---|
| HL | LL | RL | ||
| Surface irradiance | Summer | 1929 ± 67a | 553 ± 11b | 568 ± 12b |
| (μmol photons m−2 s−1) | Spring | 1668 ± 25a | 534 ± 33b | 525 ± 31b |
| Bottom irradiance | Summer | 1110 ± 99a | 354 ± 26b | 365 ± 20b |
| (μmol photons m−2 s−1) | Spring | 655 ± 39a | 262 ± 52b | 241 ± 30b |
| Fv/Fm | Summer | 0.598 ± 0.040a | 0.685 ± 0.006b | 0.682 ± 0.013b |
| Spring | 0.641 ± 0.035a | 0.671 ± 0.027a | 0.674 ± 0.033a | |
Surface and bottom irradiance levels at solar noon and maximum yield of photosynthesis (Fv/Fm) of Codium tomentosum (mean ± SD, n = 5) over a 5-week culture period during summer and spring, under control high-light (HL), low-light (LL) and red-light (RL).
Different superscript letters indicate significant differences between light treatments in each season, obtained from Tukey’s multiple comparison tests (p < 0.05).
Net productivities of C. tomentosum along the 5-week culture periods in summer and spring are represented in Figure 1. The highest productivity (> 4 g L-1 week-1) was registered in spring after 5 weeks under HL. The lowest value (– 0.65 g L-1 week-1) was also observed in HL, but during the summer period and after 3 weeks in culture. Treatments with filtered sun light (LL and RL) showed intermediate and more stable productivities, with higher values during spring (Figure 1). Net productivities were significantly lower (p < 0.001) in summer than in spring (Figure 2). In summer, significantly higher productivities were registered under LL and RL when compared to HL (p = 0.009 and p = 0.004), whereas no differences were observed between LL and RL treatments (p = 0.938). In spring, significantly higher productivities were registered in HL compared to LL (p = 0.022), whereas no significant differences were observed between control HL and RL treatment (p = 0.085). Differences in productivities in spring were also non-significant (p = 0.799) between LL and RL treatments (Figure 2).
Figure 1
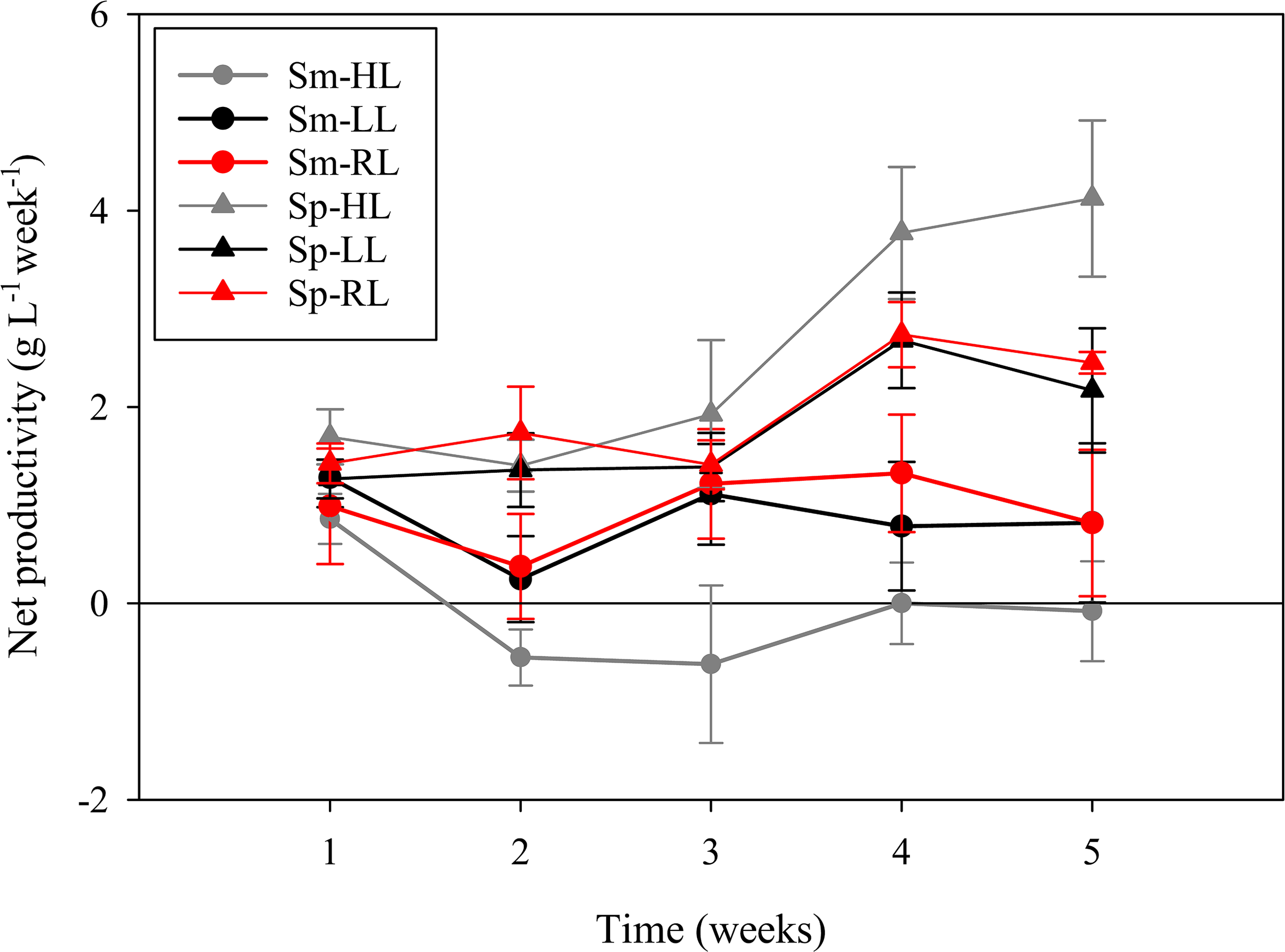
Net productivity of Codium tomentosum (mean ± SD, n = 5) over a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL).
Figure 2
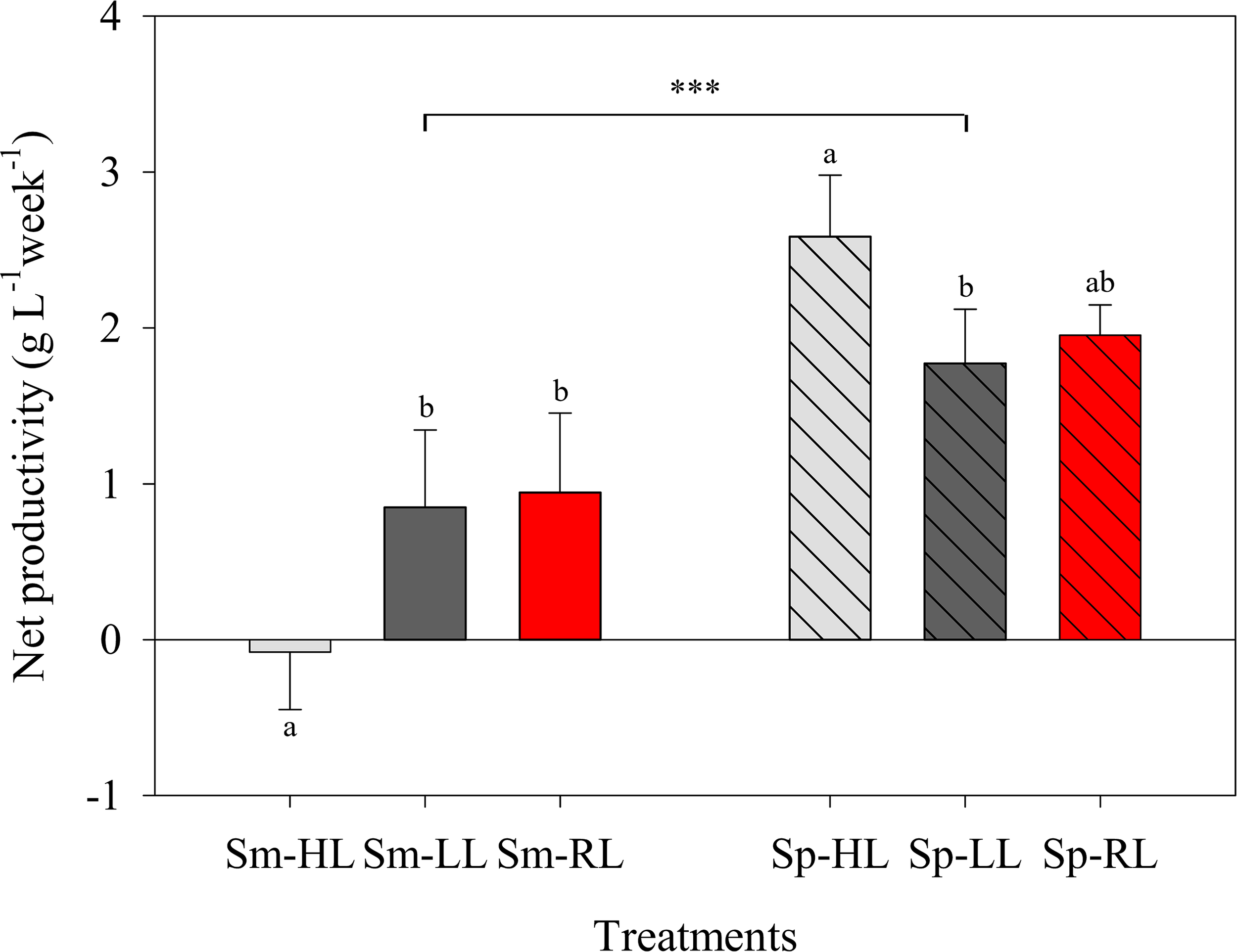
Average net productivity of Codium tomentosum (mean ± SD, n = 5) over a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL). Differences between seasons were significant at p < 0.001 (triple asterisk). Different letters indicate significant differences between light treatments within each season at p < 0.05.
Epiphytic algal biomass overgrowing C. tomentosum was monitored and quantified during the 5-week culture periods. The dominant epiphytic algae were Ectocarpus sp. (Ochrophyta), Ceramium sp., Polysiphonia sp. (Rhodophyta), and Ulva sp. (Chlorophyta). Occasionally, other algae such as Bryopsis sp. (Chlorophyta) and Porphyra sp. (Rhodophyta) were also observed, but always in very low amounts. The biomass of occurring algal epiphytes was not significantly different (p = 0.136) between summer and spring (Figure 3). However, epiphyte overgrowth changed between seasons with a clear dominance of Ectocarpus sp. during the summer period. In both seasons, epiphyte biomass was significantly higher under HL conditions (in all cases p < 0.05). Although the lowest epiphyte overgrowing biomass was recorded under RL, no significant differences in epiphyte biomass were found between LL and RL conditions (Figure 3).
Figure 3
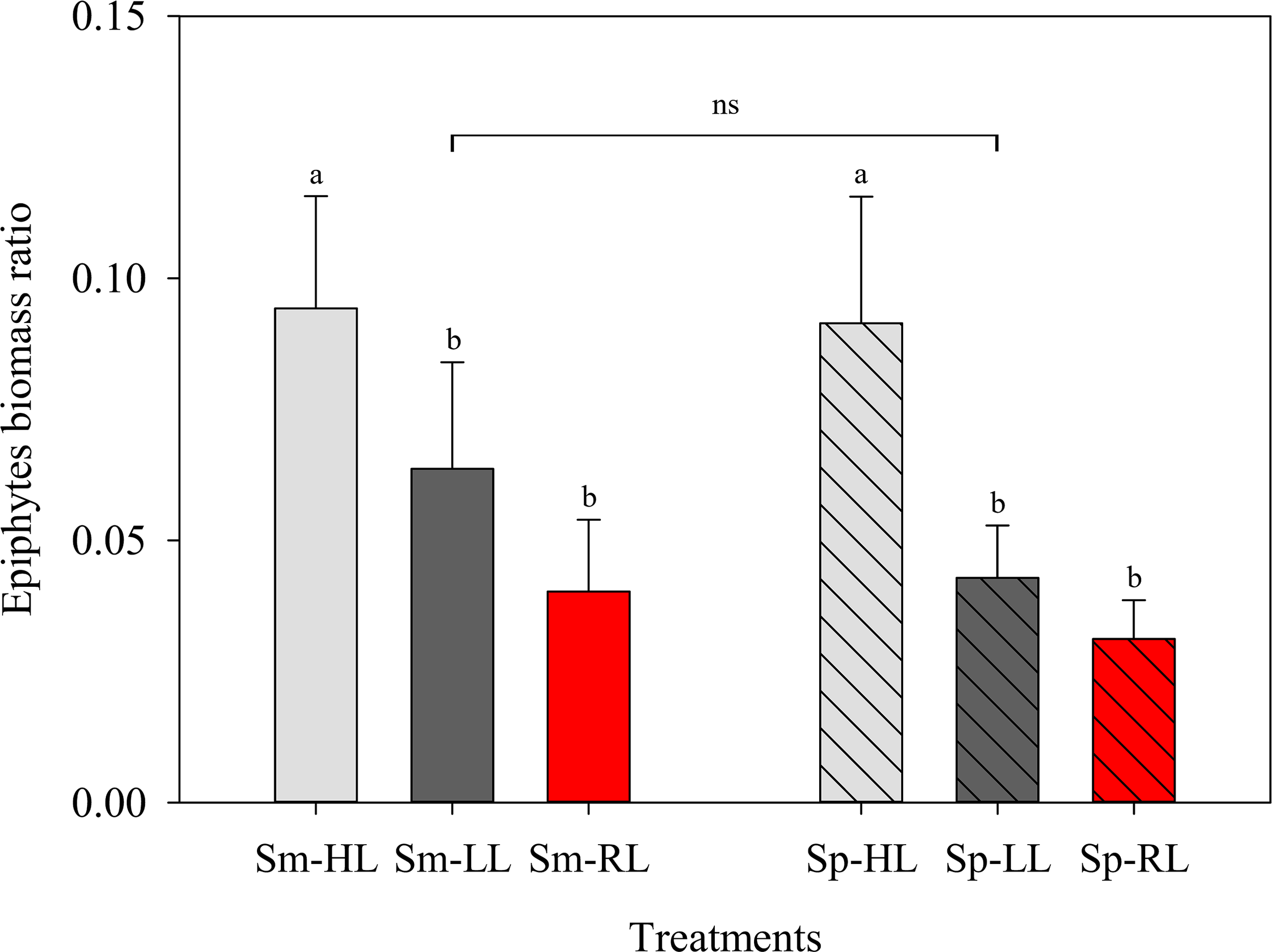
Ratio of epiphytic algal biomass (DW) per cultured Codium tomentosum biomass (DW) (mean ± SD, n = 5) over a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL). Differences between seasons were non-significant (ns). Different letters indicate significant differences between light treatments within each season at p < 0.05.
Season and light treatment significantly affected maximum photosynthetic capacity (Fv/Fm) of dark-adapted C. tomentosum. In summer, a significantly lower maximum photosynthetic capacity was found in HL, than in both LL and RL grown macroalgae (in both cases p < 0.001, Table 2). In control HL, Fv/Fm reached values of 0.598, compared to 0.685 and 0.682 under LL and RL treatments.
Pigments identified in C. tomentosum by HPLC were categorized in three main groups: light-harvesting pigments – siphonaxanthin (Siph), 9′-cis-neoxanthin (c-Neo), siphonaxanthin dodecenoate (Siph-do), chlorophyll b (Chlb) and chlorophyll a (Chla) (Figure 4); accessory xanthophylls – all-trans-neoxanthin (t-Neo), violaxanthin (Viola), antheraxanthin (Anth) and zeaxanthin (Zea) (Figure 5); and carotenes – ϵ,ϵ-carotene (ϵϵ-Car), β,ϵ-carotene (βϵ-Car), β,β-carotene (ββ-Car) (Figure 6). Main light harvesting pigments were Chla and Chlb and the carotenoid Siph. The concentrations of all light-harvesting pigments were significantly lower in summer than in spring (in all cases p < 0.001) (Figure 4). A different trend was observed for some accessory xanthophylls, with significantly higher concentrations of Anth and Zea being recorded in summer than in spring (Figure 5).
Figure 4
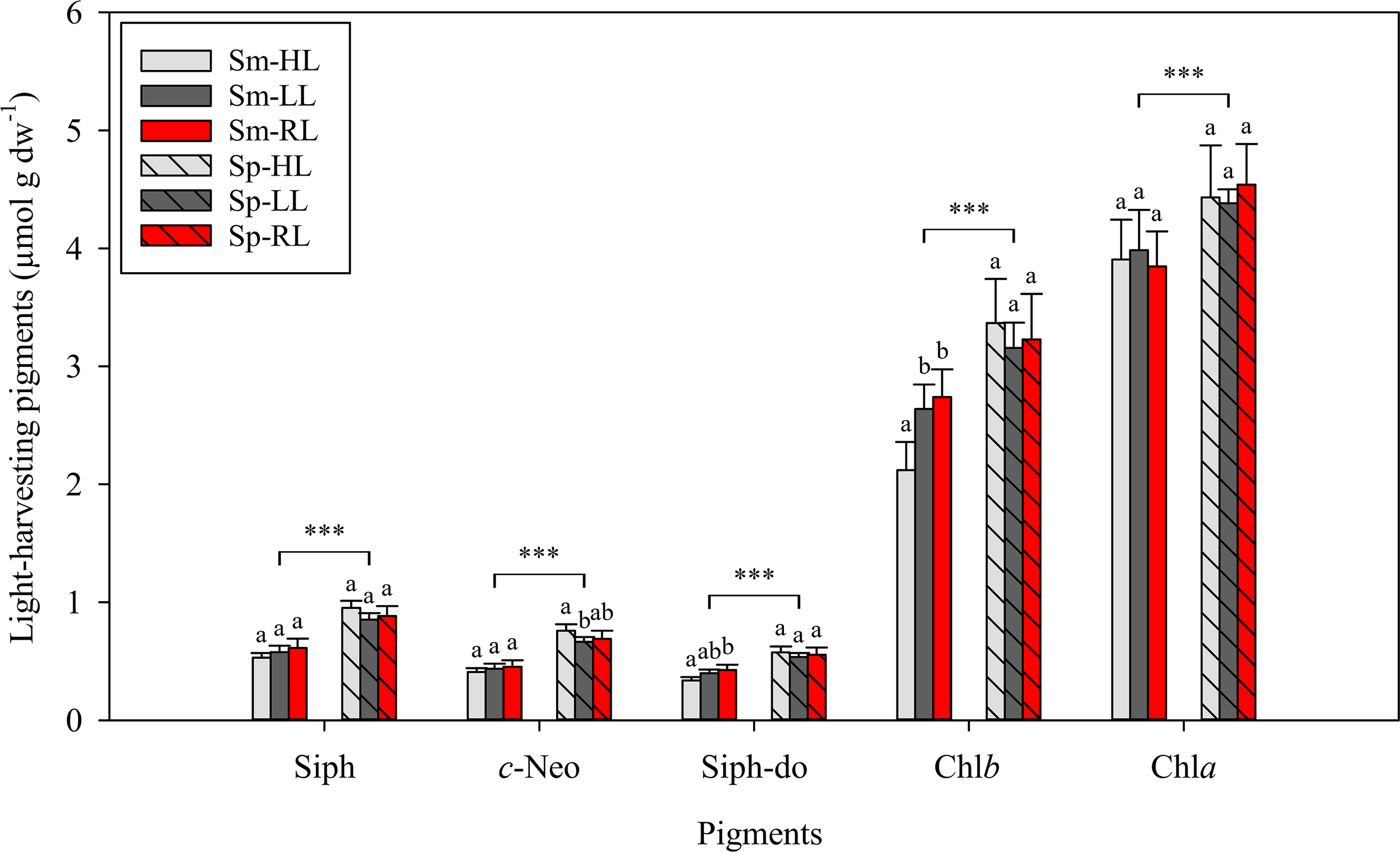
Concentrations of light harvesting pigments in Codium tomentosum (mean ± SD, n = 5) after a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL). Siph – siphonaxanthin; c-Neo – 9′-cis-neoxanthin; Siph-do – siphonaxanthin dodecenoate; Chlb, chlorophyll b; Chla, chlorophyll a. Differences between seasons were significant at p < 0.001 (triple asterisk). Different letters indicate significant differences between light treatments within each season at p < 0.05.
Figure 5
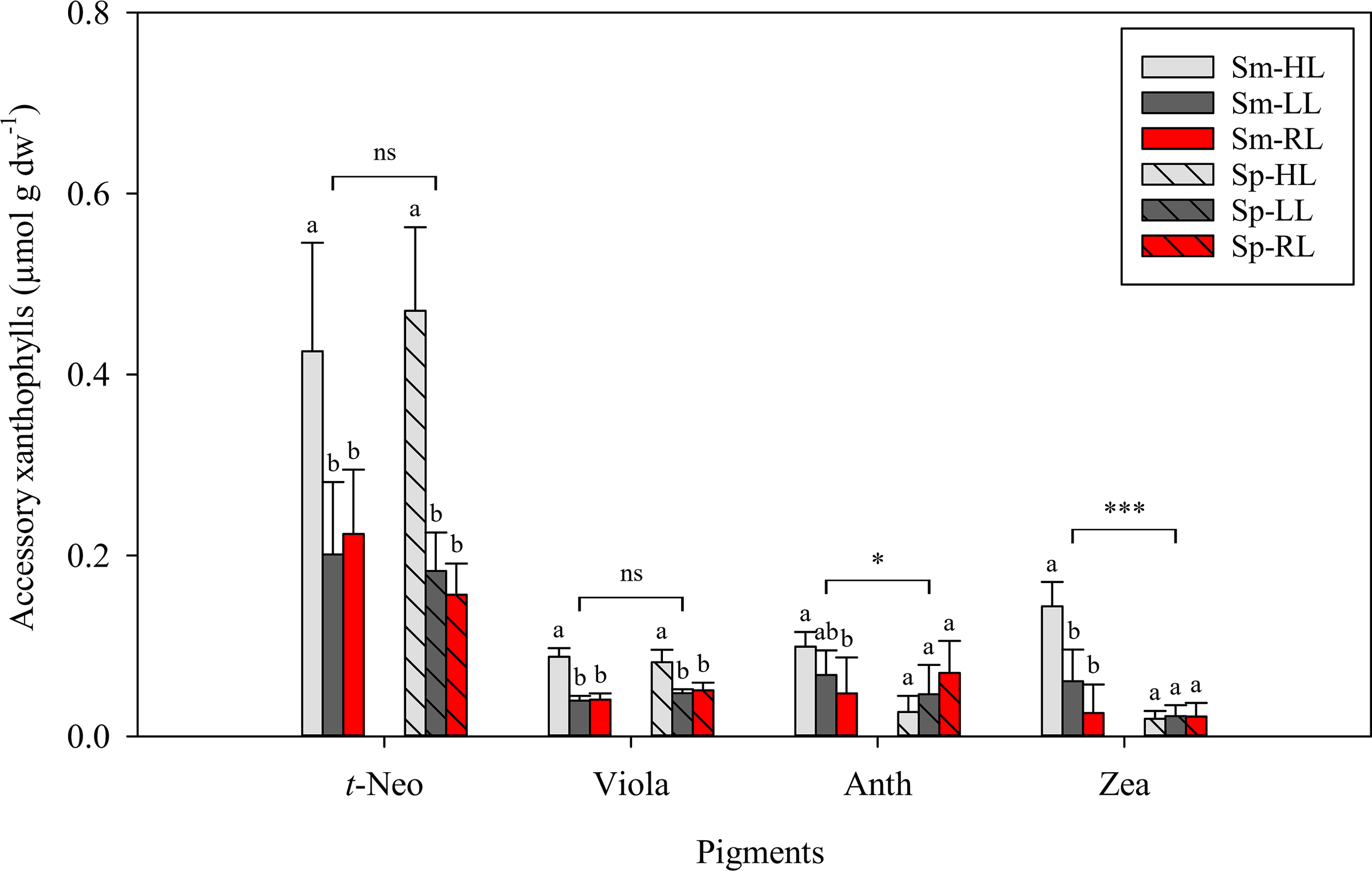
Concentrations of accessory xanthophylls in Codium tomentosum (mean ± SD, n = 5) after a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL). t-Neo – all-trans-neoxanthin; Viola – violaxanthin; Anth – antheraxanthin; Zea – zeaxanthin. Differences between seasons were non-significant (ns), significant at p < 0.05 (single asterisk) or significant at p < 0.001 (triple asterisk). Different letters indicate significant differences between light treatments within each season at p < 0.05.
Figure 6
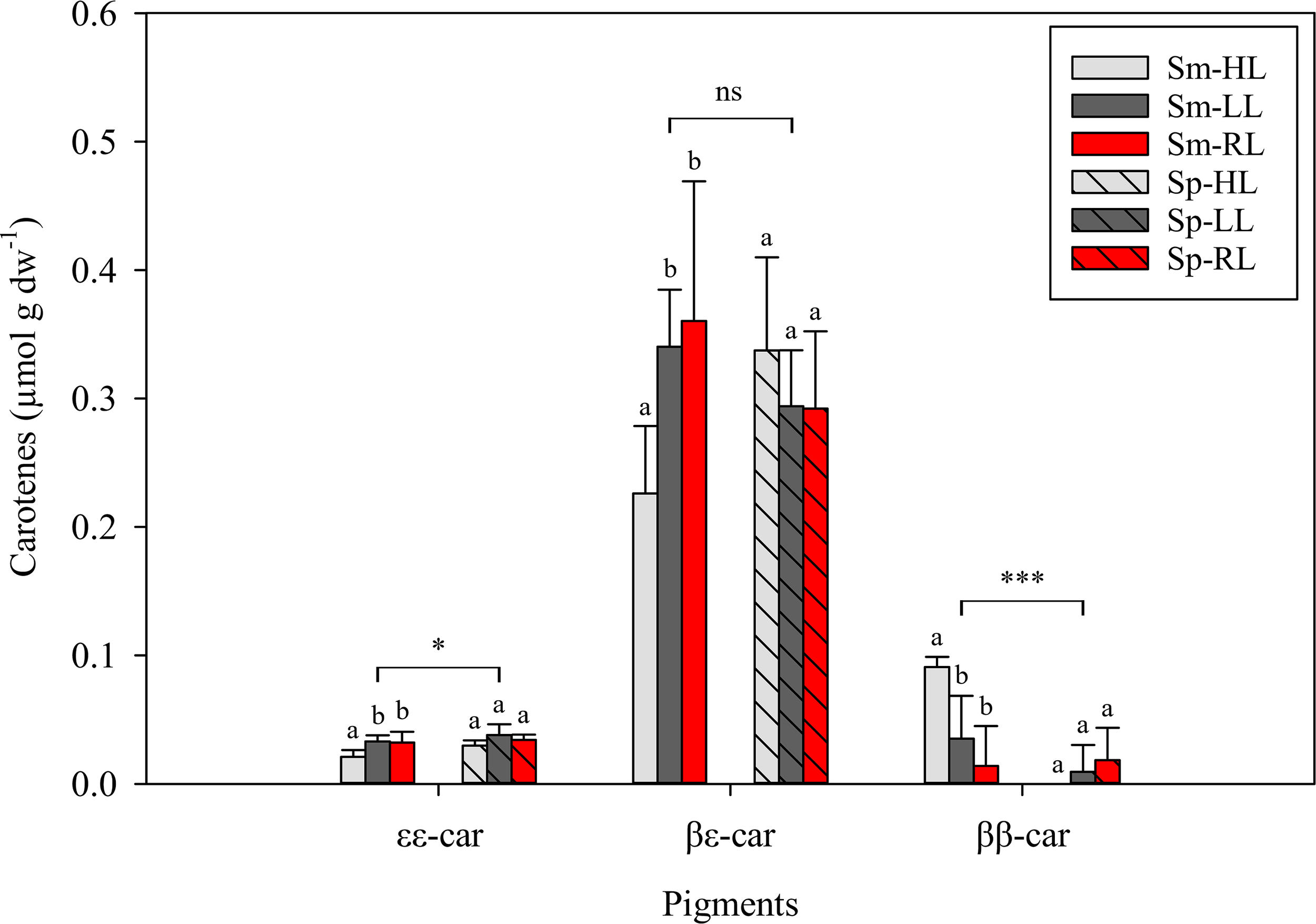
Concentrations of carotenes in Codium tomentosum (mean ± SD, n = 5) after a 5-week culture period during summer (Sm) and spring (Sp), under control high-light (HL), low-light (LL) and red-light (RL). ϵϵ-Car – ϵ,ϵ-carotene; βϵ-Car – β,ϵ-carotene; ββ-Car – β,β-carotene. Differences between seasons were non-significant (ns), significant at p < 0.05 (single asterisk) or significant at p < 0.001 (triple asterisk). Different letters indicate significant differences between light treatments within each season at p < 0.05.
Within each season, differences between light conditions were minor for main light-harvesting pigments, although significantly higher concentrations of Chlb were observed in summer for the lower irradiance treatments (LL and RL) than under control HL. Regarding accessory xanthophylls, significantly higher t-Neo and Viola were observed in HL than in LL and RL treatments within each season (in all cases p < 0.001). The same trend was observed for Zea, but only in the summer (Figure 5). The main carotene found in C. tomentosum was βϵ-Car, compared to very low concentrations of ϵϵ-Car and ββ-Car (Figure 6). No significant differences between summer and spring were observed for the main βϵ-Car (p = 0.968). In summer, significantly higher concentrations of βϵ-Car and ϵϵ-Car were found in both lower light treatments (LL and RL) than under HL (in all cases p < 0.05). Concentrations of ββ-Car in summer followed an opposite trend, with significantly higher concentrations in control HL than in LL and RL (in both cases p < 0.01). No significant differences in any of the pigment concentrations were observed between LL and RL treatments.
Discussion
Light absorption is crucial to drive photochemical reactions of photosynthesis and, therefore, light availability is a major parameter affecting macroalgal growth. On the other hand, high irradiance leads to the formation of reactive oxygen species (ROS) that damage the photosynthetic apparatus, hampering growth and reducing productivity (Murata et al., 2007; Takahashi and Badger, 2011). In the present study, C. tomentosum productivity in a land-based IMTA system was differently affected by light treatments depending on the season: in spring, higher productivities were registered in control high light (exposure to full sun light) than in lower light treatments (low and red filtered sun light), whereas the opposite was found in summer. Maximum photosynthetic capacity (Fv/Fm) in summer was lower in high light than in low and red filtered sun light cultured macroalgae, indicating photoinhibition of C. tomentosum photosynthetic apparatus in the control conditions. Irradiance at noon in HL control during the summer period averaged 2,000 µmol photons m-2 s-1, whereas values registered under the LL and RL treatments were around 550 µmol photons m-2 s-1. In an out-growing set-up using 100 m long horizontal ropes held at different water depths, Hwang et al. (2008) found that growth rates of Codium fragile were significantly higher at 1 m depth. Photosynthetic active radiation was about 630 µmol photons m-2 s-1 at this depth of the water column, corresponding to a light reduction of around 70% compared to that at water surface. These results stress the importance of controlling irradiance in macroalgal aquaculture systems in specific periods to avoid photoinhibition and considerable financial impact due to reduced productivity. Excess irradiance, however, may not be the sole responsible for the reduction of C. tomentosum productivity observed during summer. Hanisak (1979) reported a decrease in growth rate of cultured C. fragile ssp. tomentosoides for the combination of higher temperature and higher irradiance. In our study, water temperatures measured at noon exceeded 25°C in summer, whereas values were around 19°C in spring. Hence, the combination of light and thermal stress may have played a role in the lower productivities recorded for C. tomentosum grown during summer under full sunlight.
In this study, not only photon irradiance was manipulated. Spectral composition of available light was also shifted, as it too can affect macroalgal growth. Figueroa et al. (1995) cultivated the red macroalga Porphyra umbilicalis in the laboratory using fluorescent lamps and red or blue Plexiglas filters. The authors observed that red light favored growth by enhancing thallus expansion, cell division, and polysaccharide production compared to blue light. Marques et al. (2021) reported higher relative growth rates under red light than blue light in C. tomentosum grown indoors at laboratory nursery conditions. This finding was rather surprising, as C. tomentosum contains light harvesting complexes with xanthophylls siphonaxanthin and siphonaxanthin dodecenoate, as well as a higher proportion of chlorophyll b compared to most algae and plants. The pigment composition of Codium light harvesting complexes and other siphonous green algae makes them more efficient in exploiting the spectral region of blue-green light (Anderson, 1983; Benson and Cobb, 1983; Wang et al., 2013). In our study, similar productivity rates were found for C. tomentosum grown under filtered lower light treatments, using either neutral or red filters. Although comparisons between laboratory and outdoor experiments are questionable, results seem to suggest that red light is at least equally efficient in driving photosynthesis of C. tomentosum.
Consistently higher epiphyte biomass was found at high light in both spring and summer, indicating that full sun light not only affects C. tomentosum productivity, but also induces epiphyte proliferation. Growth of epiphytes is one of the most important parameters determining the market-value of macroalgal biomass (Ingle et al., 2018; Sahu et al., 2020). Furthermore, cleaning of epiphytes by hand is extremely labour-intensive and not feasible for commercial scale production. The lowest epiphyte occurrences recorded under red light in C. tomentosum indicates that spectral manipulation may be used as a tool to control algal epiphytes in macroalgal aquaculture. The use of filters (neutral or wavelength-specific) seems to be a relatively inexpensive approach to control epiphyte growth in C. tomentosum, thus contributing to improve the valorization of its biomass.
Macroalgae can adapt pigment composition in their photosynthetic apparatus, thus optimizing their light harvesting properties. A clear seasonal photoacclimation trend was observed, with higher concentrations of all light-harvesting pigments being recorded during spring. Natural pigments may exhibit various beneficial biological activities, such as antioxidant, anti-tumoral, anti-inflammatory and neuroprotective activities (Pangestuti and Kim, 2011). The carotenoid siphonaxanthin, characteristic of siphonous green algae, has been used as an anti-tumoral agent inhibiting the proliferation of cancer cells through the induction of apoptosis (Ganesan et al., 2011; Sugawara et al., 2014). In our study, we show a clear seasonal difference in the concentration of siphonaxanthin in C. tomentosum, a much valuable information in the case of farmers targeting the production of this high-value compound for biomedical applications.
Light spectra (white vs red light) had no impact on the pigment profile of C. tomentosum, but light intensity (high vs low irradiance) significantly affected pigment composition of this macroalga. Significantly higher violaxanthin and all-trans-neoxanthin concentrations were found in control HL than in LL and RL treatments. Giossi et al. (2021) showed that high light had a strong effect on the pigment composition of siphonous green algae triggering a time-dependent accumulation of violaxanthin and all-trans-neoxanthin. These changes in pigment composition can cause a rearrangement in light harvesting complexes, thus influencing the amount of energy that is transferred to reaction centers (Uragami et al., 2014; Giossi et al., 2020; Giossi et al., 2021). By reducing damages to the photosynthetic apparatus under increased irradiance, this photoprotection mechanism can enhance the fitness of Bryopsidales algae (Giossi et al., 2021).
Previous studies have shown that Bryopsidales lack a ubiquitous photoprotection mechanism in plants and most algae, the xanthophyll cycle (e.g., Raniello et al., 2004; Cruz et al., 2015; Christa et al., 2017). In green algae, the xanthophyll cycle comprises the sequential de-epoxidation of the pigments violaxanthin to antheraxanthin and zeaxanthin under high light, allowing excess energy dissipation as heat (Goss and Jakob, 2010). The reverse reactions occur under low light. Cruz et al. (2015) showed that C. tomentosum does not accumulate antheraxanthin, nor zeaxanthin, under high light and, therefore, lack this rapid photoprotective mechanism. Although zeaxanthin was detected in C. tomentosum and concentrations increased in summer under high light, a concomitant reduction of violaxanthin was not observed. Instead, several carotenoids (β,β-carotene, zeaxanthin, antheraxanthin, violaxanthin and all-trans-neoxanthin) presented increased concentrations as a result of an activation of the β-branch carotenoid biosynthetic pathway.
In conclusion, this study underlines the importance of optimizing light conditions for optimal growth of macroalgae in aquaculture systems, in particular the need for controlling high irradiance levels that can compromise biomass production. The occurrence of this light stress is seasonal and site-specific and can be controlled cost-effectively by using neutral or wavelength-specific filters as the ones used in our study. Additionally, the use of these light filters can reduce epiphyte overgrowth that decrease the market-value of macroalgal biomass, therefore contributing to the valorization of farmed macroalgae. The manipulation of irradiance also affects the biochemical composition of cultured macroalgae, which may also positively impact its market-value. We show that photosynthetic pigment composition change both seasonally and within the different light conditions tested and that the fine-tuning of light can be performed outdoors to optimize the production of high-value algal compounds. In the specific case of C. tomentosum, we recommend the use of red filters in periods of high irradiance to increase productivity and reduce epiphyte overgrowth, contributing to the valorization of this emerging aquaculture species.
Funding
This work was support by R&D project CtLight (PTDC/BIA-FBT/30979/2017) funded by the European Regional Development Fund (ERDF), through COMPETE2020 - Programa Operacional Competitividade e Internacionalização (POCI), and by national funds (OE), through Fundação para a Ciência e a Tecnologia (FCT/MCTES). Thanks are also due to FCT/MCTES for the financial support to CESAM (UIDP/50017/2020+UIDB/50017/2020+LA/P/0094/2020). S.C. was funded by the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. 949880) and by FCT/MCTES (grant 2020.03278.CEECIND). PC was funded by FCT/MCTES (grant CEECIND/01434/2018).
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
Conceptualization: RM, RC, SC, PC; Methodology: All authors; Data analysis: RM, PC; Investigation: RM, AM, SC; Supervision: PC; Project administration: PC; Funding acquisition: PC. First manuscript draft: RM. All authors contributed to manuscript revision, read, and approved the submitted version.
Acknowledgments
We thank Helena Abreu and Rui Pereira from ALGAplus for the use of company facilities. We also thank Diana Lopes for technical assistance and Francisca Brito and Rui Pereira for insights on the design of the acrylic screens.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.906332/full#supplementary-material
References
1
AndersonJ. M. (1983). Chlorophyll-Protein Complexes of a Codium Species, Including a Light-Harvesting Siphonaxanthin-Chlorophyll a b-Protein Complex, an Evolutionary Relic of Some Chlorophyta. Biochim. Biophys. Acta Bioenerg.724 (3), 370–380. doi: 10.1016/0005-2728(83)90096-8
2
BanachJ. L.Hoek-van den HilE. F.Fels-KlerxH. J. (2020). Food Safety Hazards in the European Seaweed Chain. Compr. Rev. Food Sci. Food Saf.19 (2), 332–364. doi: 10.1111/1541-4337.12523
3
BensonE. E.CobbA. H. (1983). Pigment/protein Complexes of the Intertidal Alga Codium fragile (Suringar) Hariot. New Phytol.95 (4), 581–594. doi: 10.1111/j.1469-8137.1983.tb03523.x
4
BuschmannA. H.CamusC. (2019). An Introduction to Farming and Biomass Utilisation of Marine Macroalgae. Phycologia58 (5), 443–445. doi: 10.1080/00318884.2019.1638149
5
ChristaG.CruzS.JahnsP.de VriesJ.CartaxanaP.EstevesA. C.et al. (2017). Photoprotection in a Monophyletic Branch of Chlorophyte Algae is Independent of Energy-Dependent Quenching (qE). New Phytol.214 (3), 1132–1144. doi: 10.1111/nph.14435
6
CruzS.CartaxanaP.NewcomerR.DionísioG.CaladoR.SerôdioJ.et al. (2015). Photoprotection in Sequestered Plastids of Sea Slugs and Respective Algal Sources. Sci. Rep.5, 7904. doi: 10.1038/srep07904
7
DuarteC. M.BruhnA.Krause-JensenD. (2022). A Seaweed Aquaculture Imperative to Meet Global Sustainability Targets. Nat. Sustain5, 185–193. doi: 10.1038/s41893-021-00773-9
8
FigueroaF. L.AguileraJ.NiellX. (1995). Red and Blue Light Regulation of Growth and Photosynthetic Metabolism in Porphyra umbilicalis (Bangiales, Rhodophyta). Eur. J. Phycol.30 (1), 11–18. doi: 10.1080/09670269500650761
9
FletcherR. L. (1995). Epiphytism and Fouling in Gracilaria Cultivation: An Overview. J. Appl. Phycol.7, 325–333. doi: 10.1007/BF00004006
10
GanesanP.MatsubaraK.OhkuboT.TanakaY.NodaK.SugawaraT.et al. (2010). Anti-Angiogenic Effect of Siphonaxanthin From Green Alga, Codium fragile. Phytomedicine17 (14), 1140–1144. doi: 10.1016/j.phymed.2010.05.005
11
GanesanP.NodaK.ManabeY.OhkuboT.TanakaY.MaokaT.et al. (2011). Siphonaxanthin, a Marine Carotenoid From Green Algae, Effectively Induces Apoptosis in Human Leukemia (HL-60) Cells. Biochim. Biophys. Acta1810 (5), 497–503. doi: 10.1016/j.bbagen.2011.02.008
12
GiossiC.CartaxanaP.CruzS. (2020). Photoprotective Role of Neoxanthin in Plants and Algae. Molecules25 (20), 4617. doi: 10.3390/molecules25204617
13
GiossiC. E.CruzS.ReyF.MarquesR.MeloT.DominguesM. R.et al. (2021). Light Induced Changes in Pigment and Lipid Profiles of Bryopsidales Algae. Front. Mar. Sci.8, 745083. doi: 10.3389/fmars.2021.745083
14
GossR.JakobT. (2010). Regulation and Function of Xanthophyll Cycle-Dependent Photoprotection in Algae. Photosynth. Res.106 (1-2), 103–122. doi: 10.1007/s11120-010-9536-x
15
HaftingJ. T.CraigieJ. S.StengelD. B.LoureiroR. R.BuschmannA. H.YarishC.et al. (2015). Prospects and Challenges for Industrial Production of Seaweed Bioactives. J. Phycol.51 (5), 821–837. doi: 10.1111/jpy.12326
16
HanisakM. D. (1979). Growth Patterns of Codium fragile Ssp. Tomentosoides in Response to Temperature, Irradiance, Salinity, and Nitrogen Source. Mar. Biol.50, 319–332. doi: 10.1007/BF00387009
17
HuangS.LiK.PanY.YuY.WernbergT.BettigniesT.et al. (2021). Artificial Light Source Selection in Seaweed Production: Growth of Seaweed and Biosynthesis of Photosynthetic Pigments and Soluble Protein. PeerJ9, e11351. doi: 10.7717/peerj.11351
18
HwangE. K.BaekJ. M.ParkC. S. (2008). Cultivation of the Green Alga, Codium fragile (Suringar) Hariot, by Artificial Seed Production in Korea. J. Appl. Phycol.20, 469–475. doi: 10.1007/s10811-007-9265-5
19
IngleK. N.PolikovskyM.ChemodanovA.GolbergA. (2018). Marine Integrated Pest Management (MIPM) Approach for Sustainable Seagriculture. Algal. Res.29, 223–232. doi: 10.1016/j.algal.2017.11.010
20
KimJ. K.YarishC.HwangE. K.ParkM.KimY. (2017). Seaweed Aquaculture: Cultivation Technologies, Challenges and its Ecosystem Services. Algae32 (1), 1–13. doi: 10.4490/algae.2017.32.3.3
21
KraayG. W.ZapataM.VeldhuisM. J. W. (1992). Separation of Chlorophylls c1, c2 and c3 of Marine Phytoplankton by Reversed-Phase-C18-High-Performance Liquid Chromatography. J. Phycol.28, 708–712. doi: 10.1111/j.0022-3646.1992.00708.x
22
LeandroA.PereiraL.GonçalvesA. M. M. (2019). Diverse Applications of Marine Macroalgae. Mar. Drugs18 (1), 17. doi: 10.3390/md18010017
23
MagnussonM.MataL.WangN.ZhaoJ.de NysR.PaulN. A. (2015). Manipulating Antioxidant Content in Macroalgae in Intensive Land-Based Cultivation Systems for Functional Food Applications. Algal. Res.8, 153–160. doi: 10.1016/j.algal.2015.02.007
24
MarquesR.CruzS.CaladoR.LillebøA.AbreuH.PereiraR.et al. (2021). Effects of Photoperiod and Light Spectra on Growth and Pigment Composition of the Green Macroalga Codium tomentosum. J. Appl. Phycol.33, 471–480. doi: 10.1007/s10811-020-02289-9
25
MendesC. R.CartaxanaP.BrotasV. (2007). HPLC Determination of Phytoplankton and Microphytobenthos Pigments: Comparing Resolution and Sensitivity of a C18 and a C8 Method. Limnol. Oceanogr. Meth.5, 363–370. doi: 10.4319/lom.2007.5.363
26
MoreiraA.CruzS.MarquesR.CartaxanaP. (2022). The Underexplored Potential of Green Macroalgae in Aquaculture. Rev. Aquac.14 (1), 5–26. doi: 10.1111/raq.12580
27
MurataN.TakahashiS.NishiyamaY.AllakhverdievS. I. (2007). Photoinhibition of Photosystem II Under Environmental Stress. Biochim. Biophys. Acta Bioenerg.1767 (6), 414–421. doi: 10.1016/j.bbabio.2006.11.019
28
MurchieE. H.LawsonT. (2013). Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot.64 (13), 3983–3998. doi: 10.1093/jxb/ert208
29
PangestutiR.KimS.-K. (2011). Biological Activities and Health Benefit Effects of Natural Pigments Derived From Marine Algae. J. Funct. Food.3 (4), 255–266. doi: 10.1016/j.jff.2011.07.001
30
Pérez-LlorénsJ. L.CarreroI. H.OñateJ. J. V.MurilloF. G. B.LeónÁ. (2018). Those Curious and Delicious Seaweeds: A Fascinating Voyage From Biology to Gastronomy (Cadiz: UCA Press).
31
RanielloR.LorentiM.BrunetC.BuiaM. (2004). Photosynthetic Plasticity of an Invasive Variety of Caulerpa racemosa in a Coastal Mediterranean Area: Light Harvesting Capacity and Seasonal Acclimation. Mar. Ecol. Prog. Ser.271, 113–120. doi: 10.3354/meps271113
32
ReyF.CartaxanaP.MeloT.CaladoR.PereiraR.AbreuH.et al. (2020). Domesticated Populations of Codium tomentosum Display Lipid Extracts With Lower Seasonal Shifts Than Conspecifics From the Wild –Relevance for Biotechnological Applications of This Green Seaweed. Mar. Drugs18 (4), 188. doi: 10.3390/md18040188
33
SahuS. K.IngleK. N.MantriV. A. (2020). “Epiphytism in Seaweed Farming: Causes, Status, and Implications” in Environmental Biotechnology Vol. 1. Environmental Chemistry for a Sustainable World. Eds. GothandamK. M.RanjanS.DasguptaN.LichtfouseE. (Cham: Springer), 227–242. doi: 10.1007/978-3-030-38192-9_9
34
SugawaraT.GanesanP.LiZ.ManabeY.HirataT. (2014). Siphonaxanthin, a Green Algal Carotenoid, as a Novel Functional Compound. Mar. Drugs12 (6), 3660–3668. doi: 10.3390/md12063660
35
TakahashiS.BadgerM. R. (2011). Photoprotection in Plants: A New Light on Photosystem II Damage. Trends Plant Sci.16 (1), 53–60. doi: 10.1016/j.tplants.2010.10.001
36
UragamiC.GalzeranoD.GallA.ShigematsuY.MeisterhansM.OkaN.et al. (2014). Light-Dependent Conformational Change of Neoxanthin in a Siphonous Green Alga, Codium intricatum, Revealed by Raman Spectroscopy. Photosynth. Res.121, 69–77. doi: 10.1007/s11120-014-0011-y
37
van den BurgS. W. K.DagevosH.HelmesR. J. K. (2021). Towards Sustainable European Seaweed Value Chains: A Triple P Perspective. ICES J. Mar. Sci.78 (1), 443–450. doi: 10.1093/icesjms/fsz183
38
WangH.-M,DChenC.-C.HuynhP.ChangJ.-S. (2015). Exploring the Potential of Using Algae in Cosmetics. Bioresour. Technol.184, 355–362. doi: 10.1016/j.biortech.2014.12.001
39
WangW.QinX.SangM.ChenD.WangK.LinR.et al. (2013). Spectral and Functional Studies on Siphonaxanthin-Type Light-Harvesting Complex of Photosystem II From Bryopsis corticulans. Photosynth. Res.117, 267–279. doi: 10.1007/s11120-013-9808-3
40
WardG. M.FaisanJ. P.Cottier-CookE. J.GachonC.HurtadoA. Q.LimP. E.et al. (2020). A Review of Reported Seaweed Diseases and Pests in Aquaculture in Asia. J. World Aquac. Soc51 (4), 815–828. doi: 10.1111/jwas.12649
Summary
Keywords
aquaculture, irradiance, epiphytes, light spectra, photosynthesis, photoprotection, pigments, seaweeds
Citation
Marques R, Moreira A, Cruz S, Calado R and Cartaxana P (2022) Controlling Light to Optimize Growth and Added Value of the Green Macroalga Codium tomentosum. Front. Mar. Sci. 9:906332. doi: 10.3389/fmars.2022.906332
Received
28 March 2022
Accepted
04 May 2022
Published
02 June 2022
Volume
9 - 2022
Edited by
JInghui Fang, Chinese Academy of Fishery Sciences (CAFS), China
Reviewed by
Alvaro Anibal Israel, University of Haifa, Israel; Di Zhang, Yantai University, Yantai, China
Updates
Copyright
© 2022 Marques, Moreira, Cruz, Calado and Cartaxana.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Cartaxana, pcartaxana@ua.pt
†Present Address: Rúben Marques, Scottish Association for Marine Science, University of the Highlands and Islands, Oban, United Kingdom
This article was submitted to Marine Fisheries, Aquaculture and Living Resources, a section of the journal Frontiers in Marine Science
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.