 Aurora M. Ricart1*†
Aurora M. Ricart1*† Melissa Ward1
Melissa Ward1 Tessa M. Hill1,2
Tessa M. Hill1,2 Eric Sanford1,3
Eric Sanford1,3 Kristy J. Kroeker4
Kristy J. Kroeker4 Yuihiro Takeshita5
Yuihiro Takeshita5 Sarah Merolla1
Sarah Merolla1 Priya Shukla1
Priya Shukla1 Aaron T. Ninokawa1Kristen Elsmore1Brian Gaylord1,3
Aaron T. Ninokawa1Kristen Elsmore1Brian Gaylord1,3- 1Bodega Marine Laboratory, University of California, Davis, Bodega Bay, CA, United States
- 2Department of Earth and Planetary Sciences, University of California, Davis, Davis, CA, United States
- 3Department of Evolution and Ecology, University of California, Davis, Davis, CA, United States
- 4Ecology & Evolutionary Biology Department, University of California, Santa Cruz, Santa Cruz, CA, United States
- 5Monterey Bay Aquarium Research Institute, Moss Landing, CA, United States
A Commentary on
Overstated Potential For Seagrass Meadows To Mitigate Coastal Ocean Acidification
By Van Dam, B., Lopes, C., Zeller, M. A., Ribas-ribas, M., Wang, H., and Thomas, H. (2021). Front. Mar. Sci. 8:729992. doi: .10.3389/fmars.2021.729992
Van Dam et al. (henceforth VD) published an Opinion (Van Dam et al., 2021a) and subsequent Corrigendum (Van Dam et al., 2021b) about our work regarding amelioration of low pH in seagrass ecosystems (Ricart et al., 2021). Below we discuss troubling details in the authors’ approach, an unaddressed error, misrepresentations, and problematic inferences; each contravenes VD’s argument of “overstated potential” for mitigation of low pH.
To start, VD’s original comment was rejected previously by Global Change Biology due to 1425 spurious data points and two invalid graphs. Despite being informed of these mistakes, VD submitted the identical, unchanged critique to Frontiers in Marine Science. The erroneous publication and Corrigendum resulted.
Even following correction, we disagree with VD’s two primary assertions:
1) VD claim that using ΔpH is “mathematically incorrect” because corresponding Δ[H+] values depend on initial pH, a rather strident statement given the relationship is well known (Fassbender et al., 2021; note that in our study, initial pH is that outside seagrass; i.e., Δ=measurement inside minus that outside). VD then confusingly duplicate a single set of measurements in their Figure 1A, plotting it as two separate data clusters. One cluster (their red points) improperly inverts values to show “–ΔpH” instead of “ΔpH” on the y-axis. The other, teal cluster employs within-meadow pH rather than outside-meadow pH as the independent variable, a choice unsuited to assessing whether seagrass ecosystems elevate pH relative to impinging waters, and one that is misleading. The correct relationship (Figure 1A here) demonstrates that although ΔpH and pH indeed covary, the greatest low-pH amelioration (strongest Δ[H+] depression) occurs when outside-meadow pH is low and acidification stress is high. Most importantly, key patterns of Ricart et al. (2021) remain unchanged when Δ[H+] is used instead of ΔpH (Figures 1B–D here). Therefore, our conclusions are robust to either ΔpH or Δ[H+], and pH broadens audience accessibility.
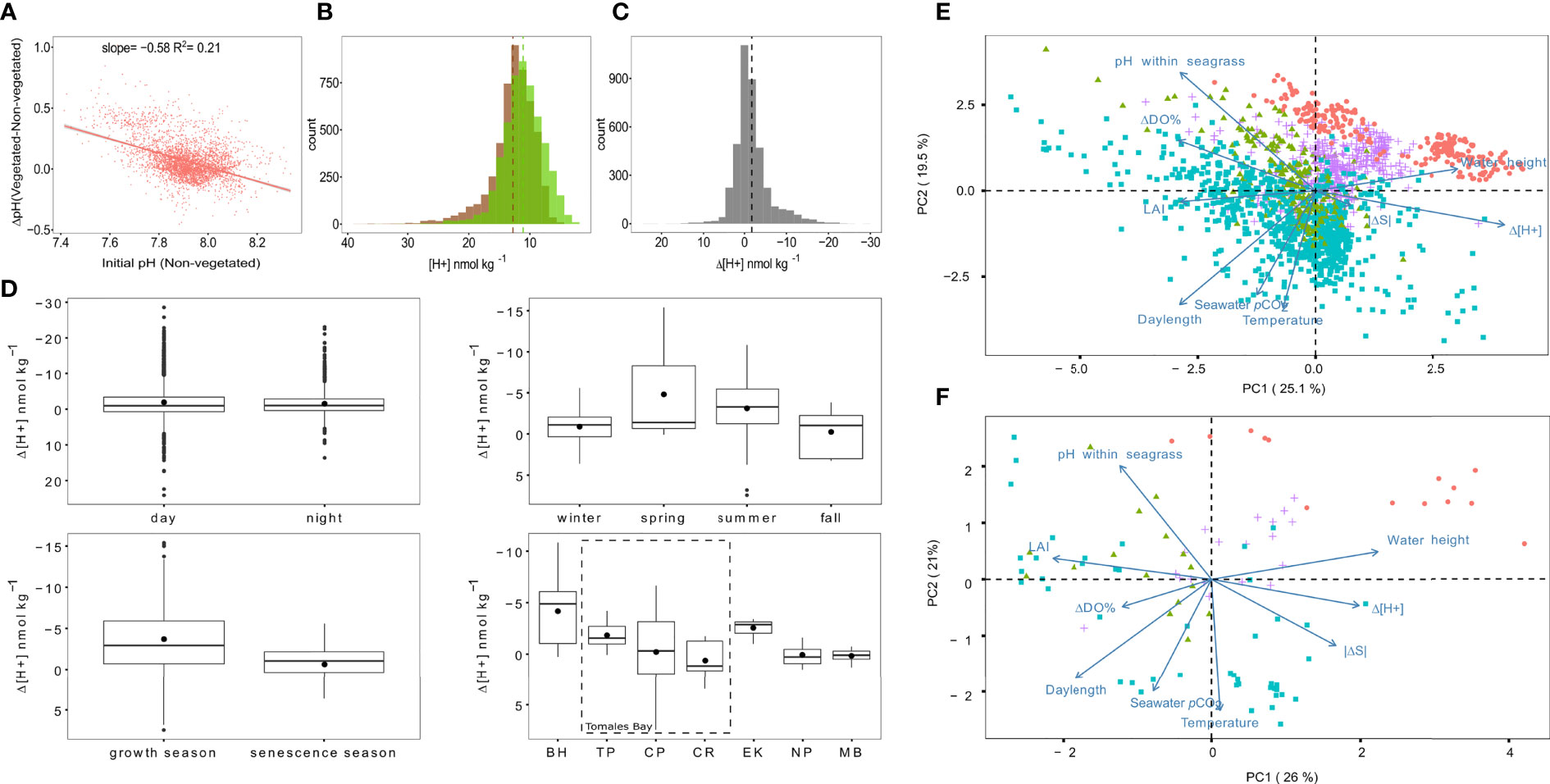
Figure 1 Corrections and clarifications to figures and assertions of VD. (A) Corrected version of VD’s panel A, showing ΔpH as a function of outside-meadow pH. Since ΔpH reflects a relative change in hydrogen ion concentration ([H+]) with respect to baseline conditions outside a seagrass meadow, it is more appropriate to use outside-meadow pH as the independent variable, rather than inside-meadow pH as was done in VD. The correction shown here results in a reversal of the slope of VD’s teal point cluster. The red point cluster in VD’s panel A depicts the same data as the teal points, except with outside-meadow pH on the x-axis. However, the red cluster is incorrect because it plots an inversion of ΔpH (i.e., -ΔpH) as the dependent variable rather than ΔpH; this mistake artificially inflates the number of negative ΔpH data at low pH in VD’s graph. Thus, the red points are simultaneously unnecessary and misleading. (B, C) Replotted versions of Figures 1A, 1B from Ricart et al. (2021), using [H+] instead of pH. Patterns here show matching trends to the figures of the original article, despite the change from logarithmic to arithmetic scaling along the x-axis. Colors indicate data from seagrass ecosystems (green), adjacent non-vegetated areas (brown), and differences between seagrass ecosystems and non-vegetated areas (grey). Dashed lines denote mean values. (D) Replotted version of Figure 5 from Ricart et al. (2021), demonstrating that key results again remain invariant if Δ[H+] is used instead of ΔpH. Note that y-axis values here run from positive at the bottom to negative at the top (expected given pH=-log[H+]), with negative Δ[H+] corresponding to amelioration of low pH. Top left, Δ[H+] during daytime compared to nighttime; top right, comparison of Δ[H+] among seasons; bottom left, comparison of Δ[H+] between seagrass productivity periods; bottom right, comparison of Δ[H+] among sites ordered latitudinally from north at left to south at right, including three seagrass meadows within Tomales Bay (dashed box). See Ricart et al. (2021) for additional details. (E, F) Principal components analyses (PCA) analogous to that of Figure 6 in Ricart et al. (2021) using Δ[H+] instead of ΔpH and incorporating ΔDO% and |ΔS| measurements. Results are shown for both the 1-h and 24-h datasets, respectively. Note that in both plots, Δ[H+] is negatively correlated with ΔDO% and positively correlated with |ΔS|, indicating that amelioration events [negative Δ(H+) values] tend to comport mainly with photosynthesis as a driver, and not with conditions characterized by strong water property gradients. RMS velocity does not appear in these plots, due to greater numbers of gaps in the velocity and ΔDO% records, which reduces their temporal overlap and prevents their simultaneous inclusion.
2) VD claim that we overstate the capacity of seagrasses to ameliorate low pH. However, we believe this stance relies too heavily on categorical thinking.
● VD state we attribute “all positive ΔpH values strictly to seagrass photosynthetic activity.” We never say this, nor do we utilize “all or none” analyses. We explicitly discuss several alternative agents of pH change.
● VD fall into artificial “yes/no” interpretations through a narrow focus on a simple model: if aerobic metabolism drives pH change, then dissolved oxygen (DO%) must rise and fall in concert. Although reasonable as a starting point, this model applies strictly to biogeochemical dynamics at a single time and place. In our study, inferences about pH amelioration derive from records differenced across space. Background patchiness in water properties creates spatiotemporal mismatches and noise in ΔpH vs. ΔDO% relationships. The same is true of tidal flows that reverse at different times depending on where a recording instrument is located relative to shore. Subsets of ΔpH and ΔDO% data will therefore deviate unavoidably from a simple photosynthesis model. What appears more relevant in VD’s analysis is that the most common conditions at northern sites, where our study suggests amelioration of low pH was strongest, do align with photosynthesis as a driver (left teal bars in their figure 1c).
● The notion that photosynthesis is important only if ΔpH and ΔDO% change in lockstep becomes especially problematic in broader interpretations of VD’s Figure 1C. The authors assume if a measurement does not uphold their simple model, then it refutes it (binary outcome). This assumption leads them to conclude that “…pH increases were most often caused by factors other than seagrass photosynthetic CO2 uptake.” However, an elementary thought experiment indicates the impropriety of this assertion. We focus on categories of their Figure 1C where spatial differences in salinity suggest distinct water masses bathed stations inside vs. outside meadows (i.e., |ΔS|>0.1; green and purple bars). Under such conditions, a water mass flowing inside a meadow could have started, prior to interacting with the seagrass ecosystem, with lower [H+] than the nearby but separate water mass flowing outside the meadow. This background gradient in pH between inside and outside stations would then inflate the perceived role of seagrass by engendering a more negative Δ[H+]. Importantly, however, the opposite is also possible: an inside meadow water mass could have started, prior to interacting with a meadow, with sufficiently high [H+] that photosynthesis could decrease [H+] and still not induce a negative Δ[H+] between inside and outside stations. In VD’s calculations, this latter scenario would count incorrectly as refuting a photosynthesis effect, even though such an effect was active. Together these contrasting situations (and analogous ones for ΔDO%) indicate that conditions where strong water property gradients exist are agnostic for revealing whether seagrass photosynthesis decreased [H+]. Employing such cases (i.e., anytime |ΔS|>0.1) as evidence against a photosynthesis effect is therefore inappropriate. Indeed, a mirroring of VD’s logic enables just as easily the claim that 1085 records were potentially consistent with a photosynthesis model. This number equals 70% of the low-pH amelioration measurements in the analysis. Spatiotemporal mismatches noted above, or additional non-photosynthetic processes, including ones active in sediments and pore waters, could contribute to the remaining fraction and warrant study (e.g., left red bars in VD’s Figure 1C). However, they too are components of “seagrass ecosystems,” the term we use preferentially in our paper over just “seagrasses.”
● VD selectively ignore evidence that conflicts with their arguments. In our study, ΔpH co-varies with leaf area index and daylength (factors associated with photosynthesis; Figure 6, Ricart et al., 2021) but is negatively correlated with water height and RMS velocity (factors that dilute and disperse signatures of photosynthesis). These trends do not align with VD’s argument that water mass differences biased patterns of ΔpH. Furthermore, if the principal components analysis is elaborated to include Δ[H+] instead of ΔpH, along with ΔDO% and |ΔS|, vectors of Δ[H+] and ΔDO% orient in opposite directions, as expected, while |ΔS| aligns with Δ[H+]. The latter suggests water mass differences were not a primary driver of the observed patterns (Figures 1E, F here); indeed, if strong water property gradients were inflating amelioration events, the |ΔS| vector would be reversed.
More issues: Our study documents that >65% of hourly records showed elevated pH within meadows. VD incorrectly report 62%, confusing matters by ignoring 1425 valid ΔpH measurements (a remarkable inversion of their original error, where they inserted an identical number of false oxygen values). They also describe their 62% as “marginally” above what “chance alone would dictate”. However, the null model underlying this statistical assertion appears inappropriate. Data for pH and DO are not black or white balls being drawn from a jar; they are continuous variables, and the null expectation of their distributions is more complicated. This distinction is one reason our study relies on analyses of actual values rather than lumped categorical tallies.
We similarly find unconvincing VD’s statement, “… the claim that OA amelioration persists overnight and ‘up to 21 days’ should be revisited”, where they assert that water masses will be completely replaced during low tides. Complete exchange of water masses does not occur in our meadows, where only shallower portions become emergent during extreme low tides (pH/DO/salinity/temperature sensors located mid-depth, submerged >97% of the time). Moreover, seawater that does exit is often returned on the next high tide and can be processed further chemically; indeed, flow reversals habitually lengthen residence times in estuaries (Fischer et al., 1979).
Additional work is indisputably necessary to dissect mechanisms of pH modification in macrophyte habitats. Regardless of those mechanisms, in our study, low pH was routinely ameliorated in seagrass ecosystems, where photosynthetic activity seemed to play an important role.
Author Contributions
AR and BG analyzed the data. AR led the writing of the manuscript. BG, TH, ES, KK, YT, MW, PS, AN, SM, and KE contributed critically to the drafts and gave final approval for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Fassbender A. J., Orr J. C., Dickson A. G. (2021). Technical Note: Interpreting pH Changes. Biogeosciences 18, 1407–1415. doi: 10.5194/bg-18-1407-2021
Fischer H., List J., Koh C., Imberger J., Brooks N. (1979). Mixing in Inland and Coastal Waters (Cambridge: Academic Press).
Ricart A. M., Ward M., Hill T. M., Sanford E., Kroeker K. J., Takeshita Y., et al. (2021). Coast-Wide Evidence of Low pH Amelioration by Seagrass Ecosystems. Glob. Change Biol. 27, 2580–2591. doi: 10.1111/gcb.15594
Van Dam B., Lopes C., Zeller M. A., Ribas-ribas M., Wang H., Thomas H. (2021a). Overstated Potential for Seagrass Meadows to Mitigate Coastal Ocean Acidification. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.729992
Keywords: ocean acidification, seagrass, carbonate chemistry, CO2 system calculations, commentary articles
Citation: Ricart AM, Ward M, Hill TM, Sanford E, Kroeker KJ, Takeshita Y, Merolla S, Shukla P, Ninokawa AT, Elsmore K and Gaylord B (2022) Commentary: Overstated Potential for Seagrass Meadows to Mitigate Coastal Ocean Acidification. Front. Mar. Sci. 9:884857. doi: 10.3389/fmars.2022.884857
Received: 27 February 2022; Accepted: 06 April 2022;
Published: 28 April 2022.
Edited by:
Antonio Cobelo-Garcia, Spanish National Research Council (CSIC), SpainReviewed by:
Matthew Liebman, United States Environmental Protection Agency (EPA), United StatesCopyright © 2022 Ricart, Ward, Hill, Sanford, Kroeker, Takeshita, Merolla, Shukla, Ninokawa, Elsmore and Gaylord. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurora M. Ricart, YW1yaWNhcnRAdWNkYXZpcy5lZHU=
†Present address: Aurora M. Ricart, Bigelow Laboratory for Ocean Sciences, East Boothbay, ME, United States