 Beatriu Tort Castro1*
Beatriu Tort Castro1* Rocío Prieto González1,2
Rocío Prieto González1,2 Seán A. O’Callaghan1
Seán A. O’Callaghan1 Paula Dominguez Rein-Loring1Eduard Degollada Bastos1
Paula Dominguez Rein-Loring1Eduard Degollada Bastos1- 1EDMAKTUB Association, Caldes d’Estrac, Spain
- 2Laboratory of Applied Bioacoustics, Polytechnic University of Catalonia (UPC), BarcelonaTech, Vilanova i la Geltrú, Spain
Ship strikes are a widespread conservation issue for many cetacean species globally. Population level impacts depend on the occurrence and severity of collisions, which may lead to life altering injuries or fatalities. Such impacts are a major concern for large, long-lived, and reproductively slow species like the fin whale. Since 2014, a seasonal feeding aggregation of fin whales has been monitored from February to June off the Catalan coast (Spain), in the northwest Mediterranean Sea. Oceanographical factors influence the occurrence and high density of krill within submarine canyons along the continental shelf, resulting in high whale abundance within a small spatial area. The study area extends 37 km offshore across a 1,944 km2 marine strip situated between the towns of Torredembarra and Castelldefels. This fin whale feeding ground is exposed to high density marine vessel traffic, given its location between the northern Mediterranean shipping lane, which links Barcelona and Tarragona Ports to the Atlantic Ocean and wider Mediterranean Basin. Ship strikes represent the greatest conservation threat for fin whales in the Mediterranean Sea. At least four fin whales have been found dead in Barcelona Port since 1986 due to ship strikes and seven live whales have been documented with injuries in the study area since 2018. Fin whale distribution was mapped with known high-risk marine vessels’ (cargo, tanker and passenger vessels) shipping lanes. Vessel density and shipping lanes characterised by speed were considered. Collision risk was estimated monthly based on the predicted fin whale occurrence and traffic density. Several shipping lanes crossed the fin whale feeding habitat every month with an average speed of 15 kn. Cargo vessels displayed the highest ship-strike risk during April, overlapping with the peak of fin whale sightings in the critical feeding area. Slower vessel speeds (8 kn) in waters <200 m depth or along the continental shelf should be implemented along the Catalan coast, during the whale season. These suggestions should be applied into the Barcelona Port transport separation scheme. Ship strike risk for this species will persist unless active management plans are adapted in the region to mitigate its risk.
1. Introduction
The fin whale (Balaenoptera physalus) (Linnaeus, 1758) is the largest cetacean species present in the Mediterranean Sea and the only baleen whale species regularly found within the Mediterranean basin year-round (Notarbartolo di Sciara et al., 2016; Aguilar and García-Vernet, 2018). Two populations have been identified within the Mediterranean Sea: the Northeast Atlantic Ocean (NENA) population and the Mediterranean subpopulation (Notarbartolo di Sciara et al., 2016). Both populations cohabit annually in the Balearic Sea off southern Spain and along the Catalan coast (Castellote et al., 2008; Castellote et al., 2012a; Gauffier et al., 2020). The NENA population moves through the Strait of Gibraltar into the Mediterranean basin between November and April and leaves between May and October (Notarbartolo di Sciara et al., 2016; Gauffier et al., 2018; Gauffier et al., 2020). On the other hand, the Mediterranean subpopulation is found in the Ligurian Sea in summer from July to September and in the Eastern Mediterranean in winter, where they have been observed feeding around Lampedusa Island in February. However, there is some evidence for the species presence in Ligurian between October and December (Canese et al., 2006; Panigada et al., 2006; Pintore et al., 2021).
Fin whales primarily feed on northern krill (Meganyctiphanes norvegica) (M. Sars, 1857) in the Mediterranean Sea (Notarbartolo di Sciara et al., 2016; Borrell et al., 2021). M. norvegica requires high levels of primary productivity that are linked to oceanic fronts (Druon et al., 2012). These oceanic fronts occur seasonally in patches across the Mediterranean and are linked to fin whale movements (Druon et al., 2012; Panigada et al., 2017; Lydersen et al., 2020). Over the past decade, during the spring, a new feeding ground has been discovered along the continental shelf off the Catalan coast, between Torredembarra and Castelldefels. Additionally, this feeding aggregation was discovered around the Cunit and Foix submarine canyons (EDMAKTUB, 2021, in press). These underwater geographic formations are particularly important due to the upwelling effect present in the area, which triggers elevated krill densities during spring (EDMAKTUB, 2021, in press; Sabates et al., 1989; Puig et al., 2000).
Distinct behavioural patterns in fin whales have been documented on the Garraf Coast. Surface feeding behaviour was observed approximately within the first 10 m of the water column, by EDMAKTUB (2018). While feeding, fin whales exhibit irregular movements in a zig-zag pattern or circular motion (Tort et al., 2017). Breathing patterns were observed to oscillate on average between two to six min between surface intervals (Tort et al., 2017). Moreover, travelling, and resting behaviour have been identified in this area. While travelling, fin whales conduct a linear path with a constant speed. Both resting and travelling have been described with a regular breathing pattern (Tort et al., 2017). Resting behaviour is defined when a whale remains stationary at the surface, or at a shallow depth just below the surface.
The fin whale is threatened by a variety of natural diseases, such as the cetacean morbillivirus (CeMV), and by anthropogenic effects including persistent pollutants, noise pollution, fishing gear entanglement, climate change and ship strikes within the Mediterranean Sea (Panigada et al., 2006; Mazzariol et al., 2016; Notarbartolo di Sciara et al., 2016). Moreover, it is a long-lived species with a high mortality rate in the first stage of life (77%), which decreases with maturity (Arrigoni et al., 2011). Female fin whales become reproductively active between 22.8 and 36.8 years, making the species highly vulnerable to natural and anthropogenic pressures. This leads to the high mortality rate recorded in the Mediterranean populations (Panigada et al., 2006; Arrigoni et al., 2011).
Fin whales are exposed to several anthropogenic threats (Panigada et al., 2006; Notarbartolo di Sciara et al., 2016). The Mediterranean Sea is one of the most heavily populated and anthropogenically impacted marine habitats globally, despite comprising only 0.8% of the global marine surface area (Micheli et al., 2013; Vaes and Druon, 2013). In this area, shipping has grown exponentially since 1992, reaching up to 30% of the global maritime traffic annually (Vaes and Druon, 2013). 220,000 ships >100 gross tons operate within it, accessing 300 ports, while following shipping lanes along the north of African coast or the southern coast of Europe (Panigada et al., 2010; Vaes and Druon, 2013).
The International Union for Conservation of Nature (IUCN) red list assessment recently classified the conservation status of the Mediterranean fin whale population as endangered (Panigada et al., 2021) and the status of the NENA population as vulnerable (Cooke, 2018). The species is protected across its range by the European Union Habitats Directive as an Annex IV species, the Marine Strategy Framework Directive (MSFD) and the Agreement on the Conservation of Cetaceans in the Black Sea, the Mediterranean Sea and Contiguous Atlantic Area (ACCOBAMS) (Vaes and Druon, 2013; Notarbartolo di Sciara et al., 2016). Within Spanish waters, cetaceans are protected from disturbance and harm with exclusion zones applied around animals under the Royal Decree 1727/2007, on December 21st. Additionally, fin whales are included in the list of wild species under special protection regime and in the Spanish catalogue of threatened species, being catalogued as vulnerable by Royal Decree 139/2011, of September 2021.
To protect migratory and resident cetacean species, the Mediterranean Cetacean Migration Corridor (CMC) declared the area as a Specially Protected Area of Mediterranean Importance (SPAMI) at the COP21, which was adopted by the Barcelona Convention in 2019 (OceanCare, 2021). The CMC was established by the Royal Decree 699/2018 in June 2018. The CMC comprises 46,385 km2 between Catalonia and Valencia within the Balearic archipelago in the Balearic Sea (OceanCare, 2021). Likewise, the Pelagos Sanctuary was designated within the Ligurian Sea between France and Italy in 1999 (Notarbartolo di Sciara et al., 2016). This designation was in response to the region’s importance for the species in the summer, as it supports the highest density of foraging fin whales in the Mediterranean basin (Notarbartolo di Sciara et al., 2016; Panigada et al., 2017). An Important Marine Mammal Area (IMMA) was also implemented across the North-West Mediterranean Sea, slope and canyon system, between the Pelagos Sanctuary and the Balearic Sea (IUCN, 2017; Torreblanca et al., 2019). This IMMA acts as a guide for the designation of marine protected areas (MPA’s) for important habitats for species warranting formal protection measures. Furthermore, the Coast of Garraf is included in the Natura 2000 Network area, which establishes a robust policy framework to ensure ecosystem sustainability and contributes to international efforts in marine conservation (Natura 2000, European Commission), Figure 1.
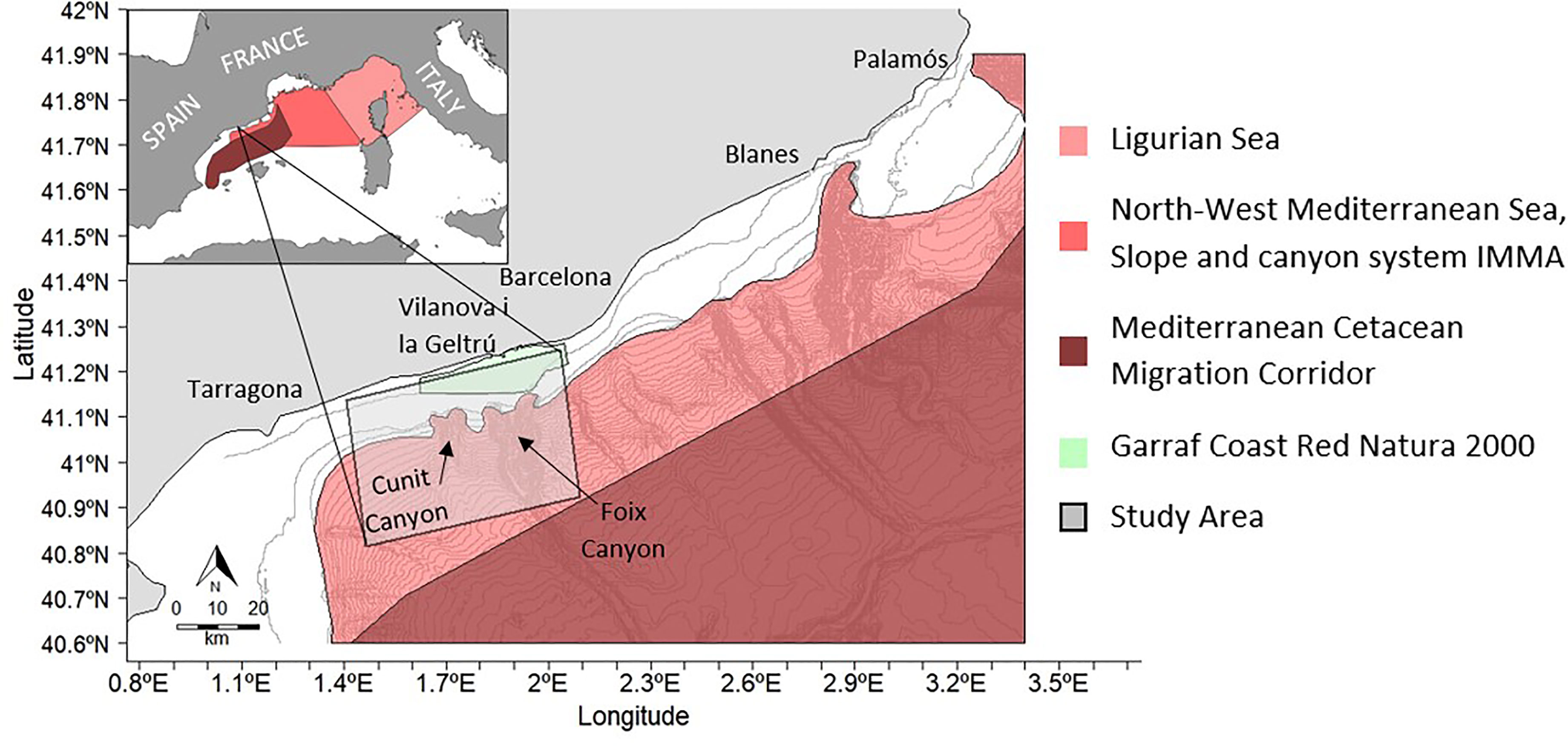
Figure 1 Fin whale project study area along the Garraf coast. The Foix Canyon (largest system) is located on the right side of the study area and the Cunit canyon is situated on the left side. The bathymetry lines have an interval of 50 meters. In the top left it can be seen an image of the W- Mediterranean in which there are represented the SPAMI areas;.In dark red the cetacean corridor, in light pink the Pelagos Sanctuary, in red the North-West Mediterranean Sea, Slope and canyon system IMMA a.nd in light green the Garraf Coast Red Natura 2000.
Ship strikes represent the current greatest threat to large whales in the Mediterranean Sea (Laist et al., 2001; Panigada et al., 2006; Vaes and Druon, 2013; Di-Meglio et al., 2018; Frantzis et al., 2019; Schoeman et al., 2020; Winkler et al., 2020). Fin whales are the cetacean species most often killed by ship strikes globally (Laist et al., 2001; Panigada et al., 2006; Winkler et al., 2020). When collided with, fin whales can be pinned onto the bow of large vessels or become dislodged from the bow and float due to decomposition gases (Laist et al., 2001; Moore et al., 2020). Injuries resulting from ship strikes vary in severity depending on the speed and size of the vessel involved in the incident (Laist et al., 2001; Panigada et al., 2006). They typically occur along the lateral and dorsal sides of whales and vary from lacerations caused by propellor movements, blunt force trauma causing broken bones and scarring to deformations and amputations of body parts (Laist et al., 2001; Panigada et al., 2006; de Reuver et al., 2021). Collisions occur most often over continental shelf waters (Laist et al., 2001). The possibility for large vessels to spot and successfully evade hitting surfacing whales is inversely proportional to the size and speed of the vessel (Laist et al., 2001). Once vessel speed reaches ≥14 kn (25.9 km/h) the likelihood of severe injuries or a fatal interaction to occur between a whale and a ship (≥ 80 m) increases greatly (Laist et al., 2001; Panigada et al., 2006; Keen et al., 2019). Damage to vessels and injuries to the crew onboard have also been reported on some occasions (Schoeman et al., 2020; Sèbe et al., 2020; Laist et al., 2001). Collision risk areas are dynamic regions where environmental factors, prey density, whale occurrence and marine traffic combine to threaten whales (Ham et al., 2021). Within the Mediterranean between 1972 and 2001, 46 (16%) of 287 stranded fin whale carcasses examined were killed by ship strikes primarily within or adjacent to the Pelagos Sanctuary, where near miss events were also noted from 2008 to 2019 (Panigada et al., 2006; David et al., 2022). The risk posed by ship strikes to fin whales has been extensively studied within the Ligurian Sea in the North-Western Mediterranean (Panigada et al., 2006; Vaes and Druon, 2013; Ham et al., 2021; David et al., 2022). In this area, an overlap of important shipping lanes and satellite tagged fin whales exhibiting foraging behaviour, demonstrated that the species uses seasonal foraging habitats in high-risk areas (Panigada et al., 2017). However, there is a lack of knowledge about the risk posed by ship strikes in the Balearic Sea.
The aim of this study was to identify critical areas where marine traffic and fin whale presence overlapped off the Catalan coast of Garraf, an area between the large ports of Barcelona and Tarragona. This study was structured in different steps. Firstly, fin whale behaviour was evaluated, and fin whale distribution was estimated in the study area. Secondly, photographed fin whales displaying collision marks and historical strandings data were accessed and evaluated. Finally, marine traffic maps to detect ship route concentrations were created considering types of vessels with known high ship strike risk. Vessel speed by vessel type and length was evaluated while route density maps were created which was used to identify potential fin whale ship strike risk areas within the study area. This study will contribute to risk mitigation and protection measures in order to promote conservation, help to make management decisions and reduce the risk of collisions occurring while fin whales are seasonally present.
2 The Methodology
2.1 Study Area
The study area comprises a 1,944 km2 region, centred around the city of Vilanova i la Geltrú and extending 37 km offshore. The area stretches between Torredembarra and Castelldefels out to 40°81.38’ N, 1°46.50’ E and 40°92.43’ N, 2°09.21’ E (Figure 1). The Cunit and Foix Canyons are situated within the middle of the survey area along the continental shelf edge. The bathymetry rapidly deepens from the continental shelf (100 m) to 2,000 m in the Foix Canyon (Puig et al., 2000). These canyon systems and the Liguro-Provençal-Catalan current create an upwelling effect which promotes primary production during spring months resulting in high densities of krill along the shelf edge (Puig et al., 2000).
The survey fieldwork used for this study was carried out in 2021, between Mach and May. A 14.3 m catamaran RV MAKTUB (model catana 471) was the research platform where dedicated visual surveying took place, on a near daily basis during the fieldwork season. The area was surveyed under engine power and by sail during suitable weather conditions (< Beaufort Sea State 4 and < 2 m swell) and with a sustained speed of five to six knots. The area being surveyed did not follow a systematic approach. The daily survey effort depended on where whale activity was detected over previous days, reported by fisherman, or determined by the weather conditions in the study area (sea state and wind strength/direction). The continental shelf edge and submarine canyon systems, where the species regularly forages, were targeted for surveying.
2.2 Fin Whale Data Collection
Fin whale sightings were obtained from dedicated research surveys. Visual effort began and finished at the 30 m in depth mark. Four observers continuously scanned the sea for whales from all 4 sides of the catamaran to have 360° coverage. Fin whales were detected either by visually spotting surfacing animals and their tall columnar blow with the naked eye or with the use of 8 x 42 binoculars. One of the observers on a break was responsible for data recording during sightings. Positions were rotated every 30 min to avoid fatigue and to minimise observer bias. These observers were trained at the beginning of the season and remained throughout it to ensure consistency in the data collection.
Once a fin whale was visually detected, survey effort to detect whales was ended and the following protocol began. Sightings occurred when the individual(s) were first seen and ended when identification data and/or biological samples were obtained. To minimise duplicating sightings of the same individuals, new sightings were recorded in three circumstances: (1) if more animals were sighted in a period over 45 min and identified as different through photo-identification, (2) if animals surfaced >3 km away from the last whale dive position and were observed in less than 10 mins from the last observation, (3) if they were sighted >2 hrs after the initial sighting. In each sighting the GPS position, species (if other cetacean species were encountered), the number of animals, the start/end time, the angle, and the estimated distance of the whale when sighted for the first time were recorded. Distance was estimated using a compass and judgement from the skipper. Environmental conditions at the beginning of the sighting were documented. During the sighting, additional data was recorded, such as photo-identification pictures, drone videos and behaviour of the animal(s).
2.3 Evidence of Ship Strikes on Live and Dead Fin Whales
Live fin whales were photographed and/or filmed with DSLR cameras (Nikon D7100 with a 150 - 400 mm telephoto lens and Canon 90D with a 70 – 200 mm telephoto lens) and a drone (DJI Mavic Pro 2/Phantom 3 Pro) during the field season for photo-identification purposes. Whales with lesions and damage consistent with ship strikes were compiled to gauge the prevalence of non-lethal ship strikes in the study area.
Strandings of fin whales were assessed from along the Garraf coast using the literature and newspaper reports to identify whales with evidence of ship strike damage such as acute abrasion and hematoma bruising on the dorsal side of the body (Laist et al., 2001; Panigada et al., 2006). Fin whales calve standings were also investigated within the study area and along the Catalan coast. In addition, records of fin whale bones recovered by bottom trawling fishermen in the study area between the ports of Ametlla de Marand Palamós were also compiled.
2.4 Fin Whale Distribution
The statistical analysis was conducted using R software version 4.1.1 (R Core Team, 2020). QGis version 3.4.3 – Madeira (QGIS Development Team, 2018) was used to map the fin whale presence and the traffic data.
Fin whale sightings recorded by dedicated research surveys and accounting for observer effort were used to determine the presence and absence of fin whales across the study area. The observer effort was defined as the time where observers were actively surveying. Only on effort transects were considered for analysis. The 2021 season was used to perform the distribution model. The aim of the model was to compute the collision risk map. The complete traffic database available was the EMODnet database (www.emodnet-humanactivities.eu) which starts in 2019 (see section 2.5). 2019 was an unusual year with few sightings and in 2020 the COVID-19 pandemic stopped the season, so data collection was not possible. Therefore, only 2021 data was used for the purposes of this study.
The data collected was assessed per month (from March to May). The data was transferred to a 3x3 km grid. To define the sampled area, only the vessel tracks, in which observers were on effort were considered. A buffer of 3.7 km perpendicular distance from the vessel track was generated, according to the maximum distance where whales were sighted during surveys. The probability of fin whale detection depended on their distance from the vessel, so a probability of detection was assigned to the buffer area. It was 1 for the first 1.8 km, where it was assumed that all fin whales would definitely be detected if present, according to the sightings recorded and as followed by other authors such as Ham et al. (2021). The detection probability then decreased with increasing distance ranges: 0.75 from 1.8 to 2.4 km, 0.5 from 2.4 to 2.7 km and 0.25 from 2.7 to 3.7 km. The buffer threshold was established according to the percentages of sightings for each distance range, being 0.85, 0.95 and 0.97 respectively. These buffers were then overlaid on the 3 km grid. A grid cell was only considered sampled if ≥50% of its area was covered by the buffer with a detection probability greater than 0.5, otherwise, it was excluded. For each sampled grid cell, a unit effort associated with the buffer was defined.
The unit effort was defined as the weighted average of the probability of detection by coverture on each pixel. A grid cell may be covered by multiple buffers during a month (e.g., a vessel track might pass more than once by the same grid cells in a month). Therefore, all unit efforts within each cell were then monthly summed. This grid sampled effort (hereafter called effort) was included into the fin whale distribution model as a correction variable.
Fin whale presence and absence within the sampled area were associated with each grid cell. When there was a sighting, it was assigned the value according to the number of animals observed. The total observations within each grid cell were then monthly summed to calculate the total number of sightings recorded in that particular cell. A zero was assigned when there were no sightings in a sampled effort cell.
Four oceanographic variables were considered as explanatory oceanographic variables to describe the fin whale distribution over the study area: chlorophyll a concentration (Chla in mg/m3), sea surface temperature (SST in K), mean depth (m) and mean slope angle (°) of the grid cell. The Chla and the SST were obtained from E.U. Copernicus Marine Services Information, both with a daily resolution of 1x1 km and 0.01°x0.01° raster respectively in a L3 processing level. Bathymetry data were derived from a fine resolution 115x115 m raster provided by the European Marine Observation and Data Network (EMODnet) (www.emodnet-humanactivities.eu). The slope angle of the seafloor was also computed through EMODnet using the slope function in QGis.
The oceanographic factors were summarised by month (from March to May) using the 75 quantile for the Chla and the mean for the SST. Then, all oceanographic values were gridded into the 3 km cells by computing the mean value with the Zonal Statistics function in QGis. Having a resultant grid with a mean Chla, SST, depth, and slope value per month.
The fin whale distribution for 2021 was modelled using Generalised Additive Models (GAMs) with Poisson distribution family and log link function and were computed using the mgcv package (Wood, 2011). All the available oceanographic variables were considered in a preliminary analysis to select the most relevant ones. A parametric linear term for each variable was checked against a smooth alternative and interactions. The variable selection was based on two criteria; (1) to determine which covariates have the strongest effects on the number of sightings, enhancing model interpretability, and (2) improving its prediction accuracy, aiming for a balance between fit and parsimony. For all the possible combinations, the prediction error criteria General Additive Cross-Validation (GACV) was used to compare models. Environmental variables that were not interpretable were discarded even if they improved the model fit.
The relationship between fin whale distribution and the recurring spring phytoplankton bloom that primarily occurs in coastal waters, continental shelf breaks and around the submarine canyons was investigated by these models. Krill does not move far from the continental shelf into shallower waters, therefore, fin whales are typically found in waters >50 m in depth, where krill aggregations can be found. Hence, a coastal correction factor (CC) was used to mask coastal areas and exclude high Chla values caused by river runoff that might affect results interpretation.
2.5 Marine Traffic Data
In this study, three complementary marine traffic databases were used for different aims. Firstly, commercial port data was acquired from the Barcelona Port authority and from the literature (OceanCare, 2021; Barcelona Port, 2022). The annual usage of Barcelona Port by cargo ships (Ro-Ro ships, Lo-Lo ships, bulk carrier and container ships), tanker vessels (oil tankers) and passenger vessels (cruise ships and ferries) were analysed from 2016 to 2021 with the aim of characterizing the marine traffic in the area. Cargo boats were defined as 70 to >200 m in length and ≤ 15 kt in speed, tankers were >80 m in length and 15 kt in speed and passenger ships were >80 m in length and <24 kt in speed. Secondly, two sources of Automated Identification System (AIS) data were analysed: (1) from the antenna located at the Laboratory of Applied Bioacoustics (LAB) of the Technical University of Catalonia, BarcelonaTech (UPC); and (2) from EMODnet. The aim, in this case, was to map and characterize the traffic corridors in the study area. The EMODnet database was used to compensate for the constraints from the LAB database, as explained below.
The AIS data was used to identify the position of vessels, their course and speed, as required by the International Maritime Organisation (IMO). The IMO requires AIS use by all vessels >500 gross tons, for any vessel >300 gross tons that is engaged on international voyages and for all passenger vessels irrespective of size (IMO, 2002). However, AIS transceivers are commercially available and are also used on vessels that do not meet the requirements by law.
High-risk ship strike vessels were selected based on criteria published in the literature considering overall length and speed (Laist et al., 2001; Panigada et al., 2006; Winkler et al., 2020; Stepanuk et al., 2021). Following Stepanuk et al. (2021), three types of vessels (cargo, tanker and passenger ships) were selected using their IMO, a unique identification number. These vessel types were previously identified as high risk in Panigada et al. (2006). The occurrence of these vessel types, seasonality and movement patterns were then assessed within the study area.
The LAB AIS data was accessible from 2011 to 2020. It was stored through the data stream of the shallow water OBSEA platform in the Mediterranean Sea. OBSEA (www.obsea.es) is a cabled seafloor underwater observatory located 4 km (41°10.92’ N, 1°45.14’ E) off the Vilanova i la Geltru coast and placed within a fishing protected area. In the AIS data from the LAB, there were gaps and much variability mainly due to two factors. Firstly, the AIS data was saved through OBSEA and therefore it was only stored when OBSEA was active. Secondly, the range of the antenna receiver decreased over time, resulting in a lack of data in recent years (e.g., in 2020, 90% of the data was below an 8 km range and less than 1% of vessels were detected at more than 15 km from OBSEA).
A two-step quality control check was implemented. Vessels detected on land or with erroneous values were deleted and then, only data from days that registered ships >42 km from the LAB, in Vilanova i la Geltrú were selected for analysis. Thus, only days with a detection range which covered the whole study area were considered for analysis, ensuring that biassed AIS data (with a partial detection range over the study area) were excluded. Given the goal of this study was to identify the principal marine traffic lines through the survey area (which remain consistent annually), all the years of data from 2011 to 2020 were pooled by month.
Shipping lanes and vessel speed within the study area from February to June were identified. Cargo, tanker, and passenger vessel shipping lanes were represented by monthly maps, characterised by three speed ranges: low (up to 8 kn), moderate (8 to 16 kn) and high (>16 kn). Vessel traffic speed through the study area was also analysed. Vessel speed distributions were compared between years and within years to evaluate the reliability of pooling all the AIS data. The normality assumption was checked to use more statistically powerful parametric tests. However, since the normality assumption failed, three non-parametric tests were computed. The Kolmogorov-Smirnoff test, a test of the equality of continuous, one-dimensional probability distributions; the Mann-Whitney-U test (also called Wilcoxon rank-sum test) which consider as null hypothesis equal medians; and the Kruskal-Wallis test by ranks, the non-parametric equivalent of the ANOVA but comparing that the medians of all groups are equal. The speed distribution by vessel type and length were also compared. We classified AIS transmissions into vessel length classes as in Panigada et al. (2006): 50−100, 100−200, 200−300, and 300−400 m. Records that contained no information on vessel classification were not included in the analyses.
Due to the constraints with the LAB data quality, a vessel density map per month and year was unfeasible but needed to model the ship strike risk assessment in 2021. Therefore, a parallel analysis using EMODnet data was implemented. Marine traffic data between March and May 2021 was acquired. Route density maps produced and provided to EMODnet by the European Maritime Safety Agency (EMSA) were obtained for cargo, tanker, and passenger vessels. The route density of vessels in the study area was then calculated by computing the mean of the number of routes per month in the same 3x3 km grid squares for each vessel type in 2021, using EMODnet dataset.
2.6 Ship Strike Risk Assessment
The critical fin whale habitat assessment was adapted from Tregenza et al. (2000). The risk of vessel strike (Ri) was defined as the probability of lethality to fin whales, from 0 (low) to 1 (high). Ri was described by this equation:
Where Wi is the mean hull width in metres according to David et al. (2011), L is the length of a whale in metres, Dcell is the mean distance travelled per pixel, T is the percentage of time near the surface, NAD is the density of fin whales according to the distribution model and VD is the density of vessels computed from the EMODnet database (see section 2.5).
Fin whale size (L) in the study area was estimated to be approximately 17 m, by using drone measures and according to (Szegedi et al., 2019). The percentage of time near the surface was set to 30, which was extracted from David et al. (2011). To convert the number of fin whales into a probability density, the predicted number of animals by grid cell was divided by the total number of fin whales estimated (NAD) in the grid for the whole season.
These variables were computed monthly in a 3x3 km2 grid, the same as in the distribution model. The Dcell was estimated following the formula from Druon (2014):
Where Dmax is the maximum distance which can be travelled by a vessel in a grid cell.
3 Results
3.1 Fin Whale Sightings
Between March and May in 2021, 149 fin whale sightings were recorded. These sightings consisted of 187 individuals from across the study area (Figure 2). Generally, single whales were recorded in each sighting (n=116), except for some groups consisting of between two to five whales (n=33), which were mostly observed in May. Sightings varied each month from a minimum of 32 in March to a maximum of 77 in April. The number of fin whale individuals sighted per month ranged from 42 to 87. April had the highest number of fin whales recorded (Figure 2). Fin whales were also photographed off Barcelona and Tarragona Ports in the presence of vessels on eight occasions during the season from March to May (Supplementary Figure S1).
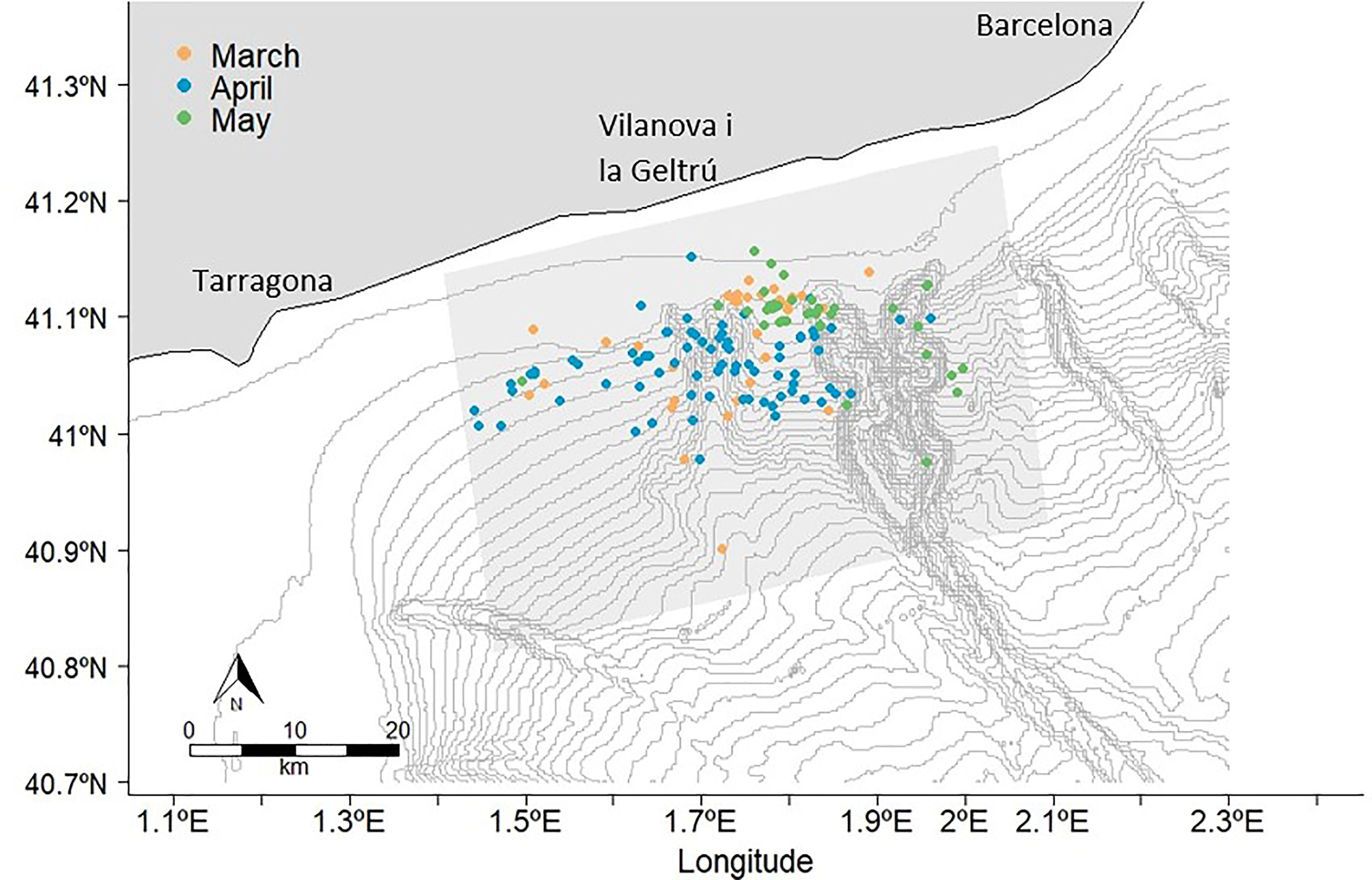
Figure 2 Fin whale sightings between March-May 2021. The bathymetry lines have an interval of 50 meters.
The percentage of fin whales observed that were engaged in surface feeding behaviour was 71% of observations, whilst 9% was travelling behaviour and 4% resting behaviour. For the remaining 16% of the sightings, the behaviour of the whale was undefined. In the first 10 m of the water column, 17 whales were filmed feeding in the area by the drone. Most feeding behaviours (55%) were observed in the evening. Resting behaviour was observed on six occasions.
3.2 Fin Whale Distribution
The selected monthly model was a GAM model using Poisson family distribution and log link function, fitted to investigate the relationship between the logarithm of the number of sightings (response variable NS) and the oceanographic variables mentioned above. This model included a multidimensional isotropic smooth function of the Chla, plus the smooth interaction between Chla and SST and the interaction between Chla and slope. The linear effect of depth and coastal correction factor, adjusted by the effort in a logarithmic scale was added. This is the resulting model (R code can be found in Supplementary Figure S2):
All the tested parameters were significant, except for the coastal correction (CC). The model had a 57.9% deviance explained. The dataset presented a zero-inflation problem which was observed in the residual plots, but it did not invalidate the model (Supplementary Figure S3). The explanatory variables plots can be found in the Supplementary Figure S4, S5. The main standard deviation of the distribution maps was 0.27, 0.37 and 0.38 for each month. The distribution maps with the standard deviation per grid cell can be found in the supplementary material (Supplementary Figure S6).
Bathymetry was found to be a strong predictor of fin whale habitat preference. The fin whale distribution in March and April was very consistent with a high concentration of sightings on the continental shelf break and between Cunit and Foix canyons as displayed in the distribution model (Figure 3). In May, the distribution started to decrease at the shelf break and moved more offshore to deeper waters (1,000 - 2,000 m) inside the Foix Canyon.
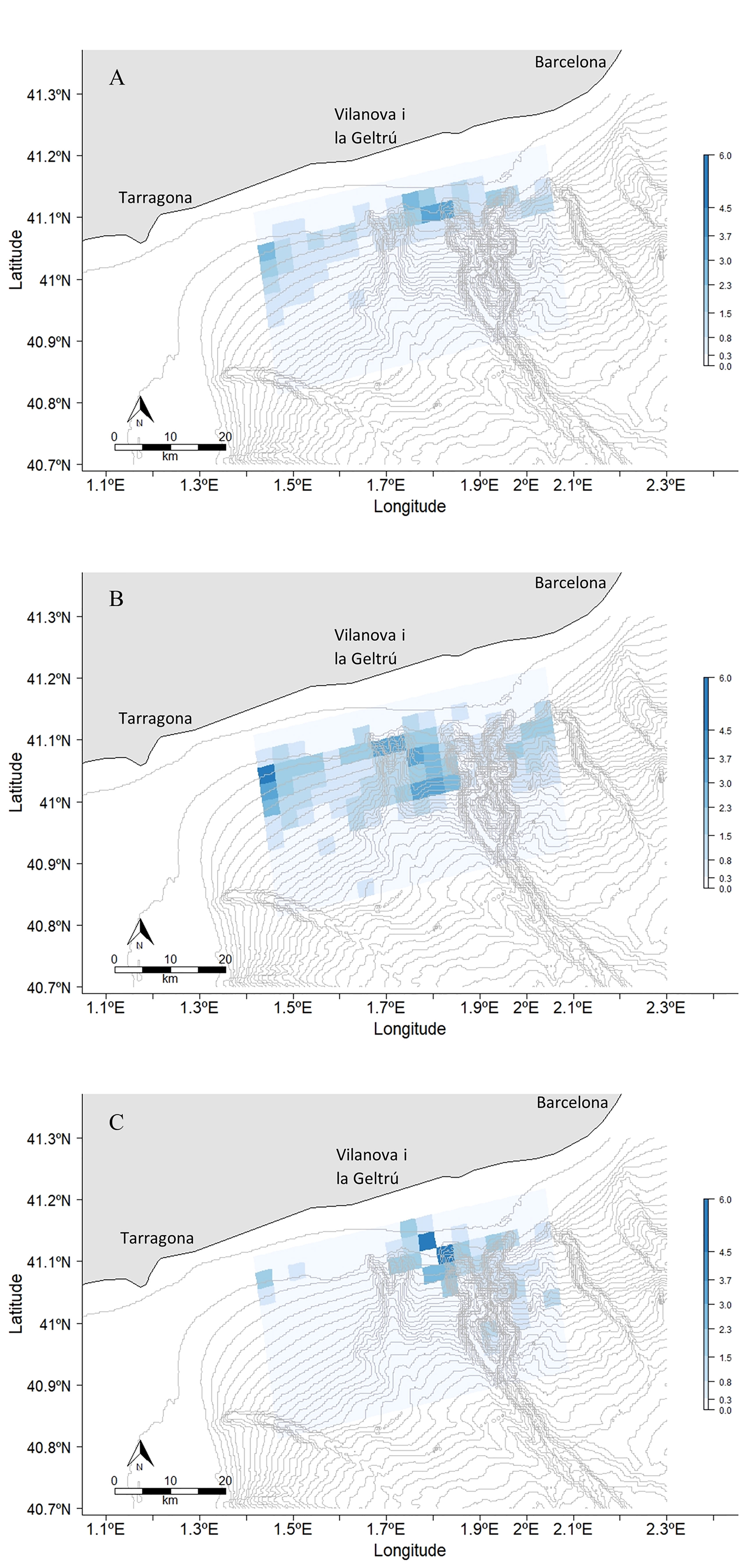
Figure 3 Fin whale distribution in (A) March, (B) April and (C) May 2021 within the study area. The bathymetric lines are drawn every 50 m depth. The Number of animals sighted per pixel varied between 0-6.
3.3 Fin Whale Ship Strike Evidence
Seven whales displaying ship strike injuries were observed. Scars were the most prevalent around the dorsal fin, both in the anterior (n = 2), and in the posterior (n = 1) sides of the fin. On one individual, a lateral scar was observed along its right flank. In addition, a deep cut with scar tissue on the tailstock of one whale was noted. However, the most severe injury recorded was a whale with a dorsal fin collapsed to its right side. Due to this injury, an ‘idiopathic’ scoliosis posterior to the dorsal fin on its right side was developed (Figure 4) (Alves et al., 2017; de Reuver et al., 2021). This injury compromised its swimming speed and diving abilities, causing it to lift the right side of its tail fluke out of the water while diving. It was in a very poor nutritional condition with its vertebral column visible (Castrillon and Bengtson Nash, 2020) (Supplementary Figures S7-S10). In the study area, only one other whale was recorded across all seasons, exhibiting injuries in April 2018. This whale also displayed ‘idiopathic’ scoliosis at its dorsal fin, where the vertebral column curved to the right. It was not re-sighted in any other season. Fin whales were seen on six occasions in the presence of cargo and tanker vessels during the 2021 season (Supplementary Figure S11). Two individuals were seen outside Barcelona Port on 19 May and one whale was seen off Tarragona Port on 10 May (Supplementary Figure S11).

Figure 4 A fin whale displaying severe damage to its vertebral column and dorsal fin on 4 April 2021. (A) lateral and (B) overhead view.Photographs by Dr. Eduard Dellogada.
Four fin whales have been recorded as ship strike fatalities in Barcelona Port since 1986 (Table 1). In 1986, two whales were found on the bow of vessels entering the port. One male whale was brought in by storm conditions on 9th of May 2002, displaying abraded skin on its central left flank posterior to the pectoral fin. Moreover, a male individual was located floating in the port in 2016. The skin along the juvenile male whale’s left flank had been removed in an acute area, with a depression and signs of haemorrhaging (Figure 5). The haemorrhaging on the body was used to indicate that the whale circulation system was still active meaning that the whale was alive when hitting. The location of the collision marks on the body also indicates that they were alive. Orange coloured faeces were found in the gastrointestinal tract indicating that the whale had recently fed on krill prior to being struck.
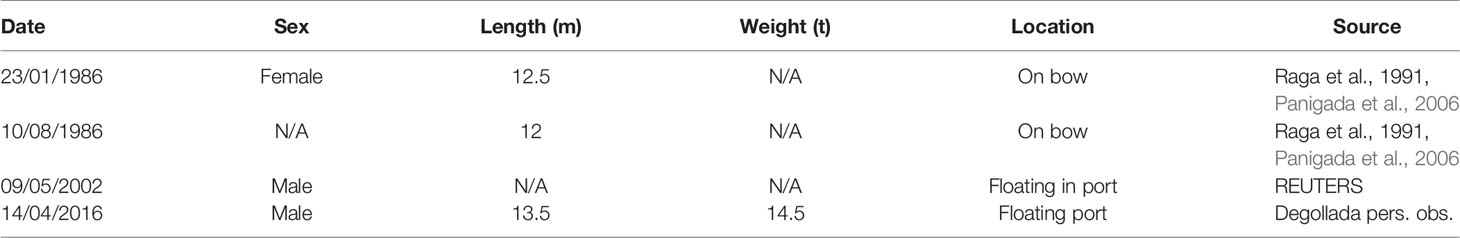
Table 1 Confirmed fin whale ship strikes at Barcelona Port since 1986.

Figure 5 Fin whale in Barcelona Port with evidence of ship strike on its leftright flank on the 14 April 2016. (A) A closeup view of the trauma and (B) an overall view of the whale. Photographs by Dr. Eduard Dellogada.
3.4 Marine Traffic
Barcelona port data was available for cargo, tanker, and passenger vessels from 2014 to 2021. Between 4,468 and 3,587 cargo ships, 852 and 1042 tanker ships and 3869 and 2156 passenger ships visit Barcelona Port annually. Most cargo ships using the port were from foreign cargo vessels (Barcelona Port, 2022). The marine traffic was evaluated during the fin whale season between 2016 and 2021 (Table 2).
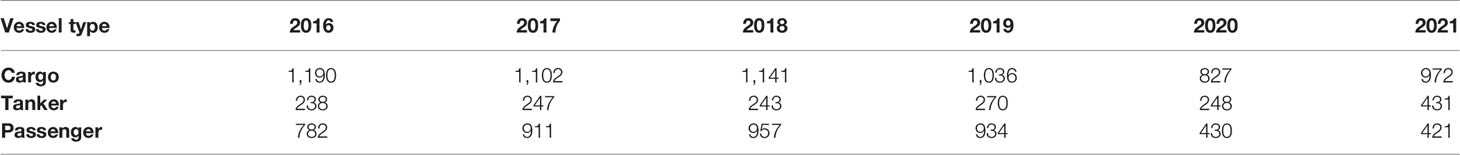
Table 2 Number of vessels that use Barcelona port annually during the fin whale season (from March to May) between 2016 and 2021. The vessel numbers for 2020 are lower due to the COVID-19 pandemic.
Well established shipping lanes were identified transiting the study area. Cargo ships were the most prevalent vessel type across the study area, especially in the northwest section, due to the proximity to the Barcelona Port. The shipping lane splits into three routes that diagonally cross the study area. Three shipping lanes to Tarragona Port in the west of the study area also transit through it (Supplementary Figures S12-S15).
Tanker vessels use both Barcelona and Tarragona Ports. One main shipping lane emerges from each port and diagonally crosses the study area from the northeast and northwest. Cargo and tanker vessels also moved parallel to the coast along the continental shelf edge (Figure 6 and Supplementary Figures S16-S19). Passenger vessels transit through the study area in its southeast corner (Figure 6 and Supplementary Figures S20-S23). This lane moves southwards towards Ibiza in the Balearic Islands (OceanCare, 2021).
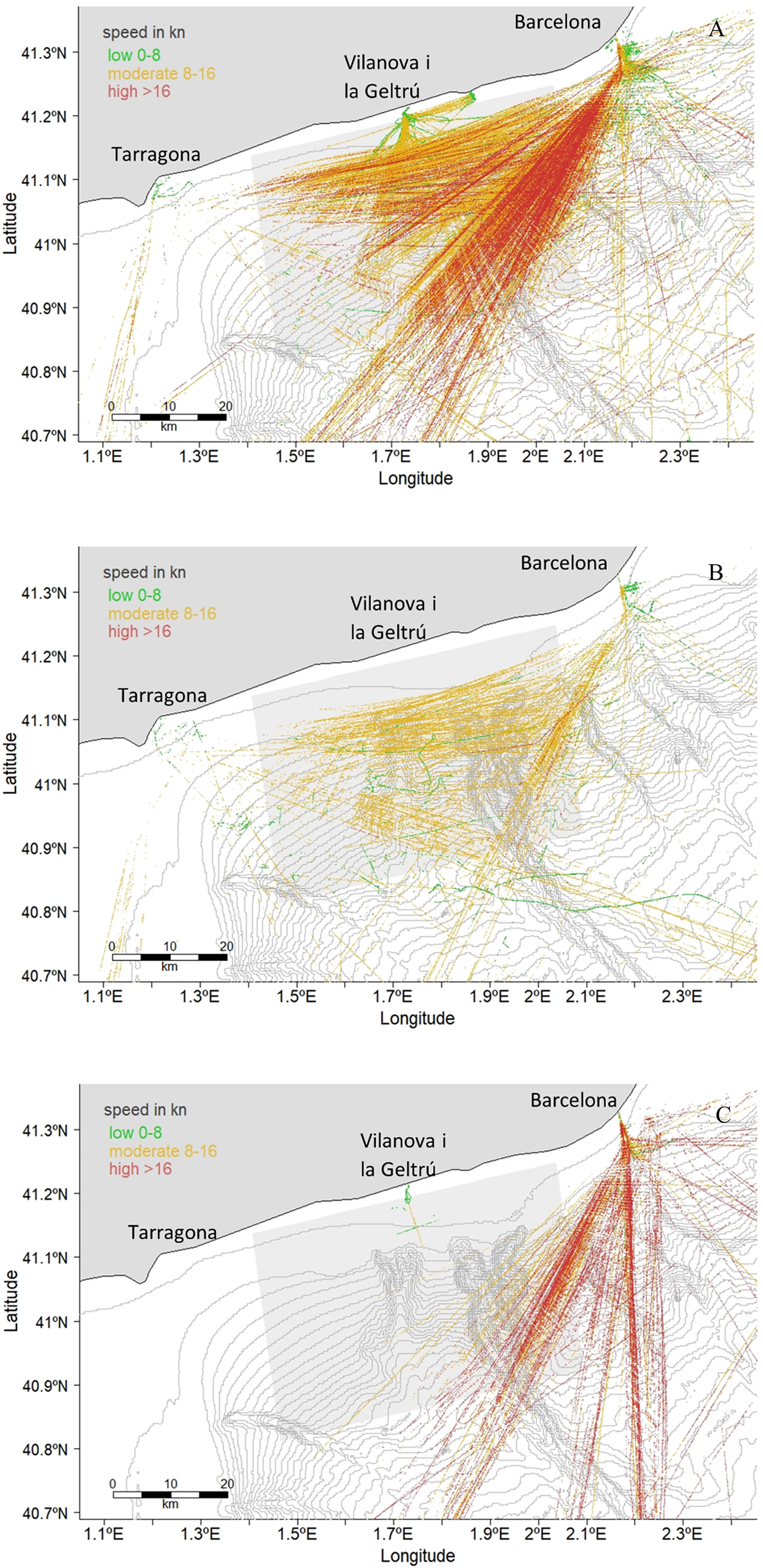
Figure 6 Vessel positions for Cargo (A), Tanker (B) and Passenger (C) vessels in April from pooled AIS data between 2011 and 2020. The bathymetry is represented by isobaths every 50 m and the study area is shaded.
3.5 Shipping Speed
The vessel speed distribution and the comparison of medians between years were significantly different due to the reception variability of the LAB AIS antenna. The p-values of the Kolmogorov-Smirnoff, the Mann-Whitney-U and the Kruskal-Wallis test, were p < 0.001. Within each year, there was also a significant difference (p < 0.001) in the vessel speed between months (Figure 7 and Supplementary Figure S24). In years when the antenna range was stable, there was a consistent pattern where the vessel speed increased from February to June. However, vessel speed was consistent across months for the different types of vessel or sizes (Supplementary Figure S25). The marine traffic lanes with the highest speed vessels that exited from or arrived at Barcelona Port were cargo and passenger vessels (Figures 6, 7 and Supplementary Figures S20-S23, S26-S29). Both vessel speed patterns, by vessel type and by vessel length, were consistent for all the months (Figure 7 and Supplementary Figures S26-S29). Cargo vessels were the most numerous vessel types that moved through the Garraf coast from February to June. The same number of tanker and passenger vessels were recorded during this five-month period. Despite the lowered numbers present, passenger vessels exhibited the highest speeds through the study area, followed by cargo ships and tankers. Approximately 80% of vessels that exceeded 20 kn were passenger vessels, travelling through the southeast corner of the study area (Figure 6). In addition, vessels between 200 and 400 m in length exhibited the highest speeds, exceeding a speed median threshold of 15 kn most frequently (Figure 7 and Supplementary Figures S26-S29).
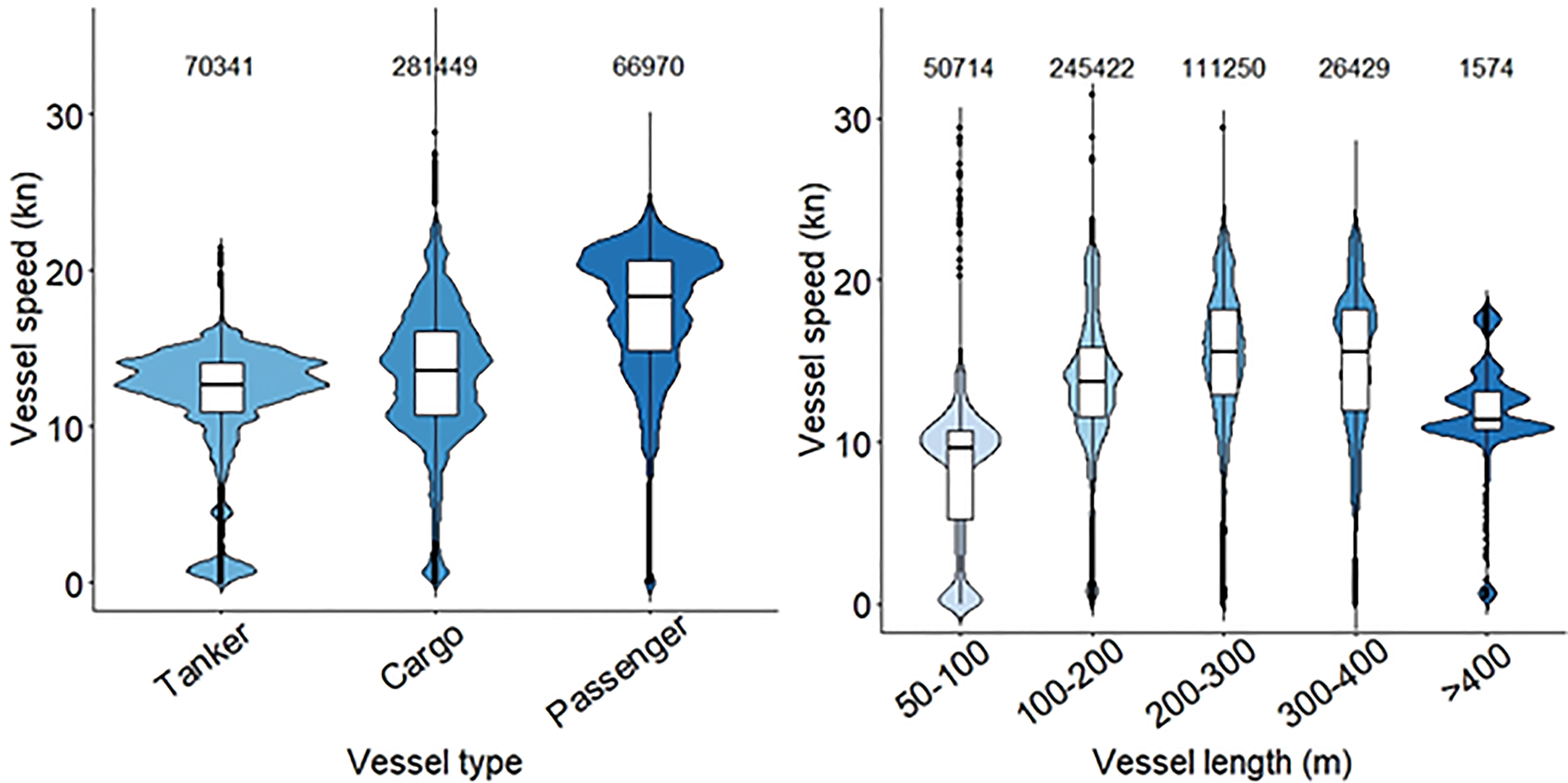
Figure 7 The distribution of vessel speeds in April (from pooled AID data between 2011 and 2020) by vessel type and vessel length in the study area, where fin whales were observed surface feeding. The number of unique vessels in each category is indicated at the top of each violin plot.
3.6 Shipping Density
Cargo vessels displayed the highest densities through the study area in all months and peaked in May. The vessel route density went from 9.75 to 10.97 with a maximum of 50.56 in April. For tanker vessels the mean route went from 3.99 to 4.26 with a maximum value of 15.89 in March. Passenger ships were the ones with the lowest density in the study area with a mean route between 0.12 and 0.19. In this case the maximum number of vessel routes per pixel was 1.9 in May (Figure 8 and Supplementary Figures S30-S32).
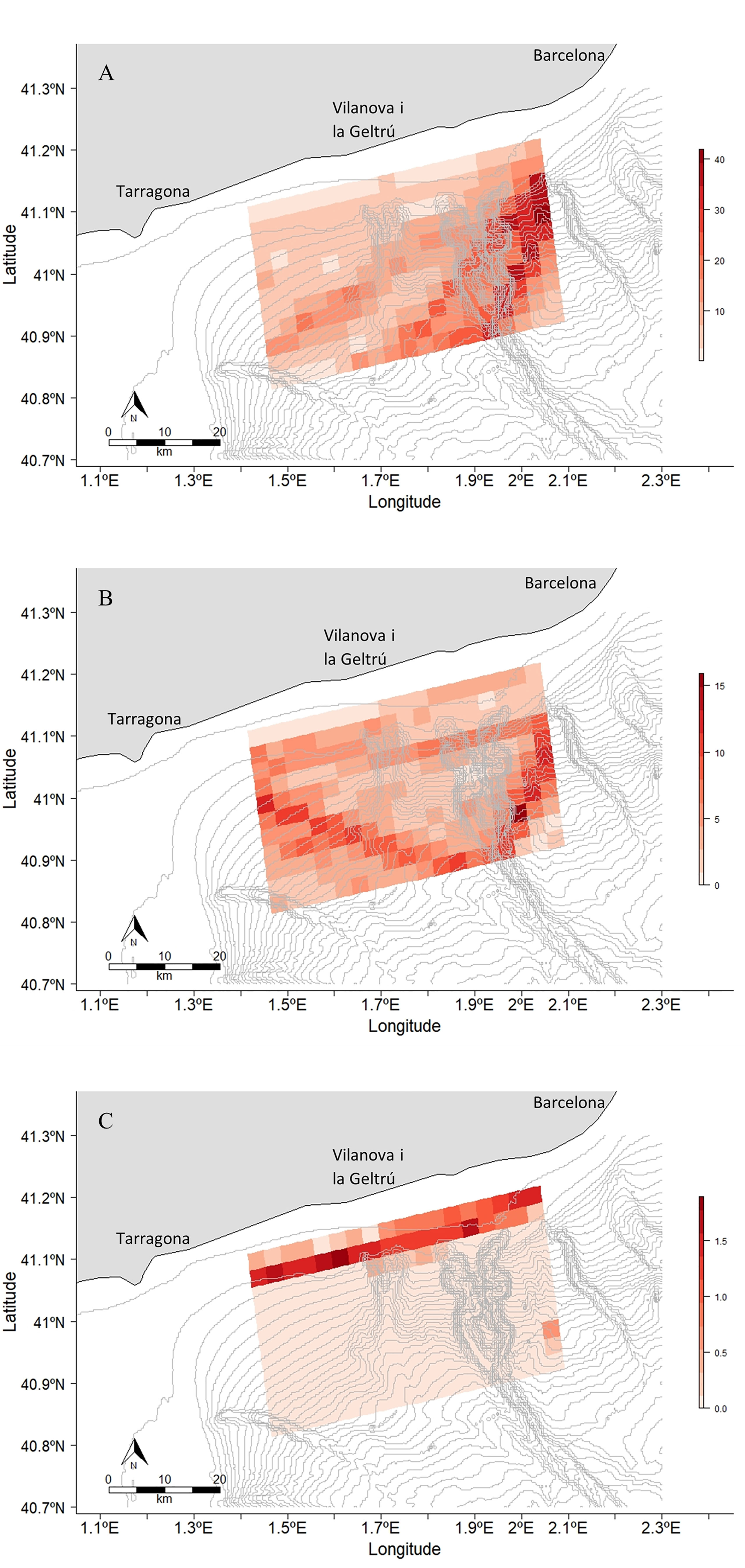
Figure 8 Vessel route density maps within the study area in 2021. The first map (A) represents the cargo ship route density in May, the second (B) represents the tanker ship route density in March and the third (C) one represents the passenger ship route density in May. These maps represent the months with highest marine traffic for each type of vessel. The maps of the months left can be found in the Supplementary Figures S29-S31.
3.7 Ship Strike Assessment
The collision risk maps performed (Figure 9 and Supplementary Figures S33-S35) indicate that April is the month with highest risk of vessel collision at a mean risk of 0.066. On the opposite, May had the lowest risk recorded with a mean value of 0.031 in the study area. The area with the highest collision risk is the left side of the Foix Canyon, because of the proximity to Barcelona port.
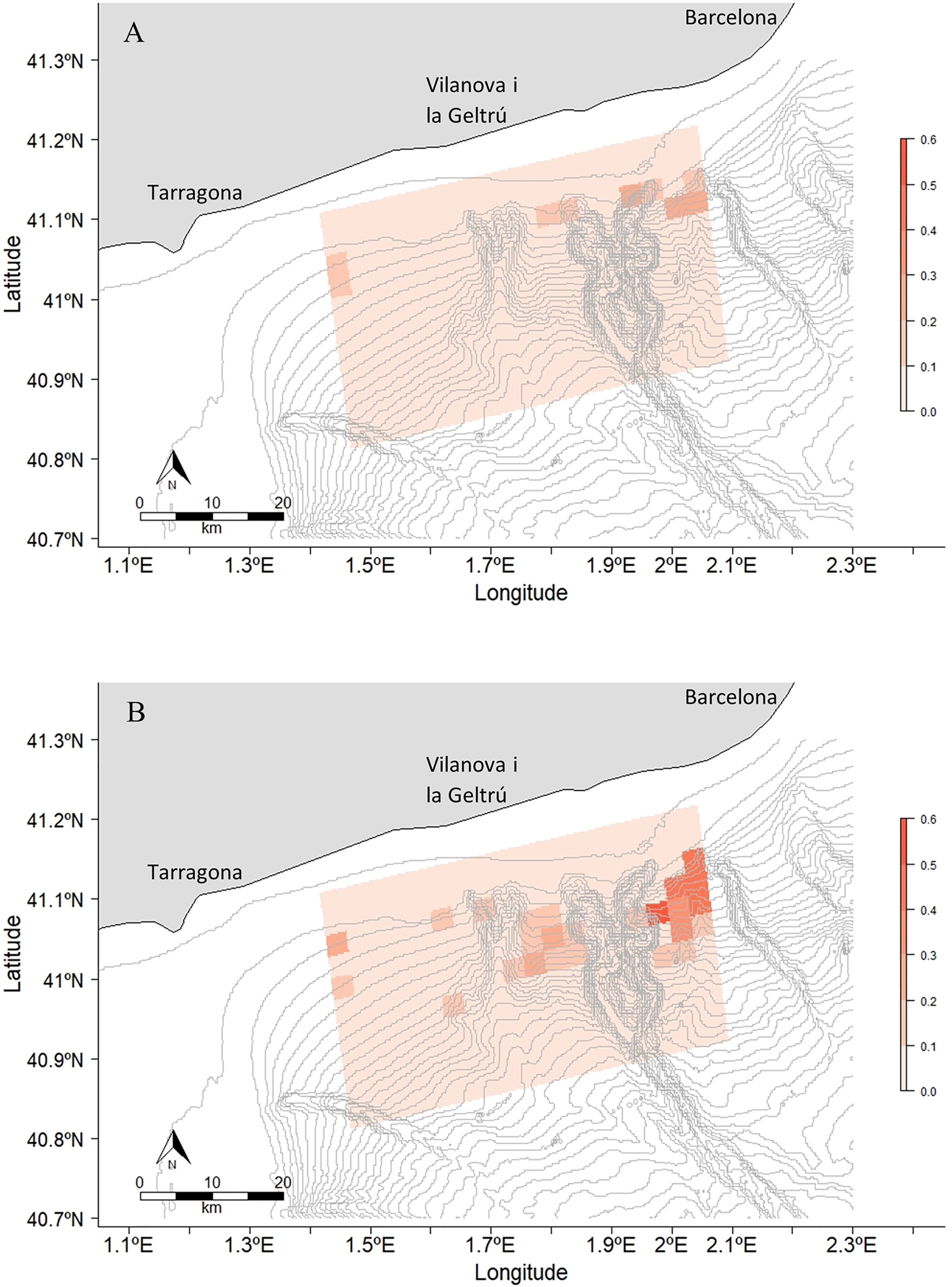
Figure 9 Risk of collision between a fin whale and a cargo ship in (A) March and (B) April. The risk range is between 0 and 0.6. April was the month with the highest risk. The other maps with the risk of collision with cargo ships in May, with tanker ships and with passenger ships can be found in the Supplementary Figures S32-S34.
4 Discussion
4.1 Fin Whale Ship Strike and Fatalities
The whales photographed in the study area generally exhibited ship strike injuries high in the dorsal side, which is the most raised body part when the animal dives. This was especially noted when the whale dove quickly (Laist et al., 2001). The injuries were similar to those reported elsewhere in the Mediterranean and Strait of Gibraltar (de Stephanis and Urquiola, 2006; Panigada et al., 2006; Gauffier et al., 2018). The aforementioned two cases of severe vertebral column deformity and likely idiopathic scoliosis in 2018 and 2021, were determined as severe welfare cases. In these, the whales were unable to swim correctly or at typical speeds, which reduced their capacity to feed efficiently (Goldbogen et al., 2006; Gough et al., 2019; de Reuver et al., 2021). Neither animal was subsequently resighted during the following seasons (2018 whale) or the remaining 2021 season (2 months).
Nine fin whales (out of 383) were photo-identified with ship strike injury evidence in the Ligurian Sea from 1990 to 2001 (Panigada et al., 2006). Only one fin whale was photographed with ship strike evidence off the Garraf coast prior to 2021, when 6 individuals were documented. This sudden increase in visual marks from ship strikes may indicate that the risk posed from this threat has increased. The four confirmed ship strike fatalities in Barcelona Port displayed typical ship strike characteristics for the species with dorsal side collision marks. Two fatal ship strikes occurred during the fin whale season (April and May) from the data available in 2002 and 2016. These whales were still young given their length range was between 12 and 14.5 m. These results are limited and there have likely been more whale strandings associated with ship strikes in proximity to Barcelona Port as indicated by the concentration of strandings on Mediterranean Database of Cetacean Strandings (MEDACES). Further work is required in evaluating the strandings from this location to assess past ship strike occurrence in the area.
Fin whales traveling in groups in the Bay of Biscay displayed lowered attentiveness rates in proximity to ferries, which may increase the risk of ship strikes (Aniceto et al., 2016). This “group effect”, the slower traveling speed of mother-calf pairs (6.6 km/hr in one study) and increased time at the surface, may increase the susceptibility of these whales to ship strikes in the study area and in the vicinity of Barcelona Port (Aniceto et al., 2016; Smultea et al., 2017). The low survival rate of calves in the Mediterranean Sea and that additional loss of sexually mature females with calves poses a high threat to the species (Panigada et al., 2006; Arrigoni et al., 2011). Poor nutritional condition and maternal separation was attributed to one live stranded calf that was euthanised and necropsied at Tarragona on 13th November 2015 after appearing in Barcelona Port on 10th/11th November (Cuvertoret-Sanz et al., 2020). Only four mother-calf pairs have been recorded present in the study area since the project began in March 2014 (EDMAKTUB unpublished data).
4.2 Fin Whale Ship Strike Risk on the Garraf Coast
Ship strike risk appears to be higher in April when the number of fin whale sightings is highest. May was the month with less risk despite it having the highest marine traffic levels. The lower risk may be due to the offshore movements of fin whales in May. The study area´s lower risk cannot be extrapolated to other close-by zones, given the proximity of Barcelona Port, the surrounding traffic routes, and the changes in oceanographic conditions along the Catalan Coast, as reported by the fisherman along seasons. Future studies are required to evaluate the whole Catalan Coast and the Balearic Sea with fin whale data in order to assess the risk of collision in other areas.
Fin whales follow a complex migratory pattern subject to food availability in the Mediterranean Sea (Geijer et al., 2016; Notarbartolo di Sciara et al., 2016; Panigada et al., 2017). The relationship between their presence and oceanographic variables (which act as indicators of productivity and consequently to food abundance) do not follow simple linear regressions. For this reason, one of the most used models for cetacean distribution is the GAM model. It offers a flexible and robust approach for the exploration and characterization of complex, non-linear relationships among variables (Panigada et al., 2008; Zerbini et al., 2015). The oceanographic model used to fit this model used Chla in order to get the primary production indicator and the SST as the indicator for column mixing which implies an enrichment of nutrients in superficial waters which implies more productivity (Croll et al., 2005; Baines and Reichelt, 2014).
The bathymetry, depth and continental shelf slope factors appear to be an important factor to correlate with chlorophyll a and sea surface temperature, as they determine where krill occurs. Northern krill is a mesopelagic species which perform a diurnal-vertical migration to feed on phytoplankton at night (Panigada et al., 1999; Kaartvedt, 2010). Fin whale surface feeding behaviours, especially lunge feeding, usually occur at dusk for this reason which was recorded often off the Garraf coast. Fin whales also remain at the surface for extended periods of time, swim slower and shallower at night (Calambokidis et al., 2019; Keen et al., 2019). These behaviours may reduce their reaction times while the crew of vessels cannot visually see animals in the dark, which substantially increases the risk of collision (Calambokidis et al., 2019; Keen et al., 2019; Guazzo et al., 2021).
In the Ligurian Sea, fin whales primarily feed in habitats reaching 2,000 m in depth and can dive to reach ~470 m to feed (Panigada et al., 1999; Panigada et al., 2005). The deep foraging dives performed in the Ligurian Sea, reduce the time that fin whales spend at the surface. 82% of the Mediterranean Sea’s fatal ship strikes were recorded between 1972 and 2001 in the Ligurian Sea area (Panigada et al., 2006). However, the shallower surface feeding behaviours exhibited by fin whales along the Catalan coast increases their presence at the surface exponentially. Surface lunge feeding in the Mediterranean has been documented off Lampedusa Island (Canese et al., 2006). This fact may expose the whales in this region to a higher risk of collision, given the whales are closer to the surface for extended periods of time, while both feeding and resting.
The intensity and visibility of fin whale blows appears to vary depending on their behaviour and the environmental conditions (Stone et al., 1992; Kopelman and Sadove, 1995; Goldbogen et al., 2006; Horton et al., 2019; Martins et al., 2020). The blow intensity may also be affected by the time of the day within the study area (Stone et al., 1992). This variation in blow visibility would influence the detection rate of whales by the crew of marine vessels.
The persistent acoustic noise from vessels has been recorded altering fin whale behaviour (Nieukirk et al., 2011; Castellote et al., 2012b). Acoustic calls to avoid masking caused by shipping noise, have also resulted in displacement behaviours while foraging in the Mediterranean Sea (Castellote et al., 2012b; Notarbartolo di Sciara et al., 2016). Adaptation to the increased noise levels within the Mediterranean Sea may make the species more susceptible to ship strikes (Castellote et al., 2012b; Notarbartolo di Sciara et al., 2016).
Of the 43 near-miss events recorded in the Ligurian Sea between 2008 and 2019, 63.4% of whales surfaced in front of ferries <50 m away (David et al., 2022). No behavioural changes were observed when whales were travelling or resting in proximity to ferries, indicating that the acoustic output of marine traffic is not suitable for the species as an avoidance signal for marine vessels (David et al., 2022). The animals observed in the study area did not show any behavioural changes that could be attributed to the presence of vessels. The species spatial awareness alters depending on their behaviour and group size which may make them more susceptible to being hit while engaging in behaviours such a surface feeding and resting (Friedlaender et al., 2015; Aniceto et al., 2016; Panigada et al., 2017; Calambokidis et al., 2019; Irvine et al., 2019). The near surface feeding behaviours recorded in the Garaff study area during the evening time, when krill migrate higher in the water column, have the potential to increase the risk of collision. During the hours of darkness whales would not be detected by visual observation unless by thermal imaging (Zitterbart et al., 2020).
Cargo vessels pose the highest ship strike risk to fin whales on the Garraf coast. They transit through important foraging areas at speeds between 10 to 20 kn, reaching a probability of having a fatal strike to around 79% (Vanderlaan and Taggart, 2007; Conn and Silber, 2013). Half of global ship strikes occur at moderate vessel speeds (below 15 kn) (Winkler et al., 2020). Death is possible even at low speeds (Kelley et al., 2021). Tanker ships recorded the slowest mean speed at around 10 kn. They crossed the study area from both sides, due to the presence of Tarragona and Barcelona Ports. In this case, even though the traffic density was low, the collision risk was high for this type of vessel, due to its prevalence in crossing the study area.
While passenger’s vessels did not display a high risk in the study area (because they transit through a small section of it), they do represent a major threat towards the species along the Catalan coast and in the Balearic Sea. They move at very fast speeds, averaging 20 kn regularly between Barcelona and the Balearic Islands, France, and Italy. In the Ligurian Sea, ship strike risk increases during the summer “high tourist season” months, when ferries move between the French and Italian mainland to Corsica and Sardinia, while fin whales forage in the area (Grossi et al., 2021; Ham et al., 2021). Pleasure boats operate in coastal waters of the study area and may move at fast speeds (>20 kn), however, it is predominantly sailing boats that occur over the Cunit and Foix Canyons due to the distance from shore. Nonetheless, sailing boats may pose a reduced risk due to manoeuvrability limitations when the sails are raised in some environmental conditions (Ritter, 2012). Further study of tourism season threat and impact on the fin whales is required off the Catalan coast for ferries and pleasure boats such as yachts (Carreño and Lloret, 2021).
4.3 Mitigation Measures to Reduce Ship Strike Risks
Ship strikes are a huge conservation concern for cetacean species globally, especially for large whales (Laist et al., 2001; Sèbe et al., 2019; Winkler et al., 2020). For this reason, a variety of mitigation measures have been implemented to minimise the risk of ship strikes worldwide (Panigada et al., 2006; Winkler et al., 2020). The most common measures used are speed reductions and the implementation of traffic separation schemes (TSS) (Guzman et al., 2013; Bezamat et al., 2015; Guzman et al., 2020; OceanCare, 2021). For example, the chances of lethal injury from ship strikes drops to 50% at 8.8 kn speed (Vanderlaan and Taggart, 2007; Conn and Silber, 2013). Thus, a speed limitation to 8 kn, along the entire Catalan coast under 200 m depth and in the areas closest to the continental shelf edge and submarine canyons between February and June, would greatly protect fin whale habitats over a wide spatial area. Moreover, slower vessel speeds would aid climate action efforts in reducing the amount of CO2 emissions being released by marine traffic (Leaper, 2019).
In order to prevent collisions with the critically endangered north Atlantic right whale (Eubalaena glacialis) (Müller, 1776) in the Gulf of Maine, the IMO subcommittee on ship strikes implemented a measure forcing ships larger than 300 gross tons to report their position, speed and destination while entering key right whale habitats: New England (foraging area) and coastal Georgia and Florida (calving area) (Conn and Silber, 2013; Cooke, 2020). Reporting ships receive an automatically generated message that provides information about right whale sightings, their vulnerability, and actions to avoid ship strikes. In addition, in the eastern Mediterranean Sea, re-routing of shipping lanes by 20 km in the Hellenic Trench, Greece was predicted to decrease the ship strike risk to sperm whales (Physeter macrocephalus) (Linneaus 1758) by 70%, with no additional cost to shipping companies transiting the area (Frantzis et al., 2019).
The use of fin whale distribution models influenced by oceanographic conditions would be useful to implement flexible regulation measures, considering that fin whale presence and behaviour is affected by foraging conditions along the Garraf coast. Marine traffic has been seasonally diverted away from areas where ship strike risk increased greatly during certain months in other countries; it has been introduced at New Scotia and Gulf of Maine, Canada coast and the Gulf of Panama (Vanderlaan and Taggart, 2007; Guzman et al., 2020). Other suggested measures include vessels moving offshore to avoid crossing known foraging habitats once they departed port. The continental shelf edge should be crossed during daylight hours, while the vessel’s master or a member of the crew actively watch for fin whales and evade any sighted animals at a far distance. This would increase the chances of the ship to avoid ship strike or collision (Gende et al., 2019). This mitigation measure could be implemented in the proposed TSS of Barcelona and Tarragona Ports.
In other areas out of the Mediterranean Sea, Marine Mammal Observers (MMO) accompany the vessels during the spring season, to help locate fin whales in the path of the vessel, and therefore, helping to avoid ship strikes (Flynn and Calambokidis, 2019; Zitterbart et al., 2020). The use of Infrared cameras may alternatively be used to detect whale blows and their body heat signature while at the surface (Zitterbart et al., 2020). Zitterbart et al. (2020) reported a 70% detection rate of whales within 2 km of vessels in Australia. Detections were also observed at night while the whales feed or rest near the surface (Calambokidis et al., 2019; Keen et al., 2019). This technology would be very useful in the Garraf study area given the area’s dusk, and the night-time fin whale foraging behaviour. The efficiency of this technique for fin whales should be evaluated in this region.
Funding should be provided to survey the Balearic Sea, particularly within the Mediterranean cetacean Migration Corridor and in the new Specially Protected Areas of Mediterranean Importance (SPAMI) area. In order to provide conservation managers with more robust data and aid in future studies about the ship strike risk for fin whales species, additional population and abundance data on fin whales within the Balearic Sea during springtime is required. Previous studies in the Balearic Sea using dedicated aerial and opportunistic boat-based surveys indicate the presence of fin whales elsewhere in the Balearic Sea. Further research is required to understand their seasonality and movement patterns (Torreblanca et al., 2019; ACCOBAMS, 2021). Consideration must also be made for the use of acoustic monitoring techniques along the Catalan coast and in the SPAMI area, in order to evaluate the noise pollution under the Marine Strategy Framework Directive descriptor 11. Evaluating this anthropogenic noise would provide opportunities to detect the cetacean presence and seasonality (including sperm whales and Cuvier’s beaked whales (Ziphius cavirostris) (Cuvier, 1823) and to monitor this region’s cetacean communities further (OceanCare, 2021).
In light of the high abundance of fin whales seasonally feeding off the Garraf coast of Catalonia and the high volume of marine traffic transiting the area, careful consideration is required to reduce the ship strike risk to this species. The endangered status of the Mediterranean fin whale population warrants an urgent need to minimise the loss of individuals from the population through anthropogenic impacts such as ship strikes. The Royal Decree protecting fin whales in Spanish waters should engage with and enforce conservation measures for this species from ship strike risk along the Mediterranean coast. The Barcelona and Tarragona Port TSS provides a unique opportunity to develop a plan with stakeholder engagement, to implement the recommendations made in this study and limit the loss of fin whales.
Data Availability Statement
The marine traffic data used in this publication was made available by the EMODnet Human Activities project, www.emodnet-humanactivities.eu, funded by the European Commission Directorate General for Maritime Affairs and Fisheries. The marine traffic AIS data used in this publication was made available by the Laboratorio de Aplicaciones Bioacústicas of the Polytechnic University of Catalonia. This data is not available to download. The bathymetric data used in this publication was made available by the EMODnet Bathymetry project, https://www.emodnet-bathymetry.eu, funded by the European Union under Regulation (EU) No 508/2014 of the European Parliament and of the Council of 15 May 2014 on the European Maritime and Fisheries Fund. The oceanographic data used in this publication was made available by the Marine Copernicus, (https://marine.copernicus.eu) funded by the European Commission (EC) and implemented by Mercator Ocean International.
Ethics Statement
The animal study was reviewed and approved by the Ministry of Ecological Transition Spain, under the ministry authorization: DGBBD/BDM/AUTSPP/17/2021.
Author Contributions
EDB, BC and SO’C assisted with the data collection. SO’C, BC and RG drafted the manuscript. BC conducted the main analysis. RG assisted and analysed the LAB AIS data. SO’C compiled present and historical collision evidence. All authors contributed perspectives and assisted with manuscript review.
Funding
The fin whale project was funded by the EDMAKTUB association with the collaboration of Ports de la Generalitat and Club Nàutic de Vilanova.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Thanks to the commercial fishermen from Ametlla de Mar to Palamós for supporting the project and to the volunteers and collaborators who make the fin whale project possible.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.867287/full#supplementary-material
References
ACCOBAMS (2021). Estimates of Abundance and Distribution of Cetaceans, Marine Mega-Fauna and Marine Litter in the Mediterranean Sea From 2018-2019 Surveys. Eds. Panigada S., Boisseau O., Canadas A., Lambert C., Laran S., McLanaghan R., Moscrop A. (Monaco: ACCOBAMS - ACCOBAMS Survey Initiative Project), 177 pp.
Aguilar A., García-Vernet R. (2018). “Fin Whale Balaenoptra Physalus,” in Encyclopaedia of Marine Mammals 3rd Edition. Eds. Würsig B., Thewissen J. G. M., Kovacs K. (Royal Society). Available at: 10.1098/rspb.2009.1135.
Alves F., Towers J. R., Baird R. W., Bearzi G., Bonizzoni S., Ferreira R., et al. (2017). The Incidence of Bent Dorsal Fins in Free-Ranging Cetaceans. J. Anat. 232, 263–269. doi: 10.1111/joa.12729
Aniceto A. S., Carroll J. L., Tetley M. J., van Oosterhout C. (2016). Position, Swimming Direction and Group Size of Fin Whales (Balaenoptera Physalus) in the Presence of a Fast-Ferry in the Bay of Biscay. Oceanologia 57, 235–240. doi: 10.1016/j.oceano.2016.02.002
Arrigoni M., Manfredi P., Panigada S., Bramanti L., Santangelo G. (2011). Life-History Tables of the Mediterranean Fin Whale From Stranding Data. Marine Ecol. 32, 1–9. doi: 10.1111/j.1439-0485.2011.00437.x
Baines M. E., Reichelt M. (2014). Upwellings, Canyons and Whales: An Important Winter Habitat for Balaenopterid Whales Off Mauritania, Northwest Africa. J. Cetacean Res. Manage. 14, 57–67.
Barcelona Port (2022) Port Authority Statistics [WWW Document]. Available at: https://www.portdebarcelona.cat/es_ES/web/autoritat-portuaria/estadisticas.
Bezamat C., Wedekin L. L., Simões-Lopes P. C. (2015). Potential Ship Strikes and Density of Humpback Whales in the Abrolhos Bank Breeding Ground, Brazil. Aquat. Conserv.: Marine Freshwater Ecosyst. 25, 573–586. doi: 10.1002/aqc.2523
Borrell A., Gazo M., Aguilar A., Raga J. A., Degollada E., Gozalbes P., et al. (2021). Niche Partitioning Amongst Northwestern Mediterranean Cetaceans Using Stable Isotopes. Prog. Oceanogr. 193, 102559. doi: 10.1016/j.pocean.2021.102559
Calambokidis J., Fahlbusch J. A., Szesciorka A. R., Southall B. L., Cade D. E., Friedlaender A. S., et al. (2019). Differential Vulnerability to Ship Strikes Between Day and Night for Blue, Fin, and Humpback Whales Based on Dive and Movement Data From Medium Duration Archival Tags. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00543
Canese S., Cardinali A., Fortuna C. M., Giusti M., Lauriano G., Salvati E., et al. (2006). The First Identified Winter Feeding Ground of Fin Whales (Balaenoptera Physalus) in the Mediterranean Sea. J. Marine Biol. Ass. 86, 903–907. doi: 10.1017/S0025315406013853
Carreño A., Lloret J. (2021). Environmental Impacts of Increasing Leisure Boating Activity in Mediterranean Coastal Waters. Ocean Coastal Manage. 209, 105693. doi: 10.1016/j.ocecoaman.2021.105693
Castellote M., Clark C. W., Lammers M. O. (2012a). Fin Whale (Balaenoptera Physalus) Population Identity in the Western Mediterranean Sea. Marine Mammal Sci. 28, 325–344. doi: 10.1111/j.1748-7692.2011.00491.x
Castellote M., Clark C. W., Lammers M. O. (2012b). Acoustic and Behavioural Changes by Fin Whales (Balaenoptera Physalus) in Response to Shipping and Airgun Noise. Biol. Conserv. 147, 115–122. doi: 10.1016/j.biocon.2011.12.021
Castellote M., Esteban J. A., Clark C. W. (2008). Fin Whale (Balaenoptera Physalus) Movements Along the Spanish Mediterranean Coast. J. Acoustical Soc. America 123, 3775. doi: 10.1121/1.2935401
Castrillon J., Bengtson Nash S. (2020). Evaluating Cetacean Body Condition; a Review of Traditional Approaches and New Developments. Ecol. Evol. 10, 6144–6162. doi: 10.1002/ece3.6301
Conn P. B., Silber G. K. (2013). Vessel Speed Restrictions Reduce Risk of Collision-Related Mortality for North Atlantic Right Whales. Ecosphere 4(4): 43. doi: 10.1890/ES13-00004.1
Cooke J. G. (2018). Balaenoptera Physalus, Fin Whale. The IUCN Red List of Threatened Species. e.T2478A50349982. doi: 10.2305/IUCN.UK.2018-2.RLTS.T2478A50349982.en
Cooke J. G. (2020). Eubalaena Glacialis, North Atlantic Right Whale. The IUCN Red List of Threated Species. e.T41712A178589687. doi: 10.2305/IUCN.UK.2018-1.RLTS.T41711A50380694.en.
Croll D. A., Marinovic B., Benson S., Chavez F. P., Black N., Ternullo R., et al. (2005). From Wind to Whales: Trophic Links in a Coastal Upwelling System. Marine Ecol. Prog. Ser. 289, 117–130. doi: 10.3354/meps289117
Cuvertoret-Sanz M., López-Figueroa C., O’Byrne A., Canturri A., Martí-Garcia B., Pintado E., et al. (2020). Causes of Cetacean Stranding and Death on the Catalonian Coast (Western Mediterranean Sea), 2012-2019. Dis. Aquat. Org. 142, 239–253. doi: 10.3354/DAO03550
David L., Alleaume S., Guinet C. (2011). Evaluation of the Potential of Collision Between Fin Whales and Maritime Traffic in the North-Western Mediterranean Sea in Summer, and Mitigation Solutions. JMATE 4, 17–28.
David L., Arcangeli A., Tepsich P., Di-Meglio N., Roul M., Campana I., et al. (2022). Computing Ship Strikes and Near Miss Events of Fin Whales Along the Main Ferry Routes in the Pelagos Sanctuary and Adjacent West Area, in Summer. Aquat. Conserv. 32 (3), 442–456. doi: 10.1002/aqc.3781
de Reuver S., IJsseldijk L. L., Homans J. F., Willems D. S., Veraa S., van Stralen M., et al. (2021). What a Stranded Whale With Scoliosis Can Teach Us About Human Idiopathic Scoliosis. Sci. Rep. 11, 7218. doi: 10.1038/s41598-021-86709-x
de Stephanis R., Urquiola E. (2006). “Collisions Between Ships and Cetaceans in Spain,” in Document Presented to the Scientific Committee of the International Whaling Commission SC/58/BC5.
Di-Meglio N., David L., Monestiez P. (2018). Sperm Whale Ship Strikes in the Pelagos Sanctuary and Adjacent Waters: Assessing and Mapping Collision Risks in Summer. J. Cetacean Res. Manage 18, 135–147.
Druon J.-N. (2014). Monitoring Pelagic Habitats to Support Future EU Policie) - Strategic View on How the Daily Monitoring of Marine Water-Column Habitats Will Support the Future EU Integrated Maritime Policy (MSP, CFP, MSFD) to Stimulate Ecosystem Health and Blue Growth. (Luxembourg: Publications Office of the European Union, 2014)
Druon J. N., Panigada S., David L., Gannier A., Mayol P., Arcangeli A., et al. (2012). Potential Feeding Habitat of Fin Whales in the Western Mediterranean Sea: An Environmental Niche Model. Marine Ecol. Prog. Ser. 464, 289–306. doi: 10.3354/meps09810
EDMAKTUB (2021) Edmaktub. Fin Whale Project Results 2021. Available at: https://www.edmaktub.org/en/results/.
EDMAKTUB (2018) Edmaktub. Fin Whale Project Results 2018. Available at: https://www.edmaktub.org/results/.
E.U. Copernicus Marine Service Information. Mediterranean Sea - High Resolution and Ultra High Resolution L3S Sea Surface Temperature Model. doi: 10.48670/moi-00171
E.U. Copernicus Marine Service Information. Mediterranean Sea - Reprocessed Surface Chlorophyll Concentration Adn Phytoplankton Functional Types From Multi Satellite Observations Model. doi: 10.48670/moi-00112
European Commission Natura 2000 Network in the Marine Environment. Available at: https://ec.europa.eu/environment/nature/natura2000/marine/index_en.htm.
Flynn K. R., Calambokidis J. (2019). Lessons From Placing an Observer on Commercial Cargo Ships Off the U.s. West Coast: Utility as an Observation Platform and Insight Into Ship Strike Vulnerability. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00501
Frantzis A., Leaper R., Alexiadou P., Prospathopoulos A., Lekkas D. (2019). Shipping Routes Through Core Habitat of Endangered Sperm Whales Along the Hellenic Trench, Greece: Can We Reduce Collision Risks? PLoS ONE 14(2):e0212016. doi: 10.1371/journal.pone.0212016
Friedlaender A. S., Goldbogen J. A., Hazen E. L., Calambokidis J., Southall B. L. (2015). Feeding Performance by Sympatric Blue and Fin Whales Exploiting a Common Prey Resource. Marine Mammal Sci. 31, 345–354. doi: 10.1111/mms.12134
Gauffier P., Borrell A., Silva M. A., Víkingsson G. A., López A., Giménez J., et al. (2020). Wait Your Turn, North Atlantic Fin Whales Share a Common Feeding Ground Sequentially. Marine Environ. Res. 155. doi: 10.1016/j.marenvres.2020.104884
Gauffier P., Verborgh P., Giménez J., Esteban R., Sierra J. M. S., de Stephanis R. (2018). Contemporary Migration of Fin Whales Through the Strait of Gibraltar. Marine Ecol. Prog. Ser. 588, 215–228. doi: 10.3354/meps12449
Geijer C. K. A., Notarbartolo di Sciara G., Panigada S. (2016). Mysticete Migration Revisited: Are Mediterranean Fin Whales an Anomaly? Mammal Rev. 46, 284–296 doi: 10.1111/mam.12069
Gende S. M., Vose L., Baken J., Gabriele C. M., Preston R., Noble Hendrix A. (2019). Active Whale Avoidance by Large Ships: Components and Constraints of a Complementary Approach to Reducing Ship Strike Risk. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00592
Goldbogen J. A., Calambokidis J., Shadwick R. E., Oleson E. M., McDonald M. A., Hildebrand J. A. (2006). Kinematics of Foraging Dives and Lunge-Feeding in Fin Whales. J. Exp. Biol. 209, 1231–1244. doi: 10.1242/jeb.02135
Gough W. T., Segre P. S., C.Bierlich K. K., Cade D. E., Potvin J., Fish F. E., et al. (2019). Scaling of Swimming Performance in Baleen Whales. J. Exp. Biol. 222(20), jeb204172. doi: 10.1242/jeb.204172
Grossi F., Lahaye E., Moulins A., Borroni A., Rosso M., Tepsich P. (2021). Locating Ship Strike Risk Hotspots for Fin Whale (Balaenoptera Physalus) and Sperm Whale (Physeter Macrocephalus) Along Main Shipping Lanes in the North-Western Mediterranean Sea. Ocean Coastal Manage. 212, 105820. doi: 10.1016/j.ocecoaman.2021.105820
Guazzo R. A., Durbach I. N., Helble T. A., Alongi G. C., Martin C. R., Martin S. W., et al. (2021). Singing Fin Whale Swimming Behavior in the Central North Pacific. Front. Marine Sci. 8. doi: 10.3389/fmars.2021.696002
Guzman H. M., Capella J. J., Valladares C., Gibbons J., Condit R. (2020). Humpback Whale Movements in a Narrow and Heavily-Used Shipping Passage, Chile. Marine Policy 118, 103990. doi: 10.1016/j.marpol.2020.103990
Guzman H. M., Gomez C. G., Guevara C. A., Kleivane L. (2013). Potential Vessel Collisions With Southern Hemisphere Humpback Whales Wintering Off Pacific Panama. Marine Mammal Sci. 29, 629–642. doi: 10.1111/j.1748-7692.2012.00605.x
Ham G. S., Lahaye E., Rosso M., Moulins A., Hines E., Tepsich P. (2021). Predicting Summer Fin Whale Distribution in the Pelagos Sanctuary (North-Western Mediterranean Sea) to Identify Dynamic Whale–Vessel Collision Risk Areas. Aquat. Conserv.: Marine Freshwater Ecosyst 31, 2257–2277. doi: 10.1002/aqc.3614
Horton T. W., Hauser N., Cassel S., Klaus K. F., de Oliveira T. F., Key N. (2019). Doctor Drone: Non-Invasive Measurement of Humpback Whale Vital Signs Using Unoccupied Aerial System Infrared Thermography. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00466
International Maritime Organization (IMO) (2002) Guidelines for the Onboard Operational Use of Shipborne Automatic Identification Sysems (AIS). Available at: http://www.navcen.uscg.gov/pdf/AIS/IMO_A_917(22)_AIS_OPS_Guidelines.pdf.
Irvine L. M., Palacios D. M., Lagerquist B. A., Mate B. R. (2019). Scales of Blue and Fin Whale Feeding Behavior Off California, USA, With Implications for Prey Patchiness. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00338
IUCN (2017). North West Mediterranean Sea, Slope and Canyon System. Important Marine Mammal Area Factsheet (IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force).
Kaartvedt S. (2010). Diel Vertical Migration Behaviour of the Northern Krill (Meganyctiphanes Norvegica Sars). Adv Mar Biol. 57, 255–275. doi: 10.1016/S0065-2881(10)57009-3
Keen E. M., Scales K. L., Rone B. K., Hazen E. L., Falcone E. A., Schorr G. S. (2019). Night and Day: Diel Differences in Ship Strike Risk for Fin Whales (Balaenoptera Physalus) in the California Current System. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00730
Kelley D. E., Vlasic J. P., Brillant S. W. (2021). Assessing the Lethality of Ship Strikes on Whales Using Simple Biophysical Models. Marine Mammal Sci. 37, 251–267. doi: 10.1111/mms.12745
Kopelman A. H., Sadove S. S. (1995). Ventilatory Rate Differences Between Surface-Feeding and Non-Surface Feeding Fin Whales (Valaenoptera Physalus) in the Waters Off Eastern Long Island, New York, U.s.a-1987. Marine Mammal Sci. 11, 200–208. doi: 10.1111/j.1748-7692.1995.tb00518.x
Laist D. W., Knowlton A. M. R., Mead J. G., Collet A. S., Podesta M. (2001). Collisions Between Ships and Whales. Marine Mammal Sci. 17, 35–75. doi: 10.1111/j.1748-7692.2001.tb00980.x
Lavanguardia (2002) Una Ballena De 12 Metros Aparece Muerta En La Bahía Del Port De La Selva. Available at: https://www.lavanguardia.com/vida/20020619/51262756893/una-ballena-de-12-metros-aparece-muerta-en-la-bahia-del-port-de-la-selva.html.
Leaper R. (2019). The Role of Slower Vessel Speeds in Reducing Greenhouse Gas Emissions, Underwater Noise and Collision Risk to Whales. Front. Marine Sci. 6. doi: 10.3389/fmars.2019.00505
Lydersen C., Vacquié-Garcia J., Heide-Jørgensen M. P., Øien N., Guinet C., Kovacs K. M. (2020). Autumn Movements of Fin Whales (Balaenoptera Physalus) From Svalbard, Norway, Revealed by Satellite Tracking. Sci. Rep. 10. doi: 10.1038/s41598-020-73996-z
Martins M. C. I., Miller C., Hamilton P., Robbins J., Zitterbart D. P., Moore M. (2020). Respiration Cycle Duration and Seawater Flux Through Open Blowholes of Humpback (Megaptera Novaeangliae) and North Atlantic Right (Eubalaena Glacialis) Whales. Marine Mammal Sci. 36, 1160–1179. doi: 10.1111/mms.12703
Mazzariol S., Centelleghe C., Beffagna G., Povinelli M., Terracciano G., Cocumelli C., et al. (2016). Mediterranean Fin Whales (Balaenoptera Physalus) Threatened by Dolphin Morbillivirus. Emerging Infect. Dis. 22, 302–305. doi: 10.3201/eid2202.150882
Micheli F., Halpern B. S., Walbridge S., Ciriaco S., Ferretti F., Fraschetti S., et al. (2013). Cumulative Human Impacts on Mediterranean and Black Sea Marine Ecosystems: Assessing Current Pressures and Opportunities. PLOS ONE 8(12): e79889. doi: 10.1371/journal.pone.0079889
Moore M. J., Mitchell G. H., Rowles T. K., Early G. (2020). Dead Cetacean? Beach, Bloat, Float, Sink. Front. Marine Sci. 7. doi: 10.3389/fmars.2020.00333
Nieukirk S. L., Mellinger D. K., Moore S. E., Klinck K., Dziak R. P., Goslin J. (2011). Sounds From Airguns and Fin Whales Recorded in the Mid-Atlantic Ocea–2009. J. Acoustical Soc. America 131, 1102–1112. doi: 10.1121/1.3672648
Notarbartolo di Sciara G., Castellote M., Druon J. N., Panigada S. (2016). Fin Whales, Balaenoptera Physalus: At Home in a Changing Mediterranean Sea? Adv. Marine Biol. 75, 75–101. doi: 10.1016/bs.amb.2016.08.002
OceanCare (2021). Quiet Waters for Whales and Dolphins: The One-Time Opportunity to Avoid, Reduce and Mitigate Noise-Generating Activities in the Mediterranean Cetacean Migration Corridor. (Switzerland: Ocean Care).
Panigada S., Donovan G. P., Druon J. N., Lauriano G., Pierantonio N., Pirotta E., et al. (2017). Satellite Tagging of Mediterranean Fin Whales: Working Towards the Identification of Critical Habitats and the Focussing of Mitigation Measures. Sci. Rep. 7, 3365 . doi: 10.1038/s41598-017-03560-9
Panigada S., Gauffier P., Notarbartolo di Sciara G. (2021). Fin Whale the IUCN Red List of Threatened Species: Balaenoptera Physalus Mediterranean Subpopulation. e.T16208224A50387979. doi: 10.2305/IUCN.UK.2021-3.RLTS.T16208224A50387979.en.
Panigada S., Leaper R., Tejedor Arceredillo A. (2010). Ship Strikes in the Mediterranean Sea: Assessment and Identification of Conservation and Mitigation Measures. J. Cetacean Res., 1–5. MANAGE.
Panigada S., Notarbartolo di Sciara G., Zanardelli Panigada M., Airoldi S., Borsani J. F., Jahoda M. (2005). Fin Whales (Balaenoptera Physalus) Summering in the Ligurian Sea: Distribution, Encounter Rate, Mean Group Size and Relation to Physiographic Variables. J. Cetacean Res. Manage. 7, 137–145.
Panigada S., Pavan G., Borg J. A., Galil B. S., Vallini C. (2008). “Biodiversity Impacts of Ship Movement, Noise, Grounding and Anchoring,” in Maritime Traffic Effects on Biodiversity in the Mediterranean Sea. Chapter 1. (Malaga, Spain: IUCN Centre for Mediterranean Cooperation).
Panigada S., Pesante G., Zanardelli M., Capoulade F., Gannier A., Weinrich M. T. (2006). Mediterranean Fin Whales at Risk From Fatal Ship Strikes. Marine Pollut. Bull. 52, 1287–1298. doi: 10.1016/j.marpolbul.2006.03.014
Panigada S., Zanardelli M., Canese S., Jahoda M. (1999). How Deep Can Baleen Whales Dive? Mar Ecol. Prog. Ser. 187, 309–311. doi: 10.3354/meps187309
Pintore L., Sciacca V., Viola S., Giacoma C., Papale E., Giorli G. (2021). Fin Whale (Balaenoptera Physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn. J. Marine Sci. Eng. 9, 966. doi: 10.3390/jmse9090966
Puig P., Palanques A., Guillén J., García-Ladona E. (2000). Deep Slope Currents and Suspended Particle Fluxes in and Around the Foix Submarine Canyon (NW Mediterranean). Deep-Sea Res. Part I 47, 343–366. doi: 10.1016/S0967-0637(99)00062-X
QGIS Development Team (2018). Qgis Geographic Information System (Open Source Geospatial Foundation Project). Available at: http://qgis.osgeo.org.
Quadern el Pais (2016) Un Pescador Troba Una Balena De 12 Metres Al Delta De l’ebre. Available at: https://cat.elpais.com/cat/2016/01/28/catalunya/1453983052_549792.html.
R Core Team (2020). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Ritter F. (2012). Collisions of Sailing Vessels With Cetaceans Worldwide: First Insights Into a Seemingly Growing Problem. J. Cetacean Res. Manage. 12 (2).
Raga J. A., Raduán A, Balbuena J. A., Aguilar A, Grau E, Borell A. (1991). Varamientos De Cetáceos En Las Costas Españolas Del Mediterráneo Durante El Período 1982–1988. Miscellania Zoologica 15, 215–226.
Sabatés A., Gili J. M., Pagés E. (1989). Relationship Between Zooplankton Distribution, Geographic Characteristics and Hydrographic Patterns Off the Catalan Coast (Western Mediterranean). Marine Biol. 103, 153–159. doi: 10.1007/BF00543342
Schoeman R. P., Patterson-Abrolat C., Plön S. (2020). A Global Review of Vessel Collisions With Marine Animals. Front. Marine Sci. 7. doi: 10.3389/fmars.2020.00292
Sèbe M., Kontovas C. A., Pendleton L. (2019). A Decision-Making Framework to Reduce the Risk of Collisions Between Ships and Whales. Marine Policy 109. doi: 10.1016/j.marpol.2019.103697
Sèbe M., Kontovas C. A., Pendleton L. (2020). Reducing Whale-Ship Collisions by Better Estimating Damages to Ships. Sci. Total Environ. 713, 136643. doi: 10.1016/j.scitotenv.2020.136643
Smultea M., Fertl D., Bacon C., Moore M., James V., Wursig B. (2017). Cetacean Mother-Calf Behavior Observed From a Small Aircraft Off Southern California. Anim. Behav. Cogn. 4, 1–23. doi: 10.12966/abc.01.02.2017
Stepanuk J. E., Heywood E. I., Lopez J. F., DiGiovanni R. A. Jr., Thorne L. H. (2021). Age-Specific Behavior and Habitat Use in Humpback Whales: Implications for Vessel Strike. Marine Ecol. Prog. Ser. 663, 209–222. doi: 10.3354/meps13638
Stone G. S., Katona S. K., Mainwaring A., Allen J. M., Corbett H. D. (1992). Respiration and Surfacing Rates of Fin Whales (Balaenoptera Physalus) Observed From a Lighthouse Tower. Report of the International Whaling Commission. 42, 739–745.
Szegedi A., Tort B., Amigó N., Degollada E. (2019). “Drone Videos: A New Method for Photogrammetry of Fin Whales (Balaenoptera Physalus)?,” in World Marine Mammal Conference (Barcelona: WMMC).
Torreblanca E., Caminãs J. A., MacIás D., Garciá-Barcelona S., Real R., Baéz J. C. (2019). Using Opportunistic Sightings to Infer Differential Spatio-Temporal Use of Western Mediterranean Waters by the Fin Whale. PeerJ. 7, e6673. doi: 10.7717/peerj.6673
Tort B., Serrallonga S., Amigó N., Degollada E. (2017). Comportamiento Del Rorcual Común (Balaenoptera Physalus) En El Mar Balear Como Zona De Alimentación (Valencia: SEC).
Tregenza N., Aguilar N., Carrillo M., Delgado I., Díaz F., Brito A., et al. (2000). Potential Impact of Fast Ferries on Whale Populations a Simple Model With Examples From the Canary Islands. Eur. Res. Cetaceans 14, 195–197.
Vaes T., Druon J.-N. (2013). Mapping of Potential Risk of Ship Strike With Fin Whales in the Western Mediterranean Sea. A Scientific and Technical Review Using the Potential Habitat of Fin Whales and the Effective Vessel Density. JRC Sci. Policy Rep. 28 pp. doi: 10.2788/8520. European Commission.
Vanderlaan A. S. M., Taggart C. T. (2007). Vessel Collisions With Whales: The Probability of Lethal Injury Based on Vessel Speed. Marine Mammal Sci. 23, 144–156. doi: 10.1111/j.1748-7692.2006.00098.x
Winkler C., Panigada S., Murphy S., Ritter F. (2020). Global Numbers of Ship Strikes: An Assessment of Collisions Between Vessels and Cetaceans Using Available Data in the IWC Ship Strike Database. Report to the International Whaling Commission, IWC/68B/SC HIM09. 33pp.
Wood S. N. (2011). Fast Stable Restricted Maximum Likelihood and Marginal Likelihood Estimation of Semiparametric Generalized Linear Models. R. Stat. Soc. (B) 73, 3–36. doi: 10.1111/j.1467-9868.2010.00749.x
Zerbini A. N., Mendez M., Rosenbaum H. C., Sucunza F., Andriolo A., Harris G., et al. (2015). Tracking Southern Right Whales Through the Southwest Atlantic: New Insights Into Migratory Routes and Feeding Grounds (San Diego, USA: IWC Scientific Committee).
Keywords: fin whale, ship strike, collision, marine traffic, injury, feeding, Catalonia, Mediterranean Sea8
Citation: Tort Castro B, Prieto González R, O’Callaghan SA, Dominguez Rein-Loring P and Degollada Bastos E (2022) Ship Strike Risk for Fin Whales (Balaenoptera physalus) Off the Garraf coast, Northwest Mediterranean Sea. Front. Mar. Sci. 9:867287. doi: 10.3389/fmars.2022.867287
Received: 31 January 2022; Accepted: 18 March 2022;
Published: 27 April 2022.
Edited by:
Antonella Arcangeli, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA), ItalyReviewed by:
Paola Tepsich, CIMA Research Foundation, ItalyRoberto Carlucci, University of Bari Aldo Moro, Italy
Copyright © 2022 Tort Castro, Prieto González, O’Callaghan, Dominguez Rein-Loring and Degollada Bastos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beatriu Tort Castro, YmVhdHJpdXRvcnRAZWRtYWt0dWIub3Jn