 Tim Awbery1,2*
Tim Awbery1,2* Aylin Akkaya1,3
Aylin Akkaya1,3 Patrick Lyne1
Patrick Lyne1 Laura Rudd1
Laura Rudd1 Gemma Hoogenstrijd1
Gemma Hoogenstrijd1 Madeleine Nedelcu1David Kniha1Mehmet Akif Erdoğan4
Madeleine Nedelcu1David Kniha1Mehmet Akif Erdoğan4 Cherisse Persad1
Cherisse Persad1 Ayaka Amaha Öztürk5
Ayaka Amaha Öztürk5 Bayram Öztürk5
Bayram Öztürk5- 1DMAD-Marine Mammals Research Association, Antalya, Turkey
- 2Marine Mammals Research Team, Scottish Association for Marine Science, Scottish Marine Institute, Oban, United Kingdom
- 3Doğal Hayatı Koruma Vakfı World Wildlife Fund (WWF)-Turkey, Istanbul, Turkey
- 4Landscape Architecture Department, Faculty of Architecture, Hatay Mustafa Kemal University, Hatay, Turkey
- 5Turkish Marine Research Foundation (TUDAV), Istanbul, Turkey
Marine traffic has been identified as a serious threat to Mediterranean cetaceans with few mitigation strategies in place. With only limited research effort within the Eastern Basin, neither baseline species knowledge nor the magnitude of threats have been comprehensively assessed. Delineating the extent of overlap between marine traffic and cetaceans provides decision makers with important information to facilitate management. The current study employed the first seasonal boat surveys within the Eastern Mediterranean Sea of Turkey, incorporating visual and acoustic survey techniques between 2018 and 2020 to understand the spatial distribution of cetacean species. Additionally, marine traffic density data were retrieved to assess the overlap with marine traffic. Encounter rates of cetaceans and marine traffic density were recorded for each 100 km2 cell within a grid. Subsequently, encounter and marine traffic density data were used to create a potential risk index to establish where the potential for marine traffic and cetacean overlap was high. Overall, eight surveys were undertaken with a survey coverage of 21,899 km2 between the Rhodes and Antalya Basins. Deep diving cetaceans (sperm and beaked whales) were detected on 28 occasions, with 166 encounters of delphinids of which bottlenose, striped and common dolphins were visually confirmed. Spatially, delphinids were distributed throughout the survey area but encounter rates for both deep diving cetaceans and delphinids were highest between the Rhodes and Finike Basins. While sperm whales were generally detected around the 1000m contour, delphinids were encountered at varying depths. Overall, two years of monthly marine traffic density were retrieved with an average density of 0.37 hours of monthly vessel activity per square kilometer during the study period. The mean density of vessels was 0.32 and 1.03 hours of monthly vessel activity per square kilometer in non-coastal and coastal waters respectively. The Eastern Mediterranean Sea has several important shipping lanes within the study area. Two priority areas for deep diving cetacean and a large priority area for cetaceans were identified in the waters between Marmaris and Finike where high cetacean encounters and dense marine traffic overlapped. The current study revealed important habitats for cetaceans within the data deficient Eastern Mediterranean Sea and delineated potential risk area where marine traffic should be limited.
Introduction
The Mediterranean Sea is a semi-enclosed body of water that contains extensive areas of abyssal waters, deep basins and trenches bounded by steep slopes. The most easterly part of the Mediterranean Sea is also known as the Levantine Sea, of which Turkey’s coastlines covers 1577km. Previous studies have reported seven species of cetaceans commonly observed within the Eastern Mediterranean Sea of Turkey; sperm whales (Physeter macrocephalus), Cuvier’s beaked whales (Ziphius cavirostris), fin whales (Balaenoptera physalus), bottlenose dolphins (Tursiops truncatus), striped dolphins (Stenella coeruleoalba), common dolphins (Delphinus delphis), and Risso’s dolphins (Grampus griseus) (Güçlüsoy et al., 2014; Akkaya Baş et al., 2016; Öztürk et al., 2016; Akkaya et al., 2020).
Sperm whales and Cuvier’s beaked whales are deep-diving species that generally live in pelagic areas characterized by deep basins and trenches or steep slopes (Heyning, 1989; Praca and Gannier, 2008; Praca et al., 2009; Notarbartolo di Sciara and Birkun Jr, 2010). As such, their distribution is closely related to bathymetry, with distinctive preference for depths of 1000m and 500-1500m for sperm whales and Cuvier’s beaked whales, respectively (Frantzis et al., 2003; Boisseau et al., 2010; Frantzis et al., 2014). Sperm whales in the Mediterranean have been described as genetically different from the Atlantic population, with estimates of less than 300 individuals for the Eastern Mediterranean (Drouot et al., 2004; Engelhaupt et al., 2009; Frantzis et al., 2014; Lewis et al., 2018; Frantzis et al., 2019). Ozturk et al. (2013) reported 43 sperm whale sightings in Turkish waters between 1999 and 2014, with the majority of the sightings occurring between Fethiye and the Rhodes Basin in the Anadolu Submarine Canyon, one of the deepest parts of the Mediterranean Sea. The whales were, however, present from Gökçeada in the northern Aegean to an eastern limit of Alanya in Turkey (Öztürk et al., 2013). Evidence suggests that the Mediterranean subpopulation is declining and is currently listed as ‘Endangered’ under the IUCN Red List (Reeves and Notarbartolo di Sciara, 2006; Notarbartolo di Sciara et al., 2012; Pirotta et al., 2021). Cuvier’s beaked whales are also regularly observed in the Eastern Mediterranean Sea, specifically in the Finike (Anaximander) Seamounts, the Antalya Canyon and the Adana Trough (Akkaya Baş et al., 2016; Cañadas and Notarbartolo di Sciara, 2018) and strandings have occurred from Gökçeada to Karataş in the east of Turkey (Öztürk et al., 2011). The species have been recently categorized as ‘Vulnerable’ on the IUCN Red List (Cañadas and Notarbartolo di Sciara, 2018). Fin whales are also considered to be ‘Vulnerable’ with a declining population (Panigada and Notarbartolo di Sciara, 2012) and scattered sightings and strandings have been reported along the Turkish Mediterranean coast (Stephens et al., 2021). In addition to deep diving species, four species of delphinid have been recorded in the Turkish waters. Populations of bottlenose dolphins have become increasingly fragmented and are listed together with striped dolphins as ‘Vulnerable’ by the IUCN (Bearzi et al., 2012; Aguilar and Gaspari, 2012). Similarly, common dolphins have a patchy distribution in the Mediterranean and are considered ‘Endangered’ on the IUCN Red List (Bearzi, 2003; Bearzi et al., 2021). Additionally, Risso’s dolphins were recently recategorized from ‘Data Deficient’ to ‘Endangered’ by the IUCN (Lanfredi et al., 2021) due in part to an estimated 50% reduction in individuals over a ten year-period in some areas of the Mediterranean (Airoldi et al., 2015; Azzellino et al., 2016). Their abundance appears even scarcer in the Eastern Mediterranean Sea, with just a handful of documented sightings and strandings (Öztürk et al., 2011; Dede et al., 2012; Kerem et al., 2012; Ryan et al., 2014).
Anthropogenic threats and their impacts on the conservation status of cetacean species in the Mediterranean are of significant concern (Boisseau et al., 2010). An increasing population has developed the necessity for increased development along the coast (both for housing and for tourism), increased exploitation of natural resources (both for food and for fuel), and increased shipping for trade. This has resulted in threats to marine mammals in the Turkish waters of the Eastern Mediterranean Sea from habitat destruction, unsustainable fishing practices (Reeves and Notarbartolo di Sciara, 2006; Notarbartolo di Sciara and Birkun Jr., 2010), chemical and noise pollution (Frantzis, 1998; Frantzis, 2004; Notarbartolo di Sciara and Birkun Jr., 2010; Notarbartolo di Sciara et al., 2012; Fylaktos and Papanicolas, 2019) and ship strikes (Laist et al., 2001; Notarbartolo di Sciara and Birkun Jr., 2010; Frantzis et al., 2014). Marine traffic may contribute to the degradation and loss of cetacean habitat through temporal or permanent changes in habitat preference (Bejder et al., 2006a; Rako et al., 2013; Campana et al., 2015), short-term changes in behavior (Jahoda et al., 2003; Aguilar Soto et al., 2006; Bejder et al., 2006b; Tyack et al., 2011), or direct physical injuries due to collisions (Panigada et al., 2006). While local traffic tends to present a threat to coastal populations, maritime transport tends to follow direct routes around land masses with a safe distance from shore, often around the 1000m depth contour (Frantzis et al., 2019). As a consequence, deep-diving cetacean species are susceptible to the high intensity of shipping in the Mediterranean (Notarbartolo di Sciara and Birkun Jr., 2010; Frantzis et al., 2019). Over 6.2% of stranded sperm whales in Italy and Greece were caused by collisions with ships, as well as 6.6% of photo-identified individuals having visible injuries attributed to ship strikes (Pesante et al., 2002; Abdulla and Linden, 2008). Additionally, in the Pelagos Sanctuary, 3% of strandings were attributed to ship strikes between 1972 and 2018 with 59% of observed injured whales showing evidence of collisions with ships (Panigada et al., 2020).
Thus far, there has been an inequality in the collection of data between the eastern and western Mediterranean (Frantzis et al., 2003; Akkaya et al., 2020) which means that it is likely that patches of cetacean habitat remain unidentified and may overlap with a range of the aforementioned anthropogenic disturbances (Akkaya et al., 2020). The current study maps the distribution of the coastal and offshore cetacean species and identifies potential risk habitats that are likely to be under pressure from maritime traffic. The Eastern Mediterranean Sea suffers from a dearth of knowledge surrounding marine megafauna. Providing more knowledge on cetacean presence, critical habitats and anthropogenic threats will offer guidance to stakeholders forming policy and aid the development of species conservation action plans for cetaceans in the Turkish waters of the Eastern Mediterranean Sea.
Materials and Methods
Study Area
The study area includes coastal zones, the continental shelf and the high seas surrounding the Turkish coast within the Eastern Mediterranean Sea (Figure 1), extending 55 km offshore and approximately 470 km along the Turkish Mediterranean coast from Marmaris to Anamur. The main survey area covered an area of 16,396 km² in the Rhodes and Finike Basins with an additional section to the east of 5,503 km² in the Antalya Basin. In the south, there was partial overlap with the most northerly section of the Special Environmental Protection Area, “Finike Seamount Special Protected Area”, and in the west, a small area of overlap with the eastern edge of the Marmaris National Park. It also overlaps with two Important Marine Mammal Areas (IMMAs). The Hellenic Trench IMMA covers the entirety of the Hellenic trench in Greece as well as the Turkish submarine canyons (IUCN-MMPATF, 2017a; Notarbatolo di Sciara and Hoyt, 2020). A small portion of the Cilician Basin IMMA (IUCN-MMPATF, 2017b), designated for the presence of the Mediterranean monk seals, overlaps the survey area in the east (Figure 1).
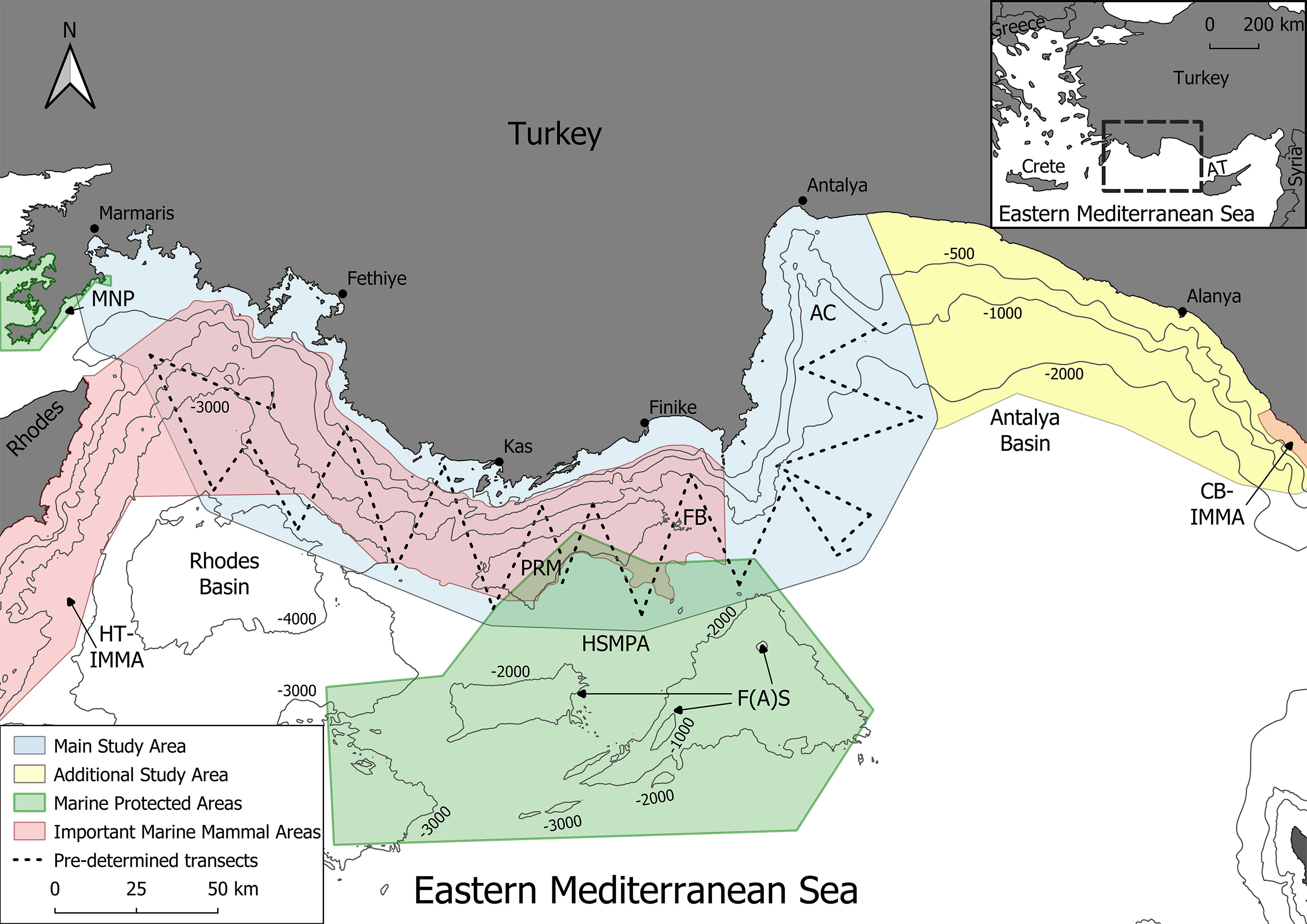
Figure 1 The main study area in the Eastern Mediterranean Sea which was surveyed using predetermined transects (shown in light blue) as well as the additional survey area (yellow), current marine protected areas (green) and Important Marine Mammal Areas (pink). The pre-determined transects are overlaid as dashed lines. Abbreviations: FB: Finike Basin, AC: Antalya Canyon, AT (inset): Antalya Trough, F(A)S: Finike (Anaximander) Seamounts, PRM: Piri Reis Sea Mountains, HSMPA: High Seas Marine Protected Area, MNP: Marmaris National Park, HT-IMMA: Hellenic Trench, Deep Divers’ Sanctuary Important Marine Mammal Area, CB-IMMA: Cilician Basin Important Marine Mammal Area.
Data Collection
Survey data were collected from a 13.95m sailing vessel with a 100hp diesel engine which typically travelled at 4 knots during search efforts. This speed was selected to avoid introducing significant hydrophone flow, propeller, or engine noise. The vessel followed equally spaced zigzag transects designed using DISTANCE software, Version 7.3 as well as the 1000m bathymetric contour line. The predetermined tracklines comprised 22 transects, with a total route of 644 km. The series of visual and acoustic surveys along these transect lines were carried out seasonally between 16th April 2018 and 14th January 2020. In addition, the eastern section of the survey area (Figure 1) was surveyed in April 2019 only.
Visual surveys were conducted during daylight hours, starting half an hour before sunrise and finishing half an hour after sunset, in Beaufort Sea states ≤4. Two observers with binoculars were stationed at the bow of the vessel, one scanning to port (270 to 10 degrees) and the other to starboard (350 to 90 degrees). The data logging software, Logger, Version 2010 was run continuously for the full duration of each survey. Effort status and environmental information were entered on an hourly basis or when conditions changed considerably. During the visual data collection, species identification, group size, group composition, behavior, group cohesion, group bearing and distance, photo-identification data and anthropogenic presence in the area were recorded.
The acoustic surveys were conducted 24 hours per day for the full duration of the survey, using a four omni-directional broadband hydrophone array towed 200m behind the vessel. The hydrophone elements can be used for high and low frequency monitoring and have a range between 10Hz – 200kHz. Acoustic signals were digitized using a Behringer U-Phoria UMC404HD sound card sampling up to 192kHz. The acoustic software PAMGuard, Version 1.15.15 Core (www.pamguard.org), was used throughout all surveys, scanning incoming signals for clicks and whistles. The PAM operators on the vessel were responsible for logging acoustic detections of cetaceans. The operator filled out an acoustic form at 15-minute time intervals for species presence, acoustic type and strength, and background noise. A scale from 0 (nothing heard) to 5 (nothing else can be heard) was used to quantify acoustic signals as in Ryan et al. (2014). Both visual and acoustic data were used in the subsequent analysis of the relative abundance of each species.
Data Analysis
During analysis, both acoustic and visual detections were classed as encounters and detected species were mapped at point of first detection for each cetacean group in QGIS (version 3.14). If a cetacean group was logged both visually and acoustically (determined by a difference in detection time of less than 2 minutes), then only the visual detection was used. This was to ensure that there was no overestimation of abundance. In order to perform spatial comparisons of different areas, a grid of hexagonal cells was created spanning the survey area. Hexagonal tessellation has shown advantages over the traditionally used square cells when visualizing data (Birch et al., 2007), as it offers a better representation of the visual and acoustic detection area around the vessel (Paradell et al., 2019). Further, hexagonal cells show a better connectivity, as the central point is the same distance from the central point of all neighboring cells (Birch et al., 2007; Paradell et al., 2019). The grid generated 286 hexagonal cells of height 10.746 km and width 10.746 km, corresponding to an area of 100 km2 per cell.
Cetacean Encounter Rates
When calculating the encounter rate, the number of groups in each hexagonal grid cell were summed using the ‘Count points in polygon’ tool in QGIS. The total distance travelled by the survey vessel in each cell was calculated using the ‘Sum line lengths’ tool in QGIS. In order calculate the encounter rates (ER), the following formula was used:
where n is the number of group encounters per cell and L is the survey effort (distance travelled by the survey vessel in kilometers). This value was then multiplied by 100 to get the encounter rate per 100km to make it comparable with other studies in the region (e.g., Boisseau et al., 2010; Ryan et al., 2014). The majority of encounters were detected acoustically, making group size more ambiguous as not all individuals in a group will necessarily be vocalizing at the same time. Thus, encounter rates were calculated as ‘number of groups of cetaceans per hundred kilometers’ rather than ‘number of individuals per hundred kilometers’. It is generally accepted that in order to avoid artificial inflation of encounter rates due to small sample biases, cells where a distance less than the diagonal of the cell has been covered by the research vessel (in this case 12.408km) should be removed from further analysis (e.g., Bearzi et al., 2006; Dinis et al., 2016). The total encounter rate was further pooled into delphinids and deep-diving cetaceans.
Depth data for each encounter was derived and integrated from the General Bathymetric Chart of the Oceans (GEBCO) using the ‘Sample from raster’ tool and distance to the nearest coast was calculated using the ‘Distance to nearest hub’ tool in QGIS.
Marine Traffic
Vessel density data were retrieved from the European Marine Observation and Data Network (EMODnet)’s Human Activities Data Portal (www.emodnet-humanactivities.eu). Full details on how vessel density by EMODnet is calculated are given in EMODnet (2019). In brief, EMODnet converts received Automatic Identification System messages from vessels (at a three-minute resolution) into reconstructions of ship track lines between these points. Each line is then intersected with a grid of cells at a 1km x 1km resolution and the length and duration attributes attached to each line are used to calculate the length of time spent by a vessel in each cell. As a result, vessel density for EMODnet data is expressed as hours per square kilometer per month (EMODnet, 2019).
Monthly raster data was retrieved from April 2018 until March 2020 (n=24 files) and data was mean averaged using the ‘Raster’ package in R (version 4.1.2) resulting in a single raster averaged across the two-year period. The zonal statistics tool in QGIS was then used to down-sample these data to match the resolution of the grid of 100km2 hexagonal cells to give the mean hours per square kilometer per month of each cell. In order to identify any seasonal variation present, monthly raster data was grouped into seasons [defined as spring: March, April, May; summer: June, July, August; autumn: September, October, November and winter: December, January, February (n = 6 files per season)]. For each season, the zonal statistics analysis used for the overall data was repeated to create four seasonal marine traffic density vector layers.
When comparing between coastal and offshore cells, coastal cells were considered those directly touching land or with a centroid that was 200m or shallower, whilst offshore cells did not intersect with land and had a centroid with a depth greater than 200m. In order to investigate which species were detected in cells with the most marine traffic, each detection was intersected with the corresponding seasonal marine traffic density vector layer using the ‘Join attributes by location’ tool in QGIS.
Potential Risk Areas
As this study represents the first two years of systematic cetacean surveys in this region, the dataset collected was not large enough to create species habitat suitability models as has previously been done when analyzing collision risk for larger datasets (e.g., Blondin et al., 2020). Thus, a simpler method was created to identify potential risk areas where both cetacean encounter rates and levels of marine traffic were high.
As deep diving cetaceans and delphinids travel considerable distances, a kernel density analysis was undertaken to create a raster surface first of deep diving cetaceans and then delphinids across the survey area. In order to do this, the centroid of each encounter rate cell created in 2.3.1 was created in QGIS. Using the encounter rate of each point as a weighting, a density raster was created using a ten-kilometer search radius and a 1km2 cell size (so as to correspond with the cell size of the existing marine traffic density raster).
Due to the skew of both marine traffic density and encounter rate data, a log-transformation was performed to coerce the data towards a normal distribution. During the log-transformation, raster cells with a value of zero for any raster, were given a ‘null’ value by QGIS as a result of trying to log-transform zero. As a zero value in any raster cell indicates there was no spatial overlap between cetaceans and marine traffic in this cell, it indicates there is no risk and so would have been given a null value in a later stage of the analysis, and therefore this was not considered an issue.
So that encounter rate and marine traffic had an approximately equal influence on the potential risk index, both encounter rate rasters were normalized between 0 and 1 using:
where zi is the ith normalized cell value in the raster, xi is the ith cell value in the original raster, min(x) is the minimum cell value in the raster and max(x) is the maximum value in the dataset. Potential risk index for each cell was then calculated as
where PRIi refers to the potential risk index for the ith cell, ERi refers to the transformed encounter rate for the ith raster cell is encounter rate >0 and MTDi refers to the transformed marine traffic density for the ith raster cell with marine traffic density >0. Values for potential risk index were then normalized between 0 and 1 to make them easier to interpret. A potential risk index was created for deep diving cetaceans and delphinids separately. All raster analyses were conducted in the raster calculator in QGIS. Due to the low percentage of cells with sufficient survey effort (>12.408km) at a seasonal scale, the potential risk index was only calculated for the total data and not for each season.
Critical Habitats
In order to identify critical habitats for deep diving cetaceans and delphinids, a kernel density analysis was again run in QGIS using the centroids of each of the hexagonal cells. The kernel density analysis was weighted using the potential risk index and used radii of 24.816km (i.e., twice the diagonal diameter of a hexagonal cell), and a cell size of 1km2. The ‘contour’ tool in QGIS was used to draw contours around the highest 20% of potential risk cells and then the ‘lines to polygons’ tool was used to convert these into polygons in order to measure the area of critical habitats for both deep diving cetaceans and for delphinids.
Results
Surveys with a total effort of 52 days were conducted from 16th April 2018 to 14th January 2020 which covered eight separate seasons (two surveys in each spring, summer, autumn and winter). Spring and summer were the seasons with the most survey effort (distance travelled by the research vessel) with 31.4% and 27.2% of the total survey effort, respectively. Autumn and winter were the least surveyed seasons with 16.2% and 24.6% of the survey effort in total (Table 1). During the study, 191 visual and acoustic detections were recorded, of which 25 were deep diving cetaceans, 25 were identified delphinids and 141 were unidentified delphinids (Table 1).
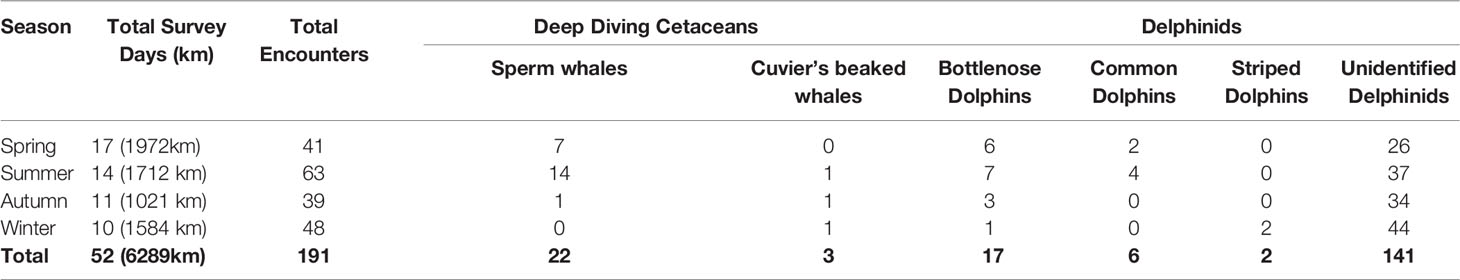
Table 1 Summary of cetacean encounters by season.
Survey Effort
Considering the zig-zag nature of transects, survey effort was generally well distributed within the core survey area with a survey effort which was greater than the diagonal of the cell (12.408km) to allow encounter rates to be calculated in 114 out of 227 cells (50.2%) in the main study area and just 2 cells in the additional survey area (Figure 2).
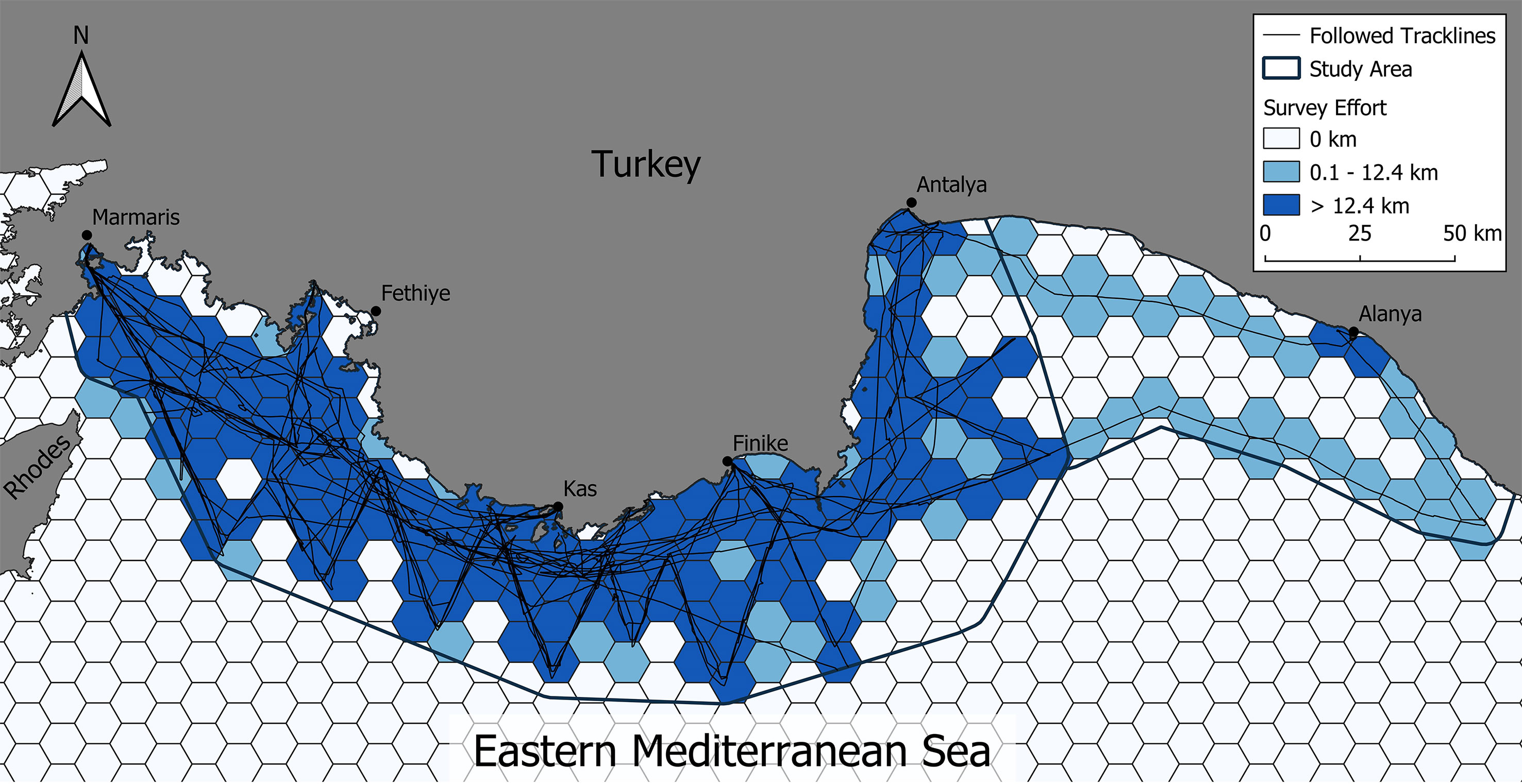
Figure 2 Survey effort in kilometres covered by the research vessel per cell in the Eastern Mediterranean Sea surrounding Turkey with darker blue cells showing areas with more than 12.408km of boat track line (suitable for encounter rate analysis), lighter blue cells showing less than 12.408km and white cells showing no survey effort.
Distribution and Encounter Rates of Deep Diving Cetaceans and Delphinids
Deep diving cetaceans were detected on 25 occasions, mainly to the west of the survey site between Rhodes and Kekova Island and consisted of sperm whales (n=22) and beaked whales (n=3). When accounting for survey effort (with insufficient survey effort removed), the median encounter rate for deep diving cells was 0 groups per 100 km due to a low number of overall detections with a mean of 0.31 groups per hundred kilometers. The highest encounter rate of 8.3 groups of deep diving cetaceans per 100km were in the waters off Fethiye. 60% of sperm whale detections (n = 13) occurred within 3km of the 1000m bathymetric contour. Sperm whale distribution had a median depth of 996.5m and a median distance from shore of 10.9km. Cuvier’s beaked whales had a slightly deeper median depth of 1,236m and were found at a similar median distance offshore of 9.6km (Figures 3, 4 and Table 2).
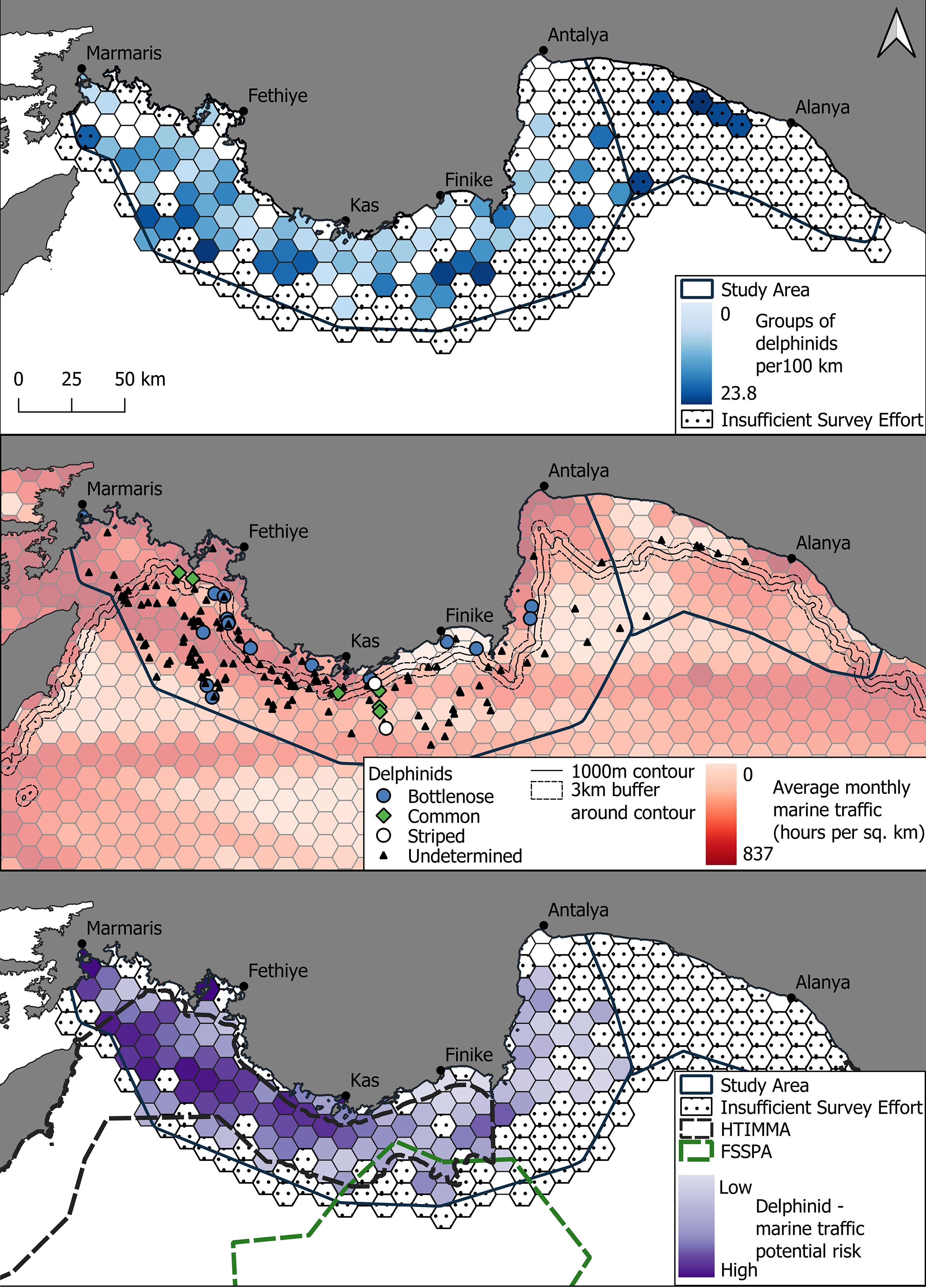
Figure 3 (Upper) Groups of delphinids per 100 km in the Eastern Mediterranean Sea with darkening blue representing an increased number of delphinid groups. (Middle) The distribution of different delphinid species with bottlenose, striped, common and undetermined groups of delphinids denoted by blue circles, green diamonds, hollow circles and filled triangles respectively. Distribution is superimposed onto the average monthly marine traffic (hours per square kilometre) with darkening red signifying higher traffic. Marine vessel information used in this figure was made available by the EMODnet Human Activities project, www.emodnet-humanactivities.eu, funded by the European Commission Directorate General for Maritime Affairs and Fisheries. The 1000m contour is shown with a 3km buffer area around the contour. (Lower) Potential risk areas where the presence of delphinids and marine traffic were both high during the study period. Darkening purple shows an increased potential for risk.
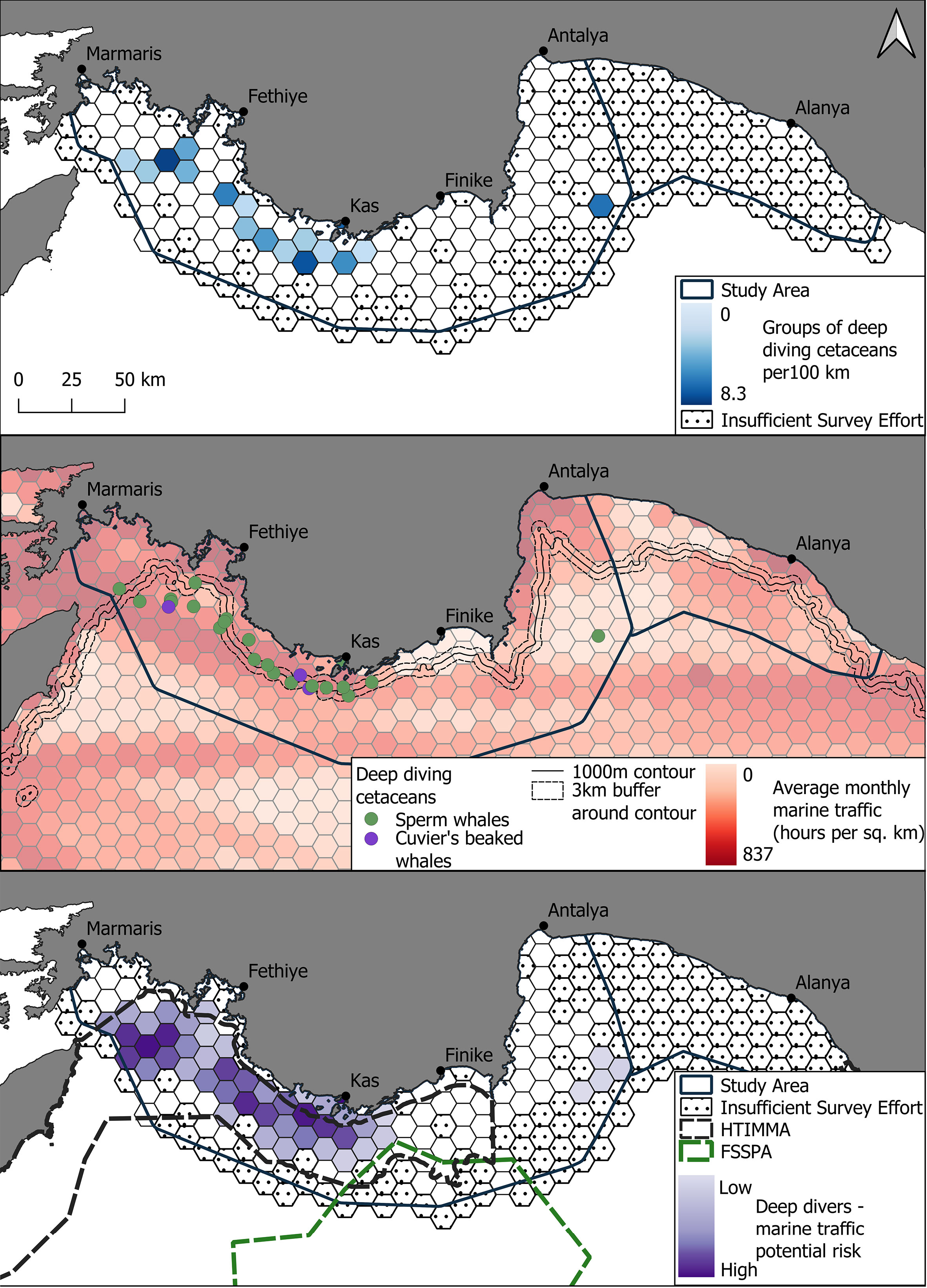
Figure 4 (Upper) Groups of deep diving cetaceans per 100 km in the Eastern Mediterranean Sea with darkening blue representing an increased number of delphinid groups. (Middle) The distribution of different deep diving cetacean species with sperm whales and Cuvier's beaked whales denoted in green and purple circles respectively. Distribution is superimposed onto the average monthly marine traffic (hours per square kilometre) with darkening red signifying higher traffic. Marine vessel information used in this figure was made available by the EMODnet Human Activities project, www.emodnet-humanactivities.eu, funded by the European Commission Directorate General for Maritime Affairs and Fisheries. The 1000m contour is shown with a 3km buffer area around the contour. (Lower) Potential risk areas where the presence of deep diving cetacean and marine traffic were both high during the study period. Darkening purple shows an increased potential for risk.
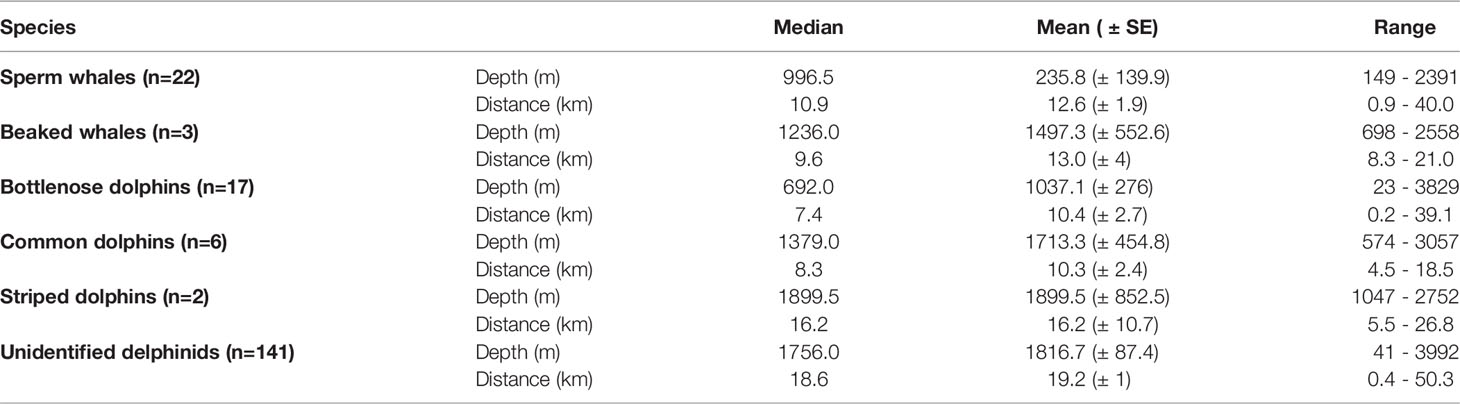
Table 2 Summary of depths and distances from the nearest shore of deep diving cetaceans and delphinid species in the study area.
Delphinids were encountered throughout the entire survey area, from Marmaris to Alanya on 166 occasions (Figures 3, 4; Table 1). Bottlenose dolphins (n = 17) were found in shallower waters than other identified cetaceans, with a median depth of 692m and were also detected the closest to shore with a median distance of 7.4km. Common dolphins (n=6) and striped dolphins (n=2) were found at considerably deeper median depths of 1379m and 1899.5m, respectively. Whilst common dolphins were encountered at a similar median distance from shore to bottlenose dolphins (8.3km), striped dolphins had a median distance from shore of 16.2km. Unidentified delphinids had a median depth of 1756m and a median distance from shore of 18.6km (Figures 3 and band 4; Table 2). In comparison to the deep diving cetaceans, there was far less of a pattern in delphinid encounter rates, with encounter rates spread throughout the survey site. The median encounter rate for delphinids of 2.3 groups per 100 km was x times higher than that of the deep diving cetaceans. The median encounter rate for all cetaceans was 2.7 groups per 100 km and the highest encounter rate of 13.4 groups per 100 km was found in offshore waters between Fethiye and Kaş. There was no clear spatial pattern between the highest overall encounter rates, but 16 of the 20 cells with the highest overall cetacean encounter rate were found in the Hellenic Trench Important Marine Mammal Area. Marine Traffic
Retrieved vessel density showed vessel presence across the entire survey area (Figures 3, 4). The average monthly hours of marine traffic per square kilometer was 0.37 (median absolute deviation (MAD): ± 0.29) with density increasing to more than 300 average monthly vessel hours per square kilometer in four cells near major ports in Marmaris, Antalya and Fethiye (n=2). Traffic was also higher around the coastline in general and a shipping lane running east-west as well as either side of Rhodes. For the entire survey area, coastal cells had a median of 1.03 average monthly vessel hours per square kilometer (MAD: ± 1.50), whilst offshore cells (those not directly connected to the coast or adjacent to the coast with a depth of less than 200m) had a median of 0.32 (MAD: ± 0.21).
In areas where marine protected areas overlapped the study area, the average monthly vessel hours per square kilometer was 0.30 (MAD: ± 0.13) and 11.53 (MAD: ± 14) for the cells in Finike Special Protected Area and Marmaris National Park, respectively. For cells in the Hellenic Trench IMMA, average monthly vessel hours per square kilometer was 0.43 (MAD: ± 0.31) whilst in the Cilician Basin it was 0.26 (MAD: ± 0.21). There were large seasonal differences in marine traffic with summer having the highest monthly vessel hours per square kilometer with 0.65 (MAD: ± 0.34) and winter the lowest with 0.09 (MAD: ± 0.11). This seasonal difference was most evident in coastal cells where summer marine traffic density was more than 13 times that of winter marine traffic (Table 3).
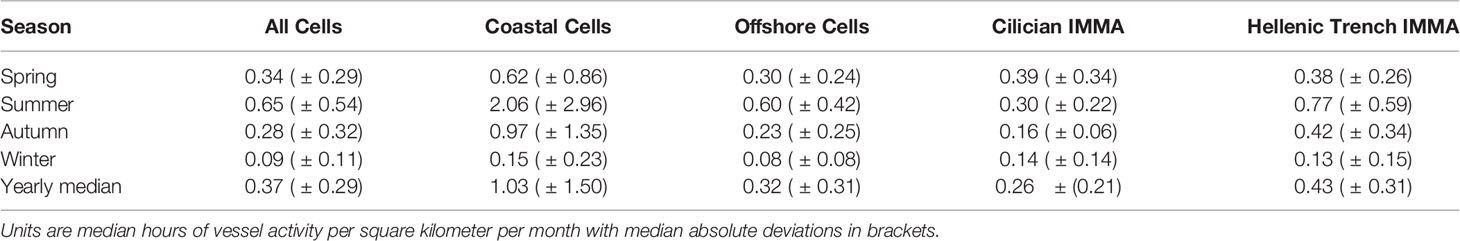
Table 3 Summary of seasonal differences in vessel density for different regions within the survey area.
Sperm whales were detected in cells with a median of 0.98 (MAD: ± 0.58) monthly vessel hours per square kilometer, whilst Cuvier’s beaked whales were found in areas with a median of 0.59 (MAD: ± 0.08). Of the delphinids, bottlenose dolphins were detected in areas that had the highest levels of marine traffic throughout the study period with a median of 0.80 (MAD: ± 0.87) monthly vessel hours per square kilometer. Common dolphins and unidentified delphinids were found in cells with relatively similar levels of marine traffic with median densities of 0.64 (MAD: ± 0.29) and 0.52 (MAD: ± 0.53) whilst striped dolphins were found in considerably less dense areas of 0.03 (MAD: ± 0.02) monthly vessel hours per square kilometer.
Potential Risk Areas
The highest potential for risk for deep divers were in the channel of water that runs between Turkey and Rhodes and in cells that surround the 1000m contour in Marmaris Bay to the West of Antalya Bay. The majority of the potential high-risk cells overlapped with the shipping lane identified during the marine traffic density analysis and also fell within the Hellenic Trench Important Marine Mammal Area. Cells further to the east had comparatively lower potential risk scores (Figure 3). Delphinids followed a similar patter with higher risk cells generally in the west of the survey area, along the same shipping lane. In addition, potential risk for delphinids was high around ports (Marmaris, Fethiye and Kaş). The potential risk index scores in Antalya Bay were generally much lower than in the west of the study area (Figure 4).
Critical Habitats
For deep diving cetaceans two critical habitats were delineated, the first in the waters between Marmaris and Fethiye, and the second in the waters off Kaş. The two critical habitats were roughly even in size (511.1 km2 and 664.3 km2 respectively). A single large critical habitat was identified spanning the waters from Rhodes Basin to the Piri Reis Sea Mountains (2516.7 km2) (Figure 5). Over 99% (1170.1 km2) of the deep diving cetacean critical habitat overlapped with the area identified as a critical habitat for delphinids. Nearly 85% (996.2km2) and 92% (2305.7km2) of deep diving cetacean and delphinid critical habitat overlapped with the Hellenic Trench IMMA respectively. None of the critical habitats overlapped with any existing protected areas (Figure 5).
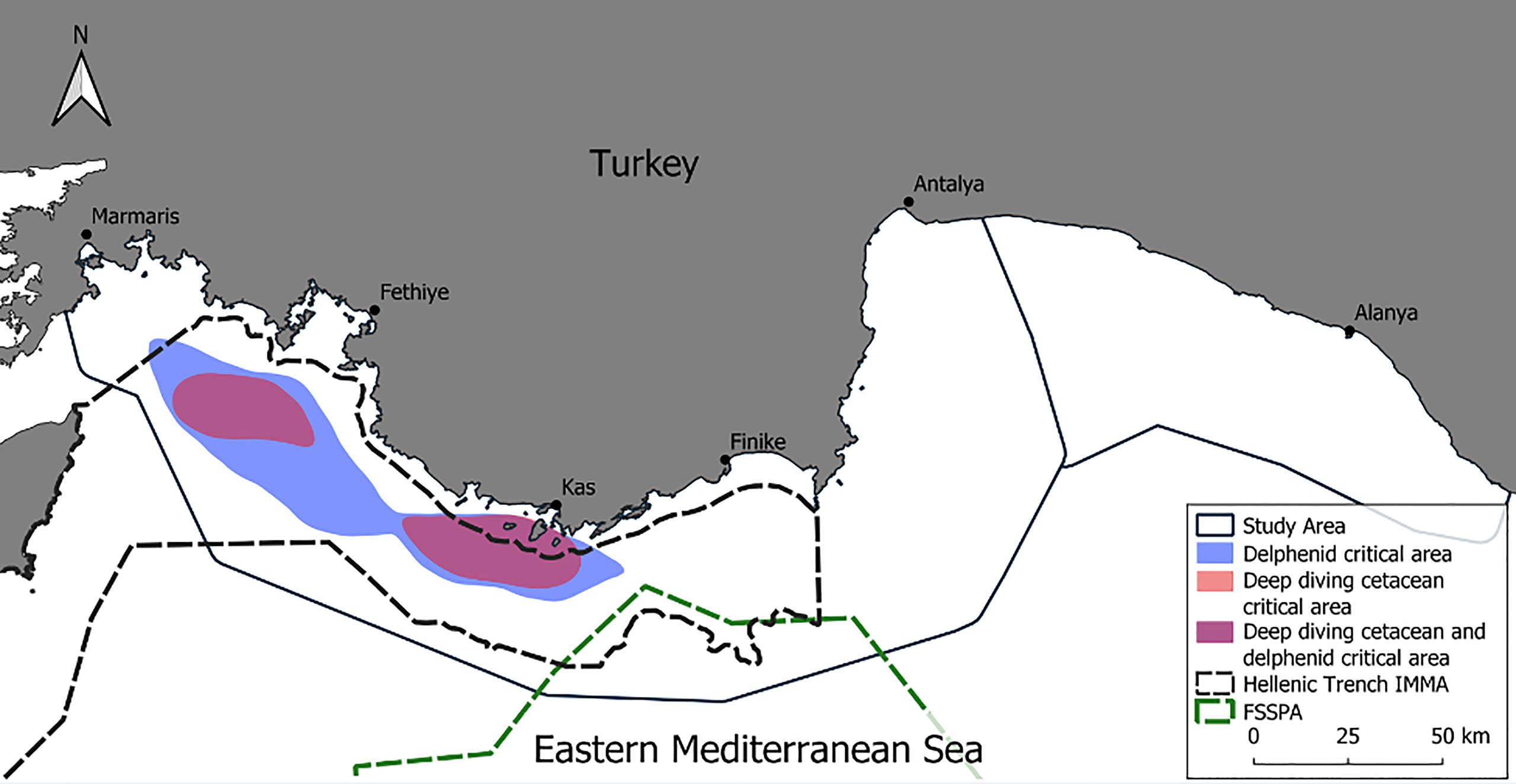
Figure 5 Areas of the Eastern Mediterranean Sea surrounding Turkey deemed critical for delphinids (blue) and deep diving cetaceans (pink). Note that both critical areas for deep diving cetaceans fell within the critical area for delphinids and thus are shown in a purple colour.
Discussion
The current study is the longest ongoing cetacean research project, spanning all seasons, in the Turkish Eastern Mediterranean Sea. This study builds off the preliminary results of Akkaya et al. (2020) to include two complete years of data collection. Of the seven cetacean species regularly found in Turkish waters (Öztürk et al., 2016), five were detected during this study, although bottlenose dolphins were the only species to be seen throughout all seasons (Table 1).
Distribution and Encounter Rates
As expected, the different species had varying distribution, albeit with considerable spatial overlap with each other and maritime traffic (Figures 3, 4). Deep diving species were encountered mainly in the west, with 23 out of 25 (92%) of the sightings occurring in the bathymetric irregularities that stretch east from the Rhodes basin to the end of the Finike Basin (Figure 3). Bathymetry has been shown to be important in relation to the distribution of the deep diving species (sperm whales and beaked whales) (Frantzis et al., 2003; Frantzis et al., 2019). The Hellenic Trench is considered by Podestà et al. (2016) to be the largest “high-density areas of occurrence” in the Mediterranean for Cuvier’s beaked whale as well as a core habitat with year-round presence of the eastern Mediterranean sperm whale population (Frantzis et al., 2011; Frantzis et al., 2019). (Frantzis. et al.’s 2019) study along the Hellenic Trench recorded 74% of the detected sperm whales to be within 3km of the 1000m contour with density decreasing as water depth gets shallower or deeper. The current study found that 60% of the deep diving species were recorded within 3km of the 1000m contour with a median respective depth of 999m, although individuals also occurred in much shallower (149m) and much deeper waters (2391m). Furthermore, in the Greek areas of the Hellenic Trench, Cuvier’s beaked whales have also shown a preference for depths of 500 to 1500m depth as well as any steep sloping bathymetry (Frantzis et al., 2003). The median value of 1236m found in this study aligned well with the findings in adjoining waters, although it should be noted that this is for just three recorded individuals, one of which was considerably deeper (2558m). Due to the close proximity and overlap of the Hellenic Trench to the current study area and similarities in depth parameters, it is more than likely that the home range of the sperm whales and Cuvier’s beaked whales extends across the waters surrounding both Greece and Turkey (Akkaya et al., 2020).
Delphinids were distributed throughout the entire survey area from Rhodes Basin to Antalya Basin, ranging from coastal waters to 50.3km offshore. In the Mediterranean, bottlenose dolphins are generally found in shallow waters along the continental shelf and in productive waters up to 600m depth (Bearzi, 2003; de Stephanis et al., 2008). However, the current study reported a median depth preference of 692m with detections of the species occurring up to a depth of 3,829m. Therefore, it is likely that as well as the commonly occurring coastal population, the Eastern Mediterranean Sea may also hold an offshore distribution of the bottlenose dolphins. Further research is needed to understand the reasons behind these offshore sightings as they may indicate that the home range of the coastal population extends to the deeper waters, that the species has separate offshore populations or that the species may show long-distance movement patterns, none of which have previously been reported in the Mediterranean Sea. Additionally, the majority of the delphinids which could not be identified to species level, were detected with a median depth of 1756m. Some delphinids depend on certain cephalopod species and abundance of these species needs to be investigated. Öztürk et al. (2007) examined the cephalopod remains from the stomachs of three striped dolphins and two Risso’s dolphins (Grampus griseus) taken as bycatch in the swordfish industry in the eastern Mediterranean Sea off the Turkish coast. In total, 478 lower beaks were identified as belonging to 14 cephalopod species, some of which are only found in deep seas. This stresses the importance of not just coastal waters, but also deep-sea ecosystems for delphinids within the Eastern Mediterranean Sea of Turkey.
In terms of seasonal distribution, sperm whales were mostly detected in spring and summer months with only one individual identified in autumn and none in winter (Table 1). The additional two seasons (autumn and winter) of data here, were further confirmation of (Akkaya et al.’s 2020) findings of seasonal presence of sperm whales in Turkish waters and corresponds with previous works in Turkey (Ozturk et al., 2013) and in Greece (Diogou et al., 2019) which both found higher numbers of sperm whales in spring and summer. When coupled with the fact that social units have been observed with calves in this area (Akkaya et al., 2020), it suggests that this area of the Eastern Mediterranean Sea may be an important breeding or nursing ground in warmer seasons. Although limited data meant that it was not possible to perform a potential risk index analysis for each season, the trends for increased marine traffic found in summer are concerning.
In contrast to Baş et al. (2016), who detected bottlenose dolphins, in just spring and summer in the coastal water of the Gulf of Antalya, this study detected bottlenose dolphins across all seasons. Common dolphins were only seen in summer months, however, this is possibly a result of limited data collection as they have been recorded year-round in the adjoining Greek (Milani et al., 2019) and Turkish (Akkaya et al., in review) areas of the Aegean Sea. There were not enough beaked whale or striped dolphin detections recorded to analyze their seasonal distribution but filling this important data gap should be considered a priority in the future.
The median encounter rate of cetaceans was 2.7 groups per 100km (2.3 groups and 0 groups per 100km for delphinids and deep diving cetaceans respectively) in the study area. This is much higher than previous recordings in the region with cetacean encounter rates of 0.68 (Boisseau et al., 2010) and 0.008 per 100km (Ryan et al., 2014). This is likely due to these studies covering different survey areas to this study and the fact that they consisted of single summer survey efforts, highlighting the importance of multi-seasonal, multi-year, local efforts (Akkaya et al., 2020).
Marine Traffic and Potential Risk
The Mediterranean is one of the world’s busiest waterways (Notarbartolo di Sciara and Birkun Jr., 2010), and thus it is important to understand the magnitude of overlap between cetaceans and marine traffic. This study highlighted the overlap in spatial usage of both the delphinids and the deep diving species with areas of heavy maritime traffic (Figure 3). The highest levels of marine traffic were along the coast, however, there was a notable presence of a shipping lane in the west of the study area along the 1000m contour of the Finike Canyon, presumably due to it being a safe depth for large boats (Frantzis et al., 2019). Whilst the median monthly hours of vessel traffic/km2 was much lower in the Finike Seamount Marine Protected Area than the rest of the core study area (0.3 as opposed to 0.37), this is likely due to the Finike Seamounts being further offshore rather than any specific protective measures. What is more concerning, is that the monthly median hours of vessel traffic/km2 within the Hellenic Trench IMMA was 0.43. When compared to the median for the entire survey area, this does not seem too high (~16% higher) but compared to other non-coastal cells (those cells not directly connected to land and with a centroid depth greater than 200m), it is a worrying 34% higher. The Hellenic Trench IMMA was ranked in the top ten most at risk of ship strike IMMAs of 114 IMMAs assessed globally (WWF-IUCN-IWC-OceanMind, 2019). The marine traffic density results in this study confirm the potential for increased ship strike. Thus, unsurprisingly, a high proportion of the cells considered to have a high potential risk index score were found within the Hellenic Trench IMMA. The western part of this IMMA has previously been proposed for a MPA (Agardy et al., 2007). The current study has now demonstrated the importance of the extension of Hellenic Trench and Anadolu Seamounts, and future protective measures must span across the international border between Greece and Turkey.
When considering the overlap between cetaceans and maritime traffic, sperm whales followed by bottlenose dolphins were detected in areas with the highest density of marine traffic. Sperm whales (Frantzis et al., 2019) and Cuvier’s beaked whales (Carrillo and Ritter, 2010) are known to be particularly vulnerable to shipping collisions (Panigada et al., 2006). The work of Frantzis et al. (2019) indicated that as a result of the major shipping route along the Hellenic Trench there is increased mortality rates of the sperm whale population due to ship strikes in this area. As there is only a low recruitment rate of sperm whales (~2.5/year) in the Eastern Mediterranean, even a small number of ship strikes would likely have population level effects (Frantzis et al., 2019). It has been found, however, that small changes in shipping routes could dramatically reduce risk (Frantzis et al., 2019). By rerouting or reducing vessel speed within these areas, the collision risk and noise pollution for sperm whales and beaked whales could be considerably reduced with minimal inconvenience for the shipping industry (Vanderlaan and Taggart, 2009; Frantzis et al., 2019). While ship strikes may be less of a direct threat to the bottlenose dolphins’ population, increased noise and pollution levels may come with direct and indirect consequences to the dolphin populations such as habitat shifts and behavioral alterations (Papale et al., 2012; Akkaya Bas et al., 2017).
Marine traffic considered within this study were retrieved from vessels using AIS which is only mandatory for ships with 300 or greater gross tonnage (International Maritime Organisation, 2021). According to the FAO, 82.9% of the fishing within the Mediterranean and Black Sea is done from small scale vessels (FAO, 2020), and this along with other small recreational boats suggest that the actual marine traffic is a lot higher than seen here, with smaller boats more likely to have a predominantly coastal distribution. The impact of small boats compared to large freight ships may vary and so it would be beneficial to have further studies looking into the combined impacts.
As well as the threat of increasing marine traffic, cetaceans are also subject to other anthropogenic disturbances. These include unsustainable fishery practices, sonar use, naval exercises and hydrocarbon exploration (Notarbartolo di Sciara and Birkun Jr, 2010; Fylaktos and Papanicolas, 2019). The latter is of particular concern with a recent rise in the number of oil and gas explorations taking place within the Eastern Mediterranean Sea. The year-round distribution and habitat use of cetaceans within Turkish waters of the Mediterranean encourages a reassessment of current levels of protections of the region to counteract excessive anthropogenic impacts.
Recommendations
The current study has delineated two habitats for deep diving cetaceans and one for delphinids that the authors deem critical where the area usage of cetaceans and marine traffic overlaps (Figure 5). The first deep diving cetacean is between Marmaris and Fethiye, the second between the Fethiye and Kaş and the delphinid critical habitat effectively encompasses the first two. These two deep diving habitats are different in terms of nutrient availability, which is important for primary production, thus consequently affecting the top predators. The cyclonic Rhodes Gyre causes nutrient enrichment, concentration of larval food distribution and local retention of eggs and larvae (Agostini and Bakun, 2002). In contrast to this, the Levantine Basin is oligotrophic, which results in extremely low values of phytoplankton abundance (Herut et al., 2017). Therefore, a research priority is to investigate the relation between oceanographic conditions and cetacean presence, especially within the Finike Basin.
Well-managed MPAs have been found to be a key tool for conservation through the regulation of anthropogenic activities in biologically valuable areas and can simultaneously be used as study sites for scientists (Edgar et al., 2007). According to Woodside et al. (2006) and Öztürk et al. (2013), the Finike (Anaximander) Seamount MPA is an important area for deep diving mammal species like sperm whales and Cuvier’s beaked whales. Whilst the current study did not detect these deep-diving species and no potential risk areas were identified within the MPA, small cetaceans were both acoustically and visually detected in this area. The lack of detections of deep diving cetaceans may be a result of very limited vessel coverage in this area and efforts should be made in the future to study these unique deep-sea habitats.
Another future research priority is the investigation of seasonal variation in the overlap between cetaceans and marine traffic as well as studying the magnitude of the impact of marine traffic on different cetacean species, particularly with the variation in seasonal marine traffic identified. Efforts have recently been made to study the Turkish waters to the east of the current study area to assess the species range and densities from west to east (Akkaya, 2021) and incorporate species like fin whales which are sighted only in the eastern part of Turkish waters (Stephens et al., 2021). Despite this, there are still few data available on potential risk areas where cetaceans are exposed to high shipping densities in the Eastern basin of the Mediterranean Sea. Dedicated long-term survey efforts prove essential to provide information on population, density patterns, and movement of cetaceans throughout the entire year and therefore reveal important cetacean habitats along the Turkish Mediterranean coast (Akkaya et al., 2020). In the meantime, this study has clearly shown the importance of the Eastern Mediterranean Sea for the cetaceans. Despite Turkey becoming a member of the Agreement on the Conservation of the Cetaceans of the Black Sea, Mediterranean Sea and contiguous Atlantic Area (ACCOBAMS), which commits the country to protect cetaceans from unregulated and uncontrolled anthropogenic activities, a lack of scientific knowledge and threat assessment persists resulting in unsustainable practices continuing within these waters (Akkaya et al., 2020). Several important international agreements including the Convention on Biological Diversity, which Turkey is a party to, require a precautionary approach to be taken and clearly there is enough evidence of the distribution of a range of cetaceans throughout this area, as well as evidence of potential risks to their prolonged survival.
Despite this study filling a considerable data gap, there is still a great dearth of information within Turkish waters. The findings of this work indicate a need for further study on cetacean response to different seasons and oceanographic variables to allow more advanced risk analyses to be performed as well as investigation into the magnitude of impact of the marine traffic on different species. It is imperative that Species Conservation Action Plans are created as soon as possible for each of these species based on current information and updated as data gaps are filled. This will allow relevant management measures such as rerouting marine traffic or creating marine protected areas to minimize exposure of cetaceans to anthropogenic threats. For highly migratory species such as cetaceans, transboundary cooperation is also essential for the sake of better conservation in this understudied area.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by Turkish Ministry of Agriculture and Forestry.
Author Contributions
TA, AAk, and PL participated in the field survey. Analysis was conducted by TA, AAk, GH, MN, and DK and the manuscript was written by all the authors. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Rufford Small Grant Foundation (Grant 27028-B), the MAVA Foundation (Contract SPA/RAC N°06/2019) and WWF-Turkey. We are grateful to the Turkish Ministry of Environment and Climate Change and Turkish Marine Research Foundation for their financial support.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to thank the Rufford Small Grant Foundation, the MAVA Foundation and WWF-Turkey for their invaluable funding contributions which allowed this research to take place. We would also like to thank Jonathan Gordon of the University of St Andrews for his continued support of the project. Finally, we would like to extend our gratitude to all of the Turkish and international researchers that helped to collect this data.
References
Abdulla A., Linden O. (Eds.) (2008). Maritime Traffic Effects on Biodiversity in the Mediterranean Sea: Review of Impacts, Priority Areas and Mitigation Measures (Malaga, Spain: IUCN Centre for Mediterranean Cooperation), 184 pp.
Agardy T., Aguilar N., Cañadas A., Engel M., Frantzis A., Hatch L., et al. (2007). “A Global Scientific Workshop on Spatio-Temporal Management of Noise,” in Report of the Scientific Workshop, 20–21. Puerto Calero, Lanzarote, Leviathan Sciences
Agostini V. N., Bakun A. (2002). ‘Ocean Triads’ İn the Mediterranean Sea: Physical Mechanisms Potentially Structuring Reproductive Habitat Suitability (With Example Application to European Anchovy (Engraulis Encrasicolus). Fish. Oceanog. 11 (3), 129–142. doi: 10.1046/j.1365-2419.2002.00201.x
Aguilar A., Gaspari S. (2012). Stenella Coeruleoalba (Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie. 2012, e.T16674437A16674052. doi: 10.2305/IUCN.UK.2012-1.RLTS.T16674437A16674052.en
Aguilar Soto N., Johnson M., Madsen P. T., Tyack P. L., Bocconcelli A., Borsani J. F. (2006). Does Intense Ship Noise Disrupt Foraging in Deep-Diving Cuvier’s Beaked Whales (Ziphius Cavirostris)? Mar. Mam. Sci. 22 (3), 690–699.
Airoldi S., Azzellino A., David L., Dhermain F., Di-Meglio N., Jimenez V., et al (2015). Final Report “Absolute Abundance of Risso's Dolphin (Grampus Griseus) in the North-Western Part of the Pelagos Sanctuary. Pelagos. Sanctuary. Convent. No. 03/2014 59 Istituto Tethys /EcoOcean Institut/GECEM/Fondazione CIMA/GIS3M. Available online at: https://www.gis3m.org/sites/gis3m.org/files/doc/final_report_grampus_pelagos.pdf
Akkaya A. (2021). “Investigating the Impact of Anthropogenic Activities on Cetaceans Within the Eastern Mediterranean Sea of Turkey,” in 5th Conference on the Conservation of Cetaceans in South Mediterranean Countries, Lebanon, 15th April.
Akkaya A., Awbery T., Lyne P., Schartmann H., Nease K., Hass C., et al. (In review) Short-Beaked Common Dolphins of Dilek Peninsula National Park, South Aegean Sea: Preliminary Results on Group Structure, Spatial Distribution, BehaviouralPatterns, Whistle Characteristics and its Behavioural Context.
Akkaya Baş A., Lagoa J., Atchoi E. (2016). New Records of Cuvier’s Beaked Whales (Ziphius Cavirostris) From the Turkish Levantine Sea. Turkish. J. Zool. 40 (3), 454–460. doi: 10.3906/zoo-1509-19
Akkaya Bas A., Christiansen F., Amaha Öztürk A., Öztürk, B. and McIntosh C. (2017). The Effects of Marine Traffic on the Behaviour of Black Sea Harbour Porpoises (Phocoena Phocoena Relicta) Within the Istanbul Strait, Turkey. PloS One 12 (3), e0172970. doi: 10.1371/journal.pone.0172970
Akkaya A., Lyne P., Schulz X., Awbery T., Capitain S., Rosell B. F., et al. (2020). Preliminary Results of Cetacean Sightings in the Eastern Mediterranean Sea of Turkey. J. Black. Sea/Mediterranean. Environ. 26 (1). https://blackmeditjournal.org/volumes-archive/vol-26-2020/vol-26-2020-no-1/5329-2/
Azzellino A., Airoldi S., Gaspari S., Lanfredi C., Moulins A., Podestà M., et al. (2016). “Risso's Dolphin, Grampus Griseus, in the Western Ligurian Sea: Trends in Population Size and Habitat Use,” in Advances in Marine Biology 75, 205–232. Eds. Notarbartolo Di Sciara G., Podestà M., Curry B. E. (Academic Press), 205–223.
Baş A. A., Erdoğan M. A., Morris N. R. C., Yeoman K., Humphrey O., Gaggioli E., et al. (2016). Seasonal Encounter Rates and Residency Patterns of an Unstudied Population of Bottlenose Dolphin (Tursiops Truncatus) in the Northwestern Levantine Sea, Turkey. Hyla.: Herpetol. Bull. 2016 (1), 1–13. https://hrcak.srce.hr/file/260618
Bearzi G., Reeves R.R., Notarbartolo Di Sciara G., Politi E., Cañadas A., Frantzis A., et al (2003). Delphinus Delphis (Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie. 2003, e.T41762A10557372. doi: 10.2305/IUCN.UK.2003.RLTS.T41762A10557372.en
Bearzi G., Politi E., Agazzi S., Azzellino A. (2006). Prey depletion caused by overfishing and the decline of marine megafauna in eastern Ionian Sea coastal waters (central Mediterranean). Biological Conservation, 127 (4), 373–382. doi: 10.1016/j.biocon.2005.08.017
Bearzi G., Fortuna C., Reeves R. (2012). Tursiops truncatus. The IUCN Red List of Threatened Species 2012: e.T22563A2782611. Accessed on 27 May 2022. Available online at: https://www.iucnredlist.org/species/22563/2782611
Bearzi G., Genov T., Natoli A., Gonzalvo J., Pierce G. J. (2021). Delphinus Delphis (Inner Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie. 2021, e.T189865869A189865884. doi: 10.2305/IUCN.UK.2021-3.RLTS.T189865869A189865884.en
Bearzi G., Reeves R. R., Notarbartolo Di Sciara G., Politi E., Cañadas A., et al. (2003). Ecology, Status and Conservation of Short-Beaked Common Dolphins, Delphinus Delphis in the Mediterranean Sea. Mam. Rev. 33 (3-4), 224–252. doi: 10.1046/j.1365-2907.2003.00032.x
Bejder L., Samuels A., Whitehead H., Gales N. (2006b). Interpreting Short-Term Behavioural Responses to Disturbance Within a Longitudinal Perspective. Anim. Behav. 72 (5), 1149–1158. doi: 10.1016/j.anbehav.2006.04.003
Bejder L., Samuels A., Whitehead H., Gales N., Mann J., Connor R., et al. (2006a). Decline in Relative Abundance of Bottlenose Dolphins Exposed to Long-Term Disturbance. Conserv. Biol. 20 (6), 1791–1798. doi: 10.1111/j.1523-1739.2006.00540.x
Birch C. P., Oom S. P., Beecham J. A. (2007). Rectangular and Hexagonal Grids Used for Observation, Experiment and Simulation in Ecology. Ecol. Model. 206 (3-4), 347–359. doi: 10.1016/j.ecolmodel.2007.03.041
Blondin H., Abrahms B., Crowder L. B., Hazen E. L. (2020). Combining High Temporal Resolution Whale Distribution and Vessel Tracking Data Improves Estimates of Ship Strike Risk. Biol. Conserv. 250, 108757. doi: 10.1016/j.biocon.2020.108757
Boisseau O., Lacey C., Lewis T., Moscrop A., Danbolt M., McLanaghan R., et al. (2010). Encounter Rates of Cetaceans in the Mediterranean Sea and Contiguous Atlantic Area. J. Mar. Biol. Assoc. Unite. Kingdom. 90 (8), 1589–1599. doi: 10.1017/S0025315410000342
Campana I., Crosti R., Angeletti D., Carosso L., David L., Di-Méglio N., et al. (2015). Cetacean Response to Summer Maritime Traffic in the Western Mediterranean Sea. Mar. Environ. Res. 109, 1–8. doi: 10.1016/j.marenvres.2015.05.009
Cañadas A., Notarbartolo di Sciara G. (2018). “Ziphius Cavirostris (Mediterranean Subpopulation),” in The IUCN Red List of Threatened Species 2018.
Carrillo M., Ritter F. (2010). Increasing Numbers of Ship Strikes in the Canary Islands: Proposals for Immediate Action to Reduce Risk of Vessel-Whale Collisions. J. Cetacean. Res. Manage. 11 (2), 131–138.
Dede A., Saad A., Fakhri M., Öztürk B. (2012). Cetacean Sightings in the Eastern Mediterranean Sea During the Cruise in Summer 2008. J. Black. Sea/Mediterranean. Environ. 18 (1), 49–57.
De Stephanis R., Cornulier T., Verborgh P., Salazar Sierra J., Pérez Gimeno N., Guinet C. (2008). Summer Spatial Distribution of Cetaceans in the Strait of Gibraltar in Relation to the Oceanographic Context. Mar. Ecol. Prog. Ser. 353, 275–288. doi: 10.3354/meps07164
Dinis A., Carvalho A., Alves F., Nicolau C., Ribeiro C., Kaufmann M., et al. (2016). “Spatial and Temporal Distribution of Bottlenose Dolphins, Tursiops Truncatus, in the Madeira Archipelago, NE Atlantic,” in Arquipélago-Life and Marine Sciences, vol. 33, pp. 45–pp. 54.
Diogou N., Klinck H., Frantzis A., Nystuen J. A., Papathanassiou E., Katsanevakis S. (2019). Year-Round Acoustic Presence of Sperm Whales (Physeter Macrocephalus) and Baseline Ambient Ocean Sound Levels in the Greek Seas. Mediterranean. Mar. Sci. 20 (1), 208–221. doi: 10.12681/mms.18769
Drouot V., Bérubé M., Gannier A., Goold J. C., Reid R. D., Palsbøll P., et al. (2004). A Note on Genetic Isolation of Mediterranean Sperm Whales (Physeter Macrocephalus) Suggested by Mitochondrial DNA. J. Cetacean. Res. Manage. 6 (1), 29–32. https://www.cetaces.org/wp-content/uploads/2009/07/Drouot04_gen-jcrm.pdf
Edgar G. J., Russ G. R., Babcock R. C. (2007). “Marine Protected Areas”. in Mar. Ecol. eds Connell S. D., Gillanders B. M. (Oxford: Oxford University Press) 27, 533–555.
Emodnet-humanactivities.eu (2019) EU Vessel Density Map. Detailed Method. Available at: https://www.emodnet-humanactivities.eu/documents/Vessel%20density%20maps_method_v1.5.pdf (Accessed 1 March 2022).
Engelhaupt D., Hoelzel R. A., Nicholson C., Frantzis A., Mesnick S., Gero S, et al. (2009). Female Philopatry in Coastal Basins and Male Dispersion Across the North Atlantic in a Highly Mobile Marine Species, the Sperm Whale (Physeter Macrocephalus). Mol. Ecol. 18 (20), 4193–4205. doi: 10.1111/j.1365-294X.2009.04355.x
FAO (2020). The State of Mediterranean and Black Sea Fisheries 2020 (Rome: General Fisheries Commission for the Mediterranean). doi: 10.4060/cb2429en Last updated 10/03/2021
Frantzis A. (2004). The First Mass Stranding That was Associated With the Use of Active Sonar (Kyparissiakos Gulf, Greece 1996). ECS. Newslett. 42 (Special Issue), 14–20. https://www.europeancetaceansociety.eu/system/files/ECS%20Special%20Publication%20Series/ECS2004_News42_Active_Sonar_Workshop.pdf#page=18
Frantzis A., Airoldi S., Notarbartolo-di-Sciara G., Johnson C., Mazzariol S. (2011). Inter-Basin Movements of Mediterranean Sperm Whales Provide Insight Into Their Population Structure and Conservation. Deep. Sea. Res. Part I.: Oceanog. Res. Pap. 58 (4), 454–459. doi: 10.1016/j.dsr.2011.02.005
Frantzis A., Alexiadou P., Gkikopoulou K. (2014). Sperm Whale Occurrence, Site Fidelity and Population Structure Along the Hellenic Trench (Greece, Mediterranean Sea). Aquat. Conservat.: Mar. Freshwat. Ecosyst. 24 (1), 83–102. doi: 10.1002/aqc.2435
Frantzis A., Alexiadou P., Paximadis G., Politi E., Gannier A., et al. (2003). Current Knowledge of the Cetacean Fauna of the Greek Seas. J. Cetacean. Res. Manage. 5 (3), 219–232. http://www.pelagosinstitute.gr/gr/pelagos/pdfs/Greek%20Cetacean%20Fauna.pdf
Frantzis A., Leaper R., Alexiadou P., Prospathopoulos A., Lekkas D. (2019). Shipping Routes Through Core Habitat of Endangered Sperm Whales Along the Hellenic Trench, Greece: Can We Reduce Collision Risks? PloS One 14 (2), e0212016. doi: 10.1371/journal.pone.0212016
Fylaktos N., Papanicolas C. N. (2019)New Technologies for Eastern Mediterranean Offshore Gas Exploration. In: European Parliamentary Research Services. Available at: http://www.europarl.europa.eu/RegData/etudes/STUD/2019/634419/EPRS_STU(2019)634419_EN.pdf (Accessed 01 November 2020).
Güçlüsoy H., Karauz E. S., Kirac C. O., Bilecenoğlu M. (2014). Checklist of Marine Tetrapods (Reptiles, Seabirds, and Mammals) of Turkey. Turkish. J. Zool. 38 (6), 930–938. doi: 10.3906/zoo-1405-81
Herut B., Segal Y., Gertner Y. (2018). The National Monitoring Program of Israel’s Mediterranean Waters—Scientific Report on Marine Pollution for 2017, Israel Oceanographic and Limnological Research; IOLR Report H50/2018; IOLR (Israel Oceanographic and Limnological Research): Haifa, Israel pp.10–102.
Heyning J. E. (1989). “Cuvier’s Beaked Whale Ziphius Cavirostris G. Cuvier,” in Handbook of Marine Mammals Vol. 4. River Dolphins and the Larger Toothed Whales. Eds. Ridgway S. H., Harrison R. J. (London and San Diego: Academic Press), 289–308.
International Maritime Organization (2021) AIS Transponders. Available at: https://www.imo.org/en/OurWork/Safety/Pages/AIS.aspx (Accessed 25/04/2021).
IUCN-MMPATF (2017a)Hellenic Trench IMMA Factsheet. In: IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. Available at: https://www.marinemammalhabitat.org/portfolio-item/hellenic-trench/ (Accessed 27/04/2021).
IUCN-MMPATF (2017b)Cilician Basin IMMA Factsheet. In: IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. Available at: https://www.marinemammalhabitat.org/portfolio-item/hellenic-trench/ (Accessed 03/03/2021).
Jahoda M., Lafortuna C. L., Biassoni N., Almirante C., Azzellino A., Panigada S., et al. (2003). Mediterranean Fin Whales (Balaenoptera Physalus) Response to Small Vessels and Biopsy Sampling Assessed Through Passive Tracking and Timing of Respiration. Mar. Mam. Sci. 19 (1), 96–110. doi: 10.1111/j.1748-7692.2003.tb01095.x
Kerem D., Hadar N., Goffman O., Scheinin A., Kent R., Boisseau O., et al. (2012). Update on the Cetacean Fauna of the Mediterranean Levantine Basin. Open Mar. Biol. J. 6 (1), 6–27. doi: 10.2174/1874450801206010006
Laist D. W., Knowlton A. R., Mead J. G., Collet A. S., Podesta M. (2001). Collisions Between Ships and Whales. Mar. Mam. Sci. 17 (1), 35–75. doi: 10.1111/j.1748-7692.2001.tb00980.x
Lanfredi C., Arcangeli A., David L., Holcer D., Rosso M., Natoli A. (2021). Grampus Griseus (Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie., e.T16378423A190737150. doi: 10.2305/IUCN.UK.2021-3.RLTS.T16378423A190737150.en
Lewis T., Boisseau O., Danbolt M., Gillespie D., Lacey C., Leaper R., et al. (2018). Abundance Estimates for Sperm Whales in the Mediterranean Sea From Acoustic Line-Transect Surveys. J. Cetacean. Res. Manage. 18, 103–117. https://view.officeapps.live.com/op/view.aspx?src=https%3A%2F%2Fore.exeter.ac.uk%2Frepository%2Fbitstream%2Fhandle%2F10871%2F27538%2F139-Abundance%2520estimates%2520for%2520sperm%2520whales%2520in%2520the%2520Med-Edited%2520Manuscript.doc%3Fsequence%3D1%26isAllowed%3Dy&wdOrigin=BROWSELINK
Milani C., Vella A., Vidoris P., Christidis A., Koutrakis E. (2019). Abundance, Distribution and Diet of the Common Dolphin, Delphinus Delphis, in the Northern Aegean Sea (Greece). Aquat. Conservat.: Mar. Freshwat. Ecosyst. 31, 76–86. doi: 10.1002/aqc.3081
Notarbartolo di Sciara G., Birkun A. Jr. (2010). Conserving Whales, Dolphins and Porpoises in the Mediterranean and Black Seas: An ACCOBAMS Status Report (Monaco: ACCOBAMS), 212.
Notarbartolo di Sciara G., Frantzis A., Bearzi G., Reeves R. R. (2006). “Sperm Whale Physeter Macrocephalus (Mediterranean Subpopulation),” in The Status and Distribution of Cetaceans in the Black Sea and Mediterranean Sea. Eds. Reeves R. R., Notarbartolo di Sciara G. (Malaga, Spain:IUCN Centre for Mediterranean Cooperation, Workshop report), 45–56.
Notarbartolo di Sciara G., Frantzis A., Bearzi G., Reeves R. (2012). Physeter Macrocephalus Mediterranean Subpopulation. In: The IUCN Red List of Threatened Species 2012. Available at: https://ww.iucnredlist.org/species/16370739/16370477 (Accessed 26 April 2021).
Notarbatolo di Sciara G., Hoyt E. (2020). Healing the Wounds of Marine Mammals by Protecting Their Habitat. Ethics. Sci. Environ. Politics. 20, 15–23. doi: 10.3354/esep00190
Öztürk A., Ayhan D., Tonay A. (2016). “Cetaceans in the Turkish Waters of the Mediterranean Sea,” in The Turkish Part of the Mediterranean Sea Marine Biodiversity, Fisheries, Conservation and Governance. Eds. Turan C., Salihoglu B., Özbek E. O., Öztürk B. (Turkish Marine Research Foundation), 566–571.
Öztürk B., Salman A., Ozturk A. A., Tonay A. (2007). Cephalopod Remains in the Diet of Striped Dolphins (Stenella Coeruleoalba) and Risso's Dolphins (Grampus Griseus) in the Eastern Mediterranean Sea. Vie. Milieu. 57, 57–63.
Öztürk A., Tonay A., Ayhan D. (2013). Sperm Whale (Physeter Macrocephalus) Sightings in the Aegean and Mediterranean Part of Turkish Waters. J. Black. Sea/Mediterranean. Environ. 19 (2), 169?177.
Öztürk A. A., Tonay A. M., Dede A. (2011). Strandings of the Beaked Whales, Risso’s Dolphins, and a Minke Whale on the Turkish Coast of the Eastern Mediterranean Sea. J. Black. Sea/Mediterr. Environ. 17, 269–274.
Panigada S., Azzellino A., Cubaynes H., Folegot T., Fretwell P., Jacob T., et al. (2020). Proposal to Develop and Evaluate Mitigation Strategies to Reduce the Risk of Ship Strikes to Fin and Sperm Whales in the Pelagos Sanctuary - Final Report (Pelagos Secretariat - Convention), 67 pp. Available at: https://www.sanctuaire-pelagos.org/it/tous-les-telechargements/etudes-scientifiques-studi-scientifici-studies/studi-italiani/1657-2018-04/file
Panigada S., Notarbartolo di Sciara G. (2012). Balaenoptera Physalus (Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie. 2012., e.T16208224A17549588. doi: 10.2305/IUCN.UK.2012.RLTS.T16208224A17549588.en
Panigada S., Pesante G., Zanardelli M., Capoulade F., Gannier A., Weinrich M.T., et al. (2006). Mediterranean Fin Whales at Risk From Fatal Ship Strikes. Mar. Pollut. Bull. 52 (10), 1287–1298. doi: 10.1016/j.marpolbul.2006.03.014
Papale E., Azzolin M., Giacoma C. (2012a). Vessel Traffic Affects Bottlenose Dolphin (Tursiops Truncatus) Behaviour in Waters Surrounding Lampedusa Island, South Italy. Marine Biological Association of the United Kingdom. J. Mar. Biol. Assoc. Unite. Kingdom. 92 (8), 1877. doi: 10.1017/S002531541100083X
Papale E., Azzolin M., Giacoma C. (2012b). Vessel Traffic Affects Bottlenose Dolphin (Tursiops Truncatus) Behaviour in Waters Surrounding Lampedusa Island, South Italy. J. Mar. Biol. Assoc. Unite. Kingdom. 92 (8), 1877–1885. doi: 10.1017/S002531541100083X
Paradell O. G., López B. D., Methion S. (2019). Modelling Common Dolphin (Delphinus Delphis) Coastal Distribution and Habitat Use: Insights for Conservation. Ocean. Coast. Manage. 179, 104836. doi: 10.1016/j.ocecoaman.2019.104836
Pesante G., Collet A., Dhermain F., Frantzis A., Panigada S., Podestà M., et al (2002). “Review of Collisions in the Mediterranean Sea,” in Proceedings of the Workshop “Collisions Between Cetaceans and Vessels: Can We find Solutions?” Eds. Pesante G., Panigada S., Zanardelli M. (15th Annual Meeting of the European cetacean Society, Roma: ECS Newsletter).
Pirotta E., Carpinelli E., Frantzis A., Gauffier P., Lanfredi C., Pace D. S., et al. (2021). Physeter Macrocephalus (Mediterranean Subpopulation). IUCN. Red. List. Threat. Specie. e.T16370739A50285671. doi: 10.2305/IUCN.UK.2021-3.RLTS.T16370739A50285671.en
Podestà M., Azzellino A., Cañadas A., Frantzis A., Moulins A., Rosso M., et al. (2016). Cuvier's Beaked Whale, Ziphius Cavirostris, Distribution and Occurrence in the Mediterranean Sea: High-Use Areas and Conservation Threats. Adv. Mar. Biol. 75, 103–140. doi: 10.1016/bs.amb.2016.07.007
Praca E., Gannier A. (2008). Ecological Niches of Three Teuthophageous Odontocetes in the Northwestern Mediterranean Sea. Ocean. Sci. 4, 49–59. doi: 10.5194/os-4-49-2008
Praca E., Gannier A., Das K., Laran S. (2009). Modelling the Habitat Suitability of Cetaceans: Example of the Sperm Whale in the Northwestern Mediterranean Sea. Deep-Sea. Res. Part I. 56, 648–657. doi: 10.1016/j.dsr.2008.11.001
Rako N., Fortuna C. M., Holcer D., Mackelworth P., Nimak-Wood M., Pleslić G., et al. (2013). Leisure Boating Noise as a Trigger for the Displacement of the Bottlenose Dolphins of the Cres-Lošinj Archipelago (Northern Adriatic Sea, Croatia). Mar. Pollut. Bull. 68 (1), 77–84. doi: 10.1016/j.marpolbul.2012.12.019
Reeves R., Noterbartolo di Sciara G. (Eds.) (2006). The Status and Distribution of Cetaceans in the Black Sea and Mediterranean Sea (Malaga, Spain: IUCN Centre for Mediterranean Cooperation), 137 pp.
Ryan C., Cucknell A. C., Romagosa M., Boisseau O., Moscrop A., Frantzis A., et al. (2014). “A Visual and Acoustic Survey for Marine Mammals in the Eastern Mediterranean Sea During Summer 2013,” in Unpublished Report to the International Fund for Animal Welfare (Kelvedon, UK: Marine Conservation Research International).
Stephens G., Akkaya A., Hardy J., Awbery T., Rudd L., Abbiss L., et al. (2021). Sightings and Stranding Reports of Fin Whales (Balaenoptera Physalus) in the Levantine Sea. J. Cetacean. Res. Manage. 22 (1), 55–60. doi: 10.47536/jcrm.v22i1.212
Taylor B. L., Baird R., Barlow J., Dawson S. M., Ford J. K. B., et al. (2012)Grampus Griseus. In: IUCN 2013. IUCN Red List of Threatened Species. Available at: www.iucnredlist.org (Accessed 26 April 2021).
Tyack P. L., Zimmer W. M. X., Moretti D., Southall B. L., Claridge D. E., Durban J.W., et al. (2011). Beaked Whales Respond to Simulated and Actual Navy Sonar. PloS One 6 (3), e17009. doi: 10.1371/journal.pone.0017009
Vanderlaan A. S. M., Taggart C. T. (2009). Efficacy of a Voluntary Area to be Avoided to Reduce Risk of Lethal Vessel Strikes to Endangered Whales. Conserv. Biol. 23 (6), 1467–1474. doi: 10.1111/j.1523-1739.2009.01329.x
Woodside J. M., David L., Frantzis A., Hooker S. K. (2006). Gouge Marks on Deep-Sea Mud Volcanoes in the Eastern Mediterranean: Caused by Cuvier’s Beaked Whales? Deep-Sea. Res. 53 (11), 1762–1771. doi: 10.1016/j.dsr.2006.08.011
WWF-IUCN-IWC-OceanMind (2019) WWF-IUCN-IWC-OceanMind: A Geospatial Analysis of Vessel Traffic in Important Marine Mammal Areas. Available at: https://www.marinemammalhabitat.org/mmpatf/activities/geospatial-analysis-of-threats/ (Accessed 27/04/2021).
Keywords: cetacean, spatial distribution, sperm whale, beaked whale, delphinids, marine traffic, AIS
Citation: Awbery T, Akkaya A, Lyne P, Rudd L, Hoogenstrijd G, Nedelcu M, Kniha D, Erdoğan MA, Persad C, Amaha Öztürk A and Öztürk B (2022) Spatial Distribution and Encounter Rates of Delphinids and Deep Diving Cetaceans in the Eastern Mediterranean Sea of Turkey and the Extent of Overlap With Areas of Dense Marine Traffic. Front. Mar. Sci. 9:860242. doi: 10.3389/fmars.2022.860242
Received: 22 January 2022; Accepted: 19 April 2022;
Published: 19 July 2022.
Edited by:
David M.P. Jacoby, University of Lancaster, United KingdomReviewed by:
David March, University of Exeter, United KingdomCaterina Lanfredi, Tethys Research Institute, Italy
Copyright © 2022 Awbery, Akkaya, Lyne, Rudd, Hoogenstrijd, Nedelcu, Kniha, Erdoğan, Persad, Amaha Öztürk and Öztürk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tim Awbery, dGltLmF3YmVyeUBzYW1zLmFjLnVr