
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 10 June 2022
Sec. Marine Conservation and Sustainability
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.867068
This article is part of the Research Topic Worldwide Survey on the Trade of Aquatic Wildmeat View all 6 articles
 Greicy F. Ruenes1,2,3*
Greicy F. Ruenes1,2,3* Larissa R. de Oliveira2,4
Larissa R. de Oliveira2,4 Renata Emin-Lima5,6
Renata Emin-Lima5,6 Alexandra Fernandes Costa5,6
Alexandra Fernandes Costa5,6 Fernanda Löffler Niemeyer Attademo7
Fernanda Löffler Niemeyer Attademo7 Fábia de Oliveira Luna7
Fábia de Oliveira Luna7 Salvatore Siciliano3,5,8
Salvatore Siciliano3,5,8In Brazil, dolphins’ body parts are commonly used as traditional amulets, named as love-charms, while dolphins’ teeth are commercialized in handcrafted necklaces and bracelets. Recent studies on forensic genetics confirmed the Guiana dolphin, Sotalia guianensis, as the principal target of the love-charms trading, with specimens incidentally captured in gillnets suggested as the primary source. As additional support for these investigations, we characterized tooth morphotypes and ages of dolphins used in the construction of traditional crafts, with the main objective of detecting the dolphin species and population groups most affected by this trade. Teeth collected from necklaces, bracelets, and earrings, sold in public markets from three major cities of Northern and Northeastern Brazil (Belém = 99, São Luís = 4, and Fortaleza =15), were analyzed using four morphological measurements: tooth total length (TL), root diameter (RD), crown length (CL), and cingulum diameter (CID). An unweighted pair group method using arithmetic means (UPGMA), with Euclidean distances as a measure of dissimilarity, multivariate analysis of variance (MANOVA), and principal component analysis (PCA) were used to detect tooth morphotypes in the samples. Ages were obtained by counting the growth layer groups (GLGs) deposited in dentine. Two different groups of teeth were detected presumably belonging to the species Sotalia guianensis and S. fluviatilis. Handicrafts showed a diverse age composition, ranging from 2 to 30 years, with a multimodal distribution (Mean =11 ± 7 SD, Median = 10, Mode = 4, 8, 19), similar to dolphin populations affected by fishery interaction. Results suggested that juveniles individuals and adults younger than 20 years, from both Sotalia species, probably incidentally captured, are the principal source of handicrafts, and raise concern about the greater vulnerability of young age categories presented in this trade.
The regular trade of cetacean products is long-standing in Brazil, still today. Folkloric beliefs from the Amazon region have the riverine dolphins (Sotalia spp. and Inia sp.) as central characters, fomenting a ritualistic usage of dolphin body parts with believed magical powers, renowned as “love charms” (Siciliano et al., 2018). These charms are commonly used and sold as traditional amulets, principally in Amazonian city markets, but have also been reported in coastal regions of the North, Northeast, and Southeast of the country (Siciliano, 1994; Sholl et al., 2008; Siciliano et al., 2018).
In Northern Brazil, the demand for boto (Inia sp.) derived amulets is high (Smith, 1996) and includes genital organs, eyes, skin, and teeth as principal products (Gravena et al., 2008; Siciliano et al., 2008). Since local traditional legends talk about boto dolphins changing to human form and seducing young women, it is believed that possessing one of these body parts brings luck in love and good sexual performance to the carrier (Cravalho, 1999; Vidal et al., 2019). However, recent research using genetic material from love charms collected in traditional markets of eight main cities from Northern to Southeastern Brazil indicated the estuarine dolphin Sotalia guianensis as the only cetacean species supporting this dolphin-derived charms trade (Siciliano et al., 2018).
Dolphins of the genus Sotalia are small-size odontocetes with geographical distribution restricted to Central and South America. The genus includes two species, Sotalia guianensis, a marine species limited to shallow coastal and estuarine habitats from Nicaragua to Southern Brazil, and Sotalia fluviatilis, a relatively smaller riverine species, exclusively found in the Amazon basin, including southern Peru, eastern Ecuador, southeastern Colombia, and northern Brazil (Flores et al., 2018) (Figure 1A). Both species are widely affected by incidental catch caused by the interaction with the intense fishing activity existing throughout their geographic range (Siciliano, 1994; Zappes et al., 2009; Meirelles et al., 2010; Iriarte and Marmontel, 2013a). Despite being fully protected by federal laws, intentional killing has been also documented for some regions of Brazil, related to their use as bait, in traditional medicine, and for human consumption (Alves and Rosa, 2008; Siciliano et al., 2008; Iriarte and Marmontel, 2013b; Barbosa-Filho et al., 2018, Dos Santos-Filgueira et al., 2021). In addition, S. guianensis and S. fluviatilis are intensely threatened by habitat degradation and pollution related to anthropic action, such as dam constructions and mining in the Amazon, besides city growth, boat traffic, and the destruction of mangroves and salt marshes in coastal areas (Flores et al., 2018; Secchi et al., 2018).
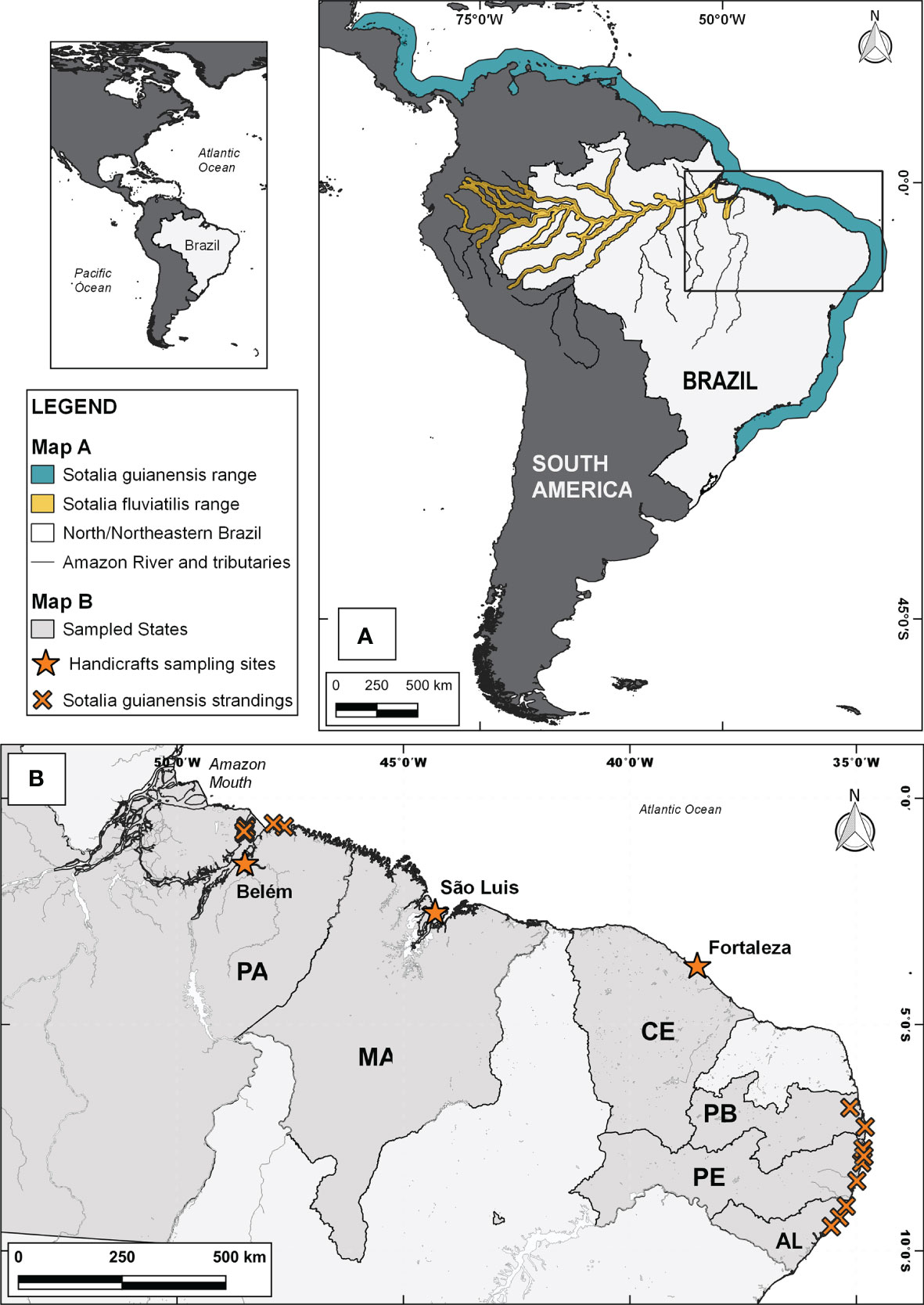
Figure 1 Distribution range of the species from the genus Sotalia in South America (Map A) and sampling points in the Northern (Map B: PA - Pará State), and Northeastern (Map B: MA - Maranhão State, and CE - Ceará State, PB - Paraíba State, PE - Pernambuco State and AL - Alagoas State) regions of Brazil were handicrafts and stranded animals were collected. Distribution range data were extracted from NatureServe and IUCN (2021).
Genetic forensic researches, such as Siciliano et al. (2018), are significantly important to assess the species more vulnerable or affected by illegal human trade, mainly when this trading uses body parts with no diagnostic characteristics or when the commercialized species are more difficult to identify. This achieves greater importance for the conservation effort of species classified as endangered, which is the case of Sotalia genus in Brazil (Flores et al., 2018; Secchi et al., 2018; Da Silva et al., 2020). As additional support to these genetic investigations, morphological characterization of specimens, as well as age determination could be valuable tools, making it possible to obtain specific information about demographic groups and populations of species that have been more exposed to this human activity. As part of the market of dolphin-derived charms, dolphin teeth are another widely traded item in the Northern Brazil region, mainly as ornaments for handmade necklaces, earrings, and bracelets, and often sold in streets and traditional markets. The morphology of teeth from aquatic mammals has been used before in the identification of populations stock (Stenella longirostris - Akin, 1988), for identification of species (Brunner, 2004), and sexual morphometric differentiation in pinnipeds (Otaria byronia - Tarnawski et al., 2014), and for ecotype characterizations based in dental alterations related to use, as in the case of Orcinus orca (Ford et al., 2011). Furthermore, the dentine and cement from aquatic mammals’ teeth are classified as recording structures, being shaped by growth layers with continuous deposition which allows accessing the exact age of individuals (Scheffer and Myrick, 1980; Hohn et al., 1989; Klevezal, 1996). These layers, named growth layer groups (GLGs), are cyclically deposited in parallel to the tissue formation’s surface, creating a pattern that can be counted. The validation of 1 year – 1 GLG is needed in order to confirm this chronological correspondence, and has been widely demonstrated for several species (Hohn et al., 1989; Da Silva, 1994; Ramos et al., 2000; Read et al., 2018). This research aimed to identify teeth from Sotalia spp. found in handicrafts sold at traditional markets of Northern and Northeastern Brazil, and assess the more frequent age classes used in their production, and therefore, affected by the trading of dolphin’s body parts.
Handicrafts, purchased by authors REL, AFC, and SS for this study, at five traditional or public markets from major cities of Northern and Northeastern Brazil were analyzed in addition to teeth obtained from stranded dolphins, used for comparison purposes only (Figure 1B). From these handicrafts, 118 teeth, being 99 acquired in Belém (Pará State), 4 in São Luís (Maranhão State), and 15 in Fortaleza (Ceará State), were used to assess the Sotalia spp. dolphins and the frequent ages commercialized in the trade of dolphin-derived artisanal objects (Table 1).
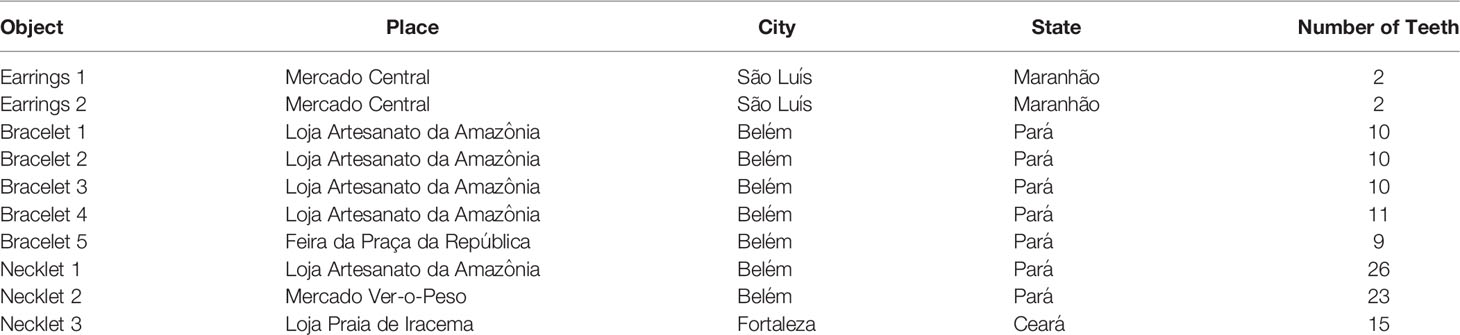
Table 1 Handicrafts collected in three major cities from the Brazil Northern and Northeastern regions.
External characteristics of the teeth adorning necklaces, bracelets, and earrings (Figures 2A–C), were used to confirm the genus/species to which they belonged. Each tooth was visually compared with teeth from small cetaceans preserved in the mammal collection at the Museu Paraense Emílio Goeldi, in Pará, Northern Brazil. This collection includes ~600 specimens of Delphinidae recovered in Pará, same geographical region where teeth are commercialized. Based on visual inspection all teeth used in this study were similar to Sotalia sp. specimens from the referred collection. Furthermore, since two different sizes of teeth from adult specimens were detected in the sample (Figures 2D, E), four external measurements were used to assess possible differences in teeth morphology that indicated different species of Sotalia genus being used in the handicrafts (Ramos et al., 2000): tooth total length (TL), measured from the apical extremity of the crow to the end of the root, root diameter (RD) measured as the maximum width of the root, crown length (CL), measured from the apical extremity of the crown to the root cingulum, and cingulum diameter (CID), measured as a diameter of the tooth at the intersection area of the crown with the root. In addition, teeth from 10 S. guianensis specimens stranded at the coast of Pará State (PA) and 10 S. guianensis specimens stranded in the Northeastern region of Brazil (NE) (Figure 1B) were measured and used for further comparisons as two different control groups (see statistical analyses section). Measurements were taken with a caliper to the nearest 0.05 mm and only teeth relatively straight and with no notable wear of the crow were analyzed.
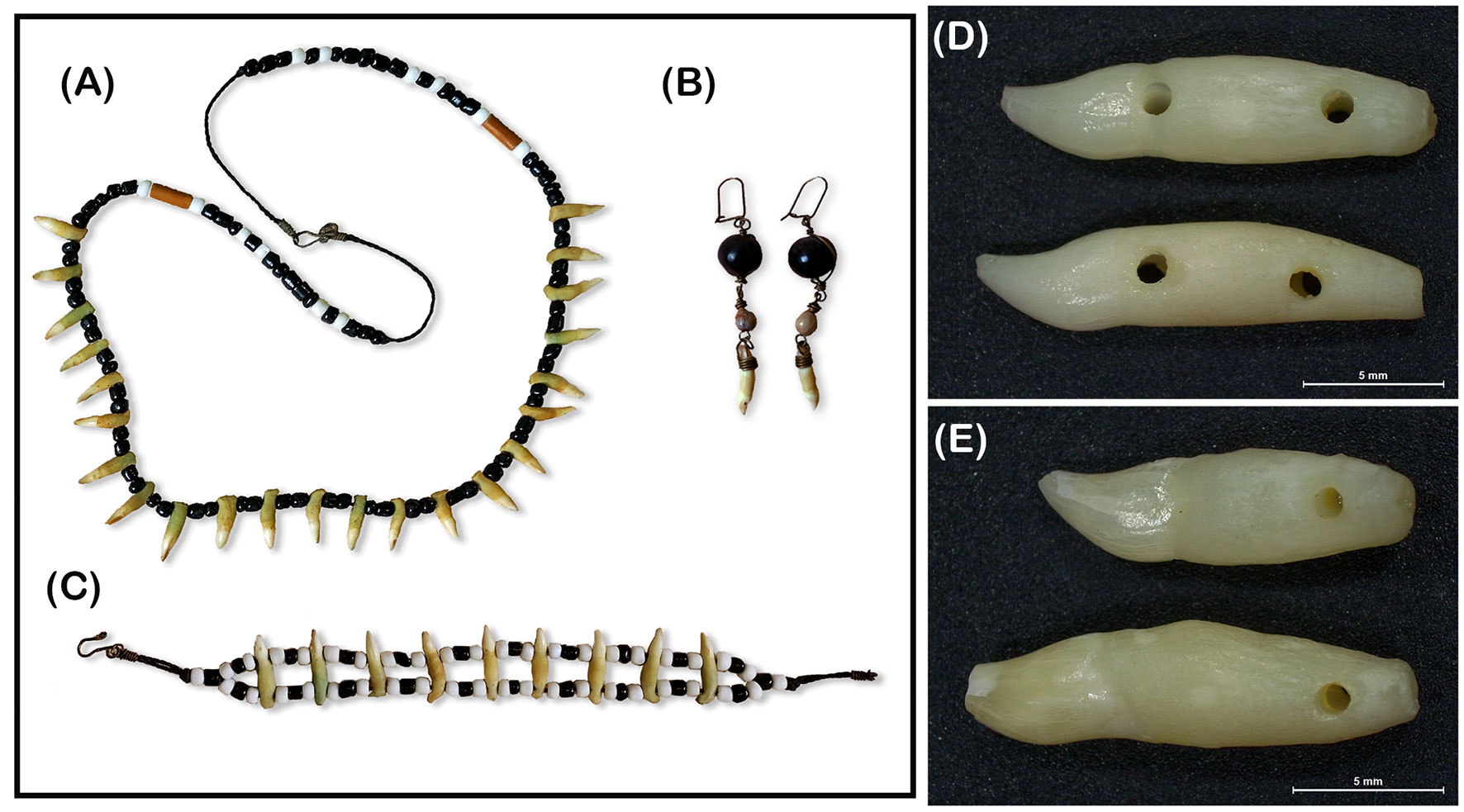
Figure 2 Sample of handicrafts manufactured with teeth of Sotalia dolphins and commercialized in the trade of artisanal objects in the North-Northeastern of Brazil: (A) necklace, (B) earrings, (C) bracelet, (D) Sotalia guianensis teeth from a bracelet, (E) Two different types of Sotalia sp. teeth detected in the necklaces.
The age frequency of the handicrafts was assessed by counting the growth layer groups (GLGs) deposited in the teeth’ dentine (Figure 3), and considering the described by Rosas et al. (2003). Each tooth was subsequently wearing, decalcified with RDO®, and cut with a manual freezing microtome in thin sections of 20-30 µm. Posteriorly, the thin sections were stained with Mayer’s hematoxylin, blued in Ammonia 2%, and finally, mounted in glycerin 100% (Rosas et al., 2003; Conversani et al., 2020). After being mounted, thin sections were photographed using a Leica S9i Stereomicroscope with an attached digital camera, and the images were processed by Leica Application Software v4.12 (Leica Microsystems, 2018) with a maximum magnification of 55x at Mammal Ecology Laboratory, Universidade do Vale do Rio dos Sinos, Brazil. Readings were made in the images three different times by two researchers separately to determine the chronological age of each tooth. The GLGs were counted in the dentine, having in consideration a correspondence of 1 GLG and 1 year of life described previously for the genus Sotalia (Da Silva, 1994; Rosas et al., 2003; Di Beneditto and Ramos, 2004). In some cases, the final age was classified as minimum age because the perforations made for the craft construction coincided with the center of the pulp cavity (Figures 2 and 3), making it impossible to read the most recently deposited GLGs. The best age estimate was defined as the coincidental readings of at least two from the three times. In the case of a discrepancy ≤ 2 GLGs between readings, final age was considered as the mean of the three different ages. If a discrepancy between readers occurs, final ages were obtained by a consensus reading involving the two researchers. Readings with differences ≥ 3 GLGs were not considered in the analysis.
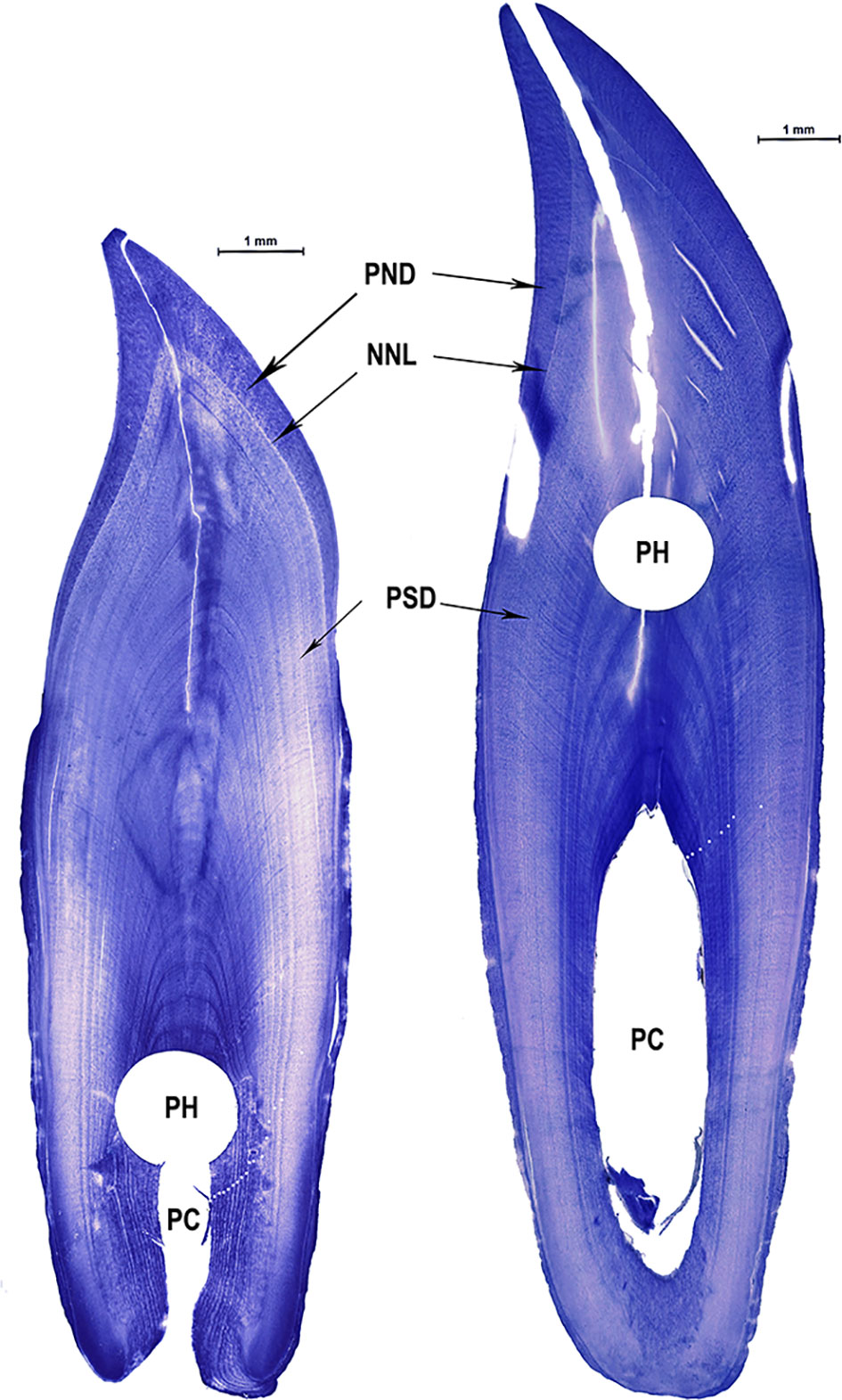
Figure 3 Thin sections from two Sotalia sp. teeth collected from handicrafts (left: tooth N1-10 with TL= 13.16 mm; right: tooth B2-5 with TL=17.4 mm). PND, Prenatal dentine, NNL, Neonatal line; PSD, Postnatal dentine; PC, Pulp cavity; PH, Holes made for craft construction. White dots indicate the GLGs counts in the dentine corresponding to the age.
Finally, based on the age readings, teeth were classified as belonging to three different age categories: juveniles (age ≤ 6), young adults (6 < age ≤ 12), and adults (13 ≥ ages), taking into account that a) the age of 7 years as the age of sexual maturity for Sotalia guianensis (Rosas and Monteiro-Filho, 2002; Rosas et al., 2003) and b) the ages between 7 and 12 years as the range when occurs the complete fusion of the sutures of the occipital complex, concurring with the age of sexual maturity in S. guianensis and S. fluviatilis (Fettuccia et al., 2009; Novais et al., 2020).
Since two species from the genus Sotalia are described in the literature (S. guianensis and S. fluviatilis), we intend to answer three questions through the statistical analysis. First, if two different groups of Sotalia sp. teeth were present in the collected handicrafts. Secondly, if some of these groups were more similar to teeth from S. guianensis specimens stranded in two different coastal localities of the sampling area (Figure 1B), and if these differences were influenced by age or if they were exclusively morphological differences. To this end, we included data from stranded specimens as two control groups (PA and NE), assuming that they adequately represented the morphological variation of the species S. guianensis, since individuals from populations with the smallest body size (PA region, Novais et al., 2020) and populations with the largest body size (NE region, Monteiro-Filho et al., 2002) described for the species were added. Finally, as a third question, we wanted to know what the most frequent ages in these products were for the detected groups.
Teeth from handicrafts were initially grouped through a cluster analysis using the four external measurements (TL, RD, RC, and CID) in order to find potential groups related to the tooth external characteristics of the two species of Sotalia. The dendrogram was constructed based on the unweighted pair group method using arithmetic means, UPGMA (Hale and Dougherty, 1988), and Euclidean distances (Podani, 2000) as a measure of dissimilarity. To evaluate the stability of the cluster, indicated by the representativeness of the structure of the data, a bootstrap resampling with 1000 iterations was performed and obtained the Jaccard coefficient of similarity between sets (Hennig, 2007). Cluster results were visualized including also the sampling localities to check if the geographical distribution of teeth was also related to the grouping.
Resulting groups from cluster analysis and teeth groups of stranded specimens (Tooth Type factor), the age category of teeth (Age class factor), and as dependent variables the four morphological measurements, were combined in a two-way multivariate analysis of variance (MANOVA). Also, the interaction between factors was evaluated in this analysis. Box-Cox transformation was used for two dependent variables (RC, and CID), to fulfill the MANOVA assumptions of homogeneity of the variance-covariance matrices and multivariate normality. Since multivariate significant differences were detected, we performed analyses of variance (ANOVAs) for each dependent variable. We used an a posteriori Tukey test to find out where the significant differences were in the ANOVA results. Likewise, to determine which morphological variables most influenced the composition of the teeth groups, a principal component analysis (PCA) was performed using also the four external measurements from teeth. Critical value for coefficients of correlation between variables and components, i.e. loading, was set to 0.5. In the graphical visualization of the PCA results, data were identified using the classification from the clusters, the two control groups, and the growth categories of teeth as a way to visualize the MANOVA results. Thus, to see if the distribution of the scores had some relation with the age or were only influenced by the morphological variables indicating similarities in teeth characteristics in the groups, and therefore, a high probability to belong to the same species.
We used p < 0.05 as the critical p-value of statistically significant. Descriptive statistics such as Range, Mean, Median, Standard deviation (SD), and 95% confidence interval (CI) were also acquired. Finally, the age distribution of the sample was obtained using the age estimates, and differences in the age distribution between age classes and groups of teeth from handicrafts were described.
All statistical analyses and graphics were conducted in the free software RStudio v.1.4.1717 (RStudio Team, 2020). The packages used were stat, from software R v.4.0.1 (R Core Team, 2020), car v.3.0-12 (Fox and Weisberg, 2019), ggdendro v. 0.1.22 (De Vries and Ripley, 2020), fpc v. 2.2-9 (Hennig, 2020), and factorextra v. 1.0.7 (Kassambara and Mundt, 2020). Graphics were created using the packages ggplot2 v.3.3.5 (Wickham, 2016) and gridExtra v.2.3 (Auguie, 2017).
Two clusters were generated by the analyses (Euclidean distance for separation = 3.843), indicating two different morphotypes of teeth from dolphins of the genus Sotalia in the sample (Figure 4). The Jaccard coefficients of the two clusters, 0.94 and 0.90, indicated good representability of the data structure by the dendrogram. The morphotype named as Type 1 included 67 teeth from handicrafts, while the morphotype named as Type 2 included 42 teeth. Related to geographical distribution, Type 1 morphotype was found in all sampled markets of Northern and Northeastern Brazil, while Type 2 was found only in two markets of Belém, the capital city of Pará State, placed at the southeastern Amazon Estuary (Figure 4).
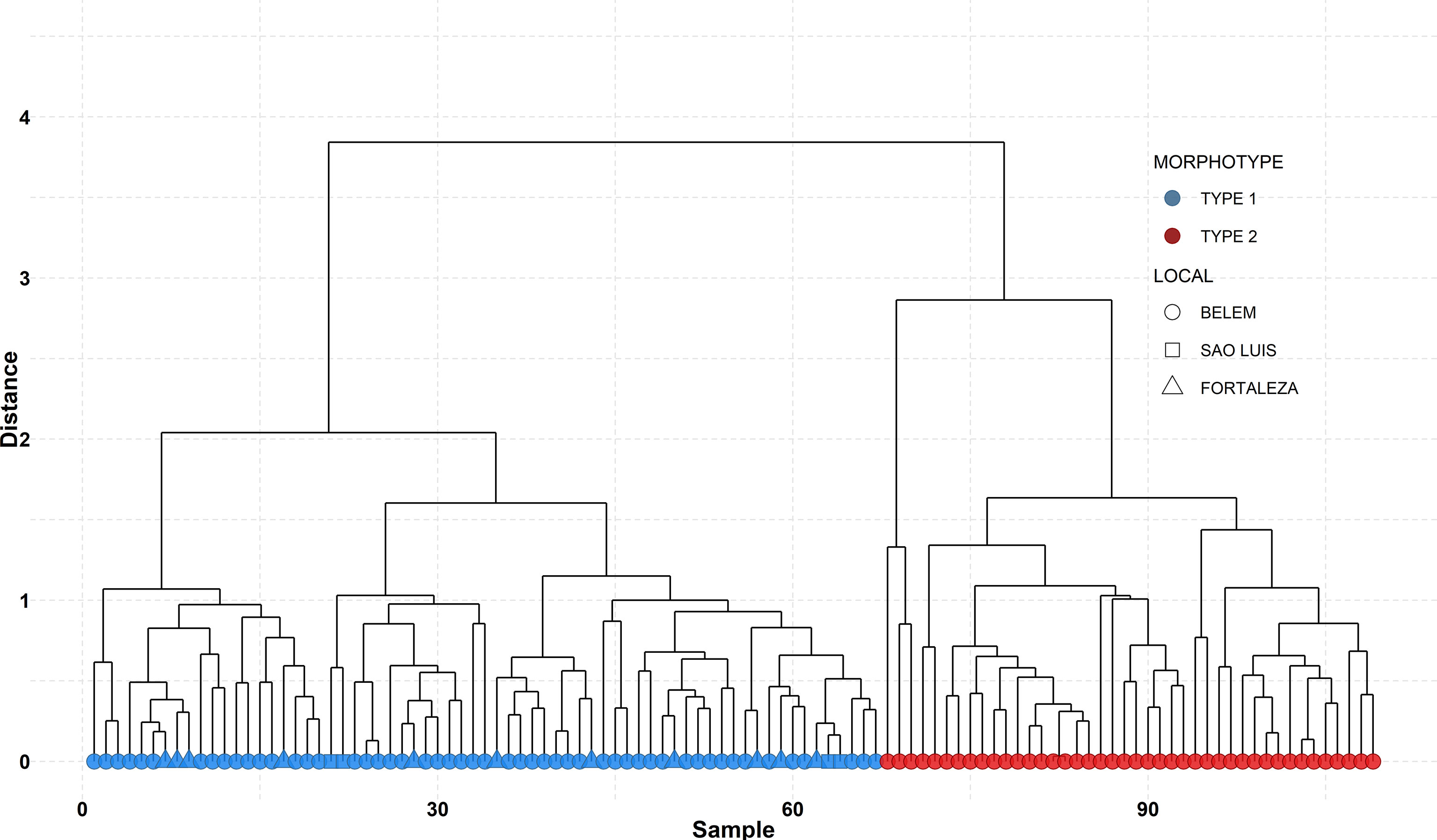
Figure 4 Dendrogram from the clustering analysis showing the two principal clusters separating tooth types. Colors indicate the teeth morphotype groups and symbols indicate the sample’s locality in the North and Northeast regions of Brazil.
Type 1 morphotype of teeth shows more similar dimensions to PA and NE control groups than Type 2 morphotype (Figure 5). The two-way MANOVA results indicated that measurements differed between the four groups of teeth (Type 1, Type 2, NE and PA) (df = 3, Pillai Trace = 1.080, approximate F = 15.900, p < 2.2e-16), age classes (juveniles, young adults, and adults) (df = 2, Pillai Trace = 0.283, approximate F = 4.600, p = 3.178e-05) and also between teeth morphotypes in relation to the ages classes (df = 6, Pillai Trace = 0.3698, approximate F = 1.900, p = 0.005). Individual ANOVAs detected statistical differences between groups of teeth for the measurements total length (TL), root diameter (RD), crown length (CL), and cingulum diameter (CID), while differences between the age classes were only related to root diameter (RD) (Supplementary Table S1). Type 1 morphotype were similar to PA control group in TL (Mean differences = -0.80, p = 0.152) and CID (Mean differences = 0.28, p = 0.971), and similar to NE control group in CID (Mean differences = 1.417, p = 0.109), and CL (Mean differences = -1.92, p = 0.052). Type 2 shows the smallest dimensions for the four measurements, being statistically different from the others groups of teeth in TL, CID and CL (Figure 5 and Supplementary Table S2). Type 2 morphotype were also different from Type 1 and NE control group in RD, but similar to PA teeth (Figure 5 and Supplementary Table S2). Related to the ages classes, juveniles (Mean differences = -0.35, p = 0.0001) and young adults (Mean differences = - 0.27, p = 0.0032) show smaller RD dimensions, being statistically different from adults (Supplementary Table S2).
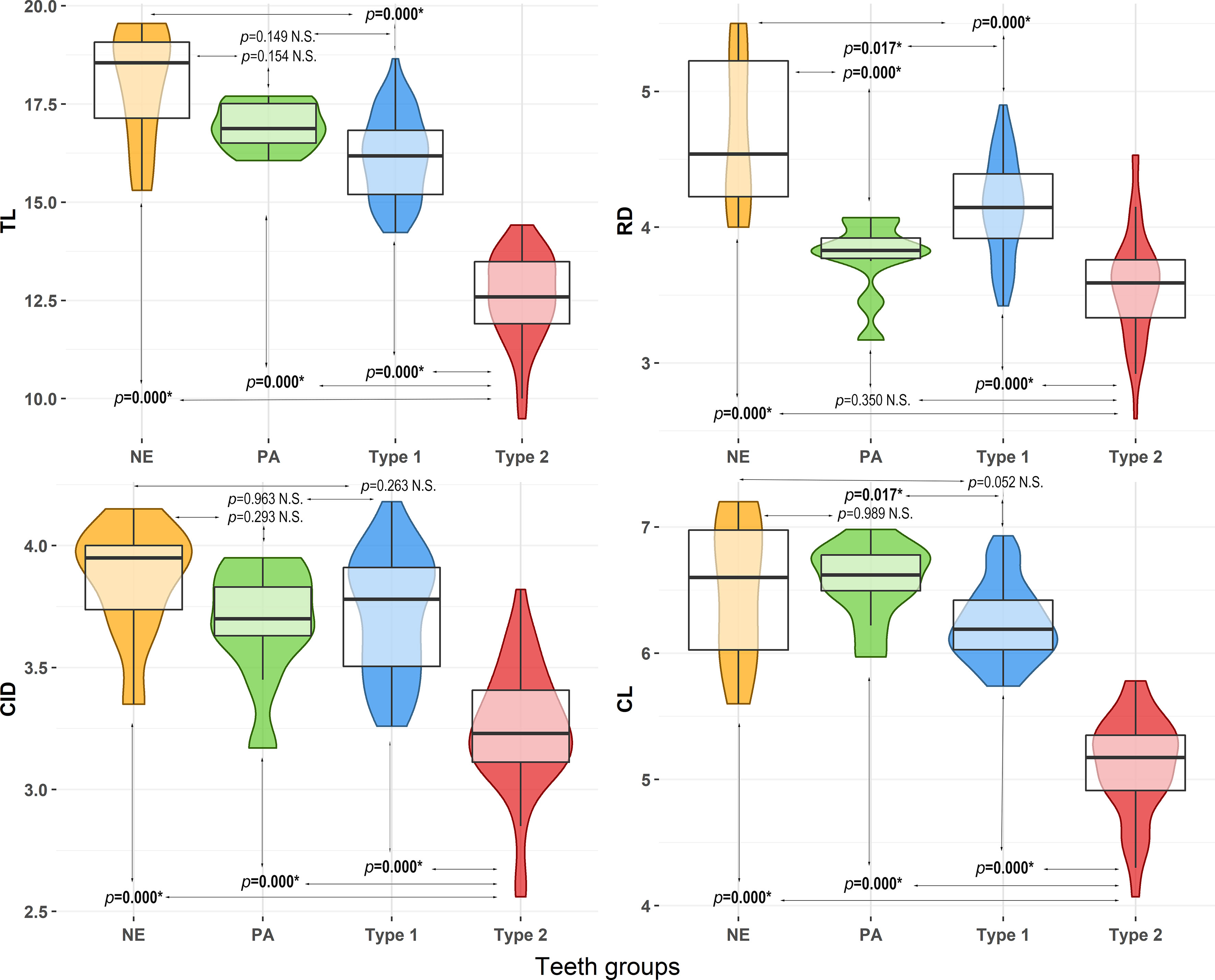
Figure 5 Violin data representation and pairwise comparison between groups of teeth for the measurements of total length (TL), root diameter (RD), cingulum diameter (CID), and crown length (CL). Type 1 and Type 2 comprised groups of teeth resulting from cluster analysis and the localities of Pará coast (PA) and Northeast Brazil (NE) comprised only stranded specimens of Sotalia guianensis used as control groups. Box represents values between the first and third quartile, the horizontal dark line represents the median, and vertical lines inside the violin shapes represent the lowers and upper values. Bold and asterisk in p-values indicate significant differences from pairwise comparisons for p<0.05; N.S., indicates non-significant differences.
Interaction between factors influenced the CID, with some age categories being different between the types of teeth, but always related with Type 2 morphotype having smaller dimensions (Supplementary Table S2). Thus, juveniles, young adults and adults teeth from Type 1 morphotype had, respectively, significant higher dimensions than juveniles (Mean differences = 2.14, p=0.00), young adults (Mean differences = 1.68, p=2.862E-03) and adults (Mean differences = 1.68, p=1.040E-05) from Type 2 morphotype. Juveniles teeth from Type 2 morphotype were also smaller than juveniles from PA (Mean differences = -2.42, p=8.267E-04) and NE (Mean differences = -2.46, p=1.210E-05) control groups, and smaller than adults from Type 1 morphotype (Mean differences =-2.19, p=0.00), PA (Mean differences= -1.90, p= 0.0162), and NE (Mean differences = -2.55, p=0.00) control groups. Young adult teeth from Type 2 were smaller than juveniles (Mean differences = -1.74, p= 0.0001) and adults (Mean differences = -1.81, p= 2.870E-05) from Type 1 morphotye, and juveniles (Mean differences = -2.13, p= 0.0038) and adults (Mean differences = -2.23, p= 1.660E-05) from NE control group. Finally, adults from Type 2 were smaller than juveniles (Mean differences = -1.61, p=5.600E-05) and young adults (Mean differences = -1.55, p=0.004) from Type 1 morphotype (Mean differences = -2.55, p=0.00), and juveniles (Mean differences = -2.02 p=0.008) from NE control group.
The PCA grouping shows a similar data structure to the clustering analysis with the first two dimensions explaining 87.1% of the data ordination (Figure 6; Supplementary Table S3). The first principal component (PC1) explained 72.9% ± 0.22 SD of the total variability. The measurement with higher loading for this PC was the TL (Loading = 0.6405), describing the size difference between the groups, followed by the CL but with a smaller correlation (Loading = 0.4818). The PC2 explained only 13.8% ± 0.09 SD of the variance, being more related to the DR (Loading = 0.8172), indicating an increase in diameter through the age categories (Figure 6) and supporting the differences between adults and the others ages classes founded in the ANOVA analysis.
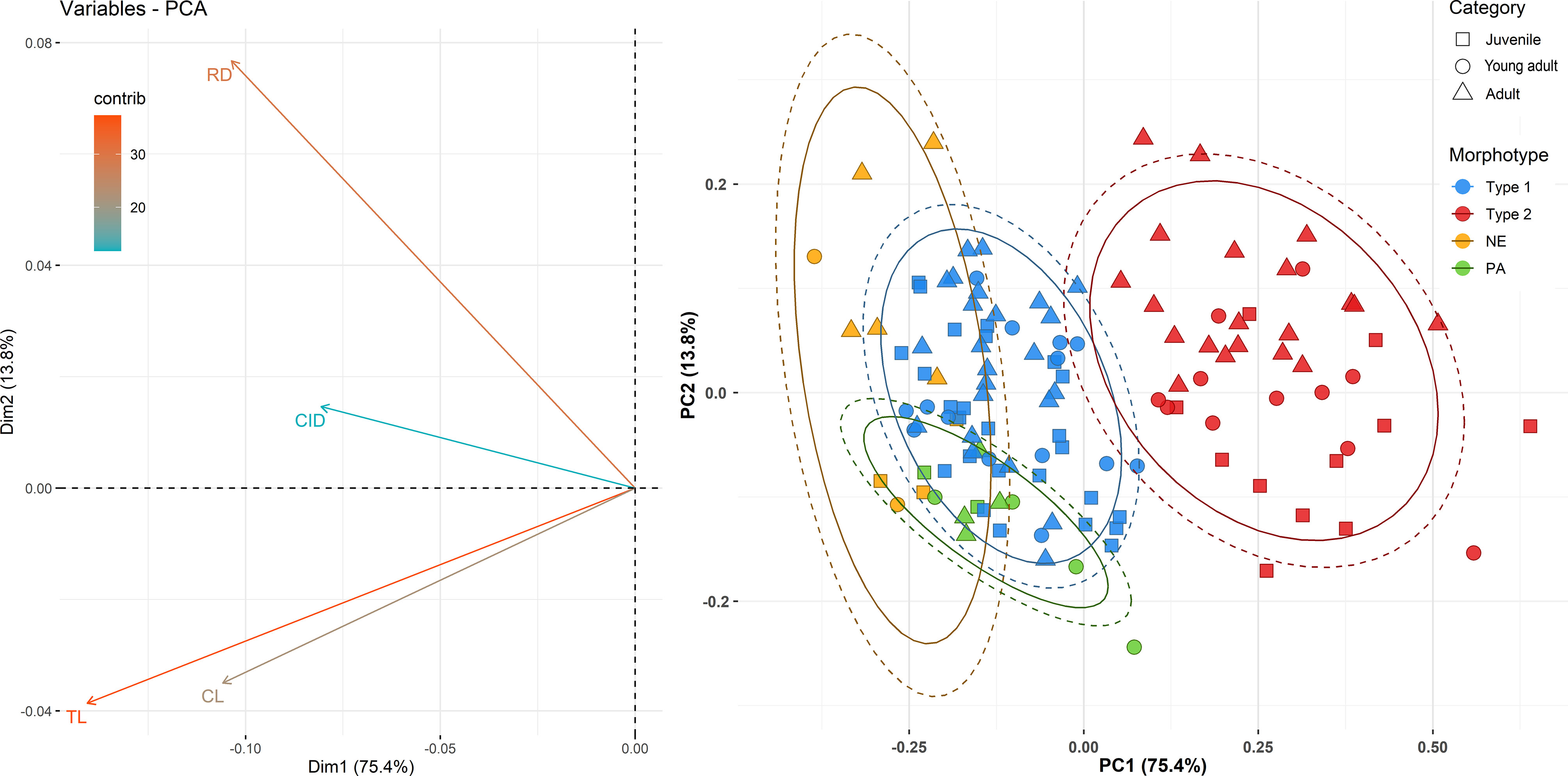
Figure 6 Principal component analysis for teeth measurements obtained from samples of Sotalia dolphins from North and Northeastern Brazil. Left: Contribution of the variables total length (TL), root diameter (RD), crown length (CL), and cingulum diameter (CID) to the principal component dimensions (PCs). Right: Scores distribution in the two dimensions of the PCs. The category indicates juveniles, young adults, and adults in the sample, and colors indicate the teeth morphotypes or species. The localities of Pará coast (PA) and Northeast Brazil (NE) included only stranded specimens of Sotalia guianensis.
Ages of 116 teeth adorning the handicrafts were possible to obtain, with 64 teeth bellowing to Type 1, 42 bellowing to Type 2, and 10 teeth with no classification. The total sample showed a diverse age composition, ranging between 2 and 30 years (Mean =11 ± 7 SD, Median = 10, Mode = 3, 4, 8, and 19 years). Some teeth from the ages 25, 28, and 29 years were aged as minimal age because of the loss of part of the pulp cavity in the construction of handicraft (see Figure 2), thus probably being older than was detected. Juveniles dolphins (ages ≤ 6) were 34% of the teeth sample, while young adults (6 < ages ≤ 12) corresponded for 27% and adults (ages ≥ 13) for 41%. Where the sample was split by teeth morphotype, Type 1 was composed of 40% juveniles, 22% by young adults, and 38% by adults. The type 2 sample was divided into 24% of juveniles, 25% of young adults, and 49% of adults. Age composition for the teeth classified as Type 1 ranged between 2 and 30 years (Mean =10 ± 7 SD), with the age class of 3 years being the most frequent in this group (17% of the sample), followed by the ages of 4 and 13 years which represented in both cases 11% of the sample. The ages of teeth classified as Type 2 ranged between 4 and 28 years (Mean=14 ± 6 SD), with the ages of 5 (12%) and 16 years (10%) being the most frequently observed. Adults, between the ages of 7 and 20 years, represent, respectively, 52% and 63% of Type 1 and Type 2 morphotypes. The distribution of ages for both morphotypes of teeth showed a multimodal frequency distribution (Figure 7; Supplementary Table S4).
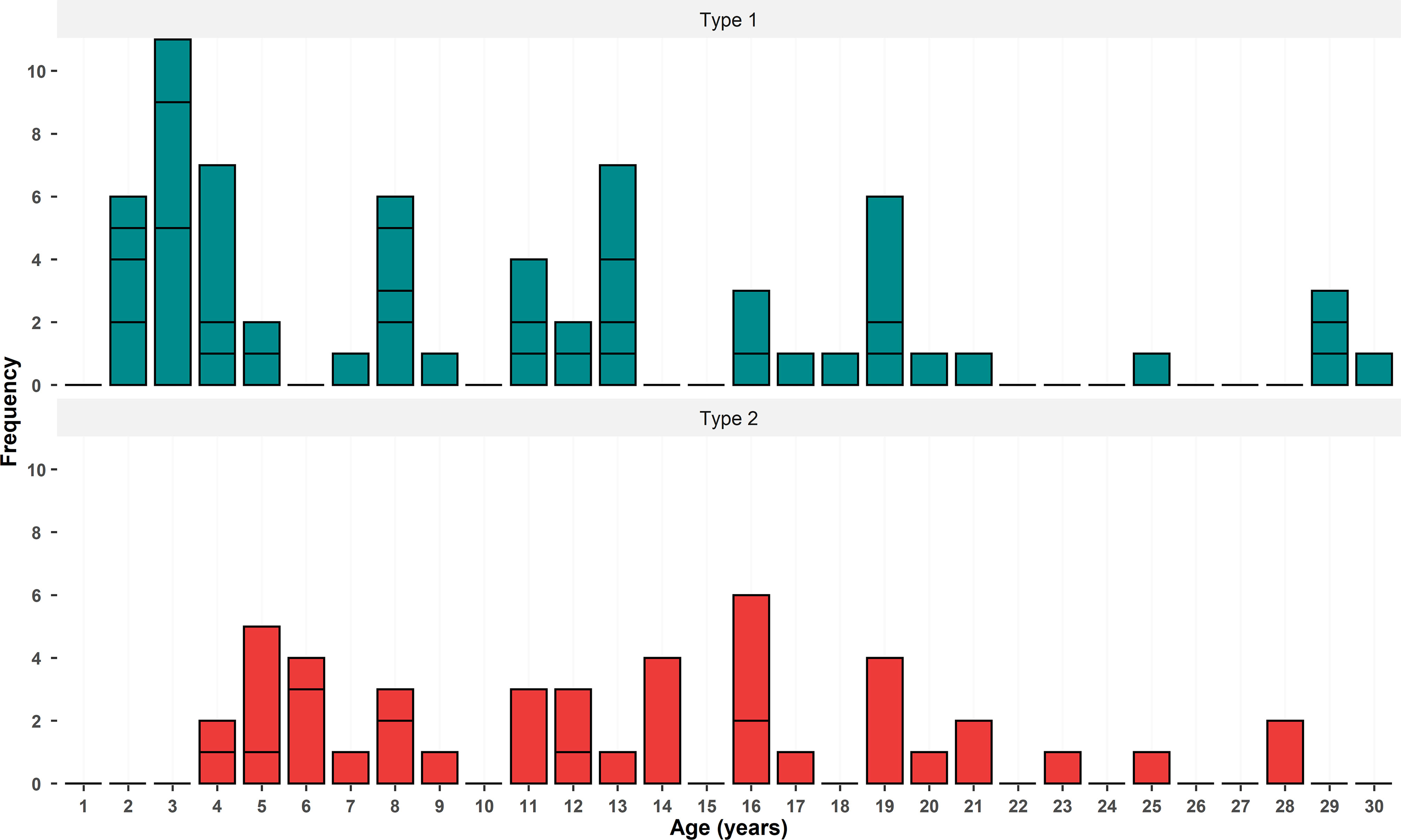
Figure 7 Age distributions for the two morphotypes of Sotalia spp. dolphin teeth detected in the trade of handcrafted objects in the North and Northeast regions of Brazil. Branches indicated the frequency of each age class in the handicraft objects.
Two morphotypes of teeth, presumably belonging to both species from the genus Sotalia, S. guianensis, and S. fluviatilis were found on handmade necklaces, bracelets, and earrings sold in public markets of three main cities of the North and Northeast coast of Brazil. Age distribution of teeth indicated that juveniles and young adult and adult dolphins, between the age of 2 and 8 years, and between the age of 11 and 20 years, are more used in these trades since were the most frequent ages observed in the different handicraft objects.
Some aspects of geographical variation in morphology of the genus Sotalia have been more frequently investigated, mainly related to general body size and osteology (Borobia, 1989; Monteiro-Filho et al., 2002; Fettuccia et al., 2009; Ramos et al., 2010; Fettuccia, 2010; Fettuccia et al., 2012; Arcoverde et al., 2014f). For these results, it is confirmed a variation in body length and cranial size for S. guianensis on the Brazilian coast, with the smallest individuals habiting the Amazon estuarine region, and a gradual increase in size for populations inhabiting coastal areas from more Northeastern, Southeastern and Southern regions (Fettuccia, 2010; Emin-Lima, 2012; Arcoverde et al., 2014). Additionally, S. fluviatilis, the riverine species of the genus, is described as significantly smaller in body dimensions compared to S. guianensis, with slower body development and with the preservation of some pedomorphic characters (Fettuccia et al., 2009; Fettuccia, 2010). In our study, it was possible to detect this geographic variation in size in the teeth collected from the handicrafts and stranded animals. Thus, teeth of stranded specimens from the Northeastern Brazilian region (NE), had the highest dimensions, while teeth from specimens stranded in the coastline of Pará state (PA), North region, were relatively smaller in size, but with no statistical difference between the two regions for the majority of measurements. Overlapping with these two groups, the teeth classified as Type 1 showed a similar interval of size, although being statistically different from those collected in Northeastern Brazil in total length and root diameter. On the other hand, the Type 2 morphotype, detected in two of the handicrafts, showed evident smaller dimensions being separated from the other three groups in the ordination analysis and with a statistically significant difference in almost all measurement comparisons. Furthermore, in the only variable where the interaction of age classes and tooth type was identified (CID), the results indicated that all age classes of the Type 2 morphotype were mostly smaller than the age classes of other tooth groups, and no differences were found within any group. Thus, suggesting that the differences found are related to the morphological characteristics of the groups and not to possible modifications throughout growth. In addition, the Type 2 morphotype was found only in the Southeastern Amazon Estuary region, while Type 1 was found in all North-Northeastern localities. Based on these results it is suggested that the Type 1 teeth morphotype belongs to the species S. guianensis, while the Type 2 morphotype of teeth belongs to the riverine species S. fluviatilis.
The use of body parts of Sotalia dolphins has been widely reported in South America. In Brazil and Venezuela, estuarine dolphins S. guianensis accidentally captured in gillnet are commonly used for human consumption and bait in longline-fishing or shark-related-fishing (Siciliano et al., 2008; Briceño et al., 2021). In addition, some practices related to Amazon traditional culture from northern Brazil, i.e., the use of dolphin parts as love’s charms (Siciliano et al., 2018), have been linked to the increased trade of genitals, eyes, dorsal fin, skin, and teeth of Amazon dolphin species in traditional markets (Siciliano, 1994; Da Silva and Best, 1996; Siciliano et al., 2008; Sholl et al., 2008). More recently, forensic genetics results have indicated that S. guianensis is the most used cetacean in the majority of dolphin’s love charms sold in traditional Brazilian markets, including the markets in the cities of Belém, São Luís and Fortaleza sampled in our research (Gravena et al., 2008; Sholl et al., 2008; Siciliano et al., 2018). Thus, our results are consistent with the geographical distribution of this trade in Northern and Northeastern Brazil and reveal specific segments of S. guianensis populations being more frequently used.
For the riverine species S. fluviatilis, its use as bait principally for the piracatinga (Calophysus macropterus) fishery, has been documented in the Amazonian regions of Peru and Brazil (Brum et al., 2015; Campbell et al., 2020), and medicinal and magical religious use has been also reported in Brazilian, Peruvian and Bolivian Amazon region (Alves et al., 2013). Based on the analysis of genetic markers on the tissue samples from love’s charms acquired in Brazilian traditional markets, Siciliano et al. (2018) did not detect the use of S. fluviatilis individuals in this kind of trade. However, handicraftsmen who were carrying a notable quantity of Sotalia teeth, interviewed near Belém, Pará, reported to the authors that they had acquired the teeth from several fishing communities in their trip through the Amazon. Therefore, it is possible the presence of S. fluviatilis teeth brought from riverine regions in handicrafts sold at public markets of Belém or another coastal region of the southeastern Amazon. Siciliano et al. (2018) also described those necklaces and bracelets made with Sotalia teeth were found in traditional open-air market squares from Belém and were the only charm found in street markets from Fortaleza, Ceará State. In this context, it could be possible that the trading of teeth and handicrafts made with them, would take a different path to these sites, more related to these travelers selling handicrafts at squares and street markets, and not with the traditional trading of other dolphin body parts. This fact could also explain the absence of S. fluviatilis in the love’s charm samples from the study conducted by Siciliano et al. (2018).
The age distribution obtained in our study indicated that teeth from juvenile individuals, between the ages of 2 and 6 years, and young adults and adults, between 8 and 19 years, are more frequently used in handicrafts. Both Sotalia species show this pattern of age distribution, being more evident in S. guianensis. Although some of the teeth may be from the same individuals, we believe that the repetition of ages on the different objects collected indicates a recurrence of animals of these age classes as a source of teeth for handicrafts. In addition, no teeth of specimens younger than two years old were found, maybe because crafts with smaller or finer teeth can be more difficult to manufacture and/or because teeth providing this trade could be collected from specimens accidentally caught in fishing gillnets, being newborns and yearlings probably discarded or less affected by this activity. This last hypothesis needs further confirmation. However, some inferences are possible to be made. In marine mammal populations under a stable age distribution, natural mortality rates are expected with the greatest frequency of yearlings (animals < 1year), followed by juveniles and adults with much less frequency (Reilly and Barlow, 1986; Barlow and Boveng, 1991; Mannocci et al., 2012). When mortality is affected by the action of bycatch (where animals are accidentally caught in gillnets during fisheries), the expected age distribution changes because not all ages are equally affected in the process (Moore and Read, 2008; Mannocci et al., 2012). Thus, an increase in juveniles’ and young adults’ mortalities relative to the other age classes is observed. This had been detected in different dolphin populations, such as striped dolphin (Stenella coeruleoalba) and common dolphin (Delphinus delphis) in the Northeast Atlantic Ocean (Mannocci et al., 2012; Brown et al., 2014), and common bottlenose dolphins (Tursiops truncatus) in North Carolina coast (Byrd and Hohn, 2017).
On the North Pará coast, S. guianensis is the most frequent marine mammal found stranded, as a result of its interactions with local fishing activity (Siciliano, 1994; Siciliano et al., 2008; Costa et al., 2017). The use of dolphin carcasses, stranded and/or incidentally captured in fisheries as a source of products found in regional markets of Northern Brazil has been previously suggested (Siciliano, 1994; Da Silva and Best, 1996; Sholl et al., 2008; Siciliano et al., 2018), as well as its use as bait or for human consumption (Siciliano, 1994; Tosi et al., 2009; Zappes et al., 2009; Dos Santos-Filgueira et al., 2021). Moreover, studies on S. guianensis bycatch for populations of Northeastern, Southeastern, and Southern Brazil indicated that juveniles and adult individuals, with body length superior to 160 cm (> 6 years old), have more vulnerability to by caught in fishing nets representing until 80% of the captures (Rosas et al., 2003; Di Beneditto and Ramos, 2004; De Moura et al., 2009; Meirelles et al., 2010; Lima et al., 2016). For S. fluviatilis, despite interactions with fishing activities are extremely difficult to record, punctual data of bycatch and intentional catches in Western Brazilian Amazon indicated a high incidence of juveniles (of ~138 cm of body length) and adults (reproductively matures animals) captured in tambaqui (Colossoma macropomum) and pirapitinga (Piaractus brachypomus) fishing gillnets (Da Silva and Best, 1996; Iriarte and Marmontel, 2013a). Furthermore, Iriarte and Marmontel (2013b), raised concern about the use of Amazonian River dolphin carcasses as a productive cost-effective means of obtaining bait for fishermen. The cost, viability, and effectiveness as drivers for the use of dolphins as bait by fishermen have been recently confirmed on the Peruvian coast, where similar human-related pressures increased the mortality of coastal small cetaceans (Campbell et al., 2020).
Whether the origin of our sample is from stranded or accidental capture, remains unknown. However, based on our results, and having into consideration the most frequent age categories observed in our sample, juveniles and adults younger than 20 years, it is possible to suggest that handicrafts commercialized as part of dolphin charm trade in the North and Northeast of Brazil and Sotalia dolphins’ bycatch seems to affect the same population segment, and they are probably related in this region. Public policies and conservation actions have been encouraged in Brazil to reduce dolphin bycatch. However, the real impact of this activity on populations of Sotalia spp. dolphins from Northern-Northeastern Brazil are still unknown. Ethnoecological studies with fishermen’s communities indicated a high incidence of this anthropic action in the region, with animals being mostly discarded and finally stranding at the coast (Siciliano, 1994; Brito, 2012; Dos Santos-Figueira et al., 2021). The overlapping in the occurrence of the dolphin population’s groups with preferential areas for gillnet placements has been suggested as one of the principal causes of the high incidence of bycatch (Martins et al., 2004; Beltrán-Pedreros and Petrere, 2010). In this scenario, the sale of body parts could be both, indicative of the high mortality of these species in the region, and a pressure that can influence this mortality, increasing the vulnerability of the local dolphin populations.
In summary, our study attempted to detect different cetaceans teeth used in the confection of handicrafts traded as artisanal dolphin-derived products in North and Northeastern Brazil using linear morphometry as a tool. We detected two different morphotypes of teeth from the genus Sotalia in the handicrafts, presumably belonging to the species S guianensis and S. fluviatilis. However further studies are needed to confirm this inference. In addition, we see the necessity of future researchers related to cetaceans’ tooth morphology, that confirm the use of teeth as an effective tool for distinguishing species where other data is unknown. We also attempted to access the segments within the Sotalia spp. populations more used in this trade by determining the age of the teeth. We found that juveniles and adults younger than 20 years are more frequently used in these handicrafts ornamented with teeth, coinciding with the age categories that have the highest incidence of cetaceans mortality related to bycatch in this region. Thus, the technic of age determination showed its applicability as a tool in gathering information about human actions, directly and indirectly, affecting specific groups of coastal populations of Sotalia dolphins in Northern and Northeastern Brazil. In addition, this result raises concerns about the increased vulnerability of the two species, but especially related to the threatened species S. fluviatilis, as scarce data have been reported on their life-history traits and only one age-related research was found available. Finally, we believe that this improved data could allow for more realistic management policies to help in the conservation of the two Sotalia species in the Amazon region, guiding enforcement actions, and improving environmental education with traders and local fishermen. However, it is imperative to assess the real impact of fisheries and bycatch on the S. guianensis population of the Northern Brazilian coast, as a way to understand how the population has been affected over the years and how this trade is supplied.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because teeth used in this study were collected from stranded carcasses or from handicrafts sold at local markets. Sampling permits were granted under licenses numbers No. 30327-1 and No. 54305-1.
GFR, LRO, and SS: study design, data analysis, and writing the manuscript. RE-L, AC, FA, FL, and SS: fieldwork. GFR: Methodology and data curation. GFR, LRO, and SS: Supply reagents, materials, and analysis tools. GFR, LRO, RE-L, AC, FA, FL, and SS: reviewing and editing the manuscript. All authors contributed to the article and approved the submitted version.
GFR was supported by Coordination of Superior Level Staff Improvement (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES) (grant number 88887.204911/2018-00). SS and LRO were supported by Brazilian National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq)
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank professor Juliano Morales Oliveira, from UNISINOS University (Universidade do Vale do Rio dos Sinos) for wisely guiding us in the statistical analysis and design. We thank to GEMAM staff (Grupo de Estudos de Mamíferos Aquáticos da Amazônia) for field support. Sampling permits were granted by ICMBio through the SISBIO (Sistema de Autorização e Informação em Biodiversidade) with licenses numbers No. 30327-1 and No. 54305-1. Many thanks to GEMM-Lagos (Grupo de Estudos de Mamíferos Marinhos da Região dos Lagos) and GEMARS (Grupo de Estudos de Mamíferos Aquáticos do Rio Grande do Sul) for the infrastructure support, and to the Mammal Ecology Lab (LEM) from UNISINOS University for the infrastructure and technical support. We also want to thank the Coordination of Superior Level Staff Improvement (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES), from Brazilian Ministry of Education, that provided the doctoral scholarship for GFR in the Ecology and Natural Resources Graduate Program (Programa de Pós-Graduação em Ecologia e Recursos Naturais, PPG-ERN) of the Darcy Ribeiro State University of Northern Rio de Janeiro (Universidade Estadual do Norte Fluminense Darcy Ribeiro, UENF). Finally, we thank to Brazilian National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq) that provided the Research Productivity grants to LRO and SS.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.867068/full#supplementary-material
Akin P. A. (1988). Geographic Variation in Tooth Morphology and Dentinal Patterns in the Spinner Dolphin, Stenella Longirostris. Mar. Mamm. Sci. 4, 132–140. doi: 10.1111/j.1748-7692.1988.tb00193.x
Alves R. R. N., Rosa I. L. (2008). Use of Tucuxi Dolphin Sotalia Fluviatilis for Medicinal and Magic/Religious Purposes in North of Brazil. Hum. Ecol. 36, 443–447. doi: 10.1007/s10745-008-9174-5
Alves R. R. N., Souto W., Oliveira R., Barboza R. R. D., Rosa I. (2013). Aquatic Mammals Used in Traditional Folk Medicine: A Global Analysis, in Animals in Traditional Folk Medicine. Eds. Alves R. R. N., Rosa I. L. (Heidelberg, Berlin: Springer-Verlag), 241–261. doi: 10.1007/978-3-642-29026-8_11
Arcoverde D., Emin-Lima R., Costa A., Di Beneditto A., Siciliano S., Sena L., et al. (2014). Evaluation of Periotic–Tympanic Bone Complex of Sotalia Guianensis (Cetacea: Delphinidae) as Tool in Identification of Geographic Variations. J. Mar. Biol. Assoc. U.K. 94, 1127–1132. doi: 10.1017/S0025315413001689
Auguie B. (2017) Gridextra: Miscellaneous Functions for "Grid" Graphics. R Package Version 2.3. Available at: https://CRAN.R-project.org/package=gridExtra.
Barbosa-Filho M. L. V., Barreto R. M. F., Siciliano S., Seminara C. I., Costa-Neto E. M. (2018). Use of Cetaceans as Bait in Southern Bahia, Brazil, by Expert Fishermen That Market Shark Fins: A Lucrative Trade and Two Threatened Zoological Groups. Ethnobiol. Lett. 9, 12–18. doi: 10.14237/ebl.9.2.2018.953
Barlow J., Boveng P. (1991). Modelling Age-Specific Mortality for Marine Mammal Populations. Mar. Mamm. Sci. 7, 50–65. doi: 10.1111/j.1748-7692.1991.tb00550.x
Beltrán-Pedreros S., Petrere M. (2010). “Fishery Activity Impact on the Sotalia Populations From the Amazon Mouth,” in Biology, Evolution and Conservation of River Dolphins Within South America and Asia. Eds. Ruiz-Garcia M., Shostell J. M. (New York: Nova Science Publishers, Inc), 219–236.
Borobia M. (1989). Distribution and Morphometrics of South American Dolphins of the Genus Sotalia. Master’s Thesis (Montreal, Quebec: Macdonald College of McGill University).
Briceño Y., Sánchez L., Trujillo F., von Fersen L., Ramírez S. (2021). Aquatic Wildmeat Consumption of Guiana Dolphins (Sotalia Guianensis) in Lake Maracaibo System, Venezuela. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.625801
Brito T. P. (2012). O Conhecimento Ecológico Local E a Interação De Botos Com a Pesca No Litoral do Estado do Pará, Região Norte - Brasil. Biotemas 25, 259–277. doi: 10.5007/2175-7925.2012v25n4p259
Brown S., Reid D., Rogan E. (2014). Characteristics of Fishing Operations, Environment and Life History Contributing to Small Cetacean Bycatch in the Northeast Atlantic. PloS One 9, e104468. doi: 10.1371/journal.pone.0104468
Brum S. M., Da Silva V. M. F., Rossoni F., Castello L. (2015). Use of Dolphins and Caimans as Bait for Calophysus Macropterus (Lichtenstein 1819) (Siluriforme: Pimelodidae) in the Amazon. J. Appl. Ichthyol. 31, 675–680. doi: 10.1111/jai.12772
Brunner S. (2004). Fur Seals and Sea Lions (Otariidae): Identification of Species and Taxonomic Review. System. Biodivers. 1, 339–439. doi: 10.1017/S147720000300121X
Byrd B. L., Hohn A. A. (2017). Differential Risk of Bottlenose Dolphin (Tursiops Truncatus) Bycatch in North Carolina, USA. Aquat. Mamm. 43, 558–569. doi: 10.1578/AM.43.5.2017.558
Campbell E., Pasara-Polack A., Mangel J. C., Alfaro-Shigueto J. (2020). Use of Small Cetaceans as Bait in Small-Scale Fisheries in Peru. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.534507
Conversani V. R., Silva D. F., Barbosa R. A., Hohn A. A., Santos M. C. D. O. (2020). Age and Growth of Franciscana, Pontoporia Blainvillei, and Guiana, Sotalia Guianensis, Dolphins From Southeastern Brazil. Mar. Mammal Sci. 37, 702–716. doi: 10.1111/mms.12763
Costa A. F., Siciliano S., Emin-Lima R., Martins B. M. L., Sousa M. E. M., Giarrizzo T. (2017). Stranding Survey as a Framework to Investigate Rare Cetacean Records of the North and North-Eastern Brazilian Coasts. ZooKeys 688, 111–134. doi: 10.3897/zookeys.688.12636
Cravalho M. A. (1999). Shameless Creatures: An Ethnozoology of the Amazon River Dolphin. Ethnology 38, 47–58. doi: 10.2307/3774086
Da Silva V. M. F. D. (1994). Aspects of the Biology of the Amazonian Dolphins Inia and Sotalia Fluviatilis. Doutoral Tese (Cambridge: Cambridge University).
Da Silva V. M., Best R. C. (1996). Freshwater Dolphin/Fisheries Interaction in the Central Amazon (Brazil). Amazoniana 14, 165–175.
Da Silva V., Martin A., Fettuccia D., Bivaqua L., Trujillo F. (2020). Sotalia Fluviatilis. IUCN Red List Threat. Species 2020, e.T190871A50386457. doi: 10.2305/IUCN.UK.2020-3.RLTS.T190871A50386457.en
De Moura J. F., Sholl T. G. C., Da Silva-Rodrigues É., Hacon S., Siciliano S. (2009). Marine Tucuxi Dolphin (Sotalia Guianensis) and its Interaction With Passive Gill-Net Fisheries Along the Northern Coast of the Rio De Janeiro State, Brazil. Mar. Biodivers. Rec 2, E82. doi: 10.1017/S1755267209000864
De Vries A., Ripley B. D. (2020) Ggdendro: Create Dendrograms and Tree Diagrams Using 'Ggplot2'. R Package Version 0.1.22. Available at: https://CRAN.R-project.org/package=ggdendro.
Di Beneditto A. P. M., Ramos R. M. A. (2004). Biology of the Marine Tucuxi Dolphin (Sotalia Fluviatilis) in South-Eastern Brazil. J. Mar. Biolog. Assoc. 84, 1245–1250. doi: 10.1017/S0025315404010744h
Dos Santos-Filgueira C. H. M., Zappes C. A., Vidal M. D., Nunes J. L. S. (2021). Traditional Knowledge of Artisanal Fishers and Sotalia Guianensis (Van Bénéden 1864) (Cetacea, Delphinidae) in the Extractive Reserve Baía do Tubarão (Brazilian Amazon Coast). Coast Manage. 210, 105700. doi: 10.1016/j.ocecoaman.2021.105700
Emin-Lima N. R. E. D. (2012). Preenchendo Lacunas Em Saúde De Ecossistemas: Estudo Morfológico E De Contaminantes Nos Botos-Cinza (Sotalia Guianensis, Van Benéden 1864) da costa Norte do Brasil (Rio de Janeiro (RJ: Escola Nacional de Saúde Pública Sergio Arouca, Fundação Oswaldo Cruz, Rio de Janeiro).
Fettuccia D. D. C. (2010). Variação Osteológica E Desenvolvimento Ontogenético Das Espécies do Gênero Sotalia (Cetacea, Delphinidae) (Manaus (AM: Instituto Nacional de Pesquisas da Amazônia).
Fettuccia D. D. C., Da Silva V. M. F., Rocha M. S., Simões-Lopes P. C. (2012). Sternum and Appendicular Skeleton: Morphometric Differences Between the Species of Genus Sotalia (Cetacea: Delphinidae). J. Mar. Biolog. Assoc. 92, 1657–1662. doi: 10.1017/S0025315412000604
Fettuccia D. D. C., Da Silva V. M. F., Simões-Lopes P. C. (2009). Non-Metric Characters in Two Species of Sotalia (Gray 1866) (Cetacea, Delphinidae). Braz. J. Biol. 69, 907–917. doi: 10.1590/S1519-69842009000400020
Flores P. A., Da Silva V. M., Fettuccia D. D. C. (2018). “Tucuxi and Guiana Dolphins: Sotalia Fluviatilis and S. Guianensis,” in Encyclopedia of Marine Mammals, 3rd ed. Eds. Würsig B., Thewissen J. G. M., Kovacs K. M. (London: Academic Press), 1024–1027. doi: 10.1016/B978-0-12-804327-1.00264-8
Ford J. K., Ellis G. M., Matkin C. O., Wetklo M. H., Barrett-Lennard L. G., Withler R. E. (2011). Shark Predation and Tooth Wear in a Population of Northeastern Pacific Killer Whales. Aquat. Biol. 11, 213–224. doi: 10.3354/ab00307
Fox J., Weisberg S. (2019). An {R} Companion to Applied Regression. Third Edition (Thousand Oaks, CA: Sage). Available at: https://socialsciences.mcmaster.ca/jfox/Books/Companion/.
Gravena W., Hrbek T., Da Silva V. F., Farias I. (2008). Amazon River Dolphin Love Fetishes: From Folklore to Molecular Forensics. Mar. Mammal Sci. 24, 969–978. doi: 10.1111/j.1748-7692.2008.00237.x
Hale R. L., Dougherty D. (1988). Differences Between Ward's and UPGMA Methods of Cluster Analysis: Implications for School Psychology. J. Sch. Psychol. 26, 121–131. doi: 10.1016/0022-4405(88)90014-3
Hennig C. (2007). Cluster-Wise Assessment of Cluster Stability. Comput. Stat. Data Anal. 52, 258–271. doi: 10.1016/j.csda.2006.11.025
Hennig C. (2020) Fpc: Flexible Procedures for Clustering. R Package Version 2. Available at: https://CRAN.R-project.org/package=fpc.
Hohn A. A., Scott M. D., Wells R. S., Sweeney J. C., Irvine A. B. (1989). Growth Layers in Teeth From Known-Age, Free-Ranging Bottlenose Dolphins. Mar. Mammal Sci. 5, 315–342. doi: 10.1111/j.1748-7692.1989.tb00346.x
Iriarte V., Marmontel M. (2013a). River Dolphins (Inia Geoffrensis, Sotalia Fluviatilis) Mortality Events Attributed to Artisanal Fisheries in the Western Brazilian Amazon. Aquat. Mamm. 39, 116–124. doi: 10.1578/AM.39.2.2013.116
Iriarte V., Marmontel M. (2013b). Insights on the Use of Dolphins (Boto, Inia Geoffrensis and Tucuxi, Sotalia Fluviatilis) for Bait in the Piracatinga (Calophysus Macropterus) Fishery in the Western Brazilian Amazon. J. Cetacean Res. Manage. 13, 163–173.
Kassambara A., Mundt F. (2020) Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. Available at: https://CRAN.R-project.org/package=factoextra.
Klevezal G. A. (1996). Recording Structures of Mammals: Determination of Age and Reconstruction of Life History. 1st Ed. Mina M.V., Oreshkin A.V. Trans. (Rotterdam: A.A. Balkema).
Lima J. Y., Carvalho A. P. M., Azevedo C. T., Barbosa L. A., Silveira L. S. (2016). Variation of Age and Total Length in Sotalia Guianensis (Van Bénéden 1864) (Cetacea, Delphinidae), on the Coast of Espírito Santo State. Brazil. Braz. J. Biol. 77, 437–443. doi: 10.1590/1519-6984.13215
Mannocci L., Dabin W., Augeraud-Véron E., Dupuy J. F., Barbraud C., Ridoux V. (2012). Assessing the Impact of Bycatch on Dolphin Populations: The Case of the Common Dolphin in the Eastern North Atlantic. PloS One 7, e32615. doi: 10.1371/journal.pone.0032615
Martins A. R., Da Silva V. M. F., Salmon D. L. (2004). Riverine Habitat Preferences of Botos (Inia Geoffrensis) and Tucuxis (Sotalia Fluviatilis) in the Central Amazon. Mar. Mammal Sci. 20, 2189–2200. doi: 10.1111/j.1748-7692.2004.tb01150.x
Meirelles A. D., Ribeiro A. C., Silva C. P. N., Soares-Filho A. A. (2010). Records of Guiana Dolphin, Sotalia Guianensis, in the State of Ceará, Northeastern Brazil. Lat. Am. J. Aquat. Res. 8, 97–102. doi: 10.5597/lajam00157
Monteiro-Filho E. L. A., Monteiro L. R., Dos Reis S. F. (2002). Skull Shape and Size Divergence in Dolphins of the Genus Sotalia: A Tridimensional Morphometric Analysis. J. Mammal. 83, 125–134. doi: 10.1644/1545-1542(2002)083<0125:SSASDI>2.0.CO;2
Moore J. E., Read A. J. (2008). A Bayesian Uncertainty Analysis of Cetacean Demography and Bycatch Mortality Using Age-at-Death Data. Ecol. Appl. 18, 1914–1931. doi: 10.1890/07-0862.1
NatureServe and IUCN (International Union for Conservation of Nature), (2021). Genus Sotalia. The IUCN Red List of Threatened Species. Version 2018-2. Available at: https://www.iucnredlist.org. Downloaded on 15 Nov.
Novais R. F., Siciliano S., Emin-Lima R., Silva J. S. (2020). Ontogeny of the Skull of Sotalia Guianensis (Cetartiodactyla: Delphinidae) on the Northern Coast of Brazil. Iheringia Série Zoologia 110, e2020024. doi: 10.1590/1678-4766e2020024
Podani J. (2000). Introduction to the Exploration of Multivariate Biological Data (Leiden (The Netherlands: Backhuys Publishing).
Ramos R. M. A., Di Beneditto A. P. M., Lima N. R. W. (2000). Relationship Between Dental Morphology, Sex, Body Length and Age in Pontoporia Blainvillei and Sotalia Fluviatilis (Cetacea) in Northern Rio De Janeiro. Brazil. Braz. J. Biol. 60, 283–290. doi: 10.1590/S0034-71082000000200012
Ramos R. M. A., Di Beneditto A. P. M., Siciliano S., Santos M. C. O., Zerbini A. N., Vicente A. F. C., et al. (2010). Morphology of the Guiana Dolphin (Sotalia Guianensis) Off Southeastern Brazil: Growth and Geographic Variation. Lat. Am. J. Aquat. Res. 8, 137–149. doi: 10.5597/lajam00163
R Core Team (2020). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Read F. L., Hohn A. A., Lockyer C. H. (2018). A Review of Age Estimation Methods in Marine Mammals With Special Reference to Monodontids. NAMMCO Sci. Publ. 10, 1–67. doi: 10.7557/3.4474
Reilly S. B., Barlow J. (1986). Rates of Increase in Dolphin Population Size. Fish. Bull. 84, 527–533.
Rosas F. C. W., Barreto A. S., Monteiro-Filho E. L. D. A. (2003). Age and Growth of the Estuarine Dolphin Sotalia Guianensis (Cetacea, Delphinidae) on the Paraná Coast, Southern Brazil. Fish. Bull. 101, 377–383.
Rosas F. C. W., Monteiro-Filho E. L. (2002). Reproduction of the Estuarine Dolphin (Sotalia Guianensis) on the Coast of Paraná, Southern Brazil. J. Mammal. 83, 507–515. doi: 10.1644/1545-1542(2002)083%3C0507:ROTEDS%3E2.0.CO;2
RStudio Team (2020). RStudio: Integrated Development Environment for R (Boston, MA: RStudio, PBC). Available at: http://www.rstudio.com/.
Scheffer V. B., Myrick A. C. (1980). A Review of Studies to 1970 of Growth Layers in the Teeth of Marine Mammals. Rep. Int. Whal. Commn. 3, 51–63.
Secchi E., Santos M. C., de O., Reeves R. (2018). Sotalia guianensis (errata version published in 2019) . The IUCN Red List of Threatened Species, 2018: e.T181359A144232542. doi: 10.2305/IUCN.UK.2018-2.RLTS.T181359A144232542.en
Sholl T. G. C., Nascimento F. F., Leoncini O., Bonvicino C. R., Siciliano S. (2008). Taxonomic Identification of Dolphin Love Charms Commercialized in the Amazonian Region Through the Analysis of Cytochrome B DNA. J. Mar. Biol. Assoc. 88, 1207–1210. doi: 10.1017/S002531540800043X
Siciliano S. (1994). Review of Small Cetaceans and Fishery Interactions in Coastal Waters of Brazil. Rep. Int. Whal. Commn. 15, 241–250.
Siciliano S., Emin-Lima N. R., Costa A. F., Rodrigues A. L. F., De Magalhães F. A., Tosi C. H., et al. (2008). Revisão do Conhecimento Sobre Os Mamíferos Aquáticos Da Costa Norte do Brasil. Arq. do Mus. Nac. 66, 1–21.
Siciliano S., Viana M. C., Emin-Lima R., Bonvicino C. R. (2018). Dolphins, Love and Enchantment: Tracing the Use of Cetacean Products in Brazil. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00107
Smith A. (1996). ”The River Dolphins: The Road to Extinction“, in ed. Simmonds J. D., Hutchinson M. P.The Conservation of Whales and Dolphins: Science and Practice. (Chichester: John Wiley and Sons), pp. 339–373.
Tarnawski B. A., Cassini G. H., Flores D. A. (2014). Allometry of the Postnatal Cranial Ontogeny and Sexual Dimorphism in Otaria Byronia (Otariidae). Acta Theriol. 59, 81–97. doi: 10.1007/s13364-012-0124-7
Tosi C. H., Magalhães F. A., Garri R. G. (2009). Meat Consumption of a Fraser's Dolphin (Lagenodelphis Hosei) Stranded Alive on the Northern Brazilian Coast. Mar. Biodivers. Rec 2, E4. doi: 10.1017/S1755267208000043
Vidal M. D., De Moura M. F., Santos-Muniz G. P. (2019). Conhecimentos E Crenças De Pescadores Artesanais Sobre Os Golfinhos Fluviais do Médio Rio Tapajós, Pará. R. Bras. Bioci. 17, 53–60.
Wickham H. (2016). Ggplot2: Elegant Graphics for Data Analysis (New York: Springer-Verlag). Available at: https://ggplot2.tidyverse.org.
Keywords: age, sotalia, guiana dolphin, tucuxi, love-charms, teeth, northern Brazil, amazon
Citation: Ruenes GF, Oliveira LRd, Emin-Lima R, Costa AF, Attademo FLN, Luna FdO and Siciliano S (2022) Who Is in Handicrafts: Tooth Morphology and Age Determination as Tools to Assess Vulnerable Dolphins Sotalia spp. Supplying the Trade of Artisanal Charms in Brazil. Front. Mar. Sci. 9:867068. doi: 10.3389/fmars.2022.867068
Received: 31 January 2022; Accepted: 02 May 2022;
Published: 10 June 2022.
Edited by:
James Scott Maki, Marquette University, United StatesReviewed by:
Hector Barrios-Garrido, University of Zulia, VenezuelaCopyright © 2022 Ruenes, Oliveira, Emin-Lima, Costa, Attademo, Luna and Siciliano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Greicy F. Ruenes, Z3JleXJ1ZW5lc0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.